














































































































































































































































































































































































































































































































































Author: Грант В.
Tags: экология растений этология растение и окружающая среда ботаника растения эволюция
Year: 1984




Text
В. Грант
Видообразование
у растений
Plant Speciation
Verne Grant
Second Edition
Columbia University Press
New York 1981
В. Грант
Видообразование
у растений
Перевод с английского
Н. О. Фоминой
под редакцией
акад. А. Л. Тахтаджяна
Москва «Мир» 1984
ББК 28.5+28.57
Г 77
УДК 581.5
Грант В.
Г 77 Видообразование у растений: Пер. с англ. — М.: Мир,
1984.—528 с., ил.
Автор монографии — известный американский ученый, уже знакомый совет-
ским читателям по книге «Эволюция организмов» (М: Мир, 1980) Настоящая
книга посвящена новейшим достижениям в области эволюционной ботаники с
широким привлечением данных смежных областей. Рассмотрено значение внутри*
видовой изменчивости, изменчивости популяций и географических рас, зваченне
гибридизации, вегетативного размножения, размножения с помощью семян, а
также других факторов в видообразовании у растений. В качестве примеров при*
ведены давные по более чем 1000 видам растений
Предназначена для ботаников, специалистов в области эволюции и система-
тики растений, для генетиков.
2004000000-454 М0_81 1
041(01)—84
ББК 28.5-j-28.57
58 + 581.4
Верн Грант
Видообразование у растений
Научный редактор М. Б. Николаева. Мл научи редактор 3. В Соллертннская
Художник В. А. Медников. Художественный редактор А. Я Мусин
Технический редактор В П Сизова Корректор В. И Киселева
ИБ № 3817
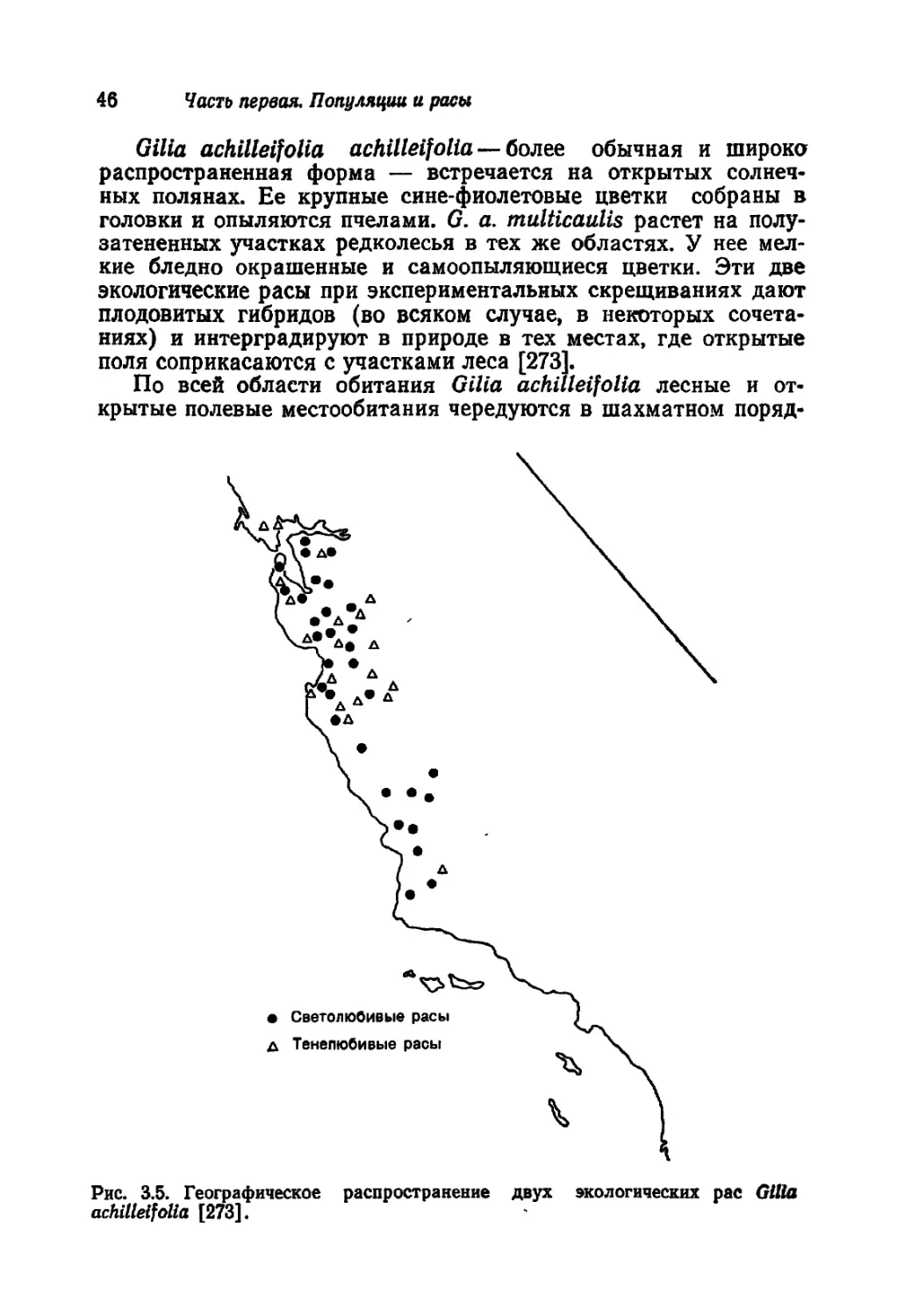
Сдано в набор 16.04.84. Подписано к печати 03 10 84. Формат бОХЭО'Лв
Бумага типографская № 1. Гарнитура литературная Печать высокая Объем 16 50 бум л.
Усл. кр.-отт. 33. Усл. печ. л. 33. Уч.-изд. л. 35. Изд. № 4/2855. Тираж 4000 экз Зак. 45.
Цена 5 руб 60 коп.
ИЗДАТЕЛЬСТВО «МИР». 129820, Москва, И-110, ГСП, 1-й Рижский пер , 2
Московская типография Ns 11 Союзполиграфпрома при Государственном комитете СССР
по делам издательств, полиграфии и книжной торговли.
Москва, 113105. Нагатинская ул., д. 1.
Редакция литературы по биологии
© 1971, 1981 Columbia University Press
© Перевод на русский язык, «Мир», 1984
Предисловие
редактора перевода
Предлагаемая вниманию читателя книга «Видообразование у
растений» принадлежит перу видного американского ботаника,
профессора Техасского университета в городе Остине и члена
Национальной академии США Верна Гранта. Грант —автор
восьми книг, в том числе «Естественная история семейства поле-
мониевых» (1959 г.), «Происхождение адаптаций» (1963 г.),
«Архитектура зародышевой плазмы» (1964 г.), «Колибри и
опыляемые ими цветки» (1968 г., совместно с Карен Грант),
«Генетика цветковых растений» (1975 г.) и «Эволюция организ-
мов» (1977 г.), вышедшая в 1980 г. в русском переводе. Многие
его работы посвящены генетическому и таксономическому изу-
чению большого американского рода Gilia из семейства Polemo-
niaceae.
Книга Гранта посвящена видообразованию у высших расте-
ний, преимущественно цветковых. Хотя Грант касается почти
всех аспектов видообразования, главное внимание он уделяет
генетике (особенно цитогенетике) и репродуктивной биологии.
В книге Гранта читатель найдет детальное и достаточно совре-
менное освещение таких вопросов, как генетические и экологи-
ческие различия видовых особенностей, роль хромосомных пере-
строек в эволюции, роль гибридизации (в том числе интрогрес-
сии) в видообразовании, эволюционная роль полиплоидии и
анеуплоидии и, наконец, роль различных специализированных
генетических систем, включая апомиксис. Все эти разделы со’-
держат много интересных и часто свежих данных и представ-
ляют собой очень ценный обзор проблемы видообразования у
высших растений.
Естественно, что в книге под таким названием автор не мог
обойти общую теорию вида, или эйдологию. Но именно в этой
части книга Гранта является наиболее спорной. Как и в преж-
них своих работах Грант придерживается концепции так назы-
ваемого «биологического вида». Как известно, эта концепция
формировалась в 40-х годах, главным образом под влиянием
6
Предисловие редактора перевода
популяционно-генетических идей Ф. Г. Добржанского, изложен-
ных в первом издании его классической книги «Генетика и про-
исхождение видов» (New York, 1937). Она стала чуть ли не не-
отъемлемой составной частью синтетической теории эволюции
и особенно пропагандировалась Эрнстом Майром. Концепцию
«биологического вида» приняли многие зоологи (в основном
специалисты по наземным позвоночным и энтомологи) и часть
ботаников. В ботанике она имела некоторый успех лишь в 40-х
и отчасти в 50-х годах (в ее пользу высказывался и автор этих
строк). Но постепенно стали выявляться все изъяны этой кон-
цепции и против нее выступил ряд крупных биологов (преиму-
щественно ботаников) как советских, так и зарубежных. В рус-
ской литературе очень убедительную ее критику дал К- М. За-
вадский в своей книге «Вид и видообразование» (Л.: Наука,
1968). Среди зарубежных ученых, выступивших против концеп-
ции «биологического вида», следует упомянуть таких крупных
ботаников, как П. Рейвн (1976) и А. Кронквист (1978).
Против концепции «биологического вида», определяемого
как репродуктивно изолированные скрещивающиеся между со-
бой популяции, можно сделать следующие основные возраже-
ния. Прежде всего, в отличие от таксономической категории
вида, категория биологического вида не универсальна и приме-
нима только к «бипарентальным видам» (термин, употребляе-
мый Грантом), что признает и Грант. Но она не универсальна
даже для бипарентальных видов. Основной критерий «биологи-
ческого вида» — репродуктивная изоляция, нескрещиваемость
с другими' даже близкими видами, — в той или иной мере при-
меним только к симпатрическим видам. Что же касается алло-
патрических видов, то здесь репродуктивная изоляция не имеет
строго облигатного характера. Общеизвестно, что во многих
случаях «хорошие» виды растений, разделенные в течение дли-
тельного времени, исчисляемого миллионами лет, непреодоли-
мыми географическими барьерами, при совместном произраста-
нии в культуре успешно скрещиваются. Таких примеров очень
много. Так, два географически очень удаленных вида плата-
на— Platanus orientalis (Восточное Средиземноморье и Запад-
ная Азия) и Р. occidentalis (восточные районы США) — при
совместном произрастании легко гибридизируют и дают впол-
не плодовитый гибрид с нормально протекающим мейозом.
Однако, несмотря на легкую скрещиваемость этих двух плата-
нов, ни один систематик не согласится объединить их в один
вид. Подобным же образом легко скрещиваются китайский вид
Catalpa ovata и восточно-североамериканский вид С. bignonioi-
des. Как и гибрид платана, гибрид катальпы вполне плодовит
и широко культивируется. С такой же легкостью скрещиваются
многие аллопатрические виды многих других растений, что хо-
Предисловие редактора перевода
7
рошо известно садоводам Таким образом, последовательное
применение критерия нескрещиваемости должно привести к
объединению многих бесспорных аллопатрических видов.
Ограничивая применение концепции «биологического вида»
только бипарентальными («двуродительскими») организмами,
Грант применяет совершенно иные критерии к унипаренталь-
ным («однородительским») группам. К ним он применяет поня-
тие «микровида» (жорданона), принципиально отличное от по-
нятия «биологического вида». Под микровидами Грант пони-
мает однотипные популяции в преимущественно унипаренталь-
ных группах растений, слегка отличающиеся морфологически
друг от друга. Он различает клональные микровиды (размно-
жающиеся вегетативно), агамоспермные микровиды, гибридные
по конституции гетерогамные микровиды (например, Rosa cani-
па) и, наконец, автогамные микровиды. Грант прав в том, что
с таксономической точки зрения эти группы растений известны
своей сложностью, и он, конечно, прав, что концепция биологи-
ческого вида к ним неприменима. Он считает, что эти группы
представляют собой «смесь», состоящую обычно из нескольких
первоначальных «биологических видов» и многочисленных
производных «микровидов» По его мнению, эта «смесь» не мо-
жет быть разложена на составляющие ее элементы «обычными
таксономическими методами». Такое утверждение — следствие
неправильного представления автора как о принятой в совре-
менной -систематике концепции вида, так и о современных ме-
тодах систематики
Грант определяет таксономический вид как «группу морфо-
логически сходных индивидуумов» и считает его (в отличие от
«биологического вида») «искусственной категорией». Мало кто
из современных систематиков согласится с определением таксо-
номического вида как группы, сходной лишь морфологически.
Прежде всего современный систематик старается не ограничи-
ваться лишь внешней морфологией, но все чаще использует
данные внутренней морфологии, включая ультраструктуру, на-
пример ультраструктуру экзины пыльцевых зерен. Но, кроме
того, для современной систематики характерно возрастающее
стремление опираться также на данные биохимии, генетики, ци-
тогенетики и даже молекулярной биологии. Наконец, для совре-
менной систематики вид есть отнюдь не чисто морфологическое,
но прежде всего морфолого-эколого-географичесьое понятие.
Таксономический вид— совершенно реальная категория, без
которой не, может обойтись ни одна область теоретической и
прикладной биологии, сельского хозяйства, лесоводства и т. д.,
а также охрана природы. Совершенно очевидно, что любой био-
" ^гический объект, с которым имеет дело исследователь или
рактик, должен быть определен до вида.
8
Предисловие редактора перевода
Вид—это основная категория таксономической иерархии и
одна из самых фундаментальных категорий биологии. Его мож-
но определить как обособившуюся в процессе эволюции систему
клонов или популяций, объединенную общими признаками .(Мор-
фологическими, экологическими, биохимическими, генетически-
ми, цитологическими и другими), общим происхождением и об-
щим географическим ареалом и достаточно четко отделенную
от близких видов как совокупностью своих признаков, так и раз-
личными (у разных видов разными) изолирующими барьерами.
Это самое общее определение вида, применимое как к бипарен-
тальным, так и к унипарентальным организмам, как к популя-
ционным, так и к клональным образованиям. Для систематики
такие категории, как «биологический вид», «микровид» (жорда-
нон) или «полувид» излишни и в таксономической иерархии им
нет места. Следует также помнить, что такие понятия, как
«сингамеон», «экотип», «климатип», «биотип», «изореагент» и
прочие, используемые в генетических и генэкологических иссле-
дованиях, также нельзя считать таксономическими категориями.
Несмотря на хорошо известную неравноценность видов у раз-
ных групп организмов, их разную структуру и разные спосо-
бы размножения, категория вида в современной биологии явля-
ется универсальной. Наибольшие сомнения вызывают виды у
облигатно-самоопыляющихся и агамных растений. Всем изве-
стны неимоверные трудности, с которыми сталкиваются систе-
матики, изучающие виды ястребинки (Hieracium) или одуван-
чика (Taraxacum). Но трудности здесь скорее технические, чем
принципиальные. Используя различные современные методы си-
стематики, включая молекулярно-биологические, и изучая объ-
екты в природе и на экспериментальных делянках, опытный
систематик почти всегда установит естественные границы вида,
его объем и связи с родственными видами. Поэтому, вопреки
мнению Гранта (и многих других авторов), здесь вполне воз-
можно объединить близкородственные «жорданоны» в настоя-
щие линнеевские виды.
Несколько слов об употреблении Грантом таксономической
номенклатуры. Он не учитывает одного важного различия двух
международных кодексов номенклатуры — ботанического и зоо-
логического. Последний регламентирует название только одной
категории таксонов рангом ниже вида — название подвида, со-
стоящее из трех слов. При этом триноминальное название пи-
шется без предшествующего третьему члену слова, обозначаю-
щего ранг (например, подвид домовой мыши — Mus musculus
domesticus). В ботанической номенклатуре признается несколь-
ко таксономических категорий рангом ниже вида (подвид, раз-
новидность, форма, подформа), требующих указания их ранга.
Грант не следует этому обязательному правилу и постоянно
Предисловие редактора перевода
9
употребляет триноминальные названия без указания ранга внут-
ривидового таксона (например, Gilia сапа speciosa). Неясно, ка-
ков таксономический ранг третьего члена.
Последняя глава книги Гранта «Вид и макроэволюция» со-
держит ряд положений, с которыми я не могу полностью согла-
ситься. Но так как предисловие — не самое подходящее место
для полемики, то я позволю себе отослать читателя, интересую-
щегося этими вопросами, к моей статье о макроэволюции в мире
растений, опубликованной в 1983 г. («Макроэволюционные
процессы в истории растительного мира». — Бот. журн., 1983,
т. 68, № 12, с. 1593—1603).
В заключение я хотел бы сказать несколько слов о почти
полном отсутствии ссылок на русскую литературу в книге Гран-
та. Давно прошли те времена, когда западному ученому можно
было более или менее обойтись без ссылок на русскую литера-
туру. Положение резко изменилось уже в первые десятилетия
нашего века, а в настоящее время любой ученый просто обязан
знать важнейшие советские работы по своей специальности. Неко-
торые видные западные ученые, например крупнейшие амери-
канские ботаники Л. Дж. Стеббинс и А. Кронквист, давно это
поняли и успешно пользуются русской литературой, добросо-
вестно ссылаясь на важнейшие работы. В этом отношении мо-
жет служить примером последняя книга А. Кронквиста «Инте-
грированная система классификации цветковых растений»
(A. Crohquist, An integrated system of classification of flowering
plants. Columbia University Press, New York, 1981), в которой
после каждого раздела приводится обширный список русской
литературы, набранный кириллицей. Поэтому вызывает некото-
рое удивление отсутствие у Гранта ссылок на книгу В. Л. Кома-
рова «Учение о виде у растений» (М. — Л., 1940), статью
Н. И. Вавилова «Линнеевский вид как система» (1931), книгу
М. А. Розановой «Экспериментальные основы систематики расте-
ний» (М. — Л., 1946) и книгу К. М. Завадского «Вид и видооб-
разование» (Л.: Наука, 1968), классическую работу А. С. Семе-
нова-Тяныпанского «Таксономические границы вида и его под-
разделения» (Зап. Имп. Акад, наук, 1910, т. 25, 1, с. 1—29), из-
данную в том же году на немецком языке в Германии. Не упо-
минаются также цитогенетические работы Г. А. Левитского.
И все же, несмотря на эти, а возможно, и некоторые другие
недостатки, выход в свет книги Гранта представляет собой
крупное событие в ботанической литературе. Я уверен, что пуб-
ликация на русском языке насыщенной фактами и оригиналь-
ной по построению книги Гранта принесет большую пользу бо-
таникам-систематикам, экологам, цитогенетикам, а также расте-
ниеводам и селекционерам и стимулирует исследования в этих
направлениях.
А. Л. Тахтаджян
Предисловие
к русскому изданию
Первое английское издание «Видообразования у растений» выш-
ло в свет в 1971 г., а переработанное второе издание — в 1981 г.
Я очень рад, что моя книга выходит в русском переводе и что
тем самым содержащийся в ней материал станет доступным
более широкому кругу читателей в Советском Союзе.
В «Видообразовании у растений» рассматривается эволюция
рас, видов и групп видов у наземных растений и в особенности
у цветковых растений. Обсуждается природа этих биологиче-
ских единиц и порождающие их эволюционные процессы.
Вид представляет собой основную единицу органической
эволюции, но, к сожалению, об этом иногда забывают. Некото-
рые биологи пытаются объяснить эволюцию как результат всего
двух процессов — мутационного процесса и отбора, не прини-
мая во внимание вид и видообразование. Разумеется, сочета-
нием мутационного процесса и отбора действительно можно
объяснить молекулярную эволюцию, в значительной степени —
эволюцию бактерий и простые изменения, происходя-
щие в процессе эволюции эукариотических организмов.
Однако этими двумя процессами нельзя адекватно объяснить
эволюцию самых сложных эукариотических организмов. Эволю-
ционные изменения, происходящие в сложных организмах, дол-
жны быть пропущены через вид. Как показано в этой книге,
это не только теоретическая необходимость, но и факт, установ-
ленный наблюдениями.
Связь между видом и эволюцией была незыблемо закрепле-
на Дарвином в заглавии его классического труда «О происхож-
дении видов» и многократно подчеркивалась многими эволю-
ционистами на всем протяжении нынешнего века.
* Высшие растения — это, безусловно, сложные эукариотиче-
ские организмы, и в процессе эволюции они проходят через ря-
ды последовательно сменяющих друг друга видов. Многие за-
коны эволюции, установленные на дрозофиле, птицах и млеко-
питающих, вполне приложимы к высшим растениям. Но в ре-
Предисловие
11
продуктивном поведении и видовой организации высших расте-
ний есть также некоторые особенности, присущие им одним, что
и определяет характер их эволюционного развития, отличный
от эволюции большинства животных. Эти особые черты эво-
люции растений описаны в моей книге.
Переходя ог общих принципов и широко распространенных
моделей к отдельным примерам, нельзя не заметить, что до-
вольно многие приводимые в книге примеры относятся к флоре
западных районов Северной Америки. Такой выбор отражает
особенности научной биографии автора. Западная часть Север-
ной Америки — это, во-первых, огромная лаборатория, предо-
ставленная нам самой природой для изучения эволюции расте-
ний, а во-вторых, это лаборатория, в которой я жил и в кото-
рой протекала большая часть моей научно-исследовательской,
работы
Я очень хорошо понимаю, что русская ботаническая и биоло-
гическая литература отражена в настоящей книге крайне недос-
таточно. В этом повинен языковый барьер Большое значение
переводов с русского на английский и с английского на русский,
состоит в том, что они позволяют преодолевать этот барьер
Я хочу воспользоваться предоставившейся мне возможностью
высказать свою признательность акад. А. Л Тахтаджяну и
проф. А. В. Яблокову, рекомендовавшим «Видообразование у
растений» к переводу, Н. О. Фоминой, сделавшей перевод, и.
акад. А. Л. Тахтаджяну, отредактировавшему его, а также из-
дательству «Мир». Книге очень повезло, что редактором рус-
ского издания оказался акад. Тахтаджян.
Остин, Техас, США Верн Грант
Предисловие
Эта книга посвящена явлениям видообразования у высших рас-
тений Видообразование рассматривается в ней как процесс,
имеющий много различных аспектов и подверженный влиянию
многих факторов. Мы начнем с описания популяций и рас
(часть первая). Затем перейдем к обсуждению природы видов
и их поведения (части вторая и третья). Далее рассматрива-
ются первичная дивергенция видов (часть четвертая), естест-
венная гибридизация и гибридное видообразование (часть пя-
тая), Полиплоидия, агамоспермия и другие специализирован-
ные генетические системы, связанные с гибридностью, а также
гибридные комплексы, в основе которых лежат такие генетиче-
ские системы, описаны в частях с шестой по восьмую вклю-
12
Предисловие
чительно. Последняя глава (часть девятая) содержит краткий
очерк макроэволюции в свете исследований, проводимых на
видовом уровне.
Короче говоря, в этой книге всесторонне трактуется эволю-
ция растений на уровне видов и групп видов, главным образом
применительно к высшим растениям.
Автор исходит из предпосылки, что естественный, или эво-
люционирующий, вид — одна из основных единиц организации
живой материи. Эволюционирующий вид, подобно всем другим
биологическим единицам, обладает рядом особых общих
свойств, которые можно выявить и объяснить. Эти свойства не
удается обнаружить, если рассматривать видовые единицы все-
го лишь как бесконечный ряд отдельных сущностей, имеющих
свои специфические особенности, т. е. так, как они рассматри-
ваются в чисто описательной таксономии, хотя подобный под-
ход позволяет заложить необходимый фундамент. Их можно
выявить, применяя те методы анализа и синтеза, которые успеш-
но используются в других разделах теоретической биологии.
Первое издание этой книги, опубликованное в 1971 г., широ-
ко используется. Однако за десять лет, прошедшие со времени
выхода в свет первого издания, в той области биологии, которая
занимается изучением растительных видов, произошло много
событий. В течение этого времени не только появлялось мно-
жество новых исследований, но и развивались мои собственные
представления об этой области биологии. Настало время для
коренного пересмотра текста.
Во втором издании несколько изменено распределение ма-
териала по главам. Добавлен ряд новых глав. Весь текст попол-
нен новыми данными. Обсуждение некоторых концепций дове-
дено до современного уровня.
Несмотря на то, что в новом издании библиография была
значительно расширена, она все еще остается далеко не полной.
Литература по этим проблемам столь обширна, что при состав-
лении обзоров приходится производить некоторый отбор. К со-
жалению, в процессе такого отбора многие хорошие работы не-
избежно выпадают из поля зрения.
Карин А. Грант, критически прочитавшая всю рукопись при
подготовке первого издания книги, прочитала все новые главы,
написанные для этого издания. Она взяла на себя также ос-
новной труд по чтению корректур.
Перепечатка рукописи была выполнена Дугласом Дьюком и
Маргарет Буленджер. Я выражаю им свою благодарность.
Д-р Викки П. Реберн и другие редакторы издательства Ко-
лумбийского университета, как всегда, оказывали мне всяческую
помощь и содействие.
Отделение ботаники В. Г.
Университет штата Техас
Январь 1981
Часть первая
Популяции и расы
1 Размножение растений
Одно из основных свойств живой материи — ее способность к
размножению. Этой способностью наделены организованные
единицы самых разнообразных размеров и сложности — от мак-
ромолекул ДНК, хромосом и клеток до отдельных организмов
и скрещивающихся популяций. В этой книге нас интересует
одна из важнейших размножающихся единиц живого мира, а
именно вид. Концепция вида как размножающейся единицы
имеет вековую давность и кратко сформулирована в известном
положении: «подобное рождает подобное».
У большинства животных и у многих растений размножение
неразрывно связано с полом. Но высшие растения произошли в
процессе эволюции от одноклеточных форм, у которых половой
процесс и процесс размножения не только обособлены, но даже
антагонистичны. Размножение у простейших происходит путем
деления: одна клетка делится на две. Половой процесс у этих
организмов происходит путем конъюгации: две клетки сливают-
ся в одну. Таким образом, весь жизненный цикл простейших
слагается из чередования фаз клеточного деления, ведущего к
увеличению численности популяции, и полового процесса, ве-
дущего к уменьшению числа особей.
Это глубокое различие между половым процессом и размно-
жением сохраняется в скрытой форме у высших растений и
животных. Наличие у высших организмов полового процесса
означает, что для осуществления размножения необходимо уча-
стие двух особей, тогда как в отсутствие этого процесса размно-
жение обеспечивает одна особь с помощью того или иного спо-
соба почкования.
У высших растений обособленность этих двух процессов
можно продемонстрировать как на физиологическом, так и на
популяционном уровне. У многих или во всяком случае у боль-
шинства многолетних растений наряду с половым наблюдается
и вегетативное размножение. Как половое, так и вегетативное
размножение приводит к истощению пищевых запасов роди-
тельских растений; оба способа требуют определенных затрат
14
Часть первая. Популяции и расы
энергии. А поскольку запасы энергии у родительских растений
ограничены, такое столкновение потребностей полового и веге-
тативного размножения разрешается у многих растений с по-”
мощью отрицательной корреляции между этими противополож-
ными способами размножения [703, 705]; см. также более позд-
ние работы, например [351]. Обильное образование семян
нередко сочетается со слабым вегетативным размножением, и
наоборот, мощное вегетативное размножение часто сопровож-
дается уменьшением количества семян.
Так, у некоторых сортов земляники удаление усов вызывает
заметное увеличение количества плодов, тогда как другие ре-
монтантные сорта выбрасывают мало усов или не образуют их
вовсе [380, 703]. Барвинок {Vinca major) обычно широко рас-
пространяется с помощью длинных ползучих побегов, но обра-
зует мало плодов; если, однако, обрезать эти ползучие побеги
и не допускать образования новых, то растения начинают завя-
зывать больше семян [703].
Важное значение размножения как такового достаточно оче-
видно. Это — необходимое условие для продолжения существо-
вания популяции или вида во времени. И это единственный спо-
соб увеличения числа особей и заселения новых территорий.
Функция полового процесса, оправдывающая его включение
в жизненный цикл, несмотря на то что он изменяет эффектив-
ность размножения популяции или вида, менее очевидна и все
еще остается предметом дискуссий. Принято считать, что поло-
вой процесс, связанный с чередованием перекрестного оплодо-
творения и мейоза, служит механизмом, обусловливающим ре-
комбинацию генов. Рекомбинация генов в свою очередь пред-
ставляет собой главный источник наследственной изменчивости
у видов с половым размножением. А наследственная изменчи-
вость обеспечивает соответствующие реакции видов на неодно-
родные или изменчивые условия среды.
Отсюда следует, что вегетативное и другие формы бесполого
размножения обладают преимуществом в более или менее по-
стоянной среде, к которой данный вид уже хорошо адаптирован,
поскольку эти формы размножения препятствуют образованию
слабо адаптированных рекомбинационных типов или сводят его
к минимуму, тогда как образование проростков в результате
полового процесса создает преимущество в изменяющейся или
неоднородной среде [280, 709, 759, 785]. Существуют среды и
того и другого типа, и необходимы оба способа размножения.
Очевидно, что в системе размножения вида, процветающего на
протяжении длительного периода времени, должны сочетаться
половой и бесполый процессы.
В самом деле, системы размножения растений сочетают в
себе и половые, и бесполые способы размножения. Каждый из
Гл. 1. Размножение растений
15
этих способов выполняет свою функцию в жизни данного вида.
Однако каждый из них оказывает при этом свое особое влияние
на природу данного вида и его структуру.
Отклонения от случайного перекрестного
оплодотворения
При проведении анализа удобно постулировать идеальное соче-
тание условий для полового размножения в некой гипотетиче-
ской популяции, а затем рассмотреть различные отклонения от
этого идеального состояния, наблюдаемые в реально сущест-
вующих популяциях.
Допустим, что для нашей гипотетической популяции харак-
терно свободное скрещивание, или панмиксия. Она не только
размножается исключительно путем перекрестного оплодотво-
рения, но при этом перекрестное оплодотворение происходит в
таких условиях, что гаметы объединяются в пары чисто случай-
ным образом. Предполагается, что любая женская гамета, воз-
никшая в этой популяции, с равной вероятностью может быть
оплодотворена любой мужской гаметой. Отсюда следует, что
при панмиксии особи, составляющие поколение потомков в дан-
ной популяции, представляют собой продукты разных пар га-
мет, взятых случайным образом из фонда гамет, созданного ро-
дительским поколением.
У реально существующих видов растений отклонения от пан-
миксии обычно выражаются в следующих четырех формах:
1) перекрестное оплодотворение происходит преимущественно
между соседними растениями (вицинизм) и приводит к скре-
щиванию между родственными особями, или инбридингу;
2) размножение происходит путем самооплодотворения (автога-
мия)— теснейшая форма инбридинга; 3) новые растения раз-
виваются из почек, образуемых родительским растением (веге-
тативное размножение); 4) новое поколение растений возникает
из семян, развивающихся на материнском растении без оплодо-
творения (агамоспермия)
Первые два из перечисленных здесь способов размножения—
вицинизм и автогамия — слагаются из оплодотворения и следую-
щего за ним мейоза и представляют собой, таким образом, фор-
мы полового размножения. Два последних способа — вегета-
тивное размножение и агамоспермия — обходятся без оплодо-
творения и представляют собой, следовательно, формы бесполо-
го или апомиктического размножения.
Всех особей, возникших из одной родительской особи в ре-
зультате митотических делений, называют членами одного кло-
на. 1аким образом, вегетативное размножение и агамоспермия
ведут к образованию клона или ряда клонов.
16
Часть первая. Популяции и расы
Существует также важное различие между первым из пере-
численных выше способов размножения и тредоя остальными.
Только этот первый способ требует перекрестного оплодотворе-
ния. В трех других случаях оплодотворения либо не происходит
вовсе, либо в нем участвуют гаметы, образованные одним роди-
тельским растением. Соответственно в первом случае размно-
жение осуществляется при участии двух родительских особей
(бипарентальное размножение), а в трех остальных—лишь од-
ной (унипарентальное размножение).
Совершенно очевидно, что популяция, состоящая из особей,
возникших в результате унипарентального размножения, будь
то клоны или инбредные линии, обладает одним важным отли-
чием от настоящей скрещивающейся популяции, члены' которой
возникают в результате настоящего полового размножения.
Случайное скрещивание в обширной популяции — идеали-
зированная ситуация, полезная в качестве некоего исходного
стандарта. Возможно, что в действительности не существует ни
одного широко распространенного вида растений, достаточно
близкого к этому состоянию. У известных нам видов растений,
размножающихся половым путем, наблюдается та или иная
степень вицинизма. Однако у некоторых реальных раститель-
ных видов обнаружено крайне резкое отклонение от случайного
скрещивания в сторону строго унипарентального размножения.
В некоторых группах растений бипарентальное размножение
может быть полностью заменено вегетативным размножением,
агамоспермией и, возможно, автогамией.
Гораздо чаще, однако, встречается некое сочетание унипа-
рентального и бипарентального размножения у одного и того
же растения. Такие растения занимают промежуточное поло-
жение между растениями со строго унипарентальным и строго
бипарентальным размножением. В сущности, как отмечает
Густафсон [317], между истинно половым размножением и
различными способами унипарентального размножения имеется
ряд переходных типов.
Вегетативное размножение
Существует много различных способов вегетативного размноже-
ния: при помощи наземных столонов и ползучих побегов, под-
земных корневищ и клубней, укорененных отводков, клубнелу-
ковиц и луковиц, придаточных почек на срезанных стеблях или
опавших листьях и выводковых почек, образующихся в цветке
или соцветии. Последний способ называют живорождением; бо-
лее правильно называть его ложным живорождением [639].
Вегетативное размножение широко распространено у много-
летних покрытосеменных, встречаясь во всех основных группах.
Гл. 1. Размножение растений
17
Среди многолетников виды, не способные размножаться вегета-
тивным путем, крайне редки. Густафсон [317] приводит данные,,
свидетельствующие о том, что в некоторых скандинавских фло-
рах примерно 80% всех видов покрытосеменных способны к
той или иной форме вегетативного размножения, а примерно у
50% многолетних покрытосеменных различные формы вегета-
тивного размножения достигли такого развития, что дают им:
возможность очень быстро расселяться.
Секвойя вечнозеленая {Sequoia sempervirens) способна да-
вать корневую поросль, представляя собой в этом отношении
исключение среди хвойных [406]. Когда старое дерево отмирает,
из корневой поросли вырастают молодые деревца, которые в
конце концов превращаются в большие деревья, окружающие
кольцом исходный родительский пень. Со временем вокруг от-
дельных деревьев, образующих такое вторичное кольцо, могут
образоваться третичные кольца. Поскольку секвойя может до-
живать до 1300 лет ([406]; см. также [352]), продолжительность,
жизни этих клонов приходится исчислять в тысячелетиях
Согласно оценкам Джепсона [405], в лесу примерно 80% зре-
лых деревьев секвойи происходят из корневой поросли и при-
мерно 20%—из семян. По нашим наблюдениям, деревья, воз-
никшие в результате полового и бесполого размножения, рас-
пределены по территории рощи и вокруг нее далеко не случай-
ным образом. В густой центральной части некоторых рощ раз-
множение происходит почти исключительно за счет корневой
поросли, тогда как проросткам удается выжить только на от-
крытых периферических участках.
У некоторых видов дубов имеются стелющиеся подземные
побеги, которые в дальнейшем могут дать начало новым расте-
ниям Например, по данным наблюдений в Техасе, молодые по-
беги Quercus virginiana иногда бывают соединены с переплете-
нием подземных побегов, расходящихся от расположенного в.
центре дерева (рис. 1.1). Зрелые деревья этого вида часто об-
разуют островки или клональные колонии [590].
Осины Populus tremuloides и Р. grandidentata дают мощ-
ные корневые отпрыски от пня. Впоследствии соединения между
корневыми отпрысками и родительским растением отмирают,
так что возникают обособленные молодые деревья. В природе
осина, как правило, встречается в виде клонов, в которые мо-
«ет входить от нескольких деревьев до большого их числа
[403].
Некоторые клоны Populus tremuloides в шт. Юта достигают
значительных размеров. Один такой клон занимал площадь в
10 га и состоял из 15000 деревьев; другой занимал более 40 га,
а которых было насчитано 47 000 деревьев, а третий — около1
га. в восточной части Северной Америки клоны Р. tremuloi-
2—45
Б
Рис. 1.1. Клонирование у Quercus virginiana (с изменениями по 1[590]).
А. Небольшая группа молодых деревьев. Б. Часть подземного побега; обра-
тите внимание на ветвление. В. Схема расположения подземных побегов у
одного клона; черными кружками показано местоположение воздушных по-
бегов.
Гл. 1. Размножение растений
19»
des не столь велики. Самый большой клон, обнаруженный в.
этом районе, занимал чуть больше 1,5 га [430].
Обширные клоны осины должны быть довольно старыми, и
некоторые из них, возможно, возникли еще в позднем плейсто-
цене. В горах шт Юта Populus tremuloides образует много жиз-
неспособных семян, во всяком случае в отдельные годы, но в
современных климатических условиях, когда в начале лета
осадков выпадает мало, доля прорастающих семян ничтожна.
В этой области осина размножается, по-видимому, только веге-
тативным способом. Поскольку климат, для которого типичны
летние засухи, сложился в Юте уже 8000 лет назад, возможно,
что и возраст некоторых клонов осины также достигает
8000 лет. На протяжении столетий или тысячелетий вегетатив-
ного размножения особой расе осины, рано покрывающейся
листвой, удалось заселить вершины гор в штате Юта, которые
прежде находились подо льдом и снегом [125].
Festuca rubra представляет собой многолетний злак, размно-
жающийся как вегетативно, при помощи корневищ, так и поло-
вым путем и являющийся ветроопыляемым и самонесовмести-
мым видом. Харберд [338] исследовал состав популяций этого'
злака в Шотландии, основываясь на выборках, взятых с обшир-
ных квадратов. Растения, обладающие различными фенотипи-
ческими признаками и плодовитые при перекрестном опылении,
рассматривались как принадлежащие к разным генотипам.
Обоюдно-несовместимые растения с одинаковыми фенотипиче-
скими признаками рассматривались как разные клоны одного-
и того же генотипа.
В результате изучения фенотипических признаков выборок и
их поведения при скрещивании Харберд [338] пришел к выводу,
что в квадрате площадью 80 м2 представлено относительно не-
много разных генотипов, но что многие растения принадлежат
к одному и тому же клону. Один из генотипов был распростра-
нен на участке диаметром около 220 м. По мнению Харберда,
для того чтобы какой-либо клон мог покрыть участок такой пло-
щади, возраст его должен достигать нескольких сотен, а может
быть, и тысячи лет. Некоторые другие клоны были небольшими.
Таким образом, изучение выборок из популяций Festuca rubra
показало, что они состоят в основном из нескольких генотипов,
которые распространились путем клонирования по большим
участкам, а также из нескольких других генотипов, образующих
лишь небольшие клоны. Перекрестное опыление при помощи
ветра у самонесовместимого злака с такой клональной структу-
рой популяции должно обычно приводить к несовместимым ком-
бинациям [338].
У родственного Festuca rubra другого многолетнего вида —
Г. ovina— наблюдается иная картина. F ovina расселяется го-
2*
20 Часть первая. Популяции и расы
раздо медленнее чем F. rubra. У F. ovina Харберд (339] обнару-
жил многочисленные генотипы на квадрате размером около 9 м2.
В этом случае в отличие от F. rubra соотношение между бес-
полым и половым размножением, по-видимому, сдвинуто в сто-
рону полового размножения. '
Прибрежноморскому злаку Spartina patens из восточной час- -
ти Северной Америки также свойственны ауткроссинг, веге-
тативное размножение корневищами и образование клонов. От-
дельные генотипы можно идентифицировать благодаря сущест-
вующим между ними различиям по изоферментам и составить
карту их микрогеографического распространения. В том районе
Северной Каролины, где проводилось изучение, на маршруте
длиной 200 м был обнаружен 101 генотип. Некоторые генотипы
образуют довольно обширные клоны, тогда как другие пред-
ставлены всего несколькими отдельными растениями [724].
Примером исключительно вегетативного размножения слу-
жит хорошо известный случай с Elodea canadensis (Hydrocha-
ritaceae) в Европе. В восточной части Северной Америки, отку-
да происходит это двудомное растение, засоряющее водоемы,
оно размножается как половым путем (семенами), так и веге-
тативно, выбрасывая побеги и образуя зимние почки. Женские
растения были завезены из Северной Америки на Британские
острова примерно в 1840 г. Ввиду отсутствия на чужой для него
территории мужских растений этот канадский сорняк мог раз-
множаться только вегетативно. Тем не менее за период с 1840
по 1880 г. ему удалось распространиться почти по всем внут-
ренним водоемам Европы [317].
Агамоспермия
При агамоспермии отдельное растение образует жизнеспособ-
ные семена, которые содержат зародыши, возникшие без опло-
дотворения. Особенности развития таких зародышей сложны и
варьируют от случая к случаю (см. гл. 31). Новый зародыш в
семени может развиваться из неоплодотворенной яйцеклетки
(партеногенез), из какой-либо другой клетки или ядра зароды-
шевого мешка (апогамия) или же из какой-нибудь соматиче-
ской клетки семязачатка (адвентивная эмбриония). Агамосперм-
ное образование семян может происходить без опыления или же
с опылением (псевдогамия). В последнем случае пыльца сти-
мулирует рост эндосперма, необходимого для нормального раз-
вития семени и зародыша [317].
Различные пути эмбрионального развития сходны в том, что
во всех случаях клеточные линии, ведущие к образованию ново-
го зародыша, обходятся как без мейоза, так и без оплодотворе-
ния. В результате, если не считать некоторых исключительных
Гл. 1. Размножение растений
21
процессов, происходит образование семян, содержащих зароды-
ши, которые генетически идентичны материнскому растению.
Агамоспермия широко распространена среди высших расте-
ний. Она встречается у многих представителей семейств Grami-
neae, Compositae и Rosaceae, а также в ряде более мелких се-
мейств Наиболее хорошо известные примеры относятся к Hiera-
ciutn, Taraxacum, Crepis, Citrus и Роа. Агамоспермные растения
обычно бывают многолетними. Их размножающиеся половым
путем родичи если и известны, то неизменно принадлежат к пе-
рекрестноопыляющимся видам, у которых перекрестное опыле-
ние обеспечивается самонесовместимостью, двудомностью или
какой-либо другой системой размножения по типу ауткроссин-
га; это позволяет считать, что для предков агамоспермных рас-
тений было характерно исключительно перекрестное оплодо-
творение [317].
В некоторых группах растений агамоспермия полностью вы-
теснила половое размножение (облигатная агамоспермия).
У других растений часть семян образуется путем агамоспермии,
а часть — в результате полового процесса (факультативная ага-
моспермия). Как и в случае вегетативного размножения, четкой
границы между половым и бесполым размножением провести
нельзя; эти два способа размножения связаны между собой ря-
дом переходных состояний [317].
При факультативной агамоспермии характер изменчивости
популяции меняется- популяция распадается на ряд групп, сос-
тоящих из идентичных особей, которые отличаются друг от дру-
га по второстепенным признакам. Такая популяция состоит из
скоплений более или менее обособленных агамоспермных мик-
ровидов.
Автогамия
Для гермафродитных покрытосеменных характерен широкий
спектр систем скрещивания — от облигатного ауткроссинга на
одном конце до фактически полной автогамии — на другом. Гер-
мафродитные цветки могут быть полностью самонесовместимы
и не завязывают семян после самоопыления, как у Gilia capita-
ta capitata (Polemoniaceae), или же самонесовместимость мо-
жет быть неполной, как у G. capitata tomentosa, у которой мно-
гие самоопылившиеся цветки образуют всего лишь по несколь-
ку семян [266]. Неполной и частичной может быть также само-
совместимость. Cheiranthus cheiri (Cruciferae) полностью само-
совместим при искусственном самоопылении; если же подверг-
нуть его в одинаковой степени самоопылению и перекрестному
опылению, то большая часть (92%) завязавшихся семян оказы-
вается результатом перекрестного опыления [46].
22
Часть первая Популяции и расы
Полностью самосовместимые покрытосеменные растения
варьируют либо в отношении деталей строения цветка, создаю-
щих преимущества для ауткроссинга или самоопыления, либо-
в отношении того и другого вместе. В природных популяциях
Clarkia unguiculata (Onagraceae), имеющей в высокой степени
протандрические цветки с выступающими наружу репродуктив-
ными органами, частота ауткроссинга, определяемая по испыта-
ниям потомства, составляет 96% [857]. В отличие от этого строе-
ние цветка С. exilis допускает значительное спонтанное само-
опыление, и в двух популяциях этого вида ауткроссинг состав-
лял 43 и 45% [856, 858].
У многих самосовместимых растений тычинки и рыльца од-
ного и того же цветка расположены очень близко друг от друга
и созревают одновременно, так что обычно самоопыление про-
исходит автоматически. Исследователи, изучающие автогамные
растения, единодушно считают, что автогамия обычно не бы-
вает настолько полной, чтобы исключить некоторый ауткроссинг.
Самоопыление может преобладать, однако время от времени
происходит перекрестное опыление, осуществляемое ветром или
насекомыми; поэтому рассматриваемую систему размножения
правильнее называть преимущественной автогамией [5, 425].
Произведенные сравнительно недавно оценки частоты аут-
кроссинга у нескольких видов преимущественно автогамных зла-
ков дали следующие результаты:
Hordeum jubatum
Hordeum vulgar в
Avena barbata
Avena fatua
Bromus mollis
-1% [32]
-1% и 4% [6,398]
-2% [398]
— от 1 до 12% и 3% [394, 398]
-4% [398]
Можно было ожидать, что существуют примеры полной или
почти полной автогамии, и в некоторых случаях такое ожида-
ние действительно оправдалось. По имеющимся оценкам часто-
та ауткроссинга у Festuca microstachys (Gramineae) составляет
0% [6]. Мелкие цветки Polemonium micranthum (Polemoniaceae)
самоопыляются еще в бутонах, а затем, уже после опыления,
раскрываются всего на несколько часов, так что никакой оче-
видной возможности для опыления их насекомыми нет. Попыт-
ки искусственного перекрестного опыления этих растений оказа-
лись безуспешными; все потомство, полученное при этих искус-
ственных скрещиваниях, было результатом самоопыления.
Имеющиеся данные позволяют считать, что некоторые популя-
ции Р. micranthum действительно полностью инбредны [301].
Автогамия или, точнее, преимущественная автогамия — обыч-
ный и широко распространенный способ размножения у покры-
Гл. 1. Размножение растений
23
тосеменных, особенно у однолетних травянистых растений [317,
780]. В сем. Polemoniaceae это наиболее часто встречающийся
способ опыления, установленный в опытах по скрещиванию
примерно у 45 видов и предсказываемый на основании полевых
наблюдений еще у 30 видов. Автогамия в этом семействе встре-
чав 1ся, насколько это известно, только у многолетних его пред-
ставителей [301].
Система размножения сказывается на характере изменчивос-
ти данной популяции. Если два или несколько гомозиготных ин-
дивидуумов, различающихся по двум или большему числу ге-
нов, размножаются путем ауткроссипга, то при этом в резуль-
тате рекомбинации сильно возрастает индивидуальная изменчи-
вость и большинство рекомбинантов остаются гетерозиготными
по крайней мере по одному гену. Непрерывное самоопыление
приводит к противоположным результатам. Если исходная по-
пуляция состоит из индивидуумов, гетерозиготных по двум или
большему числу генов, то с каждым поколением самоопыления
доля гетерозиготных индивидуумов уменьшается с равномерной
скоростью, а первоначальный ряд гетерозиготных и гомозигот-
ных рекомбинантов сводится к меньшему числу гомозиготных
типов Следует ожидать, что строго автогамная популяция дол-
жна состоять главным образом из ряда истинно инбредных чис-
тых линий.
В реальных популяциях автогамных растений эти ожидае-
мые ре'зультаты могут действительно наблюдаться или не наб-
людаться в зависимости от других факторов Прежде всего
автогамия, как мы уже видели, обычно не бывает облигатной;
время от времени в преимущественно автогамной популяции
происходят скрещивания между различными биотипами, служа-
щие источником новой изменчивости [280, 345, 782]. Кроме того,
непрерывное самоопыление не приведет к снижению в популя-
ции доли гетерозигот с ожидаемой скоростью, если эти гетеро-
зиготы обладают каким-либо селективным преимуществом по
сравнению с гомозиготами [356, 401].
Недавними исследованиями Алларда и его сотрудников было
установлено наличие у некоторых преимущественно автогамных
растений постоянной гетерозиготности, обусловленной преиму-
ществом гетерозигот. Это было обнаружено у Secale cereale [5,
401, 403]; Phaseolus lunatus [7, 344]; Avena fatua [394, 404] и
Avena barbata [108].
Фактический состав популяций у многих автогамных видов
растений оказывается, таким образом, гораздо сложнее, чем
следовало бы ожидать на основании экстраполяций от концеп-
ции чистых линий; это в последние годы подчеркивают многие
исследователи. Не следует, однако, думать, что простые модели
нереалистичны во всех случаях. Популяции некоторых автогам-
24 Часть первая: Популяции и расы
ных видов приближаются к простому составу, слагаясь из одно-
го или нескольких гомозиготных биотипов; Популяции других
видов содержат больший запас изменчивости, и частота ауткрос-
синга в них выше, так что они приближаются в этих отношениях
к популяциям видов, у которых регулярно’происходит пере-
крестное опыление.
Рекомбинационные системы
В размножении растений, как было ясно показано в предыду-
щем параграфе, участвуют как процессы, создающие изменчи-
вость, так и процессы, обеспечивающие воспроизведение геноти-
пов. Равновесие между процессами этих двух типов создается,
той частью генетической системы, которая известна под назва-
нием рекомбинационной. Рекомбинационная система устанав-
ливает различные точки равновесия для разных популяций и
видов. В диапазоне от максимальной рекомбинации до макси-
мальной репликации полезно различать три модальных состоя-
ния: открытые, ограниченные и закрытые рекомбинационные-
системы. Для них характерны соответственно широкий ауткрос-
синг, преимущественная автогамия и агамоспермия.
На рекомбинационную систему оказывают влияние многие
факторы, перечисленные в табл. 1.1. Эти факторы обсуждаются
в обзорах Гранта [294] и Солбрига [759]. Важно отметить, что
некая определенная степень регулирования рекомбинации мо-
жет быть достигнута в результате различных сочетанйй этих
регулирующих факторов.
Как указывает Солбриг [759], некоторые факторы, регули-
рующие рекомбинацию, непосредственно контролируются отбо-
Таблица 1.1. Компоненты рекомбинационной системы растений
([280], с изменениями)
I. Факторы, регулирующие количество рекомбинации на 1 поколение; ре-
гуляция осуществляется во время мейоза
1. Число хромосом
2. Частота кроссинговера
3. Стерильность гибридов
IL Факторы, регулирующие количество рекомбинации на 1 поколение; ре-
гуляция осуществляется во время оплодотворения
4. Система скрещивания
5. Система опыления
6. Потенциал расселения
7. Число семян и характер их прорастания
8. Величина популяции
9. Барьеры' для скрещивания и внешние изолирующие механизмы
III. Фактор, регулирующий количество рекомбинации в единицу времени
10. Продолжительность генерации
Гл. 1. Размножение растений
25
ром, тогда как на другие такие факторы отбор не влияет. Ком-
поненты рекомбинационной системы, непосредственно контроли-
руемые отбором, соответствуют факторам 1, 2, 4, 5, 6 и 9
(табл. 1.1). Факторы 7 и 10 контролируются силами отбора, ре-
гулирующими иные характеристики, нежели соотношение реком-
бинации и регуляции как таковое. А факторы 3 и 8 не’ подвер-
жены, согласно Солбригу [759], прямому действию отбора.
Солбриг [758, 759] выдвинул идею о том, что рекомбинаци-
онные системы сопряжены с определенными «расходами» и что
открытые рекомбинационные системы требуют больших расхо-
дов, чем ограниченные или закрытые системы. Открытая реком-
бинация связана с генетическими или так называемыми мейоти-
ческими расходами, измеряемыми числом возникающих плохо
адаптированных рекомбинантов, а также с энергетическими
расходами, связанными с приспособлениями цветков для привле-
чения опылителей и образованием избытка пыльцы для пере-
крестного опыления. Очевидно, что открытые рекомбинацион-
ные системы обладают преимуществами, перекрывающими эти
расходы.
Среднее число пыльцевых зерен на семязачаток действи-
тельно гораздо выше у перекрестноопыляющихся растений, чем
у самоопылителей. Число пыльцевых зерен на семязачаток у
видов с различными системами скрещивания составляет в сред-
нем [132]:
у облигатных перекрестноопылителей —5859
у факультативных перекрестноопылителей — 797
у преимущественно автогамных растений — 168
у облигатпо автогамных растений — 28
Impatiens capensis образует клейстогамные и хазмогамные
цветки и семена, развивающиеся в результате как самоопыле-
ния, так и перекрестного опыления. Энергетические расходы на
семена этих двух типов составляют, согласно оценкам, 135 кал
для семян из хазмогамных цветков и 65 кал для семян из клей-
стогамных цветков [876].
Экология рекомбинационных систем
Причину разнообразия рекомбинационных систем у растения
следует искать в неодинаковости требований, предъявляемых к
размножению местообитаниями разного типа. Здесь вступает в
Действие еще один спектр — от открытых местообитаний до
замкнутых сообществ.
Открытые местообитания, будь они постоянно открытыми,
как в пустыне, или периодически открытыми, как на первых
стадиях экологической сукцессии, или искусственными, как на
26 Часть первая. Популяции и расы
возделываемых полях, благоприятны для заселения. Заселению
в свою очередь способствуют ограниченные и закрытые реком-
бинационные системы. Такие рекомбинационнь^ системы обес-
печивают сохранение постоянства при размножении. Популя-
ция, адаптированная к данному открытому* местообитанию и
обладающая ограниченной или закрытой рекомбинационной
системой, может воспроизводить свой адаптивный генотип (ы) и
быстро увеличивать свою численность в пределах имеющегося
местообитания. Экспериментирование с новыми типами реком-
бинантов, которые могли бы понизить успех-размножения, сво-
дится к минимуму.
Виды растений с ограниченными и закрытыми рекомбина-
ционными системами действительно встречаются обычно в от-
крытых местообитаниях и на ранних стадиях экологической
сукцессии; например, однолетники пустынь, однолетники Среди-
земноморской области, сорняки, травянистые растения первых
стадий экологической сукцессии и агамоспермные растения в
областях, обнажившихся после окончания плейстоценового оле-
денения.
Растения с открытыми рекомбинационными системами, на-
против, встречаются в закрытых растительных сообществах; к
ним относятся, например, деревья и кустарники лесов умерен-
ной зоны, многие злаки степей и равнин и многие многолетние
травянистые растения подлеска.
В закрытых растительных сообществах вероятность выжи-
вания проростков очень мала, что связано с конкуренцией. Рас-
тительная популяция год за годом приносит урожай семян, но
новым проросткам удается сохраниться лишь в тех очень ред-
ких случаях, когда в сообществе освобождается подходящее
место. По имеющимся оценкам, за три последние столетия в
девственном лесу в Нью-Гемпшире из всех проростков бука
Fagus grandifolia сохранялось по одному на 0,04 га в 10 лет
(см. обзор [120]). В случаях, подобных этому, когда раститель-
ная популяция обладает огромной избыточной плодовитостью,
она может позволить себе иметь открытую рекомбинационную
систему и, следовательно, множество плохо адаптированных
рекомбинационных зигот.
Существуют две точки зрения относительно сил отбора, при-
ведших к развитию открытых рекомбинационных систем.
Мое первоначальное мнение [280] сводилось к тому, что вы-
сокая избыточная плодовитость растений в закрытых сообщест-
вах дает им возможность использовать долговременное преиму-
щество открытой рекомбинационной системы, а именно созда-
ние изменчивости, которая позволит справиться с будущими
изменениями условий среды. Если рассматривать это с другой
точки зрения, то можно сказать, что многим существующим в
Гл 1. Размножение растений
27
настоящее время в природе видам растений с открытыми ре-
комбинационными системами удалось сохраниться на протяже-
нии длительного времени и дожить до наших дней благодаря
такой рекомбинационной системе Наблюдаемое во многих со-
временных сообществах скопление подобных видов может быть
результатом междемового или межвидового отбора.
Более простая альтернативная гипотеза была выдвинута не-
давно Левином [472] и Солбригом [758, 759]. Растения в закры-
тых сообществах находятся в условиях непрерывно изменяю-
щейся-биотической среды. Постоянно возникают новые генотипы
патогенных организмов, растительноядных животных и расте-
ний-конкурентов. Открытая рекомбинационная система порож-
дает новые варианты, необходимые растительной популяции для
того, чтобы соответствовать изменяющимся элементам окру-
жающей ее среды. В этом случае открытые рекомбинационные
системы могли развиться в результате индивидуального отбора.
Такая гипотеза весьма привлекательна, по требует дальнейшей
проверки.
Остается вкратце коснуться некоторых терминологических
вопросов. Выражения «возникший в результате r-отбора» и
«возникший в результате /(-отбора», используемые в популя-
ционной экологии, соответствуют заселяющему виду и виду
закрытого сообщества, однако они не содержат никаких указа-
ний относительно типа рекомбинационной системы. Тем не ме-
нее различные авторы применяют эти термины в более широ-
ком смысле, распространяя их и на другие альтернативные ре-
комбинационные системы
Некоторые авторы для обозначения противоположных эко-
логических условий, связанных с рекомбинационными система-
ми разного типа, пользуются терминами «стабильные—неста-
бильные» или «предсказуемые—непредсказуемые». Одиако эти
термины могут привести к некоторой путанице. Так, среда, бла-
гоприятствующая /(-отбору, считается более предсказуемой, чем
среда, способствующая г-отбору [399, 638]. Однако, как мы
только что видели, закрытое сообщество, содержащее типы, воз-
никшие в результате /(-отбора, находится в более непредска-
зуемой биотической среде, чем если бы оно находилось в от-
крытом местообитании. Этой путаницы можно избежать, ис-
пользуя простые описательные термины — открытое местооби-
тание и закрытое сообщество.
2 Локальные расы
Расы — это популяции или системы популяций какого-либо ви-
да, статистически отличающиеся по составу своих генофондов
и по своим генетически детерминированным фенотипическим
признакам от других популяций или систем популяций, относя-
щихся к тому же виду. Это определение охватывает широкий
спектр различных популяционных единиц —от локальных скре-
щивающихся популяций до обширных географических рас. При
обсуждении этих единиц в эволюционном плане удобно разли-
чать два уровня расовой изменчивости: локальные расы и геог-
рафические расы. Первым посвящена данная глава, а вторые
будут рассмотрены в гл, 3.
Мы подходим к проблеме дифференциации локальных рас
с теоретической точки зрения. Будут рассмотрены такие факто-
ры, регулирующие этот процесс, как поток генов и создаваемые
им эффекты, взаимодействие потока генов с естественным отбо-
ром и взаимодействие между отбором и дрейфом генов. Разбор
каждой группы регулирующих факторов сопровождается изло-
жением соответствующих эмпирических данных.
Расселение и поток генов
Растения, будучи постоянно прикреплены к одному месту, спо-
собны расселяться на двух стадиях своего жизненного цикла:
пыльца переносится с помощью ветра, насекомых, птиц или
других факторов; перенос плодов или семян осуществляют раз-
личные физические или биотические факторы. Каждый ботаник,
которому приходилось наблюдать распространение семян в при-
роде, вскоре обнаруживает, что оно следует некой общей моде-
ли: большая часть производимых данным материнским расте-
нием семян, которые не были съедены животными, локализует-
ся поблизости от материнского растения, тогда как небольшое
число семян разносится на более далекие расстояния.
Такого рода общие наблюдения подкреплены в некоторых
случаях количественными данными. Солсбери [704] подсчитал
число семян Verbascum thapsus, унесенных ветром на разные
расстояния от родительского растения. Основная масса семян
упала на землю на расстоянии примерно 3,5 м от родительско-
го растения, однако за пределами этого 3,5-метрового радиуса
число семян быстро сокращалось с увеличением расстояния.
Аналогичная картина наблюдается для распространяемых вет-
ром плодов Senecio jacobaea [704].
Гл. 2. Локальные расы
2»
Такая картина распространения оказалась типичной для ря-
да растений и животных. Кривая распределения расстояний по
частоте положительно эксцессивна и асимметрична. Значитель-
ная доля единиц расселения остается вблизи родительской осо-
би. Затем с увеличением расстояния от этой исходной точки
кривая частот быстро снижается.
Приведем один конкретный пример, хорошо иллюстрирую-
щий это характерное распределение. Колуэлл [117] выбрасывал
Рис. 2 1. Распространение сосновой пыльцы ветром по двум радиусам на?
разные расстояния от источника пыльцы [117]
I — в направлении ветра; II— под углом 45° к направлению ветра
в воздух помеченную радиоактивными изотопами пыльцу сосны
Pinus coulteri и прослеживал за ее распространением по двум,
радиусам от исходной точки. На рис. 2.1 представлена кривая
распределения сосновой пыльцы, переносимой ветром. Большая:
часть пыльцы упала на землю недалеко от источника, пролетев
по ветру от 3 до 10 м. А дальше количество пыльцы быстро-
уменьшалось. На расстоянии от 45 до 120 м от источника хотя<
и было найдено некоторое количество пыльцы, но оно было
очень невелико [117].
Для изучения распространения пыльцы пчелами только что-
раскрывшиеся цветки хлопчатника (Gossyplum hirsutum и
G. arboreum) обрызгивали синей краской и прослеживали за-
последующим распространением краски по хлопковому полю-
[804]. После посещений пчел частицы краски были обнаружены
на расстоянии 25 м от обработанных цветков. Однако по мере-
удаления от места нанесения краски количество меченой пыльцы.
SO
Часть первая. Популяции и расы
равномерно убывало; это позволяет считать, что активность
пчел была сосредоточена на растениях в пределах данного со-
седства [804].
Если описанный выше характер распространения семян про-
должается на протяжении ряда последовательных поколений,
то следует ожидать, что родственные друг другу растения ока-
жутся ближайшими соседями. Если же таким образом будет
распространяться пыльца, то перекрестное опыление будет про-
исходить преимущественно между родственными растениями,
растущими друг подле друга. В результате в популяции с пере-
крестным опылением возникнет значительная степень инбри-
динга. Однако, поскольку в норме в такой популяции происхо-
дит также распространение семян и пыльцы на большие рас-
стояния, инбридинг будет дополняться небольшой, но, вероят-
но, значимой степенью свободного перекрестного опыления [280,
298].
Следует отметить, что хотя характер расселения описывается
общей для разных организмов положительно эксцессивной и
асимметричной кривой, расстояния, в пределах 'которых оно
происходит, могут сильно различаться. Короче говоря, разные
типы организмов обладают различной способностью к расселе-
нию.
Расселение представляет собой физическую основу потока
генов. Сам по себе поток генов наблюдать удается редко;
«обычно о нем судят на основе данных по расселению. Однако
при изучении межсортовых засорений у сельскохозяйственных
культур поток генов можно наблюдать и измерить.
Такие исследования были проведены на многочисленных ви-
дах культурных растений. По степени засорения между двумя
-сортами какого-либо перекрестно опыляющегося вида культур-
ных растений, растущих на соседних экспериментальных участ-
ках опытного поля в практически одинаковых условиях среды
и завязывающих семена после свободного опыления при помо-
щи ветра или пчел, можно судить о величине потока генов меж-
ду ними при данном разделяющем их расстоянии. Как показы-
вают результаты этих исследований, характер потока генов
сходен с характером расселения.
Хорошим примером служит люцерна (Medicago sativa).
Бреднер и др. [64] использовали две линии люцерны, различаю-
щиеся по окраске цветков, детерминируемой одним геном. Ли-
ния с белыми цветками гомозиготна по рецессивному гену, а
нормальная окраска — доминантный признак. Две эти линии
выращивали на отдельных экспериментальных учасФках при
различных расстояниях между участками, и они свободно опы-
лялись местными перепончатокрылыми (Botnbus и Bembix).
Процент засорения среди проростков следующего поколения на
Гл. 2. Локальные расы
3
Таблица 2.1. Засорение линии Medicago
sativa с белыми цветками генами линии с
окрашенными цветками при различных
расстояниях между экспериментальными
участками (64]
Расстояние между экспериментальными участками, м Засорение, осуществлявшееся перепончатокрылыми, %
1962 г 1963 г
50 42,9 19,6
101 30,9 9,7
201 10,0 3,3
403 6,0 6,5
805 5,2 4,4
1208 9,3 0,0
461G 6,5 0,0
участках линии с белыми цветками служит показателем потока
доминантного аллеля (табл. 2.1).
Как показывает таблица, поток доминантного маркирующего’
аллеля очень велик при небольших расстояниях (50 м), но с
увеличением расстояния снижается. Следует, однако, отметить,
что даже при расстояниях от 800 до 1600 м был обнаружен
довольно значительный поток генов [64].
Обычно при небольших расстояниях между эксперименталь-
ными участками с разными сортами степень засорения одного-
сорта другим невелика. На экспериментальных участках сред-
него размера с опыляемой ветром кукурузой (Zea mays), раз-
деленных расстоянием в 15 м, засорение составляло всего 1%
([46]; см. также [635]). У опыляемых насекомыми репы (Bras-
sica гара) и редьки (Raphanus sativus) при таком же расстоя-
нии между экспериментальными участками межсортовое засо-
рение также снижалось до небольшого уровня [46]. У перекрест-
но опыляющейся редьки межсортовое засорение при расстоянии
между участками 4,5 м составляло всего 2% [128].
Панмиктические единицы и соседства
В большой непрерывной популяции какого-либо бипарептально-
го вида панмиктическая единица — это свободно скрещиваю-
щаяся группа, в пределах которой гаметы объединяются слу-
чайным образом [909]. У организмов, у которых свободное скре-
щивание носит локальный характер, панмиктическая единица
представляет собой, в сущности, субпопуляцию. Если же сво-
32 Часть первая. Популяции и расы
*бодное скрещивание, что часто случается, не является локаль-
ным, то следует различать такую единицу, как соседство [911].
В панмиктической единице с числом особей N* происходит
некоторая доля (IJNe) инбридинга. Коэффициент 1/ЛГ® исполь-
зуют для вычисления величины соседства (ЛГв), в котором про-
исходит неслучайное скрещивание. Таким образом, величину
соседства вычисляют, вводя некую чисто гипотетическую пан-
миктическую единицу, в которой имеются отклонения от панмик-
сии [911]. Концепция соседства имеет более широкое примене-
ние, чем панмиктическая единица, а поэтому она более полезна
в популяционной биологии растений;
Область, занимаемая соседством в непрерывной популяции,
^распределенной в двухмерном пространстве, представляет собой
окружность радиусом 2о, где о — среднее квадратичное откло-
нение расстояний, на которые происходит расселение. В этой
области сосредоточено 86,5% родителей особей, находящихся
в центре окружности. Эта оценка представляет собой матема-
тическое следствие описанного выше выведенйя величины со-
седства из гипотетической панмиктической единицы [911]. Вели-
чина соседства оценивается числом размножающихся особей в
области, занимаемой этим соседством, т. е. в окружности с ра-
диусом 2ог.
Величина панмиктической единицы или соседства зависит
•от расстояний, на которые происходит расселение, и от скорости
потока генов. Если индивидуумы, принадлежащие к большой
•непрерывной популяции, обычно мигрируют на далекие расстоя-
ния, то панмиктическая единица или соседство может прибли-
жаться по величине к популяции в целом. Если же индивиду-
умы, составляющие большую непрерывную популяцию, ведут
сидячий образ жизни и расселяются в пределах небольшой об-
ласти, то скрещивания будут происходить исключительно между
соседними индивидуумами. В этом случае панмиктическая еди-
ница или соседство представляет собой небольшую локальную
труппу внутри популяции в целом. Если же расстояния, на ко-
торые происходят миграции, варьируют, так что имеет место
значительная миграция на небольшие и некоторая миграция
на большие расстояния, то панмиктические единицы и соседст-
ва будут иметь промежуточную величину [909].
Оценки величины соседства получены лишь для нескольких
видов растений и животных. По недавним оценкам величины
•соседств у Drosophila pseudoobscura ЛГе=3229 и 6479 для двух
местностей в Колорадо и Уе=4922 и 6436 — для двух местностей
в Калифорнии [132]. Соответствующие оценки для Drosophila
nigrospiracula из трех местностей в Аризоне составили Ne=
•=21256; 55483 и 145780 [415]. Такая величина скрещивающей-
Гл 2. Локальные расы
33
ся группы будет препятствовать дифференцированию локаль-
ных рас.
У тех нескольких видов растений, которые до сих пор изу-
чались с точки зрения величины соседств, скрещивающиеся
группы во всех случаях малы, и в этом опи резко отличаются
от дрозофилы. Оценки величины соседств у четырех видов рас-
тений в Иллинойсе дали следующие результаты-
Phlox pilosa (Polemoniaceae), Лге=75—282 [480]
Liatris aspera (Compositac), Ne — 3Q—191 [481]
Liatris cylindracea (Compositae), Уе=165 [712]
Lithospermum caroliniense (Boraginaceae), Ne=4 [432]
Кроме того, для Linanthus parryae (Polemoniaceae) в Кали-
форнии также, вероятно, характерны небольшие соседства, од-
нако надежных количественных данных по этому виду нет
[203, 204, 207].
Поток генов и расовая дифференциация
Величина панмиктической единицы или соседства оказывает
влияние па распределение генетических вариаций в пределах
популяции в целом. Если не учитывать действия естественного
отбора, то можно считать, что эти генетические вариации име-
ют тенденцию к равномерному распространению в результате
широкого перекрестного опыления. И наоборот, если перекрест-
ное опыление происходит внутри отдельных небольших субпопу-
ляций, то это способствует локальной дифференциации.
Райт [909] на основе математических соображений показал,
что если панмиктическая единица состоит примерно из 10 раз-
множающихся особей, то в обширной непрерывной популяции
может возникнуть значительная локальная расовая дифферен-
циация независимо от действия естественного отбора. Если чис-
ло размножающихся особей в панмиктической единице дости-
гает 100, то следует ожидать, что расовая дифференциация бу-
дет происходить не в локальных, а в региональных масштабах.
Если же свободно скрещивающаяся группа состоит из 1000 и
более особей, то расовая дифференциация невелика и происхо-
дит на значительных расстояниях, поскольку распределение ва-
риаций зависит от системы скрещивания у данного вида. Не-
значительное расселение на далекие расстояния задерживает
региональную дифференциацию в обширной непрерывной попу-
ляции [909].
Величина соседства у перечисленных выше видов растений
создает сильное отклонение от широкой панмиксии [478].
У Phlox pilosa, Liatris aspera и L. cylindracea величина Ne мог-
3-45
34 Часть первая. Популяции и расы
ла бы допускать региональную расовую дифференциацию на
основе одной лишь структуры популяции. Локальная расовая
дифференциация у Lithospermum caroliniense w, вероятно, у
Linanthus parryae может, по-видимому, быть результатом одной
лишь структуры по типу соседств.
В популяциях Linanthus parryae.и Liatris cylindracea дейст-
вительно происходит морфологическая дифференциация в ло-
кальных масштабах [203, 204, 712]. Эта дифференциация не свя-
зана ни с эдафическими факторами, ни с какими-либо иными
известными факторами среды.
Linanthus parryae (Polemoniaceae) — самонесовместимое
однолетнее растение, опыляемое насекомыми и произрастающее
в пустыне Мохаве и на примыкающих к ней горах. Популяции
в западной части пустыни Мохаве полиморфны по окраске цвет-
ков, причем в одних районах формы с голубыми и белыми цвет-
ками встречаются вперемешку, а в других они разделены.
Иногда колонии с чисто голубыми цветками растут вблизи ко-
лоний с исключительно белыми цветками или непосредственно
примыкают к ним, и такая ситуация сохраняется неизменной на
протяжении многих лет (203, 204]. Обе цветовые формы растут
на одной и той же песчаной равнине, и никаких различий в ус-
ловиях их обитания не замечено. Вполне возможно, что эти ло-
кальные расы, различающиеся по окраске цветков, — результат
ограниченного потока генов и величины соседств и что отбор,
определяемый условиями среды, не играл никакой роли в их
возникновении, однако подобный вывод ни в коей мере нельзя
считать бесспорным.
Взаимодействие между потоком генов и
отбором
Как мы видели, большая непрерывная свободно скрещивающая-
ся популяция теоретически может дифференцироваться на ряд
локальных рас на основании одного лишь характера скрещива-
ния и без участия отбора, если область расселения ограничена
и панмиктические единицы или соседства невелики [909]. Этот
теоретический вывод подтверждается некоторыми наблюдения-
ми, которые были приведены выше. В природе, однако, субпо-
пуляции, обитающие в разных частях какой-либо области, не-
редко оказываются в различных условиях среды и, следовательно,
подвергаются разным давлениям отбора. Естественный от-
бор, таким образом, действует в сочетании с системой скрещи-
вания данного вида.
В условиях свободного скрещивания, происходящего в широ-
ких масштабах в большой популяции, влияния отбора, направ-
ленного на адаптацию к той или иной конкретной локальной
Гл. 2. Локальные расы
35
среде, сводятся на нет непрерывным притоком генов из других
местностей. Давления отбора умеренной интенсивности могут
обеспечить сохранение обширных региональных, но не обособ-
ленных локальных рас. И наоборот, лишь очень интенсивный
отбор может обеспечить сохранение локальных рас в условиях
мощного потока генов.
Если, однако, популяция при такой же общей величине ока-
зывается вследствие своей системы скрещивания подразделен-
ной на мелкие панмиктические единицы или соседства, то мате-
риалом для отбора может служить генетическая изменчивость
в каждом локальном участке без вмешательства потока генов
из других участков. Подобное положение вещей благоприятно
для формирования и поддержания под действием отбора ряда
локальных рас, адаптированных к соответствующим средам.
Такая ситуация благоприятствует эволюции вида в целом, по-
скольку любая панмиктическая единица’ или соседство, обла-
дающие признаками, имеющими общую адаптивную ценность,
а не только локальное значение, может распространяться за
счет других скрещивающихся единиц, наделенных менее благо-
приятными признаками [909, 912].
Следует подчеркнуть, что именно равновесие между интен-
сивностью отбора и скоростью потока генов определяет, будет
ли происходить локальная расовая дифференциация или нет.
По своей интенсивности отбор может варьировать от слабого
до сильного, а скорость расселения — от низкой до высокой;
поэтому следует ожидать, что в природе должны встречаться
самые разнообразные сочетания эти двух факторов. Однако
некоторые авторы склонны выделять в качестве некой нормы
какое-либо одно сочетание, низкую скорость расселения и ин-
тенсивность от.бора от умеренной до высокой [196] или высокую
скорость расселения и очень интенсивный отбор [23]. В природе
существует много возможных точек равновесия и много норм.
Некоторые из них способствуют локальной расовой дифферен-
циации, другие же — нет.
Метафора «дисперсности (зернистости) среды» (environmen-
tal grain) позволяет взглянуть па эту проблему несколько ина-
че. Среда обитания данной популяции или системы популяций
обычно пространственно неоднородна. Однако эта пространст-
венная неоднородность может быть «тонкодисперсной» или
«грубодисперсной» в зависимости от величины организмов и
радиуса их расселения [486]. Приведем в качестве примера са-
ванну, где участки леса перемежаются с участками лугов. Пти-
цы одного из видов могут летать над всей саванной и кормить-
ся повсюду, не испытывая давлений отбора ни со стороны лес-
ных, ни со стороны луговых участков. Для этих птиц данная
реда тонкодисперсная Популяция прикрепленных к субстрату
3*
36
Часть первая. Популяции и расы
однолетних растений, расселяющихся на небольшие расстояния,
будет, однако, подвергаться дизруптивным давлениям отбора
лесной и луговой среды по отдельности. Для таки£ растений та
же самая среда оказывается грубодисперсной.
Микрогеографическая расовая
дифференциация
Среди перекрестно опыляющихся растений известны многочис-
ленные случаи, когда различные локальные расы встречаются
в соседних местностях, разделенных расстоянием примерно
100 м или менее, и, несмотря на это, сохраняют свои отличи-
тельные расовые признаки из поколения в поколение. В таких
случаях экологические условия в соседних колониях заметно
различаются. Возможно поэтому, что подобная локализованная
расовая дифференциация обусловлена действием дизруптивного
отбора в отдельных маленьких скрещивающихся группах. Оче-
видно, что в этих случаях дизруптивный отбор превалирует над
потоком генов.
Локальная расовая дифференциация, коррелированная с
различиями в условиях среды, наблюдалась среди прочих у
следующих перекрестноопыляющихся видов (перечисляются в
хронологическом порядке):
Hieracium umbellatum Gilia capitata Clarkia xantiana Galium pumilum Pinus monticola [840] [266, 267] [491] [186] (Скуиллас и Бинхем [766] сообщали о наличии расовых различий, но теперь они представляются фено- типическими модификациями [671])
Potentilla glandulosa Agrostis tenuis Helianthus annuus X bolanderi Diplacus longiftorus Eschscholtzia calif ornica Collinsia heterophylla Pinus albicaulls Pinus murrayana Anihoxanthum odoratum Eucalyptus vlminalis [98] [65] [790] [54] [54] [121] [881] [97] [24, 540] [457]
Гл 2 Локальные расы
37
В примере с Agrostis tenuis выявляются некоторые сущест-
венные черты. Этот свободно скрещивающийся ветроопыляемый
злак имеет более или менее непрерывное распространение на
волнистых холмах в Уэльсе, где такие факторы среды, как поч-
ва, высота, климат и интенсивность выпаса, варьируют. В соот-
ветствии с этими различиями в среде в выборках из популяций
Agrostis tenuis обнаруживаются наследственные или экотипиче-
ские различия. Следовательно, популяции реагируют на отбор,
происходящий в локальной среде, несмотря на непрерывность
распространения и перекрестное опыление с помощью ветра
Эта локальная дифференциация в некоторых случаях обуслов-
лена, вероятно, слабым или умеренным дизруптивным отбором
в сочетании с распространением пыльцы на относительно не-
большие расстояния [65].
В некоторых других местностях на популяции Agrostis te-
nuis действует иная комбинация сил."В районах свинцовых руд-
ников в Уэльсе этот злак растет на почвах с высоким содержа-
нием свинца, а также на прилегающих к ним почвах, не содер-
жащих свинец в токсичных количествах. На почвах этих двух
классов существуют экотипы, не переносящие свинец, и экотипы,
толерантные к нему. В некоторых местах эти две расы очень тес-
но соседствуют и, уж во всяком случае, находятся достаточно
близко одна от другой, чтобы было возможно их перекрестное
опыление при помощи ветра. В этом случае расовые различия,
очевидно, сохраняются благодаря дизруптивному отбору, кото-
рый достаточно интенсивен, чтобы перекрыть эффекты межра-
совых скрещиваний [402, 422, 540].
Какие факторы отбора участвуют в микрогеографической
расовой дифференциации растений? Климатические факторы
обычно изменяются постепенно по макрогеографическим тран-
сектам и поэтому вряд ли могут быть причиной резко выражен-
ной локальной расовой дифференциации. Исключения встреча-
ются на высоких горах в умеренной зоне, где интенсивность та-
кого фактора, как ветер, может изменяться при небольших из-
менениях высоты, и эти изменения коррелируют с различными
экотипами данного вида. Примерами служат Pirius albicaulis,
Р. murrayana и несколько других высокогорных древесных рас-
тений |[97].
В отличие от этого эдафические факторы часто резко изме-
няются на небольших расстояниях. По этой причине, а также
потому, что почва представляет собой чрезвычайно важный ком-
понент среды для семян и проростков, эдафические факторы
особенно хорошо соответствуют роли факторов отбора, приводя-
щего к дифференциации микрогеографических рас у растений.
У большинства растений, перечисленных в приведенном выше
списке, — от классического турессоновского примера с Hieracium
38 Часть первая. Популяции и расы
и до изученного недавно эвкалипта, а также у подробно иссле-
дованной полевицы (Agrostis) и других злаков-в Уэльсе—ло-
кальная расовая дифференциация коррелирует с изменениями
эдафических факторов среды.
Сочетание отбора с генетическим дрейфом
В популяционной системе колониального типа, состоящей из
маленьких изолированных или полуизолированных популяций,
пространственное разделение снижает интенсивность потока ге-
нов и благоприятствует действию генетического дрейфа. Если,
как это часто случается, изолированные популяции живут в раз-
ных средах, то отбор может действовать также в сочетании с
дрейфом генов. Это приводит к образованию ряда локальных
рас, специфически адаптированных к своим локальным средам.
Расовая изменчивость в таких случаях не образует посте-
пенных географических градиентов или клин, а носит несколько
беспорядочный характер. Каждая колония или группа соседних
колоний обладают собственными отличительными чертами.
Предсказанный характер расовой изменчивости наблюдается
у ряда видов растений с популяционными системами колониаль-
ного типа в областях с разнообразными условиями среды. На-
пример, различные виды Cupressus в Калифорнии часто обра-
зуют ряд изолированных рощиц; некоторые из этих рощиц об-
ладают достаточно четко выраженными отличительными приз-
наками, так что специалист по кипарисам может по одному
лишь внешнему виду данного экземпляра, не обращаясь к гер-
барной этикетке, определить, откуда он происходит [904]. Вто-
ростепенная расовая изменчивость хорошо развита также у
различных травянистых растений с колониальной структурой
популяционных систем. Примерами служат Gilia achilletfolia
в Калифорнии, секция Cheiranthus рода Erysimum и Nigella de-
genii на островах Эгейского моря [273, 749, 811].
3 Географические расы
Локальная расовая изменчивость постепенно переходит в геог-
рафическую изменчивость. Различие между этими двумя видами
изменчивости достаточно произвольно, но полезно с практиче-
ской точки зрения. Локальные расы занимают центральное мес-
то в популяционно-биологических исследованиях, тогда как бо-
лее широкие расовые группировки находятся в сфере интересов
систематиков, и их обычно рассматривают как подвиды, а иног-
да как разновидности, различаемые в формальной таксономии.
Характер внутривидовой географической изменчивости у
растений чрезвычайно разнообразен. В сущности, каждый вид
растений в неком описательном смысле неповторим. Тем не ме-
нее все многообразие типов изменчивости"' можно объединить в
несколько общих категорий. Крупные (например, нелокальные)
расы можно разбить на три категории, непрерывные географи-
ческие расы, дизъюнктивные географические расы и экологиче-
ские расы.
Непрерывная географическая изменчивость
В случае непрерывной географической изменчивости локаль-
ные популяции обычно полиморфны, что является результатом
потока генов и уравновешивающего отбора; но по географиче-
ской трансекте можно видеть постепенный сдвиг частоты дан-
ного варианта. Полиморфизм постепенно переходит в локаль-
ные расовые различия, которые в свою очередь постепенно об-
разуют различия на уровне географических рас.
Генетически детерминированные фенотипические признаки,
подверженные внутривидовой изменчивости, образуют, следова-
тельно, градиенты вдоль географических трансект. Подобные
[радиенты признаков носят название клин. Клина — это посте-
пенное изменение какого-либо признака в ряду популяций, ле-
жащих на некой географической трансекте (теоретическое об-
суждение клин см. [201]).
Клинальная, или непрерывная, географическая изменчивость
наблюдается у видов, обладающих более или менее непрерыв-
ной популяционной системой и занимающих область, в которой
наблюдается постепенный переход от одних условий среды к
Другим. Примерами служат виды равнинных злаков и деревьев
лесов умеренной зоны, имеющие широкое распространение.
junlperus virginiana в восточной части США варьирует по
своему биохимическому составу, а также по морфологическим
признакам. Флейк, Радлоф и Тернер [215], используя метод га-
40
Часть первая. Популяции и расы
Рис. 3.1. Общий вид растений и форма цветка у трех представителей группы
Mlmulus longiftorus ([541; номенклатура изменена).
А — М punlceus Б — м. longiftorus longiftorus В —M. longiftorus caly-
cinus
зовой хроматографии в сочетании co специальными методами
нумерической таксономии, проанализировали соединения из
группы терпеноидов в ряде популяций, распределенных по
трансекте, идущей от г. Вашингтона (федеральный округ Ко-
лумбия) до Техаса, на расстоянии около 240 км друг от друга.
Эти авторы обнаружили, что популяции различаются между
собой по содержанию разных терпеноидных компонентов и что,
кроме того, существует постепенная, или клинальная, изменчи-
вость по этому признаку вдоль трансекты, идущей с северо-
востока на юго-запад [215]. У этого же вида наблюдаются сход-
ные постепенные изменения в содержании летучих масел по
другой длинной трансекте — от реки Миссисипи до Небраски
[216].
Группа Mimulus longiftorus (Scrophulariaceae) состоит из
четырех таксонов кустарниковых растений, растущих в южной
Калифорнии. Три из тех форм, которые мы здесь рассмотрим,
таксономически известны как М. longiflorus longiflorus, М. I.
calycinus и М. punlceus, и принадлежат к секции Diplacus рода
Mlmulus [607]; их рассматривают также как виды выделенного
из Mimulus рода Diplacus (54]. Географическую изменчивость
этой группы изучал Бикс [54]. Нас здесь интересует, во-первых,
интерградация между М. longiflorus longiflorus и М. I. calycinus,
а во-вторых, интерградация между М. I. longiflorus и не пол-
ностью изолированным М. punlceus.
Общий вид растений этих трех форм и их цветки показаны
на рис. 3.1, а их распространение — на рис. 3.2. Mimulus puni-
Гл. 3. Географические расы
41
ceus— высокий прямостоячий кустарник с красными цветками,
сидящими на длинных цветоножках и имеющими короткие
трубки; он растет на прибрежных холмах и равнинах вблизи
океана. Mimulus longiflorus longiftorus — раскидистый кустар-
ник средней высоты с розовато-оранжевыми цветками проме-
жуточной длины; он встречается на выжженных солнцем пред-
горьях в чапаральной зоне удаленных от берега гор на юге Ка-
лифорнии. Mimulus longiftorus calycinus растет на каменистых
участках этих гор на больших высотах и в районах, гранича-
щих с пустыней Мохдве. Это — небольшое, низкорослое, разрас-
тающееся в стороны растение; цветки у него желтые, с длинной
трубкой и короткой цветоножкой. Интерградация наблюдается
между М. I. longiftorus и М. I. calycinus на промежуточных вы-
сотах на удаленных от моря горах, а также между М. I. longif-
lorus и М. puniceus в предгорьях прибрежной полосы [54].
V3’2' Географическое распространение группы Mimulus longiftorus в юж-
ной Калифорнии (по [54], с изменениями).
42
Часть первая. Популяции и расы
Расстояние, км
Рис. 3 3 Частота растений с красными, оранжевыми или желтыми цветками
в популяциях группы Mimulus longiflorus на трансекте, проходящей по юго-
западному склону гор Сан-Габриел, Калифорния (по [54], с изменениями).
Рассмотрим изменчивость окраски цветков у этих трех групп
по трансекте, проходящей по юго-западному склону гор Сан-
Габриел (рис. 3.3). Хотя, как отмечалось выше, для каждого
из этих таксонов характерна определенная нормальная окрас-
ка цветков, многие популяции, в том числе некоторые из нахо-
дящихся на данной трансекте, полиморфны по этому признаку.
Популяции, приуроченные к той части трансекты через горы
Сан-Габриел, которая соответствует предгорьям, т. е. обитаю-
щие на сравнительно небольшой высоте, состоят главным обра-
зом из экземпляров с оранжевыми цветками; эта окраска ти-
пична для М. I. longiflorus. Но в этих популяциях встречаются
экземпляры с красными цветками (типичными для М. puniceus),
а также экземпляры с желтыми цветками (типичными для
М. I. calycinus) (рис. 3.3). Чуть выше предгорий растения с
красными цветками исчезают. На средних высотах возрастает
частота растений с желтыми цветками, а на больших высотах
они вытесняют форму с оранжевыми цветками (рис. 3.3). Эту
интерградацию между расами по окраске цветков "можно пред-
ставить как сдвиг сбалансированного полиморфизма по геогра-
фической трансекте.
Один из количественных признаков цветка у этих видов так-
же подвержен внутрипопуляционной изменчивости, причем
скользящие средние лежат на той же трансекте (рис. 3.4).
Гл. 3. Географические расы
43
В этом случае сдвиг происходит в определенной зоне трансекты
на средних высотах. Ряд популяций типичных М. longiftorus
longiftorus на небольших высотах, т. е. в предгорьях, почти од-
нороден по этому количественному признаку; такая же одно-
родность наблюдается у ряда популяций М. I. calycinus в раз-
ных местах на больших высотах (рис. 3.4). По существу такие
же ступенчатые сдвиги были обнаружены при оценке различ-
ных других признаков цветка и вегетативных органов у этих
популяций, а затем все эти признаки были выражены в значе-
ниях гибридного индекса [54].
Между М. longiftorus longiftorus и М. puniceus наблюдается
также интергр^дация по различным трансектам, идущим от
удаленных от моря предгорий к прибрежным холмам и равни-
нам [54]. Если ограничиться этими данными, то М. puniceus
Айдилвайлд
Беннинг
Толл-Кэмп, 1955
Толл-Кэмп, 1960
Уотермен
Са н-Антон ио-Фоллз
Сьерра
Вершина г Потейто
Средняя часть г Потейто
Гера Потейто
Обнажение Томпсона ------
Томпсон-Сейдж —
Литтл-Дальтон, 1955 ------
Литтл-Дальтон, 1960 —«|
-30 -20 -10 О Ю 20 30
Относительная длина трубки венчика и цветоножки
Рис. 3.4. Внутри- и межпопуляционная изменчивость по одному из количе-
ственных признаков цветка у группы Mimulus longiftorus на двух трансектах,
проходящих по удаленным от побережья горам в южной Калифорнии (по
[54], с изменениями).
Оценивалась разница между длиной трубки венчика и длиной цветоножки
(в мм). Для каждой популяции представлены в графической форме среднее
значение, стандартное отклонение и диапазон изменчивости.
44 Часть первая. Популяции, и расы
можно рассматривать как одну из географических рас М. longi-
florus. Во взаимоотношениях между этими популяционными
системами имеется, однако, ряд интересных осложняющих мо-
ментов, которые мы рассмотрим в одной из последующих глав.
Дизъюнктивные географические расы
Непрерывная географическая изменчивость наблюдается в тех
случаях, когда сама популяционная система непрерывна или
почти непрерывна и достаточно велика. А это возможно при
условии пространственной непрерывности или полунепрерывно-
сти ее местообитания. Противоположная ситуация наблюдается
в тех случаях, когда местообитание данного вида разбито на
относительно небольшие области, распределенные в пространст-
ве с перерывами (например, ряд изолированных горных вершин
в случае какого-либо альпийского вида). В таких случаях из-
менчивость носит совершенно иной характер.
Популяционная система такого вида относится к колониаль-
ному типу, т. е. состоит из изолированных или полуизолирован-
ных популяций, занимающих обособленные местообитания.
Каждая изолированная колония становится специфически адап-
тированной к условиям своей локальной среды. Отдельные ко-
лонии, обитающие в различных средах, дивергируют. Всему это-
му способствует отбор в сочетании с пониженным обменом ге-
нами между колониями. И если величина колонии постоянно
или периодически бывает достаточно мала для того, чтобы
дрейф генов мог оказаться эффективным фактором,-то эти коло-
нии будут также приближаться к мономорфизму.
Вследствие этого характер изменчивости какого-либо вида,
населяющего экологические островки, резко отличается от ха-
рактера изменчивости вида с обширной и непрерывной популя-
ционной системой. Географическая изменчивость такого вида
прерывистая, географические расы разобщены, и в зависимости
от величины популяции степень полиморфизма может сильно
понизиться. И если в большой непрерывной популяции какого-
либо вида границы между географическими расами проводятся
более или менее произвольно, то у видов с популяционной сис-
темой островного типа географические расы дискретны и раз-
граничены не произвольно.
Эти противоположные типы географической изменчивости
можно наблюдать у двух видов Gilia — и. latiflora и G. leplant*
ha (Polemoniaceae). Указанные виды принадлежат к одной и
той же секции Arachnion данного рода; оба они однолетние,
оба диплоидные (2п=18), размножаются преимущественно пу-
тем перекрестного опыления и произрастают в общем в одной
и той же области в южной части Калифорнии. Оба вида поли-
Гл. 3. Географические расы
45
типические, с 4-5 географическими расами, которые рассматрива-
ются систематиками как подвиды. Как показали эксперимен-
тальные исследования [299], конспецифичные расы интерфер-
тильны у обоих видов. Однако на этом их сходство кончается.
Gilia latiflora встречается на песчаных равнинах в пустыне
Мохаве и примыкающих к ней долинах. Это местообитание не-
прерывно, как и местообитание G. latiflora. Географические
расы этого вида в большинстве случаев интерградируют на об-
ширных пространствах, и границы между ними устанавливают-
ся в значительной мере произвольно [300].
Gilia leptantha произрастает на открытых песчаных полянах
в сосновых лесах на горах южной Калифорнии. Ее местооби-
тания разбросаны по ряду изолированных горных хребтов. Та-
кие системы изолированных популяций образуют разобщенные
географические расы, сходные по фенотипическим признакам,
но разделенные хорошо выраженной прерывистостью в характе-
ре изменчивости [300].
Экологические расы
Географические расы аллопатричны, они занимают различные
географические области. Пространственные взаимоотношения
между расами могут также носить характер смежной симпат-
рии раса А и раса 5 занимают разные местообитания в одной
и той же области, скрещиваются между собой и интерградируют
в многочисленных зонах контакта, но каждая из них в своем
собственном местообитании сохраняет отличительные расовые
признаки.
Такой тип расовой изменчивости создается в тех областях,
где два или несколько местообитаний, подходящих для предста-
вителей рдпого и того же вида, распределены в пространстве в
виде пятен или в шахматном порядке. Примером служит саван-
на, в которой участки леса чередуются с участками лугов. Че-
редующиеся участки с разными типами почвы или с разными
типами материнской породы также могут быть заняты различ-
ными экологическими расами; то же самое наблюдается в го-
ристой местности па соседних горных склонах, обращенных
в разные стороны
Gilia achilleifolia (Polemoniaceae) встречается в одной из
частей Береговых хребтов (Калифорния), в саванне с участка-
ми, виргинского дуба. Популяции этого вида отличаются силь-
ной изменчивостью. Однако его различные локальные варианты
можно объединить в две большие группы или экологические
расы, известные в систематике под названиями G. a. achilleifo-
lia и G a multicaulis.
46 Часть первая. Популяции и расы
Gilia achilleifolia achiUeifolia — более обычная и широко
распространенная форма — встречается на открытых солнеч-
ных полянах. Ее крупные сине-фиолетовые цветки собраны в
головки и опыляются пчелами. G. a. multicaulis растет на полу-
затененных участках редколесья в тех же областях. У нее мел-
кие бледно окрашенные и самоопыляющиеся цветки. Эти две
экологические расы при экспериментальных скрещиваниях дают
плодовитых гибридов (во всяком случае, в некоторых сочета-
ниях) и интерградируют в природе в тех местах, где открытые
поля соприкасаются с участками леса [273].
По всей области обитания Gilia achilleifolia лесные и от-
крытые полевые местообитания чередуются в шахматном поряд-
Рис. 3.5. Географическое распространение двух экологических рас Gilia
achilleifolia [273].
Гл 3 Географические расы
47
ке. Это отражено и в характере распространения двух эколо-
гических рас данного вида (рис. 3 5).
Конспецифичные высотные расы возникают у видов, насе-
ляющих гористые области. Будут ли эти высотные расы по
своим пространственным взаимоотношениям экологическими
или географическими расами, зависит от рельефа местности.
Сплошные горные массивы с относительно пологими склонами
благоприятны для развития географических рас благодаря на-
личию обширных непрерывных пространств на разных высо-
тах. В отличие от этого в пересеченных горных массивах с
крутыми склонами те же высотные зоны представляют собой
ряд обособленных мелких пятен, нередко смежно-симпатричс-
ского типа, что благоприятствует развитию экологических рас.
Разумеется, между этими двумя основными типами рельефа су-
ществует ряд промежуточных типов.
Аналогичным образом эдафические расы могут представ-
лять собой экологические расы или мелкие локальные расы или,
наконец, нечто промежуточное в зависимости от величины тер-
ритории, занимаемой данной расой, и от ее характера.
Факторы отбора, участвующие в формировании
рас
Много лет назад Турессон [840] установил адаптивную природу
многих расовых признаков, наблюдаемых у растений. Его выво-
ды были основаны на трех группах данных. Во-первых, отличи-
тельные расовые признаки часто представляют собой адаптации
к условиям среды, преобладающим в местообитании данной ра-
сы Так, раса Atriplex litorale, обитающая на защищенном от
ветров южном побережье Швеции, представлена высокими пря-
мостоячими растениями, тогда как раса, населяющая открытое
ветрам западное побережье, — низкорослая и разрастающаяся
вширь [840]. Во-вторых, параллельные расовые различия наблю-
даются у других видов, встречающихся в тех же местообита-
ниях, например у Atriplex sarcophyllutn и А. ргаесох. И нако-
нец, в-третьих, у одной расы A litorale при выращивании ее в
среде, типичной для другой расы, в результате фенотипической
модификации развиваются признаки этой другой расы [840,
Среда, в которой обитает то или иное растение, характери-
зуется рядом аспектов, и все эти аспекты влияют на формиро-
вание расовых признаков. Наибольшее внимание уделялось
такому важному аспекту, как климат. Преобладающее большин-
ство экотипов, изучавшихся Турессоном [840, 841, 843] у Atri-
plex, Geum, Caltha, Achillea и многих других родов, относятся
именно к климатическим экотипам. Классические исследования,
48
Часть первая. Популяции и расы
проведенные Клаузеном, Кеком и Хиси [101, 103] на Potentilla
glandulosa, Achillea millefolium и других видах, также касаются
климатических рас.
Селективное воздействие климата ясно выступает в расовых
различиях по срокам цветения. В северных районах Швеции
вегетационный период короче, чем в южных. В соответствии с
этими климатическими различиями северные линии Fragaria
vesca, Campanula rotundifolia, Geum rivale и Qaltha palustris
зацветают раньше, чем южные линии тех же видов при совмест-
ном выращивании на однородных экспериментальных участках
[840, 843]. Подобным же образом расы Potentilla glandulosa и
комплекса Achillea millefolium из альпийской зоны гор Сьерра-
Невада в Калифорнии зацветают на экспериментальных участ-
ках раньше, чем расы, обитающие в тех же горах на средних
высотах [101, 103].
В формировании рас у некоторых видов явно участвуют эда-
фические факторы. Во многих случаях образование эндемичных
рас (и видов) коррелирует с отложениями гипса или выходами
известняка [847]. В гл. 2 наряду с другими примерами упомина-
лись эдафические расы Agrostis tenuis на почвах, содержащих
свинец, и на обычных почвах.
В некоторых случаях оказалось возможным продемонстри-
ровать существование зависящих от эдафических условий расо-
вых различий внутри одной климатической расы. Популяции
Achillea borealis, Gilia capitata capitata и Streptanlhus glandu-
losus, состоящие каждая из одной климатической расы, встре-
чаются в Калифорнии как на серпентинитовых, так и-на несер-
пентинитовых почвах. Как показал Круккеберг <[446], между по-
пуляциями, обитающими на тех и других почвах, имеются ра-
совые различия в отношении физиологической выносливости к
серпентинитовым почвам. Эдафическая расовая дифференциа-
ция у многих видов накладывается на климатическую расовую
дифференциацию [446, 452].
Синская [733] описала у Brassica nigra биотические расы.
В Малой Азии В. nigra встречается как в естественных усло-
виях, так и в посевах В. campestris. Эти две расы В. nigra раз-
личаются по комбинации вегетативных признаков, особенностям
цветков и семян. Более специализирована та раса, распростра-
нение которой ограничивается полями, засеянными В. campest-
ris; ее отличительные признаки дают ей возможность расти
среди растений В. campestris, а семенам — вызревать и распро-
страняться вместе с семенами последней [733]. Подобным же
образом некоторые расы Camelina sativa (Cruciferae) относятся
к неспециализированным сорнякам (С. s. sativa), тогда как
другие расы (С. s. linicola) — к специализированным сорнякам
льняных полей [780, 824].
Гл. 3. Географические расы
49>
Расовая дифференциация в отношении животных-опылите-
лей обнаружена у нескольких видов Polcmoniceae [301]. Раз-
личия в строении цветка между разными расами Gilia splendens
коррелируют с опылением их преимущественно жужжалами
(Bombylius), Eulonchus (Cyrtidae) с их длинными хоботками и>
колибри. У Eriastrum densifoliutn и Gilia achilleifolia есть расы
с широким зевом венчика, произрастающие в Калифорнии вдоль
береговой линии и опыляемые крупными видами пчел, а есть
расы с маленьким зевом, опыляемые мелкими видами пчел и
произрастающие на удаленных от моря горных хребтах. К слож-
ному переплетению расовой дифференциации могут добавиться
у многих видов растений расы, различия между которыми свя-
заны с опылением [301].
Количественных данных о скорости образования рас немно-
го. Очень интересны в этом плане исследования по расам Mimu-
lus guttatus (Scrophulariaceae) в бассейне озера Бонневиль в-
шт. Юта [860]. Бассейн этого древнего озера в течение плейсто-
ценового и голоценового периодов из территории, покрытой во-
дой и льдом, превратился в сушу, причем время этих измене-
ний можно установить.
Широко распространенный Mitnulus guttatus заселил многие
из обнажившихся при этом участков суши и образовал дивер-
гентные расы. Возраст этих популяций можно определить на ос-
новании известных возрастов их местообитаний. Таким образом
можно установить, что за 4000—14 000 лет имел место средний
уровень расовой’ дифференциации в отношении количественных
признаков и скрещиваемости [507, 860]. Это по осторожной оцен-
ке; на самом деле эволюционные изменения заняли, возможно»,
лишь часть этого времени.
4^45
Часть вторая
Природа видов
4 Биологический вид
Биологический вид — основная единица организации бипарен-
тальных живых существ, представляющая собой систему скре-
щивающихся популяций, репродуктивно изолированную от дру-
гих таких систем. Иными словами, это общая сумма скрещи-
вающихся между собой особей, а следовательно — самая обшир-
ная единица нормального бипарентального размножения. К био-
логической концепции вида приходят различными путями.
Первоначальная концепция вида возникла на основе прямых
наблюдений в природе. Первым естествоиспытателям, так же
как и их современным последователям, занимающимся популя-
ционной биологией, было очевидно, что живые организмы не об-
разуют континуум, а распадаются на более или менее дискрет-
ные скопления скрещивающихся между собой особей. Напри-
мер, члены семейства кошачьих делятся на такие дискретные
группы, как львы, тигры, рыси, домашние кошки и т. п. Каждая
группа размножается в чистоте, или, как выражались первые
естествоиспытатели, порождает себе подобных, а гибридов меж-
ду группами либо не существует вовсе, либо они столь редки,
что заслуживают особого внимания. Эта прерывистая картина
взаимоотношений и изменчивости наблюдается во многих се-
мействах животных и среди некоторых высших растений. Обо-
собленные размножающиеся внутри себя группы и есть виды.
Что касается видов в царстве растений, то лучше всего обра-
титься к тому, что писали об этом Чезальпино (Cesalpino), в
De plantis libri (1583), Джон Рэй (Ray) в Historia plantarum
(1686), а также Линней в Critica botanica [512] и Philosophia
botanica (1751; ,[513]).
Чезальпино утверждал, имея в виду виды, что подобное
всегда порождает подобное (см. [702]). Джон Рэй пошел даль-
ше, утверждая, что растения, выросшие из одного семени и
вновь продуцирующие себе подобные через семена, принадле-
жат к одному и тому же виду [702]. «Не существует более на-
дежного критерия вида, — писал он, — чем то, что он размно-
Гл. 4. Биологический вид
51
жается в чистоте, развиваясь из семени и не выходя за прису-
щие ему пределы» (см. [140]).
Линней продолжил эти высказывания, подчеркнув различия
между видами и разновидностями. «Творец Природы, создавая
виды, наложил на свои создания вечный закон плодиться и
размножаться, воспроизводя себе подобных. Правда, во многих
случаях он наделил их правом на некоторые отклонения во
внешнем виде, но не на превращение одного вида в другой»
(Critica botanica; ср. [653]). Совсем иначе обстоит дело с раз-
новидностями. «Существует столько разновидностей, сколько
существует растений, развившихся из семян данного вида»
(Philosophia botanica; ср. [653]; см. также [419])
За недостатком места, мы не можем сколько-нибудь подроб-
но изложить здесь дальнейшую историю концепции вида (см.
[245, 280, 568]). Достаточно сказать, что в XIX и в начале XX вв.
многие естествоиспытатели высказывали соображения, предвос-
хитившие биологическую концепцию вида. Современная биоло-
гическая концепция вида была сформулирована Реншем, Дю-
Рие, Майром, Добржанским и другими в период примерно с
1929 по 1957 г. Соответственно сами скрещивающиеся группы
получили название биологических видов.
Если биологические виды представляют собой обособленные
в репродуктивном отношении группы, то очевидно, что между
ними должны существовать барьеры, препятствующие гибриди-
зации; обнаружение таких барьеров послужило бы вторым до-
водом в пользу концепции биологического вида. Оказалось, что
эти постулированные барьеры, или репродуктивные изолирую-
щие механизмы, действительно существуют. Стерильность мула
вошла в легенды, и первые селекционеры иногда называли по-
лучаемые ими стерильные гибриды растительными мулами.
Бюффоп в 1749 г., а вслед за ним Кант в 1775 г. п многие дру-
гие ученые использовали стерильность потомков в качестве глав-
ного объективного критерия для разграничения отдельных ви-
дов. Однако все обстоит не так просто.
Скрещивания между биологическими видами предотвраща-
ются разнообразными изолирующими механизмами. Некоторые
пары видов действительно дают стерильные гибриды, однако от
других сочетаний видов можно искусственным путем получить
плодовитые гибриды, хотя в природе эти виды обычно не скре-
щиваются друг с другом вследствие существования различных
внешних барьеров, таких, как отвергание партнера чужого вида
или механические преграды, делающие скрещивание невозмож-
ным.
4*
Рис. 4.2 Gilia capitata (Polemoniaceae).
Изображенное здесь растение принад-
лежит к подвиду G с capitata.
Гл 4 Биологический вид
53
Пример
Gilia capitata и G. tricolor—родственные виды, принадлежащие
к одной и той же секции рода Gilia (Polemoniaceae). Это одно-
летние травянистые растения с диплоидным набором хромосом
(2п=18), размножающиеся преимущественно в результате пере-
крестного опыления.
Gilia tricolor произрастает в долинах и предгорьях централь-
ной и северной Калифорнии. G. capitata встречается в тех же
районах, но, кроме того, заходит дальше на север, юг и запад,
доходя до береговой линии (рис. 4.1). Во многих районах от-
дельные растения обоих видов растут в непосредственной бли-
зости друг от друга. Эти два вида симпатричны.
Морфологические признаки типичных представителей этих
двух видов изображены на рис. 4.2 и 4.3. Растения Gilia capita-
Рис. 4.3. Gilia tricolor
54
Часть вторая. Природа видов
ta крупные и высокие, a G. tricolor—относительно мелкие.
У G. capitata соцветие образует плотную головку, а у G. trico-
lor оно рыхлое. У G. capitata на чашелистиках имеются узкие
средние жилки, а у G. tricolor—широкие зеленые полосы. Эти
два вида различаются также по форме венчика. Кроме того,,
у G. capitata венчик одноцветный, сине-фиолетовый, а у G. tri-
color он трехцветный, с пурпурными пятнами на сине-фиолето-
вом фоне и оранжевой трубкой.
Ни тот ни другой вид нельзя, однако, назвать однородным.
G. capitata подвержен значительной географической изменчиво-
сти. Признаки цветков, коробочек и семян варьируют у разных
рас по трансекте, идущей от прибрежных песчаных дюн до гор,,
расположенных дальше от берега, в области, окружающей за-
лив Сан-Франциско (рис. 4.4). Между этими и другими расами
G. capitata наблюдается полная интерградация. Географическая;
изменчивость наблюдается и у G. tricolor, но у этого вида она
менее выражена. При этом как у G. capitata, так и у G. tricolor
Рис. 4.4. Цветки, коробочки и семена трех рас Gilia capitata, произрастающих
на трансекте, идущей от прибрежной зоны залива Сан-Франциско (Кали-
форния) к горам, расположенным на север от него.
А — Gilia capitata chamissonis. Б — G.c. tomentosa, В — G.c, capitata.
Гл. 4. Биологический вид
55
географическая изменчивость не выходит за пределы данного
вида. Промежуточные формы между этими двумя видами неиз-
вестны; ни одного гибрида между ними обнаружено не было.
Внутри каждого вида Gilia и между ними были проведены
экспериментальные скрещивания. Скрещивали семь линий Gi-
lia capitata, принадлежащих к пяти географическим расам. Эти
расы скрещиваются друг с другом во всех сочетаниях, причем
в большинстве сочетаний скрещиваются очень легко. Межрасо-
вые гибриды Fi обладают высокой или частичной плодови-
тостью, образуя от 50 до 90% полноценной пыльцы и обильные
семена. Развивающиеся из них растения F2 обычно жизнеспо-
собны, но небольшая доля растений обладает пониженной жиз-
неспособностью [266, 268]. Северная и южная расы G. tricolor
также оказались интерфертильными [269].
В отличие от этого многократные попытки провести скрещи-
вания между Gilia capitata и G. tricolor всегда оказывались
безуспешными. Перекрестное опыление между этими видами
приводит к образованию недоразвитых семян, но не дает ни
нормальных семян, ни гибридов Fi. Кроме того, установлено,
что О. tricolor не скрещивается ни с одним из других родствен-
ных видов Gilia G capitata с трудом удается искусственно скре-
стить с каким-нибудь другим близким видом, но получающиеся
при этом гибриды высоко стерильны [269, 275].
Таким образом, G tricolor и G capitata являются самостоя-
тельными биологическими видами. Между представителями ви-
да G. tricolor происходит половое размножение, приводящее к
образованию интерградирующих вариаций. Подобное же скре-
щивание с аналогичными результатами происходит между пред-
ставителями G. capitata. Однако G. tricolor и G. capitata не мо-
гут скрещиваться и действительно никогда не скрещиваются
между собой, так как этому препятствуют очень эффективные
репродуктивные изолирующие механизмы.
Биологический вид как естественное следствие
' полового размножения в биотическом
сообществе
К концепции биологического вида можно прийти и третьим
путем. Можно показать, что половой процесс, происходящий
в неоднородной среде, неизбежно должен сопровождаться орга-
низацией скрещивающихся внутри себя популяций в обособлен-
ные биологические виды Предпосылки к подобным взглядам
излагались Добржанским [165] и Кэйном [79]. Кэйн писал:
56
Часть вторая. Природа видов
«Достаточно лишь представить себе последствия неограниченной гибридиза-
ции между всеми ныне живущими животными, чтобы оценить крайнюю важ-
ность процесса видообразования. Если бы все межвидовые барьеры вдруг
каким-то чудом были устранены, то это привело бы к ужасающему столпо-
творению гибридов со всевозможными сочетаниями признаков. Ни один ин-
дивидуум не был бы должным образом адаптирован к тому или иному об-
разу жизни, а многие признаки каждого из них могли бы оказаться адапти-
рованными для почти несовместимых целей»
А Добржанский [165] утверждал, что «продолжение жизни
возможно лишь в том случае, если генные комплексы, согласо-
ванность которых проверена естественным отбором, будут пре-
дохранены от разрушения, неизбежного при неограниченной
гибридизации». Мы построим наши дальнейшие рассуждения,
использовав эти высказывания в качестве основы.
Любая пригодная для обитания локальная область на по-
верхности земного шара обычно содержит разнообразные эко-
логические ниши, занимаемые различными симпатрическими
популяциями организмов. Каждая популяция адаптирована к
своей особой нише в биотическом сообществе благодаря нали-
чию у нее определенной комбинации физиологических и морфо-
логических признаков. Каждая комбинация признаков основана
в свою очередь на определенной комбинации генов.
Половое размножение представляет собой механизм, обус-
ловливающий рекомбинацию генов. Рекомбинацию генов нель-
зя считать абсолютным благом: в одних случаях она приводит
к благоприятным результатам, а в других — к неблагоприятным.
Адаптивные свойства различных возможных комбинаций генов
широко варьируют в зависимости от входящих в них генов и
взаимодействий между ними.
Особи, принадлежащие к данной скрещивающейся популя-
ции, могут варьировать по нескольким наследственным призна-
кам, но тем не менее они способны скрещиваться друг с другом,
давая жизнеспособное плодовитое потомство. Такая популяция
полиморфна по ряду коадаптированных генов. Поясним это на
простом примере. Ген А представлен в популяции двумя алле-
лями, Аг и А2, а другой ген В точно так же представлен двумя
формами, Bi и В2. Девять диплоидных генотипов, которые мо-
гут получиться в результате полового процесса из этих двух
полиморфных генов (AiAB^, А^А^В* ..., А2А2В2В2), обла-
дают индивидуальными различиями; тем не менее все они —
нормальные представители своей популяции, адаптированные к
жизни и более или менее успешному размножению в экологиче-
ской зоне, занимаемой их популяцией. Кроме того, хорошие
адаптивные признаки нескольких рекомбинационных типов —
это не просто счастливое совпадение, а результат естественного
отбора, который через предшествующие поколения оказывал
Гл. 4. Биологический вид
57
свое действие па данную скрещивающуюся популяцию, сохра-
няя благоприятные аллели с хорошими комбинационными спо-
собностями.
Другая популяция, занимающая другую экологическую ни-
шу в том же сообществе, обладает, подобно первой популяции,
определенной адаптивной комбинацией признаков, которая под-
вержена изменчивости в известных пределах Эта популяция,
однако, полиморфна по другому набору неадаптированных ге-
нов. Допустим, что она содержит аллели Л8 и Л9 и В8 и В9, ко-
торые, комбинируясь друг с другом, дают девять диплоидных
генотипов А8А8В8В8, А8А8В8В9, , AgAgBgB^ Все эти генотипы,
несмотря на индивидуальные различия, приспособлены к жизни
в нише, занимаемой их популяцией
Последствия скрещивания между членами этих двух популя-
ций весьма различны, если рассматривать их в плане адаптиро-
вапности потомков Рассмотрим, например, девять типов потом-
ков F2, получающихся в результате скрещивания генотипов
A\AiB}B\ и А8А8В&В&. Два родительских типа, вновь появляю-
щиеся в поколении F2, несомненно, жизнеспособны; жизнеспо-
собным может оказаться и тип Fi. Остальные шесть генотипов,
представляющие собой неиспытанные рекомбинации родитель-
ских генов, могут оказаться плохо адаптированными как к раз-
личным средам одной и другой родительских особей, так и к
любой другой доступной им среде Если гены А и В не сцепле-
ны между собой, то можно ожидать, что такие жизнеспособные
или субвитальные рекомбинационные типы составят 10 из 16 зи-
гот F2.
Согласно правилам‘Менделя, при других родительских парах
из этих двух популяций, например AiAiB2B2XА8А8В9В9 и т. д.,
доля рекомбинантов в F2, не относящихся ни к P-типу, ни к
Ft-типу, будет такой же, а именно 62,5%. Поэтому в целом по-
томки, получающиеся в результате гибридизации между двумя
популяциями, различающимися по одной простой адаптивной
комбинации генов, —это преимущественно рекомбинанты с по-
ниженной приспособленностью.
Доля субвитальных рекомбинантов быстро повышается с
увеличением числа генных различий между родительскими по-
пуляциями. Допустим, что две популяции содержат различные
аллели пяти песцепленных между собой генов, от А до Е, опре-
деляющих различные адаптивные типы в каждой популяции
В таком случае скрещивание между A}Ab,.EiE\ и А8А8.. Е8Е8
Даст зиготы F2, из которых только 3,3% будут генотипически
сходны с родительскими особями или с особями F]. Остальные
96,7% зигот F2 составят рекомбинанты, жизнеспособность ко-
торых в большей или меньшей степени понижена Если роди-
тельские популяции различаются по адаптивной комбинации ге-
58
Часть вторая. Природа видов
нов, которую образуют десять независимых генов, то такие суб-
витальные рекомбинанты составят 99,9% всех зигот F2.
Иными словами, 966797 из каждого миллиона зигот F2, воз-
никших в результате гибридизации между популяциями, разли-
чающимися по адаптивной комбинации генов, состоящей из пя-
ти несцепленных между собой генов, будут обладать плохо адап-
тированной генетической конституцией. А на каждый миллион
зигот Fs, образовавшихся в результате гибридизации между по-
пуляциями, которые различаются по адаптивной комбинации
признаков, определяемой десятью независимыми генами, 999 022,
по всей вероятности, не смогут выжить ни в какой из доступных
им сред.
Конечно, реальные популяции будут так или, иначе откло-
няться от этих упрощенных моделей. Некоторые факторы, на-
пример сцепление, несколько уменьшают генетический груз,
создаваемый гибридизацией. Другие факторы, такие, как слож-
ность генетических различий между симпатрическими видами,
напротив, увеличивают этот груз, причем последние факторы,
возможно, перевешивают первые. Таким образом, вытекающие
из наших численных примеров оценки потенциального груза,
создаваемого гибридизацией между физиологически дифферен-
цированными симпатрическими популяциями, вероятно, не сле-
дует считать преувеличенными.
Если при скрещиваниях между дифференцированными сим-
патрическими популяциями наблюдаются огромные потери ре-
продуктивного потенциала, то скрещивания внутри популяции
обычно бывают благоприятными. Особи, скрещивающиеся с
другими членами своей популяции, обычно оставляют гораздо
больше потомков, чем особи, скрещивающиеся с членами чужих
популяций. В таких условиях любые генетически детерминиро-
ванные отвергания членов чужих популяций в качестве брачных
партнеров или любые барьеры, препятствующие скрещиванию,
будут подхвачены отбором и будут распространяться в каждой
симпатрической популяции.
Таким образом, мы приходим к выводу, что половое размно-
жение, если оно выходит за пределы отдельных популяций дан-
ного биотического сообщества, приводит к распаду адаптивных
комбинаций генов внутри каждой популяции. Угроза продолже-
нию существования таких симпатрических популяций тем боль-
ше, чем более свободно они могут скрещиваться друг с другом.
И наоборот, те биотические сообщества, которые состоят из
репродуктивно изолированных скрещивающихся внутри себя
популяций, или, иными словами, биологических видов, остаются
стабильными.
Гл. 4. Биологический вид
59
Концепция генофонда вида
Концепцию биологического вида можно выразить, связав ее с
представлением о генофонде. Концепция вида, основанная на
генофонде, хорошо сформулирована Добржанским [168].
«Мспделевская популяция — это репродуктивное сообщество размножающих-
ся половым путем и скрещивающихся между собой индивидуумов, облада-
ющих общим генофондом Биологический вид — это самая обширная и са-
мая емкая менделевская популяция. Надвидовые группировки, такие, как
подроды, роды и т п, не обладают общими генофондами, а поэтому лише-
ны биологической реальности, присущей менделевским популяциям Однако
виды дифференцированы на комплексы подчиненных менделевских популя-
ций, которые можно называть подвидами, расами или локальными популя-
циями»
Существенное возражение, выдвигаемое рядом авторов [195,
196, 471] против такого определения, состоит в том. что пере-
крестное оплодотворение и обмен генами происходят не на ви-
довом уровне, а внутри локальных популяций и субпопуляций.
Что касается самих этих фактов, то они верны, и мы их будем
обсуждать в следующем разделе, однако при их истолковании
упускается из виду самое главное.
Имеющиеся в данном генофонде гены доступны всем парам
способных к скрещиванию индивидуумов. Образуются ли все та-
кие пары в действительности или нет, значения не имеет. Кон-
цепция генофонда содержит некий элемент потенциальности
Две особи, находящиеся на противоположных сторонах одной
и той же обширной видовой популяции, могут объединить свои
гены; их потомки черпают гены из двух удаленных одна от дру-
гой частей обширного генофонда. Однако особь, принадлежащая
к другому, репродуктивно изолированному виду, не может чер-
пать гены из генофонда первого вида, даже если она живет с
ним на одной территории Следовательно, мы имеем полное
право представлять себе биологические виды как популяции, об-
ладающие обширными генофондами, которые доступны для всех
входящих в их состав особей.
Причины и следствия, вытекающие из концепции
Какова основа генотипического и фенотипического сходства
между особями, принадлежащими к одному и тому же биологи-
ческому виду? Согласно традиционной точке зрения, поток ге-
нов обладает значительным гомогенизирующим и интегрирую-
щим действием в обширных и свободно скрещивающихся попу-
ляционных системах на уровне видов и региональных подразде-
лений видов. Такой точки зрения придерживался еще Дарвин
[152]. «Перекрестное оплодотворение играет очень важную роль
в природе, обеспечивая сохранение особей, принадлежащих к
60
Часть вторая. Природа видов
одному и тому же виду или к одной и той же разновидности,
в чистоте и их единообразие».
В наши дни эти же взгляды усиленно пропагандирует Майр
[565, 569—571]. Приведем некоторые цитаты из его работ.
[Новогвинейские популяции Tanysiptera galatea находятся] в центре не-
прерывного потока генов, идущего в обоих направлениях через весь этот
островной материк [564].
Устойчивый и интенсивный приток генов, обусловленный процессом об-
мена генами, служит главным фактором, обеспечивающим генетическую инте-
грированность между популяциями данного вида [569].
У всех широко распространенных процветающих видов относительно по-
движных организмов, по-видимому, происходит достаточно интенсивный об-
мен генами, обеспечивающий высокую однородность генофондов всех локаль-
ных популяций [570].
«Отмечая поразительную морфологическую однородность большинства
широко распространенных видов с высокой численностью, я в... своей статье
(1954) особенно подчеркивал выравнивающее действие потока генов» [571].
Изложенная выше точка зрения была довольно широко рас-
пространена среди эволюционистов, включая и меня самого
[281, 293]. Однако за последнее время ряд исследователей вы-
сказались и против нее.
Эрлих и Рейвн [196] утверждают, что «не следует представ-
лять себе вид как эволюционную единицу, объединяемую свя-
зующими силами потока генов, [потому что] в природе поток
генов гораздо более ограничен, чем обычно принято считать...»
На это указывают и другие авторы [195, 197, 660, 753].
Эти авторы основывают свои возражения против традицион-
ной точки зрения на данных об относительно ограниченном по-
токе генов и зачастую небольших размерах соседств, о которых
мы говорили в гл. 2. Эта группа данных, несомненно, имеет
отношение к рассматриваемому вопросу.
Обратившись вновь к этой проблеме в 1980 г. [298], я при-
шел к выводу, что ни приверженцы традиционной точки зрения,
ни их нынешние противники не привели достаточно убедитель-
ных данных в пользу своих высказываний. Сторонники тради-
ционных взглядов сделали несколько правдоподобных, но до-
вольно смелых допущений, которые необходимо подкрепить
более твердыми данными. Традиционная точка зрения и в са-
мом деле уязвима.
В свою очередь в аргументации, приводимой критиками тра-
диционных взглядов, есть два слабых места. Во-первых, эти
критики рассматривают процесс расселения, ограничиваясь рам-
ками лишь одного поколения. Между тем, изучая поток генов в
масштабах географических рас и видов, необходимо рассматри-
вать более обширный промежуток времени, охватывающий мно-
го поколений. Во-вторых, цитированные выше авторы придают
слишком большое значение расселению на небольшие расстоя-
ния, которое, безусловно, представляет собой наиболее вероят-
Гл. 4. Биологический вид
6Г
ное течение событий; однако они преуменьшают роль событий,
сопровождающих расселение на далекие расстояния, которые
также могут иметь биологическое значение.
Как же обстоит дело в настоящее время? Приходится при-
знать, что окончательного решения пока нет. Имеющиеся дан-
ные можно истолковать по-разному. Я изложу здесь краткие
итоги проведенного мною недавно анализа этой проблемы [298].
Рассмотрим сначала изменение во времени расстояния, на
которое происходит расселение, для организмов с противопо-
ложными способностями к расселению. Это изменение зависит
от двух факторов, среднего расстояния, на которое происходит
расселение за 1 поколение и продолжительности генерации
Расстояние, на которое происходит расселение у Drosophila
pseudoobscura и ее близких родичей в Калифорнии, составляет
в среднем 361 м. В природе у этих мух появляется примерно по
7 поколений в год С учетом этих двух факторов можно счи-
тать обоснованной оценкой расселение на 2527 км за 1000 лет.
Это расстояние сопоставимо с длиной или шириной области рас-
пространения этого вида в западной части Северной Америки
Сравните эту величину с величиной расстояния, на которое
способно расселяться какое-либо многолетнее травянистое рас-
тение. Liatris aspera расселяется в среднем за одно поколение
па 3,5 м, а продолжительность генерации этого растения состав-
ляет 5 лет [481]. Соответственно кумулятивное расстояние, на
которое происходит расселение, составит всего 0,7 км за
1000 лет, или 7 км за 10000 лет. Это расстояние соответствует
лишь очень незначительной части ареала вида. Продолжитель-
ность существования вида может оказаться недостаточной для
его широкого расселения.
Приведенные выше основные параметры расселения типичны
для этих двух групп организмов; сходные оценки имеются для
других видов дрозофилы и для других многолетних травянистых
растений (см. [298]). Эти два организма представляют две
крайности: подвижные мухи с высокой способностью к расселе-
нию и прикрепленные к субстрату многолетние растения с низ-
кой способностью к расселению. В одном случае за период в
1000 или 10 000 лет может произойти расселение на большие
расстояния; в другом случае это, по-видимому, невозможно.
Хотя расселение представляет собой физическую основу по-
тока генов, эти процессы не идентичны, и их не следует смеши-
вать. Нас здесь интересует поток генов во времени или, точнее,
то, что мы называем генетически эффективным потоком генов
во времени. Это обмен генами между популяциями, различаю-
щимися по частоте аллелей, который происходит на протяжении
Двух или большего числа поколений и в котором участвуют три
или более популяции.
Ф2 Часть вторая. Природа видов
Между расселением во времени и генетически эффективным
потоком генов есть одно важное различие. В первом случае
суммарное расстояние, на которое происходит расселение, ли-
нейно возрастает со временем. Но во втором случае частота
иммигрирующих генов быстро снижается на каждом этапе и в
каждом поколении, пока в конце концов процесс генетически
эффективного обмена генами не Прекращается вовсе [298].
Рассмотрим гипотетический пример. Допустим, что популя-
ции I, II, III и IV распределены по трансекте, пересекающей
.ареал данного вида с запада на восток. В популяции I имеется
новый селективно нейтральный аллель Л2, частота которого
-равна 100%. Популяции II, III и IV содержат прежний аллель
Ai с частотой 100%. Обмен генами между соседними популяция-
ми происходит в обоих направлениях со скоростью т=0,10.
.Ъ таком случае частота (q) аллеля Az в этих четырех популя-
циях по прошествии трех поколений составит:
Популяции I
Популяция II
Популяция III
Популяция IV
<71=0,755
92=0,219
9з=0,025
94=0,001
Частота нового аллеля А2 резко снижается на каждом этапе
^короткого ступенчатого процесса иммиграции. На каком-то
этапе этого процесса аллель А2 становится настолько редким,
счто вероятность его включения в следующую выборку мигрантов
ничтожна. На этом этапе процесс генетически эффективного
обмена генами прекращается. Расселение как таковое продол-
жается, но генетически эффективный поток генов иссякает
*[298].
Если бы генетически эффективный обмен генами продолжал-
ся на протяжении очень многих этапов, то очевидно, что ему
»была бы необходима мощная поддержка со стороны отбора.
В таком случае нам следовало бы изменить свое допущение,
предположив, что аллель А2 селективно не нейтрален, а сох ра-
вняется отбором. При этом частота аллеля Az могла бы возрас-
тать на каждом этапе миграционного процесса, так что вероят-
ность его включения в следующую выборку мигрантов была бы
.достаточно велика. Но повышение частоты того или иного алле-
ля под действием отбора требует времени. На протяжении
гряда последовательных поколений генетически эффективный об-
мен генами, создаваемый совместным действием миграции и
-отбора, будет протекать гораздо медленнее, чем расселение как
таковое.
Гл 4. Биологический вид
6$
Рассмотрим вновь пример с дрозофилой в свете этих сооб-
ражений. Как мы уже видели, такой высоко подвижный орга-
низм с коротким временем генерации, как дрозофила, может
расселиться по области, соответствующей видовому ареалу, за
1000 лет; но для того чтобы генетически эффективный поток
генов распространился по той же области с учетом действия от-
бора на каждом этапе, вполне может потребоваться в 50 или
даже в 100 раз больше времени. Такие периоды времени при-
ближаются к историческому возрасту данного вида
Вплоть до этого момента все наши рассуждения свидетель-
ствуют в пользу антитрадициопной точки зрения. Существуют
серьезные сомнения относительно того, может ли поток генов
оказывав гомогенизирующее воздействие на внутривидовую из-
менчивость в широко распространенных видовых популяциях в
течение периодов времени порядка 10 000 лет. Эффективный
поток генов в областях, соответствующих видовому ареалу, стал-
кивается с затруднениями даже у высокоподвижных видов с
коротким временем генерации Но особенно остро стоит эта
проблема для прикрепленных к субстрату многолетних растений-
с длительным временем генерации и ограниченной способностью
к расселению.
Однако, прежде чем так легко отбросить традиционную точ-
ку зрения, следует обратить внимание на другую группу дан-
ных, свидетельствующих о том, что поток генов оказывает влия-
ние на характер видовой изменчивости. В популяционных систе-
мах с бипарентальным размножением, состоящих из множества-
географических рас, наблюдается большая однородность в пре-
делах обширных областей при непрерывной структуре популя-
ционной системы, чем при разорванной (см. гл. 3). Это разли-
чие коррелирует с наличием или отсутствием открытых кана-
лов для обмена генами. Кроме того, в бипарентальных популя-
ционных системах однородность внутривидовой изменчивости
гораздо выше, чем в унипарентальных (см. гл. 1 и 5). И снова
это различие коррелирует с наличием или отсутствием эффек-
тивного механизма для обмена генами.
Причины несоответствия между двумя группами данных не-
известны. Быть может, было бы полезно вновь рассмотреть дан-
ные по расселению. Методы, используемые при количественных
исследованиях расселения, применимы лишь на ограниченных
участках. Эти методы не позволяют выявить события, связан-
ные с расселением на большие расстояния, что, возможно, ведет
к недооценке средних расстояний, на которые происходит рас-
селение. Внесение в существующие оценки поправок, учитываю-
щих некоторое расселение на большие расстояния, могло бы
способствовать примирению этих двух групп фактических
данных.
>64 Часть вторая. Природа видов
Виды-двойники
Наиболее существенная черта биологических видов, как мы ви-
дели, связана с их взаимоотношениями при скрещивании. Спо-
собность особей успешно обмениваться гёнами, т. е. свободно
«скрещиваться между собой и производить плодовитое и жизне-
способное потомство, характеризует их как представителей
•одного и того же биологического вида, тогда как неспособность
к свободному и успешному обмену генами — признак обособ-
ленности биологических видов. Все остальное, в том числе и
морфологические различия, — это надстройка.
Родственные биологические виды действительно отличаются
друг от друга по внешним морфологическим признакам, равно
как и по лежащим в их основе физиологическим признакам, по-
скольку многие морфологические признаки входят в состав
-адаптивных комбинаций признаков соответствующих видов. Тра-
диционные таксономические методы классификации и определе-
ния основаны на этой обычной ассоциации внешних морфологи-
ческих признаков с физиологическими особенностями.
Но такая корреляция не всегда выдерживается. Известны
случаи, когда хорошие биологические виды морфологически аб-
солютно неразличимы. Такие виды получили название видов-
двойников.
Классическим примером видов-двойников служат Drosophi-
la pseudoobscura и D. persimilis. Прежде считалось, что эти мор-
фологически сходные мухи принадлежат к одному виду —
D. pseudoobscura-, однако в дальнейшем при некоторых «внутри-
видовых» скрещиваниях обнаружилось, что скрещиванию меж-
ду ними препятствует стерильность гибридов. Затем оказалось,
что эти интерстерильные группы образуют симпатрические по-
пуляции на обширных территориях в западной части Северной
Америки, не гибридизируя между собой, и в соответствии с этйм
•они были признаны отдельными видами.
После поисков, предпринятых рядом исследователей, были
в конце концов обнаружены небольшие морфологические раз-
личия в строении половых органов и крыльев у самцов, а также
некоторые поведенческие, физиологические и цитологические
различия (обзор см. [569]). Эти различия очень невелики, и их
трудно выявить обычными таксономическими методами.
Между тем мухи легко узнают друг друга. Когда самцов и
•самок Drosophila pseudoobscura и D persimilis помещают вмес-
те в популяционный ящик в лаборатории, скрещивания проис-
ходят исключительно или преимущественно между особями,
принадлежащими к одному и тому же виду; и лишь в редких
случаях в обычных лабораторных условиях самки и самцы раз-
ных видов копулируют друг с другом. Подобным же образом
Гл. 4. Биологический вид
65
в природе, где очень много возможностей для гибридизации,
особи, принадлежащие к разным видам, спариваются крайне
редко, и межвидовые гибриды неизвестны [170]. Очевидно, дро-
зофилы умеют различать виды D pseudoobscura и D persimilis
независимо от того, умеют ли это делать систематики.
Виды-двойники существуют и в других комплексах видов
дрозофилы, и во многих других группах животных (обзор см.
[569]). Виды-двойники встречаются также во многих родах выс-
ших растений [279], причем у последних это явление часто со-
четается с полиплоидией.
Тетраплоидный вид с геномом ААВВ морфологически сходен
и может интерградировать с предковым диплоидным видом АА
по двум причинам. Во-первых, что самое главное, тетраплоид-
ный вид содержит Д-геном Во-вторых, тетраплоид, вероятно,
будет выщеплять некоторое число особей родительского типа А,
особенно если это сегментный аллотетраплоид. По тем же при-
чинам этот тетраплоид будет сходен со своим вторым диплоид-
ным родительским видом ВВ. Вследствие различий в строении
хромосом диплоидные виды АА и ВВ образуют при скрещива-
нии более или менее стерильных гибридов Гд, гибриды между
тетраплоидным видом ААВВ и любым из диплоидных роди-
тельских видов, будучи триплоидами, также стерильны. Таким
образом, популяция, которую по морфологическим признакам
можно было бы принять за непрерывно интерградирующую по-
пуляцию и которую систематик мог бы рассматривать как еди-
ный вид, представляет собой на самом деле группу из трех ин-
терстерильных видов-двойников [283].
В качестве иллюстрации приведем группу видов Gilia trans-
montana, которую тщательно проанализировала Дэй [158].
В эту группу входят мелкие однолетние травянистые растения,
встречающиеся в пустыне Мохаве и на прилегающих к ней тер-
риториях в западной части Северной /Америки Это преимущест-
венно автогамные растения, однако у них происходит некоторое,
но тем не менее биологически значимое перекрестное опыление
при помощи насекомых. Общий вид этих растений показан на
рис. 4.5 и 4.6.
Группа Gilia transmontana состоит не из трех, а из пяти род-
ственных диплоидных и тетраплопдных видов. Один из тетра-
плоидных видов — G. transmontana — происходит от двух дип-
лоидов—G. minor и G clokeyl, а другой тетраплоид—G. та-
Ног — от диплоидов G. minor и G aliquanta [158]. Таким обра-
зом, у обоих тетраплоидпых видов есть один общий диплоидный
предок (G minor). Все эти пять видов в ранних таксономиче-
ских работах рассматривались как разные формы одного вида.
Результаты опытов по искусственной гибридизации свиде-
тельствуют о том, что эти пять видов, сами по себе вполне
5—45
66
Часть вторая. Природа видов
плодовитые, высоко интерстерильны в любых комбинациях.
В природе они симпатричны и растут в самых различных соче-
таниях, не скрещиваясь между собой [158, 283, 300].
Дэй [158] выращивала и сравнивала ряд линий, представ'*
ляющих известный диапазон изменчивости каждого вида, в од-
нородных условиях среды в вегетационном домике. Она обнару-
жила, что эти виды различаются, не очень сильно, но различия
между ними постоянны и затрагивают почти все части расте-
ния (см. рис. 4.5—4.7). Так, Gilia aliquanta отличается своими
несколько более крупными цветками, но у остальных четырех
видов цветки одинаково мелкие (рис. 4.6). У G. minor семена
мельче, чем у других видов (рис. 4.7). Листья, окраска венчика,
коробочки и другие признаки характеризуются незначительны-
ми межвидовыми различиями.
Однако эти внешние фенотипические различия не дают воз-
можности проводить идентификацию во всех случаях. Вернем-
ся еще раз к величине семян. У Gilia minor вес семян колеблет-
Рис. 4.5. Форма роста у группы видов-двойников рода Gilia [158J.
Гл. 4. Биологический вид
Рис. 4 6 Цветки пяти видов-двойников Gilia, изображенных на рис. 4.5
1158].
А — G. clokeyi. Б — G. transmontana. В — G minor. Г — G. malior Д — G. all-
quanta
А £ В Г Д
Рис 4 7 Семена тех же пяти видов-двойников Gilia [158].
А — G. clokeyi. Б — G. transmontana. В —G. minor. Г — G. malior. Д—G. all-
quanta
ся от 10 до 20 мг, у С?, transmontana— от 20 до 30 мг, у G. clo-
keyi— от 30 до 85 мг. Эти три вида различаются по средней ве-
личине семян (рис. 4.7), но образуют непрерывный ряд, если
рассматривать диапазон изменчивости нескольких видов [158].
В общем Gilia transmontana заполняет морфологический
разрыв между своими диплоидными предками G. minor и
G. clokeyi, так что ясно выраженные различия между этими
тремя видами найти трудно. Аналогичным образом G. malior
интерградирует с G. minor и G. aliquanta. А поскольку и
G. transmontana, и G malior содержат геном G. minor, оба
5я
68 Часть вторая Природа видов
тетраплоидных вида имеют много обших морфологических
признаков.
Проблемы таксономической идентификации в группе Gilia
transmontana не имеют, однако, отношения к вопросу о биоло-
гическом составе этой группы. Gilia transmontana, G. malior,
G. minor, G. clokeyi и G. aliquanta образуют пять отдельных
скрещивающихся групп и представляют собой, таким образом,
хорошие биологические виды независимо от того, можно ли во
всех случаях идентифицировать их но внешним морфологиче-
ским признакам или пет.
5 Видовые единицы
у унипарентальных растений
Концепция биологического вида относится к бинарен гальным
организмам. Она не распространяется па упипарентальные ор-
ганизмы, не образующие скрещивающихся групп Между тем
унипарентальное размножение обычно у растений, а также в
разных группах, принадлежащих к другим царствам; поэтому
необходимо разработать параллельные или альтернативные
концепции вида, приложимые к единицам изменчивости в этих
унипарентальных группах.
Системы унипарентальных популяций
Унипарентальные организмы образуют популяции и обширные
системы популяций. Популяции состоят из генотипически сход-
ных особей. Генотипическое сходство обусловлено происхожде-
нием от общих предковых линий и усиливается под действием
стабилизирующего отбора в данной нише или данном местооби-
тании. Эти два фактора — общие предки и стабилизирующий
отбор — приводят к тому, что родственные особи в своей среде
определенным образом интегрируются, образуя популяции [574,
731, 732].
Итак, между унипарептальными и бипарентальными организ-
мами существуют как черты .сходства, так и различия в отно-
шении характера их популяций и популяционных систем.
И у унипарентальных, и у бипарентальпых организмов наблюда-
ются следующие процессы- 1) наследование одинаковых генов
от общих предков; 2) распространение определенных генов по
всей популяционной системе или виду в результате миграции
и естественного отбора; 3) подавление распространения этих
генов в другие виды, обусловленное их неспособностью мигри-
ровать в другую среду или воспроизводиться в ней. Под дейст-
вием всех этих процессов родственные особи, несущие одинако-
вые гены, объединяются в популяции, обитающие в одной и той
же среде, независимо от того, размножаются эти особи половым
путем или нет [574, 732].
Популяции организмов, размножающихся половым путем и
при участии двух родительских особей, связаны между собой
Двумя дополнительными процессами, отсутствующими в строго
унипарентальных группах. Один из этих процессов — скрещива-
ние между особями, облегчающее распространение генов в по-
пуляционной системе. Другой процесс — репродуктивная изоля-
ция, препятствующая распространению этих генов в другие ви-
ды [732].
70 Часть вторая. Природа видов
Степени интеграции
'Существование сил, интегрирующих популяции и присущих
•только бипарентальным организмам, заставляет думать, что
'биологические виды представляют собой или могут представлять
более интегрированные единицы, чем популяционные системы
унипарентальных организмов. Можно сказать, что особи, при-
надлежащие к одной и той же популяции и одному и тому же
виду, у унипарентальных организмов связаны узами происхож-
дения от общей родительской особи, а у бипарентальных — уза-
ми общих родителей, а также узами скрещивания. Сцепления,
возникающие в результате перекрестного оплодотворения, слу-
жат фактором интеграции у популяций и видов бипарентальных
организмов, отсутствующим в унипарентальных группах.
Фактически дискретные группировки видов обычно различи-
мы у организмов с половым размножением (примером служит
большинство родов и семейств у млекопитающих и птиц), но
явно отсутствуют во многих унипарентальных группах расте-
ний, примером которых служат крупные группы видов таких ро-
дов, как Rubus и Hieracium.
Кроме того, как было показано в гл. 1, проводя различия
между бипарентальным и унипарентальным размножением, мы
не отражаем в полной мере существующее положение вещей.
У известных нам видов растений наблюдается целый спектр
систем размножения — от широкого ауткроссинга на одном кон-
це спектра через всевозможные промежуточные состояния до
строго унипарентального размножения на другом его конце. От-
сюда следует, что степень интеграции, достигаемая у этих видов,
также варьирует, и притом коррелятивно: от максимальной у
свободно скрещивающихся видов до относительно рыхлой струк-
туры в преимущественно унипарентальных группах.
Иными словами, если, следуя Мегличу [574] и Симпсону
[732], считать, что видовая структура существует как у бипарен-
тальных, так и у унипарентальных организмов, то возникает
необходимость различать разные уровни интеграции у видов с
различной биологией размножения. Хорошо интегрированные
биологические виды — одно из состояний, наблюдаемых в при-
роде. От этого состояния имеется постепенный переход к раз-
личным промежуточным уровням интеграции, существующим у
многих видов высших растений с частично, преимущественно
или исключительно унипарентальным размножением.
В системах популяций бипарентальных организмов изменчи-
вость имеет иерархическую структуру: группы локальных рас
объединяются в географические расы, а группы географических
рас — в биологический вид. В системах популяций унипарен-
тальных растений аналогичной иерархической структуры измен-
Гл. 5 Видовые единицы у унипарентальных растений 71
чивости нс наблюдается. Вместо этого у них обычно можно ви-
деть ряды морфологически различных единиц, или микровидов,
одного и того же ранга. Эти микровиды часто группируются
вокруг одного или нескольких предковых видов с половым
размножением.
Микровиды
Термином микровид (или жорданон) обозначают в ботанике
любую однородную популяцию, которая морфологически слегка
отличается от родственных ей однородных популяций. В такой
трактовке этот термин приложим к обширному кругу популя-
ций, оказываясь синонимом чистых линий, некоторых локальных
рас, а иногда видов-двойников и т п , и его действительно ис-
пользуют в этих и других случаях. Поскольку эгот термин при-
менялся широко и достаточно свободно, он, естественно, пере-
стал быть точным и его заменили другими, более строгими тер-
минами.
Однако в практике применения термина микровид есть одна
особенность, ради которой его следует сохранить. Ботаники
обычно используют этот термин для обозначения слабодиффе-
ренцированных популяций, относящихся к группам растений с
унипарентальным размножением.
Так, микровиды описаны в автогамной группе Erophila verna
и в агамоспермной секции Moriferus рода Rubus. Erophila verna
и секция Moriferus рода Rubus, конечно, очень сильно различа-
ются по систематическим связям, жизненным формам и систе-
ме размножения. На фоне таких очевидных различий между
этими двумя группами сходство их по характеру изменчивости,
достаточное, чтобы оправдать применение к ним общей концеп-
ции микровидов, приобретает важное значение. Кроме того,
этот общий элемент в характере изменчивости, к которому мож-
но приложить термин микровид, коррелирует с двумя друшми
общими свойствами, а именно с естественной гибридизацией и
унипарентальным размножением.
Не все ботаники признают эти общие свойства или хотя бы
знают о них Это особенно справедливо в отношении тех ис-
следователей, которые впервые стали применять термин микро-
вид; использование ими этого термина служит примером выде-
ления, в значительной мере интуитивного, некой популяцион-
ной единицы в ряду изменчивости известного типа Тем самым
была создана предпосылка для формулировки более опреде-
ленной концепции микровида.
Микровиды определяются здесь как популяции, принадле-
жащие к преимущественно унипарентальным группам растений,
в общем довольно однородным и лишь.незначителыю отличаю-
72 Часть вторая. Природа видов
щимся одна от другой морфологически. Их распространение не-
редко бывает ограничено относительно небольшой географичес-
кой областью, хотя это и не обязательно. Они часто имеют гиб-
ридное происхождение (прямо или косвенно) и часто навсегда
сохраняют гибридную конституцию.
В зависимости от способа воспроизведения микровиды де-
лятся на четыре главных класса: 1) клональные микровиды, вос-
производящиеся путем вегетативного размножения, например
Opuntia-, 2) агамоспермные микровиды, размножающиеся при
помощи агамоспермии, например секция Moriferus рода Rubtis;
3) гетерогамные микровиды, воспроизводящиеся при помощи
таких генетических систем, как у Oenothera biennis или Rosa
canina; 4) автогамные микровиды, которые преимущественно
автогамны и гомозиготны по хромосомам, как у Erophila.
Следует указать, что микровиды классов 1 и 2 размножа-
ются бесполым путем, а классов 3 и 4 — половым.
Автогамные микровиды в большинстве случаев не исключи-
тельно автогамны; частично у них наблюдается ауткроссинг и,
кроме того, нормальный половой цикл. Следовательно, такие
автогамные микровиды представляют собой один из типов био-
логических видов. Концепция автогамных микровидов, к которой
мы здесь подошли с одной стороны, в значительной мере сов-
падает с концепцией биологических видов-двойников, изложен-
ной в гл. 4. Дальнейшее обсуждение видов-двойников в авто-
гамных группах растений дано в гл. 11.
Проблема вида у клональных и агамоспермных микровидов,
не способных к половому размножению и не обладающих свой-
ствами биологических видов, осложняется в еще большей сте-
пени, чем у автогамных микровидов. У гетерогамных микрови-
дов, также обладающих гибридной конституцией, хотя они. и
размножаются половым путем, также возникают сложности в
связи с проблемой вида. Примеры клональных, агамоспермных
и гетерогамных микровидов описаны в соответствующих главах
восьмой части.
Гибридность и микровиды
Гибридность обычно ассоциирована с микровидами. Растения,
принадлежащие к агамоспермным и гетерогамным микроврдам,
представляют собой перманентные гибриды. Они обычно высо-
ко гетерозиготны и в случае клональных микровидов могут быть
межвидовыми гибридами. Автогамные микровиды в отличие от
микровидов трех остальных классов не сохраняют гибридной
конституции и могут быть полностью или почти полностью го-
мозиготными; однако, подобно другим микровидам, они часто
возникают как продукты гибридизации.
Гл. 5. Видовые единицы у унипарентальных растений
73
В случае гибридных микровидов гетерозиготные индивиду-
умы создают однородные популяции в результате апомиксиса
или гетерогамии В процессе своего возникновения микровид
проходит через стадию, когда его распространение сильно огра-
ничено; в случае же успешной эволюции микровид достигает
стадии широкого географического распространения. Процесс
роста популяции от гибридного индивидуума через небольшой
клоп и эндемичный микровид до широко распространенного ми-
кровида протекает непрерывно. В некий не поддающийся опре-
делению момент на поздних стадиях этого постепенного про-
цесса клональный, агамоспермный или гетерогамный микровид
приобретает право рассматриваться как таксономически само-
стоятельная единица.
Естественная гибридизация между двумя или несколькими
видами, прежде размножавшимися половым путем, за которой
следует унипарентальное размножение образовавшихся гибри-
дов и жизнеспособных вариантов, выщепляющихся в дальней-
ших поколениях, приводят к формированию многочисленных
микровидов. В группах растений, обладающих этим сочетанием
эволюционных факторов, обычно можно обнаружить ряд как
широко распространенных, так и эндемичных микровидов, ко-
торые в процессе филогенеза наложились на исходные биоло-
гические виды, затушевывая существующую между ними пре-
рывистость.
В таких группах растений изменчивость не ограничена опре-
деленными рамками, соответствующими границам вида, по ох-
ватывает два или несколько родительских видов и все возник-
шие от них микровиды. Подобные группы растений создают
огромные таксономические трудности Биологическая концепция
вида неприложима к такой группе как целому и непригодна
Для нее. Альтернативные таксономические концепции вида на
практике также оказываются несостоятельными. Причина этих
неудач заключается в том, что рассматриваемые группы расте-
ний— это смесь, состоящая обычно из нескольких исходных
биологических видов и многочисленных происходящих от них
микровидов.
Кроме того, это смесь, которую нельзя с помощью обычных
таксономических методов разделить на составные элементы.
Примеры таких гибридных комплексов, состоящих из биологи-
ческих видов и микровидов, будут приведены в главах, отно-
сящихся к восьмой части книги.
Пересмотр вопроса о гомогенности вида
Унипарентальные растения могут пролить некоторый свет на
вопрос, обсуждение которого не было доведено до конца в пре-
74 Часть вторая. Природа видов
дыдущей главе, а именно на вопрос о роли потока генов и скре-
щивания в усилении гомогенности. Как уже говорилось ранее
(см. гл. 4), известные нам скорости потока генов слишком ма-
лы, чтобы этот поток мог оказывать существенное гомогенизи-
рующее воздействие на изменчивость многолетних растений со
свободным неродственным скрещиванием на больших площадях
в течение периодов в 10000 лет.
У унипарентальных растений уровень как обмена генами, так
и скрещивания, конечно, гораздо ниже, чем у растений со сво-
бодным неродственным скрещиванием. Тем не менее гомоген-
ность наблюдается и у микровидов с обширным ареалом. Что
же определяет гомогенность у унипарентальных растений?
Сходство индивидуумов в пределах отдельных видов не обя-
зательно приписывать воздействию потока генов и скрещива-
ния. Существует другое объяснение, которое, несмотря на его
очевидность, часто упускают из виду, а именно рост численно-
сти индивидуумов, происходящих от общего предка.
Рост численности популяции происходит как у бипаренталь-
ных, так и у унипарентальных организмов. Отличить эффекты
роста численности от эффектов, создаваемых потоком генов у
бипарентальных растений, трудно. Но у унипарентальных расте-
ний сходство между индивидуумами должно быть обусловлено
главным образом происхождением всей популяции от одного
общего предка (в отличие от моих прежних взглядов; см. [293]).
Репликативный характер воспроизведения, быть может, в зна-
чительной мере обусловливает гомогенность видовой популяции
также и у бипарентальных растений, хотя у них роль этого
фактора по сравнению с ролью потока генов остается неизвест-
ной [298].
6 Вопрос о виде у растении
У высших растений наблюдается чрезвычайно широкий диапа-
зон типов организации родственных особей в популяционные
системы. В царстве растений, как и в царстве животных суще-
ствуют хорошо интегрированные биологические виды, однако
это не единственная форма организации популяций у растений.
У них имеются также большие и разнообразные популяционные
системы на уровне, таксономически соответствующем секции
или подроду, причем эти системы не подразделяются на четко
обособленные виды. Кроме того, у растений встречаются виды,
занимающие по уровню интеграции промежуточное положение
между этими двумя крайними ситуациями.
Далее в этой главе мы займемся следующими вопросами: с
какой частотой встречаются у растений эти различные типы
организации вида? Какие факторы определяют их возникнове-
ние?
Для того чтобы найти подход к решению этих вопросов, не-
обходимо установить некий эталон для сравнения. В качестве
такого эталона удобно использовать ряд дискретных и хорошо
интегрированных биологических видов, наблюдаемых у многих
или даже у большинства высших животных, например в семей-
стве кошачьих, после чего можно рассматривать сходства с этим
эталоном или отклонения от него, обнаруживаемые у растений
У растений отклонения от организации, типичной для хорошо
развитого биологического вида, принимают обычно две формы.
У бипарентальных растений между двумя или несколькими
скрещивающимися группами может существовать репродуктив-
ная изоляция, занимающая промежуточное положение между
уровнями изоляции, характерными для биологических видов и
рас. Или же бипарентальное размножение может быть заме-
щено каким-либо способом унипарентального размножения.
В том и другом случаях отклонения могут колебаться от уме-
ренных до чрезвычайно резких. И соответственно этим двум
формам отклонений от принятого нами эталона в их крайних
выражениях мы различаем два главных типа «невидов», а имен-
но— полувиды и микровиды. Микровиды были рассмотрены в
предыдущей главе, а здесь мы рассмотрим полувиды.
Как будет показано в дальнейшем, такие «невиды» (nonspe-
sics) не столь уж необычны, во всяком случае в некоторых фло-
рах, по сравнению с биологическими видами. Их весьма широ-
кое распространение приписывают отчасти природе процессов
видообразования, а отчасти основным характеристикам самих
растений.
76
Часть вторая Природа видов
Полувиды как результат постепенного
видообразования
Обычный способ видообразования у растений и животных с пе-
рекрестным оплодотворением состоит в постепенной диверген-
ции двух или нескольких популяционных систем. На ранней
стадии дивергенции эти популяционные системы ведут себя как
географические или экогеографические расы. Они занимают
соседние территории, свободно скрещиваясь друг с другом и
интерградируя в зонах контакта. На поздней стадии диверген-
ции взаимоотношения, между этими популяционными системами
приобретают характер взаимоотношений между симпатрически-
ми видами, обитающими на одной и той же территории, но не
.скрещивающимися друг с другом. Между ранней и поздней
стадиями дивергенции и репродуктивной изоляции наблюдают-
ся различные промежуточные стадии, когда популяции не пред-
ставляют собой ни хороших рас, ни хороших видов, но связаны
друг с другом благодаря незначительному скрещиванию и сла-
бому обмену генами. Популяционные системы, находящиеся на
таких промежуточных стадиях дивергенции, были удачно назва-
ны полувидами (обзор и библиографию см. [282]).
Процесс географического видообразования будет рассмотрен
в гл. 13. Здесь же нас интересует один из результатов этого
процесса. Постепенная и непрерывная эволюционная диверген-
ция от географической расы до симпатрического вида неизбеж-
но приводит к тому, что в любой данный момент можно обна-
ружить несколько популяционных систем, находящихся на про-
межуточной стадии полувидов.
Во многих родах растений мы находим дизъюнктивные алло-
цатрические популяции, которые занимают, каждая в пределах
своего ареала, сходные экологические местообитания и морфо-
логически дифференцированы до такой степени, что системати-
ки присваивают им разные видовые названия, но которые при
экспериментальных скрещиваниях оказываются интерфертиль-
ными. Примером служат Catalpa bignonioides из восточной час-
ти США и С. ovata из Китая [778]. Другой пример — Platanus
racemosa и Р. wrightii, произрастающие соответственно в оро-
шаемых горными потоками долинах Калифорнии и в каньонах
Аризоны. Arbutus menziesii и A. arizonica, произрастающие в ле-
систых районах Калифорнии и Аризоны, разделенных обширной
пустыней, служат третьим примером; в данном случае неизвест-
но, дают ли они при скрещиваниях плодовитое потомство или
нет. В этих и других аналогичных случаях отдельные популя-
ционные системы уже нельзя считать, расами, но еще нельзя
рассматривать как биологические виды. Они находятся на по-
лувидовом уровне дивергенции.
Гл 6 Вопрос о виде у растений
77
Интересную промежуточную ситуацию представляют собой
перекрывающиеся кольца рас. Такая ситуация возникает в тех
случаях, когда ряд рас интерградирует по какой-либо географи-
ческой трансекте. Терминальные расы такого ряда, различаю-
щиеся сильнее всего, сосуществуют симпатрически, не скрещи-
ваясь одна с другой в сколько-нибудь значительной степени.
Таким образом, у одной и той же популяционной системы в
разных частях ее ареала проявляются черты, свойственные ра-
се или виду. Наиболее хорошо изученные примеры кольцевого
перекрывания рас относятся к птицам и другим животным (об-
зор см. [569]). Среди растений, принадлежащих к флоре запад-
ных районов США, подобные примеры обнаружены, в частно-
сти, в следующих группах видов: Gilia capitata, Ipomopsis aggre-
gata, Aquilegia chrysantha, Mimulus longiflorus и Pinus pondero-
sa ([54, 266, 268, 270, 334], а также Grant, неопубликованные
данные).
Mimulus puniceus (Scrophulariaceae) встречается в прибреж-
ной зоне на юге Калифорнии, а М. longiflorus — в удаленных
от побережья горах (рис. 3.2). В южной части Калифорнии, па
обширном пространстве от береговой линии до расположенных
дальше от моря гор, между этими двумя единицами происходит
полная интерградация (см. гл. 3). Однако в тех частях гор
Санта-Ана, где местообитания, пригодные для М. puniceus и
М. longiflorus находятся очень близко друг от друга, популя-
ции этих двух видов сосуществуют, не интерградируя [54]. Таким
образом, М. puniceus и М. longiflorus ведут себя во многих об-
ластях как расы, а в некоторых местах как симпатрические
виды.
Полувиды как результат естественной
гибридизации
Процесс эволюционной дивергенции не обязательно неуклонно
идет лишь в одну сторону. Он обратим в довольно широких пре-
делах, поэтому полувиды могут возникать не только как еди-
ницы, находящиеся на пути превращения в виды, но также и
как продукты гибридизации между видами, которые прежде
были хорошо изолированы.
Несмотря на то что биологические виды интерстерильны и
хорошо изолированы в других отношениях, между ними могут
происходить естественная гибридизация и обмен генами, до тех
лор пока эффективность барьеров, препятствующих скрещива-
нию, не достигнет 100%. Эти барьеры между родственными
видами растений, принадлежащих к одному и тому же роду
или секции, в сущности, часто могут быть до известной степени
78
Часть вторая. Природа видов
разрушены. Нередко можно встретить два симпатрических ви-
да, различающихся и не скрещивающихся между собой на боль-
ших пространствах, но образующих гибриды в одной или не-
скольких местностях.
Межвидовая гибридизация — потенциальная угроза целост-
ности родительских видов как отдельных скрещивающихся
групп. Превратится ли эта потенциальная угроза в свершив-
шийся факт или нет —вопрос другой. Межвидовая гибридиза-
ция может привести к разнообразным конечным результатам
(см. часть пятую). Некоторые из этих результатов не нарушают
обособленность участвующих в гибридизации видов, а поэтому
в данный момент они нас не интересуют.
Здесь мы рассмотрим образование путем гибридизации но-
вых популяций, позволяющих преодолеть барьеры к скрещива-
нию, морфологический разрыв и экологическую дифференциа-
цию между первоначально обособленными родительскими вида-
ми. Такие связующие популяции гибридного происхождения де-
лятся на два класса: скопления гибридов (hybrid swarms) и
интрогрессивные популяции. Их образование поворачивает про-
цесс видообразования вспять и, если гибридизация происходит
в достаточно широких масштабах, превращает виды, бывшие
прежде самостоятельными, в полувиды.
Скопление гибридов представляет собой сложную смесь ро-
дительских форм, гибридов Fi, особей, возникших в результате
возвратных скрещиваний и расщепления. В таких скоплениях
наблюдается, как и следует ожидать, очень высокая степень
индивидуальной изменчивости. В литературе описано много при-
меров скоплений гибридов, но мы приведем здесь лишь некото-
рые из них.
Скопления гибридов между Geum rivals и G. urbanum встре-
чаются во многих местах в Англии и Европе [230, 557]. Aquile-
gia formosa и A. pubescens образуют скопления гибридов в раз-
личных зонах контакта этих двух видов в горах Сьерра-Невада
в Калифорнии [96, 270]. Колонии, образуемые тремя или четырь-
мя видами Baptista (Leguminosae) и их гибридами, встречаются
на берегу Мексиканского залива в Техасе [9—11].
При многократных возвратных скрещиваниях гибридов с
одним из родительских видов возникает интрогрессивная попу-
ляция, сходная с данным родительским видом, но проявляющая
изменчивость в сторону другого родительского вида [15, 16].
Если интрогрессия заходит достаточно далеко, то она может
привести к стиранию морфологических и экологических разли-
чий между исходными видами, которые в результате будут низ-
ведены до уровня полувидов.
В некоторые колонии Iris hexagona giganticaerulea в дельте
Миссисипи на юге шт. Луизиана интрогрессирует Iris ftilva, так
Гл. 6. Вопрос о виде у растений
79
что морфологический разрыв между нормальными незасорен-
ными формами этих двух видов заполняется их диапазоном из-
менчивости [686]. Хотя эти два вида Iris в большинстве облас-
тей, где они симпатричны, обособлены один от другого, это
обособление не полное и в некоторых зонах гибридизации они
интерградируют, соединяясь через ряд промежуточных форм. Та-
ким образом, они представляют собой полувиды гибридного
происхождения.
Определение биологического вида как наиболее инклюзив-
ной скрещивающейся группы оказывается несостоятельным в
случае естественно гибридизирующих видов и полувидов. Там,
где происходит ограниченный обмен генами между полувидами,
в остальном изолированными друг от друга, наиболее инклю-
зивной скрещивающейся единицей является не единый биологи-
ческий вид, а совокупность полувидов. Такую совокупность на-
зывают сингамеоном. Сингамеоп — самая обширная скрещиваю-
щаяся система в группе гибридизирующих видов. Сингамеон
ведет себя на своей внешней границе как хорошо изолирован-
ный биологический вид, но отличается от него более сложной
внутренней структурой.
Сингамеоны обычны у разных родов растений: Pinus, Quer-
cus, Geum, Aquilegia, Iris и др. Примеры будут описаны в гл. 18.
Масштабы проблемы вида
У бипарентальных растений полувиды гибридного происхожде-
ния достаточно обычны, во всяком случае в некоторых флорах.
Например, в результате изучения различных родов растений со
свободным неродственным скрещиванием во флоре западной
части США у меня создалось впечатление, что сингамеоны,
состоящие из гибридных полувидов, часто более обычны, чем
хорошо выраженные биологические виды. Это наблюдается у
Quercus, Ceanothus, Diplacus, Iris и у свободно скрещивающих-
ся представителей Gilia в западных областях США. У других
родов свободно скрещивающихся растений из тех же областей,
например Pinus, Aquilegia и Ipomopsis, гибридные полувиды
составляют устойчивое меньшинство.
Гибридные комплексы, содержащие многочисленные унипа-
рентальные микровиды, столь же обычны. Комплексы агамо-
спермных микровидов обнаружены у таких северных родов, как
Antennaria, Crepis, Hieracium, Taraxacum, Роа, Ranunculus, Ru-
bus и Potentilla. Густафсон [317] перечислил 73 рода растений
из всех частей земного шара, содержащих агамоспермные мик-
ровиды. Некоторые другие роды, в том числе Allium, Festuca и
Saxifraga, содержат унипарентальные гибридные формы, раз-
множающиеся путем ложного живорождения.
80 Часть вторая. Природа видов
Отклонения от эталона хороших биологических видов пред-
ставляют собой, таким образом, обычное явление у высших
растений; достаточно обычны и крайне сильные отклонения, о
чем свидетельствуют случаи распада видовой организации. Во
всех досконально изученных примерах участвует гибридизация
в.сочетании либо с унипарентальным, либо с бипарентальным
размножением. В этом заключается одно из самых важных раз-
личий между растениями и многими группами высших живот-
ных, у которых хорошие виды являются правилом, а сингамео-
ны или агамные комплексы — исключением.
Причины и следствия, вытекающие из концепции
вида у растений
Сравнивая хорошо изученные группы высших растений со мно-
гими группами птиц, млекопитающих и таких родов насекомых,
как Drosophila, мы обнаруживаем, что у растений дискретные
биологические виды в общем встречаются реже и что для них
более характерны иные ситуации. Отсюда следует, что причин-
ные факторы, определяющие различные типы организации вида
в двух царствах живой природы, также должны различаться.
Поэтому сравнительный подход к проблеме вида может пролить
некоторый свет на те факторы, от которых зависит организация
вида у высших растений.
Очевидное и коренное различие между растениями и живот-
ными—это степень сложности индивидуального организма.
Развитию относительно простого растительного организма со-
ответствует открытая система роста с серийным образованием
новых частей. Тело животного имеет гораздо более сложное и
топко сбалансированное строение, а кроме того, оно развивается
как единое целое в результате закрытой системы роста. Логично
допустить, что организмы этих двух типов различаются также
по сложности и интегрированности генетических систем, конт-
ролирующих их рост и развитие.
Одна эта предпосылка позволяет понять два главных факто-
ра, способствующих частичному или полному распаду видовой
организации во многих группах растений. Эти два фактора —
частая межвидовая гибридизация и унипарентальное размно-
жение.
Виду, обладающему высокоинтегрированным и точно сбалан-
сированным генотипом, межвидовая гибридизация нанесет отно-
сительно больший вред, чем виду с более простым и менее стро-
го координированным набором генов. Относительно простая
физиолого-морфологическая организация тела растения, отра-
жающая соответственно относительно более простой генотип,
Гл. 6. Вопрос о виде у растений 8Ь
возможно, делает растения более устойчивыми к межвидовым
обменам генами, чем большинство групп животных (279]. Сле-
довательно, из открытой системы роста растений вытекает воз-
можность разнообразных форм вегетативного и агамоспермного
размножения, в результате чего численность гибридов и их по-
томков будет увеличиваться.
Как писал Густафсон [317]: «Виды явнобрачных растений
снова и снова оказываются жертвой своих собственных черт:
своей свободной системы роста, своего вегетативного распрост-
ранения. Эти свойства‘непрерывно способствуют возобновлению
популяции, ее изменению в направлении развития полиплоид-
ное™ и апомиксиса».
Эти основные свойства растений не только сказываются на
характере растительных видов, но и оказывают глубокое влия-
ние на макроэволюцию растений. Как мы увидим в дальнейшем^
в процессе филогенеза растений неоднократно имело место об-
разование анастомозов между линиями, которые прежде были
обособленными. Если у животных видообразование обычного
типа приводит к формированию филогенетического древа, то у
растений оно нередко ведет к образованию филогенетической
сети.
6—45
7 Разные типы видов
Общий термин «вид» используется в самых разных значениях.
Поэтому, для того чтобы уменьшить число возможных недора-
зумений, желательно в каждом конкретном случае с помощью
прилагательных или приставок указывать, какой именно смысл
в него вкладывается.
Многочисленные авторы, занимавшиеся этой проблемой,
различают разные типы видов, давая им разные названия [см.
53, 79, 81, 157, 244, 279, 293, 564, 570, 732, 737, 752, 855].
Принятая здесь классификация типов видов вобрала в себя
некоторые черты более ранних систем, в частности систем Кэй-
на [79] и Симпсона £732], но распределяются они иным образом.
Настоящая система отличается, кроме того, от большинства
прежних тем, что в ней последовательно используется единст-
венный критерий — характер классифицируемых биологических
объектов.
Типы видов
Как показывает опыт, биологам необходимо различать по край-
ней мере четыре типа видов, чтобы охватить все разнообразие
•биологических объектов. Иными словами, существует не менее
четырех различных типов видовых единиц, для которых в бота-
нике и зоологии необходимо иметь четыре различные концепции
вида. В биологии растений полезно иметь еще одну, пятую кон-
цепцию вида.
Пять типов видов, различаемых в этой книге, перечислены
ниже с указанием некоторых часто употребляемых синонимов:
1. Таксономический вид (морфологический вид, фенети-
ческий вид)
2. Биологический вид (генетический вид)
3. Микроцид (агамный вид)
4. Последовательные (successional) виды (палеонтологи-
ческие виды)
5. Биосистем этический вид (эковид, ценовид)
Характеристики видов этих пяти типов приведены в табл. 7.1.
Следует указать, что каждая из перечисленных выше концеп-
ций вида имеет силу в своей собственной сфере. Однако при
использовании той или иной концепции вида за пределами этой
сферы возникают недоразумения. Как уже отмечалось, недора-
зумения могут возникнуть также в тех случаях, когда термин
«вид» используют без соответствующих пояснений, что бывает
достаточно часто.
Гл 7. Разные типы видов
8$
Таблица 7.1. Характеристики видов разных типов
Тип Классифицируемые единицы Сфера применения
Таксономический Таксоны; группы мор- фологически сходных индивидуумов Группы с морфологическими раз- личиями, используемыми в фор- мальной классификации
Биологический Системы популяций, размножающихся по- ловым путем Организмы с половым размноже- нием, принадлежащие к одному временному уровню
Микровпд Популяции упипарен гальных организмов Унипарентальныс организмы, в том числе бесполые и парасексу- альные организмы
Последовательные виты Фнлетические линии Биологические виды н микровиды во времени
Биосистемэтиче- Группы, плодовитые Группы растений с половым раз-
ский вид при скрещиваниях множением, искусственно гибри- дизированные па эксперименталь- ных участках
Биологические виды, микровиды и последовательные виды в
отличие от двух других типов — естественные единицы в том
смысле, что они существуют в природе, а не просто категории,
созданные человеком. Эти три типа можно объединить в слож-
ную единицу — эволюционный вид Симпсона [732], который мы
обсудим в одном из дальнейших разделов этой главы.
Таксономический вид
Таксономический вид, представляющий собой основную едини-
цу формальной классификации, — универсальная категория,,
встречающаяся у всех форм живой материи, как это предусма-
тривается кодексами ботанической и зоологической номенклату-
ры. Кодексы номенклатуры определяют также положение вида
в иерархии таксономических категорий В ботанике такую
иерархию составляют (в восходящем порядке), форма, разно-
видность, подвид, вид, ряд, секция, подрод, род (при этом опу-
скаются различные возможные промежуточные категории, обра-
зуемые при помощи приставок над- или под-).
Таксономический вид основан на морфологическом сходстве
и различии и представляет собой совокупность морфологически
сходных индивидуумов, отличающуюся от других таких совокуп-
ностей. Полагаться на внешние морфологические признаки прак-
тически необходимо, поскольку морфологические признаки, в
том числе окраска, — единственные признаки, которые можно
постоянно использовать при повседневной работе с любой груп-
пой организмов. Таксономический вид должен быть единицей,
6’
«4
Часть вторая. Природа видов
которую было бы удобно применять в практической работе по
классификации и определению.
Недостаток критерия морфологического сходства и различия
состоит в том,что с его помощью невозможно объективно уста-
новить уровень различия, заслуживающий видового ранга. Нет
способа, который бы позволил объективно решить, представляет
ли собой данный таксон вид, разновидность или ряд. Поэтому
разные систематики могут создавать и создают различные си-
стемы классификации для одной и той же груйпы организмов.
Несомненно, что прерывистости в ряду изменчивости, раз-
граничивающие биологические виды, можно использовать также
для разграничения таксономических видов. В тех случаях, ког-
да это возможно, следует отождествлять таксономический вид
с видом биологическим. Однако такая возможность имеется не
всегда. Во всяком случае, это нельзя считать существенным или
обязательным элементом концепции таксономического вида.
Субъективный характер таксономического вида отражен в
высказываниях разных ученых. Приведем прежде всего одну
цитату из Дарвина [152, 154]:
«Не подлежит сомнению, что до настоящего времени не удалось прове-
зти ясной пограничной черты между видами и подвидами, т. е. формами,
которые, по мнению некоторых натуралистов, приближаются к видам, но не
вполне достигают этого ранга, или между подвидами и хорошо выраженны-
ми разновидностями, или, наконец, между менее четко выраженными разно-
•видностями и индивидуальными различиями. Эти различия плавно переходят
одни в другие, сливаясь в один непрерывный ряд...
Из всего сказанного ясно, что термин «вид» я считаю произвольным, при-
думанным ради удобства для обозначения групп особей, близко между со-
бой схожих, и существенно не отличающимся от термина «разновидность»,
которым обозначают формы, менее резко различающиеся и более колеблю-
щиеся [в своих признаках]».
Эта точка зрения, доведенная до своего логического конца,
была сформулирована Риганом следующим образом (цит. по
{392J):
«Вид — это сообщество (или несколько близко родственных сообществ),
отличительные морфологические признаки которого, по мнению компетент-
ного систематика, достаточно четко выражены, чтобы оправдать присвоение
«му (или им) видового названия».
Недавно Кронквист [131] дал разумное определение таксо-
номического вида:
«Виды —это самые мелкие группы, постоянно и последовательно сохра-
няющие свою обособленность и различимые обычными способами».
Дополняя это определение, Кронквист [131] пишет: «после-
довательно сохраняющие обособленность... означает, что все или
очень большая доля рассматриваемых особей явно принадле-
жат к одной или к другой группе, а не находятся где-то посере-
дине между ними». Слова «обычными способами» подчеркивают,
Гл. 7. Разные типы видов
85
что следует применять традиционные и практичные методы.
Так, для различения видов грибов принято (и необходимо) ис-
пользовать микроскоп, тогда как при определении видов семен-
ных растений обычно пользуются простой или препаровальной
лупой.
Практическая применимость для определения и классифика-
ции представляет собой, таким образом, один из важнейших
критериев таксономического вида. А отсюда следует, что так-
сономический вид — искусственная категория, которая во мно-
гих случаях не совпадает с естественными единицами. Ибо, как
удачно выразился Бодри [53]: «Природа... развивалась, совер-
шенно не заботясь о тех практических проблемах, с которыми
лридется столкнуться ее исследователям».
Биологический вид как одна из стадий процесса
дивергенции
Биологический вид рассматривался в гл. 4 в качестве одной из
популяционных единиц. Однако эту единицу можно рассматри-
вать также как одну из стадий более или менее непрерывного
ряда уровней дивергенции популяционных систем. Узловым точ-
кам в таком ряду соответствуют локальные расы, аллопатриче-
ские полувиды, аллопатрические виды и симпатрические виды
_ Группа
ВИДОВ В
Биологический
вид В
_Аллоп^трический
пдлувид В
Географическая
раса В
Локальная
раса В
Группа ___
ВИДОВ А
Биологический^ __
вид А
Аллопатрический
полувид А
Географическая
раса А
Локальная _____
раса А
Предковая популяция__________
ПРИЗНАКИ
Рис. 7.1. Последовательные стадии процесса постепенной первичной дивер-
генции.
Часть вторая. Природа видов
86
(рис. 7.1). При гибридизации между двумя биологическими ви-
дами эти единицы вновь обретают статус симпатрических полу-
видов и'образуют новую сборную единицу — сингамеон.
Хотя теоретически иерархия популяционных систем отлича-
ется от иерархии таксономических категорий, на практике меж-
ду ними существует определенное соответствие, когда дело ка-
сается групп растений с обычным половым размножением. Для
популяционных систем приняты следующие таксономические
эквиваленты:
Локальная раса — не различается таксономически
или рассматривается как раз-
новидность
Географическая раса — подвид; некоторые авторы счи-
тают ее разновидностью
Аллопатрический, полу- —часто рассматривается как
вид таксономический вид
Симпатрический полу- — таксономический вид
вид гибридного проис-
хождения
Аллопатрический био-----таксономический вид
логический вид
Симпатрический биоло- —таксономический вид
гический вид
Группа видов —ряд
Сингамеон - — ряд, секция
Последовательные виды
Поскольку биологический вид реагирует на изменяющиеся усло-
вия среды, весьма вероятно, что с течением времени он под-
вергнется изменениям. Изменяющийся биологический вид, рас-
сматриваемый в перспективе геологического времени, превра-
щается в пару последовательных видов. Другими словами,
последовательные виды—это члены одной и той же эволюцион-
ной линии, существующие в разные периоды времени и обла-
дающие различными признаками. Вернемся вновь к рис. 7.1:
А и В на одном определенном уровне схемы — это два биологи-
ческих вида; тогда как А и В в разных точках одной линии —
два последовательных вида.
Филетическая эволюция — процесс непрерывный, а поэтому
два последовательных вида связаны, по крайней мере теорети-
чески, рядом интерградирующих форм. Таким образом, решение
о том, где провести границу между ними, неизбежно становится
произвольным. На практике принятие такого решения часто об-
легчается наличием разрывов в палеонтологической летописи.
Гл. 7. Разные типы видов
87
Для многих современных видов хвойных известны третичные
предки. Ниже перечислены некоторые пары видов, относящиеся
к сем. Pinaceae и произрастающие'на западе Северной Амери-
ки. Слева указаны виды, произраставшие в районе Большого
Бассейна в миоцене, а справа — их современные эквиваленты,
встречающиеся в хвойных лесах западных районов Америки
(31]. Члены каждой пары очень сходны морфологически и, по-
видимому, представляют собой родственные последовательные
виды.
Миоцен
Pinus alvordensis
Р florissanti
Р. quinquifolla
Picea lahontense
P. sonomensis
Tsuga mertensioides
T. sonomensis
Pseudotsuga sonomensis
Abies alvordensis
A concoloroides
A klamathensis
A. lattearpus
A sonomensis
A. toxirivus
Современная эпоха
P. contorta
P. ponderosa
P monticola
P. engetmannii
P breweriana
T. mertensiana
T. heterophylla
P. menziesii
A. recurvata
A. concolor
A. shastensis
A. magnifica
A grandis
A. amabilis
Другие примеры последовательных видов рода Pinus можно
наблюдать у третичных и современных сосен с закрытыми шиш-
ками в прибрежных районах Калифорнии [30].
Эволюционный вид
Соединив биологический, унипарентальный и последователь-
ные виды, можно получить комбинированную концепцию эво-
люционного вида.
Эволюционный вид, по определению Симпсона [731], пред-
ставляет собой популяционную систему, обладающую следую-
щими характеристиками: 1) это линия, состоящая из последо-
вательных популяций предков-потомков, существующая в прост-
ранстве и во времени; 2) эволюция этой линии обособлена от
эволюции других таких линий, т. е. от других видов; 3) эта ли-
ния обладает собственной «единственной в своем роде эволю-
ционной ролью», т. е. соответствует своей особой экологической
нише в биологическом сообществе; 4) она ивдеет собственные
эволюционные тенденции, будучи способной изменять свою эво-
люционную роль на протяжении своей истории [732].
88 Часть вторая. Природа видов
Такими характеристиками обладают как унипарентальные,
так и бипарентальные организмы. Следовательно, концепция
эволюционного вида охватывает более разнообразные системы
скрещивания и носит более широкий характер, чем концепция
биологического вида в бипарентальных группах.
Концепция эволюционного вида придает перспективность
проблеме гибридизации и интегрированности видов. Естествен-
ная гибридизация неизбежно понижает интегрированность уча-
ствующих в ней видов. Однако в свете концепции эволюционно-
го вида главный вопрос состоит не в том, происходит ли между
двумя данными видами гибридизация, а в том, утрачивают ли
два гибридизирующих вида свои различные экологические и
эволюционные роли или нет. Если, несмотря на некоторую гиб-
ридизацию, они не сливаются, то, следовательно, в эволюцион-
ной перспективе они остаются отдельными видами [732].
Концепция эволюционного вида проливает также свет на
причины гомогенности видов. Мы уже обсуждали эту проблему
в связи с биологическими и унипарентальными видами (см.
гл. 4 и 5) и имели основания усомниться в том, что поток генов
может служить главным фактором, определяющим гомогенность
во многих группах растений. Рассматривая те же самые при-
меры с точки зрения концепции эволюционного вида, мы вводим
в обсуждение исторический фактор, а вместе с ним — удовлетво-
рительное альтернативное объяснение: гомогенность видовой по-
пуляции может в значительной мере определяться происхожде-
нием от общего предка.
Биосистематические категории
Выделение биосистематических категорий основано на плодо-
витости при скрещиваниях, определяемой в экспериментах по
искусственной гибридизации. Хорошо известна следующая
иерархия этих категорий (в восходящем порядке): экотип, эко-
вид, ценовид, компариум [96, 100, 102, 136, 840, 842].
Экотип слагается из «всех членов данного вида, приспособ-
ленных к выживанию в среде определенного типа в пределах
общего ареала распространения данного вида». Два или не-
сколько экотипов «способны свободно обмениваться генами без
снижения плодовитости или мощности потомков» [102].
Эковид представляет собой совокупность экотипов. Два или
несколько эковидов разделяются барьерами неполной стериль-
ности; например, между ними возможна искусственная гибриди-
зация, причем гибриды Fi могут обладать значительной, но не
полной стерильностью.
Ценовиды —это совокупности родственных Зковидов. Раз-
личные ценовиды можно скрещивать друг с другом — обычно
Гл. 1. Разные типы видов
89
с трудом, и получающиеся гибриды Fi высоко или полностью
стерильны. Поэтому обмен генами между ценовидами очень ог-
раничен или не происходит вовсе
Компариум слагается из группы родственных ценовидов.
Различные компариумы не удается искусственно скрещивать
друг с другом, так как этому препятствует очень сильно выра-
женная несовместимость.
Представленная здесь система популяционных единиц носит
особый характер. Она основана на одном, по при этом очень
важном аспекте изоляции, а именно на внутренней репродуктив-
ной изоляции. Биосистсматические категории дают возможность
систематизировать (в тех случаях, когда они известны) взаимо-
отношения между группами по плодовитости
В «нормальной» группе растений биосистематические кате-
гории можно приравнять к соответствующим популяционным
единицам примерно следующим образом экотип=локальная
раса; эковид и ценовид=биологический вид; компариум = био-
логический вид, сингамеон или группа видов. Корреляции эти
не слишком точны в «типичных» группах и еще менее точны во
всех остальных.
Причина этого заключается в том, что преграды, обуслов-
ленные стерильностью, представляют собой лишь одну группу
изолирующих механизмов. Существуют, кроме того, внешние
изолирующие механизмы, которые могут быть весьма эффектив-
ны в природных условиях В некоторых группах растений име-
ются хорошо развитые внешние изолирующие механизмы, нс»
слабо развитые внутренние механизмы. Поэтому биосистемати-
ческие категории имеют более непосредственное отношение к
ситуациям, возникающим на экспериментальных участках, чем
к тому, что происходит в природе.
Единицы классификации культурных растений
Виды и группы видов, в которые входят культурные растения,
содержат больше отклоняющихся форм, чем аналогичные груп-
пы тех же размеров у дикорастущих растений, а использование
географических критериев затруднено, что связано с условиями
возделывания. Как указывают Харлан и де Вет [347]: «Методы
формальной таксономии не очень пригодны для классификации
культурных растений». Для этих целей необходим какой-то иной
способ классификации.
Харлан и де Вет [347] предложили иерархию категорий, в
•основе которой лежит генофонд. На видовом уровне и выше они
различают три категории генофондов GP-1, GP-2 и GP-3.
Первичный генофонд (GP-1) эквивалентен биологическому
виду Хорошо изолированные виды относят к разным катего-
90
Часть вторая. Природа видов
риям GP-1. Если растения, принадлежащие к одной и той же
группе, различаются по плоидности, то их обычно отнорят к раз-
ным GP-1.
Вторичный генофонд (GP-2) представляет собой группу био-
логических видов, которые могут быть скрещены с культурным
растением. Барьеры в виде стерильности или пониженной жиз-
неспособности гибридов существуют, но они могут быть преодо-
лены. Категория GP-2 примерно соответствует ценовиду в био-
систематике.
Третичный генофонд (GP-3) — наиболее обширная из раз-
личаемых в этой иерархии единиц; она соответствует наивыс-
шей скрещиваемости и наивышйему потенциально возможному
переносу генов. Гибриды между разными GP-3 получить труд-
но, и они нежизнеспособны или полностью стерильны. Обмен
генами между разными GP-3 достигается только с помощыо
специальных методов, таких, как выращивание зародышей в.
культуре для получения гибридов или удвоение хромосом для
восстановления плодовитости.
У Triticum к GP-1 принадлежат виды Г. monococcum, Т. iur-
gidum, Т. timopheevi и Т. aestivum. В единицу GP-2, к которой
принадлежит Triticum, входят также роды' Aegilops, Secale,
Haynaldia и три вида Agropyron. Единица GP-3, в которую вхо-
дят эти виды пшеницы и родственные ей роды, содержит, кро-
ме того, несколько других видов Agropyron и несколько видов
Elymus [347].
Некий вид GP-1, относящийся к какой-либо группе культур-
ных растений, обычно содержит не только- возделываемые, но
и спонтанные формы (либо сорняки, либо дикорастущие, либо
и те и другие вместе). Харлан и де Вет [347] предлагают ис-
пользовать для классификации этих альтернативных форм кате-
горию подвида. Вид GP-1 делится на два подвида, А и В. К под-
виду В относят сорняк и дикорастущую форму, т. е. спонтанные
расы. Подвид А сохраняют для культурных сортов.
Вид GP-1 и его подвиды А и В имеют формальный таксоно-
мический статус, и им присваиваются латинские названия.
Существует также ряд категорий рангом ниже подвида, не
имеющих формального статуса. Их можно; представить в сле-
дующем виде:
Подвид В
Раса
Подраса
Подвид А
Раса
Подраса
Культивар
Линия, клон или генотип
Гл. 7. Разные типы видов
91
Биологический вид или таксономический вид?
Разногласия по проблеме вида сосредоточены на взаимоотно-
шениях между таксономическими и биологическими видами.
Между этими двумя типами видов* имеются как сходства, так
и различия. В ранний период истории и концепции вида говорили
преимущественно о сходстве, тогда как ближе к нашему времени
стали уделять больше внимания различиям
Натуралисты раннего периода — Джон Рэй [668], Линней
[512—515], Бюффон [74] и другие — занимались высшими жи-
вотными и растениями в пределах местной фауны и флоры. Сле-
довательно, они имели дело с симпатрическнми популяциями
организмов, размножающихся половым путем и различающихся
по хорошо выраженным морфологическим признакам. Виды,
которые они выделяли, можно было одинаково хорошо опреде-
лить как по взаимоотношениям при скрещивании, так и по пре-
рывистости в изменчивости морфологических признаков. Самая
инклюзивная единица бипарентального размножения и основ-
ная таксономическая единица были для них синонимами Ины-
ми словами, между биологическими и таксономическими вида-
ми никакого различия не делалось, да и нужды в нем не было.
Для многих групп организмов, изучаемых современными
методами, также наблюдается хорошее соответствие между кри-
терием, основанным на способности к скрещиванию, и крите-
рием морфологической прерывистости. Прерывистости в измен-
чивости, разграничивающие биологические виды, делают также
возможным распознавание таксономических видов В таких
группах таксономический вид остается синонимом биологиче-
ского вида.
Однако критерии, основанные на скрещиваемости и на мор-
фологических различиях, не всегда совпадают. Расширение си-
стематических биологических исследований со времен первых
натуралистов выявило многочисленные примеры, в которых эти
два критерия не совпадают, что положило начало проблеме вида.
Существуют три часто встречающиеся ситуации, в которых
не удается или не представляется возможным приравнять таксо-
номический вид к биологическому виду. Это касается видов-
двойников, полувидов и видов с унипарентальным размноже-
нием. Каждая из этих ситуаций широко распространена у выс-
ших растений.
Система таксономических категорий универсальна. Все орга-
низмы следует распределять по родам и видам, и каждому из
них следует присваивать отдельное название. Но не все орга-
низмы размножаются половым путем. А в бесполых группах
«биологических видов не существует.
92 Часть вторая Природа видов
Кроме того, таксономическая иерархия отличается жест-
костью. В ней есть две категории — вид и подвид. Каждое рас-
тение, согласно правилам ботанической номенклатуры, относит-
ся к определенному виду. Полувидов в таксономической иерар-
хии не существует. Но некоторые популяционные системы &
природе находятся на полувидовом уровне дивергенции. Если,
угодно, их можно считать таксономическими видами,но они не
являются биологическими видами.
В связи с этим уместно напомнить, что иерархия таксоно-
мических единиц была создана до появления теории эволюции.
Лиинеевский вид был отлит в креационистскую форму. Линней
писал в 1764 г. (цит. по [768]): «Мы различаем столько видовг
сколько различных форм было сотворено изначально». Со вре-
мени Линнея никаких серьезных изменений в таксономическую-
иерархию внесено не было. Но естественные популяционные
системы эволюционируют. Их не всегда удается втиснуть в «до-
эволюционный» набор категорий. Семена, из которых выросла
проблема вида, были посеяны давно.
Третий случай, в котором критерии скрещиваемости и мор-
фологических различий вступают в противоречие, это виды-
двойники. Такие виды представляют собой настоящие биологи-
ческие виды, но их очень трудно различить по морфологическим
признакам, вследствие чего и нельзя считать хорошими таксоно-
мическими видами. Это не означает, что родственные виды-
двойники невозможно различить по фенотипическим призна-
кам. Фактически такое различие можно провести в любой груп-
пе достаточно полно изученных видов-двойников. Однако для-
того чтобы произвести идентификацию в этих группах, необхо-
димо приложить гораздо больше усилий, чем это требуется &
обычной таксономической практике. В обычной практике разде-
ление биологических видов может оказаться неудобным или да-
же неосуществимым, поэтому за единицу формальной классифи-
кации приходится принимать весь комплекс видов-двойников.
Такое положение возникает обычно в связи с аллополиплои-
дией. Рассмотрим группу трех родственных видов (двух диплои-
дов и одного тетраплоида) с геномами АА, ВВ и ААВВ. Одни
лишь внешние морфологические признаки могут оказаться не-
достаточно надежными в качестве критериев для различения
этих видов. Однако если помимо морфологических данных рас-
полагать сведениями о числе хромосом, то этого обычно бывает
достаточно для идентификации. Два диплоидных вида АА и
ВВ — морфологически крайние типы в пределах данной группы,
и их, как правило, можно отличить один от другого по внешним
признакам. Морфологически промежуточный вид ААВВ надеж-
но отличается от обоих диплоидов даже по одному лишь числу
хромосом. Таким образом, виды АА, В В и ААВВ можно идеи-
Гл. 7. Разные типы видов
93
тифицировать по совокупности наружных и внутренних феноти-
пических признаков Однако первый этап идентификации —
определение числа хромосом, для чего необходимо исследовать
живой растительный материал под микроскопом — процедура,
выходящая далеко за пределы методов, которые можно считать
стандартными для определения видов у высших растений
Здесь мы сталкиваемся с новой формой старого расхожде-
ния между естественной и искусственной системами классифи-
кации. Первая отражает генетические связи, но может оказать-
ся неудобной для применения, вторая полезна для практическо-
го использования в таксономии, но не всегда отражает биологи-
ческое родство Расхождение между этими подходами к класси-
фикации обычно затрагивает высшие категории. Однако оно
вновь возникает на видовом уровне во всех тех случаях, когда
таксономические виды, различаемые в повседневной практике,
неэквивалентны биологическим видам [283].
Таким образом, концепция вида применительно к современ-
ным организмам дуалистична, поскольку она основывается как
на морфологических критериях, так и на критерии скрещивае-
мости. Такой дуализм не создает никакой проблемы вида в
группах, в которых эти два критерия совпадают Но не во всех
группах дело обстоит так благополучно. Какому же критерию
следует отдавать предпочтение? Среди биологов нет единодуш-
ного мнения по этому вопросу. Представители различных школ
отвечают на него по-разпому, и их ответы отражают различную
практику, которой они придерживаются
Развитие концепции вида, начавшееся во времена ранних
натуралистов от общей исходной точки, в дальнейшем пошло
по разным направлениям Одно из таких направлений привело
к развитию концепции вида, принятой в таксономии. Другое
частично независимое направление привело к созданию совре-
менной биологической концепции вида. В результате при ис-
пользовании термина вид в таксономии и в популяционной био-
логии в него вкладывают различный смысл.
Во многих дискуссиях по проблеме вида не проводится раз-
личия между этими двумя употреблениями термина. Нередко
биологическую концепцию вида критикуют с позиций таксоно-
мической концепции, не признавая различий в точках зрения.
Надо ли объяснять, что подобные дискуссии лишь способствуют
сохранению путаницы.
Делались также попытки упорядочить положение, ограничив
применение термина вид и используя его лишь в одном из его
традиционных значений. К сожалению, разные авторы расхо-
дятся во мнениях о том, какое именно из этих значений следует
выбрать. Одни авторы [131, 244, 762 и др.] считают, что термин
вид следует закрепить за формальной таксономией, а для ре-
94 Часть вторая. Природа видов
продуктивно изолированных скрещивающихся групп создать
новые термины. Другие [100, 279 (но не [282]), 525 и др.] пред-
почитают сохранить термин вид для изолированных скрещиваю-
щихся групп. Ни одно из этих противоположных предложений
принято не было, и вряд ли то или другое из них найдет широ-
кое признание, поскольку обе школы исследователей пренебре-
гают исторически сложившимися традициями в применении
термина вид.
Выход из этого семантического тупика, позволяющий достичь
желанной цели —внести ясность в обсуждения, избежав при
этом опасности пойти наперекор издавна установившимся тра-
дициям в использовании термина, — состоит в том, чтобы при-
нять термины «биологический вид» и «таксономический вид»,
вкладывая в них различный смысл (79, 157, 282, 652]. Целесо-
образность разграничения этих двух типов видов была доказана
при изложении и обсуждении проблемы в этой и в предшест-
вующих главах. Задумайтесь еще раз над видами-двойниками
в свете этого обсуждения. Каждый из видов-двойников по от-
дельности представляет собой хороший биологический вид и
может быть признан за таковой исследователями, интересую-
щимися его генетическими или экологическими характеристика-
ми. В то же время группу видов-двойников в целом при обычной
“идентификации можно рассматривать как один таксономический
1ВИД.
Критика биологической концепции вида
Многие авторы предлагали концепции вида, совершенно не сов-
местимые с биологической концепцией (см., например, Беннет
{56]; Бурма [76]; Гейтс [236];* Дэвидсон [155]; Кронквист [130];
Левин [475]; Мейсон [558]; Москуин [587]; Рейвн (660, 661]; Со-
кал, Кровелло [754]; Эрлих, Холм [195]; Эрлих, Рейвн [196].
Некоторые из этих работ открыто направлены против биологи-
ческой концепции вида (Беннет, Кронквист, Москуин, Рейвн,
Сокал и Кровелло, Эрлих). Кроме того, некоторые работы, на-
правленные против биологической концепции вида, часто с одоб-
рением цитируют другие авторы (см., например, Орндаф [631]
о работе Сокала и Кровелло [754] и многие другие).
Перечисленные выше авторы в большинстве своем ботаники;
юдин (Бурма)—зоолог, публикующий свои работы в ботаниче-
ском журнале; два (Эрлих, Сокал) — зоологи, сотрудничающие
с соавторами-ботаниками. Несогласие с биологической концеп-
цией вида или возражения против нее высказывают, как показы-
вает этот перечень, в основном ботаники, но также и отдельные
зоологи.
Гл 7. Разные типы видов
95
В этом разделе мы рассмотрим некоторые из основных воз-
ражений против биологической концепции вида и попытаемся
ответить на них.
Авторы старшего поколения (Бурма, Гейтс, Дэвидсон, Мей-
сон) отстаивают номиналистическую концепцию вида' реальны
лишь индивидуумы, а виды — это просто скопления индивиду-
умов. Подобная точка зрения лишена здравого смысла приме-
нительно к бипарентальным организмам, у которых скрещиваю-
щиеся популяции действительно представляют собой реально
существующие репродуктивные единицы и группируются в более
обширные популяционные системы.
Авторы более поздних работ (Рейвн, Сокал и Кровелло,
Эрлих и Холм, Эрлих и Рейвп) подчеркивают важность скре-
щивающейся популяции, но не принимают биологический вид.
Эти авторы правы по первому вопросу. Но на границе между
популяционными системами действительно существуют барь-
еры, препятствующие скрещиванию между этими популяциями.
Точки зрения упомянутых выше авторов можно придерживаться
только в том случае, если пренебречь этим вторым аспектом или
принизить его значение
Сокал и Кровелло [754] рассмотрели поэтапно весь процесс,
необходимый для распознавания биологического вида, подверг-
нув его логическому анализу. Они приходят к выводу, что био-
логической концепцией вида трудно пользоваться па практике.
Совершенно верно, что для выделения биологического вида в
любой данной группе необходимо провести много полевых и экс-
периментальных исследований, а также проделать большую ра-
боту с музейными коллекциями. Для некоторых групп растений
такая работа проведена. Сокал и Кровелло [754], подчеркивая
эти трудности, совершенно умалчивают о том, что на самом де-
ле все необходимые операции в ряде случаев были проведены.
Левин [475], Рейвн [660], Сокал [753], Эрлих и Рейвп [196]
считают, что интеграция обширных популяционных систем видо-
вого масштаба не может быть результатом обмена генами, как
полагают другие ученые, поскольку этот процесс носит локали-
зованный характер. Вполне возможно, что это справедливо для
некоторых групп организмов, в том числе для многих растений
(обсуждение этого вопроса см. в гл. 4). Мы здесь будем исхо-
дить из допущения, что оно справедливо Затем названные выше
авторы делают заключение, что недостаточная интегрирован-
ность, обусловленная слишком обширным потоком генов, подры-
вает статус биологического вида как эволюционной единицы.
Мы можем согласиться с предпосылкой, но не с выводом.
Биологический вид состоит из действительно или потенциально
скрещивающихся между собой индивидуумов, репродуктивно
изолированных от других таких видов. Широкое скрещивание в
*96 Часть вторая. Природа видов
пределах данной популяционной системы не представляет со-
‘бой существенной особенности биологического вида; существен-
ное значение имеет его нескрещиваемость с другими популяци-
онными системами.
Многие ботаники полагают, что, согласно биологической кон-
цепции вида, все или почти все растения должны быть сгруп-
пированы в дискретные видовые единицы. Это не реализуется
во многих группах растений и в разных флорах, и такое несоот-
ветствие между наблюдением и теорией вызывает глубокое не-
довольство биологической концепцией вида. Подобное отноше-
ние к этой концепции не высказывается прямо в публикуемых
работах, и поэтому его нельзя документально подтвердить, как
это обычно принято. Тем не менее оно существует; оно прояв-
ляется при неофициальных обсуждениях и подразумевается в
некоторых опубликованных материалах [130, 660, 754, 756].
Вот еще один пример того, как в биологическую концепцию
вида вкладывают большее значение, чем это нужно. Биологиче-
ский вид представляет собой одну из стадий процесса диверген-
ции, как уже отмечалось в этой главе; существуют также и
другие стадии незавершенного видообразования и вторичного
слияния видов. Следовательно, ряд популяционных систем в
любой данный момент времени состоит как из биологических
видов, так и из полувидов. В сущности, скорее всего следует
ожидать, что в каждый данный момент своей истории не все
популяции будут сгруппированы в дискретные биологические
виды. Важно, что в каждый момент существует некоторое чис-
ло биологических видов.
Одна из характерных черт биологических единиц вообще со-
стоит в том, что процессы их размножения постепенны и тре-
буют значительного времени. Деление одной хромосомы, клетки
или особи на две дочерние хромосомы, клетки или особи прохо-
дит через ряд промежуточных стадий. Тем не менее никто не
считает хромосомы, клетки и особей, после того как они образо-
вались, произвольно выделенными единицами организации. Точ-
но так же биологический вид в процессе подразделения на два
или несколько новых видов может проходить через промежуточ-
ные стадии, и это отнюдь не должно наносить ущерб статусу
дочерних видов как реально существующих единиц па их ко-
нечных и хорошо развитых стадиях.
Общая особенность всех работ, перечисленных в начале это-
го раздела, — поиски единой универсальной концепции вида.
В них обсуждаются «виды» без всяких дальнейших уточнений.
Авторы работ, направленных против биологической концепции
вида, пытаются использовать эту концепцию в качестве удов-
летворительного универсального критерия вида и обнаружи-
вают, что она для этого не подходит. Однако, как мы отмечали.
t Гл. 7. Разные типы видов. 97
рцнее, единственный критерий вида, имеющий универсальное
применение, — это практичность и удобство.для идентификации
и классификации, и этот критерий дает нам таксономическую
единицу, которая в своей основе искусственна, и часто ле соот-
ветствует естественным единицам.
Для того чтобы сделать возможным полезный критический
диалог о виде, необходимо проводить различие между сущнос-
тями, называемыми здесь таксономическими видами и биологи-
ческими видами. Эти сущности перекрываются, но они в корне
различны. Однако в некоторых статьях, направленных против
биологической концепции вида, открыто отвергается как рас-
хождение между таксономическим и биологическим видами, так
и сами эти термины (см., например, [76]); в других же расхож-
дение просто игнорируется, а термин «вид» используется попе-
ременно в разных значениях.
Я мог бы привести многочисленные примеры работ, в кото-
рых термин «вид» без прилагательных используется в смысле
таксономического вида в одной части предложения или абзаца
и в смысле биологического вида в другой части того же самого
абзаца. Такие колебания между двумя различными концепция-
ми вида во многих случаях, по-видимому, не осознаны, но они
всегда служат источником недоразумений. Всякая логически
обоснованная критика биологической концепции вида должна
признавать различие между биологическим и таксономическим
видом.
7—45
Часть третья
Генетические и экологические
аспекты вида
8 Генетическая основа
различий между видами
Изучение наследственности при межвидовых и межрасовых
скрещиваниях проводится многими исследователями во многих
группах растений. Как показали результаты этих работ, близ-
кие виды или расы дифференцированы в отношении систем ге-
нов, различных типов хромосомных перестроек и систем сцеп-
ления. Поскольку обзор соответствующих данных уже опубли-
кован нами в другой книге [294], здесь мы ограничимся лишь
кратким их изложением.
Генные системы
Генные системы, участвующие в дифференциации видов у рас-
тений, можно классифицировать следующим образом. По систе-
мам каждого из перечисленных ниже типов имеются эмпириче-
ские данные (см. [294]).
1. Простые менделевские системы.
2. Сериальные генные системы, состоящие из двух или
нескольких генов, контролирующих последовательные
этапы какого-либо метаболического процесса или про-
цесса развития.
3. Системы множественных генов, состоящие из генов с
аддитивным действием.
4. Системы оппозиционных генов, состоящие из множест-
венных генов и (или) сериальных генов в сочетании с
контролирующими генами, обладающими негативным
действием.
5. Системы модификаторов, состоящие из главных генов
и генов-модификаторов.
6. Цитоплазматические факторы.
7. Системы, состоящие из взаимодействующих друг с дру-
гом цитоплазматических и хромосомных генов.
Простые менделевские системы встречаются не очень часто,
но они существуют. Хорошими примерами служат плодовитые
Гл 8 Генетическая основа различий между видами 99
межвидовые скрещивания у Aquilegia [648]. Цветки большинст-
ва видов имеют шпоры, но у примитивного вида A. ecalcarata
шпоры отсутствуют. Признак наличия или отсутствия шпор на-
следуется как различие, определяемое одним геном, причем на-
личие шпор — доминантный признак при скрещиваниях между
A. ecalcarata и A. vulgaris, а также между A. ecalcarata и
A. canadensis. Различие по тому же признаку наследуется как
определяемое двумя генами и при скрещиваниях A. ecalcarataX
ХА. chrysaniha и A. ecalcarataxA. alpina дает в F2 соотноше-
ние 15:1. Различие в ориентации цветка также определяется
одним геном, и при скрещиваниях A. ecalcarataXA. chrysantha
поникающие цветки доминируют над прямостоячими [648].
Преобладающая часть различий при межрасовых и межви-
довых скрещиваниях у растений наследуется как признаки, обус-
ловленные множественными генами (т. е. соответствует типам
2—5 из приведенного выше перечня). Так обстоит дело у Agui-
legia. У этой группы растений, как и у большинства других
групп, окраска цветков наследуется очень сложным образом.
А межвидовые различия по высоте растений, форме листьев,
длине шпор, величине лепестков и т. п. детерминируются у
Aquilegia множественными генами [648].
В простой системе множественных генов, контролирующих
какой-либо количественный признак, гены обладают аддитивным
действием и доминирование отсутствует. Это приводит к возник-
новению промежуточного фенотипа в Fi, непрерывной изменчи-
вости в широком диапазоне и промежуточной моды в F2 и не-
прерывной изменчивости в более узких пределах в последующих
поколениях. Классическим примером такой системы служит
наследование различий в длине венчика при скрещиваниях меж-
ду разновидностями Nicotlana longiflora [182] (рис. 8 1).
Вряд ли следует говорить, что взаимодействия между гена-
ми обычно гораздо сложнее, чем в приведенном примере. Так,
компоненты какой-либо системы множественных генов у данной
расы или вида могут обладать не только одним лишь аддитив-
ным действием на тот или иной количественный признак; одни
компоненты системы могут оказывать на развитие данного при-
знака аддитивные эффекты, а другие — негативные. Когда две
такие внутренне сбалансированные оппозиционные системы ге-
нов сочетаются в результате межрасовой или межвидовой гиб-
ридизации, то в поколениях F2, а при некоторых условиях и у
гибридов Fi, наблюдается трансгрессивное расщепление, причем
количественное выражение данного признака у гибридов выхо-
дит за пределы его изменчивости у обоих родительских типов.
В сериальных генных системах отдельные гены детермини-
руют различные этапы какого-либо процесса в развитии. Такие
системы играют важную роль в наследовании окраски цветков.
7*
100
Часть третья. Генетические и экологические аспекты вида
Рис 8 1. Распределение по частоте индивидуумов с разной длиной венчика
при скрещиваниях между различными разновидностями Nlcotiana longiflora
(по [182], с упрощениями).
При изучении генетической основы окраски цветков обычно об-
наруживается, что ее наследование определяется комплементар-
ными генами, т. е. один ген создает условия, необходимые для
действия другого гена, контролирующего следующий этап. В на-
следовании окраски цветков нередко участвуют также ингиби-
торы, роль которых состоит в оппозиционном взаимодействии с
сериальной генной системой.
Рассмотрим, например, различие между двумя близкими ви-
дами Viola, выражающееся в наличии или отсутствии одной из
особенностей в окраске цветка: для цветков V. tricolor харак-
терно темное пятно, обычно отсутствующее у V. arvensis. Кла-
узен [95, 96] показал, что у обоих видов имеются комплементар-
ные гены, определяющие образование этого пятна. Но V. arven^
Гл. 8 Генетическая основа различий между видами 101
sis содержит, кроме того, один или два гена-ингибитора, подав-
ляющие образование пятна. У потомков от скрещивания между
этими видами происходит сложное расщепление по этому при-
знаку [95].
Системы оппозиционных генов, состоящие из генов с про-
тивоположными, положительными и отрицательными воздейст-
виями на развитие данного признака, встречаются часто. О ши-
роте их распространения можно судить по результатам изуче-
ния наследования 16 многофакторных признаков у межрасовых
гибридов Potentilla glandulosa. Семь или даже восемь из этих
признаков контролируются системами оппозиционных генов
[98].
Классический пример межвидовых различий в системах мо-
дификаторов наблюдается у видов Gossypium hirsutum и G bar-
badense. Многие признаки этих двух близкородственных видов
определяются гомологичными генами. Наличие или отсутствие
пятен на лепестках, например, контролируется у каждого из
этих видов парой гомологичных аллелей гена S. При скрещива-
ниях внутри каждого вида наличие пятна доминирует над его
отсутствием, и в F2 эти признаки расщепляются в отношении
3:1. Но при межвидовых скрещиваниях между растениями с
пятнами и без пятен потомки Fi носят промежуточный харак-
тер, а в F2 наблюдается непрерывная изменчивость. Объясня-
ется это тем, что хотя главные гены, определяющие наличие или
отсутствие пятна на лепестках (и различные другие признаки) ,
у этих двух видов хлопчатника гомологичны, они несут различ-
ные наборы генов-модификаторов. Система модификаторов у
G. barbadense усиливает проявление признака пятна, а соответ-
ствующая система у G. hirsutum подавляет его [349].
Интересный тип взаимодействия генов наблюдается в тех
случаях, когда у потомков межвидовых гибридов появляются
совершенно новые и непредсказуемые признаки. Для Antirrhi-
num majus и A. glutinosum характерны типичные двугубые цвет-
ки; такие же цветки имеются у гибридов Fi между этими вида-
ми и у большинства потомков F2, однако у некоторых индивиду-
умов F2 цветки больше похожи на цветки их далекого родича,
относящегося к роду Rhinanthus [519]. Точно так же у Digitalis
при одном из межвидовых скрещиваний у некоторых потомков
появляется новый признак цветка, а именно шпоры, отсутствую-
щие у родительских видов [715].
Новые фенотипы наблюдаются также у гибридных потомков
ряда других групп растений, а не только у Antirrhinum и Digita-
lis. Следовательно, это явление носит общий характер, оно по-
лучило название макрорекомбинации (краткий обзор с библио-
графическими ссылками см. [294]). Макрорекомбинации имеют
эволюционное значение, поскольку они показывают, что меж-
102 Часть третья. Генетические и экологические асЛекты вида
видовая гибридизация может порождать не только, промежуточ-
ное формы, но также формы, не похожие ни на один из роди-
тельских видов.
Гены, кодирующие ферменты
Электрофорез позволяет выявить те генетические различия в
ферментах, которые никак не проявляются фенотипически. Эти
различия могут иметь место на любом уровне организации: на
уровне индивидуума у гетерозигот или полиплоидов; между от-
дельными особями; в пределах одного вида; между видами и
между таксонами более высокого ранга. Гены, определяющие
электрофоретическую изменчивость, отличаются от генов, кото-
рые определяют большинство внешних фенотипических призна-
ков.
В последние годы методы электрофореза используются чрез-
вычайно широко, в связи с чем накопилось огромное количество
данных по электрофоретической изменчивости как у животных,
так и у растений. Здесь мы можем лишь в общем виде коснуть-
ся этой большой проблемы. Появилось несколько новых обзоров
по электрофоретической изменчивости у растений [255, 335, 412];
есть также другие ценные обзоры, посвященные главным обра-
зом, но не исключительно, животным [28, 508].
При исследовании 13 ферментов в 6 популяциях Tragopogon
dubiuSi 3 популяциях Г. porrifolius и 3 популяциях Г. pratensis
полиморфизм по этим ферментам был обнаружен у первых двух
видов. У всех видов наблюдалось большое сходство между по-
пуляциями, о чем свидетельствуют показатели генетической
идентичности, равные 0,97—1,00. Вместе с тем между этими
тремя видами во всех попарных сочетаниях существуют четко
выраженные различия, а показатели генетической идентичности
колеблются от 0,50 до 0,62 [694].
У очень близкородственных видов Clarkia biloba и С. Ungu-
lata показатель генетической идентичности высокий (7=0,88).
В отличие от этого у менее близкородственных видов С. rubicun-
da и-С. franciscana показатель генетической идентичности ниже
(7=0,28) [252—255].
При сравнении Phaseolus vulgaris и Ph. coccineus по двум
ферментным системам — эстеразы и лейцинаминопептидазы —
методом электрофореза между ними' были обнаружены значи-
тельные различия в положении полос этих ферментов. У алло-
полиплоидного гибрида, полученного в результате скрещивания
этих видов, были обнаружены все полосы, образуемые родитель-
скими видами, что свидетельствует об аддитивности в наследо-
вании этих ферментов [884]. Такая аддитивность у аллополипло-
идов гибридного происхождения — явление обычное. Однако в
Гл. 8. Генетическая основа различий между видами ЮЗ"
некоторых случаях аддитивности не наблюдается на электро-
фореграмме аллополиплоида отсутствуют полосы, имеющиеся
у родительских видов, или, напротив, появляются новые полосы,
как, например, у искусственных аллополиплоидов Nicotiana
[669]. В таких случаях у межвидовых гибридов и у аллополи-
плоидов, по-видимому, имеют место те или иные сложные взаи-
модействия между генами.
Замечания относительно регуляции
В периодической литературе довольно широко обсуждается
вопрос о генах-регуляторах и регуляции развития как ключевых
элементах в дифференциации видов и в процессе видообразова-
ния (см., например, [77, 440, 896]).
Регуляция развития, несомненно, играет важную роль в соз-
дании фенотипических различий между видами. Это давно приз-
нано, и не об этом здесь речь.
Обсуждение рассматриваемого вопроса в приведенных выше
и в других недавно опубликованных статьях носит особый ха-
рактер В них регуляцию относят за счет действия генов-регу-
ляторов и побочных эффектов хромосомных перестроек. Здесь
нас интересуют гены-регуляторы, а хромосомные перестройки
будут рассмотрены в этом аспекте в гл. 14. Гены-регуляторы
противопоставляют структурным генам, а структурный ген —
это, в сущности, то же самое, что и классический менделевский
ген, рассматриваемый в несколько ином плане. Выдвигаемое
авторами упомянутых работ положение состоит в том, что эво-
люция «зависит главным образом от мутаций генов-регуляторов
и лишь во вторую очередь — от изменений структурных генов»
[896].
Разграничение структурных и регуляторных генов возникло
в результате изучения бактерий, и его стали широко применять
также к высшим организмам, где оно неуместно. У высших рас-
тений нет ни одного класса генов-регуляторов. Регуляция обес-
печивается гетерогенной совокупностью контролирующих генов.
В эту совокупность входят гены-ингибиторы, минус-модифика-
торы, системы оппозиционных генов, гетерохроматиновые блоки
и контролирующие элементы (которые, возможно, представляют
собой гетерохроматиновые блоки) (см. [294]). Кроме того, у
высших растений разграничение между структурными и регуля-
торными генами оказывается несостоятельным: некоторые типы
генов-регуляторов на самом деле представляют собой структур-
ные гены, как это хорошо видно на примере генов-ингибиторов.
Итак, изменения в характере развития играют важную роль
в создании межвидовых различий у высших растений. Но эти
104
Часть третья. Генетические и экологические аспекты вида
изменения нельзя связать с генетическими элементами какого-
либо одного типа. Они представляют собой результат действия
генов и генных взаимодействий многих типов.
Хромосомные перестройки
Структурные перестройки хромосом возникают в результате
разрывов хромосом и последующего воссоединения разорванных
концов каким-либо иным способом. В зависимости от числа раз-
рывов, их местоположения и способа воссоединения различают
несколько типов перестроек, главные из которых представлены
на рис. 8.2.
Во многих группах растений при цитологическом изучении
близких видов и их гибридов Fi обнаруживается, что эти виды
различаются по инверсиям, транслокациям и другим структур-
ным перестройкам. В качестве иллюстрации здесь будут описа-
ны два классических примера.
Эвери {25] провел исчерпывающий цитогенетический анализ
гибридного треугольника Nicotiana, состоящего из видов ЛА. ala-
ta, N. langsdorffii и N. bonariensis. У этих видов имеется по
9 пар хромосом, относящихся к разным классам в зависимости
от величины и морфологии в пределах данного набора и до не-
которой степени—между наборами, принадлежащими разным ви-
дам. В мейозе у гибридов Fx от скрещивания N. alata и N. langs-
dorffii хромосомы Ль Bi и D образуют цепочку, что указывает
на существование между родительскими видами различий по
двум последовательным транслокациям в этих хромосомах.
Картина конъюгации при скрещивании N. alata и N. bonaridn-
sis показывает, что эти виды различаются по реципрокной
транслокации в хромосомах Ах и Е3 и по двум последователь-
ным транслокациям в хромосомах А2, В2 и Е3. На основании
этих данных можно предсказать, что N. langsdorffii и ЛА. bona-
riensis дифференцированы по ряду транслокаций, затрагиваю-
щих все хромосомы в их наборах, за исключением хромосом С
и Е2. Картины конъюгации, наблюдаемые у гибридов, получен-
ных в результате скрещивания ЛА. langsdorffii и N. bonariensis,
подтверждают это предсказание [25].
Столь же остроумный цитогенетический анализ видов Lilium
martagon, L. hansonii и гибрида Fj между ними провел Ричард-
хон [682]. У этих двух видов лилий по 12 пар хромосом. Карти-
на конъюгации хромосом у гибрида показывает, что родитель-
ские виды различаются по небольшим парацентрическим ин-
версиям в шести из этих двенадцати -хромосом [682].
Это относительно простые и ясные примеры. В большинстве
случаев, виды , различаются по многочисленным невыявленным
1.Терминальная нехватка Ст. ABCD Изм. ABCD EFGHI 4 Парацентрическая инверсия
Ст. Изм ABCD EFGHI
t EFG ABCD °“ t t EFGHI
2.Интерстициальная делеция 5 Перицентрическая инверсия
Ст. ABCD л EFGH I Ст ABCD EFGHI
Изм ABCD I1 ' EFI Изм. t ABGFE G t DCHI
3. Дупликация 6 Транспозиция
Ст. ABCD EFGHI Ст. ABCD yy^-ghi >
Изм. ABCD f t EFGHGHI Изм. t f AD t . EFBCGHI
7.Небольшая реципрокная транслокация
Ст. ABCD EFGHI MNOP QRST
' -и- ИЗМ. ABCD t EFGHST ° 1 MNOP n ОЯ1
8. Транслокация плеча
Ct ABCD EFGHI MNOP QRST
Изм. ABCD 't QRST MNOP EFGHI
^Последовательные транслокации
Ст. ABCD EFGHI MNOP t QRST uvw WIL
1-я транслокация ABCD 3t QRST MNOP EFGHI UVW' XYZ
транслокация ABCD , QRST MNOP . . XY2 uvw л EFGHI
Рис. 8.2. Типы хромосомных перестроек [283].
В каждом случае стандартная хромосома (ст) сравнивается с хромосомой,
изменившейся в результате перестройки (изм.). Стрелками показаны точки
разрыва на стандартной хромосоме, при которых может возникнуть данная
перестройка. Центромера изображена в виде овала, располагающегося в се-
редине или почти в середине хромосомы.
106 Часть третья. Генетические и экологические аспекты вида
перестройкам, вызывающим аберрации при конъюгации в
мейозе.
Во многих родах растений были обнаружены межвидовые
•различия, обусловленные структурными перестройками хромо-
сом. Такие перестройки часто встречаются в группах травяни-
стых растений, особенно у однолетних. Хорошо известные при-
меры наблюдаются у Brassica, Clarkia, Crepis, Elytnus, Galeop-
sis, Gossypium, Gilia, Layia, Madia и Nicotiana.
Системы сцепления
Генные системы нередко характеризуются особыми типами
сцепления. Следовательно, межвидовые различия — это разли-
чия не только в генных системах, но также и в наборах сцеп-
ленных генов или в системах сцепления* Известны системы
сцепления разных типов. Помимо обычного сцепления генов в
стандартных хромосомах существуют сцепления, затрагивающие
лишь определенные типы генных систем; примерами служат:
многофакторное сцепление, сцепление М—V, сцепленные функ-
циональные генные системы, сцепленные адаптивные генные
комбинации и сбалансированные полигенные системы.
Две из этих систем имеют отношение к эволюционным про-
цессам, которые будут обсуждаться в последующих главах. Эти
системы кратко рассматриваются ниже. Обзор систем сцепле-
ния в целом можно найти у Гранта [294].
Хромосомные перестройки тесно связаны с системами сцеп-
ления. Хромосомы — это носители обычных для данного вида
групп сцепления. Структурные перестройки — источник измене-
ний обычного сцепления. Некоторые перестройки в гомозиготном
или гетерозиготном состоянии служат носителями систем сцеп-
ления определенных типов.
Многофакторное сцепление
У покрытосеменных растений модальное гаплоидное число хро-
мосом п обычно равно 7—14. Следовательно, если множествен-
ные факторы, определяющие два признака, по которым разли-
чаются две расы или два вида, распределены по набору хромо»
сом более или менее случайным образом, то некоторые из ге-
нов для каждого из этих признаков должны находиться в одной
и той же хромосоме (или хромосомах). Короче говоря, сцепле-
ние между этими двумя признаками должно быть частичным
или многофакторным. Весьма вероятно, что с одним из двух
таких признаков или с ними обоими частично связан таким же
образом третий количественный признак. Продолжая рассуж-
дать в том же направлении, можно ожидать, что все признаки,
Гл. 8. Генетическая основа различий между видами
107
находящиеся под многофакторным контролем, по которым раз-
личаются две расы или два вида, связаны, прямо или косвенно,
системой переплетающихся между собой слабых сцеплений. По-
добная система сцепления не предотвращает, но сильно ограни-
чивает рекомбинацию при межрасовой и межвидовой гибриди-
зации [98, 99, 294].
Эти ожидаемые многофакторные сцепления обнаружены в
нескольких экспериментах по гибридизации Хорошие данные
получили Клаузен и Хиси [99] при межрасовом скрещивании у
Potentilla glandulosa (2п=14). Они изучали корреляции между
14 признаками в обширной расщепляющейся популяции F2; бы-
ло известно, что 11 из этих признаков детерминируются двумя
или несколькими генами. Из 91 пары признаков у 67 наблюда-
лась слабая, но статистически значимая корреляция. Ни один
из признаков не расщеплялся независимо от всех других; на-
против, каждый признак коррелировал со многими или с боль-
шинством других 13 признаков. Корреляции между определен-
ными парами признаков, такими, как длина лепестков и длипа
чашелистиков, возможно, обусловлены плейотропным действием
генов, однако многие другие сочетания признаков, морфогенети-
чески по связанных друг с другом, как, например, опушенность
растения и вес семян, также наследовались совместно, и в этих
случаях следует предполагать наличие многофакторного сцеп-
ления [99].
Аналогичные результаты были получены позднее для ряда
количественных признаков при межвидовом скрещивании Mitnu-
lus lewisii и М. cardinalis [377].
Л4ногофакторное сцепление обусловливает корреляцию меж-
ду признаками в пределах расы или вида, получившую название
генетической связности [99], или связности признаков.
Сцепление М—V
При изучении межвидовых гибридов в нескольких родах рас-
тений были получены данные, свидетельствующие о наличии
сцепления между генами, детерминирующими различные морфо-
логические признаки, и генами жизнеспособности. Это широко
распространенное, а быть может, и всеобщее явление называют
сцеплением М— V [292]. Термин «морфологические гены» отно-
сится в данном контексте к любым генам, определяющим види-
мые фенотипические признаки, в том числе окраску разных
частей растений Под генами жизнеспособности (viability genes)
имеются в виду любые гены, оказывающие влияние на рост и
мощность
Гены жизнеспособности могут быть плейотропными, оказы-
вая действие также на морфологические признаки, и наоборот,
108 Часть третья. Генетические и экологические аспекты вида
как в случае альбиносных проростков. Нам хотелось бы иметь
возможность исключить подобные случаи плейотропии. Поэтому
мы ограничимся рассмотрением таких фенотипических призна-
ков, которые затрагивают цветки или плоды, но не могут быть
выявлены на стадии вегетативного роста и которые коррелиру-
ют с различиями в жизнеспособности. Наблюдаемые в таких
случаях корреляции позволяют предположить скорее наличие
‘Сцепления между отдельными морфологическими генами и те-
лами жизнеспособности, а не эффекты плейотропии.
Таким образом, мы здесь имеем дело с данными об аллель-
ных различиях между видами или расами растений по сцеплен-
ным генам М и V. Самая простая из возможных ситуаций —
сцепленная пара генов М—V без существенного взаимодейст-
вия между генами. Один из родительских видов имеет при этом
генотип MaVa/MaVa, а другой — генотип МьУь/МьУь. Допустим,
что генам Va и Уь соответствуют различные оптимальные для
роста условия среды. В таком случае у потомков гибрида, выра-
щиваемых в среде, благоприятствующей одному из аллелей V,
соотношение фенотипов при многофакторном расщеплении по
морфологическому маркеру М будет отклоняться от нормально-
го. Различные белее сложные примеры с участием частичного
или многофакторного сцепления между двумя или несколькими
генами V и двумя или несколькими генами М дают сходные ре-
зультаты [292].
Интерфертильные виды Mimulus cardinalis и М. lewisii
(Scrophulariaceae) встречаются соответственно на небольших
и значительных высотах в горах Калифорнии. Проростки одной
из растущих на побережье линий М. cardinalis хорошо выжи-
вают в саду на низменности (Станфорд), но в'альпинарии
(Тимберлайн) смертность среди них очень велика. Сеянцы вы-
сокогорной расы М. lewisii, напротив, хорошо выживают в Тим-
берлайне, но в Станфорде среди них наблюдается высокая
смертность [615].
Эти два вида различаются также по окраске цветков — розо-
вой у Mimulus lewisii и оранжевой у М. cardinalis. Окраска
цветков наследуется как различие по одному гену, причем розо-
вая окраска доминирует. Нобс и Хиси [616] обнаружили, что
среди потомков F2 от межвидового скрещивания при вымащива-
нии в условиях, в общем благоприятных для них, соотношение
растений с розовыми и оранжевыми цветками составляло при-
мерно 3:1.
Однако при выращивании в Тимберлайне соотношение розо-
вых и оранжевых цветков в F2 составило 8,7 к 1, а в одной пов-
торности F2, выращенной летом в Станфорде, было получено со-
отношение 1,7 к 1. Затем Нобс и Хиси сравнивали соотношения,
полученные в одном и том же саду в Станфорде зимой, т. е. в
Гл. 8 Генетическая основа различий между видами 109
прохладную погоду, и жарким летом. В зимней популяции Fa
соотношение розовых и оранжевых цветков составило 4,4 к 1,0,
а в летней— 1,7 к 1,0 [616].
Такие измененные соотношения фенотипов сопровождались
в каждом случае высокой гибелью проростков. Смертность, оче-
видно, была вызвана избирательной элиминацией типов cardina-
lis в альпийской зоне или в прибрежной полосе зимой и типов
lewisii летом у побережья. Кроме того, гены определяющие об-
щую физиологическую адаптацию проростков к условиям кли-
мата, должны быть сцеплены с геном окраски цветков, для то-
го чтобы получались наблюдаемые отклонения в соотношениях
фенотипов [616].
Сходные данные были получены при межрасовом скрещива-
нии у Potentilla glandulosa и при межвидовых скрещиваниях у
Gossypium, Lycopersicon, Phaseolus и Oryza Эти данные позво-
ляют считать, что изучавшиеся расы и виды несут разные алле-
ли сцепленных генов М и V [98, 292, 447, 448, 683, 708, 800, 801].
Во многих группах растений удается определить гены, содер-
жащиеся в гомологичных хромосомах у интерфертильных видов
или рас, однако нам почти ничего не известно о генетических
факторах, относящихся к тем затронутым перестройками уча-
сткам, по которым различаются хромосомы интерстерильных
видов. В самом деле, к изучению этой проблемы обращались
редко ввиду того, что наложение на генетический анализ ос-
ложняющих эффектов гибридной стерильности делало ее почти
неразрешимой Тем не менее для нескольких групп растений мы
теперь располагаем некоторыми предварительными сведениями
относительно того, какие гены локализованы в перестроенных
участках хромосом [287].
Эти сведения получены в результате экспериментов по гиб-
ридизации между парами видов Clarkia [329, 378, 379], Gossy-
pium [238, 241, 680, 800, 801], Triticum [219, 534, 716, 718], Zea
[551, 829], Gilia [287] и Melilotus [705]. В каждом из этих слу-
чаев данные носят косвенный характер, и анализ их сложен
Здесь можно подвести итоги только по главным моментам; бо-
лее полный обзор содержится в работах Гранта [280, 287].
Полученные данные свидетельствуют о том, что в каждой из
этих пяти групп растений морфологические гены и гены жизне-
способности лежат в участках, затронутых перестройкой. Рас-
смотрим в качестве примера результаты, полученные при гибри-
дизации видов Gilia.
Gilia malior и G. modocensis различаются по многим струк-
турным перестройкам, чем и объясняется в значительной мере
высокая стерильность их гибридов Fi. Эти виды различаются
также по генам жизнеспособности, что ведет к значительному
разрушению гибридов в F2—F6, и по многим морфологическим
ПО Часть третья. Генетические и экологические аспекты, вида
признакам. Отбор среди гибридных потомков на жизнеспособ-
ность в условиях среды, благоприятных для типов modocensis,
и отбор на плодовитость привели к возникновению трех мощ-
ных и плодовитых линий, из которых две гомологичны по струк-
турным перестройкам и плодовиты при скрещиваниях с G. mo-
docensis. Объяснить этот результат очень трудно, если не пред-
положить, что отдельные гены жизнеспособности находятся в
некоторых из участков, затронутых перестройками. Полученные
таким образом жизнеспособные линии были сходны с G. modo-
censis по большинству своих морфологических признаков, а это,
по-видимому, означает, что некоторые морфологические гены
сцеплены с генами жизнеспособности в одних и тех же пере-
строенных участках [288].
Таким образом, мы располагаем данными о том, что в не-
скольких группах растений в затронутых перестройкой участках
хромосом локализованы сцепленные между собой морфологиче-
ские гены и гены жизнеспособности. Вероятно, между система-
ми сцепления M—V у видов, гомологичных по хромосомам, и
у видов, различающихся по структурным перестройкам, нет ка-
ких-либо глубоких различий.
Интегрированный генотип
Как мы убедились, каждая часть данного организма или любой
из его количественных признаков — результат морфогенетичес-
ких процессов, контролируемых системой генов. Отсюда сле-
дует, что для нормального развития данного органа или призна-
ка необходимо гармоничное взаимодействие отдельных компо-
нентов такой системы. Кроме того, для успешного функциониро-
вания организма в целом необходима координация между его
отдельными частями и признаками. А поэтому различные систе-
мы генов, принадлежащие данному генотипу, подвергаются от-
бору на гармоничное взаимодействие друг с другом.
Функциональное и селективное значение гармоничных взаи-
модействий между генами понимали многие исследователи
(Bridges. [68] и другие). «Природные виды слагаются из инди-
} видуумов, гены которых находятся в состоянии внутреннего
равновесия, с тем чтобы обеспечивать гармоничное физиологи-
ческое и морфологическое развитие одного поколения за дру-
гим» [102J. Каждый вид обладает не только внутренним генети-
ческим равновесием, но и собственной, отличной от другой, адап-
тивной комбинацией генов. Как внутреннее генетическое равно-
весие, так и адаптивная комбинация генов могут быть
разрушены в результате межвидовой гибридизации.
В F2 и в последующих поколениях потомков межвидовых'
скрещиваний обычно бывает немало карликовых и стерильных
Гл. 8. Генетическая основа различий между видами 111
индивидуумов, индивидуумов с пониженной жизнеспособностью
или с какими-либо иными отклонениями. Растения Gilia malior,
G. modocensis и их гибриды Fi вполне жизнеспособны. Но в по-
колениях F3 — Fe 42% проростков были слабыми, чахлыми или
полулегальными; можно, кроме того, добавить к этому значи-
тельную гибель вследствие неспособности семян к прорастанию
[289]. Дисгармоничные взаимодействия между генами разных
видов могут также проявляться в F>. Хорошим примером служат
краснолистные карликовые гибриды, обнаруженные среди жиз-
неспособных растений в Fi от скрещивания Gossypium arboreum
и G. hirsutum. Такое полулегальное состояние обусловлено
комплементарным действием двух генов, и имеющихся
у некоторых линий родительских видов [239].
Иными словами, виды различаются по своим системам гене-
тического равновесия и по своим адаптивным комбинациям ге-
нов.
9 Изолирующие механизмы
Основные формы изоляции
Гибридизация и обмен генами между видами предотвращаются
или интенсивность этих процессов снижается в результате су-
ществования многочисленных и разнообразных изолирующих
механизмов. Эти механизмы можно разделить на три основных
класса: 1) пространственные, 2) средовые, 3) репродуктивные.
Классификация изолирующих механизмов представлена в
табл. 9.1. Изоляция у растений довольно подробно рассматри-
вается в ряде обзоров [282, 472, 780].
Пространственная, или географическая, изоляция сущест-
вует между любыми двумя аллопатрическими видами растений,
географические ареалы которых разделены разрывами, превы-
шающими обычный радйус распространения их пыльцы или се-
мян. Так, например, обстоит дело с Platanus occidentalis и
Р. orientalise которые распространены соответственно в восточ-
ной части США и в восточной части Средиземноморской области
и которые, несмотря на интерфертильность, очевидно, не могут
скрещиваться между собой в природных условиях [780].
Средовая, или экологическая, изоляция возникает в резуль-
тате дифференциации видов по их экологическим предпочте-
ниям. Благодаря этой дифференциации виды, сосуществующие
в одной и той же географической области, занимают в ней раз-
личные местообитания. Если между этими видами происходит
гибридизация и если их местообитания обособлены, то большая
часть их гибридных потомков не сможет выжить ввиду отсутст-
вия среды промежуточного или рекомбинационного типа -f 14].
Так, Salvia apiana и S. mellifera, которые экологически диффе-
ренцированы, но симпатричны на юге Калифорнии и которые
способны в ограниченной степени скрещиваться друг с другом,
не гибридизируют в ненарушенной природной среде, хотя иногда
образуют скопления гибридов в районах, где природные усло-
вия были искусственно изменены [17, 202, 301].
Репродуктивная изоляция создается в результате блокиро-
вания обмена генами между популяциями, а блокирование бы-
вает обусловлено генетическими различиями в строении репро-
дуктивных органов и в биологии размножения или во взаимо-
отношениях по плодовитости. Барьеры, препятствующие скре-
щиванию, могут быть внешними, действующими до перекрестного
опыления, как в случае временной, механической или этоло-
гической изоляции; или же они могут бьяъ внутренними, дейст-
вующими после перекрестного опыления, как в случае несов-
местимости, нежизнеспособности, стерильности и разрушения
гибридов. Барьеры, обусловленные несовместимостью между
Гл. 9. Изолирующие механизмы
113
Таблица 9.1. Классификация изолирующих механизмов
у растений
I Пространственные механизмы
1 Географическая изоляция
II. Средовые механизмы
2. Экологическая изоляция
III Репродуктивные механизмы
А. Внешние
3. Временная изоляция
а) сезонная
б) суточная
4. Механическая изоляция 1 Цветковая
5. Этологическая изоляция г изоляция
6 Изоляция, обусловленная автогамией J
Б. Внутренние
7. Несовместимость
а) до оплодотворения
б) после оплодотворения
8. Нежизнеснособность гибридов
9. Стерильность гибридов
10 Разрушение гибридов
родительскими видами, можно разделить на действующие до и
после оплодотворения.
Само собой разумеется, что в реальных ситуациях три ос-
новные формы изоляции могут сочетаться различным образом.
Различные виды репродуктивных изолирующих механизмов
также обычно действуют в разнообразных комбинациях.
Внешняя репродуктивная изоляция
Временная изоляция между двумя видами растений создается
в тех случаях, когда их периоды цветения и опыления приходят-
ся на разное время года или суток. Эта изоляция может быть
частичной, если пики цветения двух видов разделены достаточ-
но большим временным интервалом, но периоды цветения пере-
крываются, или же полной, если периоды цветения двух видов
совершенно не перекрываются. Соответствующие примеры мож-
но найти у Pinus, Phlox, Salvia и во многих других группах рас-
тений.
Oenothera brevipes и Ое. clavaeformis (Onagraceae), напри-
мер, растут вместе в пустыне на западе США, цветут одновре-
менно и опыляются одиночными пчелами, главным образом
андренами, однако гибриды между ними редки. Оказалось, что
цветки Ое. brevipes раскрываются до восхода солнца и их по-
сещают пчелы, активные ранним утром, тогда как цветки
Ое. clavaeformis раскрываются в позднее послеполуденное вре-
мя и их посещают пчелы, летающие в конце дня [657J.
8—45
114
Часть третья. Генетические и экологические аспекты вида
Механическая изоляция создается в результате структурных
различий между цветками двух или нескольких видов, препят-
ствующих перекрестному опылению между этими видами или
нарушающих его. Различия в строении цветков, приводящие к
механической изоляции, встречаются главным образом у тех
растений, у которых цветки имеют сложную структуру (напри-
мер, в семействах Orchidaceae, Papilionaceae, Labiatae, Scrophu-
lariaceae и Asclepiadaceae).
Так, Pedicularis groenlandica и P. attollens растут вместе в
горах Сьерра-Невада в Калифорнии, цветут одновременно и
опыляются одними и теми же видами шмелей, но не гибридизи-
руют. Отчасти это обусловлено различиями в структуре цветков
у этих двух видов. Пыльца Р. groenlandica переносится к рыль-
цу на спинке шмелей, а пыльца Р. attollens—на передней поверх-
ности головы [540, 765]. Другие способы механической изоля-
ции выработались между видами Stanhopea, Ophrys, Aquilegia,
Ascleplas, Penstemon, Mimulus, Salvia, Phlox и Cordia (обзор
см. [474]).
Этологическая изоляция, обеспечиваемая особенностями по-
ведения, благодаря которым индивидуумы предпочитают выби-
рать в качестве брачных партнеров представителей собственного
вида, играет очень важную роль в репродуктивной изоляции
у высших животных. Предполагалось, что у растений подобная
изоляция совершенно отсутствует. Тем не менее она встречается
у некоторых растений в качестве вгоричного эффекта видоспе-
цифичного поведения насекомых-опылителей, посещающих цвет-
ки этих растений. Пчелы, бражники и в меньшей степени неко-
торые другие насекомые, посещающие цветки, способны к сен-
сорным восприятиям, которые дают им возможность отличать
друг от друга разные виды цветков по запаху, форме и окрас-
ке; кроме того, они обладают инстинктом постоянства, застав-
ляющим их кормиться преимущественно на цветках одного вида
при ряде последовательных посещений. Это постоянство в посе-
щении цветков ограничивает перекрестное опыление, так что
юно не выходит, в основном или всецело, за пределы одного вида
из симпатрической смеси видов растений, различающихся по
опознавательным знакам цветков [265, 267].
Этологическая изоляция родственных видов, обусловленная
приверженностью пчел к посещению цветков одного вида, на-
блюдается у Pedicularis [765; Grant, неопубликованные данные],
Stanhopea [177], Lythrum [471], Rhinanthus [456], Cercidium
[417] и у некоторых других родов. Недавно была описана это-
логическая изоляция двух видов Anigozanthos (Haemodoraceae)
в западной части Австралии, обеспечиваемая медососами (Ме-
hphagidae) [382].
Гл. 9. Изолирующие механизмы
115
У однолетних растений часто встречаются симпатрические
смеси видов, размножающихся путем неродственного скрещива-
ния и автогамии, или же двух или нескольких преимущественно
автогамных видов. Автогамная система размножения создает
в таких случаях барьер, препятствующий перекрестному опыле-
нию между разными видами. Левин [474] рассматривает это-
как изолирующий механизм, и мы следуем его точке зрения
здесь и в табл. 9.1.
Концепция репродуктивной изоляции приложима главным
образом к организмам, для которых характерны половое раз-
множение и ауткроссинг. Ее можно расширить, с тем чтобы рас-
пространить ее на смеси автогамных видов растений с видами,
размножающимися путем ауткроссипга, а также на смеси двух
или нескольких преимущественно автогамных видов, как мы это
сделали выше; однако она лишена смысла в приложении к
строго унипарентальным группам растений и не должна исполь-
зоваться для таких групп.
Внутренняя репродуктивная изоляция
Несовместимость при скрещиваниях создается разного рода
барьерами, препятствующими развитию жизнеспособного гиб-
ридного зародыша. Некоторые из этих барьеров действуют до
оплодотворения, а другие — после него.
В некоторых случаях пыльцевое зерно не прорастает на
рыльце чужого вида, как при скрещивании Datura meteloid.es
и D stramonium, или же пыльцевая трубка не может достаточно
успешно расти в чужеродном столбике, как при скрещивании
Iris tenax и I. tenuis [25, 740]. Или же, если происходит оплодо-
творение и формируется гибридный зародыш, этот зародыш мо-
жет преждевременно погибнуть вследствие дегенерации гибрид-
ного эндосперма, как это бывает при некоторых скрещиваниях
у Datura, Iris, Gossypium и других родов.
Межвидовые гибриды Fi часто обладают нормальной мощ-
ностью, однако иногда у них наблюдаются различные конститу-
циональные дефекты, известные под общим названием нежизне-
способности гибридов, которая блокирует обмен генами между
видами в вегетативной фазе поколения Fi. Некоторые линии
Crepis tectorum при скрещивании с С. capillaris дают нежизне-
способные гибриды Fi [381]. В Fi от некоторых межвидовых
скрещиваний у Gossypium, Gilia и Papaver появляются красные
карликовые растения [239, 273—275, 459, 538]. При многих меж-
видовых скрещиваниях с участием Nicotiana langsdorffii и дру-
гих родительских типов, относящихся к роду Nicotiana, у гибри-
дов jFi развиваются генетически детерминированные опухоли
8*
116 Часть третья. Генетические и экологические аспекты вида
Гибридная стерильность, как и несовместимость, охватывает
широкий круг разнообразных явлений. В зависимости от того,
на какой стадии развития проявляется действие барьера, гиб-
ридную стерильность можно разделить на диплонтную стериль-
ность и стерильность гамет. Явления стерильности можно также
классифицировать в зависимости от факторов, которые их конт-
ролируют, на генную, хромосомную и цитоплазматическую сте-
рильность. В реальных случаях гибридной стерильности наблю-
даются разнообразные комбинации различных стадий и факто-
ров.
Взаимодействия между генами растений разных видов часто
приводят к абортивному развитию пыльников или других важ-
ных органов у гибрида. Дисгармоничные взаимодействия ядер-
ных генов с чужеродной цитоплазмой вызывают мужскую сте-
рильность у некоторых гибридов Zea, Epilobiutn, Streptocarpus,
Nicotiana, Capsicum, Solatium и в других группах растений [89,
107, 183, 312, 396, 416, 577, 619, 637, 651].
Неблагоприятные генные взаимодействия иногда проявля-
ются также на стадиях мейоза и гаметогенеза, тем или иным
образом нарушая эти тонко координированные процессы. Гене-
тически детерминированная стерильность гамет обнаружена у
гибридов Nicotiana, Geum, Gilia, Bromus и других родов [229,
476, 309, 878].
Виды растений часто различаются по хромосомным пере-
стройкам, как это отмечалось в предыдущей главе. Перестройки
могут приводить к нарушению у гибрида образования нормаль-
ных бивалентов и (или) нормального распределения хромосом в
анафазе мейоза. При этом у части продуктов мейоза возникают
дупликации или нехватки определенных участков хромосом, и
в результате из них не развиваются жизнеспособные пыльцевые
зерна и зародышевые мешки. Доля таких нежизнеспособных
гаметофитов быстро возрастает с увеличением числа структур-
ных перестроек. Например, у гибрида, гетерозиготного всего по
шести независимым транслокациям, ожидаемая фертильность
пыльцы составляет лишь 1,5%, что сравнимо по уровню сте-
рильности с высокостерильными межвидовыми гибридами.
Хорошо известно, что на поведение хромосом при мейозе
оказывают воздействие как генные факторы, так и структурные
различия в хромосомах [138, 407, 670]. Во многих случаях эти
две причины, вызывающие нарушения мейоза, а следовательно,
стерильность гамет, встречаются одновременно. В сущности
хромосомную стерильность, наблюдаемую особенно часто у. оп-
ределенных жизненных фору растений, можно рассматривать
как состояние, накладывающееся на более широко распростра-
ненное и, вероятно, играющее главную роль явление генной сте-
рильности.
Гл. 9. Изолирующие механизмы
117
Если гибрид в конечном итоге дает какое-то число потомков
Fg или Bi, то среди них, по всей вероятности, будет некоторая
доля слабых или неполноценных индивидуумов. Как ужо гово-
рилось в гл. 8, в поколениях Fa — F6 от скрещиваний Gilia та-
lior и G. modocensis примерно 42% потомков обладали понижен-
ной жизнеспособностью. В F2 от скрещиваний Layia gaillardioi-
des и L. hieracioides (Compositae) пониженная жизнеспособ-
ность наблюдалась у 80% индивидуумов [96].
Выводы
Ценные в адаптивном отношении генные комбинации, имеющие-
ся у отдельных видов, при гибридизации могут распадаться.
Разрушение гибридов — проявление распада адаптивных генных
комбинаций. Другие репродуктивные изолирующие механизмы
служат для защиты этих адаптивных генных комбинаций от
распада.
Репродуктивные изолирующие механизмы могут развиваться
как побочные продукты эволюционной дивергенции. Разрушение
гибридов, несомненно, представляет собой такой побочный про-
дукт. Эволюционная дивергенция может служить и обычно слу-
жит непосредственной причиной различных других форм репро-
дуктивной изоляции. Так, механические и сезонные различия
между Salvia mellifera и S. apiana связаны главным образом с
различием в признаках их нормальных опылителей — мелких
пчел, вылетающих весной, и крупных летних пчел соответствен-
но, и их воздействия на репродуктивную изоляцию представля-
ют собой, возможно, побочные продукты дифференциации.
Однако это не единственный способ возникновения репро-
дуктивной изоляции. Как мы увидим в дальнейшем, механизмы,
действующие в родительском поколении, могут в некоторых слу-
чаях создаваться в результате отбора па изоляцию как тако-
вую. Адаптивные генные комбинации у таких видов предохра-
няются от распада механизмами, возникающими в результате
прямого отбора, направленного на создание репродуктивной изо-
ляции.
10 Экологические взаимодействия
Между различными видами, обитающими на одной и,той же
территории, происходят генетические и экологические взаи^О:
действия. Генетические взаимодействия выражаются в свобод:
ном скрещивании, ограниченном скрещивании или полном от-
сутствии скрещивания в зависимости от степени и способа ре7
продуктивной изоляции. Что касается экологических взаимодей-
ствий, то такие виды могут вступать или не вступать в конку-
ренцию в зависимости от различных факторов; если же между
ними существует конкуренция, то она может привести к разное
образным последствиям. Кроме того, экологические взаимодей-
ствия между видами связаны с их репродуктивными взаимодей.-
ствиями.
Конечно, экология — слишком обширная область, чтобы мож-
но было достаточно подробно изложить здесь ее принципы, да
в этом и нет нужды, поскольку существует ряд хороших совре-
менных руководств по этому предмету; вместе с тем она столь
тесно связана с излагаемыми здесь проблемами, что совсем не
коснуться ее мы не можем. Поэтому нам пришлось пойти на
некоторый компромисс: в этой главе дается краткий очерк из-
бранных аспектов экологии, особенно важных для понимания
проблемы видообразования у растений. В соответствии с такой
задачей мы ограничимся замечаниями о некоторых типах взаи-
модействия между родственными видами.
Экология всегда отличалась богатством терминологии. Со-
временная экология в этом отношении также не составляет иск-
лючения; единственное отличие заключается в том, что терми-
нология ее стала новее. Так, например, в настоящее время при
обсуждении экологической ниши используются такие термины,
как ширина ниши, перекрывание ниш, гиперобъем ниши, фун-
даментальная ниша, реализованная ниша, поведенческая ниша
и т. п. Во многих случаях эти термины — просто новые названия
для существовавших ранее понятий. Для тех целей, которые мы
себе ставим в данной главе, эти термины не нужны, а поэтому
они либо не используются вовсе, либо упоминаются лишь мимо-
ходом, что дает нам возможность сосредоточить внимание на
самих экологических явлениях и процессах.
Сосуществование
Термины аллопатрия и симпатрия полезны для описания про-
странственных взаимоотношений между популяциями [564].
Аллопатрия наблюдается в тех случаях, когда популяции
обитают в разных областях. Следует различать две возможно-
Гл. 10. Экологические взаимодействия
119
сти: а) области разобщены, б) области соприкасаются (пара-
патрия).
Симпатрия наблюдается тогда, когда популяции обитают на
«одной и той же территории, так что особи одной популяции на-
ходятся в пределах нормальной области расселения другой по-
пуляции, и наоборот. Здесь опять-таки существуют две возмож-
ности [282]. а) Смежная симпатрия: популяции занимают раз-
ные ниши в одном и том же местообитании, например в саван-
не, где участки лесной растительности чередуются с открытыми
грасслендами; однако при этом особи, принадлежащие к разным
популяциям, обычно не живут в тесном соседстве. Эти популя-
ции симпатричны в генетическом смысле, коль скоро гаметы или
зиготы одной из них находятся в пределах нормальной области
расселения другой, и наоборот, по не вступают в непосредствен-
ный контакт в экологическом смысле, б) Биотическая симпат-
рия- популяции обитают в одной и той же нише, и принадлежа-
щие к ним особи часто живут в тесном соседстве; таким обра-
зом, они симпатричны не только генетически, но и экологически.
Близкородственные виды растений будут занимать сходные
или одни и те же географические области и обладать сходными
экологическими предпочтениями просто вследствие тесных фи-
логенетических связей между ними. Они могут быть аллопатри-
ческими, смежно- или биотически-симпатрическими в зависимо-
сти от различных факторов.
Биотическая симпатрия — самая тесная форма сосущество-
вания. Она может служить отправной точкой в процессе межви-
дового отбора, ведущего к разнообразным конечным результатам.
Эффекты межвидового отбора
Сосуществование родственных видов может привести и нередко
приводит к конкуренции между видами за ресурсы, необходимые
обоим видам, но имеющиеся в ограниченном количестве. Если
в этой конкуренции один вид превосходит другой, то вступает в
действие межвидовой отбор; затем извилистыми путями этот
отбор может привести к различным конечным результатам.
Один из результатов межвидового отбора — замещение ви-
дов. Это весьма обычный процесс в истории жизни. Другой ре-
зультат— развитие у одного вида механизмов, наносящих вред
виду-конкуренту (например, аллелопатия [592]). Третий воз-
можный результат — экологическая дифференциация между
двумя конкурирующими видами, позволяющая понизить уровень
прямой конкуренции между ними.
Этот последний процесс можно проиллюстрировать с по-
мощью простой модели. Допустим, что два экологически сход-
ных и симпатрических вида X и У конкурируют в гетерогенной
120
Часть третья. Генетические и экологические аспекты вида
среде (Е), имеющей три грани, или ниши (Ei, Е2, Ез). Каждый
вид полиморфен и содержит ряд различных типов (Хь Х2 и
т. д.), адаптированных к этим трем нишам. Полиморфный тип
Xi обладает превосходством в нише Еь а тип Уз —в нише Е3.
В начале процесса межвидового отбора виды распределены
следующим образом:
Среда Ei Е2 Е3
Вид X Xi (обладает пре- Х2 Х3
имуществом)
Вид У У1 Уг Уз (обладает пре-
имуществом)
К концу процесса отбора получается следующая картина:
Среда Ei Е2 Е3
Вид X Xi Х2
Вид У Y2 Уз
Каждый вид стал более узко специализированным к той ни-
ше, в которой он обладает селективным преимуществом. Оба
вида продолжают занимать общую нишу, но у каждого из них
есть теперь и отдельная ниша. Степень перекрывания ниш и
прямая конкуренция между двумя видами снизились.
Межвидовой отбор, действующий таким образом, приводит
в конечном счете к повышению специализации у родственных
симпатрических видов и к смещению признаков [73] в области
симпатрии у частично симпатрических видов.
Сосуществование без межвидового отбора
или замещение видов
Сосуществование не обязательно влечет за собой межвидовую
конкуренцию. Два вида могут сосуществовать, не вступая в
ведущую к отбору конкуренцию, если их численности снижают-
ся под действием какого-либо иного фактора, например сильно-
го хищничества. В таком случае процесс межвидового отбора
не развивается.
Возможна и такая ситуация, когда два сосуществующих вида
с одинаковыми потребностями конкурируют за какой-либо огра-
ниченный ресурс лишь в небольшой степени, потому что они
нуждаются в незначительных количествах этого ресурса. В та-
ких случаях процесс межвидового отбора протекает медленнее,
и он, возможно, менее эффективен по сравнению с другими
стохастическими факторами, воздействующими на обилие ви-
дов.
Возможны также случаи, когда сначала возникает межвидо-
вая, конкуренция, а затем следует межвидовой отбор; однако
процесс отбора не достигает такого уровня, чтобы привести к за-
мещению того или другого вида или к экологической дифферен-
Гл. 10. Экологические взаимодействия
121
циации. Межвидовой отбор — процесс длительный, и за то вре-
мя, пока он происходит, направляющий его фактор может изме-
ниться. А это может привести к реверсии относительных селек-
тивных преимуществ конкурирующих видов или разных форм
(при полиморфизме)
Если изменения факторов среды носят циклический харак-
тер, причем каждый цикл занимает меньше времени, чем процесс
полного замещения одного вида другим, то между двумя сим-
патрическими видами, находящимися в состоянии конкуренции,
может установиться неустойчивое равновесие и они будут сосу-
ществовать постоянно.
Согласно принципу конкурентного исключения [340], посто-
янное сосуществование между двумя полными конкурентами
невозможно. Этот принцип в общем виде верен как теоретиче-
ски, так и эмпирически, однако ни в одном конкретном случае
его не следует применять некритически. Как было показано вы-
ше, сосуществование не обязательно должно сопровождаться
межвидовой конкуренцией, а межвидовая конкуренция не всег-
да несовместима с сосуществованием
Эффекты экологических требований
Организмы разных типов предъявляют очень различные требова-
ния к ресурсам среды, в том числе и к ограничивающим ресур-
сам У часто встречающихся видов крупных организмов, как
правило, требования бывают более высокие, чем у часто встре-
чающихся видов мелких организмов. Вследствие этого межви-
довая конкуренция за ограничивающие ресурсы, имеющие ре-
шающее значение, обычно бывает более острой среди крупных
организмов, чем среди мелких.
Можно предсказать, что эти различия перейдут в различия
в интенсивности межвидового отбора. У крупных организмов
межвидовой отбор, вероятно, быстро приведет к замещению ви-
дов У мелких организмов, у которых межвидовая конкуренция
слабее или даже пренебрежимо мала, процесс межвидового от-
бора протекает медленнее и может не дойти до полного завер-
шения, т. е. до замещения видов, в пределах данной территории.
У таких организмов достижение симпатрии и ее сохранение осу-
ществляются легче [282, 695].
Некоторые эмпирические данные подтверждают правильность
этих предсказаний. Во флоре западных областей США близкие
виды таких древесных растений, как Pinus и Quercus, и таких
многолетних травянистых растений, как Polemonium и Calochor-
tus, обычно занимают различные экологические зоны на эко-
географических трансектах Симпатрические контакты между
родственными видами в этих группах обычно ограничиваются
122 Часть третья. Генетические и экологические аспекты вида
областями перекрывания их местообитаний. В этой флоре био-
тически-симпатрические скопления близких видов встречаются
обычно, и почти исключительно, среди однолетних травянистых
растений, таких, как Clarkia и Gilia. Например, в природе не-
редко можно встретить два, три и даже больше видов Giliar
растущих рядом [282, 283].
Сходные корреляции наблюдаются у животных. Симпатриче-
ские скопления близких видов существуют у таких мелких на-
секомых, как дрозофилы, комары и цикадки Erythroneura. Про-
тивоположную крайность представляют крупные млекопитаю-
щие, у которых близкие виды обычно аллопатричны или пара-
патричны. У чешуекрылых, пищевые потребности которых на
стадии имаго очень малы, взрослые особи, принадлежащие к
разным видам, часто занимают широко перекрывающиеся пи-
щевые ниши, тогда как усиленно питающиеся гусеницы тех же
самых видов занимают обособленные узкие пищевые нищи [282,
695].
Экологические взаимодействия и изолирующие механизмы
близких видов связаны между собой. В результате симпатриче-
ских контактов возникают ситуации, вызывающие разного рода
реакции. В некоторых группах растений эта реакция выража-
ется в экологической дивергенции и обособлении, влекущих за
собой более сильную экологическую изоляцию, тогда как дру-
гие группы реагируют усилением репродуктивной изоляции в
области симпатрии. Возможно, что во многих группах эти две
реакции объединены тем или иным образом. Важным фактором,
определяющим тип реакции у данного вида растений, является
то, в какой степени — высокой, умеренной или низкой — он нуж-
дается в ресурсах среды.
Биотическая симпатрия, сохраняющаяся на протяжении дли-
тельного времени, зависит, по-видимому, от совместного дейст-
вия двух факторов: слабой или неполной межвидовой конкурен-
ции и сильной репродуктивной изоляции.
11 Формы взаимоотношений
по плодовитости
Селекционеры давно поняли, что разные группы растений резко
различаются по той легкости, с которой происходит скрещива-
ние между видами, и по плодовитости гибридов. В некоторых
родах близкие виды скрещиваются с трудом, а гибриды между
ними, если их удается получить, обычно стерильны. Другую
крайность составляют группы растений, у которых возможны
широкие скрещивания между морфологически различными вида-
ми, секциями или даже родами, и гибриды, полученные от таких
скрещиваний, плодовиты.
Результаты различных экспериментов по гибридизации в
очень многих случаях непредсказуемы. Однако при сравнитель-
ном изучении достаточно большой выборки групп растений вы-
являются некоторые общие закономерности.
Основная часть данной главы будет посвящена обзору ряда
групп растений, главным образом из северного умеренного поя-
са, которые изучались в генетико-таксономическом плане раз-
ными исследователями. Эти растения сгруппированы здесь по
жизненным формам и системам скрещивания. В заключитель-
ных параграфах очерчены общие закономерности и указаны не-
которые корреляции.
Древесные растения
Ceanothus
Североамериканский род Ceanothus (Rhamnaceae) занимает
область от южной части Канады до Гватемалы с центром рас-
пространения на тихоокеанском побережье. Из 55 видов этого
рода 44 эндемичпы для Калифорнии. Это кустарники с мелки-
ми душистыми цветками. Опыление производят разнообразные
насекомые, обеспечивающие облигатное перекрестное оплодо-
творение у самонесовмсстимых видов и преимущественное пере-
крестное оплодотворение у самосовместимых видов. Число хро-
мосом 2гг. = 24 у всех известных видов [614].
Виды этой группы делятся на две хорошо выраженные сек-
ции Ceanothus и Cerastes. Характер взаимоотношений между
видами у калифорнийских представителей Cerastes и между
Cerastes и Ceanothus установил Нобс [614], сочетавший полевые
и экспериментальные исследования.
Виды секции Cerastes встречаются в самых разнообразных
экологических местообитаниях на тихоокеанском побережье.
Они дифференцированы как морфологически, так и экологиче-
124 Часть,третья. Генетические ц экологические аспекты вида
ски. В большинстве комбинаций эти виды (или полувиды) раз-
делены географически. В остальных сочетаниях их ареалы пере-
крываются, и они встречаются на разных, но примыкающих
друг к другу типах почв в одной и той же области [614].
Ареал Ceanothus cuneatus перекрывается с ареалами по
крайней мере 5 других видов секции Cerastes в области, рас-
положенной к северу от бухты Сан-Франциско в Калифорнии.
Ceanothus cuneatus широко распространен на песчаниках, а в
некоторых районах, где на поверхность выходят серпентиниты,
образуя отдельные пятна среди окружающего песчаника, он
встречается вместе с двумя эндемичными для серпентинитов
видами—С. jepsonii и С. pumilus. В некоторых из таких зон
контакта естественной гибридизации не происходит или почти
не происходит, тогда как в других имеет место значительная
гибридизация между С. cuneatus и видами или полувидами,
растущими на выходах серпентинитов [614].
Нобс скрещивал друг с другом 12 видов секции Cerastes;
С. cuneatus
crasslfolius
fC ramulosus
С. greggii'
perplexans
С masonii
С. verrucosus'
С. glorioeus
С rigidus>
C. Jepsonii
С. purpureus
'van albiflorus
fC confusus
C. prostratus
occidentalis
: Доведены до F2 без потери мощности
Ft жизнеспособно, семена завязываются нормально
Завязывание плодов не изучалось
Рис. 11.1. Плодовитость гибридов при скрещивании между разными видами
секции Cerastes рода Ceanothus [613].
Гл It Формы взаимоотношений по плодовитости
125
результаты представлены на рис. 11.1. Все эти виды легко скре-
щиваются и дают в F! жизнеспособные гибриды. Гибриды об-
разуют жизнеспособные семена и пыльцу; фертильная пыльца
составляет от 88 до 97% всей пыльцы. Мейоз у гибридов также
протекает нормально с регулярным формированием 12 бивален-
тов '[614].
Члены секций Cerastes и Ceanothus симпатричны на обшир-
ной территории, и виды, принадлежащие к этим двум секциям,
обычно растут в тесном соседстве в одном и том же раститель-
ном сообществе и цветут одновременно. Естественные межсек-
ционные гибриды очень редки, несмотря на обширность области
контакта [614].
В свете приведенных данных интересно было обнаружить^
что между представителями этих двух секций существует репро-
СЕКЦИЯ CERASTES
СЕКЦИЯ EUCEANOTHUS
Получены сублетальные гибриды
---------Семена не завязывались
Рис 112 Плодовитость гибридов при скрещиваниях между членами двух
секций рода Ceanothus [613].
126
Часть третья. Генетические и экологические аспекты виОа
дуктивная изоляция, обеспечиваемая эффективными внутренни-
ми барьерами. Попытки Нобса произвести межсекционные скре-
щивания в большинстве случаев оказались совершенно безус-
пешными; гибриды не давали семян (рис. 11.2), В тех редких
случаях, когда удавалось получить несколько гибридных семян
и проростков, они обычно оказывались нежизнеспособными и
логибали на ранних стадиях. Еще реже межсекционные гибриды
достигали зрелости; при этом они были стерильными (фертиль-
ная пыльца составляла от 0 до 8%), и течение мейоза было
нарушено. При конъюгации хромосом у спонтанного гибрида
между С. masonii и С. foliosus число бивалентов составляло от
2 до 11, в среднем 7 [614].
Quercus
К обширному и широко распространенному роду Quercus (Fa-
gaceae) принадлежат многочисленные виды деревьев и кустар-
ников. Все они диплоидны, причем во всех случаях, в которых
был проведен цитологический анализ, оказалось, что диплоид-
ный набор (2п) составляют 24 хромосомы. Для этих растений
характерно перекрестное ветроопыление. Все виды распределе-
ны между двумя четко разграниченными подродами — Quercus
и Erythrobalanus. Данных о взаимоотношениях между видами
дубов довольно много, но они фрагментарны и .разбросаны по
разным источникам; поэтому мы в своем изложении следуем
обзорам, составленным для некоторых географических областей
(58, 780].
Виды или полувиды, принадлежащие к одному и тому же
подроду, обычно распределены по различным, но часто заходя-
щим одна в другую, подобно переплетенным пальЦам, климати-
ческим или эдафическим зонам одной и той же области. Так,
среди калифорнийских представителей подрода Quercus вид
Quercus lobata произрастает на дне долин, a Q. douglasti—в
сухих предгорьях; Q. dur ata —къ. выходах серпентинитов, а
<?. dumosa— на окружающей территории, где серпентиниты
•отсутствуют. В Техасе Q. mohriana встречается на известняках,
Q. havardii — на песках, a Q. grisea — на выходах изверженных
пород [589].
Там, где виды или полувиды одного и того же подрода при-
ходят в контакт, они часто гибридизируют; в результате обра-
зуются обширные сингамеоны, как это будет описано в одной из
последующих глав. Естественные гибриды между видами, при-
надлежащими к одному подроду, по-видимому, более или менее
плодовиты, и мейоз протекает у них нормально [780]. Такер
[839] обнаружил,, что у гибридов между Quercus garryana и
Q. durata (весьма отдаленное скрещивание в морфологическом
Гл 11. Формы взаимоотношений по плодовитости
127
смысле) образуется от 93 до 97% фертильной пыльцы и мейоз
протекает нормально, однако семена этого гибрида стерильны;
стерильными оказались также семена гибрида между Q garrya-
па и Q. dumosa.
Дубы, принадлежащие к подродам Quercus и Ег у throb alarms,
довольно часто контактируют также и в природе. Тем не менее
гибриды между подродами в природе не обнаружены, а при
искусственных скрещиваниях возникают редко [770].
Ribes, Pinus, Eucalyptus, Prosopis
К роду Ribes (Saxifragaceae) относятся свыше 150 видов кустар-
ников. Те из них, которые были изучены цитологически, оказа-
лись диплоидами (2п=16). Этот род делится на четыре хорошо
различающихся подрода и на многочисленные секции.
Кип [429] и другие исследователи проводили множество скре-
щиваний между разными видами Ribes. Если деление на секции
привести в соответствие с данными по скрещиваниям, то выяв-
ляются следующие закономерности. Во всех испытанных комби-
нациях виды, принадлежащие к одной и той же секции, легко
скрещиваются друг с другом и дают плодовитых гибридов.
Скрещивания между представителями разных секций одного и
того же подрода бывают успешными не во всех случаях, меж-
секционные гибриды, если их удается получить, обладают час-
тичной плодовитостью. Главный барьер, препятствующий скре-
щиванию у рода Ribes, существует между разными подродами»
которые хорошо различаются также и морфологически. Репро-
дуктивная изоляция между подродами создается резко выра-
женной несовместимостью. В тех редких случаях, когда этот
барьер преодолевается, возникающие гибриды Fi стерильны
[429].
В общем сходная картина наблюдается у рода Pinus
(2п = 24), который делится на два хорошо выраженных подро-
да — Pinus (=Diploxylon) и Strobus ( = Haploxylori) Виды под-
рода Pinus успешно скрещиваются во многих комбинациях.
Успешными обычно оказываются скрещивания в пределах ка-
кой-либо группы видов, таких, как группа Pinus ponderosa или
группа Р. radiata Некоторые из таких межвидовых гибридов
плодовиты. Подобным же образом представителей подрода
Strobus можно искусственно скрещивать в некоторых комбина-
циях. Однако ни одного успешного скрещивания между членами
разных подродов провести не удалось вследствие существующе-
го между ними эффективного барьера несовместимости [129»
180, 579, 685, 906, 907].
Подобные же взаимоотношения обнаружены между видами
рода Eucalyptus (Myrtaceae) в Австралии. Этот род делится на
128 Часть третья. Генетические и экологические аспекты вида
пять подродов с многочисленными видами, у большинства из
которых число хромосом 2п=22. Прайор [650] сообщает, что ви-
ды, принадлежащие к одному и тому же подроду, обычно зани-
мают экологически различные местообитания и в ряде извест-
ных случаев интерфертильны. В отличие от этого виды, отно-
сящиеся к разным подродам, часто встречаются вместе, а .скре-
щивание между ними предотвращается высоко эффективными
внутренними барьерами [660].
Аналогичная ситуация была описана сравнительно недавно
для Prosopis (Leguminosae) [729].
Многолетние травянистые растения
с резко выраженными различиями
в механизме цветка
Aquilegia
Aquilegia (Ranunculaceae) — один из родов многолетних тра-
вянистых растений с яркими цветками; число хромосом 2п=14.
Этот род широко распространен в Евразии и Северной Амери-
ке. По всей этой области произрастают многочисленные имено-
ванные виды, многие из которых, по-видимому, представляют
собой географические расы. Разнообразные формы, относящие-
ся к этому роду, естественно, распадаются на четыре или, быть
может, пять главных групп видов [270, 646].
I) Самый примитивный вид — это Aquilegia ecalcar ata из
Гималаев с белыми или пурпуровыми цветками, лишенными
шпор. Он близок к родственному роду Isopyrum (2п=14), имею-
щему сходные цветки. 2) По всей Евразии и в северной части
Северной Америки распространено большое число форм, в том
числе группа видов A. vulgaris с поникающими синими или пур-
пуровыми цветками, снабженными длинными шпорами. Эта
группа делится на две подгруппы: подгруппа собственно A. vul-
garis с загнутыми шпорами и другая подгруппа A. alpina с пря-
мыми шпорами. 3) По всему умеренному поясу Северной Аме-
рики распространена группа A. canadensis, для которой харак-
терны поникающие красные цветки с толстыми шпорами.
4) В западных районах Северной Америки произрастают члены
группы A. chrisantha с прямостоячими желтыми цветками, не-
сущими длинные тонкие шпоры.
В различных районах Северной Америки три группы видов
распространены симпатрически. В некоторых местностях они об-
разуют естественные гибриды, сохраняя обособленность в дру-
гих районах, где они симпатрически перекрываются [270].
Первые работы,.касающиеся взаимоотношений между вида-
ми Aquilegia по .плодовитости, проведенные Андерсоном и Ша-
Гл 11. Формы взаимоотношений по плодовитости
129
фером [21], были продолжены Празмо [646—648]. Как показали
эти исследования, члены четырех описанных выше групп видов
легко скрещиваю 1ся между собой в любых комбинациях и дают
плодовитые или полуплодовитые гибриды Fi с нормальным те-
чением мейоза.
Так, Fi от скрещиваний двух далеких в пределах этого рода
видов — Aquilegia ecalcarala и A. vulgaris — дает 42% фертиль-
ной пыльцы, 19% фертильных семян и большое число потомков
F2 [645]. Fi от скрещивания A. ecalcarala и A chrysantha дает
62% фертильной пыльцы и 8% фертильных семян [647]. Потом-
ки Fi от скрещивания A. ecalcarata и A canadensis также дают
полуфертильную пыльцу и семена [649] А в Fi от скрещиваний
между членами групп A vulgaris, A. chrysantha и A canadensis
получаются полуплодовитые гибриды.
Таким образом, скрещиванию между группами видов у это-
го рода препятствуют, очевидно, главным образом внешние
барьеры. У североамериканских водосборов эти барьеры созда-
ются экологическими, механическими и этологическими изоли-
рующими механизмами. Виды, принадлежащие к группе Aqui-
legia vulgaris, обычно опыляются шмелями, виды группы A ca-
nadensis— колибри, а группы A chrysantha — бражниками [94,
270].
Скрещивания между Aquilegia и Isopyrum приводят к со-
вершенно иным результатам, нежели скрещивания внутри рода
Aquilegia Скалинска [734] пыталась скрестить Aquilegia ecalca-
rata с Isopyrum thalictroides\ при этом она обнаружила несов-
местимость па разных стадиях до и после оплодотворения.
Пыльцевые трубки Isopyium прорастают в столбике Aquilegia
на незначительное расстояние. Некоторые яйцеклетки все же
оплодотворяются Но затем, в течение двух первых педель эмб-
рионального развития, либо происходит дегенерация эндоспер-
ма, либо не развивается сам зародыш, так что в конечном ито-
ге никаких гибридных проростков не образуется [732].
Орхидные
Хорошо известно, что в группе орхидных можно производить
скрещивания между очень далекими формами Виды рода Van-
da успешно скрещиваются с представителями девяти других ро-
дов Представителей десяти родов подтрибы Laelieac (Cattleya,
Epidendrum, Laelia и другие) скрещивали примерно в 20 раз-
личных межродовых комбинациях Несколько родов подтрибы
Oncidieae успешно скрещиваются с родами, принадлежащими к
трем другим подтрибам — Cochliodeae, Comparettieac и Tricho-
centreae [463].
9-45
130
Часть третья. Генетические и экологические аспекты вида
К роду орхидных Ophrys в Средиземноморской области от-
носится ряд видов, которые при искусственных скрещиваниях
друг с другом дают гибриды, производящие фертильную пыль-
цу. Эти виды, встречаясь в природе в различных симпатриче-
ских комбинациях, не гибридизируют в сколько-нибудь значи-
тельной степени. Их изоляция в природных условиях обеспечи-
вается главным образом различиями в строении цветков,
создающими этологические и механические барьеры, препятст-
вующие межвидовому опылению, которое производят пчелы
[791].
В одной из секций орхидных рода Vanda у межвидовых гиб-
ридов происходит нормальный мейоз с полной конъюгацией
хромосом, но у гибридов между видами, относящимися к дру-
гой секции того же рода, мейоз нарушен [424]. Нормальное те-
чение мейоза наблюдается также у гибридов между некоторыми
видами Dendrob turn [721].
Секция Oncocyclus рода Iris
Секция Oncocyclus рода Iris (Iridaceae) — разнообразная груп-
па ирисов, произрастающих в юго-западных областях Азии.
Крупные цветки этих растений снабжены специализированными
механизмами, различающимися у главных групп видов; опыле-
ние производят крупные пчелиные (Xylocopa, Bombus, Antho-
phora). Изоляция разных видов в природе создается географи-
ческими, экологическими, временными и механическими барь-
ерами. Никаких внутренних репродуктивных изолирующих
механизмов у этих видов не возникло. При искусственных меж-
видовых скрещиваниях, производимых во многих комбинациях,
снижения способности к скрещиванию или плодовитости гибри-
дов по сравнению с тем, что наблюдается при внутривидовых
скрещиваниях, нет или почти нет. Почти у всех гибридов Fi фер-
тильность пыльцы была близка к 90% [27].
Antirrhinum, Penstemon, Epiphyllum
Рассмотрим кратко три другие группы многолетних растений с
яркими цветками; принадлежащие к этим группам виды интер-
фертильны в широких пределах.
Большинство видов рода Antirrhinum (Scrophulariaceae), от-
носящихся к секции Antirrhinastrum (2п=16), более или менее
легко скрещиваются друг с другом и дают плодовитых гибри-
дов. Изоляцию между этими видами создают главным образом
экологические и этологические барьеры. Но виды секции Antir-
rhinastrum изолированы от Antirrhinum orontium и от секции
Гл 11. Формы взаимоотношений по плодовитости
131
или выделенного в качестве самостоятельного (сегрегатного)
рода Asarina барьерами несовместимости [49, 560].
Три вида Penstemon секции Peltanthera (Scrophulariaceae)
обитают в разных экологических зонах, ио произрастают симпа-
трически в различных комбинациях па юге Калифорнии, к этим
видам относятся Penstemon grinnellii, Р. spectabilis и Р centran-
thifolius. Они резко различаются по строению и окраске цветков:
у Р. grinnelli венчик двугубый, с широким зевом, у Р spectabilis
цветки синеватые, с широкой трубкой, а у Р. centranthifolius—
красные, воронковидные.
При скрещиваниях па экспериментальных участках все три
вида интерфертильны. В природе они гибридизируют в некото-
рых областях, где встречаются совместно, оставаясь изолирован-
ными в других областях. Изоляция создается главным образом
механическими и этологическими факторами. Penstemon grinnel-
lii обычно опыляется пчелами-каменщицами (Xylocopa), Р. spec-
tabilis— осами-псевдомазоридами и средних размеров пчелины-
ми, а Р. centranthifolius — колибри [808].
Виды Epiphyllum (Cactaceae) также сильно различаются по
своим ярким цветкам. В пределах рода Epiphyllum, а также
между видами, которые иногда помещают в разные выделенные
из него роды, возможны отдаленные скрещивания. Так, можно
получить жизнеспособные гибриды, скрещивая между собой
вид с красными цветками, опыляемый колибри, и формы с бе-
лыми цветками, опыляемые бабочками [353].
Многолетние травянистые растения
с ярко выраженными межвидовыми различиями
в механизме цветка
Geum
Весьма обширный род Geum (Rosaceae), к которому относятся
примерно 56 видов, широко распространен по всему северному
полушарию, а также в умеренных областях южного полушария.
Это многолетние травянистые растения с перекрестным опыле-
нием. Основное число хромосом в трибе, к которой относится
Geum, х = 7. В роде Geum не известно ни одного диплоида, все
его члены — полиплоиды (от 4х до 12х) с числом хромосом от
2п = 28 до 2п = 84 [230, 231].
Гаевски [230] делит род Geum на 11 подродов. Большая
часть генетико-таксономических исследований проводилась на
двух из этих подродов, а именно — Oreogeum и Geum. К подроду
Oreogeum относятся три вида, распространение которых ограни-
чено горными районами Европы Два хорошо известных вида
9*
132 Часть третья. Генетические и экологические аспекты вида
подрода Oreogeum—G. montanum (4х) и G. reptans (6х). Бо-
лее обширный и широко распространенный подрод Geum содер-
жит 25 видов, в том числе следующие (все они гексаплоиды):
G. rivale, G. urbanum, G. macrophyllum, G. molle, G. coccineum,
G. hispidum и G. canadense [230].
В природе изоляция между этими видами создается глав-
ным образом экологическими и сезонными барьерами. Там, где
эти барьеры разрушаются, между видами происходит естествен-
ная гибридизация. Geum rivals и G. urbanum гибридизируют во
многих зонах контакта по всей Европе [228, 230].
На экспериментальных участках эти виды легко скрещива-
ются друг с другом. Любые два вида подрода Geum можно
скрещивать независимо от различий в уровне плоидности. Пред-
ставители подрода Geum успешно скрещиваются с видами Oreo-
geum и четырех других подродов. Но между группами более
высокого ранга — родом Geum и родственными родами Wald-
steinia и Coluria— вступают в действие очень эффективные
барьеры несовместимости, и все попытки провести межродовые
скрещивания оказываются безуспешными [230].
Жизнеспособность искусственных межвидовых гибридов
варьировала от гетерозисной до сублетальной при разных ком-
бинациях скрещиваемых форм. В пределах подрода Geum гиб-
риды Fi от скрещиваний Geum macrophyllum с G. rivals, G. mon-
tanum или G. canadense обладали нормальной мощностью, но
рост гибридов между G. macrophyllum и G. urbanum, G. molle
или G. hispidum был сильно подавлен. Большая часть гибридов
от скрещиваний между членами разных подродов была жизне-
способной, но гибриды Fi от скрещиваний Geum и Woronoviia
никогда не цвели [230].
Стерильность гибридов также варьировала в широких преде-
лах— от почти нормальной плодовитости до полной стерильно-
сти. Так, ^апример, в подроде Geum фертильность пыльцы гиб-
ридов Fi от скрещивания Geum rivals и G. urbanum составляла
78, а семян — 72%; высокая плодовитость наблюдалась также
в Fi от скрещиваний Geum rivals с G. coccineum, G. molle и
G. hispidum. Но гибриды G. rivale и G. macrophyllum в Fi были
высокостерильны: фертильная пыльца и завязавшиеся семена
составляли всего 1%; очень высокой стерильностью отличались
и многие другие гибриды в подроде Geum. В подроде Oreogeum
гибриды между G. montanum и G. reptans производили высоко-
стерильную пыльцу и совершенно стерильные семена [230].
Некоторые скрещивания представителей подрода Geum и
Oreogeum друг с другом давали полуплодовитых гибридов Fi;
например, у гибридов G. montanum с G. rivale фертильная
пыльца составляла 34%, а фертильные семена —15%. При дру-
гих комбинациях •представителей подрода Geum и Oreogeum,
Гл 11. Формы взаимоотношений по плодовитости
133
например G. rivalexG. reptans, получались высокостерильные
гибриды. Все прочие скрещивания между членами разных под-
родов также приводили к высокой стерильности [230].
У гибридов между членами группы видов, в которую входят
Geum rivale, G. urbanum, G. molle и G hispidum, наблюдалась
довольно высокая способность хромосом к коньюгации В раз-
ных клетках этих гибридов образовывалось от 17 до 21 бивален-
та, и их хромосомы были гомологичны или почти гомологичны.
Но в клетках стерильных гибридов от скрещивания G macro-
phyllum с G rivale конъюгация сильно нарушается, число би-
валентов варьирует от 0 до 7, а унивалепты многочисленны
Между двумя из трех геномов G. rivale (6х, подрод Geum) и
двумя геномами G. montanum (4х, подрод Oreogeum) наблюда-
ется хорошая гомология. У гибридов между членами подрода
Geum и других подродов способность хромосом к конъюгации
была понижена [230].
Судя по поведению спонтанного амфиплоида от Geum rivale
(6х) и G. macrophyllum (6х), эти виды различаются по хромо-
сомным перестройкам. У гибридов Fi конъюгация нарушена,
число бивалентов равно 0—7, и гибриды эти, как уже говори-
лось, высокостсрильпы От них были получены два растения
F2 с удвоенным числом хромосом, т. е. 12х; в клетках этих ам-
фиплоидов при мейозе формировалось до 42 бивалентов, и они
были плодовиты [229].
Есть, однако, другие факты, указывающие на то, что у гиб-
ридов Geum rivale и G macrophyllum имеются нарушения в
конъюгации хромосом, контролируемые генами. Если скрестить
G. rivale или G. macrophyllum с третьим гексаплоидным видом,
G aleppicum, то происходит полная конъюгация с образованием
21 бивалента, иными словами, хромосомы G. rivale и G. macro-
phyllum более гомологичны, чем можно было бы думать, судя по
особенностям мейоза у гибрида между этими видами |229]. Та-
ким образом, стерильность этого и других гибридов между вида-
ми рода Geum обусловлена, вероятно, не только хромосомными,
но и генными различиями [229—231].
Solarium
К секции Tuberarium рода Solanum (Solanaceae) относится
группа клубнеобразующих видов со свободным неродственным
скрещиванием, которые образуют полиплоидный ряд от 2х до
6х (х=12). Эти виды легко скрещиваются в большинстве ком-
бинаций. Некоторые межвидовые гибриды плодовиты, и все их
хромосомы образуют биваленты, другие стерильны и способ-
ность хромосом к конъюгации у них понижена, но в общем все
эти виды цитогенетически весьма близки. Имеются данные, сви-
134 Часть третья. Генетические и экологические аспекты вида
.детел ьствующие о том, что некоторые виды различаются по не-
большим структурным перестройкам [123, 311, 354, 542, 813,
<814].
Секция Apogon рода Iris
Одна из естественных трупп среди безбородых ирисов (секция
Apogon рода Iris) — ряд Californicae, или тихоокеанские ири-
сы, состоящая из 11 полувидов, распространенных по тихоокеан-
скому побережью США, от шт. Вашингтон до юга Калифорнии.
К этой группе относятся свободно опыляющиеся многолетние
травянистые растения с числом хромосом 2п=40. В окраске и
форме их ярких цветков наблюдаются межвидовые различия,
но эти различия невелики, и, кроме того, они связаны интергра-
дацией [461, 462].
В пределах общей области распространения тихоокеанских
ирисов отдельные полувиды занимают различные экологические
зоны. Например, Iris douglasiana растет вдоль береговой линии;
I. macrosiphon — дальше от берега, в предгорьях, a I. hartwe-
gii — в горах Сьерра-Невада, в зоне сосен подрода Diploxylon.
Местообитания этих и других полувидов во многих случаях не-
прерывно переходят одни в другие, что создает возможность для
краевых или смежно-симпатрических контактов между ними; в
таких случаях между подвидами происходит естественная гиб-
ридизация в многочисленных комбинациях [461, 462]. Образуе-
мый ими сингамеон будет описан в гл. 18.
Смит и Кларксон {741], а также Ленц [462] искусственно
скрещивали тихоокеанские ирисы друг с другом в разнооб-
разных комбинациях. Эти полувиды, легко скрещиваются. Гиб-
риды Fi вполне жизнеспособны, плодовиты и производят от 84
до 99% фертильной пыльцы. Мейоз у гибридов протекает нор-
мально с образованием 20 бивалентов.
Самые близкие родичи тихоокеанских ирисов — группа евра-
зийских видов с 40 хромосомами, принадлежащая к ряду Sibi-
ricae. Члены этих двух рядов легко скрещиваются друг с другом,
образуя жизнеспособные гибриды, которые, однако, стерильны
Более далекий родич тихоокеанских ирисов — Iris tenuis
(2/г=28), относящийся к ряду Evansia [462]. I. tenuis растет в
Орегоне, где он симпатричен с такими членами ряда Californi-
cae, как I. tenax. Естественной гибридизации между I. tenuis и
I. tenax не происходит; как установили Смит и Кларксон [741 j,
между ними существует эффективный барьер несовместимости.
Этот барьер создается блоками, действующими на нескольких
Гл. 11. Формы взаимоотношений по плодовитости
135
стадиях. Пыльцевые трубки в пестиках чужого вида растут мед-
ленно и нередко разрываются. Оплодотворение либо не происхо-
дит вовсе, либо задерживается. А развитие гибридных зароды-
шей в тех редких случаях, когда они образуются, прекращается
на стадии 12 клеток. Искусственных гибридов между I. tenuis
и I. tenax получить не удается.
Silene и др.
К роду Silene (Caryophyllaceae) относятся многочисленные ви-
ды многолетних травянистых растений и несколько однолетни-
ков. Их основное число хромосом х=12; у этого рода известно
много диплоидных видов (2п = 24), особенно в Европе, но до-
вольно обычны и полиплоиды.
В западных районах Северной Америки произрастают 33 ви-
да травянистых многолетников с различной плоидностью — от
2х до 8х; в большинстве случаев это тетраплоиды, но есть среди
них также диплоиды, гексаплоиды и октоплоиды [447, 450]. Эти
западноамериканские виды обладают резко выраженными мор-
фологическими различиями [451].
Крукеберг [448, 451] обнаружил, что западноамериканские
виды легко скрещиваются между собой в большинстве комбина-
ций и получающиеся при этом гибриды жизнеспособны, но, за
немногими исключениями, высокостерильны. Конъюгация хро-
мосом у гибридов очень ограничена; наблюдаются у них также
и другие нарушения мейоза [448, 451].
Обширные исследования, проведенные Викери [860] на Mi-
mulus (Scrophulariaceae), показывают, что закономерности, об-
наруженные у Geum, наблюдаются также у многолетних пред-
ставителей секции Simiolus рода Mimulus, среди которых цент-
ральное место занимают группы М. guttatus и М. glabratus. Эти
же закономерности выявляются у Vernonia (Compositae) [418].
Однолетние травянистые растения
Gilia
Одним из естественных подразделений гетерогенного рода Gilia
(Polemoniaceae) является секция Gilia, или облиственпостебель-
ные гилии. К этой секции относятся десять видов однолетних
травянистых растений Два вида гилии—Gilia capitata и G tri-
color— мы уже рассматривали в гл. 4, но в связи с другой
проблемой
Облиственностебельные гилии произрастают на тихоокеан-
136 Часть третья. Генетические и экологические аспекты вида
ском побережье Северной Америки и в умеренном поясе Южной
Америки, где ареалы разных видов перекрываются и где они
часто встречаются в тесном соседстве в различных комбинациях.
«Система размножения варьирует у разных членов группы от
свободного перекрестного опыления до автогамии. У шести ви-
дов число хромосом 2п=18, а у трех тетраплоидов — 2п=36.
Еще один, десятый, вид не изучался ни цитологически, ни гене-
тически [274, 286].
Все эти виды в большинстве комбинаций разделены мощ-
ными барьерами несовместимости. Если и удается получить
гибриды Fi, то они высокостерильны. Мейоз у гибридов нару-
шен, причем способность хромосом к конъюгации настолько по-
нижена, что в некоторых случаях образуется всего по одному
биваленту на клетку [268].
Низкая способность к конъюгации, наблюдаемая у гибридов,
обусловлена главным образом различиями между родительски-
ми видами по структурным перестройкам, о чем свидетельст-
вуют особенности искусственных амфиплоидов. У стерильных
гибридов с нарушенным мейозА!, полученных на эксперимен-
тальных участках путем скрещивания разных видов в четырех
различных комбинациях, произошло спонтанное удвоение хро-
мосом; в результате возникли амфиплоидные потомки, хромосо-
мы которых нормально конъюгировали с образованием бивален-
тов, а плодовитость колебалась от средней до высокой [268,
286]. В первых поколениях амфиплоидных потомков наблюда-
лось расщепление как по поведению хромосом, так и по плодо-
витости; это служит веским доказательством того, что в данном
случае, помимо хромосомных перестроек, имеют также место
нарушения мейоза, контролируемые генами [271].
Подобная же картина наблюдается в близкой к предыдущей
секции Arachnlon, состоящей также из многочисленных видов
однолетних травянистых растений, которые часто встречаются
в природе симпатрически. Виды эти обычно разделены эффек-
тивными барьерами несовместимости, а гибриды между ними
в тех случаях, когда их удается получить, высокостерильны.
Стерильность гибридов имеет в основном хромосомную, а час-
тично-генную природу [158, 285, 300].
Layla
К роду Layla (Compositae) принадлежит 15 видов однолетних
трав, произрастающих в предгорьях и долинах Калифорнии и
цветущих весной. В этой области тесная симпатрия 2 — 3 ви-
дов— обычное явление. По числу хромосом эти виды можно
разбить на три серии: у шести диплоидных видов число хромо-
Гл 11. Формы взаимоотношений по плодовитости
137
------- Стерильные гибриды
Скрещивание провести не удалось
Рис 11.3 Плодовитость гибридов при скрещиваниях между разными вида-
ми Madia [96].
сом 2п=14, у восьми диплоидных видов — 2п=16 и у одного
тетраплоидного — 2п=32 [428],
Виды, имеющие по 7 пар хромосом, успешно скрещиваются
между собой. Большинство видов с 8 парами хромосом также
способны скрещиваться друг с другом. Однако между видами с
7 и видами с 8 парами хромосом, а также между некоторыми
видами с 8 парами существуют эффективные барьеры несовме-
стимости [96, 102].
138 - Часть третья. Генетические и экологические аспекты вида
Гибриды между некоторыми видами, например между Layla
gaillardioides и L. heterotricha, нежизнеспособны, а такие скре-
щивания, как L. gaillardioidesy.L. hieracioides, сопровождаются
разрушением гибридов. При многих межвидовых скрещиваниях
наблюдается стерильность гибридов, сопровождающаяся нару-
.шениями мейоза {96, 428].
Способность хромосом к конъюгации у межвидовых гибри-
дов сильно различается в разных комбинациях скрещиваемых
видов. При скрещивании диплоидных видов, принадлежащих к
одной и той же группе, у потомков Fi конъюгируют все или поч-
ти все хромосомы. В отличие от этого у гибридов от скрещива-
ний между членами различных групп видов способность хромо-
сом к конъюгации понижена. Так, у гибридов между Layla hete-
>rotricha (п=8) и L. glandulosa (n=8) образуется от 0 до 2 би-
валентов, а у гибридов между £. heterotricha (п==8) и L. pla-
tyglossa (n=7) —от 0 до 3 бивалентов [96, 428].
Клаузен [96] полагает, что снижение способности хромосом
к конъюгации у межвидовых гибридов Layia обусловлено ген-
ной дисгармонией между родительскими видами, а не структур-
ными перестройками в их хромосомах. Однако имеются данные,
свидетельствующие о том, что стерильность у Layla вызывается
как хромосомными, так и генными различиями. У гибридов Fi
от скрещивания Layla platyglossa (п=7) и L. pentachaeta
(n=8) образуется от 0 до 3 бивалентов. Эти гибриды путем
удвоения хромосом дали некоторое количество тетраплоидных
потомков с числом хромосом 2п=30. У тетраплоидных растений
хромосомы нормально конъюгировали, образуя 14—15 бивален-
тов; мультивалентов и вторичных ассоциаций, по-видимому, не
было. Последние стадии мейоза были, однако, нарушены, и эти
тетраплоиды оказались стерильными (102, 428].
Сходную картину Клаузен и его сотрудники [96] обнаружили
у родственного Layla многолетника Madia (Compositae)
(рис. 11.3).
Clarkia и др.
Род Clarkia (Onagraceae) содержит много видов однолетних
трав, которые часто встречаются в природе симпатрически. Как
и в случаях с Gilia и Layia, эти виды изолированы мощными
барьерами несовместимости. При некоторых скрещиваниях гиб-
риды оказываются нежизнеспособными. Если же гибриды дости-
гают зрелости, то они бывают более или менее стерильными.
Стерильность в значительной степени обусловлена хромосом-
ными различиями [496].
В сущности такая же картина наблюдается у многих других
родов однолетних растений, например у Brassica (Cruciferae,
Гл. 11. Формы взаимоотношений по плодовитости 139
[916]), Spergularia (Caryophyllaceae, [656]), в группе Atriplex
triangularis (Chenopodiaceae, [320]), в однолетних секциях Ni-
cotiana (Solanaceae, [26, 250]), Collinsia (Scrophulariaceae, [232,
233, 235]), Microseris (Compositae, [92]), Helianthus (Composi-
tae, [374]), Stephanomeria (Compositae, [251]), в секции Cera-
tochloa рода Bromus (Gramineae [332, 794, 795]), Avena (Gra-
mineae, [653]).
Автогамные однолетники
Gilia incons plena
Комплекс Gilia inconspicua образует группа низкорослых авто-
гамных видов с мелкими цветками, относящихся к секции Агас-
hnion рода Gilia (Polemoniaceae). Известно 25 видов, принад-
лежащих к этому комплексу. Они широко распространены по
всем аридным областям западной части Северной Америки, где
встречаются 23 вида, а также в умеренном поясе Южной Аме-
рики. Основное число хромосом у них % = 9 Это обычно диплои-
ды и тетраплоиды; известен также один октоплоид [283].
Виды, входящие в этот комплекс, сходны, хотя и неидентич-
ны по своим морфологическим признакам, и их трудно опреде-
лять обычными таксономическими способами. В гл. 4 были рас-
смотрены и проиллюстрированы черты сходства между пятью
видами комплекса Gilia inconspicua, а именно G transmonta-
na, G. malior, G. minor, G. aliquanta и G. clokeyi. Кроме того,
эти растения часто образуют симпатрические скопления из 2,
3 или 4 видов, вследствие чего некоторые из ранних системати-
ков пришли к логическому заключению, что все эти формы от-
носятся к одному изменчивому виду или двум иптерградирую-
щим видам. В настоящее время на основе данных, полученных
в полевых и экспериментальных исследованиях, установлено,
что группа G. inconspicua представляет собой комплекс видов-
двойников [283].
^Между всеми этими видами почти в любых комбинациях
действуют эффективные барьеры несовместимости (рис. 114).
Если удается получить межвидовые гибриды, то они почти во
всех случаях высоко стерильны (рис 11.4); конъюгация хромо-
сом у гибридов выражена слабо. Несколько групп данных сви-
детельствуют о том, что виды этой группы различаются по хро-
мосомным перестройкам, т. е. о том, что стерильность гибридов
имеет хромосомную природу; однако имеются и другие данные,
говорящие о наличии также и генной стерильности [158, 283,
285,300]. •
140
Часть третья. Генетические и экологические аспекты вида
—-------Скрещивание совершенно не удалось
•——— Гибриды высокостерильны (0 — 9% фертильной пыльцы)
* 1 Гибриды полустерильнЫ (12 — 28% фертильной пыльцы).
Рис. 11.4. Фертильность гибридов при скрещиваниях между видами-двойни-
ками из группы Gilia inconspicua [283].
INT — G. interior; AUO — G. austrooccidentalis; JAC — G. Jacens; MIN —
G. minor; BRE — G. brecciarum; DIEG — G. diegensis; TWE — G. tweedy I;
INC —G. inconspicua sens str.; TRA —G. transmontana; MAL —G. malior;
FLA — G. flavocincta; OPH — G. ophthalmoides; S II — G. sinuata II; S I -
G. sinuata I; MOD —G. modocensis; TET —G. tetrabreccia; C 4x — G. crassi-
folla 4x; C 8x — G. crassifolia 8x; OCH — G. ochroleuca; MEX — G. mexicana;
CLO — G. clokeyi-, BRA — G. «bradbury»-, ALI — G aliquanta.
Erophila, Elymus и др.
Комплекс Erophila verna (Cruciferae) — группа морфологически
сходных автогамных однолетних трав, произрастающих в Евро-
пе. Основное число хромосом х—7. В этом комплексе есть дип-
лоиды (2л =14), полиплоиды и различные анеуплоиды с числом
хромосом вплоть до 2п=64 [901].
Всю эту группу было принято рассматривать как один таксо-
номический вид — Draba verna, или Erophila verna. Ее генети-
Гл. И. Формы взаимоотношений по плодовитости 141
ко-таксономичсский состав анализировали несколько исследова-
телей, а в особенности Винге [901], который считает, что она
состоит из четырех видов-двойников. Эти виды встречаются в
Европе симпатрически в разных попарных комбинациях. Винге
(901] показал, что между видами группы Erophila verna сущест-
вует барьер стерильности, создаваемый несовместимостью и
различиями в структуре хромосом.
Как показывают экспериментальные данные, симпатрические
виды-двойники, разделенные барьером хромосомной стерильно-
сти, существуют также у автогамного многолетнего злака Е1у-
mus glaucus [750, 751].
Судя по цитотаксономическим данным, такие же симпатри-
ческие виды-двойники имеются по крайней мере в трех других
группах автогамных однолетников- Vicia sativa (Leguminosae)
[337], Camissonia micrantha (Onagraceae, [658]) и Mentzelia
albicaulis (Loasaceae).
Общие модели
Наша следующая задача заключается в том, чтобы обобщить
все данные, представленные в этой главе При этом можно вы-
явить следующие общие модели (в качестве примеров приведе-
ны те таксоны, которые подробно рассматривались в предыду-
щих разделах главы).
1. Модель Ceanothus Древесные растения со свободным не-
родственным скрещиванием и более или менее неспециализиро-
ванными цветками. Разные виды совместимы друг с другом,
плодовиты при скрещиваниях, и их хромосомы гомологичны в
широких пределах. В природе виды изолированы или полуизо-
лированы посредством экологических и других внешних факто-
ров Между разными секциями или подродами существует силь-
ный барьер несовместимости. Ceanothus, Quercus, Pibes, Pinus,
Prosopis, Eucalyptus -
2. Модель Aquilegia. Многолетние травянистые растения co
свободным неродственным скрещиванием и специализированны-
ми цветками. Разные виды совместимы друг с другом, плодови-
ты при скрещиваниях, и их хромосомы гомологичны в широких
пределах. Изоляция между видами создается механическими и
этологическими барьерами, а также экологическими и другими
внешними факторами. Между разными секциями, подродами или
родами существует сильная несовместимость. Aquilegia, Antir-
rhinum, Penstemon, Epiphyllum, секция Oncocyclus рода Iris,
-орхидеи.
3. Модель Geum Многолетние травы со свободным нерод-
ственным скрещиванием и умеренными межвидовыми различия-
ми в строении цветков. Разные виды совместимы друг с другом
в широких пределах, но между секциями или подродами суще-
ствуют эффективные барьеры несовместимости. Виды, принад-
лежащие к одной и той же группе видов, интерфертильны и их
хромосомы гомологичны; гибриды между видами, относящимися
к разным группам, характеризуются хромосомной или генной
стерильностью. Родственные виды изолированы в природе эко-
логическими или другими внешними факторами. Изоляция сход-
на с наблюдаемой у типа 2, но она в большей мере обусловлена
хромосомной и генной стерильностью, чем механизмами, созда-
ваемыми строением цветка. Geum, Solatium, секция Apogon рода
Iris, Silene секция Simiolus рода Mimulus, Vernonia.
4. Модель Madia. Однолетние травы с разнообразными систе-
мами размножения — от свободного неродственного скрещива-
ния до инбридинга. Близкие виды обычно изолированы барьера-
ми несовместимости, а также хромосомной и генной стерильно-
сти. Brassica, Spergularia, Atriplex, Clarkia, Gilia, Nicotiana,
Collinsia, Layia, Madia, Microseris, Helianthus, Stephanomeria,
Bromus, Avena.
5. Модель Gilia inconspicua. Однолетние или, реже, много-
летние травы с автогамной системой размножения. Отдельный
таксономический вид состоит из нескольких или многих видов-
двойников, изолированных барьерами несовместимости, а также
хромосомной и генной стерильностью. Gilia inconspicua, Его phi-
la verna, Elymus glaucus, Vicia sativa, Camissonia micrantha,
Mentzelia albicaulis.
Большая часть генетико-таксономических исследований про-
водилась на группах растений, встречающихся в северном уме-
ренном поясе. О природе барьеров, создаваемых стерильностью,
и их распространении среди растений южного умеренного и тро-
пического поясов известно гораздо меньше. Это отразилось и
на подборе родов, рассмотренных в данной главе. Поэтому те
типы, которые здесь были выделены, отнюдь не исчерпывают все
возможности, и в будущем число их, несомненно, возрастет в
результате дальнейших генетико-таксономических исследований.
Тропический американский древесный род Theobroma (Ster-
culiaceae) отличается высокой межвидовой несовместимостью,
подобно умеренным однолетним родам, и значительной нежиз-
неспособностью гибридов [2].
В Австралии некоторые группы древесных растений, такие,
как Podocarpus, Proteaceae, некоторые Myrtaceae и некоторые
Epacridaceae, претерпели дифференциацию в отношении струк-
туры хромосом, сравнимую с наблюдаемой во многих группах
травянистых растений северных флор (325, 413, 414, 746, 747].
Цитологические и морфологические данные позволяют считать,
что австралийской флоре, возможно, присуще некоторое своеоб-
разие, заключающееся в наличии в ней древесных растений со
специализированными цветками, опыляемыми животными, и с
дифференциацией по структуре хромосом внутри родов и меж-
ду ними.
Другая интересная и своеобразная черта австралийской и
новозеландской флоры связана с родом Epilobiutn (Onagraceae),
к которому относятся автогамные многолетние травы. Плодови-
тость при скрещиваниях и гомологичность хромосом — общее
правило для большинства межвидовых комбинаций в этой груп-
пе [666].
Рассматривая модели, которые мы идентифицировали и об-
судили выше, следует отметить ряд общих черт в генетических
системах филогенетически неродственных друг другу родов.
Черты сходства между Ceanothus, Quercus и Pinus или между
Madia, Clarkia и Gilia, а также коллективные различия между
этими группами родов выражены очень резко. Выделенные ти-
пы, по-видимому, отражают подлинные конвергенции между
членами разных семейств растений.
Общие корреляции
Для того чтобы создать более широкую основу для наших обоб-
щений, можно использовать более обширный набор групп рас-
тений, чем тот, который мы рассматривали до сих пор в этой
главе. Несколько лот назад я просмотрел литературу по 72 груп-
пам (родам, секциям или подтрибам) растений, которые изуча-
лись в генетико-таксономическом аспекте. Я распределил эти
группы растений по разным категориям на основе наличия или
отсутствия несовместимости и барьеров стерильности между
родственными группами. Эти же самые филады можно также
сгруппировать в три класса в соответствии с жизненными
формами
Корреляция между жизненной формой и типом взаимоотно-
шений по плодовитости представлена в табл 111. Рассмотрим
сначала распределение барьеров несовместимости. Виды, при-
надлежащие к одной и той же секции или группе видов, обычно
легко скрещиваются между собой у древесных растений и мно-
голетних трав, но бывают разделены барьерами несовместимости
у однолетних трав (табл. 11.1).
В распространении барьеров стерильности выявляется та же
самая корреляция с жизненной формой Гибриды Fi, получен-
ные при скрещиваниях между родственными видами, обычно
плодовиты или частично плодовиты у древесных растений и
многолетних трав, но у однолетних трав они в большинстве слу-
чаев стерильны (табл 11.1).
К подобным же выводам пришли и другие авторы [143, 144,
144 Часть третья Генетические и экологические аспекты вида
Таблица 11.1. Распространение барьеров
несовместимости и стерильности в группах растений,
относящихся к различным жизненным формам (по
[280] с изменениями)
Жизненная форма Число филад, в которых скрещивания между близкими видами обычно приводят к следующим результатам
скрещиваемость стерильность гибридов
высокая низкая высокая низкая
Деревья и кустарники 19 2 11 2
<7 Многолетние травы 28 0 15 6
Однолетние травы 3 19 3 19
194, 783], основываясь на независимом изучении имеющихся
данных.
В нескольких случаях оказалось возможным сравнивать раз-
ные секции одного и того же рода в отношении корреляции
между жизненными формами и эффективности барьеров сте-
рильности. У рода Helianthus многолетние виды, составляющие
группу Н. giganteus, оказались интерфертильными, тогда как
однолетники, относящиеся к группе И. annuus, интерсте-
рильны [374].
У Crepis (Compositae) члены трех секций однолетних трав,
а именно Hostia, Phytodesia и Netnaucherces, при скрещивании
друг с другом дают стерильные гибриды. В отличие от этого
скрещивания между многолетними видами секции Lepidoseris
в большинстве случаев дают плодовитые гибриды; однако в
другой секции многолетников (Berinia) межвидовые гибриды во
многих случаях стерильны [33].
Однолетние виды рода Knautia (Dipsacaceae), относящиеся
к секциям Tricheroides и Knautia, хорошо различаются по своим
геномам и изолированы друг от друга эффективными внешними
и внутренними репродуктивными барьерами. Многолетние же
виды, принадлежащие к секции Tricherа, напротив, сходны по
своим геномам, и репродуктивная изоляция между ними выра-
жена слабо [190].
Так случилось, что- большинство деревьев, кустарников и
многолетних трав, включенных в наш обзор, размножаются пу-
тем свободного неродственного скрещивания, тогда как многие
из рассмотренных групп однолетников содержат частично или
преимущественно самоопыляющиеся формы. Возможно поэтому,
что наблюдаемая в табл. 11.1 корреляция между стерильностью
и жизненной формой отражает на самом деле более глубокую
зависимость между типом системы размножения и образованием
барьера стерильности.
Гл 11 Формы взаимоотношений по плодовитости 14S
Выводы
Многие группы многолетних растений северной умеренной зоны
характеризуются своеобразным типом изоляции: близкие виды
обычно плодовиты при скрещиваниях друг с другом, и их изоля-
ция в природе обеспечивается главным образом экологическими
и другими внешними факторами; но изоляция между более да*
лекими видами, принадлежащими к разным секциям, подродам
или родам, обеспечивается внутренними барьерами, блокирую-
щими скрещивание или же успешное развитие гибридных за-
родышей.
Эти барьеры, создаваемые несовместимостью и нежизнеспо-
собностью и контролируемые генами, возникают в таксономиче-
ской структуре данной группы растений внезапно. Внутренние
каналы, по которым происходит обмен генами между видами,,
открыты в довольно широких таксономических пределах, но за
этими пределами они становятся закрытыми.
Для многих однолетних и некоторых многолетних трав уме-
ренной зоны северного полушария характерен иной тип изоля-
ции. У них между близкими видами обычно действуют внутрен-
ние изолирующие механизмы, создающие несовместимость и
гибридную стерильность. Кроме того, в этих группах преобла-
дает стерильность, обусловленная структурой хромосом. Неред-
ко у них существуют также внешние изолирующие механизмы
и генная стерильность, однако эти последние играют лишь вто-
ростепенную роль.
Можно считать, что группы видов, плодовитые при скрещи-
ваниях, и группы видов, стерильные при скрещиваниях, облада-
ют некоторыми общими для тех и других основными механизма-
ми изоляции. Экологическая изоляция, репродуктивная изоля-
ция, создаваемая внешними механизмами, и различные барьеры,
контролируемые генами, имеются и у тех и у других групп расте-
ний. Одно из главных различий между ними — интерполяция
хромосомной стерильности в пределах некоторого круга родст-
венных форм в одном случае и отсутствие этого—в другом [280].
10-45
&
Часть четвертая
Эволюционная дивергенция
12 Способы видообразования
Виды различаются, как мы видели в гл. 8, по адаптивным приз-
накам, контролируемым сложными комбинациями генов. Сле-
довательно, видообразование сопряжено с возникновением но-
вых и различных комбинаций генов в обособленных популяциях.
Этот аспект процесса видообразования относится к эволюцион-
ным видам вообще. Что же касается биологических видов, со-
стоящих из перекрестно оплодотворяющихся организмов, то у
них видообразование связано, кроме того, с развитием репродук-
тивных изолирующих механизмов.
При обсуждении видообразования удобно рассматривать
упомянутые выше события — развитие новых генных комбина-
ций и изолирующих механизмов — как завершение процесса ви-
дообразования. Теперь мы можем перейти к рассмотрению сил,
обусловливающих процесс видообразования, и полей, на кото-
рых протекает этот процесс.
Силы и поля
Эволюционные силы, участвующие в видообразовании, можно
разделить на две группы: силы, создающие изменчивость, и си-
лы, закрепляющие возникшие изменения в дочерних популя-
циях. Ниже перечислены силы, создающие изменчивость и иг-
рающие определенную роль в процессе видообразования; каж-
дой из этих сил дано буквенное обозначение, которым мы будем
пользоваться в дальнейшем.
Изменчивость, существующая в обычных условиях (и)
Гибридизация (Л)
Новые мутации (и)
Силы, которые закрепляют изменчивость и значение которых
для видообразования либо установлено, либо предполагается
^некоторыми учеными, сводятся к следующим.
Отбор с широким свободным неродственным скрещива-
нием ($)
Гл. 12 Способы видообразования
147
Инбридинг в небольших популяциях (и отбор) (tp)
Инбридинг в результате самооплодотворения (и отбор)
(if)
Инбридинг в результате ассортативного скрещивания
(М
Силы, создающие изменчивость, и силы, закрепляющие эту
изменчивость, могут комбинироваться самыми разнообразными
способами, в результате чего возникают различные двустадий-
ные процессы, например v—s, v—ip и т. п.
Поля, на которых могут происходить процессы видообразо-
вания, бывают аллопатрическими, смежно-симпатрическими и
биотически-симпатрическими. В результате сочетания этих трех
полей с различными комбинациями сил возникают многочислен-
ные способы видообразования.
Способы видообразования
Число возможных комбинаций из 3 типов сил, создающих из-
менчивость, 4 типов сил, закрепляющих эту изменчивость, и
3 типов полей, конечно, достаточно велико, по многие из этих
возможных способов нереалистичны и ими можно пренебречь.
Остается ряд способов, из которых одни известны, другие веро-
ятны, но существование их не установлено, а третьи хотя и спе-
кулятивны, но часто упоминаются в текущей литературе.
Ниже перечислены, с краткими замечаниями, известные, ве-
роятные или серьезно обсуждаемые способы видообразования.
1. Аллопатрическое поле, v—s Географическое видооб-
разование. Хорошо известно.
2. Аллопатрическое поле, v—ip. Квантовое видообразо-
вание. Известно.
3 Аллопатрическое поле, v—if. Аллопатрическое видо-
образование, связанное с самооплодотворением. Сход-
но с квантовым видообразованием Известно у расте-
ний.
s 4. Смежно-симпатрическое поле, v—ip Смежно-симпат-
рическое видообразование. Весьма вероятно у насеко-
мых со свободным неродственным скрещиванием и у
других беспозвоночных, занимающих разные узкие,
нищи и сосуществующих в одном и том же местооби-
тании.
5 Биотически-симпатрическое поле, h—u—i. Аллопо-
липлоидное видообразование. Хорошо известно у рас-
тений.
6. Биотически-симпатрическое поле, h—i Рекомбинаци-
онное видообразование и гибридное видообразование
148 Часть четвертая. Эволюционная дивергенция
с сегрегацией внешних барьеров. Весьма вероятно у
растений.
7. Биотически-симпатрическое поле, v—if. Биотически-
симпатрическое видообразование у самооплодотво-
ряющихся организмов. Вероятно для некоторых рас-
тений, беспозвоночных и протистов.
8. Биотически-симпатрическое поле, v—s или v—ia.
Биотически-симпатрическое видообразование у сво-
бодно скрещивающихся организмов при действии диз-
руптивного отбора или ассортативного скрещивания.
Некоторые ученые рассматривают этот способ как
реально возможный.
Обсуждение
Описанная выше система классификации способов видообразо-
вания на основе поля и действующих сил взята из моей более
ранней работы [282]. Эта система носит общий характер и при-
ложима к животным, растениям и протистам. Со времени ее
первого опубликования в 1963 г. появились три другие общие
классификации способов видообразования [77, 894, 913].
Система Буша [77] основана на географическом поле; систе-
ма Уайта [894] — на поле и наличии или отсутствии хромосом-
ных перестроек; система Райта [913] —на поле, а также на
том, является ли дивергенция первичной или она связана с гиб-
ридизацией. Кроме того, между этими системами имеются па-
раллелизмы, перекрывания и различия, так что подробное срав-
нение их завело бы нас слишком далеко. И все же здесь сле-
дует сделать несколько частных замечаний.
Один из выделяемых Бушем [77] способов видообразова-
ния— это парапатрическое видообразование. Я не пользуюсь
этим термином, потому что парапатрия, в сущности, представ-
ляет собой одну из форм аллопатрии, и, следовательно, в этом
термине нет необходимости. Парапатрическое видообразование
в том смысле, какой в него вкладывает Буш [77], по-видимому,
соответствует моему «аллопатрическому видообразованию у си-
дячих организмов» (см. [282]), а это один из подтипов географи-
ческого видообразования. В системе Уайта {894], претендующей
на приложимость ко всем высшим организмам, упущено не-
сколько способов видообразования, играющих важную роль у
высших растений, а поэтому она не совсем применима при изу-
чении эволюции растений.
Уайт [894] выделяет особый способ видообразования, кото-
рый он называет стасипатрическим видообразованием. При ста-
сипатрическом видообразовании в области, занимаемой данным
лидом, появляется новая хромосомная перестройка, которая об-
Гл. 12. Способы видообразования 149
разует в этой области новую гомозиготу по структурной пере-
стройке. Этот способ видообразования Уайт противопоставляет
аллопатрическим способам видообразования. Я считаю данные,
на основе которых выделяется стасипатрическое видообразова-
ние, неубедительными, а сам способ гипотетическим, хотя Уайт,
очевидно, со мной не согласится. По крайней мере некоторые
из приводимых им примеров можно так же хорошо или даже
лучше объяснить как случаи квантового видообразования, свя-'
занные с хромосомными перестройками.
Наша собственная система, изложенная выше, дает приемле-
мую основу для обсуждения способов видообразования у расте-
ний. Обратите внимание на то, что способы 5 и 6 в нашем пе-
речне представляют собой типы гибридного видообразования
у растений. Они будут обсуждаться в дальнейших главах, в пя-
той части книги. Другие способы —это аспекты первичной эво-
люционной дивергенции, которые рассматриваются в следую-
щей главе под названием первичного видообразования.
13 Первичное видообразование
Под первичным видообразованием здесь имеется в виду про-
цесс эволюционной дивергенции между популяциями до видово-
го уровня. Мы относим к первичному видообразованию все явле-
ния, обусловливающие первичную эволюционную дивергенцию,
и исключаем из него явления гибридного видообразования и
усиления изоляции, которые обсуждаются отдельно в последую-
щих главах.
Процессу видообразования в качестве начального условия
необходим запас генетической изменчивости. Такая изменчи-
вость представляет собой всеобщее явление в органическом ми-
ре. Существует три уровня изменчивости, важных для видообра-
зования; полиморфизм, локальная расовая изменчивость и геог-
рафическая изменчивость. Эти типы изменчивости уже были
описаны в гл. 2 и 3.
Соотношение между разными уровнями изменчивости опре-
деляет путь первичного видообразования. Наиболее хорошо
известен и документирован путь от полиморфизма через локаль-
ные, а затем географические расы до видов. Этот путь хорошо
описывается географической теорией видообразования, которую
мы рассмотрим более подробно в следующем разделе этой гла-
вы. Следует также рассмотреть альтернативные сокращенные
путивозникновения новых видов. Путь, при котором выпадает
формирование географических рас, носит название,_квантового
видообразованияГв таких случаях новый вид развивается из
какой-либо локальной расы. Цуть биологически-симпатрическо-
ХО^видообраэювания~ббхЪДО? как локальные, так и географиче-
ские~тгасБГ~Предполагается, что. новый вид развивается непо-
средственно из какой-либо вариации_,п^Ш1мбрфного вида. Пер-
вйчноё~вигДбобразование в биотически-симпатрическом поле —
дискуссионная проблема, которую мы попытаемся разобрать в
одном из дальнейших разделов этой главы.
Географическое видообразование
Географическая теория видообразования имеет давнюю исто-
рию. Ее основные концепции неоднократно формулировались с
разными дополнениями и разъяснениями рядом зоологов, в том
числе такими выдающимися учеными, как М. Вагнер [863],
К. Джордан [421]' Д. С. Йордан [420], Ренш [676J, Добржан-
ский [166] и Майр3 [564]. Обзор этой проблемы в историческом
плане представлен в работе Майра [569].
Гл. 13. Первичное видообразование
151
Теория географического видообразования в ее современной
форме утверждает, что географические расы — это предшествен-
ники видов в непрерывном процессе эволюционной дивергенции
(см рис. 7.1). Пространственная изоляция популяций на расо-
вой стадии дивергенции создает” возможность для развития и
поддержания у отдельных популяций комбинаций генов, опреде-
ляющих отличительные морфологические и физиологические
признаки этих популяций. На этой же стадии начинает закла-
дываться репродуктивная изоляция. А формирование видов
представляет собой дальнейшее развитие указанных процессов.
Репродуктивные изолирующие механизмы, развивающиеся в
течение периода пространственной изоляции двух или несколь-
ких популяций, позволяют этим популяциям сосуществовать
друг с другом, нс скрещиваясь в тех случаях (если таковые воз-
никают), когда в результате расширения своих ареалов эти по-
пуляции оказываются в одной и той же области.
Данные в пользу этой теории можно вкратце суммировать,
следуя Майру [564]: 1) между признаками, разделяющими геог-
рафические расы, и признаками, разделяющими виды, нет ка-
чественных различий; 2) во многих группах обычны системы
популяций, занимающие промежуточное положение между геог-
рафическими расами и видами, т. е. полувиды; 3) известны слу-
чаи, когда в одной части ареала данного вида ряд интергра-
дирующих рас замещают друг друга географически, а в другой
его части симпатрически перекрываются; 4) между географиче-
скими расами одного вида часто можно обнаружить частичную
репродуктивную изоляцию.
Примеры промежуточных стадий между географическими
расами и видами, о которых говорится в пунктах 2 и 3, приво-
дились в гл. 6 Пункт 4 обсуждается более подробно далее в
настоящей главе Здесь мы опишем пример, в котором все ста-
дии дивергенции, требуемые географической теорией видообра-
зования, можно обнаружить в пределах одной естественной
группы растений
Этим примером служит группа родственных между собой рас
и видов диплоидных однолетних растений, распространенных в
пустынных и гористых районах на юго-западе США и принад-
лежащих к более обширной секции Arachnion рода Gilia. Мор-
фологические, экологические, географические и цитогенетиче-
ские взаимоотношения между этими расами и видами рас-
смотрены Грантом и Грант [263, 300].
Наиболее ярко выраженные географические расы Gilia lati-
flora хорошо дифференцированы морфологически. Эти расы ип-
терфертильны и непрерывны, а поэтому они связаны в природе
рядом интерградирующих форм (рис. 13 1, А). Близкий к G. 1а-
tiflora вид G leptantha состоит из ряда географических рас, ко-
О 75 150
Километры
Рис 13.1. Пять стадий
дивергенции у видов ги-
лий секции Arachnion
(Polemoniaceae) в юго-
западной части Север-
ной Америки.
А. Непрерывные геогра-
фические расы. Б. Дизъ-
юнктивные географиче-
ские расы. В. Маргиналь-
по-симпатрические полу-
виды. Г. Аллопатрические
виды. Д. Симпатрические
PHAWt
Гл. 13. Первичное видообразование
153
торые также различаются морфологически и интерфертильны,
но занимают разобщенные области, в связи с чем между ними
нет непрерывной интерградации (рис. 13.1, Б).
Для Gilia latiflora, G. leptantha и родственного им третьего
полувида G tenuiflora характерно в значительной степени алло-
патрическое распространение с перекрыванием в пограничных
районах (рис 13.1,В). Морфологические и экологические раз-
личия между этими тремя полувидами более выражены, чем
между расами в каждом из них. Способность к скрещиванию
между разными подвидами и плодовитость образующихся при
этом гибридов также понижены по сравнению с тем, что наблю-
дается при межрасовых скрещиваниях. Тем не менее пути для
обмена генами остаются открытыми. В некоторых областях сим-
патрического контакта эти полувиды гибридизируют; в других
областях симпатрии они остаются обособленными.
Gilia ochroleuca и G. mexicana занимают совершенно отдель-
ные области (рис. 13 1, Г) Эти два вида разделены мощным
барьером стерильности и сильно выраженной прерывистостью
морфологических признаков
Gilia ochroleuca симпатрична на юге Калифорнии с синга-
меопом G. tenuiflora—leptantha—latiflora, и особи всех этих
форм во многих местах растут вместе и цветут одновременно
(рис. 13.1, Д). Морфологическая прерывистость между этими
двумя популяционными системами сохраняется на протяжении
всей обширной зоны симпатрии. Преграды, препятствующие
скрещиваниям между этими двумя формами, и стерильность
гибридов выражены в значительной степени.
Квантовое видообразование
Квантовое видообразование заключается в том, что от предко-
вого вида какого-либо свободно скрещивающегося организма
отпочковывается новый и дивергентный вид, проходя через про-
межуточную стадию географически изолированной или полуизо-
дированной периферической популяции. Силами, закрепляющи-
ми изменчивость, служат отбор и дрейф генов, действующие
совместно в маленькой периферической дочерней популяции.
Такое определение процесса квантового видообразования соот-
ветствует определению, данному ранее Грантом [282J.
Желательно помнить это первоначальное определение кван-
тового видообразования, поскольку за последнее время в лите-
ратуре появилось несколько совершенно ошибочных определений
этого процесса [29, 8921. Ошибки возникли из-за того, что Уайт
[894] не читал работу Гранта [284], а Айала и Валентайн [29]
не читали книгу Добржанского и др [171].
154
Часть четвертая. Эволюционная дивергенция
Квантовое видообразование было впервые описано Майром
[565] в его оригинальной и важной статье. Формулировке*этого
способа предшествовали две более ранние концепции Майра, а
именно — концепция аберрантных признаков в периферических
популяциях и принцип основателя [564], а также его новая в то
время концепция генетических революций [565].
Обсуждая способы видообразования в работе 1963 г. [282],
я принял модель Майра [565], придавая ей очень большое зна-
чение, но присоединил к ней также две другие концепции, со-
вершенно очевидно имеющие самое непосредственное отношение
к этому способу видообразования. Эти ранние концепции, кото-
рым Майр [565] не уделил должного внимания, —концепция
Райта [906] о взаимодействии отбора с дрейфом генов и кванто-
вая эволюция Симпсона [730]. Обе концепции относились к мел-
ким изолированным популяциям и обе предсказывали, что в
таких популяциях создаются чрезвычайно благоприятные усло-
вия для быстрых и коренных эволюционных изменений. При
создании этих концепций их никак не связывали с процессом
видообразования, однако они были вполне совместимы с теория-
ми видообразования. Концепция взаимодействия отбора с дрей-
фом генов предусматривала движущую силу, необходимую для
эволюционной дивергенции. Термин квантовое видообразование,
как мне казалось, соответствует комбинации участвующих в
этом процессе факторов, поэтому в 1963 г., обсуждая указанный
процесс в своей книге, я впервые ввел этот термин [282].
Квантовое видообразование можно сравнить с географиче-
ским видообразованием. Но если географическое видообразова-
ние — процесс постепенный и консервативный, то квантовое ви-
дообразование вызывает быстрые и коренные изменения гено-
типа или фенотипа или же одновременно и того и другого.
Мы исходим из того, что поток генов в более или менее неп-
рерывных центральных популяциях какого-либо перекрестно-
опыляющегося вида оказывает поглощающее действие на новые
вариации. Кроме того, эти условия благоприятствуют генам, об-
ладающим хорошей комбинационной способностью, что позво-
ляет им смешиваться с чужими генами, поступающими с непре-
рывным потоком генов. Если же несколько особей из такой
популяции оказываются в стороне от основного русла потока
генов в виде полуизолированной периферической популяции (на-
пример, вновь основанной колонии), то это ведет к инбридингу
с сопровождающими его резко выраженными генетическими и
фенотипическими последствиями. В большинстве случаев такие
резкие изменения скорее будут иметь низкую адаптивную цен-
ность, что повлечет за собой вымирание этой популяции. Однако
в одной или н^кольких изолированных периферических попу-
ляциях могут произойти радикальные изменения, которые при-
Гл 13 Первичное видообразование
155
вед>т к возникновению нового хорошо адаптированного состоя-
ния, и в результате подобная популяция даст начало отклоняю-
щемуся дочернему виду [565, 569].
Этот новый дочерний вид, если он сможет выжить в природе,
будет разрастаться, и по мере увеличения его численности нач-
нет возрастать запас его изменчивости Со временем он даст
начало новому поколению дочерних видов, которые будут воз-
никать путем квантового видообразования.
Характер изменчивости, наблюдаемой у многих видов или
групп видов, свидетельствует в пользу квантового видообразо-
вания В основной части такого вида, занимающей центральную
область ареала, наблюдается географическая изменчивость кли-
нального и консервативного типа. Но по периферии ареала
предкового вида в географически изолированных местах встре-
чаются расы и виды с отклоняющимися признаками. Майр
[564, 565, 569] приводит в качестве примера ракетохвостых зи-
мородков (Tanysiptera galatea), обитающих на Новой Гвинее
и прилежащих островах, и дронго (Dicrurus hottentotus).
Позднее Карсон [83—86] получил цитогенетические и био-
географические данные о квантовом видообразовании у Drosop-
hila и, в частности, у дрозофил Гавайских островов.
Квантовое видообразование у Clarkia
В результате биосистематических и цитогенетических исследо-
ваний однолетнего рода Clarkia (Onagraceae), проведенных
Люисом и его сотрудниками [493, 494, 496, 497, 499, 859], было
обнаружено несколько пар видов, взаимоотношения между ко-
торыми явно относятся к типу родители — потомки. Так, совер-
шенно очевидно, что Clarkia biloba и С. Ungulata филогенети-
чески представляют собой соответственно предковый и дочер-
ний виды [499]. То же самое относится к таким парам видов,
как С. rubicunda и С. franciscana; С. unguiculata и С exilis;
С. mildrediae и С. virgata-, С. mildrediae и С. stellata [493, 494,
859].
s Дочерний вид отличается от предкового по морфологическим
признакам, экологическим предпочтениям, расположению участ-
ков в хромосомах или же по всем трем характеристикам одно-
временно и нередко занимает ограниченную географическую об-
ласть, расположенную на периферии ареала предкового вида
[493, 499].
Люис [493] отмечает, что периферические популяции в эко-
логически краевых местообитаниях данного вида в климатиче-
ски неблагоприятные годы бывают обречены на полное или поч-
ти полное вымирание. Снижение численности, близкое к выми-
ранию, с последующей регенерацией популяции из одной или
156 Часть четвертая. Эволюционная дивергенция
нескольких генотипически аберрантных выживших особей иног-
да непосредственно ведет к быстрому образованию дивергент-
ного дочернего вида (493]. Иными словами, быстрое видообра-
зование бывает иногда результатом резких колебаний числен-
ности популяций в периферических областях. Люис [493] припи-
сывает такие быстрые эволюционные изменения действию край-
не сильного, или «катастрофического», отбора, однако правиль-
нее рассматривать эти изменения как результат совместного
действия отбора и дрейфа генов.
Очень часто можно обнаружить, что в пределах одного вида
Clarkia периферические популяции отклоняются по морфологи-
ческим и хромосомным признакам. Такие полуизолированные
периферические локальные расы служат примером популяций,
которые иногда могут дать начало новому дочернему виду с
отклоняющимися признаками [493, 859].
Хромосомные перестройки, по которым различаются разные
виды Clarkia, не похожи на те, по которым различаются геогра-
фические расы одного и того же вида. И это еще раз свидетель-
ствует о том, что многие новые виды Clarkia возникли не из
географических рас, а из отклоняющихся периферических попу-
ляций [493].
Пример Clarkia biloba и С. Ungulata был подробно проана-
лизирован Люисом и Робертсом [499]. Это самосовместимые, но
преимущественно свободно скрещивающиеся однолетние травы.
Они очень близки морфологически и генетически, но разделены
барьером хромосомной стерильности. Главное морфологическое
отличие касается формы лепестков, которые у С. biloba сердце-
видные, а у С. Ungulata в виде язычков. Число хромосом у
С. biloba 2п=16, а у С. Ungulata 2п=18 [499]. Эти виды очень
сходны также по электрофоретическим характеристикам {254].
На филогенетическую взаимосвязь между этими двумя ви-
дами указывают число хромосом и экологические
предпочтения. У Clarkia biloba и=8, что близко к основному
числу х=7 для рода Clarkia, тогда как у эволюционно более
продвинутого С. Ungulata n=9. С. biloba занимает более влаж-
ные местообитания, чем С. Ungulata, и, таким образом, она и
экологически ближе к предковой форме. Цитотаксономические
и экологические исследования предоставляют нам достаточно
веские данные, позволяющие считать, что вид С. Ungulata про-
изошел от С. biloba [499].
Географические взаимоотношения между этими видами поз-
воляют сделать дальнейшие заключения относительно того, как
происходило их образование в филогенезе. Clarkia biloba зани-
мает обширную область в северной части Сьерра-Невады и при-
лежащих $ ней районах Калифорнии. С. Ungulata, напротив,
занимает очень небольшую область в одной из речных долин на
Гл. 13. Первичное видообразование
157
южной оконечности ареала С biloba. Такое ограниченное и пе-
риферическое распространение С. Ungulata позволяет предпо-
лагать, что этот вид возник от предкового вида С. biloba очень-
быстро и совсем недавно [499].
Цитогенетический анализ межвидовых гибридов выявляет
характер и степень хромосомных изменений, обусловливающих
дифференциацию Clarkia Ungulata от С biloba. Четыре из
хромосом, входящих в хромосомные наборы этих двух видов,
гомологичны и образуют при мейозе биваленты. Две другие
хромосомы каждого набора образуют у гибрида кольцо из че-
тырех хромосом, что указывает на существующие у этих двух
видов различия по реципрокной транслокации именно в этих
хромосомах [499].
Остальные хромосомы образуют у гибрида цепь из пяти хро-
мосом. Дополнительная девятая хромосома Clarkia Ungulata
располагается в середине цепи; это свидетельствует о том, что
она представляет собой третичный трисомик. Таким образом,
у предков С. Ungulata должна была произойти вторая и неза-
висимая транслокация, и одна из участвующих в ней хромосом-
сохранилась в гомозиготном состоянии, т. е. в виде третичного
тетрасомика, в результате чего у современной С. Ungulata по-
лучился набор из девяти хромосом. Кроме того, лишняя девятая
хромосома С. Ungulata, очевидно, несет гены, детерминирующие
характерную для этого вида форму лепестков [499].
Итак, в генетической дифференциации Clarkia biloba и
С. Ungulata важную роль играют две отдельные транслокации.
Их совместное действие и создает в основном барьер стериль-
ности между этими двумя видами. Та транслокация, которая
сохранилась в виде третичного тетрасомика, вызвала анеуплоид-
ное увеличение числа хромосом от ц = 8 у С. biloba до п = 9 у
С. Ungulata. Лишняя девятая хромосома у С. Ungulata обуслов-
ливает основное морфологическое различие между этим видом
и предковым видом С. biloba [499].
Солбриг [755] также постулировал происхождение некоторых
видов Gutierrezia (Compositae) в результате квантового видо-
образования.
Аллопатрическое видообразование, связанное
с самооплодотворением
У многих самосовместимых и преимущественно свободно скре-
щивающихся растений тем не менее в какой-то степени проис-
ходит самооплодотворение. У таких растений эффекты инбри-
динга в дочерних популяциях могут быть обусловлены системой
размножения, а не величиной популяции. Результатом является
358 Часть четвертая. Эволюционная дивергенция
видообразование (способ 3; см. гл. 12), очень сходное с кванто-
вым видообразованием (способ 2). Очевидное различие между
этими способами состоит в сочетании сил: v — if вместо v — ip.
Кроме того, у растений, способных к самоопылению, эффекты
инбридинга выражены обычно менее резко, чем у растений с
облигатным перекрестным опылением.
Во многих случаях прохождение популяции через «бутылоч-
ное горлышко» низкой численности и самоопыление могут про-
исходить одновременно и совместно воздействовать на изменчи-
вость в дочерней популяции. На периферии предкового вида в
результате закрепления изменчивости вследствие инбридинга
или ограниченных размеров популяции или того и другого мо-
жет возникнуть новая локальная раса или узко эндемичный вид.
Clarkia biloba и С. Ungulata самосовместимы, и вполне возмож-
но, что в процессе формирования С. Ungulata наряду с ограни-
чением размеров популяции происходило некоторое самоопыле-
ние.
Аллопатрическое видообразование у растений, связанное с
•самоопылением, также очень близко к симпатрическому видооб-
разованию с самоопылением (способы 3 и 7; см. гл. 12). В неко-
торых случаях имеющихся данных недостаточно, для того чтобы
провести различие между этими двумя способами.
Из этих двух возможностей в большинстве случаев более ве-
роятно видообразование на каком-либо периферическом
участке за пределами основного ареала предкового вида. Если
дочерняя популяция в какой-то степени способна к свободному
•скрещиванию, как это характерно для большей части автогам-
ных растений, то в случае симпатрии с предковым видом ее от-
клоняющиеся вариации будут поглощаться, а в случае аллопат-
рии они могут сохраниться и закрепиться. Кроме того, в неко-
торых случаях биогеографические данные свидетельствуют в
пользу какого-либо аллопатрического периферического участка.
Вполне возможно, однако, что существуют примеры действи-
тельно симпатрического видообразования при участии самоопы-
ления и следует вести поиски таких примеров.
Вероятным примером способа 3 служат виды Gilia tenuiflora,
<?. austrooccidentalis и G. jacens. Эти три диплоидных вида
•встречаются в южной части Береговых хребтов в Калифорнии
(рис. 13.2); они очень близки друг другу, но морфологически
хорошо различимы. Во всех трех возможных комбинациях они
дают полуплодовитые гибриды Fi со слегка нарушенной конъюга-
цией хромосом [300]. По сравнению с преимущественно свобод-
но скрещивающимся видом G. tenuiflora два других вида не-
сколько упрощены как морфологически, так и в отношении си-
стемы размножения, представляя собой небольшие автогамные
•однолетники. Кроме того, они занимают более аридные место-
Гл. 13. Первичное видообразование
15»
Рис. 13.2. Географическое распространение G. tenuiflora и двух дочерних ав-
тогамных видов в южной Калифорнии.
обитания, чем G. tenuiflora. Следовательно, эволюция, вероятно,,
шла от некой предковой формы, сходной с G. tenuiflora, к
G. austrooccidentalis и G. jacens в качестве дочерних форм.
Географическое распространение этих видов показано на
рис. 13.2. Как можно видеть, Gilia austrooccidentalis и G. jacens
занимают ограниченные и более или менее изолированные обла-
сти к югу от более обширного ареала G. tenuiflora. Принимая
во внимание все изложенные здесь обстоятельства, следует сде-
лать вывод, что G. austrooccidentalis и G. jacens дивергировали
от предкового вида G. tenuiflora в качестве видов, приспособлен-
ных к более аридной среде, и что отправными точками диверген-
ции послужили какие-то краевые в географическом и экологи-
ческом смысле популяции этого предкового вида.
Другой возможный пример — различающаяся по хромосомам
пара видов Clarkia rubicunda и С. franciscana. С. franciscana
представляет собой узко эндемичный автогамный вид, область-
распространения которого примыкает к области распростране-
ния его вероятного предка С. rubicunda в северной части Кали-
форнии [497]. Хотя электрофоретические данные и не подтверж-
дают прямо такое происхождение [253], они и не исключают его.
Еще одним примером служат диплоидные автогамные одно-
летние представители Vicia (Leguminosae) [576], Mimulus;
160
Часть четвертая. Эволюционная дивергенция
(Scrophulariaceae) [435] и Stephanomeria (Compositae) [253,
255, 257]. Mimulus nasutus считается автогамным дочерним ви-
дом свободно скрещивающегося М. guttatus [435]. A Stephanome-
ria malheurensis— автогамный вид, происходящий от широко
распространенного и свободно скрещивающегося 3. exigua сого-
naria [253, 256, 257].
Готлиб [253] высказывает мнение, что Stephanomeria mat-
Sieurensis — автогамный дочерний вид симпатрического проис-
хождения. Действительно, вид S. malheurensis известен только
из одной местности на северной окраине ареала S. exigua сого-
naria в юго-восточной части шт. Орегон. Необходимо обследо-
вать обширную область к северу и востоку от этой местности,
поскольку вполне вероятно, что там будет обнаружен S. mal-
heurensis. Это изменило бы всю биогеографическую картину.
Возможно, что 3. malheurensis зародился в какой-то симпатри-
ческой области, однако более вероятно (по приведенным выше
причинам), что он возник аллопатрически где-либо на перифе-
рии ареала предкового вида, став впоследствии маргинально-
симпатрическим с S. exigua.
Биотически-симпатрическоег видообразование
Стоящая перед нами проблема заключается в том, чтобы уста-
новить, может ли первичная дивергенция на видовом уровне
происходить в пределах одной скрещивающейся популяции. Это
давний, много обсуждавшийся и все еще не решенный вопрос.
Добржанский [168] и Мазер [561] отмечают, что полимор-
физм и сймпатрические виды — это проявление альтернативных
реакций живых организмов на среду, в которой на общей тер-
ритории имеются разнообразные ниши. Далее Мазер (но не
Добржанский) высказал мнение, что виды могут возникать не-
посредственно из полиморфных типов при наличии некоторых
определенных условий отбора. Тодэй и другие исследователи
развивали эту точку зрения, получив некоторые эксперимен-
тальные данные, подтверждающие ее [572, 752, 810, 825, 827].
Майр {565, 569, 570], напротив, возражает против возможности
первичного видообразования в биотически-симпатрической сре-
де. Все высказывания за и против продолжаются на протяжении
многих лет, но проблема до сих пор остается нерешенной.
Попытаемся проанализировать все, что с нею связано, и нач-
нем наш анализ с перечисления и определения круга нерешен-
ных вопросов.
Первичное симпатрическое видообразование ведет от поли-
морфизма к видам по кратчайшему пути, на котором нет проме-
жуточной стадии географической и микрогеографической изоля-
&
Гл. 13. Первичное видообразование
161
ции. Поэтому, во-первых, следует выяснить, возможен ли теоре-
тически такой путь эволюции, во-вторых, вероятен ли он теоре-
тически; и, наконец, в-третьих, действительно ли он существует
в природе. Тодэй и его сотрудники, а также их последователи
занимались двумя первыми вопросами, а Майр — третьим.
Теоретические затруднения, связанные с первичным симпа-
трическим видообразованием, наиболее велики в том случае,
когда популяция основателей состоит из перекрестно оплодотво-
ряющихся особей. В этом случае процессу дивергенции непре-
рывно противостоит скрещивание между дивергентными поли-
морфными типами. В условиях строгого самооплодотворения эти
трудности в значительной мере сглаживаются. Следовательно,
обсуждение теоретической возможности первичного видообразо-
вания следует начинать с рассмотрения этой проблемы в том
виде, в каком она существует в системе, размножающейся при
помощи перекрестного оплодотворения.
Дивергенция между разными полиморфными типами в пе-
рекрестнооплодотворяющейся популяции усиливается дизруп-
тивным отбором [561, 570, 810, 825, 827]. Этот тип отбора дей-
ствует в условиях, когда один аспект гетерогенной среды благо-
приятен для одного полиморфного типа, а другой ее аспект —
для другого. Некоторые эксперименты, проведенные на Droso-
phila melanogaster, показали, что сильный дизруптивный отбор
ведет к дивергенции, несмотря на случайное скрещивание [579,
810, 825, 827]. В проведенных в конце шестидесятых и начале
семидесятых годов экспериментах на комнатной мухе (Musca
domestica) были получены сходные результаты [640, 752].
Напротив, в перекрестнооплодотворяющейся, но панмиктиче-
ской популяции воздействие дизруптивного отбора может про-
должаться все время благодаря системе ассортативного скре-
щивания Тенденция генетически сходных индивидуумов скре-
щиваться друг с другом усиливает дизруптивный отбор.
Для проявления действия дизруптивного отбора в условиях
случайного скрещивания необходимы очень сильные давления
отбора. Давления отбора, применявшиеся в экспериментах на
дрозофиле, варьировали от очень сильных [810] до слабых [242],
которые, однако, в природных условиях все еще рассматрива-
лись как сильные.
Для того чтобы сопоставить результаты экспериментов по
дизруптивному отбору с ситуацией в панмиктических природ-
ных популяциях, следует выяснить, происходит ли сильный от-
бор также и в природе. Данные о существовании в природных
популяциях селективных ценностей порядка 20% или выше бы-
ли получены Фордом [218], который пришел к выводу, что такая
высокая селективная ценность — явление достаточно общее и
обычное.
11-45
162
Часть четвертая. Эволюционная дивергенция
Мне кажется, что вывод Форда основан на ряде примеров,
выбранных для такого специального исследования именно по-
тому, что в них особенно заметно проявляются реакции на дей-
ствие естественного отбора. В природе высокая селективная
ценность — скорее исключение, чем правило. Но как бы то ни
было, сильный отбор преобладает по крайне мере в некоторых
природных популяциях, и в таких случаях дизруптивный отбор
может оказывать свое действие, несмотря на случайное скре-
щивание.
Мы упомянули об ассортативном скрещивании в качестве
возможного вспомогательно фактора. Ассортативное скрещива-
ние имеет потенциально важное значение, потому что оно дает
дизруптивному отбору возможность вызвать эволюционную ди-
вергенцию при более низких интенсивностях отбора, чем это
необходимо в условиях панмиксии. Комбинация ассортативного
скрещивания с дизруптивным отбором открывает более широкие
возможности для симпатрической дивергенции в природе.
Быть может, нам следовало бы, однако, подойти к проблеме
симпатрического видообразования с другой точки зрения. Мы
вправе усомниться в допустимости экстраполяции от полимор-
физма к видообразованию. Эти две формы симпатрической ге-
нетической дифференциации, несомненно, имеют некоторые об-
щие черты. Но между ними существуют также качественные
различия.
Полиморфизм представляет собой изменчивость такого типа,
при котором различия между видами обусловлены различиями
в сложных комбинациях генов. Скрещивания между двумя хо-
рошо адаптированными полиморфными формами, по всей веро-
ятности, дадут некоторое количество менее хорошо адаптиро-
ванных потомков. Однако при скрещивании между двумя хоро-
шо адаптированными видами доля плохо адаптированных потом-
ков будет несравненно выше. Отсюда следует, что плата за диз-
руптивный отбор, оцениваемая по генетической смерти, при ис-
тинном симпатрическом видообразовании должна быть гораздо
выше, чем при поддержании и усилении полиморфной системы,
и вполне может оказаться чрезмерно высокой для большинства
природных популяций.
Итак, мы не располагаем биогеографическими данными, ко-
торые свидетельствовали бы о существовании у перекрестно-
оплодотворяющихся организмов биотически-симпатрического
видообразования, хотя для географического и квантового видо-
образования такие данные имеются. Кроме того, постулируя
видообразование путем дизруптивного отбора в перекрестно-
оплодотворяющейся популяции, мы сталкиваемся с очень боль-
шими затруднениями теоретического характера, которые сгла-
живаются при рассмотрении самооплодотворяющейся популя-
Гл. 13. Первичное видообразование
163
ции. Первичное биотически-симпатрическое видообразование у
перекрестнооплодотворяющихся организмов не было ни доказа-
но, ни исключено, в связи с чем по этому вопросу желательно
собрать больше данных.
Репродуктивная изоляция как побочный продукт
дивергенции
Следует ожидать, что образование различных генных комбина-
ций, детерминирующих различные комбинации адаптивных при-
знаков в дивергирующих популяциях, будет во многих случаях
в порядке побочного действия снижать возможность скрещива-
ний между такими популяциями. Короче говоря, можно ожидать
возникновения репродуктивной изоляции как побочного продук-
та эволюционной дивергенции. В пользу такого способа возник-
новения репродуктивной изоляции, на который впервые указал
Дарвин [152, 153], а впоследствии и многие другие авторы, сви-
детельствуют многочисленные данные как по растениям, так и
по животным.
Между разными расами одного вида обычно существует час-
тичная репродуктивная изоляция. Ранее в этой книге мы рас-
сматривали примеры расовой дифференциации в отношении
климата, агентов опыления и других факторов внешней среды.
Частичная сезонная изоляция нередко наблюдается между раз-
ными климатическими расами, принадлежащими к одному виду.
А между расами, для которых характерны разные агенты опы-
ления, может возникнуть частичная механическая и этологи-
ческая изоляция.
Salvia mellifera и S. apiana относятся к отдельным симпат-
рическим видам, обитающим в южной части Калифорнии. В их
способности сохранять свои отличительные признаки в областях
сосуществования главную роль играет механическая изоляция.
Эти два вида очень сильно различаются по строению цветков,
приспособленных к опылению разными пчелами: цветки S. tnel-
lifera опыляют такие формы, как Chloralictus и Anthophora,
имеющие средние размеры, а цветки S. apiana опыляют круп-
ные пчелы-каменщицы рода Xylocopa [264]. Сезонная изоляция
возникла, возможно, как побочный продукт этих различий в спо-
собе опыления. Salvia mellifera цветет ранней весной, в период
пика активности своих опылителей, a S. apiana — в конце весны
и в Начале лета, в период вылета пчел-каменщиц. Этологиче-
ская изоляция — другой побочный эффект тех же самых разли-
чии в строении цветков, поскольку пчелы научаются посещать
только те цветки, строение которых обеспечивает им возмож-
ность успешно выполнить свои функции. Наконец, некоторые
11*
164
Часть четеертаЯ. Эвдлюциопная дивергенция
данные указывают на существование между этими видами1 барь-
ера частичной несовместимости, которая может быть также
связана с различиями в строении их цветков [264].
Длина столбика по отношению к длине пыльцевой трубки —
фактор, играющий важную роль в совместимости при перекрест-
ном опылении у цветковых растений. В пределах данной расы
рост пыльцевой трубки координирован с нормальной для этой
расы длиной столбика. Но две расы, различающиеся по вели-
чине цветков, нередко бывают разделены, кроме того, барьером
частичной несовместимости. Так, при искусственном перекрест-
ном опылении между двумя расами Gilia ochroleuca с крупны-
ми цветками у них образуются многочисленные коробочки и
семена; но между расами с крупными и расами с мелкими
цветками этого же вида существует ясно выраженный барьер,
препятствующий их успешному скрещиванию. Расы G. ochro-
leuca с крупными и мелкими цветками различаются также по
легкости скрещивания с чужеродным видом G. сапа [300].
Большое число искусственных перекрестных опылений было
проведено у диплоидных видов секции Arachnion рода Gilia
[300]. Полученные результаты, сгруппированные по уровням
таксономической дивергенции, представлены в табл. 13.1. Как
видно из таблицы, у растений этой группы перекрестная совме-
стимость снижается с увеличением эволюционной дивергенции.
Виды рода Picea в одних комбинациях успешно скрещива-
ются друг с другом и дают гибриды, однако в других случаях
скрещивания оказываются безуспешными. Райт [905] обнару-
жил, что не удаются в большинстве случаев скрещивания меж-
ду видами, ареалы которых сильно удалены один от другого и
которые хорошо дифференцированы морфологически. Напротив,.
Таблица 13.1. Эффективность барьеров, создаваемых несовместим
мостыо, на разных уровнях дивергенции среди диплоидных
представителей секции Arachnion рода Gilia [300]
Тип скрещивания Число испы- танных комби- наций Среднее число налившихся семян на один опыленный цветок Число гибрид- ных индивид дуумов на 10 опыленных цветков
Между членами одной популяции 4 17,8 22
Между разными географическими расами одного и того же вида 26 15,2 12
Между разными видами, относящи- мися к одной и той же секции 103 3,7 3
Между гилиями секции Arachnion и диплоидными видами других сек- ций А 41 0,004 0,038
Гл 13 Первичное видообразование' .
виды Picea, занимающие соседние ареалы, в большинстве слу-
чаев скрещиваются довольно легко Такая корреляция позво-
ляет считать, что барьеры несовместимости возникли, вероятно,
в процессе дивергенции [905].
Генные комбинации, лежащие в основе межрасовых разли-
чий, создаются в процессе отбора на основании своих гармонич-
ных и сбалансированных воздействий па развитие в пределах
каждой расы. Однако многие межрасовые гибриды получат по
наследству генотипы, лишенные внутреннего равновесия.
И в самом деле, у межрасовых гибридов многих видов растений
в той или иной степени наблюдаются пониженная жизнеспособ-
ность, частичная плодовитость и разрушение.
Крукеберг [449] сравнивал плодовитость гибридов' Ft, полу-
ченных при скрещиваниях между разными популяциями Strtp-
tanthus glandulosus (Cruciferae) в 334 различных комбинациях.
При этом он обнаружил статистически значимую отрицатель-
ную корреляцию между плодовитостью гибридов и географиче-
ским расстоянием, разделяющим родительские популяции Пло-
довитость гибридов понижается с увеличением расстояния меж-
ду родительскими расами, а после того, как это расстояние
превысит 80—120 км, она резко падает [449].
Все способы репродуктивной изоляции и изоляции, созда-
ваемой средовыми факторами, существующие между хорошими
биологическими видами, существуют также, во всяком случае
в зарождающейся форме, между расами одного вида. Для того
чтобы произошло усиление изоляции от уровня, типичного для
рас, до уровня, типичного для видов, необходимо лишь продол*
жение процесса дивергенции.
Выводы
В гл 11 были рассмотрены некоторые результаты процесса ви-
дообразования. В данной главе мы рассмотрели одну из фаз
этого процесса — фазу, обозначаемую как первичное видообра-
зование. Попытаемся теперь связать то, что было сказано в
этой главе, с материалом гл. 11. Какие группы явлений видооб-
разования из описанных в гл. И можно считать результатом
первичного видообразования?
’У древесных растений с беспорядочной системой опыления,
примером которых служат Ceanothus, Quercus, Pinus и другие
роды, наблюдаются взаимоотношения, указывающие на посте-
пенную дивергенцию, связанную с дли1сльпо существующими
экологическими условиями. В результате такой дивергенции
образуются расы, полувиды и, в конечном итоге, группы видов,
между которыми существуют внутренние изолирующие механиз-
мы и которые рассматриваются таксономически как секции или
166
Часть четвертая. Эволюционная дивергенция
подроды. Естественная гибридизация часто происходит на ран-
ней и промежуточной стадиях дивергенции. Для того чтобы
объяснить тип взаимоотношений, существующих между видами
Ceanothus, нет нужды постулировать какие-либо иные процессы,
кроме процессов первичного видообразования и простой естест-
венной гибридизации, рассматриваемой как процесс, обратный
первичной дивергенции.
У многолетних трав со специализированными системами опы-
ления, подобных Aquilegia, Antirrhinum, Penstemon (частично)
и многим орхидным, наблюдаются в общем сходные взаимоот-
ношения. У них также выявляется постепенная дивергенция
через ряд стадий до состояния полной внутренней изоляции,
причем на ранних и средних стадиях нередко происходит ревер-
сия, вызываемая естественной гибридизацией. Дивергенция
«обусловлена различиями в агентах опыления, а также длитель-
но существующими экологическими условиями. Тип диверген-
ции, наблюдаемый у Aquilegia, так же как и тип, характерный
для Ceanothus, можно удовлетворительно объяснить как ре-
зультат первичного видообразования.
У таких многолетних травянистых растений, как Geum и
виды секции Apogon рода Iris, у которых нет сильно выражен-
ных межвидовых различий в системе опыления, также имеются
некоторые общие черты с типами Aquilegia и Ceanothus, но при
этом у них имеется и ряд отличий, как, например, хромосомная
стерильность между группами видов. Логично приписать общие
черты некой общей причине в процессе первичного видообразо-
вания. Что же касается отличительных явлений, характерных
для типа Geum, в частности наличия различных хромосомных
перестроек у разных групп видов, то они требуют иного объяс-
нения и будут дополнительно обсуждаться в одной из дальней-
ших глав.
Тип Madia, обнаруженный у многих родов однолетних трав,
еще сильнее отличается от типов Ceanothus и Aquilegia, так как
у принадлежащих к этому типу растений наблюдается хромо-
сомная стерильность между близкими видами. Эту черту нель-
зя отнести полностью за счет сил, способствующих диверген-
ции как таковой и обсуждавшихся в настоящей главе. Однако
у типа Madia есть другие черты, указывающие на действие про-
цессов первичного видообразования.
14 Структурные изменения
хромосом при видообразовании
На протяжении XX в. дифференциация видов растений и жи-
вотных в отношении структуры хромосом изучалась с различ-
ных точек зрения. В первые десятилетия были описаны межви-
довые различия в структуре хромосом у таких родов растений,
как Nicoiiana, Triticum и Viola. Исследовались различные типы
перестроек и их поведение при конъюгации. Была установлена
зависимость между поведением структурных гетерозигот в про-
цессе мейоза и стерильностью межвидовых гибридов, а также
генетическим поведением аллополиплоидов и других форм гиб-
ридного происхождения. Результаты исследований этого перио-
да суммированы Дарлингтоном [138, 139].
На следующем этапе стало возможным попытаться понять
смысл обнаруженных ранее цитогенетических явлений, задав-
шись следующим вопросом, какова функциональная роль видо-
специфичного кариотипа в жизни самого вида? Поиски ответа
на этот вопрос велись в различных направлениях. Структурные
перестройки связывали с благоприятными эффектами положе-
ния [248], благоприятными системами сцепления генов [150, 790,
795] или с обоими этими факторами [277].
С расширением цитогенетических исследований и вовлече-
нием в них большего числа родов растений стало также воз-
можным установить, в каких таксономических группах встреча-
ется изменение последовательности участков в хромосомах. Дар-
лингтон [143, 144, 150], Стеббинс [790, 794], Грант [280], Эрен-
дорфер [191] и Люис [490] выявили некоторые общие тенденции
и корреляции у высших растений. Основные выводы, сделан-
ные этими авторами, были изложены в гл. 11.
Позднее интерес сосредоточился на процессах видообразо-
вания, ведущих к формированию видоспецифичных кариотипов.
Этой проблемой занимались исследователи нескольких школ.
В 50—60-х годах она обсуждалась применительно к растениям
ЛхОисом [493, 494, 497] и мною [284], а применительно к насе-
комым— Уайтом [890—892], Карсоном [84], Джоном и Люисом
[406, 407]. Обсуждения продолжались и в 70-х годах (см., на-
пример, [85, 87, 293, 483, 611, 645, 894]).
Статьи, в которых рассматриваются эволюционные аспекты
изменения последовательности участков в хромосомах, очень
различны по своей научной ценности Многие авторы, по-види-
мому, занимались этой проблемой, не будучи знакомы со взгля-
дами других исследователей, высказанными ранее. Во многих
статьях уделяется внимание лишь какой-либо одной стороне
проблемы.
468 Часть четвертая, Эволюционная дивергенция
' В этой главе мы обсудим два взаимосвязанных вопроса. Это,
«во-первых, — функциональная роль видоспецифичной структуры
хромосом и, во-вторых, способ установления нового кариотипа
в процессе видообразования. Конечно, первую проблему можно
было бы обсуждать, не связывая ее со второй. А некоторые ав-
торы пытаются обсуждать вторую проблему, почти или совершен-
но не касаясь первой. Однако мы в своем изложении исходим
из того, что проблема создания новой последовательности уча-
стков в хромосоме неотделима от проблемы ее функциональ-
ного значения.
Теория эффекта последовательности участков
в хромосомах
Фенотипическое выражение данного гена нередко зависит от
оего пространственных взаимоотношений с соседними генами в
хромосоме. Это явление хорошо известно под названием эффек-
та положения гена. Гольдшмидт [247—249] расширил концеп-
цию эффекта положения, распространив ее на определенную
последовательность участков хромосом в геноме в целом. Каж-
дая последовательность участков обладает собственным инди-
видуальным «эффектом последовательности» (pattern effect) и
создает свой собственный характерный фенотип [248, 249]. От-
сюда следует, что изменение этой последовательности служит
источником новых эффектов последовательности, а тем самым
средством для создания новых комбинаций признаков
[248, 249].
Затем Гольдшмидт установил корреляцию различий между
видами по физиологическим и морфологическим признакам с их
различиями во взаиморасположении хромосомных участков.
Между этими двумя типами межвидовой дифференциации су-
ществует эволюционная связь. Формирование нового вида на-
чинается с крупномасштабного мутационного события в хромо-
сомах—с «системной мутации», которая перемешивает и пере-
распределяет хромосомные участки. В результате такого пере-
распределения (если оно не оказывается гибельным) возникает
новый вид (248, 249].
Ряд эволюционистов (обзор см. [282]) выдвинули серьезные
возражения против гольдшмидтовской теории видообразования.
Системные мутации должны возникать вначале в гетерозигот-
ном состоянии, и несущие их особи, будучи полными структур-
ными гетерозиготами, окажутся стерильными. Кроме того, си-
стемные мутанты — в гомозиготном или гетерозиготном состоя-
нии — скорее всего будут нежизнеспособны. Даже если данный
системный мутант сможет преодолеть «бутылочное горлышко»
начальной стерильности и нежизнеспособности, что весьма сом-
Гл. 14. Структурные изменения хромосом при видообразовании 169
нительно, то как он сможет утвердиться в популяции, состоящей
из немутантных особей?
Взгляды Гольдшмидта в их первоначальной форме не встре>-
тили широкой поддержки со стороны эволюционистов. Они бы-
ли приняты, однако, с существенными исправлениями некото-
рыми учеными, занимающимися видообразованием у растений.
К их числу относятся Лионе и Рейвн [493, 494, 497]
Люис и Рейвн постулировали ряд хромосомных пере-
строек — не столь многочисленных, чтобы вызвать полную сте-
рильность у гетерозигот, или с не столь резко выраженными
фенотипическими эффектами, чтобы привести к нежизнеспособ-
ности,— и тем самым устранили одно из главных возражений
против теории Гольдшмидта. Они высказали мнение, что какой-
то фактор — быть может, гены-мутаторы [493, 497] или вынуж-
денный инбридинг [494] — в определенные периоды в истории,
растительной популяции может вызвать разрывы хромосом и
структурные перестройки. Некоторые из новых геномов могут
обладать такими эффектами последовательности, которые по-
рождают новые физиологические свойства и дают своим носи-
телям возможность адаптироваться к новым экологическим
местообитаниям [497]. Такие ценные в адаптивном отношении
гено^- югут закрепиться в гомозиготном состоянии в какой-
’болыпой дочерней популяции, а затем распростра-
'ревратиться в новый дочерний вид [494, 497].
была предложена еще одна модификация теории
а Уилсон и его сотрудники [440, 645, 896] высказа-
1то хромосомные перестройки служат источником
характере генной регуляции, по которым различа-
эффекта последовательности участков обладает
тоинствами с теоретической точки зрения. Име-
е количество данных, свидетельствующих о том;.
1й, так и у животных функционально связан-
гены нередко локализованы в хромосомах
ъно, структурные перестройки разрывают
новые пространственные взаимоотношения
сказывается на фенотипе. С эмпирической
'ельства в пользу того, что эффект после^-
л важную роль в эволюции, менее убеди-
Т| ' ожепия гена у растений наблюдаются
04 оомосомные перестройки в большинстве
у1- чаметпых фенотипических изменений.
51 'ффекты положения встречаются чаще».
онитс _ —я вредными.
eol>xs последовательности в качестве полного или-
д же осно. ..иго объяснения межвидовых различий в кариотипе
!70 Часть четвертая. Эволюционная дивергенция
у растений имеет, как я уже отмечал в других работах [284,
293], один фатальный недостаток. У всех жизненных форм рас-
тений дивергирующие виды характеризуются различными ком-
бинациями адаптивных признаков. Если бы эти межвидовые
различия адаптивных свойств определялись эффектом последо-
вательности, то можно было бы ожидать, что им будут сопут-
ствовать соответствующие межвидовые различия кариотипов во
всех группах растений. Однако на самом деле этого нет. Как
мы убедились в гл. 11, в некоторых таксономических группах
и у некоторых жизненных форм растений изменение последо-
вательности участков в хромосомах связано с видообразованием,
а в других — нет. Поэтому главное эволюционное значение из-
менения последовательности участков в хромосомах следует
искать не в прямых эффектах последовательности, а в чем-то
ином.
Теория, основанная на сцеплении генов
Другое основное объяснение значения изменения последователь-
ности участков в хромосомах для видообразования состоит в
том, что происходящие при этом перестройки создают возмож-
ность для сцепления генов. На эту мысль наводит прежде всего
установленный цитогенетикой факт, что инверсии и транслока-
ции в гетерозиготном состоянии служат механизмами, позво-
ляющими выключить некоторые гены из процесса эффективной
рекомбинации. Логично было приписать сходную функцию сцеп-
ления перестройкам, закрепившимся в гомозиготном состоянии
в .процессе видообразования, как это и сделали Дарлингтон
[142], Дарлингтон и Мазер [150], Стеббинс [790] и Грант [277].
- Теория, объясняющая изменения последовательности участ-
ков в хромосомах на основании сцепления генов, подтверждает-
ся также данными о том, что этот процесс постоянно сопровож-
дает видообразование в тех группах растений, особенно одно-
летних и некоторых многолетних, у которых сильно ограничен-
ная рекомбинация, по-видимому, обладает высокой селективной
ценностью по другим причинам.
Как указывает Стеббинс [780, 782], травянистым растениям,
первыми заселяющим новые местообитания, в особенности од-
нолетним, необходима относительно высокая однородность при
размножении семенами, для того чтобы быстро увеличить чис-
ленность любого данного генотипа, доказывающего свою адап-
тивную ценность; такое требование репродуктивного постоянства
обычно обеспечивается у однолетников небольшим числом хро-
мосом или низкой частотой хиазм, а также самоопылением. Пе-
речень факторов, ограничивающих рекомбинацию у однолетних
ра$адцй,.был впоследствии расширен за счет включения в него
Гл 14 Структурные изменения хромосом при видообразовании
171
также и межвидовых различий в последовательности участков
[277, 280, 785].
На основе данных, полученных в некоторых экспериментах по
гибридизации, считалось, что различия между видами растений
в последовательности хромосомных участков, обусловленные пе-
рестройками, не зависят от морфологических и физиологических
различий между родительскими видами [780]. С таких позиций
было трудно усмотреть какую-либо генетическую связь между
изменениями последовательности участков в хромосомах и меж-
видовыми различиями по адаптивным признакам. Однако в
дальнейшем были получены данные [287, 707], указывающие на
то, что в нескольких группах растений сцепленные группы ге-
нов, определяющих цепные в адаптивном отношении морфоло-
гические и физиологические признаки, локализуются в участках,
подвергшихся перестройкам (см. гл. 8). Эти данные устраняют
существовавшее прежде теоретическое препятствие.
Дополнительные данные в пользу теории изменения после-
довательности участков, основанной на сцеплении генов, были
получены Бэбкоком и его школой в результате изучения редук-
ции основного числа хромосом у Crepis в процессе филогенеза.
Бэбкок и его сотрудники [33, 34, 37, 793] путем сравнительных
морфологических и цитологических исследований установили,
что в филогенезе рода Crepis преобладающим направлением был
переход от многолетних травянистых растений с основным чис-
лом хромосом х=6 к однолетникам с х=3. Стеббинс [780] вы-
сказал правдоподобное предположение, что адаптивная ценность
низкого основного числа хромосом состояла в усилении сцепле-
ния генов у возникших в результате однолетних видов.
Далее стало возможным показать, что в механизме умень-
шения числа хромосом у Crepis участвовали неравные реципрок-
ные транслокации [33, 610, 834]. Тем самым была установлена
корреляция между изменением последовательности участков в
хромосомах и изменениями в группах сцепления.
Закрепление новых структурных гомозигот
Закрепление хромосомных перестроек у структурных гомозигот
в дочерней популяции сопровождается в качестве побочного
эффекта образованием барьеров хромосомной стерильности
между новой и старой популяциями. Процесс этот представляет
собой, таким образом, особый случай первичного видообразова-
ния. Это один из аспектов видообразования, который все еще
остается малопонятным.
Если принять теорию изменения последовательности участ-
ков в хромосомах, основанную на сцеплении генов, то становит-
ся логически необходимым постулировать для родительской по*
172 Часть четвертая. Эволюционная дивергенция
Пуляции некое предковое состояние генного и связанного с
лим хромосомного полиморфизма. Такой генный и структурный
«полиморфизм мог быть либо сбалансированным и длительным,
•либо переходным и кратковременным.
Другой необходимый постулат состоит в том, что одна из
«возможных гомозиготных комбинаций генов, встречающаяся в
полиморфной родительской популяции в качестве одного из ва-
риантов, имеет наивысшую адаптивную ценность в каком-либо
Ловом местообитании, доступном для этой популяции. В таком
случае выщепенцы, гомозиготные по этому сочетанию генов,
могут заселять новое местообитание и успешно размножаться
в нем Образующаяся в результате дочерняя популяция будет
отличаться от основной части родительской популяции как по
своим генам, так и по структуре хромосом [284, 894].
«Конечно, этот процесс может происходить независимо в раз-
ных дивергирующих линиях. Два или несколько различных но-
вых местообитаний, доступных для данной популяции, могут
оказаться местом рождения двух или нескольких генных или
структурных гомозигот, происходящих от одной и той же измен-
чивой родительской популяции. В результате возникнут адаптив-
ные различия и связанные с ними изменения последовательно-
сти участков в хромосомах у трех или большего числа попу-
ляций.
У дрозофилы, прямокрылых и Clarkia (Onagraceae) типы
хромосомных перестроек, по которым различаются близкие ви-
ды, не похожи на те, которые встречаются в пределах вида в
«качестве обычных полиморфных или расовых вариантов [493,
•497, 770, 875, 892]. У дрозофилы, кроме того, полиморфизм по
строению хромосом лучше всего развит в центральных популя-
щиях данного вида, тогда как периферические популяции часто
бывают мономорфными в отношении последовательности генов
$83, 84, .88]; Эти факты свидетельствуют о том; что виды, у кото-
рых сейчас наблюдается широко распространенный хромосом-
ный полиморфизм, когда-то в прошлом были мономорфными
£84, 88, 408, 770,891}.
Какиё'способы видообразования обусловливают закрепление
новой последовательности участков в хромосомах? Имеющиеся
«ведения носят косвенный характер, и их нельзя считать окон-
чательными. Некоторые исследователи отдают предпочтение
-симпатрическому способу видообразования [252, 253, 894].
Однако следует ожидать, что сохранение и размножение особей
с новыми последовательностями в области, занятой особями с
последовательностями предкового типа, будет затруднено. Дру-
гие исследователи, в том числе и я, считают, что новый вид,
как правило, „возникает из какой-либо изолированной моно-
МОрфцой;периф€^!Шеской. популяции предкового, вида [86, 87,
Гл. 14. Структурные изменения хромосом при видообразовании
173
Выводы
Как было показано в гл. 11, дифференциация близких видов
по структуре хромосом — одна из ярко выраженных особенно-
стей однолетних растений, относящихся к типам Madia и Gilia
inconspicua. Процессы первичного видообразования на основе
генной изменчивости не позволяют объяснить эту особенность.
Но теория, основанная на сцеплении генов, дает удовлетвори-
тельное объяснение преобладанию у однолетних растений из-
мененных последовательностей участков в хромосомах.
Однолетние растения обычно играют в своих сообществах
роль пионеров. Эта экологическая роль благоприятствует рас-
тениям с ограниченными рекомбинационными системами, а хро-
мосомные перестройки служат эффективными факторами, огра-
ничивающими рекомбинацию. Близкие виды однолетников, пер-
выми осваивающие новые местообитания, обладают различными
наборами сцепленных адаптивных генных комбинаций, связан-
ных с различной последовательностью участков в их хромосо-
мах
Тип Geum, как это было также показано в гл. 11, занимает
промежуточное положение между типами Madia и Gilia incon-
spicua, с одной стороны, и типами Ceanothus и Aquilegia — с
другой. Близкие виды многолетних трав, относящиеся к типу
Geum, генетически дифференцированы и репродуктивно изоли-
рованы при помощи внешних механизмов, как у типов Ceanot-
hus и Aquilegia, и эти общие черты можно в целом объяснить
первичной дивергенцией на основе генной изменчивости. Но хро-
мосомная стерильность обнаружена между группами видов в
родах, относящихся к типу Geum. Таким образом, здесь тоже
имело место изменение последовательности участков в хромосо-
мах, но на более высоком таксономическом уровне, чем у родов,
относящихся к типам Madia и Gilia inconspicua.
Можно сделать вывод, что процесс дивергенции в отношении
расположения участков в хромосомах, наблюдающийся в со-
временный период у быстро эволюционирующих однолетних
-растений, происходил в каком-то более раннем цикле видообра-
зования у многолетних растений, принадлежащих к типу Geum.
Иными словами, у Geum и у подобных ему родов самой недав-
ней стадии видобразования, не связанной с изменением после-
довательности участков в хромосомах, предшествовала стадия,
.сопровождающаяся изменениями этой последовательности. Быть
может, в более ранний период видообразования такие многолет-
ние роды играли в сообществах роль пионеров, сравнимую с
ролью Однолетников в некоторых недавних сообществах
Древесные растения северного умеренного пояса, очевидно,
претерпели изменения последовательности участков в хромосо-
174 Часть четвертая. Эволюционная дивергенция
мах в еще более ранние периоды своей эволюционной истории.
Целые роды или семейства в настоящее время цитологически
стабильны; все известные виды, которые к ним относятся, имеют
одно и то же основное число хромосом, отличаясь, однако, от
родственных родов или семейств по этому числу. Так, у Betula-
сеае х=14, у Fagaceae я =12, а у Corylaceae х= 14, 11 и 8 [151];
В австралийской флоре также известны группы древесных рас-
тений, как, например, Proteaceae и Myrtaceae, у которых, судя
по имеющимся данным, в далеком прошлом имело место изме*
некие последовательности участков в хромосомах £747].
В заключение следует отметить, что нам известно достаточ-
но много относительно изменения последовательности участков
в хромосомах и его роли в видообразовании, _однако целый ряд
проблем все еще остается невыясненным.
15 Отбор на репродуктивную
изоляцию
Репродуктивная изоляция в ее разных формах может разви-
ваться и развивается как побочный продукт эволюционной ди-
вергенции между популяциями, о чем уже говорилось в гл. 13.
На этот основной процесс накладывается другой, комплементар-
ный, способ возникновения изоляции. Некоторые типы механиз-
мов репродуктивной изоляции создаются в результате прямого
отбора на изоляцию как таковую в случае, если это состояние
выгодно для тех или иных конкретных популяций. Предполага-
ется, что те особи из двух симпатрических популяций, которые
производят нежизнеспособные или стерильные гибриды внесут
меньший вклад в будущие поколения, чем сестринские особи
из тех же родительских популяций, которые не гибридизируют.
Следовательно, у каждого из двух видов частота генетических
факторов, детерминирующих какое-либо препятствие или эвер-
сию к межвидовой гибридизации, будет из поколения в поколе-
ние повышаться. Происходящий в этих направлениях естествен-
ный отбор приведет, таким образом, к усилению репродуктив-
ной изоляции, первоначально возникшей как побочный резуль-
тат дивергенции.
Особые условия
На способность отбора усиливать зарождающуюся репродук-
тивную изоляцию оказывают влияние разные условия. Некото-
рые из этих условий необходимы, а другие — благоприятны для
отбора на изоляцию.
Совершенно очевидно, во-первых, что этот особый процесс
отбора возможен лишь в том случае, если участвующие в нем
виды симпатрические или по меньшей мере маргииально-симпат-
рические
Во-вторых, результаты гибридизации должны быть вредны-
ми. Некоторые или все гибриды в Fi или в последующих поко-
лениях должны быть стерильными или нежизнеспособными. Та-
кая ситуация обычно наблюдается при межвидовой гибридиза-
ции у растений, как мы уже отмечали в предшествующих гла-
вах; однако она не распространяется на все случаи гибридиза-
ции у растений.
Какой-либо гибридный генотип в Fi или в последующих по-
колениях может быть стерильным, но обладать в том местооби-
тании, в котором он встречается, более высокой вегетативной
мощностью, чем любой из родительских видов. Если хорошо
адаптированное гибридное растение способно к вегетативному
176 Часть четвертая. Эволюционная дивергенция
или агамоспермному размножению, то оно может затем рас-
пространиться и повысить свою численность по сравнению с ро-
дительскими видами в среде, обычной для этих последних. В ря-
де групп растений наблюдались реальные примеры гибридов,,
численность которых превышала численность родительских форм
в определенных местообитаниях. Отбор на изоляцию происходит
в обычной комплементарной ситуации, когда в результате меж-
видовой гибридизации действительно значительно снижается
число хорошо адаптированных потомков, производимых гибриди-
зирующими родительскими особями.
Еще одно условие, способствующее эффективному действию
отбора на репродуктивную изоляцию, состоит в том, что утра-
та репродуктивного потенциала, происходящая при гибридиза-
ции, весьма невыгодна для участвующих в гибридизации орга-
низмов. Селективная невыгодность снижения репродуктивного
потенциала сильно варьирует у организмов разных классов. На
одном конце находятся организмы с короткой продолжитель-
ностью жизни, такие, как поденки и однолетние растения, особи
которых имеют возможность размножаться лишь один раз и
жизни. А на противоположном конце находятся долгоживущие
древесные или травянистые растения замкнутых сообществ, ко-
торые обычно из года в год, десятилетие за десятилетием произ-
водят огромный избыток семян. И, разумеется, множество рас-
тений и животных занимают промежуточное положение между
этими двумя крайностями.
Селективная невыгодность утраты репродуктивного потен-
циала велика в случае однолетних растений и других эфемер-
ных организмов, но для долгоживущих растений замкнутых
сообществ она пренебрежимо мала [780]. Отсюда следует, что-
отбор на репродуктивную изоляцию может быть особенно силь-
ным у однолетников и относительно неэффективным у многолет-
них растений с большой продолжительностью жизни [169, 785]~
Некоторые многолетние травы, возможно, приближаются к од-
нолетникам, по своей способности реагировать на отбор, направ-
ленный на создание изоляции [479].
Далее, отбор на репродуктивную изоляцию не обязательно*
ведет к полному предотвращению гибридизации, как указал
Эрман [198]. Репродуктивная изоляция будет усиливаться до
уровня, необходимого для того, чтобы предохранить популяции
от суммарных вредных эффектов гибридизации. Этот уровень
зависит от толерантности популяций к селективной элиминации.
Эрман [198] считает, что минимально необходимый уровень ре-
продуктивной изоляции — это уровень, при котором «обмен ге-
нами между видовыми популяциями не превышает той скоро-
сти, с которой рассеянные гены могут быть элиминированы есте-
ственным отбором».
Гл. 15. Отбор на репродуктивную изоляцию
17Т
И, наконец, отбор на создание репродуктивной изоляции мо-
жет действовать только на изолирующие механизмы, сущест-
вующие в родительском поколении. Особенно эффективно отбор*
усиливает этологические барьеры и барьеры несовместимости.
При этом такой действующий до оплодотворения барьер, как.
этологическая изоляция, сохраняет репродуктивный потенциал-
родительских видов более успешно, чем какой-либо барьер, всту-
пающий в действие, подобно несовместимости, после оплодотво-
рения.
Важная роль этологической изоляции у животных может
быть частично или в значительной мере приписана отбору, на-
правленному на создание изоляции У растений же одной это-
логической изоляции обычно недостаточно, чтобы предотвра-
тить гибридизацию, потому что приверженность насекомых-
опылителей к одним и тем же цветкам время от времени нару-
шается, и в таких случаях несовместимость может оказался?
чрезвычайно существенной В связи с этим следует ожидать,,
что у растений отбор на изоляцию создает не только этологи-
ческие барьеры, но и барьер несовместимости
Барьеры несовместимости состоят у растений из целого комп-
лекса блокирующих механизмов, действующих до и после опло-
дотворения. Механизмы, действующие до оплодотворения, имеют
высокую селективную ценность, так как сохраняют яйцеклетки
для внутривидовых скрещиваний. Однако механизмы, действую-
щие после оплодотворения и препятствующие образованию зре-
лых гибридных семян, также обладают селективным преимуще-
ством по сравнению с отсутствием таких механизмов, поскольку*
они позволяют сохранить питательные вещества, количество ко-
торых ограничено, для запасания в негибридных семенах [288].
Эксперименты по искусственному отбору
Гипотеза о том, что отбор способен усиливать репродуктивную
изоляцию, была подвергнута проверке в экспериментах с искус-
ственным отбором.
Первые такие эксперименты были проведены на дрозофиле.
Смешанные культуры, содержавшие два вида (Drosophila pseu-
doobscura и D. persimilis) или две линии одного и того же вида
(D melanogaster) подвергались па протяжении ряда поколений
отбору, направленному против гибридов В процессе такого
искусственного отбора этологические барьеры между видами
или линиями усиливались [443, 445, 874].
У растений аналогичные эксперименты по отбору провел
Патерниапи [634]. Патерниани использовал два сорта Zea mays,
различающиеся по окраске зерен. Формы с белыми и желтыми*
зернами высевали вперемешку, создавая им возможность дляс
12—45
ff78
Часть четвертая. Эволюционная дивергенция
перекрестного опыления. Затем проводили искусственный отбор,
сохраняя початки, содержавшие главным образом негибрид-
иые семена. После такого отбора на протяжении пяти поколе-
ний уровень спонтанной межсортовой гибридизации упал с 47
до 3% у сорта с желтыми семенами и с 36 до 5% у сорта с бе-
лыми семенами. Усиление репродуктивной изоляции было час-
тично обусловлено большими различиями между сортами по
«срокам цветения, а частично — развитием барьеров несовмести-
мости, действующих до оплодотворения.
Данные по природным популяциям
Как предсказывает теория, отбор, направленный на создание
изоляции, действительно происходит, о чем свидетельствуют экс-
периментальные данные. Следовательно, наша задача состоит
п том, чтобы оценить роль этого процесса в природе. Для групп
видов характерны три типа географического распространения,
позволяющие сравнивать родственные аллопатрические и сим-
патрические популяции по относительной степени их изоляции
в родительском поколении (рис. 15.1)'.
В простом случае ареалы двух близких видов перекрывают-
«ся, так что некоторые расы одного вида аллопатричны по от-
ношению к другому виду, а другие — симпатричны (рис. 15.1,А).
Аллопатрические и симпатрические виды можно сравнивать по
степени изоляции между ними.
В самых первых работах по изучению отбора на изоляцию
в природе была использована именно такая ситуация. Исследо-
вания проводились главным образом на парах видов плодовой
мушки и лягушек и касались этологической изоляции. Оказа-
лось, что в большинстве случаев этологическая изоляция выра-
жена сильнее между симпатрическими расами какой-либо пары
видов, чем между аллопатрическими расами тех же самых двух
А Б в
Рис. 15.1. Три типа географического распространения в группе видов, в ко-
торой возможно сравнивать относительный уровень репродуктивной изоля-
ции между аллопатрическими и симпатрическими популяциями (объяснения
<см. в тексте).
Гл. 15. Отбор на репродуктивную изоляцию
17&
видов (см. (199, 200] и приведенную в этих работах библиогра-
фию).
Другим источником данных послужили сравнения между ви-
дами в целом в группах видов, которые состоят из нескольких
симпатрических видов, занимающих центральную часть ареала,
и находящегося на периферии географически изолированного
вида (рис. 15.1,Б).
Сравнения степени этологической изоляции в группах видов
с распространением подобного типа проводились у разных жи-
вотных, Приведем в качестве примеров группы видов Drosophi-
la guarani [439], D. willistoni [172], Erebia tyndarus [518] и Gam-
busia affinis [385]. В каждой из этих групп видов этологическая
изоляция была выражена относительно сильно между симпатри-
ческими видами и слабее между периферическим видом и одним
из центральных видов.
В подобной ситуации (рис. 15.1,5) по крайней мере один
вид в каждой паре видов оказывался в условиях симпатрии, а
следовательно, подвергался отбору на усиление изоляции. Это
осложняет картину. Более четкие данные можно получить, срав-
нивая эффективность барьеров между двумя или несколькими
симпатрическими видами с барьерами между двумя или не-
сколькими аллопатрическими видами (рис. 15.1, В). Такие срав-
нительные исследования мощности барьеров несовместимости
проводились на облиствепностебельных гилиях, о чем пойдет
речь в следующем разделе.
Барьеры несовместимости у Gilia
У растений барьеры несовместимости могут возникнуть и воз-
никают как побочные эффекты эволюционной дивергенции. На-
это указывает наличие у многих видов явной, хотя и слабо вы-
раженной несовместимости между расами. Данные о том, что
такая перекрестная несовместимость у видов облиственносте-
бельных гилий является побочным эффектом дивергенции, были
приведены в гл. 13. Но перекрестная несовместимость может
создаваться также как механизм, усиливающий изоляцию
Облиственностебельные гилии — естественная группа одно-
летних растений, произрастающих на тихоокеанском побережье-
Северной Америки и в умеренном поясе Южной Америки (см.
гл- 11). Девять из десяти видов, составляющих эту группу, изу-
чались с цитотаксономической, генетической, а также экологи-
ческой точек зрения. Все полученные межвидовые гибриды вы-
сокостерильны, и мейоз у них нарушен. Имеются косвенные дан-
ные, указывающие на то, что все девять видов окажутся интер-
стерильными во всех возможных комбинациях. Между некоторы-
12*
Часть четвертая. рволюциошюяЯивереенщья
180
зли- видами существуют достаточно мощные барьеры несовмести-
мости; между другими эти барьеры менее эффективны' [274,
Облиственностебельные гилии представляют собой однолет-
ние травянистые растения, которые цветут весной, а в конце
весны или начале лета, образовав семена, отмирают. Поэтому
•снижение продукции семян в результате возникновения стериль-
ных гибридов крайне невыгодно этим растениям' в селективном
отношении. В некоторых популяциях Gilia обнаружен полимор-
физм по скрещиваемости с чужим видом, что создает возмож-
ность для действия отбора [290].
Девять видов облиственностебельных гилий, изученных гене-
тически, можно разделить на две подгруппы на основе их геог-
рафического распространения.
Пять видов — Gilia capitata, G. tricolor и другие — встреча-
ются в предгорьях и долинах Калифорнии и прилежащих райо-
нах. Ареалы этих видов сильно перекрываются (рис. 4.1). Особи,
принадлежащие к двум или нескольким видам этой подгруппы,
растут и цветут в тесном соседстве во многих естественных мес-
тообитаниях (рис. 15.2).
ПРЕДГОРНЫЕ И ДОЛИННЫЕ ВИДЫ
Фис. 15.2. Пары видов облиственностебельных гилий, растущие в природе в
тесном соседстве [290].
Гл. 15. Отбор на репродуктивную изоляцию
181
Остальные четыре вида — Gilia millefoliata, G. nevinii и дру-
гие— встречаются на морском берегу в четырех далеко отстоя-
щих одна от другой местностях в Северной и Южной Америке,
Эти приморские виды полностью аллопатричны по отношению
друг к другу. Два из них имеют редкие симпатрические контак-
ты с некоторыми предгорными и долинными видами, а ареалы
двух других целиком лежат за пределами области распростра-
нения видов первой подгруппы (рис. 15.2).
Эти две географические подгруппы облиственностебельных
гилий заметно различаются по эффективности барьеров несов-
местимости. Эффективность барьера несовместимости можно
оценить количественно по среднему числу гибридных семян, про-
изводимых па один цветок при перекрестном опылении чужим
видом в контролируемых условиях.
Симпатрические предгорные и долинные виды разделены
очень мощными барьерами несовместимости. У этих видов про-
дукция гибридных семян на один цветок колебалась от 0,0 до
1,2 при средней 0,2 для многочисленных скрещиваний между
разными видами В отличие от этого приморские виды легко
скрещиваются между собой. Скрещивания между разными при-
морскими видами давали от 7,7 до 24,8 гибридных семян на один
цветок, а средняя для межвидовых скрещиваний во всех комби-
нациях составила 18,1 семян на цветок [290].
Как отмечалось выше, в качестве побочного эффекта дивер-
генции следует ожидать некоторой несовместимости при скрещи-
ваниях Но уровень дивергенции между видами в обеих геогра-
фических подгруппах этих гилий примерно одинаков. Различия
в скрещиваемости, наблюдаемые между двумя географическими
подгруппами видов, нельзя объяснить на основе одной лишь
эволюционной дивергенции. Высокую степень несовместимости
при скрещиваниях между симпатрическими видами предгорий
и долин наиболее разумно отнести за счет отбора, направлен-
ного на создание изоляции [290].
Подтверждение этому дает изучение гилий, принадлежащих
к секции Arachnion В этой группе нельзя проводить сравнения
третьего типа (рис. 15.1,В), так как в ней нет подгруппы алло-
патрических видов, но сравнения второго типа (рис. 15.1, Б)
провести можно. В результате многочисленных скрещиваний
между симпатрическими видами в этой группе была выявлена
-сильная несовместимость, выразившаяся в том, что среднее чис-
ло гибридных семян на цветок составило 3,4. Однако при скре-
щиваниях между центральными и периферическими видами не-
совместимость слабее и среднее число гибридных семян на цве-
ток равно 7,6 [290].
182 Часть четвертая. Эволюционная дивергенция
Опознавательные знаки цветков
Пчелы и различные другие насекомые-опылители обладают ин-
стинктом постоянства, в силу которого они всякий раз посещают
преимущественно цветки растений одного вида. Этот инстинкт,
как уже было сказано в гл. 9, лежит в основе этологической
изоляции между разными видами растений, которые опыляются
пчелами или другими насекомыми.
Классическими экспериментами Фриша было показано, что
медоносные пчелы способны распознавать определенные цвета,
формы и запахи, по которым различаются цветки разных видов
во многих группах растений [221, 222]. Такие же способности
обнаружены у других пчел и насекомых, посещающих цветки
[206, 253]. Облетая цветки и постоянно выбирая цветки одного
вида, насекомое руководствуется их отличительной окраской (в
частности, ее узором), формой и (или) запахом.
Несколько лет назад был проведен обзор многочисленных
родов покрытосеменных с целью оценить относительное значение
признаков цветков в систематике растений с различной систе-
мой опыления. Оказалось, что 38% признаков, используемых
систематиками для различения близких видов растений, опы-
ляемых пчелами, относятся к лепесткам и другим внутренним
частям цветка. Примерно такой же процент признаков цветка
используется в систематике тех видов растений, которые опы-
ляются другими насекомыми, также неизменно посещающими
одни и те же цветки. Что же касается тех растений, которые
беспорядочно опыляются многими видами неспециализирован-
ных насекомых, то у них лишь 15% признаков, используемых
для различения близких видов, относятся к цветку [265].
При этих подсчетах межвидовые различия по тем признакам
цветка, которые связаны с механической изоляцией, не отделяли
от различий по признакам, связанным с этологической изоля-
цией. Тем не менее можно с достаточной уверенностью допус-
тить, что какая-то часть этой внушительной массы признаков
цветка у покрытосеменных, постоянно опыляемых одними и те-
ми же видами насекомых, служат опознавательными знаками
для опылителей. В подтверждение такого допущения можно
привести результаты, полученные при более подробном изучении
некоторых частных случаев.
Так, например, Gilia capitata capitata и G. с. chamissonis
различаются помимо прочих признаков по форме лепестков и по
запаху цветков. На экспериментальном участке, где высеяны
вперемешку эти две формы гилии и где их посещают пчелы, от-
дельные насекомые обычно сохраняют верность одной или дру-
гой форме. Такое поведение пчел позволяет считать, что при вы-
Гл. 15. Отбор на репродуктивную изоляцию
183
•боре цветков они руководствуются формой лепестков, запахом
цветков или одновременно обоими этими признаками [265, 267].
Дополнительные данные, свидетельствующие о возможном
адаптивном значении опознавательных знаков цветков для соз-
дания этологической изоляции, были получены позднее Левином
и Керстером [478], а также Левином и Шаалем [484] на флок-
сах. Распространение этих видов соответствовало типу, изобра-
женному на рис. 15.1, Л.
В восточной части США ареалы Phlox pilosa и Р. glaberrima
перекрываются, и во многих местах эти виды биотически сим-
патричны. Эти два вида несовместимы, но между ними нет дос-
таточно надежной механической или сезонной изоляции. Их
опыляют одни и те же виды чешуекрылых, что приводит в сме-
шанных колониях к довольно значительному опылению одного
вида другим [478, 479].
И у Phlox pilosa, и у Р. glaberrima цветки обычно розовые.
У Р. pilosa встречаются также формы с белыми цветками в ка-
честве полиморфных вариантов. В большинстве полиморфных
популяций Р. pilosa форма с белыми цветками встречается ред-
ко, но в некоторых популяциях она преобладает.
Географическое распространение популяций Phlox pilosa с
.преимущественно белыми цветками представляет большой инте-
рес. Эти популяции всегда симпатричны с популяциями Р. gla-
berrima, у которого цветки розовые. Такая корреляция позволяет
•считать, что белая окраска цветков в смешанных колониях по-
могает бабочкам-опылителям распознавать разные виды этого
растения [478].
Бабочки, перелетающие от Phlox glaberrima к Р. pilosa в
смешанных колониях, явно предпочитают Р. pilosa с розовыми
цветками и избегают форму с белыми цветками. Подсчет пыль-
цевых зерен Р. glaberrima на рыльцах Р. pilosa показывает, что
-опылители переносят в пять раз больше пыльцевых зерен этого
вида на розовые цветки Р. pilosa, чем на такое же число белых
цветков Р. pilosa [479]. Таким образом, в смешанных колониях
форма Р. pilosa с белыми цветками, очевидно, обладает селек-
тивным преимуществом по сравнению с формой с розовыми цвет-
ками. Этим, по-видимому, объясняется высокая частота, дости-
гаемая формой Р. pilosa с белыми цветками в области перекры-
вания с Р. glaberrima [479].
У нескольких групп растений, произрастающих в Скалистых
горах в штате Колорадо и опыляемых одними и теми же насе-
комыми, наибольшая дивергенция по признаку высоты соцветия
наблюдается в условиях симпатрии. Высота, на которой нахо-
дится соцветие, оказывает влияние на постоянство посещения
опылителей. Дивергенция по высоте соцветия представляет со-
бой, возможно, результат отбора на изоляцию [862J.
184
Часть четвертая. Эволюционная дивергенция
Исторические аспекты
Первые высказывания по теории отбора, направленного на соз-
дание изоляции, в новейший период эволюционных исследований
принадлежат Фишеру [213], Добржанскому (167] и Хаксли
[392]. Их формулировки были краткими, но весьма многозначи-
тельными и послужили стимулом для последующих исследова-
ний.
Одна из задач, возникших вслед за этим, состояла в более
точном указании условий, в которых можно ожидать действия
предполагаемого процесса отбора. Другая задача заключалась
в получении убедительных доказательств того, что процесс этот
действительно происходит в природе. В шестидесятых годах на-
шего века эти задачи были выполнены. Дальнейшие исследова-
ния, проводившиеся в семидесятых годах, еще больше укрепили
представление о роли отбора в усилении изолирующих механиз-
мов, и в настоящее время оно завоевало всеобщее признание.
С исторической точки зрения интересно отметить, что и Дар-
вин, и Уоллес еще в XIX в. рассматривали возможность отбора
на создание репродуктивной изоляции. Дарвин [153] писал:
«Можно допустить..., что зарождающемуся виду было бы выгодно, если
бы он оказался в какой-то небольшой степени бесплодным при скрещива-
нии с родительской формой или с какой-либо другой разновидностью, по-
тому что таким образом возникло бы меньше метисных и ухудшенных потом-
ков, могущих затем примешивать свою кровь к новому виду, находящемуся
в процессе формирования. Но тот, кто даст себе труд поразмыслить над те-
ми стадиями, проходя через которые эта начальная степень бесплодия мо-
жет усиливаться под действием естественного отбора,.. убедится в чрезвычай-
ной сложности этой проблемы. По зрелом размышлении мне кажется, что
это не могло быть достигнуто путем естественного отбора».
Подобный же взгляд Дарвин высказал в «Происхождении
видов» [154].
Уоллес [873], однако, считал, что естественный отбор мог
создавать преграды скрещиванию между дивергирующими сим-
патрическими видами. В статье, озаглавленной «Влияние есте-
ственного отбора на стерильность и плодовитость», Уоллес [873]
писал:
«Многим может показаться, что поскольку бесплодие должно быть вы-
годно зарождающимся видам, когда они занимают одну и ту же или со-
седние области, так как оно нейтрализует эффекты межвидовых скрещива-
ний, то оно должно усиливаться под действием естественного отбора; и это
покажется еще более вероятным, если допустить, и на то, как мы убеди-
лись, есть основания, что плодовитость бывает подвержена изменчивости,
пожалуй, столь же часто, как и другие признаки... Особенно следует отме-
тить, что подобный эффект возникнет не в результате сохранения бесплод-
ных вариантов по причине их бесплодия, но в результате неполноценности
гибридных потомков, которые более малочисленны, менее способны обеспе»
Гл. 15. Отбор на репродуктивную изоляцию
185
чить продолжение существования свой расы и менее приспособлены к дан-
ным условиям среды, чем любая из чистых форм Именно эта неполноцен-
ность гибридных потомков и ecib главное; и поскольку число таких гибри-
дов будет всегда ниже там, где бесплодие будет наибольшим, постольку те
части двух форм, в которых бесплодие достигнет наибольшей степени, будут
обладать преимуществом и в конечном счете выживут в борьбе за сущест-
вование».
Уоллес считал также, что различным симпатрическим видам
растений, опыляемых насекомыми, постоянно посещающими
цветки одного и того же вида, выгодно иметь на цветках разные
опознавательные знаки, так как подобные знаки снижают часто-
ту опыления пыльцой чужого вида. Приведем еще одну цитату
[873].
«Быть может, именно для того, чтобы помочь насекомым посещать вся-
кий раз лишь один цветок, что чрезвычайно важно для сохранения данного
вида, у растений, зацветающих в одно и то же время в смешанных колони-
ях, цветки обычно сильно различаются по форме и окраске Эту особенность
локального распределения окраски цветков можно сравнить в отношении ее
назначения с опознавательной окраской у животных Точно так же как ок-
раска у этих последних дает возможность особям разного пола узнавать
друг друга и тем самым избегать стерильных союзов между разными вида-
ми, форма и окраска цветков у каждого вида растений, отличающая их от
цветков других видов, обычно растущих вокруг, помогает насекомым-опыли-
телям избегать переноса пыльцы одного цветка на рыльца чужого вида»
Взгляды Уоллеса опередили его время, поэтому его идея не
привлекла внимания, а затем была забыта В основном та же
мысль, по сформулированная в генетических терминах, была
вновь высказана Фишером и другими в начале современного пе-
риода изучения эволюции, о чем уже говорилось выше. Я счи-
тал уместным выразить запоздалое признание гипотезе Уоллеса,
назвав отбор, направленный на создание репродуктивной изоля-
ции, эффектом Уоллеса [290].
После того как я в 1966 г. поставил вопрос относительно
обоснованности модели Уоллеса о происхождении стерильности,
возникли некоторые проблемы. Отбор на создание репродуктив-
ной изоляции не может оказывать влияние на гибридную сте-
рильность. Все зависит от истолкования того, какой смысл вкла-
дывал Уоллес в термин стерильность. Уоллес и Дарвин применя-
ли этот термин не в современном его смысле, т. е. для них он
означал не только стерильность гибридов, но и то, что мы те-
перь называем несовместимостью. Так, Дарвин [152, гл. 9], рас-
сматривая бесплодие, проводит различие между «бесплодием
Двух видов при первом скрещивании» и «бесплодием происшед-
ших от них гибридов»; Уоллес [873] также проводил это раз-
личие
Я расцениваю весь этот абзац Уоллеса [873, с. 173—186] сле-
дующим образом Уоллес иногда имел в виду гибридную сте-
186 Часть четвертая. Эволюционная дивергенция
рильность, и в этих случаях он был неправ, но иногда он имел’ в
виду несовместимость при скрещиваниях, и в таких случаях о»
был на верном пути. Я склонен делать упор на этих последних
случаях, пренебрегая первыми, и отдавать должное Уоллесу за>
его первые шаги в этой трудной и сложной проблеме.
Эффект Уоллеса и смещение признаков
Эффект Уоллеса и смещение признаков создают сходную карти-
ну изменчивости в группах видов с географическим распростра-
нением типа А (рис. 15.1), а именно — максимальную дифферен-
циацию признаков в зоне симпатрии. В связи с этим высказы-
вались мнения, что эффект Уоллеса «может быть включен в бо-
лее широкую концепцию смещения признаков» [474].
Однако здесь возможны различные толкования. Я рассмат-
риваю эффект Уоллеса и смещение признаков не как концеп-
ции, одна из которых включает в себя другую, а как две парал-
лельные концепции. Эффект Уоллеса возникает в результате ре-
продуктивных взаимодействий между видами, а смещение приз*
наков — в результате экологических взаимодействий. Эффект
Уоллеса — продукт отбора, направленного на снижение межви-
довой гибридизации, смещение признаков — результат отбора,
направленного на снижение межвидовой конкуренции.
В классической работе Брауна и Уилсона [73] о смещении
признаков все внимание сосредоточено почти исключительно на
различиях в признаках, не имеющих отношения к размножению^
и на экологических взаимодействиях. Авторы лишь вкратце ка-
саются различий по признакам, участвующим в создании ре-
продуктивной изоляции. Концепция отбора, направленного на
создание репродуктивной изоляции, возникла в результате рас-
суждений в совершенно ином направлении, нежели то, которое
развивается в качестве основной темы в статье Брауна и Уил-
сона.
Недавно термин смещение репродуктивных признаков был
использован Валеном [886] в связи с некоторыми примерами, от-
носящимися к растениям. Это как бы перекинуло мостик между
концепциями эффекта Уоллеса и смещения признаков. Выше я
отметил, что эти две концепции параллельны, однако можно
также сказать, что в некоторых случаях они сливаются.
Один из примеров Валена [886] касается величины цветков
у пар видов Solanum в Мексике. Цветки S. tumholtzianum и
о. grayi имеют одинаковую величину в аллопатрических частях
своих ареалов, но в симпатрических областях они заметно раз-
личаются по величине. То же самое наблюдается у другой пары
видов — S. citrullifolium и S. rostratum. Величина цветков у этих
растений коррелирует с размерами опыляющих их пчел. Это
\ Гл. 15. Отбор на репродуктивную изоляцию 187
смещение признаков объясняют тем, что в областях симпатрии
оно снижает конкуренцию за пчел, относящихся к одному виду.
Такая дифференциация, по-видимому, ведет к развитию механи-
ческой изоляции между видами [886].
Я согласен с тем, что это — пример смещения признаков и
что он удачно назван смещением репродуктивных признаков,
поскольку дифференциация, очевидно, возникает в результате
конкуренции за жизненно важный фактор, в данном случае — за
опылителей. Репродуктивная изоляция при этом, несомненно,
возникает, однако она, по-видимому, не является продуктом от-
бора на создание изоляции как таковой.
Параллельный случай, относящийся к окраске цветков и дру-
гим их признакам, описан для пары видов Fuchsia parviflora и
F. encliandra [87]. Здесь также различия в признаках цветков,
наблюдаемые между двумя видами в области симпатрии, связа-
ны. с более выраженными различиями между их опылителями
в этой области, чем в аллопатрических областях.
Во многих реальных случаях, касающихся цветковых расте-
ний и их опылителей, два селективных процесса — отбор на сни-
жение конкуренции за опылителей и отбор на изоляцию цвет-
ков, — вероятно, переплетаются друг с другом.
Часть пятая
Естественная гибридизация
и ее результаты
16 Естественная гибридизация
Проблема естественной гибридизации в животном и раститель-
ном царствах всегда привлекала особое внимание человека-
Греческая мифология изобилует самыми причудливыми и неве-
роятными примерами гибридов между животными; составители
средневековых травников описали несколько подлинных гибри-
дов в европейской флоре. Возникновение научного подхода к
вопросам гибридизации датируется серединой XVIII в.
Появление трудов, принадлежащих школам Кельрейтера и
Линнея, которые придерживались противоположных взглядов,,
следует считать началом научного исследования проблемы гиб-
ридизации у растений. «Vorlaufige Nachricht» Кёльрейтера [444J
публиковалась с 1761 по 1766 г. Труд Линнея «Disquisitio de se-
xu plantarum» [513] впервые увидел свет в 1760 г. В 1751 г. один
из учеников Линнея, придерживавшийся его взглядов, — Хаарт-
ман [321}—опубликовал свой труд «Plantae Hybridae».
После Кёльрейтера и Линнея вопрос о гибридизации расте-
ний в природе обсуждался многими авторами на всем протяже-
нии XIX и XX вв., что привело к накоплению обширнейшей лите-
ратуры по данному вопросу. Объем литературы столь велик, что
дать ее полный обзор не представляется возможным; поэтому
мы и не пытаемся это сделать.
Большая часть литературы по гибридизации в природе носит
описательный характер и состоит из сообщений о гибридах, об-
наруженных в той или иной группе растений. Подобного рода
специальные сведения занимают надлежащее им место в архи-
вах систематики. Биолога-эволюциониста больше интересуют об-
щие выводы, подкрепляемые частными данными. В соответствии
с этим в нашем изложении основное внимание уделяется общим
принципам.
На протяжении длительной истории изучения естественной
гибридизации у растений рассматривались две общие проблемы:
1) являются ли естественные гибриды уродцами или представ-
ляют собой нормальное явление и какова частота их появления
Гл. 16. Естественная гибридизация
18»
в разных группах растений или флорах? 2) приводит ли есте-
ственная гибридизация к каким-либо значительным эволюцион-
ным последствиям? Иными словами, возникают ли иногда из-
естественных гибридов новые расы или новые виды? Эти проб-
лемы обсуждались многими авторами, принадлежавшими к раз-
ным поколениям, причем нередко разные авторы, основываясь-
на одних и тех же данных, приходили к различным выводам.
В этой и в последующих главах мы попытаемся разобрать две-
сформулированные здесь обширные проблемы.
В настоящей главе рассматривается образование спонтанных
гибридов в природе. Гл. 17 посвящена эволюционным продук-
там таких гибридов, вплоть до расового уровня. В гл. 19 обсуж-
дается возникновение новых видов в результате гибридизации.
Многим читателям будет полезно, если мы перечислим здесь-
некоторые из наиболее значительных обзоров. Очень ценным
справочным изданием может служить книга Робертса «Гибри-
дизация растений до Менделя» [688], в которой собраны выдерж-
ки из трудов ранних исследователей — от Линнея и Кёльрейтера
до Фокке. Современные обзоры по естественной гибридизации у-
растений с соответствующими литературными ссылками принад-
лежат Хейзеру [365, 372], Андерсону [15, 16], Стеббинсу [780,
786], Бейкеру [39], Стейсу [767] и Левину [476]. Сводки по
естественной гибридизации у животных опубликованы в числе-
прочих Майром [569, 570] и Ремингтоном [672].
Гибридизация как процесс, обратный
дивергенции
Образование двух или нескольких репродуктивно изолирован-
ных популяций из общей предковой скрещивающейся популя-
ции — процесс чрезвычайно длительный. Дивергенция между от-
дельными линиями может в конце концов достигнуть уровня
полной репродуктивной изоляции, когда скрещивание между ни-
ми становится уже совершенно невозможным. У высших расте-
ний такая стадия абсолютной репродуктивной изоляции, вероят-
но, почти во всех случаях бывает обусловлена развитием полной
несовместимости между линиями, поскольку всегда существуют
способы обойти полную стерильность гибридов, а внешние изо-
лирующие механизмы бывают совершенными лишь в редких
случаях, а быть может, и никогда.
Между начальной и конечной стадиями видообразования
происходит неуклонное снижение интенсивности скрещивания
дивергирующих популяций и, напротив, прогрессивное развитие-
изоляции, создаваемой средовыми факторами, и репродуктивной
изоляции. Спонтанное скрещивание между популяциями, кото-
рые в прошлом дивергировали до уровня дизъюнктивных рас„
490
Часть пятая. Естественная гибридизация и ее результаты
полувидов или видов и которые разделены частичной экологиче-
ской или репродуктивной изоляцией или как той, так и другой,
представляет собой естественную гибридизацию.
Термин гибрид используется чрезвычайно широко, в том чис-
.ле для обозначения продуктов скрещивания между особями с
различными генотипами, принадлежащими к одной и той же по-
пуляции, и продуктов прививок, в которых привои и подвои при-
надлежат к разным видам или родам. Для того чтобы можно
•было плодотворно обсуждать естественную гибридизацию с эво-
люционной точки зрения, необходимо несколько ограничить при-
менение этого термина. Стеббинс [780] определяет гибридизацию
как «скрещивание между индивидуумами, принадлежащими к
«отдельным популяциям, которые обладают различными адаптив-
ными нормами». В определении Стеббинса подразумевается
имевшая место в прошлом эволюционная дивергенция, а в оп-
ределении, которое мы сформулировали выше, о ней говорится
прямо.
Гибридизация в этом смысле представляет собой процесс,
«обратный эволюционной дивергенции. Процесс этот может быть
локальным или распространиться более широко в зависимости
от того, сколь велика охватываемая гибридизацией географиче-
ская область. Гибридизация может происходить между симпа-
трическими видами или между расами, которые были прежде
разобщены и дискретны, но затем восстановили контакт по ка-
кому-либо новому географическому фронту. В последнем случае
результатом будет полоса изменчивых промежуточных типов,
известная под названием зоны вторичной интерградации, или
гибридной зоны (см. [672]). В конечном счете результаты есте-
ственной гибридизации могут быть очень кратковременными
или, как мы увидим в дальнейшем, длительными.
Гибриды и скопления гибридов
Встречающиеся в природе межвидовые гибриды (по-видимому,
jгибриды первого поколения) обнаружены во всех крупных груп-
пах растений и во всех хорошо изученных флорах. Хаартман
[321] в своей книге «Plantae Hybridae», опубликованной в 1751 г.,
«описал ряд естественных гибридов, а также спонтанные гибри-
ды, возникшие в садах. Одним из них был естественный гибрид
между Trifolium repens и Т. pratense, встречающийся в разных
частях Швеции (см. [688]).
К 1891 г. Кернер [431] имел возможность привести список
евстречающихся в природе гибридов европейской флоры, в кото-
рый вошло свыше 1000 межвидовых гибридов, относящихся ко
многим группам растений — от мхов, папоротников и хвощей до
□свойных и сережкоцветных деревьев и травянистых покрытосе-
Гл. 16 Естественная гибридизация
1915
менных. Более новые списки естественных гибридов опубликова-
ны Алленом [4], Хейзером [365] и Стейсом [767].
Ботаники уделяют особенно большое внимание деревьям и.
кустарникам того или иного региона, а поэтому у крупных дре-
весных растений гибридные индивидуумы, даже если это еди-
ничные экземпляры, обычно бывают выявлены, и сообщения о-
них публикуются довольно часто. Кернер [431] обнаружил на
Дунае гибридную иву, вероятно имевшую конституцию Salix:
incanaxdaphnoides, в 1852 г., т. е. в то время, когда существо-
вание естественных гибридов все еще оспаривалось ботаниками.
Конечно, и после 1852 г. в природе было обнаружено множест-
во гибридных ив, возникших в результате самых разнообразных
межвидовых комбинаций (ср. [736]).
Многие авторы описали естественные гибриды дубов, проис-
ходящие от разных родительских видов (см., например, [57, 122^
343, 633, 839, 866, 903]). В Калифорнии изредка встречаются
гибриды между Pinus coulteri и Р. jeffreyi, а в шт. Миннесота —
между Picea mariana и Р. glauca [516, 917]. У Ceanothus доволь-
но обычны внутрисекционные гибриды; кроме того, в природе
встречаются отдельные межсекционные гибриды [537, 614]. Спи-
сок естественных гибридов древесных и травянистых растений
можно продолжать бесконечно.
Гибрид Fi может быть плодовитым, частично плодовитым,
высокостерильным или полностью стерильным. Во всех случаях,
за исключением последнего, он производит некоторое количест-
во потомков последующих поколений Различие между высокой
степенью гибридной стерильности и полной стерильностью мо-
жет быть очень невелико, составляя доли процента, по, по мне-
нию многих ученых, имеет при этом важное биологическое зна-
чение.
Стеббинс [786] приводит один численный пример, который
тем но менее достаточно реалистичен Допустим, что у нас есть
гибрид многолетнего травянистого растения, образующего корне-
вища, например Elymus, и что этот гибрид высокостерилен —
фертильность его семян равна всего 0,001% Со временем такой
гибридный индивидуум может дать начало клону, выбрасываю-
щему ежегодно по 1000 цветоносных побегов с 200 цветков на
каждом. Это составит в общей сложности 200 000 цветков, а сле-
довательно, 200 000 потенциальных семязачатков в год А посту-
лированная 0,001 %-ная фертильность семян означает, что этот
гибридный клон может дать два семени в год или 200 семян за
столетие [786].
Частично плодовитые гибриды Fi способны размножаться-
половым путем в результате самоопыления, родственных скре-
щиваний с сестринскими гибридными растениями или возврат-
ных скрещиваний с одним или обоими родительскими видами.
«92 Часть пятая. Естественная гибридизация и ее результаты
-Получающиеся при этом потомки F2 могут затем продолжать
скрещиваться друг с другом или с исходными, родительскими
^растениями. В результате образуется скопление гибридов —
•крайне изменчивая смесь видов, гибридов, беккроссов и реком-
бинантов, возникающих в более поздних поколениях.
Скопления гибридов известны во многих группах растений.
Для наших целей достаточно перечислить несколько типичных
примеров. У древесных растений скопления гибридов описаны
-в числе прочих у Juniperus [209, 697], Quercus (59, 343], Aescu-
lus [341, 342], Eucalyptus [115] и Opuntia [304]. Среди травяни-
стых покрытосеменных аналогичными примерами служат Carex
1179], Viola [66, 700], Geum [230, 557], Aquilegia [94, 270], Mi-
.mulus секция Diplacus [54], Epilobium [666] и Helianthus [366].
Среди папоротников скопления гибридов можно обнаружить у
.Рteris, исследованного Уокером [870].
Полный список естественных гибридов и скоплений гибридов
у растений начинался бы с Achillea и кончался бы Zinnia. Слу-
чайная или спорадическая естественная гибридизация —нор-
мальное явление в царстве растений, возникающее в тех слу-
чаях, когда два или несколько родственных видов приходят в
.контакт. Но явление это не универсально. Привлекают внимание
те исключительные случаи, в которых два или несколько близ-
•ких видов произрастают на одной и той же территории, не гиб-
ридизируя (насколько об этом можно судить по данным поле-
вых исследований).
Gilia tricolor не гибридизирует со своими симпатрическими
.родичами в Калифорнии [269]. В Великобритании Ranunculus
•acris, R. repens и bulbosus часто встречаются вместе, однако
гибриды между ними неизвестны [350]. Papaver dubiutn и Р. 1е-
coquii также сосуществуют на Британских островах, не гибри-
дизируя [539]. Lotus corniculatus и близкие ему виды сохраняют
обособленность в Калифорнии и на севере Европы, но, по-види-
мому, гибридизируют в Средиземноморской области [306]. Семь
.видов Leavenworthia симпатричны на юго-востоке США, но не
образуют гибриды [692]. У Sedum в центральных районах Мек-
сики и у Silene на западе Северной Америки гибриды очень ред-
ки [116, 451]. Juniperus virginiana и J. ashei не гибридизируют
в различных зонах симпатрического контакта на плато Озарк
на юге центральной части США, хотя в более ранних работах
л сообщалось о существовании гибридов между ними fl, 215].
Внутренний контроль над гибридизацией
•Самая важная причина отсутствия гибридизации между родст-
венными симпатрическими видами — это несовместимость в ши-
роком смысле, т. е. наличие не только внутренних барьеров, дей-
Гл 16 Естественная гибридизация
193
ствующих до оплодотворения, но также барьеров, действующих
на ранних стадиях после оплодотворения.
Если говорить о перечисленных в предыдущем разделе слу-
чаях отсутствия гибридизации, то имеются экспериментальные
данные, указывающие на существование очень мощных барьеров
несовместимости между видами групп Gilia tricolor, Ranunculus
acris, Lotus corniculatus и Leavenworthia [269, 306, 350, 692].
Естественные гибриды между членами разных секций редки
у Ceanolhus и не известны у Quercus. Кроме того, как показали
экспериментальные скрещивания, у этих родов существуют очень
мощные барьеры несовместимости между видами, принадлежа-
щими к разным секциям, о чем уже говорилось в гл. 11 (см.
[614])
Однако барьеры несовместимости — это лишь часть пробле-
мы Papaver dubium и Р. lecoquii легко скрещиваются на экспе-
риментальных участках, по, как мы уже отмечали, в Великобри-
тании в естественных условиях они не гибридизируют [638]. Кру-
кенбергу [451] удалось добиться успеха почти во всех скрещива-
ниях, которые он пытался произвести между 28 западноамсри-
канскими видами Silene, по в природе эти виды сохраняют обо-
собленность и лишь в редких случаях образуют гибриды.
Внешний контроль над гибридизацией
Известно много примеров, когда данная пара родственных видов
растений или животных не гибридизирует в природе, но легко
образует гибриды в ботаническом саду или зоопарке Кельрей-
тер писал об этом еще в 1761 г. [444, 688]:
«[Маловерояшо, чтобы] при том упорядоченном устройстве растительно-
го царства, которое создала природа, могло возникнуть гибридное растение
Природа которая всегда, даже в случаях кажущегося величайшего беспо-
рядка, привержена к самому великолепному порядку, заранее исключила эту
путаницу [Однако] в ботанических садах, где в тесном пространстве собра-
ны всевозможные растения со всех частей земного шара, гибридные растения
могут возникнуть, особенно если разместить их в соответствии с их система-
тическим положением, а следовательно, поместить рядом растения, облада-
ющие наибопьшим сходством По крайней мере здесь человек в известном
смысле предоставляет растениям ту возможность, которую он дает своим
животным, привезенным из очень удаленных одна от другой частей земного
шара, которых он вопреки природе держит в плену в зоопарке или в еще
более ограниченном пространстве».
Позднее Кернер [431] сформулировал положение о роли мес-
тообитания в ограничении естественной гибридизации более кон-
кретно.
«Если данный вид в какой-либо местноеги благоденствует, представлен
большим числом индивидуальных растений и возобновляется в потомках,
которые в основном не изменены, то можно допустить, что организация это-
го вида соответствует почве и климату данной местности В отсутствие га-
13—44
194 Часть пятая, Естественная гибридизация и ее. результаты
кой гармоничной связи не может быть и речи о процветании вида; напротив,
он рано или поздно должен вымереть. Это соответствие климата и почвы ор-
ганизации растения, проявляющееся в его внешней форме, должно сущест-
вовать также и для вновь возникшего гибрида, с тем чтобы те несколько
растений, которые появляются в каком-либо месте, могли выжить в своих
первоначальных местообитаниях и дать начало многочисленным потомкам.
Иногда такое соответствие существует, а иногда его нет. В последнем слу-
чае гибрид исчезает, едва появившись на свет. Но если даже его организа-
ция адаптирована к почве и климату того места, где он возник, ему пред-
стоит еще выдержать борьбу с теми видами, которые уже обосновались в
этой местности, а в особенности со своими собственными родительскими ви-
дами. Если эти последние пышно растут в данной местности и численность
их велика, то новой форме нелегко завладеть территорией».
Кернер приходит к выводу, что гибридные растения могут
обосноваться в местообитании своих предков только в том слу-
чае, если они в адаптивном смысле превосходят родительские
виды или если они могут найти подходящее свободное местооби-
тание, не занятое этими последними. Подобные условия встреча-
ются редко. Кернер [431] приводит в качестве примеров Rhodo-
dendron, Salvia и Nuphar, гибридные формы которых успешно за-
селили новые местообитания, отличные от местообитаний роди-
тельских видов.
В начале современного периода изучения естественной гиб-
ридизации важное значение факторов среды в предотвращении
или допущении естественной гибридизации подчеркивали Дю
Рие [181], Виганд [895], Эплинг (202], Андерсон [14, 15], Хейзер
[365], Стеббинс [780] и многие другие исследователи.
Виганд [895] указывал на то, что естественная гибридизация
между разными видами растений нередко бывает ограничена
определенными местностями, в которых среда была нарушена в
результате вмешательства человека. На Ньюфаундленде и в
шт. Мэн, например, гибридные формы Amelanchier и Rubus
встречаются в нарушенных местообитаниях вдоль железнодо-
рожных путей, тогда как в нетронутых лесах или в лесах, гра-
ничащих с этими путями, произрастают родительские виды в
чистом виде [895].
Это наблюдение было распространено на многие другие груп-
пы растений. У ирисов естественная межвидовая гибридизация
происходит главным образом в местообитаниях, нарушенных в
результате деятельности человека — земледелия, постройки до-
рог, лесных разработок и т. п., — как в дельте Миссисипи, так
и на тихоокеанском побережье [15, 462, 655, 686, 860]. Salvia
apiana и S. mellifera, которые симпатричны на обширной терри-
тории в южной части Калифорнии, остаются обособленными там,
где их полынно-кустарничковое сообщество находится в своем
естественном состоянии, но в тех местах, где среда была искус-
ственно нарушена, между ними наблюдается локальная гибри-
дизация [17, 202, 264].
Гл. 16. Естественная гибридизация
195
В 1891 г Кернер дал объяснение корреляции между гибри-
дизацией и разрушением местообитаний, которое в несколько
расширенном виде сейчас принимается большинством исследо-
вателей. В замкнутом стабильном сообществе для образующих-
ся время от времени гибридных зигот подходящих местообита-
ний пет. Стабилизирующий отбор элиминирует их почти сразу
же после их появления. Но в тех местах, где естественное со-
общество нарушается в результате строительства дороги, пере-
выпаса и т. п , что приводит к созданию новых свободных место-
обитаний, гибриды могут утвердиться и это действительно про-
исходит. Иными словами, изоляция, создаваемая средой, подав-
ляет гибридизацию между совместимыми друг с другом видами
в стабильном замкнутом сообществе, но в открытом местообита-
нии она частично утрачивает свою эффективность.
Гибридизация местообитания
Андерсон [14, 15] расширил концепцию контроля над гибридиза-
цией со стороны среды, распространив ее на второе и последую-
щие поколения гибридов. Он указывал на существование тако-
го же расщепления по физиологическим различиям, как и по
различиям морфологическим. Если Fi носит промежуточный ха-
рактер и обладает единообразием в отношении экологических
предпочтений, то F2 и последующие поколения отличаются боль-
шим разнообразием рекомбинантов по физиологическим призна-
кам. Если родительские виды различаются по нескольким неза-
висимым физиологическим признакам, то последующим поколе-
ниям их гибридных потомков потребуются десятки или сотни
экологических ниш. Андерсон [14, 15] приходит к выводу, что
потомки Fi могут удовольствоваться промежуточным местооби-
танием, но что для выживания сколько-нибудь представитель-
ной выборки поколения F2 потребуется разнообразный набор
экологических ниш — своего рода «гибридизация местообита-
ния».
Как писал Андерсон [15, гл. 17].
«Хорошо известно, что гибридам, для того чтобы выжить, необходимы
промежуточнье местообитания Не учитывалось, однако, то, что для сохра-
нения не только первого гибридного поколения, но и дальнейших поколений
необходимы нс просто промежуточные местообитания, а местообитания,
предоставляющие также все возможные сочетания контрастирующих разли-
чий первоначальных местообитаний Только путем гибридизации местообита-
ния могут сохраниться в природе гибридные рекомбинации»
Промежуточные местообитания встречаются в природе не
слишком редко. Но местообитания, которые в достаточной степе-
ни гибридизированы, чтобы допустить относигсльпо свободное
1 Q*
196 Часть пятая Естественная гибридизация и ее, результаты
воспроизведение межвидового растительного гибрида, гораздо
менее обычны. И такие гибридизированные местообитания соз-
даются в современном мире главным образом в результате дея-
тельности человека, например строительства дорог, сельскохо-
зяйственных работ и лесоразработок [15].
Замечу мимоходом, что необходимо проводить различие меж-
ду гибридизированными и свободными местообитаниями. Второе
и последующие поколения межвидовых гибридов могут выжить
как в тех, так и в других местообитаниях, но по разным причи-
нам: в открытом местообитании — вследствие ослабления меж-
видовой конкуренции и стабилизирующего отбора, а в гибриди-
зированном местообитании — вследствие разнообразного набора
новых ниш. Можно представить себе скопление гибридов, произ-
растающих либо в однородном открытом местообитании, либо
в смешанном, но более или менее замкнутом местообитании.
Следовательно, гибридизация местообитания представляет со-
бой не единственное условие, которое создает возможность для
обоснования расщепляющихся гибридных потомков.
Основной эффект деятельности человека, упоминавшейся вы-
ше, состоит в частичном удалении природной растительности
и создании тем самым открытых местообитаний. Часто эти дей-
ствия сопровождаются искусственной гибридизацией местооби-
тания вследствие смешения почвы разных типов, создания новых
градиентов затененности или влажности и т. п. А в тех местах,
где такая гибридизация происходит, создаваемые ею возможно-
сти для воспроизведения гибридов играют, вероятно, второсте-
пенную роль по сравнению с теми, которые создает наличие сво-
бодных участков.
Открытые местообитания наряду с гибридизацией местооби-
таний или без нее —обычный результат деятельности человека
в современном мире. Но на протяжении всей истории земли,
вплоть до современности и включая ее, многократно происхо-
дили разрушения биотических сообществ и возникновение но-
вых местообитаний под действием чисто природных факторов.
Обвалы, наводнения, извержения вулканов и пожары, вызван-
ные ударом молнии, создавали новые открытые местообитания
локального масштаба. Процессы горообразования, поднятия су-
ши из моря, ледниковые периоды и периоды засухи оказывали
на природные сообщества воздействия крупного масштаба. Да-
леко идущие последствия имели также биотические факторы,
подобные иммиграции новых крупных растительноядных орга-
низмов или новых -возбудителей заболеваний.
Климатические и геологические изменения обусловливают
миграции растений, приводящие к новым симпатрическим кон-
тактам между видами. В то же самое время, как полагают
Андерсон [14, 16] и другие авторы, эти физические изменения
Гл 16 Естественная гибридизация
197
или изменившиеся биотические факторы или тс и другие могут
открыть новые незанятые местообитания, которые будут слу-
жить местом размножения для естественных гибридов
Сопоставление высших растений с высшими
животными
Естественные гибриды обнаружены в разных крупных группах
высших животных Соответствующие списки для птиц состав-
лены Кокремом [116] и Грэем [308], а для млекопитающих —
Грэем [307]. Ценные обзоры естественной гибридизации у живот-
ных принадлежат Хэббсу [386], Майру [569, 570] и Моору [584].
Подробное обсуждение сложной проблемы гибридизации у
животных и освещение различных связанных с ней частных во-
просов читатель найдет в этих сводках и в приведенных в них
обширных библиографиях Здесь же нас интересует нс гибриди-
зация у животных как таковая, а широкое сравнение между жи-
вотными и растениями в отношении частоты гибридизации и ее
эволюционной роли Эти два царства действительно различают-
ся по указанным параметрам
Установлено, что естественная гибридизация оказывает влия-
ние на характер изменчивости в отдельных группах животных,
в частности у некоторых бабочек, цикадок, рыб и птиц [383, 386,
387, 672, 696, 722, 723]. Перечень примеров можно было бы про-
должить. Вместе с тем во многих других группах животных, та-
ких, как двукрылые или млекопитающие, естественная межви-
довая гибридизация, очевидно, встречается редко и не влечет
за собой никаких последствий.
Но увеличение числа тех или других примеров не поможет
нам глубже понять роль гибридизации в эволюции животных.
Как было показано в этой главе, естественная гибридизация
наблюдается в одних группах растений, по отсутствует в дру-
гих Широкие сравнения между царствами растений и живот-
ных в отношении важности гибридизации пока еще нельзя про-
водить на удовлетворительной количественной основе, и поэто-
му приходится искать другие подходы к этой проблеме
Несколько поколений ботаников считали, что гибридизация
играет важную роль в эволюции растений (см., например, [14,
16, 163, 181, 205, 317, 365, 431, 519, 609, 780]). А несколько поко-
лений зоологов пришли к выводу, что гибридизация не играет
важной общей роли в эволюции животных (см., например, [165,
214, 564, 569, 882, 889])
Я привожу здесь высказывания различных авторов не для
того, чтобы решить научный вопрос подсчетом голосов или апел-
ляцией к авторитетам Суть дела состоит в том, что авторы-бо-
тапики и авторы-зоологи выражают свои взгляды, основываясь
198 Часть пятая. Естественная гибридизация и ее результаты
на большом запасе знаний, который не следует легко отбрасы-
вать.
С этим связан другой вопрос, мнения по которому также до-
вольно единодушны, а именно вопрос об относительной частоте
хороших биологических видов в царстве животных и царстве
растений. Как отмечалось в гл. 6, отклонения от идеального
стандарта дискретных биологических видов очень обычны у
высших растений, но гораздо менее обычны у высших животных.
Эти отклонения от организации биологического вида в случае
высших растений обычно сопровождаются естественной гибри-
дизацией.
Другой подход состоит в сравнении частоты полиплоидии в
двух царствах. Хорошо известно, что примерно половина видов
высших растений — полиплоиды, тогда как среди животных по-
липлоидия известна лишь в нескольких группах. Кроме того,
большинство встречающихся в природе полиплоидных расте-
ний— аллополиплоиды {102]. Это служит явным указанием на
важное значение гибридизации в эволюции растений; аналогич-
ные указания в отношении царства животных отсутствуют.
Этологические барьеры, препятствующие гибридизации, у
высших животных развиты гораздо сильнее, чем у растений.
У животных эти барьеры, препятствующие скрещиванию, игра-
ют главную роль и высоко эффективны, если не считать тех слу-
чаев, когда создаются аномальные условия. В большинстве
групп растений подобные этологические барьеры совершенно от-
сутствуют. А в тех группах, в которых они существуют, — в эво-
люционно наиболее продвинутых семействах цветковых расте-
ний— они носят вторичный характер, находясь в зависимости
от поведения посещающих цветки насекомых, и поэтому менее
эффективны, чем непосредственные предпочтения при выборе
брачного партнера, существующие у животных.
Хэббс [386] и Майр [569] отмечают, что среди животных гиб-
ридизация обычна главным образом в группах с наружным
оплодотворением, в частности у рыб и у некоторых амфибий, но
не у рептилий, птиц, млекопитающих и насекомых, для которых
характерно внутреннее оплодотворение. Правда, и здесь не об-
ходится без редких исключений. Царство растений в целом срав-
нимо с той частью царства животных, которой свойственно на-
ружное оплодотворение, поскольку у всех растений перекрестное
оплодотворение обеспечивается внешними агентами. С этим
коррелирует также сходство в отношении частоты гибридиза-
ции у растений и у животных с наружным оплодотворением.'
17 Интрогрессия
Воспроизведение естественных гибридов в очень многих слу-
чаях следует по определенному пути, известному под названием
интрогрессии Интрогрессивная гибридизация, по определению
^AHjTgp г л н я это повторные—вози р а £ны е скрещивания _ ё^тест-
венно^ю-шбдида^с одной или обеими родительскими популяция-
ми^ Она приводит к переносу генов от одного вида или полувида
к другому, что сопровождается преодолением барьеров, препят-
ствующих скрещиванию fl5, 16, 191.
Ранние работы Дю Рие [181] и Марсден-Джонса [557] по-
могли понять явление интрогрессии, хотя эти авторы и не ис-
пользовали данный термин [293, 365]. Марсдеп-Джопс [557] про-
вел генетическое и полевое исследование Geum urbanum, G. riva-
le и их гибрида, G. intermedium, в Англии. Родительские виды
широко распространены в Англии и экологически разделены;
гибриды также часто встречаются в некоторых местообитаниях.
Марсден-Джопс [557] обнаружил два места, в которых формы,
получающиеся в результате возвратных скрещиваний G. rivaleX
XG. intermedium, замещали чистых G rivale Он утверждает,
что это «пример возникновения дикой популяции нового типа в
результате межвидовой гибридизации» [557].
Дю Рие провел популяционные исследования групп видов у
Dracophyllum, Coprosma и Salix в новозеландской и скандинав-
ской флорах. По его данным, члены какой-либо пары видов гиб-
ридизируют в определенных местах, а кроме того, у популяций
тех же родительских видов наблюдается конвергенция по мор-
фологическим признакам в зонах симпатрического перекрыва-
ния Дю Рис объяснял эту конвергенцию гибридизацией, за ко-
торой следуют возвратные скрещивания. В результате такого
процесса один вид, как он выразился, «засоряется» определенны-
ми генами другого вида [181].
Факторы, способствующие интрогрессии
Рассмотрим колонию, состоящую из одного или двух видов и их
гибридов Fi. В такой колонии действуют генетические и эколо-
гические факторы, которые способствуют возвратным скрещива-
ниям, а тем самым направляют воспроизведение гибридов по
пути интрогрессии. Рассмотрим сначала популяционно-генети-
ческие факторы.
Естественный гибрид обычно бывает представлен одним или
несколькими растениями, находящимися в окружении обширной
популяции одного или обоих родительских видов Далее, гибри-
200 Часть пятая. Естественная гибридизация и ее результаты
ды Fi обычно бывают по крайней мере полустерильными, а час-
то и высокостерильными. В таких условиях они вносят в гамет-
ный фонд колонии очень небольшое число жизнеспособных га-
мет по сравнению с вкладом родительских видов. Вследствие
этого в очень большой части случаев эффективных оплодотворе-
ний, в которых участвует какой-либо гибрид, вторым партнером
окажется растение одного из родительских видов, что приведет
к появлению потомков от возвратных скрещиваний, т. е беккрос-
сов [780].
Экологические факторы, способствующие интрогрессии, пред-
ставляют собой естественный результат условий в местообита-
ниях, которые были описаны в гл. 16. Каждый родительский вид
хорошо адаптирован к той экологической нише, которую он
обычно занимает. В открытом или промежуточном местообита-
нии могут поселиться гибриды Ft и некоторые потомки F2 про-
межуточного типа. Открытое местообитание или местообитание,
до некоторой степени гибридизированное, может дать приют не-
которым рекомбинантам, возникшим к результате естественной
гибридизации. Однако большую часть возможностей для за-
крепления гибридных потомков в обычном месте встречи между
двумя видами или полувидами получают те генотипы, которые по
своим физиологическим признакам приближаются к одной или
другой родительской форме. Необходимыми экологическими пред-
почтениями скорее всего будут обладать потомки от возвратных
скрещиваний, для которых существующие в этом месте условия
среды, по всей вероятности, окажутся благоприятными [15, 19].
Бэтке и Олстон [38] проанализировали состав одной гибрид-
ной колонии Baptista в Техасе. В качестве критериев они ис-
пользовали морфологические и биохимические признаки. На ос-
нове этих критериев они установили, что колония имеет следую-
щий состав:
Baptista ieucophaea 470 растений
Baptista sphaerocarpa 561 растение
Гибриды Fi 83 растения
Продукты возвратных скрещиваний с В leuco- 37 растений
phaea
Продукты возвратных скрещиваний с В. sphaero- 19 растений
сагра
Таким образом, в естественных условиях растения, происходя-
щие от гибридов, распадаются на два класса реципрокных бек-
кроссов [38].
Описанные выше популяционно-генетические и экологические
факторы продолжают действовать в последующих поколениях
гибридов и ведут к повторным возвратным скрещиваниям. В ре-
Гл. 17. Интрогрессия
201
зульгате некоторые гены или хромосомные участки одного вида
проникают в генофонд другого вида-. А вследствие этого полу-
чившие их популяции вида-реципиента конвергируют в направ-
лении вида-донора по разным морфологическим, биохимическим
и физиологическим признакам [15, 16, 19, 181]. .
Методы
Два самых старых и наиболее широко распространенных мето-
да изучения интрогрессии основаны на использовании гибридно-
го индекса и «многомерных» диаграмм разброса (pictorialized
scatter diagrams). Получающиеся при этом графики двух ти-
пов дают наглядное представление о том, что происходит в ин-
трогрессивных популяциях; таким образом, эти графики служат
как для анализа, так и для описания процесса интрогрессии.
Вначале Андерсон и его сотрудники пользовались гибридным
индексом. Многомерные диаграммы разброса — существенное
усовершенствование, введенное ими позднее [15, 18].
Основная причина, обусловившая использование этих стан-
дартных методов и их последующее усовершенствование, связа-
на с многофакторным сцеплением. Как уже говорилось в гл 8,
оно представляет собой частичное и переплетающееся сцепление
между различными количественными признаками. Многофактор-
ное сцепление ведет к слабой ассоциации признаков, или связ-
ности признаков, у расщепляющихся гибридных потомков.
Андерсон [13, 15, 16] показал, что многофакторное сцепление и
связность признаков создают основу для обнаружения и коли-
чественной оценки результатов интрогрессии.
Следует ожидать, что в результате возвратных скрещива-
ний и интрогрессии возникнет изменчивая популяция, прибли-
жающаяся по своим фенотипическим признакам к одному из
родительских видов. Гипотезу о том, что такое сочетание измен-
чивости и конвергенции фактически обусловлено интрогрессией,
можно проверить, использовав критерий ассоциации признаков.
Этот критерий применим в случае признаков, не связанных меж-
ду собой морфогенетически, как, например, опушенность стебля
и окраска цветков; он неприменим к таким плейотропным приз-
накам, как форма листьев и форма чашелистиков.
Если какие-либо гены, определяющие один признак, ввести
от вида-донора в интрогрессивную популяцию другого вида, то
тогда в результате многофакторного сцепления рекуррентная
родительская форма получит вместе с ними другие генные сис-
темы и детерминируемые ими признаки Поэтому интрогрессив-
ная популяция будет походить на рекуррентную родительскую
форму, но будет варьировать в направлении вида-донора не
202 Часто пятая. Естественная гибридизация и eei результаты
только по одному количественному признаку, а по многим или
по всем таким признакам [15, 16].
Метод гибридного индекса предполагает рассмотрение опре-
деленного числа, например десяти, отдельных признаков, разли-
чающихся у двух предполагаемых родительских видов. Припи-
шем экстремальному состоянию каждого признака, обнаружен-
ного у вида А, по произвольной шкале оценку 0, противопо-
ложному его состоянию у вида Б — оценку 2, а промежуточному
состоянию у известных или предполагаемых гибридов Fi —
оценку 1. Или же некоторым морфологическим признакам при-
дается относительно больший вес, чем другим, для того чтобы
отразить их относительную полезность в качестве показателей
лежащих в их основе генетических детерминант. В первом слу-
чае общая оценка, или гибридный индекс, отдельного растения,
принадлежащего к виду А и измеренного по десяти признакам,
будет равна нулю, а значение гибридного индекса для растения,
принадлежащего к виду Б, будет равно 20. Эти противополож-
ные оценки служат эталоном для определения промежуточности
и изменчивости гибридной, популяции.
Далее следует определить значения гибридного индекса для
каждого отдельного растения в выборке, взятой из гибридной
колонии, и представить распределение значений индекса по
частоте в виде гистограммы. Колония, состоящая из родитель-
ских видов и гибридов Fi, представлена на гистограмме рядом
пиков в экстремальных и промежуточных областях. В отличие
от этого популяция, подверженная сильной интрогрессии, пред-
ставлена значениями гибридного индекса в диапазоне от зна-
чений, характерных для гибридов Fi, до значений, характерных
для одной из родительских форм. Наконец, у популяции, подвер-
женной слабой интрогрессии, выявляется заметный сдвиг моды
от одного из экстремальных состояний к промежуточному, сопро-
вождаемый усилением индивидуальной изменчивости.
В следующем разделе будет описано применение метода гиб-
ридных индексов к реальным примерам интрогрессии у Iris и
Helianthus (рис. 17.2 и 17.5).
Метод с использованием гибридного индекса объединяет ре-
зультаты измерения отдельных признаков. Два растения могут
иметь одинаковые промежуточные оценки вследствие разных,
но компенсаторных отклонений от некой экстремальной комби-
нации признаков, в связи с чем с помощью этого метода анали-
за сведений относительно корреляции признаков получить почти
не удается. Напротив, при использовании метода многомерных
диаграмм разброса эти сведения сохраняются, так что во многих
случаях предпочтение отдают последнему методу.
Обычная диаграмма разброса, в которой два количественных
признака представлены осями координат, а каждое отдельное
Гл. 17 Интрогрессия
203
растение — точкой, отражает изменчивость данной популяции по
двум признакам. В многомерной диаграмме разброса дополни-
тельные признаки представлены длиной усиков, отходящих от
этих точек. Подобная диаграмма почти с одного взгляда позво-
ляет охватить всю картину изменчивости в популяции, варьиру-
ющей одновременно по трем или большему числу признаков
Затем можно перейти к построению ряда многомерных диаг-
рамм разброса для разных популяций предполагаемых роди-
тельских видов и популяций, происходящих от них в результате
интрогрессии. Такие ряды дают возможность непосредственно
сравнивать несколько популяций по диапазону и коррелирован-
ное™ их изменчивости В следующем разделе это показано на
примере Iris. Использование коэффициентов корреляции для пар
изменчивых признаков в гибридных популяциях, казалось бы,
должно давать определенное преимущество, поскольку эти ко-
эффициенты количественно оценивают меру связности призна-
ков. Я пользовался этим методом [270], однако он так и не по-
лучил широкого распространения
Сравнительно недавно несколько исследователей, хорошо
знакомых со статистикой или математикой, внесли ряд усовер-
шенствований в метод гибридных индексов, используя классифи-
цирующие функции [216, 608], различные метрические методы
[608, 882], метод главных компонент [608, 629], многомерный
статистический анализ [629] и кластерный анализ [126]. Все эти
методы довольно сложны. Более подробное их описание можно
найти в перечисленных здесь работах.
В ранних работах по исследованию интрогрессии использо-
вались морфологические признаки Еще одно относительно не-
давнее новшество— использование биохимических признаков,
выявляемых методами электрофореза, хроматографии и други-
ми. Включение биохимических признаков, конечно, позволило
расширить ту фактическую основу, на которую опирается изу-
чение интрогрессии.
Среди ранних исследований, использующих биохимические
данные, следует назвать работы Тернера и его сотрудников на
Baptisia (Leguminosae) [9, 38, 845]. Вслед за этими работами
были опубликованы первые общие обзоры по этой проблеме
[8, 9]. В настоящее время использование биохимических призна-
ков вошло в повседневную практику. В следующем разделе мы
опишем исследование интрогрессии у Juniperus; в этом исследо-
вании были использованы как биохимические, так и другие
сложнейшие методы анализа.
204 Часть пятая. Естественная гибридизация и ев| результаты
Примеры
Из многочисленных хороших примеров интрогрессии мы рас-
смотрим здесь только четыре. Выбранные нами примеры иллю-
стрируют различные аспекты этого процесса и разные подходы
к его изучению.
Iris
Ирисы, растущие в дельте Миссисипи в южной части шт. Луи-
зиана, изучали Райли [686] и другие. В ненарушенных прирбд-
ных местообитаниях в этой области Iris fulva и I. hexagona gi-
ganiicaerulea1 симпатричны и не гибридизируют. /. hexagona gi-
ganticaerulea (=1. giganiicaerulea) растет на заболоченных поч-
вах, a I. fulva — на более сухих соседних участках и в лесах.
Там, где природные местообитания разрушаются человеком и
вместо них возникают пастбища и канавы, между двумя этими
ирисами происходит гибридизация и появляются интрогрессив-
ные популяции [686].
Райли выбрал четыре колонии и подробно изучил их. Коло-
ния G— это чистая популяция Iris hexagona giganiicaerulea,
растущая на нетронутых болотах. Колонии Н-1 и Н-2— более
или менее гибридизированные популяции, растущие на пастби-
ще в 150 м от колонии G. Колония F— чистая популяция I. ful-
va, растущая в дубовом лесу в нескольких километрах от коло-
нии G. Незаверенные популяции (G и F) служат эталонами для
сопоставления гибридизированных популяций {Н-1 и Н-2).
Из каждой колонии Райли собрал по 23 растения; для каж-
дого растения были проведены измерения по 7 признакам, отно-
сящимся к цветку (рис. 17.4). Затем результаты измерений бы-
ли оценены по произвольной шкале, по которой экстремальное
/ц/па-подобное состояние получает оценку 0, a giganticaerulea-
подобное состояние — оценки 2, 3 или 4. Оценки по всем семи
признакам для каждого растения объединяли затем в значения
гибридного индекса, колебавшиеся от 0 до 17. Распределения
значений индекса по частоте для каждой колонии представляли
в виде гистограмм (рис. 17.2).
1 В отличие от «Международного кодекса зоологической номенклатуры»
«Международный кодекс ботанической номенклатуры» признает несколько
таксономических категорий рангом ниже вида. Поэтому после названия вида
необходимо указывать таксономический ранг внутривидового таксона, если
он приводится (см. Ч. Джеффри, Биологическая номенклатура, Изд-во
«Мир», М, 1980, стр. 27—28). Так, после названия Iris hexagona должно
следовать «subsp.», «var» или «forma» в зависимости от того, какой ранг
придается названию giganiicaerulea — Прим. ред.
Гл. 17. Интрогрессия
205
Как показывают эти гистограммы, колония Н-2 по своим
морфологическим признакам в значительной мере сходна с
Iris giganticaerulea, но содержит несколько гибридных растений.
Колония Н-1, однако, представляет собой интрогрессивную по-
пуляцию, и ее диапазон изменчивости в значительной степени
Рис. 17 1 Цветки и чашелистики (увеличены) Iris fulva (А) и I hexagona
giganticaerulea (Б) [15]
заполняет разрыв в морфологических признаках между I. gi-
ganticaerulea и I fulva [686].
Позднее Андерсон [15] представил эти же данные Райли
[686] в виде многомерных диаграмм разброса (рис. 17.3 и 17.4),
эти диаграммы выявляют корреляцию признаков в родительских
гибридных популяциях.
Спустя еще некоторое время Рэндолф и его сотрудники [655]
вновь изучили этот случай. Проведя исчерпывающий анализ из-
менчивости по 13 признакам у многочисленных аллопатрических
и симпатрических популяций этих двух видов, они пришли к
выводу, что эффекты интрогрессии локализованы в нарушенных
местообитаниях в пределах зоны симпатрии. Насколько можно
было установить, каждый из видов Iris в значительной части
своего ареала не испытывал воздействия генов другого вида.
206
Часть пятая. Естественная гибридизация и ее результаты
. J
I___I_I_I_I_I—I—I—I—I—I___I—I_I_I__I_I I I
0123456789 1011 121314151617
Значения индекса
Рис. 17.2. Распределение по частоте значений гибридных индексов для
23 растений в каждой из четырех колоний ирисов в южной части Луизианы
[686].
F — Iris fulva. G. — I. hexagona giganticaerulea. H-l — популяция, подверг-
шаяся сильной интрогрессии. Н-2 — популяция, подвергшаяся слабой интро-
грессии.
Heliant has
Наш следующий пример — виды подсолнечника Helianthus
annuus и Н. bolanderi, изученные Хейзером [366]. Масличный
подсолнечник (Н. annuus) растет как сорняк по всему умерен-
ному поясу Северной Америки В Калифорнии Н. atinuus не яв-
ляется одним из элементов местной растительности; вероятно,
он сравнительно недавно был интродуцирован индейцами.
Н. bolanderi встречается в Калифорнии и Орегоне как в есте-
ственных местообитаниях, так и в зарослях сорняков. Местная
раса Н. bolanderi растет в этих двух штатах на выходах сер-
/ fulva
Колония H'2
R
RV
Размеры лепестков
Рис. 17 3 Многомерные диаграммы разброса Iris julva, I hexagona gigan
ticaerulea и двух гибридных колоний (из [15], по данным [686]).
Те же самые популяции, что и на рис. 17.2
208
Часть пятая. Естественная гибридизация и eet результаты
Трубка X ♦ •
Желтая Зеленоватая Зеленая
Форма лепестков
Обратно- Промежуточные Лопатчатые
яйцевидные,
почти
без ноготков
Тычинки | ф •
Выступающие Почти равные Включенные
ветвям столбика
Ветви столбика <
Маленькие Средние Большие
Гребень •
Отсутствует Слабо развит Хорошо развит
Размеры, лепестков----
Рис. 17.4. Объяснение символов, использованных на рис. 17.3.
пентинитов в предгорьях, тогда как сорная раса распростране-
на в центральной долине Калифорнии [366].
Эти два вида подсолнечника различаются по многим морфо-
логическим признакам. Helianthus annuus— высокое растение
с овальными листьями, а Н. bolanderi— растения от низкой до
средней высоты с ланцетовидными листьями. Кроме того, они
различаются также по опушенности, обертке соцветия,- централь-
ной части цветоложа, язычковым цветкам и. семянкам. Такие
морфологические различия служат маркерами, позволяющими
производить оценку отдельных растений и анализировать попу-
ляции методом гибридных индексов [366].
Как показали эксперименты по гибридизации, Helianthus
annuus и Н. bolanderi легко скрещиваются между собой в обоих
направлениях. Гибриды Fi промежуточны по морфологическим
признакам, конъюгация хромосом в мейозе у них нарушена, а
пыльца и семена высокостерильны [366].
Helianthus annuus и Н. bolanderi приходят в контакт в раз-
ных районах Калифорнии. Периоды цветения у них перекрыва-
ются, и в ряде мест они гибридизируют. На рис. 17.5, Д, пока-
зана картина изменчивости в одном скоплении гибридов в до-
лине р. Сакраменто в Калифорнии^ На основе морфологических
Гибридный индекс
Рис. 17.5. Частота распределения значений гибридного индекса в пяти попу-
ляциях Helianthus annuus и Н. bolanderi [365].
А. Раса Я. bolanderi с серпентинитовых почв предгорий в Калифорнии.
Б. Раса И. annuus из штата Миссури. В Сорная раса Н. bolanderi из долин
Калифорнии Г И. annuus из долины Сан-Хоакин Д. Скопление гибридов из
долины реки Сакраменто.
14—45
210
Часть пятая. Естественная гибридизация и ее результаты
признаков и коррелированной с ними фертильности пыльцы был
сделан вывод, что эта гибридная колония состоит из двух роди-
тельских видов, нескольких гибридов Fi и двух типов реципрок-
ных беккроссов [366].
Признаки неинтрогрессированных популяций Helianthus из
Сент-Луиса (шт. Миссури) и Н. bolanderi, произрастающих на
выходах серпентинитов в северной части Калифорнии, показаны
на рис. 17.5, А й Б. На рис. 17.5, Г и Д изображены для сравне-
ния популяции тех же двух видов из центральной долины Кали-
форнии, где они растут как сорняки. Можно видеть, что в каж-
дой из двух последних популяций изменчивость направлена в
сторону другой. Диапазоны изменчивости этих популяций нахо-
дятся в пределах изменчивости беккроссов из скоплений гибри-
дов; поэтому был сделан логический вывод, что они представ-
ляют собой продукты интрогрессивной гибридизации [366].
Распространение расы Helianthus bolanderi, экстремальной
по морфологическим признакам, ограничено серпентинитовыми
почвами предгорий, тогда как интрогрессивная раса того же ви-
да встречается как сорняк в центральной долине вместе с
Н. annuus. Такое экогеографическое распространение с доста-
точным основанием позволяет предположить, что вслед за ин-
тродукцией, вероятно индейцами, Н. annuus в Калифорнию на-
чалась гибридизация между этой формой и местной дикорасту-
щей формой Н. bolanderi [366]. Интрогрессия от Н. annuus к
Н. bolanderi привела затем к образованию долинной расы Н. bo-
landeri, которая смогла распространиться по всем возникшим
в это же время сорным местообитаниям. По всей вероятности,
Н. annuus в центральных районах Калифорнии испытал в свою
очередь воздействие интрогрессии со стороны Н. bolanderi Воз-
можно, что возраст интрогрессивной долинной расы Н. bolande-
ri составляет всего несколько сотен «лет [366].
Оливьери и Джейн [629] еще раз проанализировали этот слу-
чай, использовав более обширный набор признаков, в том числе
и биохимических, и проведя более сложный статистический ана-
лиз имеющихся данных. Полученные ими гистограммы, как и
следовало ожидать, слегка отличаются от гистограмм Хейзера
(рис. 17.5), но, по-видимому, соответствуют его первоначальной
интерпретации.
Helianthus annuus перекрывается и гибридизирует с други-
ми однолетними видами подсолнечника в других районах США,
например с Н. debilis и Н. argophyllus в Техасе и с Н. petiolaris
на Великих равнинах и на юго-западе.'Я. annuus дал начало
интрогрессивным расам, изменчивость которых направлена в
сторону Н. debilis в Техасе и Н. petiolaris — в центральных райо-
нах США. Н. petiolaris существует также в качестве интрогрес-
сивной расы [364, 368—371, 373].
Гл 17 Интрогрессия
211
Juniperus
Juniperus virginiana широко распространен на востоке США, а
J. scopulorum— на западе Их ареалы в основном аллопатричны
и переходят один в другой с несколькими небольшими участка-
ми контакта на границах (рис 17 6). Фассет [209] сообщает о
скоплениях гибридов, выявленных им на основе изучения морфо-
логических признаков
Сравнительно недавно несколькими исследователями [216]
был проведен анализ популяции этих можжевельников на длин-
ной трансекте от восточной части .Миссури до западной части
Монтаны (рис 17.6) с использованием биохимических призна-
ков— 43 содержащихся в листьях соединений, входящих в со-
став летучих масел. Полученные данные были подвергнуты дис-
криминантному анализу
На основе оценок, полученных в результате такого анализа,
авторы этой работы построили шкалу гибридности, которую
можно было использовать для пострения гистограмм Характер
изменчивости в ряде гистограмм — по одной для каждой попу-
ляции на трансекте от А до Z— показывает, что интрогрессия
происходит от Juniperus scopulorum к J. virginiana в северо-
западной части области, занимаемой этим последним [216]. Че-
тыре такие гистограммы представлены на рис. 17.7.
Pinus
Интересный пример интрогрессии в рамках исторического вре-
мени, основанный на палеоботанических данных, описан Мейсо-
ном [558]. Этот пример относится к двум видам сосны, Pinus
remorata и Р. muricata, произраставшим в прибрежной части
Калифорнии в период от плейстоцена до современной эпохи.
В плейстоцене популяции Pinus remorata на острове Санта-
Крус морфологически сильно отличались от родственной Р. mu-
ricata, которая в то время была распространена на материке
в Калифорнии. В позднем плейстоцене или в постплейстоцеио-
вос время Р. muricata мигрировала на остров Санта-Крус. 1Меж-
ду этими двумя видами происходила естественная гибридизация,
продолжавшаяся и в современную эпоху, что привело к изме-
нению признаков у некоторых популяций Р. remorata. Ископае-
мые данные, собранные в Калифорнии, свидетельствуют о про-
исходившей в плейстоцене интрогрессии в обратном направле-
нии— от Р remorata к Р. muricata-, этот процесс продолжается
и в настоящее время (457].
14*
Рис. 17.6. Географическое распространение Juniperus virginiana и J scopulorum [216].
Буквами показано местонахождение изучавшихся популяций.
Гибридная шкала
Гибридная шкала
Рис. 17.7. Гистограммы гибридных шкал для популяций Juniperus иа трансек-
те, идущей от шт. Миссури до шт Монтана |[216]
Величины для построения гибридной шкалы получены в результате дискри-
минантного анализа 43 летучих масел, содержащихся в листьях и стеблях.
В, С и D — популяции J. virginiana из Миссури и юго-восточной части
Небраски X— популяция J. scopulorum из Моитаны. И и № —две интро-
грессивные популяции из пограничной зоны между этими двумя видами
Положение на карте этих и других популяций на данной трансекте показано
на рис. 17 б.
«. Связность признаков как критерий гибридности
Андерсон [13, 15, 16] рассматривал связность признаков — сла-
бую ассоциацию между разными .признаками в изменчивой по-
пуляции — как диагностический признак гибридности, о чем уже
говорилось выше. Это заключение было основано как на теоре-
тических, так и на эмпирических данных. Связность признаков—
ожидаемый результат многофакторного сцепления в любой рас-
щепляющейся гибридной популяции. Это было подтверждено
экспериментально на искусственных гибридных популяциях.
214 Часть пятая. Естественная гибридизация и ее результаты
Обычно связность признаков наблюдается также в природных
гибридных популяциях.
Заключение Андерсона на протяжении многих лет пользо-
валось широким признанием. Однако недавно было показано,
что такое слишком широкое обобщение неоправданно [297].
Рассмотрим сначала эмпирические данные, относящиеся к
природным гибридным популяциям. Эти данные касаются почти
исключительно многолетних растений, обычно, травянистых.
У таких растений особи одного или обоих родительских расте-
ний сохраняются достаточно долго, чтобы иметь возможность
смешиваться с гибридами Fi и последующими поколениями.
Если они попадают в выборки из популяций, как это часто бы-
вало в ранних исследованиях, то они искажают результаты в
пользу связности признаков.
При повторном анализе данных по нескольким старым при-
мерам— Iris Aquilegia, Oxytropis и другим группам растений,
у которых была описана связность признаков, — в каждой вы-
борке были обнаружены особи родительского типа. После иск-
лючения этих особей из выборки и проведения нового анализа
одной только гибридной субпопуляции связность признаков за-
метно снижалась [297].
Уровень связности признаков можно измерить по коэффи-
циентам корреляции для разных пар признаков. А степень сни-
жения связности признаков в гибридной субпопуляции по срав-
нению с популяцией в целом можно измерить по различию меж-
ду коэффициентами корреляции для одной и той же пары приз-
наков в выборках из той и другой.
Дальнейшими исследованиями в том же направлении было
установлено, что в разных случаях степень снижения'связности
сильно варьирует. Так, например, в одной гибридной популяции
Opuntia при анализе популяции в целом наблюдалась статисти-
чески значимая корреляция между признаками почти во всех
попарных комбинациях, однако в гибридной субпопуляции такая
корреляция была обнаружена лишь для одной пары признаков.
Данные о связности признаков в этой гибридной популяции —
артефакт, обусловленный способом взятия выборки. В отличие
от этого в гибридной популяции Iris уровень статистически вы-
являемой связности признаков хотя и ниже, чем предполагалось
прежде, но все же достаточно высок [297].
Итак, связность признаков — обычная, но не всеобщая чер-
та, характеризующая природные гибридные популяции расте-
ний. Она проявляется, например, у некоторых гибридных попу-
ляций Iris и Aquilegia при правильном взятии выборок, но не
наблюдается у некоторых гибридных популяций Opuntia, Oxyt-
ropis и Gilia [297]. Первоначальное обобщение претендовало на
слишком большую универсальность.
Гл. 17. Интрогрессия
215
Для того чтобы понять это, пам придется вновь вернуться
к теоретическим допущениям, на которых было основано преж-
нее обобщение Некоторые из этих допущений относятся к внеш-
ним факторам, определяющим связность признаков, а другие —
к внутренним.
Самый важный внешний фактор — естественный отбор.
Предполагалось, что отбор в природных гибридных популяциях
всегда будет благоприятствовать генотипам, приближающимся
к родительским типам Это допущение, несомненно, состоятельно
во многих или даже в большинстве случаев. Но нет никаких
причин, по которым можно было бы считать, что оно всегда
справедливо. В тех случаях, когда имеются новые и разнообраз-
ные среды, доступные для заселения, отбор будет благоприятст-
вовать также новым рекомбинантным типам.
Что касается внутренних факторов, то реальной силой сле-
дует считать многофакторное сцепление, и оно способствует
связности признаков; но рекомбинация генов — также реальная
сила, которая действует в противоположном направлении, вызы-
вая рекомбинации признаков. В результате любая данная гиб-
ридная популяция представляет собой некое равновесие между
этими противоположными тенденциями Положение точки рав-
новесия в каждом отдельном случае в значительной мерс зави-
сит от рекомбинационной системы данного растения (см. гл. 1).
В общем следует ожидать, что у растений с высоким основным
числом хромосом связность признаков в гибридных популяциях
будет слабее, чем у растений с низким основным числом; кро-
ме того, у однолетних растений связность слабее, чем у много-
летних
Экологическая сегрегация морфологических
типов'
В гибридных популяциях происходит расщепление не только по
морфологическим и биохимическим признакам, но также и по
физиологическим признакам, определяющим экологические пред-
почтения Однако в отличие от первых двух точно оценить фи-
зиологические признаки трудно и сведения о них публикуются
редко. Тем не менее полевые наблюдения свидетельствуют о том,
что в гибридных популяциях происходит сегрегация по экологи-
ческим предпочтениям.
Интрогрессивная популяция, или скопление гибридов, часто ’
растет в неоднородных или до некоторой степени гибридизиро-
ванных местообитаниях, расположенных между местообитания- ’
ми популяций родительских видов. В таких случаях микрогеог-
рафическое распределение беккроссов разных типов бывает не-
216 Часть пятая. Естественная гибридизация и ее результаты
случайным. Напротив, как и следовало ожидать, особи, прибли-
жающиеся по своим морфологическим признакам к одному из
родительских видов, обычно имеют тенденцию скапливаться в
тех участках гибридного местообитания, которые больше всего
сходны с нормальной для этого вида средой. И наоборот, гиб-
ридные выщепенцы противоположного типа и беккроссы зани-
мают главным образом ниши, приближающиеся к местообита-
ниям другого родительского вида. Я наблюдал такую экологи-
ческую сегрегацию различных морфологических форм в гибрид-
ных популяциях Aquilegia, Mimulus sect. Diplacus, Ipomopsis и
Gilia. Несомненно, другие ботаники наблюдали то же самое и у
других растений. Это явление было описано Бенсоном, Филлип-
сом и Уайлдером [59] в гибридной популяции Quercus.
В этом случае родительскими полувидами были Quercus
douglasii и Q. turbinella subsp. californica. Q. douglasii— листо-
падное дерево с сизо-голубоватыми листьями, растущее в пред-
горьях центральной и северной Калифорнии на противополож-
ной от моря стороне. Quercus turbinella subsp. californica — веч-
нозеленый кустарник, растущий на пустынных и полупустынных
горных склонах в центральной и южной Калифорнии. Эти два
полувида встречаются вместе и гибридизируют в разных местах
в предгорьях южной и центральной Калифорнии. Популяция,
подробно проанализированная Бенсоном, Филлипсом и Уайлде-
ром [59], произрастает в этой зоне контакта, на перевале Техон.
Техонская популяция занимает экологически разнообразную
область, где склоны холмов обращены к разным сторонам све-
та. Жаркие сухие склоны, обращенные на юг и юго-восток, боль-
ше всего соответствуют обычному местообитанию Quercus tur-
binella subsp. californica. Склоны, обращенные на север и севе-
ро-восток, также сухие, но получают меньше солнечных лучей
и приближаются по своим условиям к обычной среде более се-
верного вида Q. douglasii.
Бенсон, Филлипс и Уайлдер [59] провели измерения по 11 ве-
гетативным признакам в выборке из техонской популяции и
оценили их по методу гибридных индексов. При этом оказалось,
что гибридная популяция в целом содержит большое число бек-
кроссов, приближающихся к Quercus douglasii, а также бек-
кроссов, приближающихся к Q. turbinella subsp. californica, при-
чем относительные количества тех и других примерно одинако-
вы. Однако при повторном анализе данных с учетом микрогеог-
рафического местоположения отдельных растений выявилась
другая картина. На склонах, обращенных на север и на северо-
восток, растения морфологически напоминали Q. douglasii.
В отличие от этого на склонах, обращенных на юг и юго-восток,
было много растений, приближавшихся по своим гибридным
индексам к Q. turbinella subsp. californica [59].
Гл. 17. Интрогрессия.
217
Сцепление М— V в связи с интрогрессией
Гены, детерминирующие у высших растений, различные морфо-
логические признаки, в том числе те, которые раньше других
выявляются у цветков, плодов или на поздних стадиях вегета-
тивного развития, обычно сцеплены с генами, воздействующими
на рост и мощность на ранних стадиях развития. Близкие виды
растений часто несут аллели сцепленных друг с другом морфо-
логических генов и генов жизнеспособности. Примером такого
сцепления служит сцепление М—V, которое мы вкратце обсуж-
дали в гл 8 Кроме того, имеются данные о существовании сис-
тем сцепления М—V в перестроенных участках, по которым
различаются хромосомы видов растений, стерильных при скре-
щиваниях, а также в гомологичных участках у рас или видов,
плодовитых при скрещиваниях (см. гл 8).
Сцепление М—V, несомненно, широко распространенное, а
быть может, и универсальное явление в мире растений Здесь мы
обсудим отношение этого широко распространенного типа сцеп-
ления к анализу интрогрессии. Допустим, что мы обнаружили
популяцию растений, характеризующуюся следующими черта-
ми: 1) популяция подвержена изменчивости; 2) растения по
комбинации морфологических признаков приближаются к неко-
ему виду А, но изменчивы в направлении другого вида Б; 3) они
приближаются также к виду А по своим экологическим пред-
почтениям; 4) они плодовиты при скрещивании с видом А, но
их гибриды с видом Б обладают хромосомной стерильностью.
Используя стандартные методы анализа интрогрессии, мы
пришли бы к выводу, что наша гипотетическая популяция пред-
ставляет собой продукт интрогрессии определенных генов от ви-
да Б к виду А Мы постулировали бы имевшую место в прош-
лом гибридизацию АхБ, за которой следовали возвратные скре-
щивания с родительским видом А, сопровождавшиеся отбором
беккроссов, хорошо соответствовавших среде вида А
Интрогрессивная гибридизация, несомненно, будет приво-
дить к описанным выше эффектам Однако следует указать, что
такие же конечные результаты могут быть получены при другом
способе гибридизации Если гены, определяющие морфологиче-
ские признаки, обычно сцеплены с генами жизнеспособности и
если виды растений различаются по аллелям генов, входящих
в эти системы сцепления М—V, то межвидовая гибридизация,
за которой следуют непрерывный инбридинг и отбор инбредных
продуктов, приведет к комбинации признаков, перечисленных в
одном из предыдущих абзацев. Следовательно, такую комбина-
цию признаков нельзя считать абсолютным критерием интро-
грессии [292].
218
Часть пятая. Естественная гибридизация и ее результаты
Я наблюдал в эксперименте с Gilia проявление эффектов, по-
добных тем, которые возникают при интрогрессии, но в отсут-
ствие последней. Родительскими видами служили Gilia modocen-
sis и G. malior. Они представляют собой автогамные тетрапло-
идные виды, принадлежащие к комплексу G. inconspicua, с ко-
торым мы уже познакомились в гл. 11. Здесь пас интересуют
некоторые инбредные линии, происходящие от гибридов Fb по-
лученных при скрещивании G. malior и modocensis, в частности
линии, обозначенные на рис. 17.8 как Ветвь II [288, 292].
Гибриды Fi от скрещивания между Gilia malior и G. modo-
censis высоко, но не полностью стерильны, а конъюгация хромо-
сом у них очень сильно нарушена. Помимо нарушения конъюга-
ции у гибридов, имеются и другие цитогенетические данные,
указывающие на то, что родительские виды различаются по
многочисленным независимым перестройкам в разных участках
[285, 289].
Эти гибриды Fi дали некоторое число потомков без измене-
ния плоидности, которых размножали путем инбридинга на про-
тяжении ряда поколений. Полученные таким образом инбредные
линии подвергли отбору на плодовитость и мощность в среде,
благоприятной для типов, сходных с Gilia modocensis. Морфо-
логические признаки как таковые при отборе не учитывались.
Q. malior G modocensis Ветвь II, F,,
Рис 17 8 Два интерстерильных вида Gilia и один из гибридов между ни-
ми [292]
Этот гибрид был получен в результате инбридинга и отбора па жизнеспо-
собность По морфологии и плодовитости при скрещиваниях он близок к
одному из родительских видов.
Гл. 17. Интрогрессия
219
Растения, получившиеся после отбора в ряде поколений, были
плодовитыми и мощными [288, 289].
Одна группа родственных линий, происходивших от одной и
той же родительской особи из Fa и названная Ветвью II, оказа-
лась морфологически очень близкой к Gilia modocensis
(рис 17 8) В ней, однако, наблюдалась изменчивость в сторону
G. malior
Плодовитые растения из Ветви II использовали для возврат-
ных скрещиваний с обоими родительскими видами. Гибриды, по-
лученные от скрещивания между гибридом из Ветви II и Gilia
modocensis, были плодовиты, и все их хромосомы конъюгирова-
ли с образованием бивалентов, тогда как при возвратном скре-
щивании с G. malior получались высокостерильные гибриды с
нарушенной конъюгацией [288].
Допустим теперь, то растения Ветви II были бы обнаруже-
ны в природе и изучены с морфологической, экологической и ци-
тогенетической точек зрения Очевидно, их определили бы как
отклоняющуюся расу Gilia modocensis, получившую некоторые
гены от G malior в результате интрогрессии и обладающую по-
этому соответствующими этим генам морфологическими призна-
ками. Однако в данном случае нам известна вся родословная
Ветви II, и мы можем утверждать, что никакого возвратного
скрещивания у ее предков не происходило [289, 292].
Таким образом, как из экспериментальных исследований, так
и из теоретических рассуждений следует, что эффекты, не отли-
чимые от эффектов интрогрессии, могут возникать и иным путем
Существующие методы анализа интрогрессии не дают возмож-
ности различать: а) возвратное скрещивание и отбор и б) отбор
во взаимодействии со сцеплением М—V [292].
Учитывая все это, становится необходимым пересмотреть кри-
терии, на основе которых определяют интрогрессию, а также не-
которые примеры, идентифицированные при помощи этих крите-
риев Быть может, наилучшим критерием на данном уровне на-
ших знаний служит система скрещивания растений, о которых
идет речь. У преимущественно автогамных растений размноже-
ние гибрида путем возвратных скрещиваний крайне маловеро-
ятно Относить наблюдаемую у таких растений изменчивость,
сходную с интрогрессией, за счет интрогрессии недопустимо
У некоторых гибридизирующих автогамных растений, в част-
ности у группы Elymus glaucus и группы Aegilops variabilis
[751, 781, 919], изменчивость, сходная с наблюдаемой при интро-
грессии, в самом деле была истолкована как интрогрессия
Однако в свете того, что нам известно о сцеплении М—V, при-
нятые в этих случаях интерпретации следует, возможно, пере-
смотреть
220 Часть пятая. Естественная гибридизация и ее результаты
В отличие от этого в группах растений, размножающихся при
помощи ауткроссинга, наиболее вероятный способ размножения
гибридов — возвратные скрещивания. Это прежде всего следст-
вие самой системы размножения. Кроме того, это обусловлено
соотношением числа гамет, поставляемых в гаметный фонд мно-
гочисленными плодовитыми растениями родительских видов и
редкими стерильными гибридными растениями, соответственно,
о чем уже говорилось выше в этой главе.
Если, обнаружив у свободноопыляющихся растений картину
изменчивости, сходную с интрогрессией, сделать вывод о нали-
чии у них интрогрессии, то такой вывод скорее всего окажется
правильным, соответствуя точке зрения Андерсона [15, 16],
Стеббинса [780] и других авторов. Все примеры интрогрессии,
выбранные в качестве иллюстраций для данной главы (Iris, He-
lianthus, Juniperus и т. п.), относятся к перекрестноопыляющим-
ся растениям.
Следует ожидать, что в группах растений с системой раз-
множения, промежуточной между ауткроссингом и инбридингом,
размножение гибридов происходит разнообразными смешанны-
ми способами. Судя по данным Пази и Цохари [636], так, воз-
можно, обстоит дело в некоторых частях группы Aegilops varia-
bilis.
Трансгрессия барьеров хромосомной
стерильности
Интрогрессия между видами растений, различающимися по
структурным особенностям кариотипов, должна быть связана с
прохождением генов через барьер хромосомной стерильности.
В этом разделе нам предстоит рассмотреть механизм, с по-
мощью которого происходит трансгрессия барьеров хромосом-
ной стерильности. Мы изложим здесь в пересмотренном и рас-
ширенном виде материалы, публиковавшиеся ранее [280, 282].
Начнем с рассмотрения некоторых предварительных усло-
вий. Представим себе некую гибридную популяцию, состоящую
из особей двух видов и гибридов между ними. Растения размно-
жаются при помощи ауткроссинга. Хромосомы родительских ви-
дов различаются по структурным перестройкам, ведущим к сни-
жению конъюгации или несбалансированности гамет у гибри-
дов или же одновременно к тому и другому, в результате чего
у гибридов наблюдается частичная или полная хромосомная
стерильность. Некоторые гены, которые у одного из родитель-
ских видов были локализованы в этих хромосомах, попав в ге-
нотип другого родительского вида, образуют рекомбинанты, бо-
лее адаптированные к среде этого другого вида. Изложенные
Гл. 17. Интрогрессия
221
здесь условия, по всей вероятности, часто встречаются в при-
роде.
Процесс трансгрессии стерильности, происходящий, согласно
ожиданиям, при этих условиях, сложен. Он слагается из несколь-
ких отдельных субпроцессов, действующих одновременно. Во-
первых, растения, составляющие гибридную популяцию, размно-
жаются при помощи ауткроссинга Во-вторых, гомсологичпые
хромосомы структурных гетерозигот образуют биваленты, и по
крайней мере в некоторых из материнских клеток пыльцы и ма-
теринских клеток мегаспор наблюдается кроссинговср Отбор
гамег происходит так, чтобы элиминировать образовавшиеся
при мейозс клетки с нехватками или дупликациями и сохранить
генетически сбалансированные гаметофиты. В то же время есте-
ственный отбор среди членов данной популяции благоприятст-
вует сохранению некоторых межвидовых рекомбинационных ге-
нотипов в среде одного из родительских видов
Мы рассмотрим сначала процесс трансгрессии стерильности
применительно к одной определенной модели В этой модели
имеются вид-реципиент (Я) и вид-донор (£>), вводящий некото-
рые из своих генов в первый вид путем интрогрессии. Рекомби-
национные генотипы, содержащие некоторые гоны вида D на ге-
нетическом фоне вида R, имею, высокую адаптивную ценность
в среде вида R Адаптивная цет юсть таких межвидовых реком-
бинационных типов в этой среде выше, чем ценность комбина-
ции соответствующих аллелей у нсинтрогрессированпого ви-
да R.
Гены, входящие в обладающую адаптивной ценностью ком-
бинацию, находятся в хромосомах, различающихся у этих двух
видов по структурным перестройкам
Для того чтобы упростить обсуждение, мы сосредоточим вни-
мание па одной хромосоме (/), присутствующей у родительских
видов в двух разных структурных формах (7, и Хромосомы
!Г и Ц отличаются по одной транспозиции, показанной на
рис 17 9, А. В процессе филогенетической истории хромосомы
Id один из участков в левом плече поменялся местами с одним
из участков в правом плече, но в хромосоме 1Г эта перестройка
не имела места, в результате чего хромосомы этих двух типов
различаются по своей структуре, как это показано на рис 17.9, А.
Центральные, или интерстициальные, участки хромосом 1Г и
Id гомологичны (рис 17 9, А) Мы постулируем, что гены вида
D, образующие с генами вида R ценную в адаптивном смысле
рекомбинацию, локализованы в этих интерстициальных участ-
ках хромосомы 1а. Гены вида R, входящие в ту же самую цен-
ную комбинацию генов находятся в соответствующих гомоло-
гичных участках хромосомы 1Г или же в этой, а также и в дру-
гих хромосомах данного набора
Вид R
Рис. 17.9. Цитогенетическое поведение пары хромосом, различающихся по
своей структуре, в гибридной популяции.
Родительские виды и D несут и вводят в данную популяцию две частично
гомологичные формы этой хромосомы (обозначенные соответственно 1Г и Id),
различающиеся по одной транспозиции. А. Родительские формы. Б. Продук-
ты мейоза в Fx или у гибридов, полученных при возвратных скрещиваниях,
с кроссинговером между плечами. В. Продукты мейоза в Fi или у гибридов,
полученных при возвратных скрещиваниях, с двойным кроссинговером
между участками, подвергшимися перестройке
Гл. 17. Интрогрессия
223
У гибридов 5x5 конъюгация хромосом / может быть нару-
шена в различной степени вследствие отсутствия полной гомоло-
гии между ними Следует ожидать, что у^биваленгов LL в тех
случаях, когда они образуются, будет довольно регулярно про-
исходить кроссинговер в гомологичных интерстициальных участ-
ках (рис. 17 9,5). При таком кроссинговсре половина клеток,
образующихся в результате мейоза, несет нехвапш и дуплика-
ции по смещенным участкам (рис. 17.9,5), и из них развиваются
нежизнеспособные гаметофиты, что обусловливает полусгсриль-
ность гибридов Иными словами, когда в результате кроссинго-
вера смещенный участок попадает в гомологичную хромосому,
он, создавая нехватку и дупликацию, выступает в роли фактора
стерильности.
Хромосомы с нехватками и дупликациями будут выпадать из
фонда гамет в каждом поколении. То же самое будет происхо-
дить с генами вида D, локализованными в иптерсищиалытых
участках этих хромосом
Функциональными окажутся лишь те гаметы, которые несут
некроссоверные хромосомы 5 или 5 (рис 17 9,5). Следователь-
но, гибрид Fi после скрещивания со своими сибсами дает по-
томков F2, расщепляющихся на два родительских и один гиб-
ридный класс (IrIr> Lid, LL), а при возвратном скрещивании с
видом R происходит расщепление на один родительский я один
гибридный классы (55, 55/) Генотипы 55 полустсрильны в
F2, 5i и 52, так же как и в Fi В отличие от этого средн потом-
ков последующих поколении плодовиты лишь те, которые обла-
дают родительскими генотипами — 55 и 55-
Таким образом, при обычном кроссинговере в хромосоме I
(рис 17.9,5) гибридная популяция расщепляется по морфоло-
гическим и физиологическим признакам, которые определяются
генами, локализованными в эюй хромосоме, на родительские
типы и тип Fj Интро1рсссия генов, лежащих в хромосоме 2, от
вида D к виду R блокирована, так как эти гены сцеплены с фак-
торами стерилыюс1и, находящимися в этой же самой хромо-
соме
Если, однако, кроссинговер в структурной гстерозпготе про-
исходит в точках, отделяющих смещенные участки от гомоло-
гичных интерстициальных участков, то генетические результаты
гибридизации окажутся совершенно иными. Двойной кроссинго-
вер, который приведет к такому отделению, схематически изоб-
ражен на рис 17.9,5. Такого рода кроссинговер может происхо-
дить или не происходить в поколении Fr, если он не произойдет
в Fi, то он может произойти в любой структурной гетерозиготе
в любом последующем поколении от возвратного скрещивания.
В результате такого двойного кроссинговера возникнут крос-
соверные хромосомы типа 5d (рис. 17.9, 5) Эти хромосомы ге-
224 Часть пятая. Естественная гибридизация и ее результаты
нетически сбалансированы, в связи с чем содержащие их гаме-
ты жизнеспособны. По своей структуре они гомологичны хромо-
сомам 1г, поэтому способны образовать фертильные гетерозиго-
ты Jrbd и тем самым включиться в хромосомный фонд вида R
Сбалансированные хромосомы Ьа, несут также в своих цент-
ральных интерстициальных участках обширные блоки генов ви-
да D. Поэтому хромосомы Ird, которые внутренне сбалансиро-
ваны и одновременно гомологичны хромосомам 1Г, могут служить
мостиком для перехода генов вида D к виду R. Вслед за этим
в результате естественного отбора может произойти увеличение
частоты новых хромосом Ьа или их частей в среде вида R.
Мы рассмотрели процесс трансгрессии стерильности на при-
мере упрощенной модели с одной транспозицией. Можно пока-
зать, что этот процесс будет происходить таким же образом при
наличии перестроек других типов, в частности транслокаций и
реинверсий, и соответствующих кроссинговеров. Кроме того, что
весьма примечательно, этот процесс будет действовать при нали-
чии большего числа перестроек в отдельных хромосомах или
плечах хромосом, с тем чтобы обходить как мощные, так и сла-
бые барьеры хромосомной стерильности.
Естественный отбор, благоприятствующий сохранению новой
межвидовой комбинации генов, протаскивает важные гены ви-
да-донора через барьер стерильности, состоящий из хромосом-
ных перестроек, число и типы которых могут быть различны.
Такой отбор может привести к созданию новой расы, сходной
по своим морфологическим и физиологическим признакам с ви-
дом-донором, но в отношении плодовитости принадлежащей к
группе вида-реципиента.
Остается рассмотреть роль генов, локализованных в пере-
строенных участках, в процессе интрогрессии через барьер хро-
мосомной стерильности. Эти гены ограничивают упомянутую
выше конвергенцию.
Те определяющие морфологические признаки гены, которые
локализованы в перестроенных участках вида D, будут элими-
нированы вместе с самими этими участками в результате отбо-
ра гамет в гибридной популяции. Если гены морфологических
признаков вида D сцеплены с генами жизнеспособности этого
вида в тех же участках, то они будут элиминироваться также и
естественным отбором в среде вида R. Сцепления двух типов —
сцепление между морфологическими генами и генами хромосом-
ной стерильности и сцепление генов М—V в перестроенных
участках — препятствуют слишком сильному сближению интро-
грессивной расы с видом-донором по морфологическим призна-
кам. Иными словами, перестроенные участки ограничивают инт-
рогрессию чужих генов и играют роль тормозов в процессе кон-
вергенции морфологических и физиологических признаков.
Гл. 17. Интрогрессия
225
Интрогрессия и расообразование
Во многих группах видов путем интрогрессии образуются отдель-
ные расы Показательным примером служит группа Helianthus
annuus, о которой мы уже говорили.
Расы интрогрессивного происхождения описаны в числе про-
чих у следующих родов Dracophyllum [181], Coprosma [181];
Abies [563] (обзор см [780]); Tradescantia [119]; Cistus [137];
Helianthus [366]; Gilia -[266, 300]; Pinus [334] и Purshia [786].
Во многих случаях расы интрогрессивного происхождения сим-
патричны, по крайней мере частично, с видом- или полувидом-
донором.
Образование рас путем интрогрессии весьма часто происхо-
дит у южнокалифорнийских диплоидных гилий с крупными
цветками, принадлежащих к секции Arachnion. Виды и полуви-
ды этих гилий встречаются симпатрически во многих разных
комбинациях, занимая образовавшиеся недавно аридные и по-
луаридные местообитания. Расы интрогрессивного происхожде-
ния, по-видимому, хорошо адаптируются к некоторым из этих
местообитаний. Судя по морфологическим, экологическим и ци-
тогенетическим данным [300], перечисленные ниже географиче-
ские расы, получившие подвидовые названия, происходят от сле-
дующих родительских форм-
G latiflora davyi G
G leptantha transversa G
G сапа speciosa G
G ochroleuca vivida G
latiflora интрогрессия от G. tenuiflora
leptantha leptantha интрогрессия от
G. latiflora
сапа интрогрессия от G. leptantha pur-
push
ochroleuca bizonata интрогрессия от
G. leptantha leptantha
Интрогрессивная гибридизация — фактор, вызывающий кон-
вергенцию по морфологическим признакам и экологическим
предпочтениям в зоне симпатрического контакта между видами
или полувидами. Другие факторы действуют в противоположном
направлении
При наличии подходящих условий симпатрические контакты
могут также привести к отбору на репродуктивную изоляцию
и (или) смещение признаков, как было отмечено в гл 15. В та-
ких случаях в симпатрической зоне усиливается дивергенция
между видами Симпатрия — эго состояние, вызывающее разно-
образные реакции
Смещение признаков, хорошо известное у животных, у рас-
тений описано лишь в отдельных случаях По мнению некоторых
ботаников, растения в этом отношении не изучались достаточно
15-45
226
Часть пятая. Естественная гибридизация и ее результаты
тщательно. Возможно, что это отчасти справедливо. Мне, одна-
ко, хотелось бы подчеркнуть роль интрогрессии: конвергенция
по морфологическим и физиологическим признакам, обусловлен-
ная интрогрессией, встречается у растений часто и, возможно,
преобладает над смещением признаков во многих или даже в
большинстве случаев.
Выводы
Естественная гибридизация между симпатрическими видами или
полувидами растений происходит довольно часто при наличии
подходящих экологических условий. Имеется много описатель-
ных, но не количественных данных, свидетельствующих о том,
что естественная гибридизация чаще встречается у растений, чем
у высших животных с внутренним оплодотворением.
Гибридизация у растений недостаточно коррелирует с эффек-
тивностью внутренних репродуктивных барьеров между роди-
тельскими видами, вплоть до предельной точки, т. е. абсолют-
ной перекрестной несовместимости. Вместе с тем наблюдается
тесная корреляция между гибридизацией и наличием открытых
местообитаний, в которых могли бы обосноваться гибриды и их
потомки. В современном мире такие открытые местообитания
создаются в широких масштабах в результате деятельности че-
ловека, но они возникали также на протяжении всей геологиче-
ской истории в результате различных природных процессов.
Воспроизведение естественных гибридов в группах растений,
размножающихся при помощи ауткроссинга, обычно происходит
путем возвратных скрещиваний и интрогрессии. Инфильтрация
генов от вида-донора к виду-реципиенту гораздо сильнее ограни-
чена в тех случаях, когда родительские виды различаются по
хромосомным перестройкам, а не несут совершенно гомологич-
ные хромосомы.
В группах автогамных растений гибридизация может при-
вести к конечным результатам, очень сходным с теми, которые
обычно считают указанием на интрогрессию. Альтернативный
путь слагается из инбридинга (а не возвратного скрещивания)
расщепления и отбора.
Интрогрессивная гибридизация играет ключевую роль в фор-
мировании рас у многих преимущественно свободноопыляющих-
ся групп растений. Расы интрогрессивного происхождения об-
наружены в группах видов, принадлежащих к родам Pinus,
Abies, Quercus, Purshia, Cistus, Coprosma, Dracophyllum, Helian-
thus, Gilia, Tradescantia и другим.
18 Сингамеон
Локализованная естественная гибридизация и ограниченный об-
мен генами между полувидами, которые обычно бывают репро-
дуктивно изолированы, приводят к объединению этих полувидов
в более объемлющую (inclusive) скрещивающуюся единицу.
Эта более крупная единица представляет собой сингамеон.
Подобная ситуация была описана Лотси [520, 521] для одной
из групп рода Betula На европейском континенте произрастает
много форм Betula различного и к тому же спорного таксоно-
мического ранга, и все эти формы гибридизируют в широких
масштабах. Лотси [520] пришел к выводу, что «Betula пред-
ставляет собой очень большое скрещивающееся сообщество —
единый сингамеон»
Дю Рие [181] дал обзор многочисленных аналогичных слу-
чаев у Nothofagus на Новой Зеландии, Geum — в Европе, Ме-
landrium — в Швеции, Salix — в северной части Европы и т. п.
Все эти группы, по его мнению, представляют собой «большие
и высокополиморфные «гибрид»-сингамеоны, в которых виды
более или менее утратили свою самостоятельность..» (с. 384).
Позднее рядом исследователей была разработана концепция
сингамеона [53, 134, 279]. В Современных терминах сингамеон
можно определить как «общую сумму видов или полувидов, ко-
торые связаны между собой частой или эпизодической гибриди-
зацией, происходящей в природе; [таким образом] это любая
гибридизирующая группа видов...» [279, с. 67].
Сингамеон — самая «объемлющая» скрещивающаяся едини-
ца в гибридизирующей группе видов. Он может иметь относи-
тельно простую внутреннюю структуру и состоять из двух-трех
полувидов, но иногда он бывает и очень сложным. Образовав-
шиеся в некоторых группах растений сингамеоны представляют
собой обширные сети полувидов, состоящие из многочисленных
таксономических видов и занимающие ареалы, сравнимые по
своей протяженности с целыми континентами.
По внешнему виду и по репродуктивным взаимодействиям с
другими популяционными системами сингамеон сходен с биоло-
гическим видом, т е. он является репродуктивно изолированной
единицей; вместе с тем он отличается от биологического вида
своей более сложной внутренней структурой Следует отметить,
что основные составные части сипгамеонов в формальной систе-
матике рассматриваются как виды, и это совершенно правиль-
но; они обычно действительно представляют собой хорошие так-
сономические виды.
15*
228 Часть пятая. Естественная гибридизация и ее результаты
Сингамеоны существуют во многих родах растений. Выше
уже были перечислены некоторые давно известные примеры из
флоры Европы и Новой Зеландии. В западной части Северной
Америки много сингамеонов обнаружено у Pinus, Juniperus,
Quercus, Mimulus. sect. Diplacus, Iris, Aquilegia, Gilia sect.
Arachnion и у других родов. В предшествующих главах уже бы-
ли описаны некоторые аспекты этих сингамеонов. Здесь мы рас-
смотрим структуру сингамеонов в целом на примере двух групп
растений.
Тихоокеанские ирисы
Самый сложный сингамеон, тщательно изученный одним иссле-
дойателем,— это, вероятно, ирисы, произрастающие на тихооке-
анском побережье Северной Америки |[462].
Ряд Californicae секции Apogon рода Iris представляет собой
естественную группу, состоящую из 11 таксономических видов,
которые распространены от южной Калифорнии до шт, Вашинг-
тон. Их взаимоотношения по плодовитости обсуждались в гл. И.
Между ними не существует сильной внутренней изоляции.
Рис. 18.1. Структура сингамеона у ирисов тихоокеанского побережья [462].
Толстые линии, соединяющие разные виды, означают высокую степень гиб-
ридизации; тонкие линии —умеренную гибридизацию; прерывистые линии —
незначительную гибридизацию. Буквы в кружках — сокращенные названия
разных видов и полувидов рода Iris. BRA — /. bracteaia; CHR — I. chrysophyl-
la; DOU —7. douglasiana; PER —I. fernaldii; HAR —7. hartwegil; INN —
7. innominata; MAC — I. macrosiphon; MUN — I. munzii; PUR —7. purdyi;
TEN — I. tenax; 71AA — I. tenuissima; XTH —7. thompsonii=I. douglasia-
na x 7. innominata.
Гл. 18. Сингамеон
229
Эти виды или полувиды занимают разнообразные экологиче
ские зоны в пределах их общего ареала. Так, Iris douglasiana
встречается вдоль берега, / macrosiphon — в предгорьях и т. д.
Местообитания этих и других полувидов часто переходят одно
в другое, создавая возможность для маргинальных или смежно-
симпатричсских контактов. Кроме того, во многих местах эколо-
гические барьеры разрушаются в результате лесных разработок,
прокладки дорог и других нарушений среды, вызванных деятель-
ностью человека. Как следствие этого между полувидами начи-
нается естественная гибридизация в многочисленных комбина-
циях [461, 462].
Все эти таксономические виды, за исключением / munzii,
связаны между собой в результате происходящей между ними
естественной гибридизации. Иными словами, из 11 таксономи-
ческих видов лишь один — I. munzii—представляет собой изо-
лированный вид, а 10 остальных — это связанные друг с другом
полувиды, образующие сингамеоны, как показано па рис. 18.1
[462].
Некоторые полувиды, как, например, / douglasiana, зани-
мают в сингамеоне центральное положение, будучи связаны со
многими другими полувидами, тогда как другие, подобно / hart-
wegii, являются периферическими и связаны лишь с одним по-
лувидом. Некоторые пары полувидов гибридизируют в очень зна-
чительной степени, другие — в умеренной, а третьи — в очень
слабой (рис. 18 1).
Белые дубы
В обоих подродах дуба — Quercus (белые дубы) и Егу-
throbalanus (черные дубы)—сингамеоны образовались неза-
висимо Между этими двумя сингамеонами существует сильная
репродуктивная изоляция Сингамеон белых дубов наиболее об-
ширен как географически, так и таксономически.
Белые дубы в США изучались многими ботаниками. Лите-
ратура по естественной гибридизации этих д>бов очень велика
и разбросана по разным изданиям Пальмер [633J собрал све-
дения по всем известным для Северной Америки гибридным
комбинациям, однако после выхода его обзора (1948 г.) появи-
лось много новых статей на эту тему, а других общих обзоров
сделано нс было. Я попытаюсь обрисовать здесь в основных
чертах сингамеон белых дубов в США При этом будут приве
лены ссылки на ключевые работы, поскольку дать полный об-
зор литературы не представляется возможным.
Для удобства изложения разделим сингамеон белых дубов
на региональные группы Мы начнем наш обзор с калифорний-
230
Часть пятая. Естественная гибридизация и ее результаты
Рис. 18.2. Структура сиигамеона белых дубов (подрод Quetcus} в Ка-
лифорнии.
ской группы, а затем проследуем через всю страну к восточ-
ным штатам.
Ареалы семи видов белых дубов, встречающихся в Калифор-
нии, перекрываются в различных комбинациях. В некоторых
частях перекрывающихся областей, где экологическая изоляция
нарушена, они гибридизируют. На рис. 18.2 эти полувиды рас-
положены в порядке, примерно соответствующем их географи-
ческому распространению с севера на юг, а соединяющие их
линии отражают степень гибридизации между ними.
Калифорнийская группа этого сингамеона интересна тем,
что она объединяет дубы, резко различающиеся по своим мор-
фологическим и экологическим признакам. Quercus garryana —
лесное дерево с большими рассеченными листьями, распростра-
ненное в северо-западной части тихоокеанского побережья США
Q. dumosa и Q. turbinella —низкорослые кустарники с мелкими
колючими листьями, растущие соответственно в чапаральных
и пустынных фитоценозах. Несмотря на такие резкие различия,
эти формы связаны либо прямо, гибридизируя друг с другом,
либо косвенно, через промежуточные популяции.
В определенных местах на севере Калифорнии лесное дере-
во Q. garryana гибридизирует с формой засушливых предго-
рий, Q. douglasU [780], и с видами кустарниковых зарослей,
Гл. 18 Сингамеон
231
Q. dumosa и Q. durata [837]. На юге Калифорнии Q. douglasii
гибридизирует с чапаральным Q. dumosa, a Q. dumosa — с пус-
тынным Q. turbinella [57, 836]. Эти и другие комбинации пока-
заны на рис. 18.2.
Судя по морфологическим признакам, одной из родительских
форм единственного гибридного дуба, растущего на пустынных
горах в южной Калифорнии, был Quercus lobata [838]. Q. loba-
ia, распространенный в предгорных долинах, в настоящее время
не встречается вблизи того места, где растет гибрид; возможно,
однако, что он встречался там во время одной из плювиальных
фаз плейстоцена. Раньше считалось, что другой родительской
формой был обильный в этой местности Q. turbinella [838]. Прав-
да, основываясь на данных более поздних исследований, можно
думать, что другой родительской формой был новый юго-запад-
ный вид, названный Q. cornelius-mulleri (К. Nixon, личное сооб-
щение; см. также Madrono).
Область распространения сингамеона белых дубов тянется
дальше на юго-запад, от Аризоны и Юты до западной части
Техаса, где он представлен формами, показанными на рис. 18.3.
Эти полувиды расположены на схеме также в соответствии с их
географическим распространением. Гибридизация между пус-
тынной формой Q. turbinella и растущей в Скалистых горах
формой Q. gambelii в настоящее время происходит на севере
Аризоны и происходила в Юте в прошлом, о чем свидетельству-
ют старые изолированные гибридные клоны [125]
^0 gambelii
(о turbinella)
Q ajoensis
Q havardi
Q rugosa
Рис. 18.3. Структура сингамеона белых дубов (подрод Quercus) в юго-
западной части США
232
Часть пятая. Естественная гибридизация и ее результаты
Хотя Близстоящее время Quercus gambelii Скалистых гор и
крупноплодный дуб (Q. macrocar ра) восточной части США ге-
ографически изолированы, в прошлом, вероятно в плейстоцене,
они были связаны гибридизацией, о чем свидетельствует нали-
чие промежуточных признаков у некоторых аберрантных попу-
ляций [573].
На юго-западе Quercus turbinella гибридизирует также с
Q. grisea, который в свою очередь гибридизирует с Q. mohriana,
а этот последний — с Q. havardi [589] (рис. 18.3).
В Техасе Quercus siellata занимает центральное положение
в сингамеоне белых дубов. Здесь он гибридизирует с Q. havardi,
Q. mohriana, Q. virginiana, Q. minima, Q. oleoides, Q. margaretta,
Q. alba, Q. lyrata, Q. macrocarpa, Q. prinus и Q. sinuata. Некото-
рые из этих полувидов гибридизируют также друг с другом, на-
пример Q. virginiana с Q. minima [589, 593].
Кроме того, на востоке США Quercus stellata гибридизирует
с этими же полувидами, а также с другими, например с Q. bico-
lor и Q. prinoides [343, 632].
В восточной части Северной Америки белый дуб Quercus alba
занимает центральное положение в сингамеоне, гибридизируя
* fИ f ♦
° alba Гибриды Q bicolor
♦ WV н
Q. alba Гибриды Q prinus
Рис 18 4. Листья Quercus alba и его двух гибридов [343].
Гл 18. Сингамеон,
233
Рис 18 5 Структура сингамеона белых дубов (подрод Quercus) в во-
сточной части США [343]
здесь с 11 другими полувидами: Q. michauxii, Q. muehlenbergii,
Q. prinoides, Q. lyrata, Q. macrocarpa, Q. stellata, Q. bicolor,
Q. austrina, Q. prinus, Q. margaretta, и Q. robur {343]. Форма
листьев и ее изменчивость у двух из этих гибридных комбина-
ций показаны на рис. 18.4.
Хардин [343] провел прекрасное исследование сингамеона
белых дубов в восточной части Северной Америки, уделив особое
внимание Quercus alba. Многие растущие здесь виды имеют ши-
рокое распространение, причем области их распространения в
значительной степени перекрываются с областями распростране-
ния других видов В зонах симпатрии время от времени проис-
ходит естественная гибридизация. Структура образовавшегося
в результате сингамеона схематически показана на рис. 18.5.
234 Часть пятая. Естественная гибридизация и ее результаты
Как можно видеть, 14 из 16 различных видов — члены этого
сингамеона [343].
Черешчатый дуб (Quercus robur) выращивают в восточной
части Северной Америки, где он спонтанно гибридизирует в не-
скольких местах с Q. alba [343]. На Британских островах, на
своей родине, Q. robur гибридизирует с Q. petraea [126, 767].
Таким образом, сингамеон белых дубов перекидывает мост че-
рез Атлантический океан и продолжается в Европе.
19 Гибридное видообразование
К числу важнейших проблем эволюции растений относится
проблема, связанная с ролью гибридизации в видообразовании,
которую обсуждало не одно поколение исследователей. Служит
ли естественная гибридизация одним из механизмов возникно-
вения новых видов растений?
Некоторые исследователи раннего периода отвечали на этот
вопрос утвердительно. Однако, как мы увидим далее, эти же
самые авторы упускали из виду некоторые аспекты проблемы,
в особенности расщепление в гибридном потомстве, и их утвер-
дительные ответы были поэтому преждевременными и в значи-
тельной мере основывались на интуиции. Тем не менее эти пер-
вые обсуждения поддерживали интерес к проблеме до тех пор,
пока у современных исследователей не появилась возможность
подойти к ней с позиций генетики.
В одной из первых глав было показано, что новые расы воз-
никают иногда в результате естественной гибридизации Это
процесс гибридного образования рас, но необязательно —про-
цесс гибридного видообразования Новая раса, образовавшаяся
в результате процессов, включающих гибридизацию, может в
дальнейшем продолжать дивергировать до видового уровня в
условиях географической изоляции от исходной предковой расы.
Это особый случай географического видообразования, при кото-
ром гибридизация создала необходимую изменчивость на одной
из промежуточных стадий. Дальнейшее обсуждение этого слу-
чая пас здесь не интересует
Под гибридным видообразованием мы имеем в виду возник-
новение нового вида непосредственно из какого-либо естествен-
ного гибрида Такое определение сразу выдвигает на первый
план самую суть проблемы, ясно, что тот самый половой про-
цесс, который привел к образованию естественного гибрида, вы-
зовет также разрушение содержащейся в нем комбинации генов
в результате расщепления в последующих поколениях. Следова-
тельно, существенную часть механизма гибридного видообразо-
вания составляет стабилизация репродуктивного поведения гиб-
ридов
Стабилизация воспроизведения гибридов
Расщепление в Fi или в любом из последующих поколений, воз-
никших в результате межвидового скрещивания, может быть
полностью подавлено или сильно ограничено Известно много
236 Часть пятая. Естественная гибридизация и ее результаты
разных механизмов, с помощью которых это достигается: 1) ве-
гетативное размножение; 2) агамоспермия, 3) перманентная
гетерозиготность по транслокациям; 4) перманентная нечетная
полиплоидия; 5) амфиплоидия; 6) рекомбинационное видообра-
зование; 7) выщепление нового типа, изолированного внешними
барьерами.
Способы 1 и 2, объединяемые под общим названием апомик-
сиса, вкратце описаны в одном из последующих разделов этой
главы и более подробно рассматриваются в части восьмой. Спо-
собы 3 и 4 также рассматриваются в части восьмой на примере
генетических систем группы Oenothera biennis и группы Rosa
canina соответственно. Способ 5, амфиплоидия или аллополи-
плоидия, описан в общих чертах в этой главе, а затем подробно
разбирается в части шестой.
Способы 6 и 7 в отличие от способа 5 связаны с рекомбина-
цией на гомоплоидном уровне. Способ 6, названный рекомбина-
ционным видообразованием, заключается в образовании нового
рекомбинанта, гомозиготного по факторам хромосомной стериль-
ности. Общее представление о нем дается в этой главе, а в
гл. 20 он рассмотрен более подробно. Способ 7 заключается в
образовании нового рекомбинанта, изолированного внешними
барьерами. Мы называем этот способ гибридным видообразова-
нием при наличии внешних барьеров и вкратце рассмотрим его
в одном из последующих разделов данной главы.
Все перечисленные здесь способы создают возможность для
размножения любого данного генотипа гибридного происхожде-
ния. Это может быть генотип либо самого гибрида Fi, либо одно-
го из происходящих от него последующих поколений, и он мо-
жет быть гетерозиготным или гомозиготным.
При способах 6 и 7 рекомбинант гибридного происхождений
достигает состояния, при котором он размножается в чистоте,
благодаря тому что он, по крайней мере на время и по крайней
мере частично, становится гомозиготным. При способах 1—4 ге-
нотип гибридного происхождения размножается в чистоте, со-
храняя свое высокогетерозиготное состояние. Способ 5 представ-*
ляет собой некое компромиссное решение, приводящее к комби-
нации признаков, присущих гомозиготности, с признаками гете-
розиготности. При этом способе форма гибридного происхожде-
ния гомозиготна в том, что касается мейоза, и поэтому
размножается в чистоте, но сохраняет противоположные генные
системы двух родительских видов.
В любом случае конкретная генная комбинация гибридного
происхождения, если она обладает адаптивной ценностью в ка-
кои-либо доступной для нее среде, способна увеличить свою чис-
ленность и образовать собственную популяцию в местности со-
ответствующей этой благоприятной среде. Такая производная
Гл 19 Гибридное видообразование
237
популяция может представлять собой новый вид или микровид
гибридного происхождения
Способ стабилизации воспроизведения гибрида определяет
тип вида, возникающий в результате гибридизации. Вегетатив-
ное размножение и агамоспермия — бесполые способы воспро-
изведения В результате этих способов унипарентального раз-
множения может возникнуть клональный или агамоспермный
микровид. Перманентная структурная или нумерическая гиб-
ридность (способы 3 и 4) также приводят к возникновению уни-
парентальных микровидов В отличие от этого амфиплоидия,
рекомбинационное видообразование и гибридное видообразова-
ние при наличии внешних барьеров представляют собой способы
стабилизации, действующие в условиях полового размножения.
Следовательно, они приводят к образованию новых биологиче-
ских видов
Согласно концепциям, рассмотренным в гл. 7, новые виды,
возникающие от гибридов или от форм гибридного происхожде-
ния,—это во всех случаях эволюционные виды Но лишь в том
случае, если половое размножение и свободное неродственное
скрещивание сохраняются на всем протяжении критических ста-
дий формирования, можно говорить о гибридном происхождении
новых биологических видов. Трудности гибридного видообразо-
вания достигают наивысшей степени в этом последнем случае,
где половое размножение остается в силе. Возникновение нового
биологического вида в результате гибридизации — теоретически
самый сложный, но вместе с тем и самый интересный процесс
История вопроса
В доменделевский период возможность гибридного происхожде-
ния новых видов растений обсуждали Линней в 1744 и 1760 гг,
Уильям Герберт в 1820 г, Шарль Нодэн — в 1863 г. и Антон
Кернер в 1891 г. В начале послеменделевского периода эту ли-
нию рассуждений продолжили Лотси, Хайата и другие. Взгляды
перечисленных исследователей нашли отражение в их работах-
Линней [514], Герберт [376], Нодэн [609], Кернер [431], Лотси
[519] и Хайата [355], обзор см. Дю Рие [181].
Линней и его ученик Рудберг впервые высказали мысль о
гибридном происхождении новых видов растений в 1744 г в свя-
зи с Peloria— одной из форм Linaria, характеризующейся ра-
диально-симметричными цветками В «Disquisitio de Sexu Plan-
tarum» [514] Линней вновь говорит о гибридном видообразова-
нии «Невозможно сомневаться в том, что существуют новые
виды, возникшие в результате гибридизации».
Кернер [431] основывал свои рассуждения на известных фак-
тах о том, что скрещивание порождает новые формы и что во
238 Часть пятая. Естественная гибридизация и ее результаты
многих группах растений встречаются естественные межвидо-
вые гибриды; из этого он делал вывод, что «гибриды могут да-
вать начало новым видам». Конечно, в природе образуется мно-
го гибридов, но лишь немногим из них удается стать новыми
видами. По мнению Кернера, важным ограничивающим факто-
ром в данном случае является среда.
Только в тех местах, где имеется подходящее и открытое
местообитание, не занятое родительскими видами, гибридные
растения могут закрепиться и увеличить свою численность.
Однако такие открытые местообитания встречаются в природе
повсюду, так же, как и виды гибридного происхождения. В ка-
честве примеров Кернер приводит Rhododendron, Salvia и Nu-
phar в Европе.
Так, Rhododendron intermedium представляет собой продукт
гибридизации между R. ferrugineum и R. hirsutum. В некоторых
местах в Альпах /?. intermedium изредка встречается вместе с
родительскими видами, но в других местах он образует обшир-
ные собственные популяции и значительно превосходит по чис-
ленности родительские типы. В таких местах R. intermedium
достигает статуса хорошего вида [431].
Линней и Кернер были выдающимися ранними сторонника-
ми идеи о том, что естественные гибриды могут служить отправ-
ными точками для возникновения новых видов. Анализ их рас-
суждений показывает, что оба они не уделяли должного внима-
ния двум важным аспектам этой проблемы, без учета которых
невозможно создать удовлетворительную гипотезу гибридного
видообразования, а именно — стерильности и расщеплению
Линнею и Кернеру не было известно явление расщепления.
Кернер, собственно, постулирует, что «гибриды передают свою
форму потомкам в неизменном виде... при условии исключения
пыльцы от других видов». Линней проводил мало эксперимен-
тальных исследований по гибридизации растений, а эксперимен-
тальные исследования Кернера касались других аспектов раз-
множения растений. Поэтому ни тот, ни другой не могли себе
представить, что при переходе от естественного гибрида к дочер-
нему виду в случае плодовитых межвидовых скрещиваний воз-
никают какие-то проблемы, связанные с передачей наследствен-
ных признаков.
^Список первых авторов, занимавшихся гибридизацией расте-
ний, которые знали о существовании явления расщепления, но>
не обсуждали формирование видов в результате гибридизации,
достаточно велик: от Кёльрейтера |[444] до Менделя, Дарвина и
Фокке [217].
Большой интерес представляет статья Нодэна [609], посколь-
ку в ней обсуждается и расщепление, и гибридное видообразо-
вание. Нодэн рассматривал первое как установленный факт, а
Гл 19. Гибридное видообразование
239
второе — как некую возможность Он понимал, что явление рас-
щепления создает трудности для теории гибридного видообразо-
вания Тем не менее он допускал возможность исключительных
случаев, в которых гибридные признаки закреплялись в после-
дующих поколениях, и считал, что такие случаи могут привести
к возникновению новых видов {609].
Осложнение, создаваемое стерильностью, в ранний период
изучения гибридного видообразования также отбрасывалось
Было известно, что некоторые пары видов при скрещиваниях
дают стерильных гибридов, а другие — плодовитых; поэтому
считалось, чго гибридное видообразование возможно лишь в тех
случаях, когда гибриды плодовиты [431]. Таким образом, вни-
мание было сосредоточено только на одном особом аспекте всей
проблемы, а проблема гибридного видообразования в целом ка-
залась проще, чем она есть на самом деле.
Создание теории современной генетики в первые десятиле-
тия нашего века дало, наконец, возможность правильно поста-
вить старую проблему гибридного видообразования, учитывая
при этом не только морфологические и физиологические призна-
ки, но также барьеры, создаваемые стерильностью гибридов, и
расщепление. Для того чтобы решить эту проблему, необходимо
было найти некий механизм, с помощью которого из межвидо-
вого гибрида мог бы возникнуть новый, внутренне изолирован-
ный константный вид без утраты способности к половому раз-
множению
Первым крупным достижением в этом направлении была ги-
потеза Винге [898, 899], касающаяся амфиплоидии (сам этот
термин появился гораздо позднее). Согласно этой гипотезе, но-
вый константный вид может возникнуть от гибрида между дву-
мя существовавшими ранее видами в результате удвоения у
него числа хромосом. Гипотеза Винге была вскоре подтверждена
экспериментальными данными, полученными другими исследо-
вателями для Nicotiana, Raphanus-Btassica, Galeopsis и других
растений [105, 417, 596, 597].
Амфиплоидия —это способ видообразования, связанный с
однонаправленным увеличением числа хромосомных наборов,
тогда как эволюция многих видов растений происходила, оче-
видно, на диплоидном уровне, и поэтому оставался неясным во-
прос о том, возможно ли гибридное видообразование без изме-
нения плоидности
Сам Винге вернулся к этому вопросу в своих более поздних
работах, проводившихся па Tragopogon и Erophila [900, 901], а
Лампрехт [458] — в работе на Phaseolus Эксперименты на Tra-
gopogon, Erophila и Phaseolus проводились со специальной
целью: установить, возможно ли возникновение новых плодови-
тых видов или микровидов в результате гибридизации, не со-
240 Часть пятая. Естественная гибридизация и ее результаты
провождающейся амфиплоидным удвоением. Эти эксперименты
проложили путь другим ученым, исследовавшим иной, отличный
от амфиплоидии процесс гибридного видообразования, извест-
ный теперь под названием рекомбинационного видообразования.
Введение в амфиплоидию
Хромосомная стерильность создается в результате структурных
перестроек между родительскими геномами, что приводит к на-
рушениям, видимым или невидимым, течения мейоза у гибрида:
гомеологичные хромосомы либо не конъюгируют нормально с
образованием бивалентов и не расходятся, как положено, к по-
люсам в анафазе, либо конъюгируют, но расщепляются, образуя
при этом дочерние ядра, несущие нехватки и дупликации. В лю-
бом случае какая-то часть продуктов мейоза оказывается несба-
лансированной и не дает полноценные споры и гаметы Эта
часть нежизнеспособных спор, а тем самым и степень стериль-
ности, быстро возрастает с увеличением числа гетерозиготных
перестроек (см. гл. 8 и 9).
Допустим, что у гибридов, обладающих хромосомной стериль-
ностью, произошло удвоение числа хромосом. В таком случае,
если отвлечься на время от других возможных осложняющих
факторов, мейоз у этих гибридов будет протекать нормально, с
образованием фертильных гамет.
Такое восстановление фертильности на новом полиплоидном
уровне наиболее ясно выражено в первом из указанных выше
случаев, в котором хромосомы диплоидного гибрида не обра-
зуют нормальных бивалентов Обозначим хорошо различающие-
ся по своей структуре геномы родительских видов буквами А и
В. Два диплоидных вида имеют при этом конституцию АА и
ВВ, а гибрид между ними — конституцию АВ. У хромосом, при-
надлежащих к набору А, нет гомологичных партнеров, с кото-
рыми они могли бы конъюгировать при мейозе у гибрида, нет
гомологичных партнеров и у хромосом из набора В Однако у
образующегося при этом аллотетраплоидного гибрида (или ам-
фидиплоида) с геномом ААВВ положение совершенно иное.
У него имеется по гомологичному партнеру для каждой хромо-
сомы каждого генома, поэтому мейоз и образование гамет могут
протекать нормально.
Во втором из приведенных выше случаев гомеологичные хро-
мосомы гибрида образуют биваленты, но расщепляются и дают
генетически несбалансированные продукты мейоза. Диплоидные
родительские виды содержат различные субгеномы—Л3 и Af.
Происходящий от них гибрид Fi имеет конституцию AsAt и ха-
рактеризуется хромосомной стерильностью при явно нормаль-
ном мейозе. В этом случае также у тетраплоидного производного
Гл. 19. Гибридное видообразование
241
плодовитость по всей вероятности, восстановится, по
крайней мере частично, в результате того, что Дарлингтон [138J
назвал дифференциальным сродством (differential affinity).
Каждая Д8-хромосома и каждая Л-хромосома у этого тстра-
плоида имеет полностью гомологичного партнера Такие гомо-
логичные хромосомы предпочтительно конъюгируют друг с дру-
гом и расходятся к противоположным полюсам, образуя струк-
турно и генетически сбалансированные продукты мейоза.
Тетраплоидная форма, происходящая от гибрида, обладаю-
щего хромосомной стерильностью, независимо от того, имеет ли.
она геном ААВВ или AsA3AtAtt не только плодовита, но и ре-
продуктивно изолирована от своих диплоидных родительских
видов Скрещиванию между тетраплоидными и диплоидными
растениями нередко препятствуют барьеры несовместимости, и
гибриды, если они образуются, представляют собой триплоиды,
а поэтому обычно стерильны.
Классические примеры Primula kewensis, Raphanobrassica и
Galeopsis tetrahit иллюстрируют различного типа геномы, встре-
чающиеся у экспериментальных амфиплоидов
Гибрид Fi от скрещивания между Primula floribunda
(2п=18) и Р. verticillata (2п=18), широко известный под назва-
нием Р kewensis, — диплоидное многолетнее травянистое расте-
ние, подобно своим родительским видам У этого гибрида про-
исходила конъюгация с образованием нормальных бивалентов^
но он был высокостерилен. В три разных года это в остальном
стерильное гибридное растение спонтанно выбрасывало фер-
тильные побеги, которые, как показало цитологическое иссле-
дование, представляют собой тетраплоиды (2п = 36) и при конъ-
югации хромосом у них образуются преимущественно бива-
ленты.
Очевидно, у этих тетраплоидных побегов предпочтительно
конъюгировали полностью гомологичные хромосомы Эти фер-
тильные побеги возникали в результате удвоения хромосом в
сомашчсских клетках почки или какой-либо части почки. Они
давали семена, из которых развивались плодовитые и вполне
однородные потомки F2 [612 849]
Противоположная ситуация была обнаружена у Raphano-
brassica— амфиплоида, полученного в результате скрещивания
Raphanus sativus и Brassica oleracea. Оба родительских вида —
диплоиды с 2/г = 18 хромосом. У гибридов Fj конъюгация хромо-
сом полностью нарушена, и они высоко стерильны Эти гибри-
ды дали некоторое количество нередуцировапных диплоидных
гамет От слияния таких гамет были получены теграплоидные
(2п = 36) растения F2, у которых мейоз протекал без нарушений
с образованием нормальных бивалентов и которые по большей
части были вполне плодовитыми, давая морфологически одно-
16—45
242 Часть пятая. Естественная гибридизация и ее результаты
родное поколение F3. Эта новая плодовитая размножающаяся в
чистоте линия изолирована барьером стерильности от диплоид-
ных родительских видов [417].
Galeopsis tetrahit (2п=32) в отличие от Primula kewensis и
Raphanobrassica представляет собой тетраплоидный вид, встре-
чающийся в природных условиях в северной части Европы и в
Азии. Мюнтцинг [596, 597] показал, что это однолетнее травя-
нистое растение является амфиплоидом, возникшим в Европе
от других родственных диплоидных видов: G. pubescens (2п=16)
и G. speciosa (2п=16).
У гибридов Fi, полученных в результате искусственного скре-
щивания между Galeopsis pubescens и G. speciosa, мейоз нару-
шен, и они высоко стерильны; фертильная пыльца составляет у
них всего 8%, а число образующихся бивалентов — от 5 до 8.
От гибридов Fi, несмотря на их высокую стерильность, было
получено поколение Fo, состоявшее из нескольких стерильных
диплоидов и одного стерильного триплоида. Последний возник
в результате слияния одной нередуцированной и одной редуци-
рованной гаметы Fj. Этот триплоид при возвратном скрещива-
нии с G. pubescens образовал одно семя, из которого в F3 раз-
вилось тетраплоидное растение. Вероятно, оно возникло в ре-
зультате оплодотворения нередуцированной Зп-яйцеклетки нор-
мальным lft-спермием. У этого тетраплоидного растения F3 в
метафазе мейоза образовалось 16 бивалентов, а фертильная
пыльца составила 70%. Оно дало плодовитое тетраплоидное по-
томство F4 [596, 597].
Этот искусственный аллотетр а плоид сходен с возникшим в
природе Galeopsis tetrahit как по морфологическим признакам,
так и по числу хромосом. Кроме того, подобно дикорастущему
G. tetrahit, он изолирован от G. pubescens и G. speciosa барь-
ером несовместимости. Его можно было бы рассматривать как
синтетическую форму G. tetrahit. Чтобы проверить это предполо.-
жение, Мюнтцинг скрестил искусственный тетраплоид с природ-
ным G. tetrahit. Скрещивание прошло без затруднений, а расте-
ния Fi были плодовиты, и конъюгация хромосом протекала у
них нормально. Это был первый пример экспериментального
создания существующего в природе амфиплоидного вида [596,
597].
Введение в рекомбинационное видообразование
Рекомбинационное видообразование — термин, принятый для
обозначения одной из наименее известных форм гибридного
видообразования (291]. Этот процесс происходит в рамках систе-
мы полового воспроизведения и ведет к образованию дочернего
вида, изолированного от родительских видов, как и при амфи-
Гл 19. Гибридное видообразование 243>
плоидии, барьером хромосомной стерильности. Однако в этом
случае в отличие от амфиплоидии дочерний вид остается на том
же уровне плоидности, что и родительские формы.
Такой результат достигается благодаря возникновению и за-
креплению в последующих поколениях гибридов новой рекомби-
национной формы, гомозиготной по разным независимым хро-
мосомным перестройкам, по которым различались родительские
виды Такая новая гомозиготная рекомбинационная форма пло-
довита при скрещиваниях внутри себя, по стерильна при скре-
щиваниях с любым из родительских видов.
Процесс рекомбинационного видообразования может привес-
ти к созданию дочерних видов, окруженных барьерами стериль-
ности, различающимися по своей эффективности. Степень изо-
ляции дочернего вида прямо пропорциональна степени стериль-
ности при скрещиваниях между родительскими видами.
Основную идею, на которой построена гипотеза рекомбина-
ционного видообразования, высказал Мюнтцинг [595, 598, 601].
Согласно предложенной им концепции, отдельные пары факто-
ров, обусловливающих хромосомную стерильность, могут реком-
бинироваться, в результате чего образуется новая плодовитая
форма; однако дальнейшей разработкой этой концепции Мюнт-
ципг не занимался. Герасимова [237] подтвердила правильность
концепции Мюнтцинга в экспериментальных работах по трансло-
кациям у Crepis. Стеббинс [776, 777, 780] рассматривал в общем
плане рекомбинацию факторов стерильности в связи с гибрид-
ным видообразованием. Позднее Стеббинс [782] и Грант [280]
предложили конкретную генетическую модель рекомбинацион-
ного видообразования. Еще позднее, в 1963 г, я опубликовал
обзор по этой проблеме, собрав все имевшиеся к тому времени
фактические данные [282].
Итак, мы вкратце ознакомили читателя с рекомбинационным
видообразованием, с тем чтобы сопоставить этот способ с дру-
гими способами гибридного видообразования В следующей гла-
ве оно будет рассмотрено более подробно
Отметим мимоходом, что процесс рекомбинации способен
создавать новые барьеры генной стерильности из пред существо-
вавших барьеров совершенно с тем же успехом, с каким он соз-
дает барьеры хромосомной стерильности. Имеются некоторые
данные, указывающие на образование путем рекомбинации но-
вых барьеров генной стерильности у Phaseolus и новых контро-
лируемых генами барьеров несовместимости у Gilia [273, 458].
Об этой стороне проблемы известно, однако, слишком мало, что-
бы она заслуживала специального обсуждения в этой книге.
В общем этот процесс в значительной мере сходен с процессом,
в котором участвуют факторы хромосомной стерильности и ко-
торый будет описан в дальнейшем.
16*
244 Часть пятая. Естественная гибридизация и ее результаты
Гибридное видообразование при наличии
внешних барьеров
Гибридизация между двумя плодовитыми при скрещивании
видами растений может привести к образованию разнообразных
рекомбинантов с разными комбинациями морфологических и
физиологических признаков, по которым различались родитель-
ские формы. Некоторые из этих новых комбинаций признаков
могут обусловить создание внешней изоляции того или иного
типа между гибридным потомством и родительскими видами.
Если такие рекомбинанты, изолированные внешними барьерами,
хорошо адаптированы в какой-либо доступной им среде, то они
могут затем увеличивать свою численность на территории, зани-
маемой родительскими видами.
В одном из первых разделов этой главы мы уже приводили
исторический пример возможного гибридного видообразования
у Rhododendron, изучавшийся Кернером [431]. Кернер постули-
ровал, что R. intermedium произошел от гибрида между R. fer-
rugineum и R. hirsutum и сумел обосноваться в некоторых мес-
тах в Альпах. Этот дочерний вид гибридного происхождения
отличается от родительских видов по окраске цветков и до не-
которой степени — по предпочитаемым им почвам. На основе
того, как излагает этот пример Кернер, следует считать, что
R. intermedium частично изолирован от R. ferrugineum и R. hir-
sutum поведением опыляющих эти растения пчел, а, возможно,
также и эдафическими факторами. Более новое описание гибри-
да R. intermedium и его родительских видов дал Хеги [359].
Delphinium recurvatum, D. hesperium и D. gypsophilum со-
ставляют группу плодовитых при скрещивании друг с другом
диплоидных видов, распространенных в предгорьях и долинах
Калифорнии. Судя по морфологическим и экологическим приз-
накам, D. gypsophilum возник в результате выщепления из
скопления гибридов от скрещивания между D. recurvatum и
D. hesperium. По экологическим предпочтениям и морфологиче-
ским признакам D. gypsophilum занимает промежуточное поло-
жение между этими предполагаемыми родительскими видами.
Среди потомков F2, полученных при искусственном скрещива-
нии между D. recurvatum и D. hesperium, некоторые экземпля-
ры морфологически сходны с D. gypsophilum. В природе D. gyp-
sophilum сохраняет обособленность от двух других видов глав-
ным образом благодаря экологической изоляции [495].
В Калифорнии обитают три родственных диплоидных
(2п=16) вида Penstemon (Р. centranthifolius, Р. grinnellii и
Р. spectabilis), до некоторой степени плодовитые при скрещива-
ниях друг с другом. Они различаются по строению цветков, спо-
Гл 19 Гибридное видообразование
245
собам опыления и экологическим особенностям Сопоставление
морфологических признаков этих трех видов, данные экологи-
ческих исследований в природных условиях и признаки естест-
венных гибридов между ними позволяют считать, что Р. specta-
bilis возник в результате гибридизации между Р. centrantijolius
и Р grinnellii Нельзя, однако, исключить возможность того, что
вид Р. spectabilis возник в процессе первичного видообразова-
ния. В природе эти три вида изолированы друг ог друга меха-
ническими, этологическими и экологическими барьерами [808,
809J.
Стеббинс и Ферлан [791] постулировали гибридное проис-
хождение Ophrys murbeckii от скрещивания между О. fusca и
О. lutea в Средиземноморской области. В этом случае обособ-
ленность предполагаемых дочернего и родительских видов под-
держивается главным образом этологическими и экологическими
изолирующими факторами [791].
Такер и Сауэр [839] описали ряд диплоидных популяций
Amaranthus в дельте р. Сакраменто в Калифорнии, где в настоя-
щее время, возможно, происходит гибридное образование
диплоидных видов. В этом регионе обнаружено несколько хоро-
шо выраженных видов (Л. powellii, A. caudatus, A retroflexus)
и одновременно несколько отклоняющихся локальных популя-
ций, у которых наблюдаются в разных комбинациях признаки
названных выше видов. Эти отклоняющиеся популяции или не-
которые из них, по-видимому, представляют собой различные
диплоидные гибридные производные уже существующих видов
Amaranthus [839].
На Аляске Carex rostrata обычно встречается в пресноводных
озерах, а С rotundata — на торфяных болотах. В переходных зо-
нах между этими двумя местообитаниями обнаружены скопле-
ния гибридов, образовавшихся в результате скрещиваний меж-
ду названными выше видами В некоторых местах из скоплений
гибридов выщепляется новая, морфологически отличающаяся
плодовитая популяция, и численность ее по сравнению с числен-
ностью родительских видов возрастает Эта новая форма рас-
сматривается как дочерний вид гибридного происхождения и
получила название С. paludtvagans [179]
Новая Зеландия — один из центров видообразования для ро-
да Epilobium с 37 местными видами, занимающими широкий
спектр местообитаний Это многолетние травы, для которых ха-
рактерны вегетативное размножение, мелкие цветки и преиму-
щественно автогамная система размножения Epilobium — гомо-
плоиды с 2п = 36 Продукты скрещивания между разными вида-
ми плодовиты в большинстве комбинаций Симпатрия представ-
ляет собой обычное явление; при этом репродуктивная изоляция
246 Часть пятая. Естественная гибридизация и ее результаты
обеспечивается автогамией, а также экологической дифферен-
циацией [666].
Однако иногда репродуктивные барьеры разрушаются, так
что гибриды и скопления гибридов хорошо известны. Обнаруже-
но также несколько однородных популяций гибридного проис-
хождения. Некоторые из них находятся на расовом, а другие —
на видовом уровне. Так, предполагается, что Epilobium purpura-
tum на острове Южный — результат скрещивания между Е. eras-
sum и Е. porphyrium. Трудно сказать что-либо определенное от-
носительно этого и некоторых других случаев, но в общем пред-
ставляется вероятным, что разнообразие видов Epilobium на
Новой Зеландии увеличилось путем гибридного видообразова-
ния [666].
Недавно был описан интересный пример гибридного видооб-
разования у древовидных папоротников (Cyatheaceae) в Пуэрто-
Рико [118]. Виды Alsophila bryophila, A. dryopteroides и Nephe-
lea portoricensis (у всех 2n«69 II) гибридизируют во всех воз-
можных комбинациях. Образующиеся при этом гибриды плодо-
виты и дают в F2 гомоплоидных выщепенцев, часть которых со-
храняется в природе.
Одно из гомоплоидных гибридных производных от скрещива-
ния между Alsophila dryopteroides и Nephelea portoricensis обра-
зует однородную популяцию, превосходящую по численности по-
пуляции родительских видов в той же местности. Эта гибридная
популяция размножается половым путем. Никаких данных о ве-
гетативном размножении нет [118].
Размножение в чистоте у этой формы гибридного происхож-
дения обеспечивается, по-видимому, следующей системой вос-
произведения. Гибридный папоротник Fi производит рекомби-
нантные споры, из которых развиваются обоеполые гаметофиты.
Мужские и женские гаметы развиваются на гаметофите в ре-
зультате митотических делений, а поэтому они генетически иден-
тичны. В результате самооплодотворения внутри гаметофита об-
разуется гомозиготный спорофит, несущий такую же генную ком-
бинацию, как и та рекомбинантная спора, из которой этот гаме-
тофит развился. Такой спорофит дает начало спорам, из кото-
рых развиваются новые гаметофиты, так что в конце концов
возникает колония идентичных индивидуумов данного нового
типа ,[И8]-
Изолированность такой популяции гибридного происхожде-
ния обеспечивается внешними экологическими барьерами и са-
мооплодотворением (118].
Гл. 19. Гибридное видообразование
247
Введение в апомиктическое видообразование
Иногда оказывается, что один из межвидовых гибридов в F]
или в каком-либо из последующих ранних поколений обладает
исключительной адаптивной ценностью Его высокогетерозигот-
ный генотип в таком случае может быть наделен гетерозисным
эффектом или гомеостазом или одновременно и тем и другим.
Подобный гибрид потенциально заслуживает воспроизведения
и увеличения численности Но размножение межвидового гибри-
да, превосходящего по своей адаптивной ценности родительские
виды, сопряжено с двумя очевидными затруднениями. Первое из
них — гибридная стерильность, второе — неспособность размно-
жаться в чистоте. Существует, однако, несколько способов, поз-
воляющих обойти эти затруднения, на что уже давно указал
Дарлингтон [138, гл 16].
Любая гетсрозигота, произошедшая в результате межвидо-
вого скрещивания, может преодолеть внутренне присущие ей
стерильность и расщепление, затрудняющие воспроизведение, и
обеспечить размножение своего особого гетерозиготного геноти-
па при помощи нескольких способов стабилизации, перечислен-
ных в начале этой главы (способы 1—5). Здесь в связи с гиб-
ридным видообразованием мы упомянем о двух первых спосо-
бах, объединяемых под общим названием апомиксис В этом
разделе дается лишь общее предс!явление об апомиктическом
видообразовании, а более подробно оно будет рассматриваться
в части восьмой
Opuntia fulgida и О spinosior — два различных вида какту-
сов, растущих в пустынях на юго-западе США и в Мексике. Они
время от времени скрещиваются, давая гибриды с высоко сте-
рильными семенами. Несмотря на стерильность семян, гибрид-
ные растения достигли большого обилия вдоль реки Хила в
Аризоне благодаря своей способности к вегетативному размно-
жению опавшими члениками стебля [304].
Европейские ежевики, принадлежащие к секции Moriferus
рода Rubus, образуют большой комплекс гибридизирующих ви-
дов, размножающихся половым путем, и их апомиктических
производных. Их гибриды размножаются как путем агамоспер-
мии, так и вегетативно. Гибриды каждого типа могут размно-
жаться и распространяться при помощи семян, образующихся
бесполым путем, или в результате укоренения опавших кусочков
стебля, многие микровиды возникли именно такими способами
[315].
Гибрид, обладающий хорошо выраженными морфологически-
ми признаками и экологическими предпочтениями, может таким
образом распространиться апомиктически и создать свою соб-
248 Часть пятая. Естественная гибридизация и ее результаты
ственную популяцию. Такая популяция, состоящая из одинако-
вых гибридных индивидуумов апомиктического происхождения,
если она недостаточно хорошо обособлена морфологически или
узко эндемична, или же характеризуется одновременно и тем и
другим, представляет собой клональный или агамоспермный.
вид. Если же гибридные производные обладают комбинацией
признаков, легко распознаваемой в обычной таксономической
практике, и если этой комбинации признаков удалось распрост-
раниться путем апомиксиса по определенной географической об-
ласти, то образовавшаяся в результате популяция достигает ста-
туса таксономического вида.
Выводы
Полезно кратко охарактеризовать известные способы гибридно-
го видообразования у растений.
I. Гибридное видообразование в сочетании с половым вос-
произведением; новый биологический вид возникает непосредст-
венно из межвидового гибрида.
1. Гибридное видообразование при наличии внешних
барьеров; формирование нового рекомбинационного типа
в потомстве гибрида, отделенного от родительских видов
внешними изолирующими механизмами.
2. Рекомбинационное видообразование; гибрид, характе-
ризующийся хромосомной стерильностью или полусте-
рильностью, дает начало гомоплоидным или неудвоившим-
ся производным, обладающим новой гомозиготной реком-
бинацией различных независимых хромосомных пере-
строек, по которым различаются родительские виды; но-
вая плодовитая линия характеризуется собственной ком-
бинацией признаков и стерильна при скрещиваниях с ро-
дительскими видами.
3. Амфиплоидия; гибрид, обладающий хромосомной сте-
рильностью, путем удвоения числа хромосомных наборов
.дает начало плодовитой полиплоидной форме, которая
изолирована от родительских видов барьером, создавае-
мым хромосомной стерильностью.
II. Гибридное видообразование в сочетании с бесполым или
почти бесполым размножением; размножение гибрида при помо-
щи механизмов, обходящих нормальный половой или хромосом-
ный цикл, и распространение возникшей гибридной формы по
определенной географической области, так что по величине и
составу популяции она достигает уровня унипареитальногоГ мик±
ровида илисдида. ..... ' . - - >
Гл. 19. Гибридное видообразование 249
4. Апомиктическое видообразование; новый клональный
или агамоспермный микровид развивается соответственно
из межвидового гибрида путем вегетативного размноже-
ния или агамоспермного образования семян (часть вось-
мая).
5. Перманентная структурная или нумерическая гибрид-
ность; какая-либо гетерозигота по транслокации размно-
жается по генетической системе типа Oenothera biennis
или численность какого-либо нечетного полиплоида уве-
личивается по генетической системе Rosa canina, в ре-
зультате чего образуется популяция, соответствующая
по величине гетерогамному микровиду (часть восьмая).
20 Рекомбинационное
видообразование
Рекомбинационное видообразование — это формирование и за-
крепление в потомстве гибрида, характеризующегося хромосом-
ной стерильностью или полустерильностью, нового гомозиготно-
го рекомбинанта по двум или нескольким независимым пере-
стройкам, которыми различаются родительские виды.
В результате процесса рекомбинационного видообразования
могут возникнуть дочерние виды, изолированные слабыми или
сильными барьерами стерильности. Дать объяснение гибридному
происхождению новой гомоплоидной линии, которая сама пло-
довита, но высокостерильна при скрещиваниях с другими пред-
ставителями своего рода, несомненно, труднее. Однако, если мы
сможем найти удовлетворительное объяснение этому случаю, у
нас окажется все необходимое, для того чтобы понять теоре-
тически более простые случаи, в которых функционируют слабые
барьеры стерильности.
В этой главе мы опишем гипотетический механизм рекомби-
национного видообразования, дадим обзор соответствующих экс-
периментальных данных и попытаемся понять возможную роль
этого процесса в природе.
Гипотеза
Довольно четкую модель, описывающую интересный с теорети-
ческой точки зрения случай рекомбинационного видообразова-
ния с возникновением сильных барьеров стерильности, предло-
жили Стеббинс [782] и Грант [280]. Эта модель предусматривает
возникновение барьера хромосомной стерильности между двумя
гибридизирующими родительскими видами в результате незави-
симых друг от друга структурных перестроек. Модель основана
на более ранних работах Мюнтцинга и других авторов, о чем го-
ворилось при вводном обсуждении этой проблемы в гл. 19.
Допустим, что два родительских вида изолированы друг от
друга барьером хромосомной стерильности, создаваемым двумя
или несколькими независимыми хромосомными перестройками.
Гибрид между этими двумя видами, обладающий хромосомной
стерильностью, хотя и неполной, может дать начало одному или
большему числу новых гомозиготных рекомбинантов по струк-
турным перестройкам. Сами эти новые формы плодовиты, но
проявляют хромосомную стерильность при скрещивании друг
с другом и с родительскими видами [280, 282, 780, 782].
Такой процесс рекомбинации можно проиллюстрировать на
простом примере. Допустим, что два родительских вида разли-
Гл. 20. Рекомбинационное видообразование
251
Гаметы F,
Несбал ансиро ван н ые
ABCD.
ABCdD
ABaCD
AB.CD.
ABaCdD
ABaCdDe
AbBCD
AbBCDe
AbBCdD
AbBCdD.
AbBe CD.
AbBaCdD
Сбалансированные
ABCD
ABCdD.
AbBBCD
AbBoCdD.
Гомозиготы F2
AA BB CC DD
A A BB CdCd D.D.
AbAb ВвВв CC DD
AbAb ВвВв CdCd D.D.
Рис. 20.1. Поведение при скрещивании гибрида, гетерозиготного по двум не-
зависимым транслокациям.
Первая транслокация затрагивает участки а и b в хромосомах А и В, а вто-
рая— участки с и d в хромосомах С и D. У родительского вида Pi последо-
вательность участков стандартная во всех четырех хромосомах, тогда как
вид Р2 несет обе транслокации. Гибриды Fi образуют гаметы 16 классов.
У гамет 12 классов имеются нехватки и дупликации по участкам а — Ь, уча-
сткам с — d или обеим парам участков. Гаметы четырех других классов сба-
лансированы и жизнеспособны. Эти четыре класса функциональных гамет
продуцируют в F2 различные типы гетерозигот по транслокациям (на схеме
не показаны) и четыре типа структурных гомозигот (показаны). Дальнейшие
объяснения см. в тексте.
чаются по двум транслокациям в четырех хромосомах. Они со-
держат хромосомные наборы ЛА В В CCDD и AbAbBaBaCdCdDcDc
соответственно (см. рис. 20.1). Гибрид Fi от скрещивания
между ними имеет конституцию AAbBBaCCdDDc, гетерозиготен
252
Часть пятая. Естественная гибридизация и ее результаты
по двум независимым транслокациям, и поэтому 25% его гамет,
как и следовало ожидать, фертильны (см. рис. 20.1).
В число функционирующих гамет входят гаметы двух роди-
тельских типов, которые нас здесь не интересуют, и гаметы двух
рекомбинационных типов (ABCdDc и AbBaCD), которые интере-
суют нас, поскольку они могут дать в F2 начало новым гомо-
зиготным типам АА ВВ Cdpd DCDC и AbAbBaBaCCDD (рис. 20.1).
Эти новые гомозиготные рекомбинанты полностью плодовиты
при скрещиваниях внутри себя, но частично стерильны при скре-
щиваниях друг с другом и с родительскими видами.
Разумеется, если число независимых транслокаций больше,
то они создают более мощный барьер хромосомной стерильности.
Так, следует ожидать, что гетерозигота по шести независимым
транслокациям даст всего 1,56% фертильных гамет. Межвидо-
вые гибриды растений, производящие примерно такие количест-
ва фертильной пыльцы, считаются высокостерильными. В об-
щем доля фертильных гамет, производимых гетерозиготами по
транслокациям, составляет l/2rt, где п —число независимых
транслокаций.
Кроме того, по мере усиления барьера, создаваемого хромо-
сомной стерильностью, число новых фертильных типов гомози-
гот, которые могут возникнуть в F2 или в последующих поколе-
ниях, также возрастает. И эти новые, гомозиготные по структур-
ным перестройкам рекомбинанты или большинство из них будут
изолированы от родительских видов мощными барьерами сте-
рильности.
Вернемся вновь к примеру гибрида, гетерозиготного по шес-
ти независимым транслокациям. Этот гибрид может дать в пос-
ледующих поколениях 62 новых плодовитых гомозиготных ре-
комбинанта. Такие рекомбинанты при возвратных скрещиваниях
с родительскими видами дадут гибридов, производящих от 3 до
50% фертильных гамет. При этом гибриды, полученные от воз-
вратных скрещиваний рекомбинанта, отличающегося самой вы-
сокой частотой, с любым из родительских видов, дадут 12,5%
фертильных гамет.
Структурные перестройки иных типов помимо транслокаций,
в частности транспозиции и реинверсии, также создают возмож-
ность для усиления барьеров хромосомной стерильности путем
рекомбинации. Транспозицией называют обмен участками в пре-
делах одной хромосомы, как это показано на рис. 8.2. Реинвер-
сия представляет собой результат двух последовательных инвер-
сий в одной и той же области хромосомы, причем одна из ин-
версий заключена в другой. Если ABCDEF — последователь-
ность генов в стандартной хромосоме, то в хромосоме с реинвер-
сией она имеет вид AECDBF. У структурных гетерозигот крос-
синговер между транспозициями или в пределах реинверсии при-
Гл 20 Рекомбинационное видообразование
253
водит к образованию хромосом с нехватками и дупликациями.
Поэтому множественные независимые транспозиции или реин-
версии в гетерозиготном состоянии могут создать высокую cie-
пень хромосомной стерильности при условии, что кроссипговер
всегда происходит между транспозициями или ^внутри реипвер-
сий Функциональные гаметы представляют собой нскроссовер-
ные типы по транспозициям или реинверсиям
Среди функциональных гамет будет несколько таких, у кото-
рых отдельные транспозиции или реинверсии, затронувшие раз-
ные хромосомы, рекомбинированы. Эти гаметы могут дать в F2
или в более поздних поколениях новые плодовитые типы струк-
турных гомозигот, которые будут изолированы от родительских
видов барьерами стерильности Транспозиции и реипверспи, по-
добно транслокациям, могут рекомбинироваться, давая новые
плодовитые изолированные липни.
В процессе рекомбинационного видообразования важную
роль играет система скрещивания участвующих в нем растений.
Система скрещивания, существующая в гибридной популяции,
будь то свободное неродственное скрещивание или самооплодо-
творение, оказывает решающее влияние на образование этой
гибридной популяцией новых плодовитых видов, изолированных
внутренними барьерами Инбридинг способствует рекомбинаци-
онному видообразованию [280].
Непосредственный конечный результат рекомбинационного
видообразования — возникновение структурной гомозиготы ново-
го типа Структурная гомозиготность достигается быстрее всего
потомками какой-либо структурной гетерозиготы в условиях ин-
бридинга Новая гомозиготная форма, коль скоро она образова-
лась, также лучше всего закрепляется и сохраняется системой
инбридинга. Свободное неродственное скрещивание в гибридной
популяции, напротив, сильно снижает вероятность как возник-
новения новой структурно-гомозиготной формы, так и после-
дующего сохранения такой формы в качестве самостоятельной
линии, размножающейся в чистоте [280, 282].
Поучительно сопоставить легкость закрепления новой струк-
турной гомозиготы в простом гипотетическом случае при про-
тивоположных системах скрещивания Допустим, что родитель-
ские виды различаются по двум независимым перестройкам, как
на рис 20 1 В результате гибридизации между этими видами
возникнет смешанная колония, состоящая, например, из двух
растений Fi и четырех растений Рь Допустим далее, что вос-
произведение гибридов даст ряд форм F2, которые будут одина-
ково жизнеспособны и одинаково плодовиты Какова же будет
частота одной определенной повой структурной гомозиготы (т е
АА ВВ CdCd DCDC па рис 20 1) в гибридном покотении F3 при
самооплодотворении и при случайном скрещивании'?
254 Часть пятая. Естественная гибридизация и ее результаты
При самооплодотворении эта гомозиготная форма составит
11% поколения F3. Если, однако, растения размножаются путем
•свободного скрещивания и если гибриды Fi случайно скрещива-
ются друг с другом и с растениями Pi в гибридной колонии, то
результаты будут совершенно иные. В этих условиях частота той
же самой новой гомозиготы среди гибридов F3 составит от О
до 5% (в зависимости от того, какие гибриды F2 будут воспро-
изводиться) , с модой в левой части этого диапазона.
При свободном скрещивании в типичной гибридной колонии,
состоящей из смеси растений Fi и растений родительских типов,
возникновение определенной новой структурной гомозиготы воз-
можно. Но вероятность появления такого генотипа невелика, а
кроме того, если он и возникнет, то он не будет размножаться
в чистоте. В гибридной колонии такого же состава самооплодо-
творение во много раз более эффективно в смысле создания и
закрепления нового вида.
Нам осталось вкратце обсудить судьбу различий по феноти-
пическим признакам между родительскими видами в процессе
рекомбинационного видообразования.
При рекомбинации участков хромосом рекомбинируют также
локализованные в них гены. И когда новая рекомбинация хро-
мосомных перестроек закрепляется в гомозиготном состоянии,
то закрепляются и входящие в нее гены. В результате дочерний
вид может унаследовать новую комбинацию морфологических и
физиологических признаков родительских видов, и эта новая
комбинация признаков будет размножаться в чистоте.
Экспериментальный подход
Гипотеза рекомбинационного видообразования создавалась на
протяжении 30 лет. В течение этого времени на представителях
ряда групп растений проводились эксперименты, относящиеся к
рекомбинационному видообразованию. Самые первые экспери-
менты принадлежали Герасимовой [237], Винге [901] и Лампрех-
ту [458].
Эти первые эксперименты ставились не для проверки отдель-
ных аспектов гипотезы рекомбинационного видообразования, ко-
торые были разработаны позднее. По сути дела, некоторые из
этих первых фактических данных были получены в эксперимен-
тах, которые и не были настоящими экспериментами в том, что
касается гипотезы рекомбинации. Взаимоотношения между тео-
рией рекомбинационного видообразования и экспериментом не
всегда складывались так, как это должно было быть, если исхо-
дить из той картины развития науки, какую обычно описывают
в учебниках. Тем не менее со временем накопилось довольно
много экспериментальных данных, если понимать термин экспе-
Гл 20. Рекомбинационное видообразование
255-
римент в широком смысле и если учитывать как первоначальные-
фактические данные различных исследователей, так и выведен-
ные ими заключения
Экспериментальные данные по рекомбинационному видооб-
разованию были получены в исследованиях на Crepis [237]г
Erophila [901], Nicotiana [742, 744], Elymus [782], Collinsia [162,
233] и Gilia [291]. Все эти данные подтверждают правильность
по крайней мере одного аспекта гипотезы рекомбинационного
видообразования. К числу более полных экспериментов, в кото-
рых исследовалось одновременно несколько аспектов гипотезы,
относятся эксперименты на Nicotiana, Elymus и Gilia. Большин-
ство проведенных исследований имело дело со слабыми барьера-
ми стерильности. Только в экспериментах с Elymus и Gilia были
предприняты попытки проверить гипотезу рекомбинационного-
видообразования в условиях очень мощных барьеров хромосом-
ной стерильности.
Ранпие эксперименты
Герасимова [237] проводила свои исследования на четырех ли-
ниях Grepis tectorum (2п = 8), различавшихся по гомозиготным
транслокациям в одной или в обеих парах хромосом. Нормаль-
ная линия С tectorum характеризуется стандартной последова-
тельностью участков во всех четырех хромосомах- А, В, С. D.
У мутантной линии I имеется гомозиготная транслокация, затра-
гивающая хромосомы А и D, а у мутантной линии II — хромо-
сомы В и С. Эти две линии, плодовитые сами по себе, оказались
частично стерильными при скрещиваниях друг с другом, чего и
следовало ожидать. Полустерильный гибрид Fj от скрещивания
между линиями I и II, которые были гетерозиготны по двум не-
зависимым транслокациям, дал поколение F2, содержавшее це-
лый ряд гетерозигот и гомозигот по транслокациям
Одно из растений F2, названное линией nova, содержало обе
транслокации в гомозиготном состоянии и было плодовитым.
Линия nova отличается от линий I и II одной транслокацией,
поэтому можно было предположить, что при скрещивании со-
своими непосредственными родительскими формами она даст
частично стерильных гибридов. От нормальной линии Crepis tec-
torum линия nova отличается двумя независимыми транслока-
циями, а поэтому следует ожидать, что скрещивания между эти-
ми линиями будут еще более стерильными. Герасимова [237J
подтвердила это, получив гибрид Fi между линиями tectorum и
nova, который оказался полустерильным; мейоз у него был на-
рушен, а фертильные семена составляли 30%.
256
Часть пятая. Естественная гибридизация и ее результаты
Это был первый случай экспериментального получения новой
изолированной плодовитой формы путем рекомбинации незави-
симых факторов, создающих хромосомную стерильность. Барьер
стерильности в этом случае 'был слабым и фенотипически линии
не различались. (
Группа Erophila verna (х=7) представляет собой комплекс
-автогамных однолетних видов-двойников с различным, полипло-
идным или анеуплоидным, числом хромосом 1[901], о чем уже
говорилось в гл. 11. Винге скрещивал две следующие формы
Е.verna: Е. confertifolia (п=15) и Б. violacea-petiolata (п=32).
У гибридов конъюгация хромосом была сильно нарушена и'от-
мечалась высокая стерильность как пыльцы, так и семян; фер-
тильность семян составляла всего 3% нормальной [901].
Поколение F2 состояло из 451 растения, среди которых наб-
людалось расщепление по морфологическим признакам, плодо-
витости и числу хромосом. Четыре растения F2 послужили осно-
вателями для четырех семей F3 и их инбредных потомков вплоть
до поколения F9. Линии продолжали расщепляться, достигая
•единообразия в разное время — от поколения F5 до F9. Восемь
линий, которые были подвергнуты цитологическому анализу,
достигли в F7—F9 стабильности по числу хромосом, которое во
всех случаях отличалось от исходного и было различным в раз-
ных линиях. От скрещивания (п= 15) X (и=32) произошли ли-
нии с п=22, 23, 25, 29, 31 и 34. Эти линии были плодовитыми,
морфологически единообразными и представляли собой новые
анеуплоидные типы [901].
Эксперименты Винге показали, что гибридизация между ро-
дительскими видами, находящимися на разных уровнях плоид-
ности, может привести к возникновению новых плодовитых кон-
стантных анеуплоидных типов. Предполагается, хотя соответст-
вующих возвратных скрещиваний не производилось, что гибри-
ды от скрещиваний этих анеуплоидов с родительскими видами
будут до некоторой степени стерильны. Этот эксперимент остав-
ляет открытым вопрос о том, может ли гибридизация давать так-
же начало новым микровидам на гомоплоидном уровне.
Последующие эксперименты при существовании
слабых барьеров стерильности
‘Следующая группа экспериментов, которую нам предстоит рас-
смотреть, проводилась на Nicotiana, Collinsia, Elymus и Gilia;
они отличаются от первых экспериментов тем, что ко времени
их проведения формулировка гипотезы рекомбинационного видо-
образования уже стала вырисовываться более четко Это позво-
ляло планировать эксперименты таким образом, чтобы с их по-
мощью можно было действительно подвергнуть проверке гипо-
Гл 20 Рекомбинационное видообразование
257
тезу пли ее наиболее существенные части. В этом разделе мы
рассмотрим эксперименты, проводившиеся с Nicotiana и СоШп-
sia в условиях существования слабых барьеров стерильности
Смит и его сотрудники [742, 744] провели длительный селек-
ционный эксперимент на гомоплоидных пнбредных линиях, про-
исходящих ог скрещивания между разными видами Nicotiana.
Оба родиюльских вида, N. langsdorfii и N sanderae, — дипгтои-
ды с 2п=18. Они различаются по величине цветков (у N. lang-
sdorfii трубка венчика короткая, а у N sanderae — длинная), а
также по вегетативным и другим признакам.
У гибрида Fi пыльца была полустерильная^ (число фертиль-
ных пыльцевых зерен соиавляло в среднем 55%), а семена —
довольно фертильными Мейоз был несколько нарушен. Большая
часть хромосом конъюгировала, образуя биваленты, но некото-
рые из них соединялись в цепочку или оставались одиночными;
у таких хромосом наблюдалось отставание в анафазе [744].
Снижение фертильности пыльцы, вероятно, обусловлено главным
образом этими нарушениями конъюгации
В F2 были заложены три селекционные линии, которые про-
должали вести нхтем самоопыления oiдельных родительских рас-
тений вплоть до Fio Искусственный отбор производили по дли-
не трубки венчика В одной липни вели отбор на длинную труб-
ку, как у родительского вида Nicotiana sanderae, в другой — на
короткую трубку, как у N. langsdorfii, а в третьей — на трубку
промежуточной длины Все три линии реагировали па отбор по
величине цветков, но при этом стали различаться также но дру-
гим признакам цветка и вегетативным признакам, не подвергав-
шимся отбору. Они стали отличаться морфологически как друг
от др\1а, так и от родительских видов [744].
Отобранные таким образом линии оставались диплоидными,
подобно родительским видам Наибольший интерес для нас
представляют диплоидные производные с короткими венчиками
и другими связанными с этим признаками, так как у них в по-
колениях F6 — F9 фертильность пыльцы повысилась поч1и до
нормального уровня, тогда как две другие линии в поздних по-
колениях оставались или становились полустер ильным и [744].
Формы с мелкими цветками при скрещивании с обеими ро-
дительскими линиями давали полустсрильпых гибридов. Фер-
тильность пыльцы у 1ибридов, полученных при возвратном скре-
щивании с Nicotiana langsdorfii, составляла 46—58%. а при воз-
вратном скрещивании с A-’. sanderae — 60—61%. Как у тех, так
и у арутх гибридов отмечались некоторые отклонения в мейо-
зе Таким образом, линия с мелкими цветками была стерильна
при скрещивании с пюбой из родительских линий, так же как и
эти липин при скрещивании одна с другой [742, 744J.
Г-
258 Часть пятая. Естественная гибридизация и ее результаты
Гарбер и его сотрудники изучали гибридов и гибридных по-
томков от скрещиваний Collinsia corymbosaxC. tinctoria и С. со-
rymbosaXC. multicolor [162, 233]. Оба родительских вида ди-
плоидны (2п=14) и различаются по ряду морфологических
признаков.
Межвидовые гибриды F] содержали транслокационные коль-
ца из 6 хромосом и характеризовались высокой плодовитостью.
У растений Fa и F3 мейоз протекал нормально, с образованием
бивалентов. Большинство растений, составлявших поколения F2
и F3, имели геномы родительских типов. Однако в этих поко-
лениях были три линии, представляющие каждое из двух перво-
начальных скрещиваний, которые обладали новыми рекомбина-
ционными геномами, отличавшимися от родительских по транс-
локациям. Кроме того, эти линии отличались новыми комбина-
циями морфологических признаков [162, 233].
Более поздние эксперименты при существовании
мощных барьеров стерильности
Первый эксперимент по рекомбинационному видообразованию
при наличии очень мощного барьера хромосомной стерильности
был проведен на гибридном травянистом растении из комплек-
са Elymus glaucus. Родительскими видами этого гибрида были
Elymus glaucus и Sitanion jubatum; у обоих видов число хро-
мосом 2п=28. Этот эксперимент, поставленный Стеббинсом, опи-
сан в его кратком сообщении [783].
У этого межвидового гибрида происходит конъюгация хро-
мосом с образованием 11—14 бивалентов, но он высоко стери-
лен: количество фертильной пыльцы не достигает 1%, а число
семян равно примерно 1 на 10 000 цветков. Цитогенетический
анализ искусственных аллополиплоидов, происходящих от этого
гибрида, показывает, что стерильность его имеет частично хро-
мосомную природу и обусловлена мелкими перестройками, не
очень сильно нарушающими образование бивалентов, а частич-
но— генного характера [783, 786].
Гибриды Fi, происходящие от указанных выше родительских
видов, получали несколько раз от скрещиваний между разными
микровидами группы Elymus glaucus и разными линиями Sita-
nion jubatum или родственного ему S. hist г ух. Затем проводили
возвратные скрещивания растений Fi, полученных от различных
гибридных комбинаций, с Е. glaucus; доля семян и растений Bj
была при этом очень невелика: в результате нескольких скрещи-
ваний было получено в общей сложности 173000 цветков, кото-
рые принесли всего 15 семян Bi и 11 неудвоенных беккроссов.
Растения Bi в большинстве случаев представляли собой по мор-
Гл 20 Рекомбинационное видообразование
259
фологии и плодовитости реверсии к Е. glaucus, а поэюму они
пас здесь не интересуют [783].
Одно из растений Bi обладало, однако, морфологическими
признаками как гибрида Fb так и Elymus glaucus, послуживше-
го отцовским растением, и было полустсрильпым. При самоопыле-
нии оно дало в поколениях I] и 12 мощное и полностью плодо-
витое потомство. Эти инбредные семьи характеризовались рас-
щеплением по морфологическим признакам с некоторой измен-
чивостью в направлении Sitanion jubatum [783].
Одно растение из поколения Ь скрестили затем с исходной
родительской линией Elymus glaucus, чтобы выяснить их вза-
имоотношения по фертильности Гибриды F], полученные в ре-
зультате такого тесшрующего скрещивания, оказались высоко
стерильными, дав всего 0—3% фертильной пыльцы [783].
Работа, проведенная па Elymus, продвинула еще на один
шаг экспериментальное доказательство рекомбинационного ви-
дообразования. Она подтверждает эту гипотезу на важном при-
мере с очень мощным барьером стерильности, имеющим, по
крайней мере частично, хромосомную природу. Изолированное
гомоплоидное производное было получено, однако, путем воз-
вратного скрещивания в поколении Fi, а не путем прямого ин-
бридинга. При такой постановке эксперимента снижается его
применимость к тем реально существующим в природе ситуа-
циям, в которых скорее всего можно ожидать рекомбинационно-
го видообразования
Следующий эксперимент, который нам предстоит рассмот-
реть, был поставлен с парой видов из 1р>ппы Gilia inconspicua
и их гибридными производными [291]. Родительские виды G. ma-
lior и G. modocensis — автогамные однолетние растения с чис-
лом хромосом 2п = 36. Гибриды Fi от скрещивания между ними
высокостерильны фертильная пыльца у них составляет всего
2%, а всхожесть семян — менее 1% Конъюгация хромосом у
гибрида сводится к образованию 1 — 10 бивалентов Имеются не-
зависимые данные, свидетельствующие о том, что такое нару-
шение конъюгации в значительной степени обусловлено струк-
турными перестройками между родительскими видами, так что
гибридная стерильность имеет главным образом хромосомную
прI!роду [289, 291].
Этот гибрид, несмотря на свою щерилыюсть, дал некоторое
число потомков F2 и последующих поколений на гомоплоидном
уровне Значительно больше половины растений в поколениях
F2 и F6 характеризовались пониженной жизнеспособностью или
стерильностью или же одновременно и тем и другим. Растения
F2 были использованы для закладки ряда пнбредпых линий, в
которых на протяжении последующих поколений проводился от-
бор на мощность и плодовитость в сочетании с нормальным те-
17*
260 ^Часть пятая. Естественная гибридизация и ее результаты
чением мейоза. Этот отбор оказался успешным в нескольких ли-
ниях, которые в конечном итоге достигли в последующих поко-
лениях полной мощности и плодовитости при нормальном тече-
нии мейоза и без изменения плоидности [289, 291].
Плодовитые гомоплоидные растения, полученные от стериль-
ных межвидовых гибридов, скрещивали затем с родительскими
видами и между собой, для того чтобы выяснить их генетиче-
ские взаимоотношения. Одна группа линий, известная под наз-
ванием «Ветвь II», оказалась как бы восстановленной формой
Gilia modocensis, соответствуя ей как по своим фенотипическим
признакам, так и по плодовитости при скрещиваниях. Ветвь II
не служит подтверждением гипотезы рекомбинационного видо-
образования.
Другое плодовитое гомоплоидное производное, названное
«Ветвь III», унаследовало морфологические признаки родитель-
ских видов в новой комбинации. Ветвь III была сходна с Gilia
modocensis по четырем признакам, с G. malior — по трем и с
промежуточной, подобно гибридам Fi — по четырем другим при-
знакам [291]. Характер ветвления и форма листьев у этой груп-
пы линий и родительских видов показаны на рис. 20.2.
При скрещиваниях Ветви III с Gilia malior, G. modocensis и
Ветвью II наблюдалась хромосомная стерильность. У гибридов
Fj от скрещиваний G. malior с Ветвью III, например, конъюга-
ция хромосом была нарушена (в среднем образовывалось 13 би-
валентов), а фертильная пыльца составляла 4—18%. Всхожесть
семян была ниже 1%. Таким образом Ветвь III изолирована от
родительских и сестринских форм мощным барьером стериль-
ности [291].
Поведение Ветви III по всем существенным аЪпектам соот-
ветствует тому, чего следовало ожидать на основании гипотезы
рекомбинационного видообразования. Она была получена от
межвидового гибрида с высокой хромосомной стерильностью пу-
тем чистого инбридинга. Шаг за шагом в результате отбора в
ряде последовательных поколений опа достигла полной плодо-
витости в сочетании с нормальным мейозом и без изменения
плоидности. При скрещиваниях с родительскими видами
Ветвь III дает стерильных гибридов. Она обладает собственным
особым сочетанием морфологических признаков родительских
видов, а также их факторов хромосомной стерильности.
Аналогичные экспериментальные исследования проводятся в
настоящее время на видах рода Oryza. У О sativa и у О. glaber-
rima число хромосом 2п=24. Гибриды Fi между ними высоко-
стерильны. Путем возвратных скрещиваний этих гибридов с ро-
дительскими видами можно получить дальнейшие поколения
потомков. Среди инбредпых потомков от этих последующих
скрещиваний было несколько форм с новыми рекомбинациями
Гл. 20. Рекомбинационное видообразование
261
Рис 20.2 Характер ветвления и форма листьев у двух иптерстерильных ви-
дов Gilia и у Ветви III — плодовитой изолированной гомоплоидной формы,
происходящей от гибрида между ними [291].
См также рис 17 8, относящийся к тем же родительским видам и происхо-
дящей от них Ветви II.
признаков родительских видов. Это позволяет предполагать ре-
комбинационное видообразование Особенности этих новых лц-
ний в отношении плодовитости пока еще не выяснены [708].
Роль блочного наследования в рекомбинационном
видообразовании
Некоторые экспериментальные данные, относящиеся к Gossy-
pium, Clarkia и Gilia, позволяют предположить, что перестроен-
ные участки хромосом у близких видов несут различные аллели
генов, оказывающих влияние на рост и мощность. У пар видов,
принадлежащих к этим и другим родам, в перестроенных участ-
ках хромосом можно установить локализацию генов, детермини-
562 Часть пятая. Естественная гибридизация и ее результаты
рующих морфологические признаки. Фактические данные, ка-
сающиеся блочного наследования систем генов М—V, были из-
ложены в другой моей работе [283], а краткий обзор этих дан-
ных приведен в гл. 8.
Здесь нам предстоит рассмотреть влияние на рекомбинаци-
онное видообразование генов М—V, локализованных в пере-
строенных участках, по которым различаются родительские ви-
ды. Допустим, что между двумя видами, которые различаются
по блокам генов М— V, имеющим такую же протяженность, как
и структурные перестройки, происходит естественная гибриди-
зация. Допустим далее, что гибридное потомство растет и раз-
множается в местообитании одного из родительских видов (Pi),
но не в отличающемся по экологическим условиям местообита-
нии другого вида (Р2).
Очевидно, что в такой ситуации отбор под действием среды
будет благоприятствовать гибридным выщепенцам, несущим
аллели жизнеспособности вида Рь Но, кроме того, можно ожи-
дать, что в этой гибридной популяции действие отбора будет
направлено на сохранение целых блоков генов или, что то же
самое, перестроенных участков хромосом вида Pi в ущерб соот-
ветствующим участкам вида Р2. Таким образом, можно ожидать,
что в данном случае гомоплоидные гибридные производные по
своей зависящей от хромосом плодовитости, критическим эколо-
гическим предпочтениям и связанным с ними морфологическим
признакам будут ревертировать к генотипу одного из родитель-
ских видов (Pi).
Именно такой результат наблюдался при контролируемых
экспериментальных условиях в потомстве гибрида от скрещи-
вания Gilia malior и G. modocensis, обладавшего хромосомной
стерильностью. Мы имеем в виду гомоплоидные линии, назван-
ные Ветвью II. Эти линии в условиях среды, типичной для
G. modocensis, подвергали отбору на мощность, но не на мор-
фологическое сходство с G. modocensis или плодовитость при
скрещиваниях с последним. Тем не менее в результате такого
отбора была получена форма, гомологичная по своим хромосо-
мам G. modocensis, плодовитая при скрещивании с этим роди-
тельским видом и морфологически очень сходная с ним [288].
Пример с Ветвью II был рассмотрен в гл. 17 и проиллюстриро-
ван на рис. 17,8.
Из всего сказанного можно сделать вывод, что гены, опреде-
ляющие жизнеспособность и локализованные в участках, за-
тронутых перестройками, потенциально способны тормозить или
сдерживать процесс рекомбинационного видообразования. Этот
потенциальный тормоз может начать действовать, если гибрид-
ная популяция растет в местообитании одного из родительских
видов
Гл 20 Рекомбинационное видообразование
265
Наиболее подходящим местом для рекомбинационного видо-
образования служит, вероятно, какое-нибудь повое, измененное
или «рекомбинационное», местообитание, находящееся поблизо-
сти от гибридной популяции Такая среда может оказаться не-
обходимой в качестве местообитания для индивидуумов с новой
комбинацией перестроенных участков и может послужить фак-
тором отбора, способствующим сохранению этих участков, если
они несут блоки генов, обладающие адаптивными свойсшами.
Таким образом, следует ожидать, что рекомбинационное видо-
образование будет происходить главным образом в тех место-
обитаниях, которые по своим экологическим характеристикам
отличаются от местообитаний родительских видов.
О роли рекомбинационного видообразования
в природе
Конечная цель проведения экспериментов по рекомбинационно-
му видообразованию заключается в том, чтобы установить осу-
ществимость этого процесса и его роль в эволюции растений
Попытаемся теперь выяснить роль рекомбинационного видообра-
зования в природе в свете имеющихся экспериментальных дан-
ных.
Теоретически рекомбинационное видообразование можно
сравнить с явлением амфнплоидии, о котором известно гораздо
больше Эти два процесса представляют собой два альтернатив-
ных пути от гибрида с хромосомной стерильностью к новому
птодовигому изолированному дочернему виду Как показывают
многочисленные экспериментальные и цитотаксономические дан-
ные, амфиплоидный путь наблюдается в эволюции растений
очень часто. Логично предположить, что рекомбинационное ви-
дообразование также представляет собой один из обычных про-
цессов, происходящих в природе; оно, быть может примерно
столь же обычно, как и амфиплоидия, но в отличие от последней
пе оставляет следов, которые можно было бы выявить путем
цитотаксономических исследований.
Эксперименты, проводившиеся на Gilia, — единственные, в
которых формы, происходящие от высокостерильпого гибрида,
были получены путем чистого инбридинга, — могут преподать
нам урок в этом отношении. В первых поколениях потомков от
скрещивания G. malior и G modocensis наблюдалось значитель-
ное разрушение гибридов Эти линии едва удавалось сохранить
в благоприятных искусственных условиях; что же касается их
выживания в естественных условиях, то оно еще менее вероятно
[289]. У некоторых из этих линий, которым удалось преодолеть
барьер, создаваемый разрушением гибридов, происходил, кроме
того, возврат к одному из родительских видов [2881. В отличие
'264
Часть пятая. Естественная гибридизация и ее результаты
от них единственный новый дочерний вид, которому удалось воз-
никнуть, на протяжении своей долгой истории в условиях экспе-
риментальной среды оставался конституционно слабым [291].
Эксперименты с Gilia показали, что при гибридизации между
высоко интерстерильными и хорошо дифференцированными ви-
дами преодолеть барьер, создаваемый разрушением гибридов,
крайне трудно.
В прежних формулировках гипотезы рекомбинационного ви-
дообразования эффективность барьера, обусловленного разруше-
нием гибридов, не учитывалась. В теоретических и эксперимен-
тальных исследованиях этого способа видообразования все вни-
мание было сосредоточено на одном аспекте проблемы — гиб-
ридной стерильности. Амфиплоидия представляет собой один
из способов, позволяющих избежать хромосомной стерильности
у межвидового гибрида, а рекомбинационное видообразование
можно рассматривать как альтернативный выход из того же
тупика. Однако при этом оставалось недостаточно ясным, поче-
му амфиплоидия дает также возможность преодолеть и барьер,
создаваемый разрушением гибридов, тогда как рекомбинацион-
ное видообразование такой возможности лишено.
Наиболее подходящий вывод, к которому мы можем прийти,
исходя из имеющихся в нашем распоряжении эксперименталь-
ных данных, состоит в том, что рекомбинационное видообразо-
вание— гораздо менее обычный способ видообразования у рас-
тений, чем амфиплоидия.
Экспериментальные исследования показали, однако, что в
искусственных условиях рекомбинационное видообразование
имеет место. Это в свою очередь позволяет допустить, что такой
процесс может иногда происходить и в природе.
Примером естественного вида, который, вероятно, возник в
результате рекомбинационного видообразования, служит Gilia
achilleifolia' (2п=18). Этот диплоидный вид встречается в Ка-
лифорнии, в южной части Берегового хребта. Считается, что он
возник в результате гибридизации между какими-то древними
представителями групп диплоидных видов G. capitata и G. ange-
lensis [273].
В пользу этой филогенетической гипотезы имеются достаточ-
но убедительные данные. Прежде всего растения Gilia achillei-
folia по морфологическим признакам, относящимся ко всем их
частям, занимают промежуточное положение между двумя ро-
дительскими видами; кроме того, они крайне изменчивы по сво-
им признакам, и такая изменчивость выражается в форме диф-
ференциации на локальные расы. Некоторые расы G achilleifo-
lia морфологически близки к G. capitata, тогда как другие близ-
ки к G. angelensis; систематики иногда даже путали первых с
G. capitata, а вторых с G. angelensis [273].
Гл. 20 Рекомбинационное видообразование 265
Два предполагаемых родительских вида в настоящее время
гибридизируют в природе в некоторых районах на юге Калифор-
нии Весьма важно, что возникшие в результате этой гибриди-
зации интрогрессивные формы морфологически сходны с Gilia
achilleifolia [273].
Известно, что часть хромосом Gilia achilleifolia гомологич-
на, а часть негомологична хромосомам членов групп видов
G capitata и G. angelensis [274]. Соотношение геномов всех этих
форм совместимо с гипотезой рекомбинационного происхожде-
ния G. achilleifolia. Кроме того, G. achilleifolia самосовместима,
а некоторые се расы преимущественно автогамны. Таким обра-
зом, система размножения этой формы благоприятствует закреп-
лению нового генома рекомбинационного типа.
Необходимо провести дополнительные генетико-таксономиче-
ские исследования с целью увеличить число вероятных приме-
ров. Необходимы также более чистые экспериментальные ис-
следования по рекомбинационному видообразованию. Мы еще
очень далеки от того, чтобы оценить, сколь велика роль реком-
бинационного видообразования в эволюции растений.
21 Гибридные комплексы
У многих групп видов, принадлежащих к разным родам и раз-
ным семействам растений, наблюдается сходный характер из-
менчивости. Этому общему типу изменчивости присущи следую-
щие черты. Группа видов содержит многочисленные и разнооб-
разные формы, интерградирующие по морфологическим призна-
кам и экологическим предпочтениям. Ввиду такой интерграда-
ции эти разные формы можно было бы считать членами одного
политипического вида, но диапазон морфологической и экологи-
ческой изменчивости группы в целом сильно превышает ее диа-
пазоны у обычных политипических видов, относящихся к тому
же роду. Кроме того, некоторые из этих форм в тех или иных
зонах симпатрического контакта остаются более или менее обо-
собленными. Интерградация между ними не похожа на интер-
градацию между расами.
Таксономически такая группа является «критической» и
«трудной». Ни одна система классификации, созданная на осно-
ве общепринятых гербарных и полевых методов, не дает воз-
можности удовлетворительно произвести обычное определение.
Если же проанализировать данную группу с применением тех
же рутинных морфологических и географических методов, но не
с таксономической, а с популяционно-биологической точки зре-
ния, то выясняется, что она представляет собой комплекс отно-
сительно слабо выраженных видов, полувидов и (или) микрови-
дов. Генетико-таксономические данные, если их удается полу-
чить, нередко подтверждают концепцию о группе взаимосвязан-
ных видов, поскольку при этом обнаруживается существование
барьеров стерильности или других репродуктивных барьеров
между некоторыми морфологически различными и симпатриче-
скими формами.
У групп растений, обладающих такой характерной изменчи-
востью, нет ни одной общей для всех генетической системы. Они
могут состоять из диплоидных или полиплоидных популяций
или же одновременно из тех и других. Размножение может быть
половым, почти половым (субсексуальным) или бесполым. Раз-
личные группы растений, создающие затруднения для система-
тиков, сходны в общих чертах по характеру изменчивости, но
отнюдь не все они сходны по типу своих генетических систем.
Существует, однако, и другой фактор, общий для всех групп
растений, обладающих описанным выше типом изменчивости.
Этот фактор — широко распространенная естественная гибриди-
зация в пределах данной группы и закрепление значительного
числа форм гибридного происхождения. В соответствии с этим
такие группы можно объединить под общим названием гибрид-
I л. 21 1 ибридные комплексы
267
ных комплексов. Гибридный комплекс — это группа видов, со-
стоящая из трех или большего числа исходных предковых видов
и их гибридов или потомков этих гибридов, в которой в резуль-
тате естественной гибридизации морфологическая прерывистость
между исходными формами оказалась стертой [272].
Классификация
Гибридные комплексы можно классифицировать в зависимости
от способа стабилизации воспроизведения естественных гибри-
дов или их потомков [272]. В одной из предыдущих глав мы пе-
речислили известные способы гибридного видообразования;
здесь мы вновь приведем этот перечень, с тем чтобы на него бы-
ло удобнее ссылаться.
Известны следующие способы стабилизирующего размноже-
ния гибридов- 1) вегетативное размножение, 2) агамоспермия;
3) перманентная гетерозиготность по транслокациям; 4) перма-
нентная нечетная полиплоидия; 5) амфиплоидия 6) рекомбина-
ционное видообразование; 7) гибридное видообразование при
наличии внешних барьеров. Эти способы создают возможность
для размножения в чистоте высокогегерозиготного генотипа (1 —
5) или гомозиготного выщспенца (6—7) в условиях бесполою
(1—2) или полового (3—7) размножения.
Если процесс гибридного видообразования независимо повто-
ряется в разных линиях, так что из трех или более исходных
видов образуется два или более гибридных вида, то возникает
гибридный комплекс. Тип гибридного комплекса соответствует
способу стабилизации воспроизведения гибридов.
1. Гомогампый комплекс Формы гибридного происхождения
размножаются половым путем с нормальным течением мейоти-
ческого цикла. Дочерние виды гибридного происхождения ди-
плоидны или по меньшей мере неудвоившиеся и гомоплоидные
по отношению к исходным родительским видам _
2. Полиплоидный комплекс. Формы, возникшие в результате
естественной гибридизации, представляют собой четные поли-
плоиды (обычно амфиплоиды), размножающиеся половым пу-
_тсм. --------------------------------- ' —
3. Агампый комплекс Увеличение численности гибридов или
форм гибридного происхождения полностью или частично осу-
ществляется при помощи неоплодотворенных семян. Эту концеп-
цию можно расширить, с тем чтобы включить в нее бесполые
папоротники у которых гибриды размножаются путем апога-
метии.
4 Клональный комплекс. Численность гибридов или форм
гибридного происхождения увеличивается бесполым путем, при
помощи различных способов вегетативного размножения.
268
Часть пятая. Естественная гибридизация и ее результаты
5. Гетерогамный комплекс. Гибриды или формы гибридного
происхождения представляют собой перманентные гетерозиготы
по транслокациям или перманентные нечетные полиплоиды.
В гомогамных комплексах дочерние виды возникают от роди-
тельских видов в результате рекомбинационного видообразова-
ния или гибридного видообразования при наличии внешних
барьеров. В соответствии с этим гомогамный комплекс может
состоять из видов, разделенных барьером хромосомной стериль-
ности, или же из видов, плодовитых при скрещиваниях между
собой.
У двух генетических систем — перманентной гетерозиготности
по транслокациям и перманентной нечетной полиплоидии —
имеется несколько общих черт, а именно продолжающаяся гете-
розиготность по хромосомам, комплементарные геномы в муж-
ских и женских гаметах и половое размножение, нередко само-
оплодотворение. Структурная или нумерическая гибридность
поддерживается благодаря половому размножению, нередко са-
мооплодотворению, из поколения в поколение. Эти две генетиче-
ские системы также дают начало гибридным комплексам сход-
ных типов. Вследствие такого сходства по ряду признаков для
обозначения создаваемых этими системами эволюционных про-
дуктов (гетерогамный микровид и гетерогамный комплекс)
можно использовать общую терминологию. Тем не менее гете-
рогамные комплексы делятся на два подтипа: комплексы, со-
держащие диплоидные гетерозиготы по транслокациям, и комп-
лексы, состоящие из нечетных полиплоидов.
Приведенная выше классификация гибридных комплексов
носит аналитический характер. Реальные гибридные комплексы,
развивающиеся в той или иной группе растений, могут содер-
жать смесь разных типов. Многие высокие и некоторые низкие
полиплоиды являются агамоспермными. Поэтому агамные комп-
лексы нередко накладываются на полиплоидные комплексы,
комбинируясь с ними. Подобным же образом в разных ветвях
одной и той же группы растений может иметь место как реком-
бинационное видообразование, так и амфиплоидия. В таких слу-
чаях возникает частично гомогамный, а частично полиплоидный
гибридный комплекс. В реально существующих клональных
комплексах наряду с полиплоидией, агамоспермией или поло-
вым размножением гибридов нередко встречается вегетативное
размножение.
Роль системы размножения
Естественные гибриды образуются между видами растений, об-
ладающими любой системой размножения. Однако результат
естественной гибридизации в очень сильной степени зависит от
Гл 21 Гибридные комплексы
269
системы размножения участвующих в ней растений, и влияние
этой системы может сказаться на типе развивающегося в конеч-
ном итоге гибридного комплекса
У однолетних растений, гибриды которых располагают лишь
коротким периодом времени для размножения, гибридизация в
условиях свободного скрещивания приводит к интрогрессии, а в
условиях самооплодотворения — к амфиплоидии [275]. Эти ожи-
даемые результаты очень четко подтверждаются у однолетних
гилий, у которых формы с перекрестным оплодотворением ха-
рактеризуются сильно выраженной интрогрессией, тогда как
автогамные формы даюг амфинлоидов. Таким образом в гибри-
дизирующих группах однолетних растений автогамная система
размножения способствует образованию полиплоидных комплек-
сов.
У многолетних цветковых растений, способных к вшетатив-
ному размножению, полиплоидия, как мы увидим в части шес-
той. встречается относительно часто, и, разумеется, такие расте-
ния также образуют дочерние клоны Поэтому следует ожидать,
что широкая гибридизация в группе многолетних растений, раз-
множающихся вегетативным способом, приведет к образованию
полиплоидных и клональных комплексов
У папоротников, которые являююя многолетними растения-
ми, полиплоидия гораздо чаще встречается в равпоспоровых
группах, чем в разноспоровых [442]. По-видимому, у равноспо-
ровых значительное место занимает самооплодотворение [442].
Таким образом, следует ожидать, что равноспоровость и само-
оплодотворение способствуют развитию полиплоидных комплек-
сов в гибридизирующих группах папоротников.
Как было отмечено в предыдущей главе, самооплодотворение
у однолетних и у многолетних растений служит фактором, спо-
собствующим рекомбинационному видообразованию. Таким об-
разом, следует ожидать, что гомогамные комплексы, состоящие
из видов и микровидов, характеризующихся хромосомной сте-
рильностью, при наличии подходящих местообитаний будут раз-
виваться в гибридизирующие группы преимущественно автогам-
ных растений.
У большинства агамоспермных растений их предковые фор-
мы, размножающиеся половым путем, бывают двудомными, од-
нодомными или самонесовместимыми [317, 318]. Гушафсон [317,
318] отметил, что растения, обладающие той или"иной из этих
систем размножения, в особенности гибридные растения, б\дут
иметь очень небольшие шансы продолжить свое существование
путем полового размножения в изолированных насаждениях,
тогда как агамоспермия и вегетативное размножение — относи-
тельно легкие пути, позволяющие выйти из тупика стерильности.
Облигатное перекрестное опыление у гибридов представляет со-
270
Часть пятая. Естественная гибридизация и ее результаты
бой, таким образом, фактор, благоприятствующий развитию
агамных и клональных комплексов.
Автогамия преобладает у перманентных гетерозигот по транс-
локациям группы Oenothera biennis и, по-видимому, у перма-
нентных нечетных полиплоидов группы Rosa canina. Каждая из
этих групп возникла от предковых форм, для которых были ха-
рактерны нормальный мейоз и свободное опыление. В каждой
группе самооплодотворение составляет один из компонентов ге-
нетической системы, обеспечивающей поддержание перманент-
ной гибридности. В экологическом отношении формы гибридного
происхождения особенно успешно развиваются в сорных или
окультуренных местообитаниях; так обстоит дело, в частности,
в группе Oenothera.
Не исключено, что предковые популяции обладали совершен-
но необходимыми им гетерозисными свойствами и поддерживали
эти свойства при помощи непрерывного перекрестного опыления.
В таком случае при заселении нового местообитания, где не бы-
ло подходящих опылителей, у них развилась в качестве компен-
сации система перманентной гетерозиготности, основанная на
самооплодотворении при участии неодинаковых гамет [144, 146].
Согласно этой гипотезе, главным стимулом, вызвавшим эволю-
цию по крайней мере некоторых гетерогамных комплексов, была
продолжавшаяся зависимость популяций от гетерозиса на фоне
обеднения среды опылителями; происходившее одновременно из-
менение системы размножения —от свободного опыления к са-
мооплодотворению— явилось частью механизма, обеспечиваю-
щего подгонку всей генетической системы к новым условиям су-
ществования.
Гомогамный комплекс
Гомогамный комплекс представляет собой гибридный комплекс,
в котором новые производные виды возникли в результате про-
цессов гибридного видообразования, не сопряженных с резки-
ми изменениями в генетической системе. Нам уже известны про-
цессы рекомбинационного видообразования при наличии барь-
еров хромосомной стерильности и гибридного видообразования
при наличии внешних барьеров. Третий возможный процесс, ко-
торым следует заняться в будущих исследованиях, —формиро-
вание новых, контролируемых генами барьеров стерильности и
несовместимости в результате рекомбинации. В зависимости от
того, какой из этих различных процессов ответствен за видооб-
разование, возникающие виды могут быть изолированы один от
другого и от родительских видов барьерами хромосомной сте-
рильности, внешними барьерами, генной стерильностью и несов-
Гл 71 Гибридные комплексы
271
местимостью или же одновременно и внутренними и внешними
барьерами.
В любом случае производные виды остаются на том же уров-
не плоидности, что и предковые виды. Кроме того, у гибридных
производных сохраняются нормальный половой цикл и нормаль-
ное течение мейоза Если родительские виды представляют со-
бой диплоидов, размножающихся половым путем, то их дочерние
виды — также диплоиды с половым размножением.
В гибридных комплексах всех других типов производные ви-
ды можно отличать от родительских но их цитогене шческим приз-
накам или системе размножения. Однако в гомогамных комп-
лексах это невозможно, поскольку гибридные производные со-
храняют исходную генетическую систему предковых видов.
Вследствие этого анализировать гомогамные комплексы очень
трудно, хотя и возможно. Гомогамный комплекс легко может
остаться незамеченным, и многие из них, вероятно, так и нс бы-
ли распознаны. Этим, вероятно, и объясняется немногочислен-
ность известных примеров.
Гомогамный комплекс облиствеиностебельных
гилий
Облиственностебельные гилии (секция Gilia рода Gilia) группа
стерильных при скрещиваниях друг с другом однолетних травя-
нистых растений, широко изучавшихся как таксономически, так
и цитогенетически Большинство видов этой группы диплои
дны (2п=18), а некоторые —тетраплоидны (2п = 36) Все виды
размножаются половым путем, а системы размножения варьиру-
ют от облигатного перекрестного опыления до преобладающего
инбридинга [272, 274].
Взаимоотношения между Gilia capilala и G tricolor в этой
группе были описаны в гл 4 как пример пары хороших биологи-
ческих видов. В гл. 9 был дан обзор плодовитости при скрещи-
ваниях в пределах этой секции в целом, чтобы проиллюстриро-
вать некий общий тип взаимоотношений, обычных для однолет-
них растений, — тип Madia. Здесь будет описана сетчатая фило-
гения этой группы как пример гомогамного комплекса
Мы рассмотрим виды облиственностебельных гилий, произ-
растающих в Калифорнии — центре распространения и вероят-
ном центре происхождения этой секции, а точнее виды, произ-
растающие в материковой Калифорнии, без близлежащих остро-
вов Эти виды перечислены на рис 21.1 Вес они диплоидны, за
исключением G clivorum (2/г = 4х = 36)
Облиствепностебельные гилии делятся на три группы видов,
различающиеся по морфологическим признакам и экологическим
предпочтениям: 1) группа Gilia tricolor, в которую входят G. tri-
272
Часть пятая. Естественная гибридизация и ее результаты
Рис. 21 1 Филогения облиственностебельных гилий в Калифорнии [272].
Сплошными линиями показаны основные направления дивергенции; преры-
вистыми—виды и популяции гибридного происхождения. Все эти формы —
диплоиды, за исключением тетраплоидкого G. clivorum.
color и G. angelensis-, 2) группа G. millefoliata и родственные
формы; 3) группа G. capitata. Эти три группы видов отражают
главные ветви в филогении рассматриваемой секции (рис. 21.1).
Крайние члены группы — G. capitata и G. tricolor, противополож-
ные признаки которых были описаны в гл 4 (см. рис. 4.2 и 4.3).
Gilia capitata представляет собой сложный политипический
вид, состоящий из необычайно разнообразных и многочисленных
форм, которые распадаются на две крайние группы и гибридов
между ними, занимающих промежуточное положение. Крайние
формы G. capitata дивергировали в прошлом до видового уровня
и все еще сохраняют, подобно видам, способность оставаться
обособленными в некоторых зонах симпатрического контакта.
В других областях, однако, крайние формы связаны между со-
бой расами гибридного происхождения [266, 268].
Gilia achilleifolia по морфологическим и экологическим приз-
накам занимает промежуточное положение между определенны-
ми ветвями групп G. capitata и G. tricolor. Кроме того, у этого
вида наблюдается необычайно сильная изменчивость в направ-
лении обеих предполагаемых родительских групп. Некоторые из
Гл. 21. Гибридные комплексы 27$
таких вариантов соответствуют естественным гибридам, извест-
ным между этими двумя группами видов. Все имеющиеся дан-
ные свидетельствуют о гибридном происхождении G achilleifo-
lia, показанном на рис. 21.1 [274].
Цитогенетические и морфологические данные с достаточным
основанием позволяют предположить, что Gilia clivorum (2п=
= 36)—амфиплоид, происходящий от G. achilleifolia и G. mil-
lefoliata, как это показано на рис. 21.1 [271, 274].
Популяции, промежуточные по морфологическим признакам
между Gilia capitata и G. angelensis, встречаются в каньоне
Санта-Ана, на юге Калифорнии, а популяции, морфологически
промежуточные между G. capitata и G. millefoliata,— на мысе
А1ендосипо, на севере Калифорнии. Таксономический статус этих
популяций неясен, и на рис. 21.1 они просто зафиксированы как
таковые.
Рис. 21.1 позволяет понять, как в результате гибридизации
ветвящаяся филогенетическая схема превращается в сетчатую,
без изменения генетических систем у.форм гибридного происхож-
дения. На ней показаны также последовательные уровни гибри-
дизации: виды, которые сами возникли в результате гибридиза-
ции, становятся предками более позднего поколения видов и по-
пуляций гибридного происхождения.
Еще неизученные гомогамные комплексы, несомненно, суще-
ствуют у Pinas, Quercus и у многих других групп растений, в ко-
торых имеются диплоидные или гомоплоидные виды, иногда
гибридизирующие друг с другом. Филогенетический анализ дру-
гих примеров таких комплексов заслужива-ет внимания в качест-
ве предмета дальнейших исследований.
18-45
Часть шестая
Полиплоидия
22 Полиплоидия:
распространение и частота
Исторический очерк
Полиплоидией называют особое арифметическое соотношение
между числом хромосом у родственных организмов, различаю-
щихся по этому числу. В простейшем случае у полиплоидного
организма число хромосом вдвое больше, чем у какой-либо род-
ственной .формы. В других случаях число хромосом у полиплоида
увеличено не вдвое по сравнению с гаплоидным их числом
у одной из родственных форм, а в большее число раз, например
в 3 или 6 (Зх или 6х). Или, наконец, число хромосом у поли-
плоида равно сумме их чисел у двух родственных форм, обла-
дающих меньшим числом хромосом, как в ряде 2л=10, 12, 22.
Во всех этих случаях у полиплоидного организма наблюдается
«более высокое число хромосом в результате прибавления доба-
вочных целых хромосомных наборов, имеющихся у одного или
нескольких предковых организмов. Короче говоря, полиплои-
дия— это наличие у одного организма трех или большего числа
хромосомных наборов.
Явление полиплоидии было открыто в начале нашего века,
когда цитогенетика и цитотаксономия растений находились на
стадии первых ранних поисков. Главную роль в его открытии
сыграли Винклер [902], предложивший термин полиплоидия, и
Винге [898], частично объяснивший сущность этого явления.
Винклер [902] изучал вегетативные прививки и химеры у
Solatium. Он обнаружил, что из каллуса, развивающегося на
-стебле на поверхности разреза, может регенерировать новый
побег, причем из стеблей диплоидных S. nigrum развивались
таким образом тетраплоидные растения. Винклер предположил,
что такие тетраплоидные растения возникают в результате со-
матического удвоения в первоначально диплоидной ткани.
Винге [898] сравнивал родственные виды, принадлежащие к
одному и тому же роду, по числу хромосом. Он использовал ре-
зультаты собственных подсчетов хромосом у Chenopodium, а
также опубликованные ранее данные подсчетов Тахары [820] на
Chrysanthemum. При этом Винге обнаружил, что число хромо-
Гл 22. Полиплоидия распространение и частота 275
сом у разных видов образует упорядоченные ряды. У разных
видов Chenopodium 2п=18 и 36, а у разных видов Chrysanthe-
mum 2п=18, 36, 54, 72 и 90. Таким образом, у близких видов
число хромосом оказалось кратным некоторому общему основ-
ному числу.
Для объяснения этих наблюдений Винге [898J предложил
оказавшуюся крайне плодотворной гипотезу удвоения числа хро-
мосом у межвидовых гибридов. По его мнению, полиплоидный
ряд возникает в результате следующей последовательности со-
бытий. Межвидовая гибридизация дает начало гибридным рас-
тениям, которые стерильны вследствие нарушения у них конъ-
югации хромосом. Иногда у гибрида или у его потомков проис-
ходит спонтанное удвоение числа хромосом, и они сыновятся
плодовитыми Затем такие плодовитые полиплоидные растения
могут размножиться и превратиться в новый константный вид.
Таким образом, два диплоидных вида путем гибридизации мо-
гут дать начало новому тсграплоидному виду; в результате гиб-
ридизации между диплоидом и тетраплоидом может возникнуть
гексаплоидпый вид, а при гибридизации между другими форма-
ми — полиплоиды с более высокой плоидиостыо [898].
Гипотеза Винге вскоре была подтверждена экспериментально
путем искусственной межвидовой гибридизации у Nicoliana,
Raphanobrassica и Galeopsis [105. 427, 596]. Классические приме-
ры Raphanobrassica и Galeopsis были описаны в гл 19
Совершенно очевидно, что между явлениями полиплоидии, с
коюрыми имел дело Винклер и которыми занимались Винге и
его последователи, имеются существенные различия В первом
случае это внутривидовая негибридная полиплоидия, а во вто-
ром— полиплоидия, связанная с межвидовой гибридизацией
Полиплоидия первого типа возникает в результате умножения
одного и того же набора хромосом, а полиплоидия второго ти-
па— в результате удвоения структурно несходных хромосомных
наборов у межвидового гибрида Эти различия стали яснее бла-
годаря терминам автополиплоидия и аллополиплоидия, введен-
ным Кихарой и Оно [438]. Термин амфиплоидия— удобный си-
ноним термина аллополиплоидия [102].
Масштабы проблемы и ее суть
Изучение полиплоидии необычайно расширилось со времени
первых исследований, и это обусловило появление огромного
количества литературы Анализ плоидности играет важную роль
в таких разнообразных областях исследований, как цитогенети-
ка, физиология, селекция растений, цитотаксономия и биогеогра-
фия Совершенно очевидно, что рассмотреть здесь проблему по-
липлоидии в ее полном обьеме невозможно
is*
276 Часть шестая. Полиплоидия
Хорошие обзоры различных аспектов полиплоидии составили
Мюнтцинг [599, 600, 602], Дарлингтон [139, 148], Клаузен, Кек
и Хиси [102], Лёве А. и Лёве Д. [528, 532], Стеббинс [780, 787],
Тышлер [831], Ригер [684], Готтшалк [258] и Люис [505].
В этой книге основное внимание уделено эволюционным ас-
пектам полиплоидии у высших растений. Для того чтобы выпол-
нить эту задачу, необходимы лишь некоторые данные, относя-
щиеся к различным областям науки, в которых занимаются
полиплоидией. Поэтому мы изложим здесь выборочные цитотак-
сономические, цитогенетические и биогеографические данные.
При отборе этих данных мы руководствовались их важностью
для стоящего перед нами вопроса.
В ближайших разделах этой главы мы рассмотрим данные,
•которые показывают, что полиплоидия действительно играла
очень важную роль в эволюции высших растений, особенно по-
крытосеменных и папоротников. В пользу такого заключения сви-
детельствуют две группы фактов: значительная часть видов выс-
ших растений — полиплоиды; различные роды достигают высо-
ких уровней плоидности. Совершенно очевидно, что в связи с
этим возникает вопрос: чем объяснить наблюдаемые факты?
В чем причины такого успеха полиплоидного состояния в эволю-
ции растений? Мы попытаемся выяснить этот вопрос в после-
дующих главах..
Полиплоидные ряды
В целях иллюстрации мы перечислим несколько хорошо извест-
ных примеров полиплоидии у покрытосеменных. Примеры эти
«сгруппированы по типу полиплоидных рядов.
Простейший случай — род, секция или группа видов, в кото-
рую входят диплоиды и тетраплоиды. Классическими примера-
ми служат Dahlia, Gossypium и группа Nicotiana tabacum.
У нескольких диплоидных видов Dahlia 2п=32, тогда как са-
довая, или изменчивая, георгина (D. variabilis) — тетраплоид
(2п=64) (подробности и библиографию см. [151]). Многочис-
ленные дикорастущие виды Gossypium и выращиваемые в Ста-
ром Свете G. arboreum и G. herbaceum — диплоиды, тогда как
выращиваемые в Новом Свете хлопчатники, принадлежащие к
видам G. hirsutum и G. barbaden.se, — тетраплоиды. Число хро-
мосом равно у них соответственно 2п=26 и 2п = 52, а основное
число я=13 [391]. Группа Nicotiana tabacum (х=12) содержит
ряд диплоидных видов (2п=24), в том числе N. glutinosa, а так-
же тетраплоидный (2п=48) настоящий табак, N. tabacum [250].
Следующий тип — просто продолжение предыдущего. У чле-
нов полиплоидных рядов этого типа основное число хромосом
Тл 22 Полиплоидия: распространение и частота
277
Рис 22 1. Полиплоидные ряды у Chrysanthemum (х=-9) [602]
Первая метафаза мейоза в материнских клетках пыльцы Л — С mankinoi
(2х, и=9). Б — С. indicum (4х, п=18) В —С japonense (6х, л=27). Г —
С ornatum (8х, п=36) Д — С. yezoensis (Юле, л = 45). Е— С. pacificum (10я,
п=45).
увеличивается в четное число раз, т. е. равно 2х, 4х, 6х, 8х.
Примером такого ряда, достигающего гексаплоидного уровня,
служат виды рода Triticum с числом хромосом 2п=14, 28 и 42
при основном числе х=7. Более длинный полиплоидный ряд об-
наружен у Chrysanthemum (х=9), что уже отмечалось ранее.
В этом роде встречаются формы с числами хромосом, равными
2х, 4х, 6х, 8х и 10х (рис. 22.1), и даже более высокими. Еще
длиннее полиплоидный ряд у Poientilla (х = 7), содержащий фор-
мы с числами хромосом, кратными всем четным числам — от 2х
до 16х (фактические данные и библиографию см. [151]).
Полиплоидные ряды, в которые входят формы с числами хро-
мосом, кратными как четным, так и нечетным числам, можно
проиллюстрировать па примерах Rosa и Crepis. У диплоидных
роз 2п=14. В группе Rosa canina есть, кроме того, тетраплоид-
ные, пентаплоидные и гексаплоидные формы с числом хромо-
сом 2п = 28, 35 и 42 (см [151]). В группе Crepis occidentalis
(х=11) полиплоидный ряд длиннее: в него входят формы 2х,
Зх, 4х, 5х, 7х и 8х [36]. Нечетные полиплоидные виды в этих и
278
Часть шестая. Полиплоидия
других подобных группах размножаются бесполым или субсек-
суальными способами.
У дибазических полиплоидов число хромосом равно сумме
двух анеуплоидных диплоидных чисел [144]. Примером, относя-
щимся к роду Brassica, служит тетраплоидный вид В. napus
(2п=38), происходящий от В. oleracea (2п=18) и В. campestris
(2п=20) [102, 848]. Другой дибазический полиплоидный ряд об-
наружен в одной из групп рода Clarkia с гаметным числом хро-
мосом п=8, 9, 17 и 26. С. prostrata (2п=6х=52), вероятно, про-
изошел от С. davyi (2п=4х=34) и С. speciosa (2и=2х=18);
С. davyi в свою очередь — тетраплоидное производное двух
диплоидных видов с 2п=16 и 2п=18 соответственно [496].
Ядра полиплоидов содержат много дуплицированного гене-
тического материала, поэтому они часто могут выдержать поте-
рю одной или нескольких пар хромосом. В результате получа-
ются модифицированные полиплоидные ряды, приводящие в ко-
нечном итоге к тому, что Дарлингтон назвал полиплоидным
сбросом (polyploid drop). Такого рода измененный ряд обнару-
жен у Hesperis (Cruciferae), где у разных видов гаметное чис-
ло п=7, 14, 13 и 12 [144, 148, 151].
У папоротников встречаются полиплоидные ряды, в которых
основное число хромосом х=37, 41 или другим столь же высо-
ким числам. Например, в группе Poly podium vulgar е (х=37)
есть диплоидные, тетраплоидные и гексаплоидные формы с
2п = 74, 148 и 222. В основе другого полиплоидного ряда у Polys-
tichum лежит х=41. В таких случаях может оказаться, что ос-
новное число, выведенное из существующего полиплоидного ря-
да, на самом деле не соответствует истинному основному числу
в филогенетическом смысле. Существующие в настоящее время
диплоидные члены ряда представляют собой, возможно, давно
возникшие полиплоиды из какого-либо более раннего эволюцион-
ного цикла [554].
Аналогичные рассуждения справедливы и в отношении неко-
торых сравнительно высоких основных чисел, обнаруженных у
других покрытосеменных. В трибе Pomoideae1 сем. Rosaceae
основное число х=17. В других трибах этого семейства основ-
ные числа хромосом не выходят за пределы, обычные для дип-
лоидов: Rosoideae (х=7), Prunoideae (х=8) и Spiraeoideae
(х=9). На основе этих данных Дарлингтон высказал предполо-
жение, что триба Pomoideae представляет собой модифицирован-
ное тетраплоидное производное более примитивных представите-
лей Rosaceae с 7 парами хромосом [138]. Позднее Стеббинс
[780], исходя из морфологических и цитологических признаков,
предположил, что Pomoideae (х= 17)— это, вполне возможно, ал-
Правильнее Maloideae. — Прим. ред.
Га 22 Полиплоидия- распространение и частота
279
лотетраплоиды, возникшие в результате гибридизации Prtinoi-
deae (х = 8) и Spiraeoideae (х — 9)1.
Возможно, что число хромосом х=13 у Gossypium — вторич-
ное основное число, первоначальным основным числом у этого
рода было х = 7. Такое основное число характерно и для многих
других родов Malvaceae. От него в результате удвоения и поли-
плоидного сброса могли произойти состоящие из 13 пар хромо-
сомные наборы ныне существующих диплоидных видов Gossy-
pium [832].
Распространение полиплоидии
в разных таксономических группах
Полиплоидия широко распространена среди покрытосеменных.
Она часто встречается у представителей Rosaceae, Compositac,
Rubiaceae, Iridaceae, bramineae, и многих других семейств.
Определенно, хотя и относительно редко, она встречается в та-
ких семействах, как Caesalpinaceae, Passifloraceae и Polemonia-
сеае. В ряде других семейств, например у Fagaceae и Berberida-
сеае, полиплоидные формы существуют, но они редки.
Стеббинс [780] обратил внимание на межродовые различия
частоты полиплоидии в нескольких семействах покрытосемен-
ных. Так, в сем. Salicaceae полиплоидия обычна у Salix, но ред-
ко встречается у Populus. В сем Betulaceae полиплоидные фор-
мы обычны у Betula, но относительно редки у Alnus Полиплои-
дия часто встречается у Thalictrum, но редко — у Aquilegia (сем.
Ranunculaceae), часто — у Tulipa, но редко — у Liliutn (Lilia-
ceae) [780].
У голосеменных полиплоидия возникает редко и спорадиче-
ски [433, 831]. У гинкго и саговников она неизвестна. Сообща-
лось о тетраплоидин у Gnetum и Welwitschia, но в настоящее
время эти сообщения считаются сомнительными [159]. У Ephed-
ra, однако, тетраплоидия встречается часто.
Среди хвойных один из очень немногих подлинных естествен-
ных полиплоидов — это Sequoia sempervirens (Taxodiaceae)
(2п = 6х=66); близкий к нему род Metasequoia — диплоид
(2п = 22) Другой пример—Fitzroya cupressoides (Cupressaccae)
с 2п = 4л = 44 [326]. У Juniperus известны триплоидные и тетра-
плоидные культивары (см. гл. 34).
У Pteridophyta в отличие от голосеменных полиплоидия рас-
пространена очень широко и достигает самых высоких уровней,
1 Иной точки зрения придерживается В И Гладкова (см ее статью
«О происхождении подсехейства Maloideae, Ботанич. жури, 1972, 57, 42—49).
По мнению А. Кронквиста (A Cronquist, «Ап Intergrated System of Classifi-
cation of Flowering Plants», Columbia Univ. Press, NY, 1981), гипотеза Глад-
ковой ближе к истине — Прим. ред.
280 Часть шестая. Полиплоидия
известных в царстве растений. Большинство Polypodiaceae—вы-
сокие полиплоиды. Полиплоидия обнаружена у обоих родов Psi-
lotales — Psilotiim и Tmesipteris. Виды Equisetum гомоплоидны
с очень высоким числом хромосом (2п«216), что свидетельст-
вует о давно возникшей полиплоидии. Среди Lycopodiales высо-
кий уровень плоидности встречается у Lycopodium и Isoetes, но
Selaginella1 сохраняет низкое диплоидное число хромосом
(2п=18), Хорошие сводки данных по числу хромосом у Ptery-
dophyta составили Ментон [554] и Лёве и др. [533].
Среди Bryophyta полиплоидия очень часто встречается у
мхов, но довольно редко — у печеночников. Полиплоидия также
хорошо известна у зеленых водорослей и харовых, но необычна
у бурых и красных водорослей [505].
В царстве грибов и в царстве животных полиплоиды крайне
редки. Обзор данных по грибам содержится в работе Люиса
[505]. Обзоры по полиплоидии у животных см. Бунгенберг [75],
Уайт (893, 894] и Люис [505].
Среди беспозвоночных возможные случаи полиплоидии обна-
ружены у Lumbricius и других дождевых червей {75, 588]. Наи-
более хорошо известные примеры относятся к членистоногим:
Artemia salina (Crustacea), Trichoniscus elizabethae (Isopoda),
Otiorrhynchus spp. (Curculionidae, Coleoptera), Solenobia triquet-
rella (Psychidae, Lepidoptera) и Ochthiphila polystigtna (Cha-
maemyiidae, Diptera) [75, 517, 769].
Среди позвоночных полиплоидия обнаружена у некоторых
рыб (713, 714]. Что касается полиплоидии у других групп рыб,
например Salmonidae, то единого мнения по этому вопросу не
существует [713, 714, 888]. Несколько случаев полиплоидии из-
вестно у амфибий и ящериц, в частности у ящериц рода Chemi-
dophoras [63].
Таким образом, полиплоидия спорадически встречается в
царствах животных и грибов, но характерна она для царства
растений.
Диапазон хромосомных чисел
Число хромосом у покрытосеменных колеблется от 2п=4 до
2п«500. И то и другое крайние значения обнаружены у дву-
дольных: низкое число у Haplopappus gracilis (2п=4), а высо-
кое—у одного из видов Kalanchoe (2п«500) (40, 395]. Другой
высокий полиплоид среди двудольных — Buddleja colvilei, вид,
1 В современных системах эти три рода относят к трем самостоятельным
порядкам — Lycopodiales, Selaginellales и Isoetales. — Прим. ред.
Гл 22 Полиплоидия- распространение и частота
281
у которого 2n«30G (см. [151]). У однодольных1 число хромо-
сом колеблется от 2п=6 до 2п=226.
Часто!а распределения видов покрытосеменных с разным
гаплоидным числом хромосом показана на рис. 22 2 Приведен-
ные на этом рисунке кривые построены на основе выборки из
17 138 видов, перечисленных в «Хромосомном атласе» [149, 151].
Анализ этой выборки и сведение полученных данных в таблицы
было проведено д-ром Хауордом Латимером и мною в 1951 г.
В настоящее время эти таблицы можно было бы значительно
расширить за счет многочисленных новых подсчетов хромосом,
проведенных начиная с середины пятидесятых годов и собран-
ных воедино в сводках Федорова [210] и Мура [583]. Однако вы-
борка, по которой построены кривые на рис 22 2, достаючно
велика и репрезентативна, так что этот рисунок хорошо отра-
жает главные статистические особенности числа хромосом у
цветковых растений.
Как показывает рис. 22.2, существуют тысячи видов покры-
тосеменных с 14—15 и более парами хромосом. Более высокие
числа образуют почти непрерывный анеуплоидный ряд от п=14
до ц=85, это1 ряд тянется далее с многочисленными разрывами
до п= 154, а затем наблюдается скачок к п = 250. У небольшого
числа видов, как и можно было ожидать, обнаружены еще бо-
лее высокие хромосомные числа.
Наивысшее число хромосом в царстве растений характерно
для папоротников и близких к ним форм [521, 522, 533, 554]. Со-
матические клетки Tmesipteris tannensis содержат свыше 400
хромосом [554]. Еще выше число хромосом у Ophioglossum По-
липлоидный ряд у этого рода начинается с'2п = 240, характер-
ного для 7 видов У О vulgatum и О thermale 2ц=480; у О. azo-
ricum 2и=720 (рис.22 3). У О. nipponicum 2п = 960, а наивысшее
число хромосом, 2п=1260, обнаружено у О reticulatum (резуль-
таты некоторых новых подсчетов и ссылки на прежние публика-
ции по Ophioglossum см [521, 522])
Диапазон уровней плоидности
Обращаясь от обсуждения диапазона по хромосомным числам
к обсуждению диапазона по уровням плоидности, мы нередко
сталкиваемся с определенным осложнением. Хромосомы можно
сосчитать. Что же касается уровня плоидности, то во многих
группах растении его можно точно определить методом сравне-
1 Спет , о I, юдопъных найме ъ ’ее число хромосом (2/.’-=4) обнаружено
у Ztngena bieberslciniana (Цвсчев Н Н, Бопховских 3 В, Ботанич журн, 50,
9, 1965) и у Colpoduiht versicolor (Соколсвсхая А П , Пробатова Н С.,
Ьотанпч журн, 62, № 2 1977) —
282
Часть шестая. Полиплоидия
ния, однако во многих других группах его нельзя определить не-
посредственно, а можно лишь оценить, исходя из некоего гипо-
тетического основного числа [139, 832].
Если род состоит из видов с числом хромосом 2я=14, 28, 42
и 56, то нетрудно установить, что он содержит тетраплоиды, гек-
саплоиды и октоплоиды. Сомнения возникают в том случае, если
род содержит только формы с 2м=28 и 2я=56. Казалось бы,
носителей такого числа хромосом следует считать соответствен-
но диплоидами и тетраплоидами, но подобное заключение мо-
жет быть ошибочным; более вероятно, что это тетраплоиды и
окгоплоиды, но и такой вывод остается всего лишь гипотезой.
Гаплоидное число хромосом
Гл 22 Полиплоидия- распространение и частота 283
Тем не менее в качестве основы для определения уровня плоид-
ности или частоты полиплоидов в данной группе растений гипо-
теза, которая, возможно, верна, лучше, чем факт, который, воз-
можно, неверен.
По данным Ментон [554], вСо 13 видов Equisetum, принад-
лежащие к двум секциям этого рода, имеют 2гс»216. Решив
на основе одного этого факта, что у Equisetum полиплоидия от-
сутствует, мы допустили бы ошибку. На самом деле все виды
Equi'setum находятся, по-видимому, на одном и том же высоком
уровне плоидности. Надеяться на то, что когда-нибудь будет
установлено основное число хромосом у этого таксономически
изолированного и гомоплоидного рода, мы не можем и, следо-
вательно, не можем оценить этот уровень плоидности [554].
Каких уровней достигает полиплоидия у покрытосеменных?
Начнем рассмотрение этого вопроса с перечисления некоторых
родов покрытосеменных, содержащих высокие полиплоиды, у
которых основное число хромосом известно по непосредствен-
ным цитотаксономическим данным, и поэтому верхний уровень
плоидности можно определить с достаточной степенью достовер-
ности.
50- 40
40- В
Числб ВИДОВ
30-
Гаплоидное число хромосом
Рис. 22 2 Частота распределения 17 134 видов двудольных по гаплоидному
числу хромосом [282]
А и Б Распределение в диапазоне от п=2 до п=61 В и Г Распределение
в диапазоне от п=37 до и=120 Еще у четырех видов, которые на графиках
не показаны, число хромосом лежит в пределах от п=132 до п = 250 А и
В — однодольные, Б и Г — двудольные
Часть шестая. Полиплоидия
Рис 22 3 Материнская клетка споры Ophioglossum azoricum примерно с
360 бивалентами [522]
Полиплоидный ряд у Potentilla (х = 7) имеет диапазон от
2п=14 до 2n=112 Potentilla haematochroa, у которой 2гг=112,
представляет собой 16-плоид. Полиплоидный ряд Chrysanthe-
mum (х=9) содержит формы с числом хромосом, кратным всем
четным числам от 2х до 10х, а затем, после некоторого разры-
ва,— формы с такими высокими числами, как 2п=160 и 198.
Ch. lacustre, у которого 2п=198, по-видимому, представляет со-
бой 22-плоид. В одной из групп рода Senecio с основным числом
хромосом х=5 имеются виды, находящиеся на нижнем и сред-
нем уровнях плоидпости—от 2х до 16х. У S. roberti-friesii, от-
носящегося к этой группе, 2п«180, т. е. он представляет собой
примерно 36-плоид (фактические данные и библиографию см.
[1511).
По сообщению Хэйра и Бейценберга [328], у Роа litorosa из
Новой Зеландии 2п=263—265 Известно, что у Роа основное
число х=7. Следовательно, Р. litorosa — 38-плоид, у которого
Гл 22 Полиплоидия распространение и частота 285
произошел полиплоидный сброс Это наивысший уровень плоид-
носги, известный у однодольных [328].
Buddleja (Loganiaceac) и Kalanchoe (Crassulaceae) служат
примерами родов покрытосеменных с очень высоким числом хро-
мосом, у которых основное число, а следовательно, и уровень
плоидности можно определить лишь косвенным путем. В роде
Buddleja есть виды с числом хромосом 2%, 4х и 6х, если считать
основным числом х=19. В таком случае В colvilei с 2п^300 —
примерно 16-плоид. Возможно, однако, что х=19 у Buddleja и
близких к нем} родов представляет собой вторичное основное
число давнего тстраплоидного происхождения. Тогда В. colvilei
уже не 16-плоид, а 32-плоид. У представителей рода Kalanchoe
(Crassulaceae) преобладающее основное число х = 17. При этом
виды Kalanchoe с 2н^500 следует считать примерно 30-плоида-
ми с анеуплоидпыми отклонениями. Но если, что представляется
вероятным, х=17 получено от одной из старых тетраплоидных
форм, то приведенную оценку уровня плоидности следует
удвоить, Kalanchoe, по-видимому, достигает наивысшего уровня
плоидности — примерно 60% (фактические данные и библиогра-
фию см. [151]).
У папоротников и близких к ним групп фактические или из-
вестные основные числа как уже отмечалось, в общем высокие.
Бентон [554] приводит следующие основные числа для чешрех
родов Polypodiaceae- Asplenium (% = 36), Polypodium (x=37),
Dryopteris (x = 41), Polystichum (% = 41). В каждом из этих ро-
дов есть полиплоидный ряд с указанным основным числом.
Так, например, в группе Polypodium vulgare (х = 37) имеются
диплоиды, тетраплоиды и гсксаплоиды; у последних 2п = 222
Однако нынешние основные числа почти наверное соответствуют
полиплоидным числам далекого прошлого Исходные, или пер-
воначальные, основные числа у этих родов папоротников оста-
ются неизвестными Можно, однако, без большого риска допус-
тить, что современные полиплоидные папоротники находятся па
достаточно высоких уровнях плоидности [554].
Ophioglossum (Ophiog ossaceae) образует полиплоидный ряд,
начинающийся с 2х=2п = 240 и доходящий через 4,г, 6% и 8х до
более чем 10% у О reticulatum, у которого 2м = 1260 Было выска-
зано правдоподобное предположение, что у Ophioglossum исход-
ное, пли первоначальное, основное число %=15. Из эюго сле-
дует, что современные «диплоидные» виды — 16-плоиды, а
О reticulatum — 84-плоид [521, 613].
Леван [467] изучат жизнеспособность различных искусствен-
ных полиплоидных форм Phleum pratense (% = 7) Он установил,
что 11-плоидпыс формы тимофеевки, содержащие 77 хромосом,
жизнеспособны, тогда как у 12-плоидов с 84 хромосомами жиз-
неспособность сильно понижена В этом случае пороговое для
286
Часть шестая. Полиплоидия
жизнеспособности число хромосом лежит в пределах между 77
и 84. У растений с 12х и 13х митоз был нарушен; это позволяет
предположить, что течение митоза играет важную роль в опре-
делении этого порога {467].
Со сходными цитологическими и физиологическими затрудне-
ниями встречаются, вероятно, высокие полиплоиды, возникаю-
щие в природе. Но все же уровни полиплоидии, обнаруженные
в природе, значительно превосходят верхние пределы, дости-
гаемые в экспериментальных условиях. Несомненно, что высокие
полиплоиды, существующие в природе в настоящее время, — со-
храненная отбором выборка из древних линий, которые успеш-
но и постепенно преодолевали разнообразные физиологические
затруднения.
В большинстве случаев полиплоидии, известных у животных,
она остается на низких уровнях плоидности, лишь изредка дости-
гая умеренно высоких уровней. Так, полиплоидные формы у
Cnemidophorus и у других ящериц — триплоиды; у амфибий —
по большей части триплоиды и тетраплоиды, хотя у трех видов
известны гексаплоиды и октоплоиды [63]. У долгоносиков (Сиг-
culionidae) большинство полиплоидов также триплоиды, хотя
известно несколько тетра-, пента- и гексаплоидов. Такой же
диапазон уровней плоидности характерен для других семейств
насекомых, в которых встречаются полиплоиды [517].
Частота
Наша следующая задача состоит в том, чтобы оценить долю
полиплоидных видов среди высших растений. Дать точную оцен-
ку частоты полиплоидии так же трудно, как и оценить ее уров-
ни и по тем же причинам. Если, желая оставаться на твердой
почве фактических данных, вычислять процент полиплоидных
форм по данным цитотаксономического анализа полиплоидных
рядов, то мы неизбежно упустим из виду многочисленные древ-
ние полиплоидные виды и тем самым серьезно занизим частоту
полиплоидии. Единственный способ избежать такого искаже-
ния— принять некоторые допущения относительно исходных,
или первоначальных, основных чисел.
Среди ранних оценок частоты полиплоидии у покрытосемен-
ных укажем на оценки, приведенные Мюнтцингом, Дарлингто-
ном и Стеббинсом. Мюнтцинг [599, 600] и Дарлингтон [139]
привели численные данные, свидетельствующие о том, что при-
мерно половина видов покрытосеменных — полиплоиды. По
приближенной оценке Стеббинса [780], полиплоиды составляют
30—35% всех видов покрытосеменных.
Позднее я попытался подойти к этой проблеме, сопоставив
|имеющиеся данные по частоте распределения видов покрытосе-
Гл 22 Полиплоидия: распространение и частота 287
менных с различным числом хромосом (рис. 22.2) с правдопо-
добным и широко распространенным допущением о том, что для
этой группы первичное основное число х = 7, 8 и (или) 9 [282].
В таком случае гаплоидное число хромосом п=14 может слу-
жить удобным пограничным пунктом- виды с 14 парами хромо-
сом и более при всех статистических расчетах относят к поли-
плоидам, а виды с меньшим числом пар хромосом — к дипло-
идам.
Такой подход также нс свободен от ошибок, что связано с
осложнениями, создаваемыми анеуплоидией. Среди покрытосе-
менных имеется несколько тетраплоидных видов, у которых
2п=12, 10 или даже меньшему числу пар; эти виды происходят
от нисходящего анеуплоидного ряда Некоторые другие виды с
2п=14 или большим числом пар, напротив, представляют собой
диплоидные продукты восходящей анеуплоидии. Мы не знаем,
сколько видов в нашей выборке принадлежат к каждой из этих
категорий. Следует, однако, отметить, что эти два источника
ошибок компенсируют друг друга.
При помощи вычислений, произведенных описанным выше
методом, были получены следующие оценки относительной час-
тоты полиплоидных видов у покрытосеменных [282J-
Дзудоть^ые 43%
Однодольные 58%
Покрытосеменные в целом 47%
Недавно Голдблат [246] вновь вычислил частоту полиплоидов у
однодольных, воспользовавшись более полными данными, при-
веденными в сводках Федорова [210] и Мура [583], и получил
оценку 55% (вместо 58%).
Голдблат [246] считает, что я провожу пограничную линию
слишком высоко (между н=13 и 14) и поэтому моя оценка час-
тоты полиплоидии слишком занижена. На самом деле при пер-
вом анализе этого вопроса я провел эту границу ниже и получил
другие оценки, но публиковать их не стал. Если постулировать,
что формы, у коюрых п=13 или выше, — полиплоиды, то на ос-
нове тех же самых фактических данных получаются следующие
оценки относительной доли полиплоидных видов:
Двудольные 49%
Однодольные 60%
Покрытосеменные в целом 52%
Я не решился бы однако, провести границу ниже, чем на уровне
и=13, так как считаю, что восходящая анеуплоидия ”игра<ч
важную роль вплоть до п=12, а на уровнях п=13, 14 и т. д
встречается с довольно значительной частотой Быть может, не
288 Часть шестая. Полиплоидия
тинная частота полиплоидных видов у покрытосеменных нахо-
дится приблизительно посередине между двумя приведенными
выше оценками.
В отличие от покрытосеменных у хвойных только 1,5% поли-
плоидных видов, а у саговниковых не обнаружено ни одного по-
липлоидного вида [433].
Для Pteridophyta подходящей выборкой может служить пе-
речень хромосомных чисел, составленный Ментон [554]. В этом
перечне таксономические виды, содержащие формы двух или
нескольких уровней плоидности, рассматриваются для интере-
сующих нас здесь целей как два или несколько видов, а гибри-
ды в расчет не принимаются. Составленный таким образом пе-
речень содержит 103 вида, и в нем представлены все крупные
труппы Pteridophyta. Только пять из этих видов, в том числе три
вида Selaginella с 2п=18, — первичные диплоиды. Полученная
при этом оценка частоты полиплоидов у Pteridophyta равна 95%.
23 Типы полиплоидов
Разработка классификации
Между автополиплоидами и аллополиплоидами (или амфиплои-
дами) существует фундаментальное различие. Кихара и Оно
[438] первоначально определили это различие в общем виде:
авгополиплоидия в отличие от аллополиплоидии является ре-
зультатом удвоения одного и того же набора хромосом.
В первом издании «Современных успехов цитологии» Дар-
лингтон [138] сформулировал это несколько иначе: автополип-
лоид представляет собой полиплоид, возникший путем удвоепия
или сложения хромосомных наборов какой-либо структурной
гомозиготы; следовательно, это организм, содержащий три или
большее число наборов гомологичных хромосом В отличие от
этого аллогюлиплоид возникает в результате удвоения числа
хромосом у межвидового гибрида; следовательно, это полиплоид,
содержащий отдельные наборы негомологичпых хромосом |138].
Тем, кто начинал изучение этой проблемы, было ясно, что
между гомологичными и пегомологичными хромосомами, между
структурными гомозиготами и структурными гетерозиготами и
между гомозиготными индивидуумами и межвидовыми гибрида-
ми существует полная интерградация Таким образом, автопо-
липлоидия и аллополиплоидия, согласно приведенному выше
определению, — крайние звенья непрерывного ряда, члены кото-
рого постепенно переходят друг в друга. Во mhoihx случаях мо-
жет оказаться полезным дальнейшее подразделение этих проти-
воположных классов и выделение некоторых промежуточных ти-
пов.
Мюнтцинг [599], рассмотрев различные определения автопо-
липлоидов, предложил следующую концепцию. Автополиплоиды
содержат множественные хромосомные наборы, которые иден-
тичны в структурном отношении, но могут различаться по ген-
ному составу. В дальнейшем Клаузен, Кек и Хиси [102] выдели-
ли два основных типа автополиплоидов: 1) автополиплоиды,
образующиеся из плодовитых межрасовых гибридов, и 2) авто-
полиплоиды, образующиеся из обычных негибридных индиви-
дуумов
Дарлингтон [138] различает три основных типа аллотстраплои-
дии в зависимости о г степени структурной дифференциации
между входящими в их состав геномами Примерами этих трех
типов служат Raphanobrassica, Primula kewensis и Crepis rubra-
foetida
Наиболее важные данные относительно Raphanobrassica и
Primula kewensis были приведены в гл. 19, в которой читатель
найдет интересующие его подробности.
19—45
290
Часть шестая. Полиплоидия
В экстремальном случае, как у Raphanobrassica, структурные
различия между геномами очень велики, поэтому у диплоидного
гибрида хромосомы не конъюгируют, однако у тетраплоидов,
происходящих от этих гибридов, они образуют биваленты. Для
промежуточного состояния, примером которого служит Primula
kewensis, характерна несколько менее сильная структурная диф-
ференциация хромосом, так что они конъюгируют и образуют
биваленты и у диплоидного гибрида, и у его тетраплоидного
производного. Слабая структурная дифференциация между ро-
дительскими геномами, такая, как между Crepis rubra и С. foe-
tida, делает возможным образование бивалентов у диплоидного
гибрида и усиленное образование квадривалентов у происходя-
щего от него тетраплоида. Следовательно, аллополиплоиды это-
го третьего типа по своим цитогенетическим признакам прибли-
жаются к автополипиоидам [138].
Последующие авторы продолжали разработку этой общей
темы. Клаузен, Кек и Хиси [102] классифицируют амфиплоиды
на основе двух взаимосвязанных признаков, а именно эффектив-
ности барьера стерильности между родительскими видами и
степени конъюгации между геномами родительских форм. По их
системе Raphanobrassica представляет собой интерценоспеци-
фичный амфиплоид с неконъюгирующими геномами; Primula
kewensis — интерэкоспецифичный амфиплоид с конъюгацией
между геномами. Между этими двумя типами существуют раз-
личные промежуточные комбинации. Для обозначения амфипло-
идов типа Р. kewensis и Raphanobrassica соответственно Стеб-
бинс [778, 780] ввел полезные термины сегментный аллополи-
плоид и геномный аллополиплоид.
На гексаплоидном, октоплоидном или более высоких уров-
нях плоидности растение может быть одновременно и авто- и
аллополиплоидом. Полиплоиды такого типа были удачно назва-
ны автоаллополиплоидами. Они возникают от гибридов между
двумя родительскими формами, принадлежащими к разным ви-
дам и имеющими различные геномы, одна из которых или обе —
автополиплоиды. Так, в результате гибридизации между авто-
тетраплоидом и отличающимся от него по геному диплоидом об-
разуется триплоидный гибрид, который при удвоении дает гек-
саплоид, содержащий четыре генома одного вида и два генома
другого вида.
В заключение этого раздела перечислим главные типы поли-
плоидов с указанием их генетической конституции. В соответст-
вии с принятой практикой каждый геном обозначен одной из
букв алфавита. Между геномами Л, В и т. д. существуют боль-
шие структурные различия, поэтому у межвидовых гибридов
конъюгация хромосом обычно бывает нарушена. Геномы или
субгеномы Л3, At и т. д. по своей структурной дифференциации
Гл 23 Типы полиплоидов
291
занимают промежуточное положение, вследствие чего как у ди-
плоидного гибрида, так и у его амфиплоидного производного
наблюдается частичная или полная конъюгация хромосом. Ди-
плоидные виды, которые самостоятельно или в комбинации друг
с другом образуют разного рода полиплоиды, имеют геномы АА,
AtAt, ВВ и т. д.
I. Автополиплоиды
1. Истинный автополиплоид АААА
2. Межрасовый автополиплоид АААА
II. Амфиплоиды
3. Сегментный аллополиплоид A&ASA/At
4 Геномный аллополиплоид ААВВ
5. Автоаллополиплоид ААААВВ
Обратите внимание па то, что типы 2, 3, 4 и 5 являются гиб-
ридными полиплоидами и только тип 1 негибридного происхож-
дения
Цитологические критерии
Основными критериями, по которым различаются автополиплои-
ды и амфиплоиды, служат поведение хромосом, плодовитость,
соотношения при расщеплении и морфология. В отдельных слу-
чаях все эти критерии могут оказаться несостоятельными. Не-
сомненные амфиплоиды часто проявляют цитогенетические свой-
ства автополиплоидов и наоборот
Наиболее очевидная особенность поведения хромосом, по ко-
торой, как следует ожидать, должны различаться автополиплои-
ды и амфиплоиды, — это наличие или отсутствие мультивален-
юв при мейозе.
Образование мультивалснтов ограничивается, однако, часто-
той хиазм, которая в свою очередь зависит от величины хромо-
сом и генетических условий. Вследствие этих ограничивающих
факторов у автотетраплоидов число квадривалентов почти всег-
да бывает ниже максимально возможного. Так, у автотетрапло-
идной ржи (Secale cereale, х = 7) образуется от 0 до 6 квадрива-
лентов на клетку, тогда как максимальное их число равно 7 IV,
а средняя частота квадривалентов варьирует от 2 до 4 IV у раз-
ных линий [397, 604].
Вместе с тем у сегментных аллополиплоидов возможно и
часто наблюдается образование некоторого числа мультивален-
тов. Например, тетраплоид Primula kewensis, как уже говори-
лось выше, обычно образует биваленты, но время от времени у
него появляются квадриваленты. Возникновения квадривален-
тов у этого и других сегментных аллополиплоидов следует ожи-
дать, поскольку между хромосомами, принадлежащими к раз-
личным субгеномам, существует частичная гомология.
19*
2?2
Часть шестая. Полиплоидия
Если эти гомеологичные хромосомы могут конъюгировать у
диплоидного гибрида, то они могут иногда конъюгировать и у
его амфиплоидного производного. Иными словами, избиратель-
ная конъюгация может оказаться не вполне эффективной. Такая
межгеномная (или так называемая гетерогенетическая) конъ-
югация может просто привести к образованию бивалентов. Но
если гетерогенетическая конъюгация сочетается с нормальной
гомогенетической конъюгацией с участием тех же партнеров, то
образуются ассоциации более высокого порядка — триваленты
и квадриваленты. У сегментного аллотетраплоида, имеющего ге-
ном AsAsAtAt, четыре хромосомы, принадлежащие к любому
данному хромосомному типу I, могут время от времени конъюги-
ровать, образуя квадривалент 1А3-1Ав-1А/-1А#.
Генетические критерии
Что касается плодовитости, то Дарлингтон [138] сформулировал
следующее общее правило* между плодовитостью диплоидного
гибрида и плодовитостью его тетраплоидного производного су-
ществует отрицательная корреляция. Плодовитые гибриды дают
стерильных тетраплоидов, стерильные гибриды — плодовитых
тетраплоидов, а гибриды, обладающие промежуточной плодо-
витостью, дают тетраплоидов с промежуточной плодовитостью.
В таком случае автополиплоидов можно отличить от амфиплои-
дов по стерильности первых и плодовитости вторых.
Это обобщение логически вытекает из ожидаемых различий
в поведении хромосом между автополиплоидами и амфиплоида-
ми, о чем говорилось в предыдущих разделах. У автополиплоида
неравномерное расщепление по гомологичным хромосомам, по-
лученным от мультивалента, ведет к стерильности гамет. В про-
тивоположность этому у амфиплоидов гомологичные хромосомы,
полученные от бивалентов, расщепляются равномерно, в резуль-
тате чего образуются фертильные гаметы.
Мы видели, однако, что на самом деле поведение хромосом
у полиплоидов нередко отклоняется от ожидаемого. Автополи-
плоиды, у которых конъюгация бивалентов генетически детерми-
нирована, могут быть высокоплодовитыми, а амфиплоиды могут
быть стерильными по разным причинам. У амфиплоида, обра-
зующего мультиваленты, может происходить неравномерное рас-
щепление хромосом, как у типичного автополиплоида, что при-
водит к формированию несбалансированных и нежизнеспособных
гамет. Или же у амфиплоида наблюдается нормальная — в цито-
логическом отношении — конъюгация, но образующиеся бива-
ленты гетерогенетичпы; иными словами, конъюгировавшие хро-
мосомы расщепляются равномерно в количественном отношении,
Гл. 23. Типы полиплоидов
293
но образуют генетически несбалансированные продукты реком-
бинации. Амфиплоид снова дает стерильные гаметы. Или. на-
конец, генетическая стерильность может сочетаться у диплоид-
ного гибрида с хромосомной стерильностью и передаваться его
амфиплоидным потомкам.
Экспериментальные данные по плодовитости полиплоидов
разных типов согласуются с реальным положением вещей. Об-
щее правило о наличии отрицательной корреляции по плодови-
тости справедливо во многих случаях, однако из него имеются
также многочисленные исключения. Несомненно, известно много
примеров автотетраплоидов с пониженной плодовитостью и
аллотетраплоидов с повышенной плодовитостью Но существует
также и много противоположных примеров.
Сирс [716] создал ряд диплоидных гибридов и производных
аллотетраплоидов между десятью диплоидными видами Triti-
сшп, Aegilops и Haynaldia (п = 7) В 18 гибридных комбинациях
ему удалось сравнить соответствующие Fi и аллотетраплоиды в
отношении конъюгации хромосом и плодовитости. У разных
диплоидных гибридов уровень образования бивалентов варьиро-
вал от высокого до низкого. У аллотетраплоидов плодовитость
колебалась от 100 до 0% Однако никакой постоянной зависимо-
сти между степенью конъюгации у диплоидов Ff и степенью
плодовитости их тетраплоидных производных обнаружено не бы-
ло Гибриды Fi с низким уровнем конъюгации в некоторых слу-
чаях производят плодовитых амфпплоидов, а в других — сте-
рильных в зависимое!и от прочих факторов [716].
Ожидаемое различие между автополиплоидами и а.мфиплои-
дами по поведению при скрещиваниях также выводится из раз-
личных экстремальных типов поведения хромосом, описанных
выше У любого автотетраплоида следует ожидать тстрасомною
наследования тех признаков, по которым он гетерозиготен, тогда
как амфиплоид должен сохранять постоянство и размножаться
в чистоте.
Теоретически выделенные для полиплоидов соотношения при
расщеплении сильно отличаются от соотношений у диплоидов
Рассмотрим гетерозиготный автотстраплоид с генетической кон-
ституцией TTtt по гену Т, локализованному вблизи центромеры
в четырех хромосомах которые распределяются по гаметам слу-
чайным образом. Следует ожидать, что соотношение зигот в F2
от этой гетсрозиготы составит 17777:34 гетерозиготы • Itttt
Если ген 7 находится на расстоянии 50 единиц хромосомной
карты от центромеры, то теоретическое соотношение зигот в F2
должно быть 17777*19,8 гетерозиготы * \tttt Соотношения, на-
блюдаемые в экспериментах, лежат между этими двумя теорети-
ческими оценками [511, 580]. В любом случае частота класса
гомозигот по рецессивному гену в F2 гораздо ниже при тетра-
*294
Часть шестая. Полиплоидия
сомном наследовании (между 1/22 и 1/36), чем при дисомном
наследовании (1/4).
В отличие от автополиплоидов у аллотетр апл оида, несущего
гены ТТ в одной паре хромосом в геноме А и гены tt в другой
паре хромосом в геноме В, вообще не следует ожидать расщеп-
ления по гену Т.
Тетрасомное наследование, однако, не всегда происходит у
всех автотетраплоидов и не обязательно отсутствует у алло-
тетраплоидов. Оно может наблюдаться у сегментного аллотетра-
плоида в случае образования у него мультивалентов. В свою
очередь у автотетраплоида, в особенности если он образовался
давно, могла произойти генетическая дивергенция, в результате
которой прежде гомологичные аллели превратились в отдельные
пары дупликатных генов.
У амфиплоида может происходить некоторое расщепление,
но не в таких соотношениях, как при тетрасомии. Допустим, что
у нас есть сегментный аллотетраплоид, несущий пару алле-
лей ТТ в одной паре хромосом одного субгенома и пару аллелей
tt в другом субгеноме. Такой сегментный аллотетраплоид обычно
образует гаметы Tt и зиготы TTtt. Однако иногда у него образу-
ются гетерогенетические пары (T/f и T/t) с последующим неза-
висимым распределением аллелей t't в результате возникает не-
которое число гамет tt, которые, объединяясь, дают некоторое
число зигот tttt. Таким образом, сегментный аллотетраплоид из*
редка дает выщепенцев, соответствующих диплоидной родитель-
ской форме tt.
Герстель и Филлипс [240] провели ряд остроумных экспери-
ментов с синтетическими амфиплоидами Gossypium и Nicotiana,
которые дали им возможность установить корреляцию между
соотношениями при расщеплении и близостью геномов. У неко-
торых амфиплоидов, происходящих от видов, не связанных близ-
ким родством, расщепления не наблюдается. У других амфипло-
идов, происходящих от близкородственных видов, как, например,
Gossypium arboreum и G. herbaceum, наблюдается расщепление
в соотношениях, характерных для тетрасомиков. Третьи амфи-
плоиды, различающиеся не полностью, но в достаточной степени,
изредка расщепляются по некоторым генам-маркерам [240].
Морфологические критерии
Часто считают, что по морфологическим признакам автополи-
плоид должен походить на диплоидный родительский вид, тогда
как амфиплоид должен быть сходен с межвидовым гибридом.
Однако экспериментальные автополиплоиды не всегда сходны с
диплоидными родительскими видами. Ясно, кроме того, что если
какой-либо сегментный аллополиплоид может расщепляться в
Гл. 23. Типы полиплоидов
295
Таблица 23 1. Амфиплоиды, которых путали с одной или с
обеими родительскими формами [102]
Амфиплоид Принимали за
Spartina townsendli Phleum pratense Роа annua Iris versicolor Eriogonum fasciculatum ssp. foliolo- sum Rumex acetosella Brassica napus Galeopsis tetrahit Penstemon neotericus Madia citrigracilis Artemisia douglasiana S. alterniflora и S stricta P nodosum P. exilis и P. supina I virginica E f typicum и E. f. polifolium R angiocarpus и R tenuifolius В olei acea и В. campestris G speciosa и G. pubescens P laeius и P. azureus M gracilis A suksdorfii и A. ludoviciana
направлении одного диплоидного родительского вида по одному
гену, то он может таким же образом расщепляться и по другим
генам и в конечном счете окажется очень сходным с диплоид-
ным видом как морфологически, так и экологически. Автоалло-
полиплоиды также часто проявляют морфологическое сходство
с определенным диплоидным видом.
У природных амфиплоидных видов и в самом деле обычно
наблюдается изменчивость в сторону одного или обоих диплоид-
ных родительских видов. Клаузен, Кек и Хиси в 1945 г. [102]
составили список амфиплоидов, которых систематики ошибочно
принимали за одну из родительских форм или как за одну, так
и за другую; мы приводим этот список в табл. 23.1. Позднее
было обнаружено много других аналогичных примеров, и мож-
но считать, что изменчивость подобного рода достаточно обычна
(см. [283]). Это показывает, что сильное морфологическое сход-
ство между данным полиплоидом и данным диплоидом не мо-
жет служить надежным критерием автополиплоидии
Относительная частота автополиплоидов
и амфиплоидов в природе
Определить, что данный природный полиплоид представляет
собой автополиплоид или амфиплоид,— отнюдь не легкая зада-
ча, поскольку различные критерии, которыми мы располагаем,
в пограничных случаях оказываются ненадежными. Сегментные
аллополиплоиды и автоаллополиплоиды по морфологическим
признакам и по поведению хромосом могут быть похожи на ав-
тополиплоидов. Даже после проведения значительного числа
296
Часть шестая. Полиплоидия
экспериментов и цитогенетических исследований не всегда уда-
ется дать окончательный ответ на этот вопрос. Тем не менее
проблему эту не следует считать неразрешимой. Если при изу-
чении данного полиплоида принять во внимание все критерии
и ресинтезировать его экспериментально из его предполагаемых
родительских форм, то можно определить природу и происхож-
дение этого полиплоида достаточно удовлетворительно.
Сейчас, оглядываясь назад, можно сказать, что в первых
работах по естественной полиплоидии проблему эту чрезмерно
упрощали. И Дарлингтон [138, 139], и Мюнтцинг [599] считали,
что автополиплоиды часто встречаются в природе и играют важ-
ную роль в эволюции растений. Такое заключение было основа-
но главным образом на широком распространении у многих так-
сономических видов растений морфологически сходных диплоид-
ных и полиплоидных форм [138, 139, 599]. В его пользу свиде-
тельствовало также образование мультивалентов у многих из
тех же полиплоидных форм [599]. Однако в настоящее время эти
две группы данных нельзя рассматривать как достаточно веские
аргументы в пользу подобного заключения.
Клаузен, Кек и Хиси [102] опубликовали критический обзор
данных примерно по 28 тщательно изученным группам расте-
ний, содержащим природные полиплоиды. Здесь мы изложим их
заключения [102, гл. 6—8] в обобщенном и упрощенном виде.
Из 28 примеров природных полиплоидов только три признаны
несомненными и три других — возможными автополиплоидами.
Несомненные автополиплоиды — это Galax aphylla, Biscutella
laevigata и Zea peretinis; возможные автополиплоиды — Vacci-
nium uliginosutn, Eragrostis pallescens, Galium mollugo и G. ve-
rum. Таким образом, преобладающее большинство природных
полиплоидов в этой выборке хорошо изученных и подвергнутых
повторному критическому анализу случаев являются амфиплои-
дами [102].
Позднее Стеббинс [778] еще раз проанализировал все эти
примеры и перевел некоторые из возможных случаев в категорию
сомнительных. Единственным несомненным автополиплоидом
Стеббинс считает Galax aphylla.
По данным Ментон [554], у папоротников и близких к ним
групп широко распространены гибридизация и амфиплоидия.
Однако, по ее мнению, естественная автополиплоидия у Pterido-
phyta отсутствует, за одним возможным исключением — Psilo-
tum.
Справедливости ради следует добавить, что некоторые из бо-
лее поздних авторов все еще продолжают считать, что автопо-
липлоидия играет или могла бы играть более важную роль в
природе, чем это вытекает из рассмотренных выше исследований
[143, 146, 543]. За последние годы перечень известных амфипло-
Гл. 23. Типы полиплоидов 297
ядов сильно увеличился» тогда как список возможных природ-
ных автополиплоидов увеличился лишь незначительно.
Наиболее обоснованный вывод, который можно сделать из
имеющихся фактических данных по сосудистым растениям, со-
стоит, в том, что амфиплоидия встречается гораздо чаще и рас-
пространена гораздо шире, чем автополиплоидия. Кроме того,
из немногочисленных твердо установленных природных автопо-
липлоидов по крайней мере некоторые представляют собой меж-
расовых автоплоидов, как, например, Biscutella laevigata и So-
lanum tuberosum. Таким образом, в полиплоидии у растений
тем или иным способом бывает обычно замешана гибридизация.
24 Факторы, способствующие
возникновению полиплоидии
Вопрос о том, почему полиплоидия играет столь важную роль
в эволюции растений, очень сложен. Его многочисленные и раз-
нообразные аспекты широко обсуждались многими авторами.
Были попытки установить корреляции между полиплоидией и
климатом, географической широтой, высотой над уровнем мо-
ря, типом местообитания, жизненной формой, системой размно-
жения, гибридным происхождением, величиной клеток, величи-
ной хромосом, их структурой, характером и поведением половых
хромосом, генотипом и другими факторами. Некоторые из этих
факторов рассматриваются в настоящей главе.
По-видимому, не существует какой-то одной причины, опре-
деляющей эволюционный успех полиплоидии. Вместе с тем раз-
личные факторы, способствующие полиплоидии, не равноценны
по своему значению.
В предпринятых мною попытках дать здесь анализ явления
полиплоидии в эволюционном плане оказалось полезным раз-
делить регулирующие его факторы на три категории: 1) перво-
степенные, или фундаментальные, факторы; 2) второстепенные,
или факторы, играющие значительную роль; 3) третьестепенные,
или относительно маловажные факторы. Именно в этом поряд-
ке в последующих разделах этой главы рассматриваются усло-
вия, благоприятствующие полиплоидии. Такой подход к пробле-
ме позволяет сформулировать ряд объяснений.
Преимущества, внутренне присущие
амфиплоидному состоянию
Хорошо известно, что гибриды между вполне дифференцирован-
ными расами или видами часто обладают повышенной мощ-
ностью или жизнеспособностью или же повышенным физиологи-
ческим гомеостазом. Генетические механизмы, лежащие в основе
гетерозиса, усиленно изучались и обсуждались. Предполага-
лось, что у гибридов диапазон ферментативной активности ши-
ре, чем у их родительских форм, и что они более разнообразны
в биохимическом отношении. Нас здесь интересуют, однако, не
генетические причины, а фенотипические результаты гибридпо-
сти. Гетерозис и физиологический гомеостаз — признаки, кото-
рые можно наблюдать у многих гибридов в царствах животных
и грибов, а также и в растительном царстве. Гетерозисные и го-
меостатические свойства многих гибридов, по всей вероятности,
имеют адаптивную ценность для своих носителей.
Г л 24 Факторы, способствующие возникновению полиплоидии 299
При нормальном половом размножении гибриды не могут
воспроизводить в чистоте свое гетерозиготное состояние. Если
гибрид стерилен, то он вообще не способен к половому размно-
жению. Если же он плодовит, то в последующих половых поко-
лениях он утрачивает свои ценные свойства — гетерозис и го-
меостаз. Однако амфиплоидия представляет собой такую гене-
тическую систему, которая позволяет выйти из тупика [138].
Амфиплоид восстанавливает утраченную плодовитость и может
размножаться половым путем, сохраняя способность воспроиз-
водить в чистоте свой высокогетерозиготный генотип. Он обла-
дает преимуществами гибридности в сочетании с преимущест-
вами, которые дает размножение в чистоте.
Поэтому в тех случаях, когда данный природный гибрид об-
ладает гетерозисными и гомеостатическими свойствами, амфи-
плоидия дает возможность воспроизводить эти ценные в адап-
тивном отношении свойства в ряду последующих половых поко-
лений. Кроме того, амфиплоидия не препятствует полностью
расщеплению и рекомбинации, если не считать случаев крайне
резко выраженных геномных аллополиплоидов, но допускает
появление некоторых новых вариантов, создавая тем самым воз-
можность для возникновения более тонких адаптивных призна-
ков и их закрепления в последующих поколениях
Высокая частота полиплоидных видов у высших растений,
преобладающее большинство которых—амфиплоиды, а осталь-
ные— главным образом мсжрасовые автополиплоиды, свиде-
тельствует о том, что данная генетическая система успешно
справляется с воспроизведением адаптивных гибридных гено-
типов в рамках полового размножения.
Другая черта полиплоидов, которая может оказаться выгод-
ной во многих ситуациях, — это большая забуференность их ге-
нотипов по сравнению с диплоидами, обеспечиваемая наличием
многочисленных дупликаций Полиплоидный индивидуум лучше
забуферен в физиологическом отношении, чем соответствующий
диплоид И он забуферен лучше, чем диплоид в отношении раз-
множения, так как благодаря тетрасомному наследованию он
медленнее расходует имеющиеся у пего запасы изменчивости.
Такими буферными свойствами обладают как автополиплоиды,
так и амфиплоиды.
Способы образования
Существует два основных способа возникновения полиплоидов:
соматическое удвоение при митозе и отсутствие редукции хромо-
сом при мейозе [362].
300
Часть, шестая. Полиплоидия
При соматическом удвоении хромосомы во время митоза де-
лятся, но не расходятся к полюсам и оказываются все в одном
дочернем ядре. В результате соматическая клетка с числом хро-
мосом 2п дает начало дочерней клетке с числом хромосом 4п.
Это соматическое удвоение может затронуть некоторые вегета-
тивные ткани, не имеющие отношения к развитию цветков, на-
пример корневые клубеньки; такие случаи нас здесь не интере-
суют. Но если удвоение произойдет в зиготе или в зародыше,
находящемся на ранних стадиях развития, то оно приведет к
изменению всего растения. Или же оно может произойти в одной
из почек и, следовательно, оказать влияние на одну из ветвей
растения. Тетраплоид Primula kewensis, о котором мы уже го-
ворили в гл. 19, возник в результате соматического удвоения,
происшедшего у некоторых цветоносных ветвей диплоидного
гибридного растения.
Отсутствие редукции связано с тем, что при мейозе не про-
исходит образования клеточной стенки. Если это имеет место в
диплоидных материнских клетках пыльцы и материнских клет-
ках зародышевого мешка, то образуются диплоидные споры и
гаметы. В таком случае после слияния двух нередуцированных
гамет могут сразу образоваться тетраплоидные зиготы. Именно
так возникла Raphanobrassica, как было отмечено в гл. 19.
Иногда отсутствие редукции может затронуть лишь одну ли-
нию половых клеток, что приведет к развитию либо диплоидных
пыльцевых зерен, либо диплоидных яйцеклеток. В таких слу-
чаях образование нового тетраплоидного индивидуума происхо-
дит в два этапа. На первом этапе в результате слияния 2п- и
ln-гамет образуется Зп-зигота, а в следующем поколении — на
втором этапе — нередуцированная Зп-гамета, сливаясь с нор-
мальной ln-гаметой, дает тетраплоид. Искусственный аллотетра-
плоид Galeopsis tetrahit был получен именно в результате таких
последовательных этапов (см. гл. 19).
Менее обычный способ возникновения полиплоидии связан с
полиспермией, т. е. оплодотворением яйцеклетки двумя мужски-
ми ядрами. Этот процесс описан у Listera ovata и у некоторых
других орхидей (324].
Амфиплоиды обычно образуются из диплоидных гибридов,
как в случаях Primula kewensis и Raphanobrassica. Высказыва-
лись также предположения, что в некоторых группах, в частно-
сти у Zauschneria и Galium, амфиплоиды возникли в результате
гибридизации между автополиплоидами, принадлежащими к
разным видам [102]. Если эта интерпретация верна, то в подоб-
ных случаях сначала происходил процесс удвоения, а гибриди-
зация имела место позднее.
Гл 24 Факторы, способствующие возникновению полиплоидии
301
Полиплоидия и жизненная форма
Мюнтцинг [599] провел анализ числа хромосом у 48 родов тра-
вянистых растений, относящихся к покрытосеменным, среди ко-
торых были как многолетние, так и однолетние виды Он уста-
новил наличие корреляции между числом хромосом и характе-
ром роста Как правило, у многолетних видов данного рода чис-
ло хромосом выше, чем у однолетних его представителей. Во
многих случаях более высокое число хромосом у многолетних
видов служит явным свидетельством полиплоидного происхож-
дения. Так, в родах Helianthus, Euchlaezea, Sorghum и других
имеются многолетние полиплоиды и однолетние диплоиды [599].
Стеббинс [775] подтвердил заключения Мюнтцинга и допол-
нил его обзор, добавив к нему 202 рода двудольных Он соста-
вил таблицы числа хромосом для растений, относящихся к каж-
дому из трех классов жизненных форм, а именно к древесным
растениям, многолетним травянистым растениям и однолетним
или двулетним травянистым растениям. Высокая частота поли-
плоидных видов обнаружена у 52% родов травянистых и у 36%
древесных родов растений Таким образом тенденция к поли-
плоидии сильнее выражена у травянистых двудольных Если же
выборку травянистых родов разбить на подгруппы многолетних
и однолетних, то станет ясно, что полиплоидия чаще встречает-
ся у многолетних родов [775].
Густафсон также установил, что на севере Европы частота по-
липлоидии статистически выше у многолетних Gramineae, Ra-
nunculaceae, Cruciferae, Leguminosae и Compositae, чем у одно-
летних представителей этих семейств [317]. Следует отметить,
что Pteridophyta, у которых Ментон [554] обнаружила такую
высокую частоту полиплоидии, — преимущественно многолетние
травянистые растения
Мюнтцинг [599] пытался объяснить корреляцию, наблюдае-
мую между полиплоидией и многолетним образом жизни, до-
пустив, что полиплоидпость оказывает влияние на жизненную
форму. Он указал, что у экспериментальных автополиплоидов
скорость роста в общем ниже, чем у диплоидных родительских
форм. Такое замедление роста могло бы во многих случаях
привести к превращению однолетнего растения в многолетнее
при удвоении числа хромосом [559, 654].
Более поздние авюры предпочли представить эту зависи-
мость, поменяв местами причину и следствие. По их мнению,
многолетний образ жизни с его возможностями для вегетатив-
ного размножения благоприятствует полиплоидии [143, 146, 317,
318, 775]. Веским доводом в пользу такой ишерпретации факти-
ческих данных служит корреляция, подмеченная Дарлингтоном
Один из о^ень немногих полиплоидов среди хвойных, Sequoia
302
Часть шестая. Полиплоидия
sempervirens, представляет собой также один из крайне редких
видов хвойных, способных регенерировать из корневых отпрыс-
ков [143].
Каким образом вегетативное размножение благоприятствует
полиплоидии? На этот вопрос разные авторы отвечают по-раз-
ному. По мнению Дарлингтона [146], вегетативное размножение
благоприятствует выживанию уже возникших полиплоидов. По
мнению же Густафсона [317], клональное размножение снижает
нормальную рекомбинацию, но облегчает функционирование
нередуцированных гамет.
Более простое и, по-видимому, более общее объяснение со-
стоит в том, что большая продолжительность жизни многолет-
них трав, а в особенности тех, которые способны к вегетативно-
му размножению, сильно повышает вероятность возникновения
соматического удвоения у гибридного растения [276, 775]. Мно-
голетний травянистый гибрид Primula kewensis в первый год
своего существования не образует фертильных тетраплоидных
ветвей, однако в последующие годы это наблюдалось в трех
отдельных случаях; следовательно, мы вправе допустить, что
этот гибрид, возможно, никогда не дал бы начала плодовитым
амфиплоидам, если бы продолжительность его жизни была ко-
роткой.
Факторы, имеющие первостепенное значение
Относительно природы и распространения полиплоидии сделано
три важных обобщения. Во-первых, полиплоидия представляв г
собой характерную особенность царства растений и только этого
царства, причем и здесь она встречается главным образом у
многолетних растений с большой продолжительностью жизни.
Во-вторых, полиплоидия у растений обычно носит форму амфи-
плоидии. И в-третьих, амфиплоидия представляет собой генети-
ческую систему, дающую возможность воспроизводить при по-
мощи полового размножения гибридный генотип, обладающий
ценными адаптивными свойствами. В сущности, это, вероятно,
наилучшая генетическая система для выполнения столь сложной
задачи. Теперь, руководствуясь этими обобщениями, можно
определить основные факторы, способствующие возникновению
полиплоидии.
Полиплоидии способствует сочетание трех основных факто-
ров: 1) большая продолжительность жизни и наличие возмож-
ностей к вегетативному размножению; 2) первичное видообразо-
вание, сопровождающееся хромосомными перестройками; 3) вы-
сокая частота естественной межвидовой гибридизации. В преды-
дущих главах книги эти три фактора рассматривались как
самостоятельные явления вне связи с полиплоидией.
Гл 24 Факторы, способствующие возникновению полиплоидии
303
Важно понимать, что полиплоидия, как весьма частое и ши-
роко распространенное эволюционное событие, зависит от сов-
местного действия всех трех факторов. Отсутствие любого из
этих факторов в данной группе организмов сильно задерживает
или полностью исключает формирование полиплоидных видов.
Второй и третий факторы играют особо важную роль. Первый
фактор важен, но не столь необходим; как мы увидим ниже, у
однолетних травянистых растений его отсутствие может компен-
сироваться другими второстепенными факторами.
Перечисленные три основных фактора особенно характерны
для высших растений И именно у высших растений часто воз-
никают полиплоидные виды, достигающие пышного развития.
В царстве животных, где полиплоидия — явление чрезвычайно
редкое, эти три фактора обычно отсутствуют или же встречают-
ся как исключения. Таким образом, как это указывает Густаф-
сон [317], глубокие различия, существующие между высшими
растениями и высшими животными, привели к глубоким разли-
чиям в характере их эволюции, и одно из этих различий — явле-
ние полиплоидии.
В царстве растений полиплоидия распространена неравно-
мерно, и во многих случаях ее наличие или отсутствие коррели-
рует с наличием или отсутствием одного из основных факторов.
В тех группах растений, у которых видообразование происхо-
дит по типу Ceanothus, полиплоидия в основном отсутствует.
В гл. 11 были описаны примеры, относящиеся к Quercus, Ribes,
Ceanothus и другим родам покрытосеменных. Кхошу [433] ука-
зывал на многочисленность неполиплоидных видов среди хвой-
ных. Все это — древесные растения с большой продолжитель-
ностью жизни отдельных индивидуумов, у которых часто наблю-
даются вегетативное размножение и естественная гибридизация.
Но близкие гибридизирующие виды не различаются по хромо-
сомным перестройкам |[33]. Короче говоря, факторы 1 и 3 нали-
цо, по фактор 2 отсутствует.
Мюнтцинг [599] показал, что частота полиплоидии определен-
но ниже у однолетних травянистых растений по сравнению с
многолетними, принадлежащими к тому же роду. Хейзер [367]
установил, что полиплоидия наблюдается относительно редко
среди сорной флоры штата Индиана, и показал, что это корре-
лирует с преобладанием среди процветающих сорняков расте-
ний с однолетним жизненным циклом. Густафсон [317, 318] так-
же обращает внимание на то, что среди однолетних растений
вообще и в сорняковой флоре, состоящей главным образом из
однолетних видов, в частности процент полиплоидных видов ни-
зок
Межд) однолетними травяниешми растениями, относящими-
ся к род\ Madia, существуют межвидовые различия по структу-
304
Часть шестая. Полиплоидия
ре хромосом-. Естественная гибридизация происходит у них час-
то, но индивидуальная продолжительность жизни невелика и
возможности для вегетативного размножения отсутствуют. Та-
ким образом, у них наличествуют факторы 2 и 3, но нет фак-
тора 1.
Широта и высота местности
Давно известно, что частота полиплоидии у высших растений
коррелирует с географической широтой. Процент полиплоидных
видов во флоре данной области закономерно возрастает в на-
правлении от низких широт к высоким. Такая тенденция впер-
вые была замечена Хагерупом [323] и Тышлером [830]. В ранних
данных по этому вопросу имелись разного рода ошибки, которые
были устранены в более поздних исследованиях, однако общие
выводы оставались прежними [833].
Новые исследования оказались также значительно более
обширными по сравнению с ранними обзорами. Региональные
флоры, упоминавшиеся в этих ранних работах, подверглись бо-
лее полному повторному изучению, а, кроме того, к их числу
добавились новые флоры. Оригинальные исследования слишком
многочисленны, чтобы их можно было здесь перечислить; обзо-
ры, содержащие обширные библиографии, опубликовали Лёве А.
и Лёве Д. [528, 530] и Ханельт [336]. Более новые данные под-
тверждают сделанные ранее обобщения.
В табл. 24.1 приведен процент полиплоидных покрытосемен-
ных во флорах ряда областей, лежащих от 36 до 84° с.ш. в
Евразии и Арктике. Данные взяты из работ Лёве А. и Лёве Д.
[530], а также Ханельта [336] и несколько обобщены; для выяс-
нения дальнейших подробностей читатель может обратиться к
этим обзорам. Все перечисление в табл. 24.1 региональные фло-
ры были подвергнуты исчерпывающему цитотаксономическому
изучению и расположены в порядке возрастания широты мест-
ности. Можно видеть, что частота полиплоидных видов возрас-
тает с 37% в Средиземноморской области до 74% на Шпицбер-
гене и что она вообще выше в арктической и субарктической
зонах [336, 524, 528, 530, 8331.
Одна из последних оценок частоты полиплоидных видов сре-
ди сосудистых растений арктического региона в целом состав-
ляет 60% в низких и 70% в высоких северных широтах [532].
На тихоокеанском побережье Северной Америки частота по-
липлоидов также увеличивается с возрастанием широты от Ка-
лифорнии до Аляски [387]. Имеется ряд данных, указывающих
на то, что подобная же тенденция к увеличению доли полиплои-
дов в высоких широтах наблюдается также в Южном полуша-
рии [325, 336].
Гл. 24. Факторы, способствующие возникновению полиплоидии
305
Таблица 24.1. Частота полиплоидных видов покрытосеменных в
разных широтных зонах в Евразии и Арктике (обобщено по
данным Лёве А. и Лёве Д. [530] и Ханельта [336])
Область Град с. ш. Полиплоиды, %
Сицилия 36—38 37,0
Румыния 44—47 46,8
Венгрия 46—49 48,6
Пардубице (ЧССР) 50 52,3
Центральная Европа 46—55 50,7
Шлезвиг-Гольштейн 54—55 54,5
Дания 54-58 53,5
Англия 50—61 52,8
Юго-западная часть Гренландии 60—62 74,0
Фарерские о ва 62 71,0
Исландия 63—-66 71,2
Швеция 55—69 56,9
Финляндия 60—70 57,3
Норвегия 58—71 57,6
Северо-западная часть Аляски 68 59,3
О-в Девон 75 76,0
Шпицбер! ен 77—81 74,0
Земля Франца-Иосифа 80—82 75,0
Земля Пири 82—84 85,9
Частота полиплоидных видов относительно велика в высо-
ких горах. По данным Ханельта [336], процент полиплоидов во
флорах различных высоких гор Евразии, Америки и Новой Гви-
неи составляет от 45 до 85. Иногда частота полиплоидов бывает
выше на высоких горах, чем на близлежащих низменностях, но
это случается не всегда [336].
Лёве А. и Лёве Д. [531] сравнивали частоту полиплоидии в
соседних альпийских и лесных флорах на г Вашингтон и в го-
рах Уайт-Маунтинс в шт. Нью-Гэмпшир. В альпийской зоне,
выше верхней границы лесной растительности, 63,6% таксонов
сосудистых растений — полиплоиды, тогда как среди соседней
лесной растительности полиплоиды составляют только 45%. Та-
ким образом, частота полиплоидии в альпийской зоне сущест-
венно выше [531].
Эти факты достаточно определенны. Но что они означают?
Им давали различные интерпретации. Чтобы показать все раз-
нообразие этих интерпретаций, я вкратце изложу их здесь.
Подробно они обсуждаются Лёве А. и Лёве Д. [528, 531], Стеб-
бинсом [780. 787], Джонсоном, Пэкером и Ризе [410], а также
Ханельтом [336.
1. В суровом холодном климате полиплоиды образуются под
действием температурных шоков [323].
20—45
306
Часть шестая. Полиплоидия
2. Полиплоиды лучше адаптированы к холодному климату
и к другим связанным с этим климатом условиям среды, чем
диплоиды {524, 528, 531].
3. Корреляция между полиплоидией и широтой местности
или высотой над уровнем моря носит косвенный характер. По-
липлоидия чаще всего встречается у многолетних травянистых
растений, размножающихся вегетативным путем, и эта жизнен-
ная форма преобладает в арктических и альпийских флорах
{317, 318].
4. Изменения климата в плейстоцене, в том числе продвиже-
ние и отступление ледяных щитов и ледников, вызывали изме-
нения в распространении существующих видов. Возникали но-
вые контакты, происходила естественная гибридизация и обра-
зовывались гибридные полиплоиды [554].
5. Изменения климата и рельефа местности в плейстоцене
способствовали естественной гибридизации в новых комбина-
циях, как было сказано выше. С отступлением льдов открылись
новые местообитания, которые можно было заселять. Амфиплои-
ды особенно хорошо подходят для заселения этих новых свобод-
ных местообитаний [192, 780, 787].
Первое из этих объяснений теперь уже считается несостоя-
тельным. Решающей стадией является не образование поли-
плоидов, а их закрепление в статистически значимых соотно-
шениях, выявляемое при сравнении флор в целом. Третье объяс-
нение также можно отбросить. Корреляция между полиплоидией
и географической широтой действует в пределах группы много-
летних травянистых растений в разных флорах [528].
' Все остальные объяснения достаточно правдоподобны, одна-
ко на основе тех статистических и флористических данных, ко-
торыми мы располагаем в настоящее время, им нельзя дать
окончательную оценку. Необходимы более подробные исследо-
вания.
Экологические факторы
Один из подходов к очерченной выше проблеме состоит в том,
чтобы изучить частоту полиплоидов в различных экологических
группах данной северной флоры.
Джонсон и Пэкер [409J собрали данные о частоте полиплоид-
ных покрытосеменных в разных частях одной из долин в север-
ной части Аляски на 68° с. ш. Частота полиплоидов в пределах
этой долины сильно различалась в разных местообитаниях, по-
вышаясь в нарушенных местообитаниях с влажной почвой и
неглубоким залеганием вечной мерзлоты и понижаясь в более
стабильных местообитаниях с более сухой почвой и более глу-
боким залеганием мерзлоты.
Гл 24. Факторы, способствующие возникновению полиплоидии
307
Эрендорфер [193] проанализировал различные растительные
сообщества во флоре низменных районов Австрии с точки зре-
ния частоты полиплоидов. Самый высокий процент полиплоидов
был обнаружен на лугах, недавно созданных в этой области
человеком, тогда как в лесных сообществах частота полиплои-
дов была ниже.
Другой подход к этой проблеме состоит в том, чтобы подроб-
но изучить географическое и экологическое распространение
родственных диплоидных и полиплоидных видов в отдельных
группах растений.
Как у покрытосеменных, так и у папоротников наблюдается
следующая общая картина* полиплоидные представители данной
группы имеют широкое распространение, тогда как их диплоид-
ные родичи распространены менее широко [554, 780]. У папорот-
ников, относящихся к родам Cystopteris, Polypodium, Dryopteris
и Asplenium, полиплоиды многочисленны, а диплоиды либо ред-
ки, либо представляют собой реликтовые или вымершие формы
[554]. Подобное же различие между диплоидами и полиплоида-
ми наблюдается в группе Biscutella laevigata (Cruciferac) [552,
553].
В комплекс Gilia inconspicua (Polemoniaceae) входят много-
численные диплоидные и тетраплоидные виды, произрастающие
в пустынях и горах западной части Северной Америки В этой
группе и географическое, и экологическое распространение дип-
лоидных видов в общем ограничено по сравнению с их амфи-
плоидными производными. Многие из диплоидных видов опреде-
ленно представляют собой реликш. В огличис от диплоидов
полиплоиды имеют широкое распространение и очень обильны
на обширных и разнообразных по своим условиям территориях
[283]. Тетраплоидные гилии при выращивании их на экспери-
ментальных участках отличаются также от своих диплоидных
родичей более высокой мощностью.
Данные, полученные при использовании описанных здесь
двух подходов, свидетельствуют в пользу объяснений 4 и 5.
Однако эти данные нельзя считать решающими, и проблема все
еще остается открытой.
По моему мнению, подтверждаемому результатами моих соб-
ственных исследований на нескольких группах полиплоидов и
диплоидов как в природе, так и па экспериментальных участ-
ках, амфиплоиды и в самом деле часто характеризуются более
высокой мощностью, гомеостатическими буферными свойствами
и способностью к адаптации по сравнению со своими диплоид-
ными родичами. Вследствие этого амфиплоиды в природе не-
редко обладают адаптивным превосходством над предковыми
диплоидными формами. Появление на территории предковых
видов новых пригодных для заселения местообитаний, возникаю-
20*
308
Часть шестая. Полиплоидия
щих в результате изменений климата, разрушений, причиняе-
мых деятельностью человека, или других нарушений среды, соз-
дает те экологические предпосылки, при которых амфиплоиды
могут воспользоваться своими преимуществами.
Полиплоидия и система размножения
Амфиплоиды у однолетних растений возникают главным обра-
зом в результате отсутствия редукции и последующего слияния
нередуцированных гамет [102]. Если однолетнее гибридное рас-
тение размножается при помощи ауткроссинга, то вероятность
соединения двух нередуцированных гамет очень невелика. Та-
кой гибрид может начать интрогрессию, но вряд ли он за свою
короткую жизнь сумеет произвести амфиплоида,- Однако если
однолетнее гибридное растение размножается при помощи авто-
гамии, то вероятность соединения двух нередуцированных гамет
значительно повышается [278].
Две однолетние секции рода Gilia —Gilia и Arachnion
(х=9) —насчитывают вместе 39 видов; все эти виды, за исклю-
чением одного, были изучены цитотаксономически и генетичес-
ки. Что касается плоидности, то среди 38 цитологически изучен-
ных видов обнаружено 23 диплоида, 14 тетраплоидов и 1 окто-
плоид [281, 283, 286]. В отношении размножения у этих же ви-
дов наблюдается весь диапазон систем — от облигатного
ауткроссинга до автогамного инбридинга. Весьма примечатель-
но, что все 15 полиплоидных видов автогамные [278]. У одного
в целом автогамного тетраплоидного вида имеется раса, размно-
жающаяся при помощи ауткроссинга.
На ряде примеров была прослежена родословная однолетних
амфиплоидных видов Gilia и установлено их происхождение от
определенных автогамных диплоидных видов. Оказалось, что от
автогамных диплоидных видов Gilia происходит относительно
больше амфиплоидов, чем от диплоидных Gilia с перекрестным
оплодотворением.
Такая же корреляция между автогамией и возникновением
полиплоидов обнаружена и у других родов, в секциях, состоя-
щих из однолетних видов. Большинство полиплоидных видов
среди однолетних представителей родов Galeopsis, Madia, Mic-
roseris, Clarkia, Mentzelia, Atnsinckia и Eschscholtzia — автогам-
ные формы [278]. Наблюдаемая корреляция согласуется с пред-
ставлением о том, что автогамия является одним из второсте-
пенных факторов, способствующих возникновению полиплоидии,
который в известной степени компенсирует короткую продолжи-
тельность жизни каждого индивидуального растения у однолет-
них видов.
Гл. 24 Факторы, способствующие возникновению полиплоидии 309
Многолетние травянистые растения, у которых полиплоидии
широко распространена, обычно размножаются при помощи
перекрестного опыления и часто бывают самонссовмсстимыми
Корреляцию между тремя факторами — характером роста, по-
липлоидией и системой размножения — можно в этом случае
объяснить несколькими разными способами
Густафсон [317] вслед за Люисом [489] счшает, что у само-
несовместимых диплоидов в столбике создаются условия, кото-
рые благоприятствуют функционированию нередуцированных
пыльцевых зерен, образуемых этим же самым растением; это
приводит к формированию тетраплоидных или по меньшей мере
триплоидных зигот По моему мнению, совершенно достаточно
более простое и более прямое объяснение. Многолетние гибрид-
ные растения располагают достаточным временем, а следова-
тельно, и большими возможностями, чтобы претерпеть полипло-
идное удвоение, обычно путем соматического удвоения, по иног-
да также путем соединения нередуцированных гамет. Таким об-
разом, по Густафсону, причинная связь сущее 1вует между само-
несовместимостью и полиплоидией, а многодетность — коррели-
рованный с ними фактор, тогда как, с моей точки зрения при-
чинная связь существует между многодетности) и полипло-
идией, а коррелированный фактор — свободное скрещивание.
На эту проблему можно взглянуть и под иным углом зрения
У растений, обладающих системой самонесовместимости гамето-
фитного типа или типа Nicotiana, полиплоидизация превращает
самонесовместимый диплоид в самосовместимый тетраплоид
[488 494] У Solarium, и Nicotiana (сом Solanaceae) и у некото-
рых представителей сем. Scrophulariaccac и Leguminosae дипло-
идные виды обычно самонесовместимы, а природные полиплоид-
ные виды самосовместимы [494].
Мёллер [594] высказал мнение, что установление полиплоид-
ного состояния должно быть особенно затруднено у раздельно-
полых организмов. Предполагалось, что главная причина этого
заключается в нарушении механизма определения пола у тетра-
плоидов с половыми хромосомами XXVY Меллер ]594] сделал
такое предположение, надеясь, что оно послужит объяснением
редкости полиплоидии у живошых, в большинстве своем раз-
дельнополых, и се широкого распространения у растений, кото-
рые по большей части гермафродитпы.
Полученные в дальнейшем фактические данные частично под-
тверждают, а частично противоречат гипотезе Меллера. Твер-
до установленные примеры полиплоидии у животных действи-
тельно обнаружены либо в группах, размножающихся партено-
генетичсски (Artemia salina, равноногие раки, чехлоноски, турец-
кий скосарь, Ochthiphila}, либо у гермафродитов (дождевые
черви) [75, 769]. Вместе с тем полиплоидия возникла и у некото-
310
Часть шестая. Полиплоидия
рых двудомных растений как в природе (Salix, Rumex, Galium),
так и в экспериментальных условиях (Melandrium) [161, 522,
780, 885]
Влияние величины хромосом
Дарлингтон давно высказал предположение, что механика де-
ления хромосом и механика клеточного деления, возможно, иг-
рают роль факторов, ограничивающих полиплоидию. Между ве-
личиной клеток, величиной хромосом и полиплоидным числом
хромосом существуют реципрокные зависимости [138].
Известно, что скорость роста у автополиплоидов часто бы-
вает ниже, чем у родственных им диплоидов; это обусловлено,
по крайней мере частично, задержкой клеточного деления [599].
На некотором уровне плоидности, различном для разных групп
растений, искусственный полиплоид переступает некий порог
жизнеспособности, который опять-таки частично определяется
особенностями митоза (см. гл. 22).
Клетки, образующие питательные ткани в зародышевых меш-
ках и пыльниках, часто или обычно содержат эндополиплоидные
ядра. Такая клетка — продукт многократных эндомитозов; она
не претерпевает клеточных делений. Эти эндополиплоидные
клетки достигают гораздо более высоких уровней плоидности,
чем растения в целом. У некоторых представителей Ranuncula-
ceae, Papaveraceae и Compositae находящиеся в зародышевом
мешке клетки-антиподы могут достигать 64-плоидности [602].
Наивысшая степень плоидности обнаружена у Phaseolus cocci-
neus (=Р. multiflorus), зародышевый мешок которых содержит
4096-плоидные ядра (147].
Отдельные клетки, которые не должны делиться, явно могут
функционировать при гораздо более высоких уровнях плоидно-
сти, чем целые растения, рост которых осуществляется путем
клеточных делений. Это различие между терминальными члена-
ми клеточных линий и растущими растительными организмами
в смысле толерантности к высоким уровням плоидности согла-
суется с гипотезой Дарлингтона.
Отсутствие или редкость полиплоидии у некоторых Liliaceae
и у некоторых древесных растений объясняются, возможно, тре-
бованиями, предъявляемыми клеточной механикой. У Lilium,
Fritillaria и у некоторых других родов лилейных с крупными
хромосомами полиплоиды либо не достигают высоких уровней
плоидности, либо отсутствуют вообще. Быть может, в таких слу-
чаях хромосомы просто слишком велики, так что при высокой
плоидности клеточное деление становится невозможным [138,
139].
Гл 24 Факторы, способствующие возникновению полиплоидии
311
У древесных двудольных камбиальные клетки узкие, и это,
по-видимому, ограничивает как число хромосом, так и их вели-
чину. Относительно низкая частота полиплоидии у древесных
двудольных по сравнению с многолетними травянистыми дву-
дольными и полное ее отсутствие в некоторых родах и семейст-
вах двудольных, возможно, обусловлены именно этой особен-
ностью камбиальных клеток [139, 775].
Влияние генотипа
Нарушение конъюгации хромосом у межвидового гибрида соз-
дает необходимые условия, делающие возможным отсутствие
редукции и образование амфиплоидов. Нарушение конъюга-
ции— результат различий между родительскими геномами по
хромосомным перестройкам. Однако, как показали многочислен-
ные исследования [139, 502, 670, 687, 868], поведение хромосом
во время мейоза регулируется также определенными генами.
Отсюда следует, что на частоту отсутствия редукции, а тем са-
мым и на возникновение полиплоидов у ряда гибридов с хромо-
сомной стерильностью, могут оказывать влияние генотипические
различия по тем или иным генам, контролирующим мейоз [286,
816, 817].
Гены, детерминирующие различные отклонения в мейозе, на*
пример асинапсис или формирование аномального веретена, из-
вестны у кукурузы, дрозофилы и других генетически хорошо изу-
ченных организмов. Нас здесь особенно интересуют те из генов,
регулирующих мейоз, которые вызывают нарушение второго
мейотического деления, в результате чего образуются нередуци-
рованные гаметы; такие гены были обнаружены у Datura stra-
monium и Zea mays [25, 679, 709].
У Datura stramonium имеется рецессивный мутантный ген,
известный под названием dyad, который вызывает образование
к концу мейоза диад нередуцироваппых спор как в мужской, так
и в женской линиях. Диплоидное растение Datura, гомозиготное
по гену dyad, при самооплодотворении дало начало некоторому
числу тетраплоидных потомков [25, 709]. Растения Zea mays,
несущие рецессивный аллель гена elongate в гомозиготном сос-
тоянии, дают некоторое число нередуцировапных яйцеклеток, по
нормальную гаплоидную пыльцу. Поэтому первое поколение по-
томков растений elongate состоит главным образом из трипло-
идов. В последующих поколениях эти триплоиды могут давать
тетраплоидов и более высоких полиплоидов [679].
У диплоидных гибридов Fi от скрещивания Rumex thyrstflo-
rus и R acetosa наблюдается индивидуальная изменчивость в
312
Часть шестая. Полиплоидия
отношении мейоза. У некоторых растений Ft мейоз протекает
нормально, и они образуют тетрады; у других второе мейоти-
ческое деление отсутствует, и они образуют диады нередуциро-
ванных гамет. Выщепление форм, образующих диады, происхо-
дит также в поколениях F2 и Вь Потомки F2 от гибридных инди-
видуумов с нормальным мейозом представляют собой диплоидов
(2п=14). Но потомки F2 от гибридных растений, образующих
диады, — тетраплоиды или почти тетраплоиды (2/г«25) [816',
817].
Восемь видов секции Gilia рода Gilia скрещивали искусст-
венно в различных комбинациях; было получено 15 гибридов,
достигших стадии цветения. У четырех из этих 15 типов межви-
довых гибридов произошло спонтанное удвоение числа хромо-
сом путем соединения нередуцированных гамет, в результате че-
го было получено плодовитое или полу плодовитое амфиплоид-
ное поколение F2. Все эти четыре гибрида, давшие амфиплоид-
ных потомков, имели в качестве одного или обоих родительских
видов Gilia miUefoliata и (или) G. valdiviensis. G. millefoliata и
G. valdiviensis — пва близкородственных диплоидных (2п=18)
вида, принадлежащих к одной и той же геномной группе. Таким
образом, выборка искусственных спонтанных амфиплоидов сек-
ции Gilia рода Gilia отличается необычно высокой концентра-
цией геномов millefoliata [286].
У одного из гибридов между Gilia millefoliata и G. achilleifo-
lia, выращиваемых на экспериментальных участках, заложенных
в трех повторностях, наблюдалось удвоение хромосом. В отли-
чие от этого на участках, которые также были заложены в не-
скольких повторностях, но на которых были высеяны гибриды,
не имевшие в качестве родительских форм ни G. millefoliata, ни
G. valdiviensis, амфиплоиды не появлялись [286]. В свете этих
данных представляется знаменательным, что из трех известных
природных тетраплоидных видов, относящихся к секции Gilia
рода Gilia, два несут геном millefoliata [286].
Левин <[470] обнаружил аналогичное явление у рода Phlox
в восточной части Северной Америки В этой области встреча-
ются пять аллотетраплоидных видов Phlox. Четыре из этих пяти
тетраплоидов несут геном диплоидного вида Phlox pilosa [470].
Таким образом, создается впечатление, что родительские ви-
ды генотипически различаются по способности производить ам-
фиплоидных потомков как в природе, так и на эксперименталь-
ных участках при более или менее одинаковых прочих условиях.
Возможно, что гены, входящие в состав различных геномов,
представляют собой один из третьестепенных факторов, воздей-
ствующих на распределение полиплоидов в пределах данного
рода или секции, в состав которых входят разные симпатричес-
кие виды.
Гл 24. Факторы, способствующие возникновению полиплоидии
313
Общие выводы
Полиплоидия представляет собой генетическую систему, часто
встречающуюся и широко распространенную среди высших рас-
тений. В разных таксономических группах сосудистых растений
полиплоидия распределена неравномерно, и распространение ее
зависит от Климата, жизненной формы растения, его системы
размножения, структуры хромосом, их величины и от других
факторов. Причины эволюционного успеха полиплоидии у выс-
ших растений кроются в этих разнообразных корреляциях, и их
можно выявить, проведя соответствующий анализ.
Огромное большинство встречающихся в природе полиплоид-
ных видов растений, подвергнутых генетико-таксономическому
анализу, оказались амфиплоидами. Амфиплоидия представляет
собой определенного типа перманентную гибридность. Эта гене-
тическая система дает возможность межвидовому гибриду пре-
одолеть барьер стерильности и воспроизводить в чистоте свою
гибридную конституцию. Кроме того, она позволяет растениям
использовать преимущества, присущие высокогетерозиготному
генотипу, в частности гетерозис и физиологический гомеостаз, в
многочисленных гибридных комбинациях, которые содержат эти
выгодные свойства.
Способность какой-либо группы растений использовать в
своем эволюционном развитии преимущества, предоставляемые
амфиплоидной генетической системой, зависит от разнообразных
факторов, различающихся также по своему значению. Одни фак-
торы оказывают влияние на возникновение нового полиплоида, а
другие — на последующее создание им природной популяции.
Как в возникновении, так и в закреплении нового полиплоида
участвуют разного рода внутренние и внешние факторы
Первостепенное и фундаментальное значение имеет сочета-
ние трех условий Первое — наличие диплоидных видов, несу-
щих различные геномы или субгеномы. Второе — естественная
гибридизация между этими видами Третье—большая продол-
жительность жизни многолетних растений, повышающая вероят-
ность соматического удвоения, или, в качестве частичной компен-
сации у короткоживущих однолетних видов, — автогамная си-
стема размножения, повышающая вероятность соединения нерс-
дуцироваппых гамет. В случае отсутствия у данной группы рас-
тений любого из этих трех условий следует ожидать, что поли-
плоидов в ней нет или они редки Изучение распространения по-
липлоидии в зависимости от хромосомных перестроек, гибрид-
ности и многолетнего образа жизни (или автогамии, заменяю-
щей его у однолетников) показало, что полиплоидия и в самом
деле часто встречается только в тех группах растений, которые
обладают этими признаками.
314 Часть шестая. Полиплоидия
Не во всех группах растений, отвечающих этим -трем глав-
ным условиям, полиплоидия распространена одинаково равно-
мерно или встречается одинаково часто. Неравномерное рас-
пространение полиплоидии среди разных групп растений, срав-
нимых в отношении факторов первостепенного значения, свиде-
тельствует о существовании второстепенных и третьестепенных
факторов.
Нарушение местообитания и стрессовые условия среды, часто
в холодном климате, принадлежат к числу внешних факторов
второстепенного значения, благоприятствующих возникновению
полиплоидии. К второстепенным внутренним факторам, препятст-
вующим возникновению полиплоидии, относятся в числе прочих
хромосомы, имеющие слишком крупные по сравнению с клеткой
размеры, и клетки, величина которых мала по сравнению с раз-
мерами и числом содержащихся в них хромосом. Наличие или
отсутствие генов, вызывающих нарушение второго деления мейо-
за, а следовательно обусловливающих образование нередуциро-
ванных гамет, — третьестепенный фактор, который, возможно,
позволяет объяснить относительно неравномерное участие раз-
личных диплоидных видов в создании амфиплоидов в некоторых
родах.
25 Полиплоидный комплекс
Нередко те самые факторы, которые способствуют образова-
нию одного нового тетраплоидного вида, способствуют также
возникновению других новых тетраплоидов из других диплоидов,
относящихся к этой же группе видов. Эти же факторы могут
затем привести к образованию тетраплоидами гексаплоидов и
октоплоидов. В результате такой непрерывной полиплоидизации
по разным ветвящимся направлениям получается полиплоидный
комплекс.
Одна из простейших возможных структур полиплоидного
комплекса представлена на рис. 25 1. Изображенный на этом
рисунке комплекс состоит из трех диплоидных видов, из двух
возможных аллотетраплоидпых производных и одного аллогек-
са плопда. Вместе они представляют собой шесть видов-двой-
ников, изолированных барьерами хромосомной стерильности, а
возможно, и другими, связанными с последними, барьерами;
однако эти виды содержат много одинакового генетического ма-
териала, и поэтому диапазоны изменчивости по морфологичес-
ким признакам и экологическим предпочтениям у них перекры-
ваются.
Другие возможные типы этого простого полиплоидного комп-
лекса могут состоять из автотетраплоидов и аллотетраплоидов,
автоаллогексаплоидов и геномных аилогексаплоидов или алло-
октоплоидов и гексаплоидов. При наличии достаточно обширного
ряда диплоидных видов в качестве основного фонда эволюция
путем полиплоидии может создать очень большой полиплоидный
комплекс, содержащий многочисленные тетраплоидные и гекса-
плоидные виды, а некоторые потомки этих последних достигают
в нем еще более высоких уровней плоидности
Методы анализа
Полиплоидный комплекс можно идентифицировать при помощи
цитотаксономических методов Филогенетические взаимоотно-
шения между входящими в него видами-двойпиками можно
установить при помощи цитогенетических и генетико-таксономи-
ческих методов. Филогенетический анализ полиплоидного комп-
лекса можно довести до любой степени полноты. Для наших
целей полезно разбить анализ полиплоидного комплекса на че-
тыре главные стадии.
Первая стадия — это идентификация полиплоидного комп-
лекса как такового. Перед нами большая и таксономически
трудная группа растительных видов Судя по морфологическим
316
Часть шестая. Полиплоидия
____I________।---------1________I_________[______
Диапазон изменчивости морфологических признаков
Рис, 25.1. Структура простого полиплоидного комплекса.
признакам и по географическому распространению, их можно
считать видами-двойниками, но таксономические границы этих
видов, их ареалы и генетическая природа неизвестны, поэтому
следует применить цитотаксономические методы. Проведение
ряда предварительных подсчетов числа хромосом может выявить
наличие в этой группе растений полиплоидного ряда, что послу-
жит первым указанием на то, что она представляет собой поли-
плоидный комплекс.
Расширение цитотаксономических исследований позволит нам
далее выяснить многие детали и завершить картину. Затем сле-
дует перейти ко второй стадии анализа, на которой иногда мож-
но обнаружить, что разные растения, обладающие сходными
морфологическими признаками и встречающиеся вместе в одной
и той же местности, имеют различные четные числа хромосом.
Эти данные позволяют предположить, что между видами-двой-
никами, находящимися на разных уровнях плоидности, сущест-
вует тесная филогенетическая связь. Или же можно обнаружить
ряд форм с разными и неинтерградирующими морфологически-
ми признаками, находящихся на одном и том же основном уров-
не плоидности и составляющих, таким образом, вероятную ос-
нову изучаемого полиплоидного комплекса. По мере накопле-
ния данных по подсчету хромосом в природных популяциях
можно начать устанавливать корреляции между тем или иным
уровнем плоидности и определенными морфологическими приз-
наками, биохимическими компонентами или величиной пыльце-
вых зерен и устьиц. Установив такие корреляции, можно путем
экстраполяции в известных пределах предсказывать таксономи-
ческие и географические границы некоторых видов-двойников.
Теперь на основе обширных цитотаксономических данных мож-
но описать в общих чертах наш полиплоидный комплекс.
Для третьей стадии необходимо прибегнуть к эксперимен-
тальным методам, хотя и в несколько предварительном плане.
Гл 25. Полиплоидный комплекс
317
Для этого проводятся скрещивания между некоторыми видами-
двойниками и определяется плодовитость полученных гибридов.
Эта третья, или биосистсматическая, стадия анализа необходи-
ма для того, чтобы убедиться в правильности некоторых родст-
венных отношений, предсказанных ранее на основе цитотаксоно-
мических данных Биосистсматические методы дают возмож-
ность определить некоторые филогенетические связи внутри по-
липлоидного комплекса.
Четвертая стадия состоит в приложении экспериментальных
генетико-таксономических методов ко всем членам изучаемого
комплекса. При помоши обычных методов искусственной гибри-
дизации, анализа геномов и в тех случаях, когда это возмож-
но, — ресинтеза прослежнваекя родословная одного тстраплоид-
ного вида от его диплоидных предков Используя те же методы,
следует далее усыновить родословные других тетраплоидных
членов комплекса Наконец, этими же методами устанавлива-
ются родословные тех его членов, которые обладают более вы-
сокой плоидпостыо. Теперь становится возможным проследить
филогенетическое происхождение определенных полиплоидных
видов от определенных диплоидных видов. Постулированную та-
ким образом филогению можно подкрепить данными цитогене-
тического анализа искусственных межвидовых гибридов, полу-
ченных при скрещиваниях друг с другом разных полиплоидных
видов Филогенетический анализ полиплоидного комплекса за-
вершен
Здесь полезно провести аналогию с химическими методами
анализа Мы можем сказать, что тайное неизвестное соединение
представляв! собой белок, установив, что в его состав входят
такие элементы, как азог и сера. Затем, применяя более тонкие
аналитические методы, можно определить его химическую фор-
мулу. При помощи еще более сложных и трудоемких методов
можно вывести его структурную формулу, отражающую химиче-
ские связи между составляющими его атомами Химические фор-
мулы известны для многих белков, однако лишь для очень не-
многих из них полностью установлена структурная формула.
Так обстоит дело и с изучением полиплоидного комплекса
Идентифицировать ту или иную группу растений как полиплоид-
ный комплекс (стадия 1) относительно легко Достаточно серь-
езный цитотаксономический анализ (стадия 2) провести значи-
тельно труднее, и он требует многих лет работы. Еще более
трудную задачу представляет биосистематический анализ (ста-
дия 3), который едва ли можно осуществить в одиночку. Излиш-
не говорить, что полный генетико-таксономический анализ одно-
го полиплоидного комплекса (стадия 4) —чрезвычайно трудоем-
кая задача, поглощающая массу времени.
318
Часть шестая. Полиплоидия
Заслуживают упоминания некоторые часто допускаемые ме-
тодологические ошибки. Специалисты по цитотаксономии иног-
да переоценивают надежность описательных цитотаксоном иче-
ских методов в применении к полиплоидному комплексу, недо-
оценивая в то же самое время важность генетико-таксономиче-
ского подхода. Разница между цито- и генетико-таксономиче-
скими методами по затрате труда и времени очень велика; одна-
ко для сколько-нибудь надежного филогенетического анализа
полиплоидного комплекса дополнительные усилия, связанные с
применением экспериментального генетико-таксономического
подхода, абсолютно необходимы.
Многие цитотаксономисты неверно интерпретируют получен-
ные ими результаты, рассматривая их в свете таксономической,
а не биологической или эволюционной концепции вида. Цитотак-
сономист обнаруживает группу диплоидов и тетраплоидов со
сходными морфологическими признаками; он рассматривает их
как различные «хромосомные расы» одного и того же вида
Другая группа диплоидов и тетраплоидов, сходных между собой,
но отличающихся по морфологии от первой группы, рассматри-
вается как другой вид, состоящий из двух «хромосомных рас».
В результате получается таксономическая система, в которой час-
то бывают смешаны естественные родственные связи и взаимо-
отношения по плодовитости. Морфологически различные тетра-
плоидные типы могут быть на самом деле противоположными
выщепенцами одного и того же сегментного аллотетраплоида
(ЛвАзЛИг) и в качестве таковых должны рассматриваться как
относящиеся к одному и тому же виду. В таких случаях погра-
ничную черту, разделяющую разные виды, следует проводить не
в середине комплекса AsAsAtAt, а между этим комплексом в це-
лом и диплоидными видами и A/At.
Многие группы растений были идентифицированы как поли-
плоидные комплексы. Довольно большое число полиплоидных
комплексов уже подверглось детальному цитотаксономическому
анализу, а некоторые из них—и предварительному биосистема-
тическому анализу. И лишь для нескольких полиплоидных комп-
лексов был проведен обширный, хотя и неполный генетико-так-
сономический анализ.
В двух следующих разделах рассматриваются главные осо-
бенности некоторых полиплоидных комплексов, изученных цито-
таксономически и биосистематически. Далее будут рассмотрены
полиплоидные комплексы Triticum—Aegilops, Bromus и Gilia,
по которым были проведены обширные генетико-таксономиче-
ские исследования.
Гл 25 Полиплоидный комплекс
319»
Цитотаксономические примеры
Эрепдорфер и его сотрудники провели цитотаксономпческпй ана-
лиз полиплоидных комплексов в нескольких ветвях рода Galium
fRubiaceae). Такие полиппоидные комплексы описаны в группе
G pumilum в Европе [184], в группе G. anisophyllum также в
Европе [191], в группе G. incanutn в Турции и соседних с ней об-
ластях [185] и в группе G. multiflorum в западной части Север-
ной Америки [161, 189].
Грхппа Galium pumilum (х=11) в Европе содержит поли-
плоидный ряд от диплоидов до октоплоидов Диплоидный член
комплекса, G austriacum, встречается как реликт ледникового
времени в разобщенных районах Альп, где входящие в этот
комплекс полиплоиды занимают обширные и непрерывные ареа-
лы Г184].
Полиплоидный комплекс в группе Galium anisophyllum r
Европе содержит все четные полиплоиды от 2х до 10х Здесь
снова липлоиды встречаются в ряде убежищ, разбросанных по
горам Европы, а тетраплоиды занимают непрерывный и обшир-
ный ареал. Распространение видов с высоким уровнем плоид-
ности показано па рис. 25 2 [191].
Sanicula crassicaulis и родственные ей формы (Umbelliferae)
образуют еще один полиплоидный комплекс с основным числом
хромосом х-^8 [551 В таксономический вид S crassicaulis вхо-
дят тетраплоиды. гексаплоиды и октоплоиды, занимающие об-
ширную область вдоль тихоокеанского побережья Северной
Америки и Чили Полагают, что диплоидные виды S. laciniala и
S hoffmannii, встречающиеся в прибрежных районах Калифор-
нии, служат вероятными предками тетраплоидного S. crassicau-
lis Другой диплоидный вид, S bipinnatifida, судя по его морфо-
логическим признакам, возможно, был одной из предковых форм
некоторых октоплоидпых членов этого комплекса [55].
Группа папоротников, принадлежащих к роду Asplenium
(Polypodiaccac) и распространенных в восточной части Север-
ной Америки, образует еще один полиплоидный комплекс при
основном числе хромосом х^Зб [8641. В этом комплексе есть
диплоидные виды, обладающие крайними морфологическими
признаками, — A. montanum, A. platyneuron и A. rhizophyllum
(2п = 72) В некоторых случаях промежуточные по морфологи-
ческим признакам формы являются стерильными диплоидными
или триплоидными гибридами. Другие — плодовитые — проме-
жуточные формы представляют собой тетраплоиды (2п = 144)
Таким образом, тетраплоид A. pinnatifidum занимает по морфо-
логическим признакам промежуточное положение между А топ-
tanum и A. rhizophyllum и, вероятно, является амфиплоидным
производным этих двух диплоидных видов [864].
Рис. 252. Географическое распространение диплоидов и полиплоидов, при-
надлежащих к группе Galium anisophyllum [191].
Гл 25 Полиплоидный комплекс
321
Родственные взаимоотношения в этой группе Asplenium,
установленные Вагнером [864] при помощи цитотаксономических
методов, были затем проверены с применением хемотаксономи-
ческих методов [740]. Методом хроматографии на бумаге было
исследовано содержание определенных химических соединений
у разных видов. Оказалось, что естественные гибриды и алло-
тетраплоиды содержат в разных комбинациях вещества, имею-
щиеся у их предполагаемых диплоидных родительских видов
[740]. Хемотаксономические данные подтвердили филоюнию,
разработанную на основе цитотаксономических данных.
Другие полиплоидные комплексы, изученные циготаксономи-
ческими методами, — это группа Polypodium, vulgare (Polypo-
diaceae) в Европе и Северной Америке и группа Trillium camsch-
tatcense (Liliaceae) в Японии [454, 554—556]. Среди многочис-
ленных полиплоидных комплексов в сем Gramineae, исследо-
ванных цитотаксономическими методами, укажем группу Огу-
zopsis-Stipa в Северном полушарии, циркумполярную группу
Роа alpina и группу Bouteloua curtipendula североамериканских
равнин [262, 411, 627].
Биосистематические примеры
Группа Phacelia heterophylla состоит из свободно скрещиваю-
щихся многолетних и двулетних травянистых растений, широко
распространенных по всей западной части Северной Америки и
южной части Южной Америки. Эта группа сложна в таксономи-
ческом отношении, так как содержит много разных, но ингер-
градирующих морфологических форм. Хекард [358] установил,
что данная группа предсгавляет собой полиплоидный комплекс
(х= 11), состоящий из шести известных диплоидных видов, мно-
гочисленных тетраплоидов и нескольких гексаплоидов
Как показывают результаты искусственной гибридизации,
между диплоидными видами существуют хорошо развитые барь-
еры, препятствующие скрещиванию, и барьеры, создаваемые
стерильностью гибридов. В отличие от диплоидов различные
тетраплоидные формы в большинстве случаев довольно легко
скрещиваются друг с другом, давая полуплодовигых или плодо-
витых гибридов. Эти различия в степени внутренней изоляции
коррелируют с различиями в характере изменчивости На дип-
лоидном уровне таксоны обычно разделены разрывами в мор-
фологических признаках, тогда как на тетраплоидном уровне
таксоны часто образуют непрерывный ряд благодаря интергра-
дации по этим признакам [358].
Полиплоидные представители группы Phacelia heterophylla,
вероятно, представляют собой амфиплоидов, возникших в ре-
зультате гибридизации между диплоидными видами в различ-
21-45
322
Часть шестая. Полиплоидия
ных комбинациях; такое толкование послужило бы дальнейшим
объяснением их интерградации по морфологическим признакам.
У нас пока нет никаких данных относительно происхождения
различных тетраплоидных и гексаплоидных форм.
Хекард [358] объединил морфологически близкие диплоидные
и тетраплоидные формы в один вид. Следовательно, единицы,
которым были даны видовые названия, являлись во многих слу-
чаях таксономическими видами, но отнюдь не биологическими
или эволюционными видами. Полученные Хекардом генетические
данные иногда не согласуются с морфологическими данными,
чего и следовало ожидать Так, диплоидный вид Phacelia nemo-
ralis не удалось искусственно скрестить с тетраплоидным Ph. пе-
moralis, однако тетраплоидный Ph. nemoralis скрещивается с
тетраплоидным Ph. californica, образуя высокоплодовитые гиб-
риды. Дальнейшие этапы анализа этого интересного полиплоид-
ного комплекса должны состоять в том, чтобы выделить вчерне
биологические виды на тетраплоидном и гексаплоидном уровнях
и установить их родословнзпо.
Род Clarkia (Onagraceae) в западной части Северной Амери-
ки и в умеренной зоне Южной Америки содержит более обшир-
ный полиплоидный комплекс [496]. У диплоидных видов 2/г=10,
14, 16 и 18 хромосомам при первоначальном основном числе
х=7. Эти диплоидные виды распадаются на несколько ясно вы-
раженных групп видов, которые по праву возведены в ранг от-
дельных родов: Godetia, Euchardium и Clarkia sensu stricto.
Однако те филогенетические линии, которые на диплоидном
уровне дивергируют, на тетраплоидном и гексаплоидном уров-
нях сливаются в различных комбинациях. Такие анастомозы
служат основанием для того, чтобы рассматривать всю эту со-
вокупность как один род Clarkia sensu lato (496].
Постулированная сетчатая филогения рода Clarkia так, как
ее представляли в 1955 г., схематически показана на рис. 25.3.
После 1955 г. Люис и его сотрудники получили много дополни-
тельных данных. На рис. 25.3 изображена вероятная родослов-
ная различных полиплоидных видов Clarkia, построенная на ос-
нове морфологических и цитологических данных [496].
Группа A chiilea millefolium (Compositae)—полиплоидный
комплекс (х=9), широко распространенный в Северном полу-
шарии [102, 188]. Собственно A. millefolium в Старом Свете и
A. borealis в Новом Свете являются гексаплоидами; A. collina
и A. virescens в Старом Свете и североамериканский A. lanulo-
sa— тетраплоидами. В Европе встречаются два европейских
диплоидных вида —A. asplenifolia (на влажных лугах) и A. se-
tacea (в сухих степях) (102, 189].
Ареалы диплоидных видов Achillea asplenifolia и A. setacea
перекрываются, и, несмотря на сильные внутренние изолирую-
to
Рис. 25 3. Филогенетические взаимоотношения в полиплоидном комплексе рода Clarkia [496].
Прописными буквами обозначены секции; цифры в скобках рядом с видовыми названиями указывают нормальное число
хромосом в гаметах у данного вида.
324
Часть шестая. Полиплоидия
шие барьеры, эти виды гибридизируют в природе, хотя и в ог-
раниченной степени. В Альпах происходит интрогрессия от
A. setacea к A. asplenifolia на диплоидном уровне. Тетраплоид
A. collina— амфиплоид, происходящий от этих двух диплоидов.
Затем A. collina гибридизирует с другим тетраплоидным видом
A. virescens. В некоторых сочетаниях естественная гибридизация
происходит также между тетраплоидными и амфиплоидными
видами: A. collina (4х) X A. millefolium (6х) и A. lanulosa (4х)Х
ХА. borealis (6х); при этом образуются анеуплоидные потомки,
что еще больше осложняет характер изменчивости всей этой
группы [189].
Цохари и Фельдман [918] обнаружили сходную ситуацию в
полиплоидном комплексе, в который входят Triticum и Aegilops
(Gramineae). Здесь так же, как и у Achillea, разные аллотетра-
плоидные виды с различной геномной конституцией гибридизи-
руют в природе, что еще больше усложняет и без того сложную
изменчивость этой группы [918].
Комплекс Gilia inconspicua
Группа Gilia inconspicua (х=9)—это часть секции Arachnion
рода Gilia, которая состоит из. автогамных однолетников. Эта
группа широко распространена в западной части Северной
Америки и на юге Южной Америки. В Северной Америке изве-
стно 23 вида этого комплекса, а в Южной — 2 вида; есть веские
основания считать, что в каждой из этих областей существуют
и другие еще неизвестные виды. Из 23 североамериканских видов
13 диплоидны (2п=18) и 19 тетраплоидны (2п=36). Эти тетра-
плоиды представляют собой амфиплоидов, и в большинстве слу-
чаев их предки установлены.
В гл. 11 группа Gilia inconspicua была приведена в качестве
примера одного из типов межвидовых взаимоотношений, обыч-
ных среди автогамных однолетних растений. Часть этой группы,
к которой относятся О. transmontana и родственные ей виды,
уже рассматривалась в гл. 4 в связи с проблемой видов-двойни-
ков (см. рис. 4.5). Здесь мы вновь рассмотрим эту группу как
полиплоидный комплекс, который подвергся широкому генети-
ко-таксономическому изучению, но все еще не проанализирован
до конца.
Диплоидные виды этой группы распадаются на 5 геномных
групп. Родственные виды, принадлежащие к одной и той же ге-
номной группе, обычно различаются по субгеномам [300].
Гибриды Fb полученные от 17 скрещиваний между тетра-
плоидными видами в разных комбинациях, были подвергнуты
цитогенетическому анализу. У большей части этих гибридов
конъюгация хромосом в мейозе протекала на среднем уровне.
Гл. 25. Полиплоидный комплекс
325
Так, в Fi от скрещивания Gilia malior и G. transmontana наблю-
далось в среднем по 9 бивалентов при диапазоне от 7 до 15 II,
тогда как при полной конъюгации образуется 18 бивалентов.
Уровень конъюгации хромосом у этого и у сходных с ним гиб-
ридов позволяет предположить, что два участвовавших в гибри-
дизации тетраплоидных вида имеют один общий геном, но раз-
личаются по второму геному [285].
Для некоторых пар тетраплоидных видов это предположение
подтвердилось; общий диплоидный геном идентифицировали,
получив соответствующие триплоидные гибриды и установив, что
конъюгация хромосом происходит, как и предполагалось, по ти-
пу Drosera. Следовало ожидать, что у триплоидных гибридов,
полученных в результате скрещиваний между тетраплоидпыми
и диплоидными видами, принадлежащими к одной из линий
предков — потомков, конъюгация будет происходить по ти-
пу >9 П+<18 I.
Именно такой тип конъюгации и был обнаружен у триплоид-
ных гибридов Gilia sinuata (4х) X G. latiflora (2х) и G. то do cert-
sis (4x)xG latiflora (2х) [285]. Следовательно, геном G latiflo-
ra, или геном Т, имеется как у G. sinuata, так и у G modo*
censis. Па основе морфологических и экологических данных мож-
но сделать вывод, что именно G latiflora, а не какой либо дру-
гой член Т-геномной группы послужил одним из диплоидных ро-
дительских видов и для G. sinuata, и для G. modocensis. Что же
касается видов, сыгравших роль второй родительской формы
для этих двух тетраплоидных видов, то они различаются как по
своим геномам, так и морфологически.
Дэй [158] провела подробное таксономическое исследование
двух тетраплоидных видов и их предполагаемых диплоидных ро-
дительских видов в подгруппе Gilia transmontana. Морфологи-
ческие, экологические, цитологические и цитогенетические дан-
ные свидетельствовали о том, что родословная этих двух тетра-
плоидов, G transmontana и G. malior, носи г сетчатый характер
и что они происходят от трех диплоидных видов, как эго пока-
зано на рис. 25 4. Дэй затем подтвердила правильность такой
филогении, искусственно ресинтезнровав G transmontana и
G malior из их диплоидных родительских форм и показав, что
каждый искусственный амфиплоид дает плодовитых потомков
при скрещивании с соответствующим природным тетраплоид-
ным видом [158].
Сходные данные были получены для родословной тетрапло-
идного вида, известного под названием Gilia ophthalmoides III.
Считалось, что этот вид происходит от скрещивания между
G. clokeyi (2х) и G. mexicana (2х). Сломанные тетраплоидные
потомки, полученные от этого диплоидного межвидового гибри-
да, оказались хорошими G. ophthalmoides, что подтвердило пред-
326
Часть шестая. Полиплоидия
полагавшуюся филогению ('[300], а также неопубликованные
данные V. и A. Grant). Имеются и другие данные, указывающие
на частично общую, а частично отличающуюся родословную
близкого к G. ophthalmoides III тетр апл оидного вида—G. opht-
halmoides I.
Эту сетчатую систему родственных связей можно расширить,
включив в нее Gilia sinuata, G. modocensis и другие тетр апл оид-
ные виды. Сетчатая родословная двух частей этого полиплоид-
ного комплекса изображена на рис. 25.4.
Triticum и Aegilops
Хорошо известный полиплоидный ряд пшеницы составляет часть
более крупного полиплоидного комплекса, в который входит
Aegilops. Triticum и Aegilops связаны между собой на полипло-
идном уровне. Разграничение старых родовых единиц — пшени-
цы и эгилопса, — основанное на различиях в признаках колоса
и колосков, оказывается, таким образом, несостоятельным. Оче-
видно, Triticum и Aegilops составляют один род, и специалисты
по генетике пшеницы склонны перенести виды, относившиеся до
сих пор к Aegilops, в род Triticum. Однако эта таксономическая
и номенклатурная реорганизация еще не закончена, и пока что
Гл. 25. Полиплоидный комплекс
327
Таблица 25 1 Система классификации и геномная конституция
пшениц (Triticum) (основана па данных Фельдмана [211];
данные о распространении по Харлану и Цохари [348], а также
по Цохари и др. [919])
1. Т monococcum Однозернянка. 2х. АА
а) Дикорастущая раса Т. m boeoticum
Ближним Восток ог Турции до Ирана и южная часть Балканского
полуострова1
б) Культурная раса Т m monococcum
2 Т turgidum Культурная двузернянка и твердая пшеница 4х ААВВ
а) Дикорастущая раса Т t dicoccoid.es
Ближний Восток ог Турции и Израиля до Ирана
б) Культурная пленчатая раса Т t dicoccum
в) Культурные голозерные расы. Т t durum, turgidum, polonicum,
carthlicum
3 T iimopheevii 4x AAGG
а) Дикорастущаяа paca T. t. araraticum
На юге СССР, в Грузии и Армении2
б) Культурная раса
4 Т aestivum (=Г. vulgare). Мягкая, или обыкновенная, пшеница, бх.
AABBDD
а) Культурные пленчатые расы Т ае spelta, macha, vavilovii
б) Культурные голозерные расы Т. ае. aestivum, compactum, sphaero-
coccum
1 T boeoticum встречается также в Южном Крыму, в южней части Западного За-
кавказья и в Южном и. Восточном Закавказье Близкий вид, Т urartu, встречается в Ар-
мении и в северо-восточной части Турции (Цвелев Н Н .ос cit • — Лрим ред
1 Т araraticum встречается также в северо-восточной чао и Турции и в Северном
Иране — Прим ред
мы имеем дело с двумя перекрывающимися наборами родовых
и видовых названий.
Triticum—Aegilops — группа однолетних злаков, принадле-
жащих к подтрибе Triticinae трибы Hordeae1 Как у Triticum—
Aegilops, так и у трибы в целом основное число хромосом х = 7.
В отношении рода Triticum в прежнем узком смысле в пос-
ледние годы наблюдается тенденция к низведению многих групп,
прежде считавшихся видами, до ранга подвидов, или культива-
ров, что упрощает систематику пшениц. В табл. 25.1 представ-
лена система пшениц, основанная на работе Фельдмана [211].
Геномный анализ пшениц проводили Кихара [436, 437], Сирс
[718, 719] и многие другие. Полиплоидная пшеница получила
геном А от диплоидного вида Triticum monococcum Источник
генома В у тетраплоидной Т. turgidum неизвестен; мы вновь вер-
немся к этому вопросу чуть ниже. Геном D у гексаплоидной
Т. aestivum происходит от диплоидного вида Aegilops squarro-
1 Согласно правилу приоритета, эту трибу следует называть Triticeae
(см Цвелев Н. Н, Злаки СССР, Изд-во «Наука», 1976, с. 105). — Прим ред.
328 Часть шестая. ПолиплоиОия
sa(=T. tauschii)1, занимающего обширный ареал в западной
части Азии и на юге Средней Азии.
Геном В имеется как у Triticum turgidum, так и у Г. aesti-
vum. Попытки найти диплоидный вид с конституцией ВВ оказа-
лись безуспешными. Одно время полагали, что Aegilops speltoi-
des(=Triticum speltoides) является носителем генома В, однако
цитогенетические и электрофоретические данные противоречат
этой гипотезе, и в настоящее время она в своей первоначальной
форме отвергнута. Теперь считается, что Ае. speltoides содержит
геном SS. Что касается необнаруженного диплоида ВВ, то воз-
можно, что он вымер. Другая возможность состоит в том, что
диплоида В В никогда и не существовало. Геном В, имеющийся
у Т. turgidum, мог возникнуть у тетраплоидной Т. turgidum в
результате дивергенции от какого-нибудь другого родственного
генома, возможно S [211].
Филогению полиплоидного ряда, ведущего к мягкой пшени-
це, можно представить следующим образом: 1) гибридизация
между дикорастущей однозернянкой (АА) и неидентифициро-
ванным диплоидом, несущим геном В, или каким-то предшест-
венником В, с образованием амфиплоидной дикорастущей дву-
зернянки (ААВВ); 2) превращение дикорастущей двузернянки
в культурную пленчатую, а затем в культурную твердую пшени-
цу в результате изменений, вызванных селекцией в условиях
доместикации; 3) гибридизация между культурной двузернянкой
и Aegilops squarrosa' (DD) с образованием амфиплоидной
мягкой пшеницы (AABBDD); 4) селективные изменения при
культивировании, приведшие к превращению пленчатой пшени-
цы в голозерную мягкую пшеницу.
Эти изменения происходили в западных и юго-западных
районах Азии. При археологических раскопках в этих районах
в ряде мест найдены карбонизированные зерна Triticum mono-
coccum, Т. turgidum и Т. aestivum. Возраст древнейших найден-
ных зерен этих трех видов соответствует 9000 лет до н.э. [211].
Дикорастущая двузернянка встречается в настоящее время
на Ближнем Востоке от Израиля и Ливана до реки Тигр и Кас-
пийского моря 1[348]. Твердые пшеницы выращивают главным
образом в Средиземноморской области, где климат относитель-
но мягкий [919]. Aegilops squarrosa распространен от прикаспий-
ского региона до среднеазиатских степей и доходит на востоке
до Ташкента и Алма-Аты [919]. Этот вид, очевидно, представлен
рядом экотипов, в том числе экотипами, адаптированными к
1 Н Н. Цвелев (loc cit) считает Aegilops squarrosa синонимом A triun-
cialis subsp triuncialis распространенного от Средиземноморья до Средней
Азии, а Ае. tauschii он признает самостоятельным видом и даже относит к
другой секции рода Aegilops —Прим, ред
Гл 25. Полиплоидный комплекс
329
резко континентальному климату. Экологические адаптации
Ае squarrosa составляют самый важный вклад этого вида в
формирование мягких пшениц. Наличием у гексаплоидной мяг-
кой пшеницы генома D объясняется ее широкое экологическое
разнообразие и ее способность расти в холодном и сухом степ-
ном климате ([919]; Харлан, личное сообщение)
Интересно отметить, что этот процесс продолжается у искус-
ственно синтезированного аллооктоплоида Triiicale — амфипло-
идного производного Triticum aesiivum (6x)xSecale cereale (2x),
который можно выращивать в северных широтах и в горных
районах
Triticum titnopheevii обладает геномом AAGG (табл. 25.1)
и представляет собой, таким образом, еще одно тетраплоидное
производное Т. monococcum. Другой диплоидной родительской
формой мог быть Aegilops speltoides (SS) или такой близкий
к нему вид, как Ае. bicornis (SbSb). Геном S близок к геному
G. Эта филогенетическая гипотеза допускает, что геном S прев-
ратился в геном G у тетраплоида Т. titnopheevii [211]
Среди большого числа видов, относимых к старому роду
Aegilops, встречаются диплоиды, тетраплоиды и гексаплоиды.
У Aegilops различают пять геномов и еще большее число субге-
номов Пять геномных групп, имеющихся у диплоидных видов,
по большей части совпадают с таксономическими секциями это-
го рода. Главные геномы, субгеномы и их диплоидные носители
перечислены в табл 25 2 Некоторые субгеномы (Л1сг, D2,SV и
т п), известные только у полиплоидов, в табл. 25.2 не вклю-
чены.
Геномы полиплоидных видов Aegilops приведены в табл. 25.3,
из которой видно перекрещивание филогений этих видов. Так,
Таблица 25.2. Геномы и их диплоидные носители у
Aegilops [437, 918]
Геном Субгеном Диплоидный вид
Mt Ае mutica 1
S Sb Ае bicornis
5* Ае sharonensis, longissima
5 Ае speltoides, ligustica
D Ае squarrosa
С С Ае caudata
С“ Ае. umbellata
м м Ае cornosa
Ми Ае uniaristata
’) Aegilops mutica в настоящее время выделяется в отдельный моно-
типный род Amblyopyrum (см Цвелсв II Н op cit , с 154) — Прим ред.
330
Часть шестая. Полиплоидия
Таблица 25.3. Геномная конституция
полиплоидных видов Aegilops (918]
Полиплоидный вид Геном
Ае. crassa (4х) DDMcrMer
Ае. crassa (6х) DDD2D2McrMcr
Ае. Juvenalis DDCaCaM!M!
Ае. ventricosa DDM”M”
Ае cylindrica DDCC
Ае variabilis и Ае. kotschyl Qt&S’S*
Ае. triuncialis CUC“CC
Ае. columnaris CuCuMeMe
Ае. biuncialis C“C“MbMb
Ае triaristata (4х)
Ае triaristata (6х) СиСиМ*М‘М*2М‘2
Ае ovata C“C“M°M°
два диплоидных вида, несущих геномы DD и ММ, дали начало
тетраплоидам с конституцией DDMM. Один из этих диплоидов
(DD) и другой диплоид (СС) дали тетраплоид DDCC и т. д.
Разные тетраплоидные виды нередко имеют по одному общему
диплоидному предку и различаются по другому. Процесс реком-
бинации геномов продолжается на уровне гексаплоидов.
Сетчатый характер взаимоотношений в группе Triticum—
Aegilops представлен здесь как сложение различных геномов
в результате амфиплоидии. Однако это лишь часть проблемы.
Имеются данные, как прямые, так и косвенные, свидетельствую-
щие о естественной гибридизации и интрогрессии между некото-
рыми участвующими во всех описанных выше процессах видами.
В результате такой гибридизации изменялись морфологические
признаки, а, возможно, и геномы этих видов I[i211, 636, 918, 919].
Старые полиплоидные комплексы
С течением времени диплоидные виды, входящие в данный
полиплоидный комплекс, могут вымереть. Могут вымереть так-
же и низкие полиплоиды, так что единственными живыми пред-
ставителями комплекса останутся формы с более высокой пло-
идностью.
Группа Brotnus carinatis (Gramineae) служит примером по-
липлоидного комплекса, далеко продвинувшегося в' своем разви-
тии. В этой группе есть формы, достигшие высоких уровней пло-
идности, например В. arizonicus (12х), однако многие ее дипло-
идные и тетраплоидные представители вымерли. Стеббинс и его
сотрудники (779, 781, 794, 795] установили филогению 12х- и
8х-видов этой группы.
Гл 25. Полиплоидный комплекс
331
Комплекс Bromus carinatus принадлежит к секции Cerato-
chloa рода Brotnus, но входящие в него виды содержат геномы
двух близких секций, Bromopsis и Neobromus. Все эти расте-
ния— многолетние злаки, нередко рыхлокустовые, произрастаю-
щие в Северной и Южной Америке. Основное число хромосом
х = 7.
В секции Ceratochloa рода Brotnus^ диплоидные или тетра-
плоидные виды неизвестны. В. catharticus и его родичи, произ-
растающие в Южной Америке, — аллогексаплоиды, несущие
геномы, происходящие от диплоидных видов, которые вымерли
(рис 25 5).
Bromus carinatus и В. margittatus, растущие в Северной Аме-
рике, — аллооктоплоиды. Три из имеющихся у них четырех ге-
номов гомологичны геномам южноамериканских гексаплоидов.
Четвертый геном L, судя по цитологическим и морфологическим
данным, происходит от какого-то североамериканского диплоид-
ного вида, относящегося к секции Bromopsis (рис. 25.5). Эти
октоплоиды амфиплоидного происхождения возникли, вероятно,
в западной части Северной Америки в конце третичного перио-
да, возможно в плиоцене. Приходится допустить, что в прош-
лом в Северной Америке существовали какие-то гсксаплоидные
виды, гомологичные современным южноамериканским гексапло-
идам [779, 794].
Bromus arizonicus — амфиплоид с числом хромосом 12х. Он
также гомологичен южноамериканским гексаплоидам по трем
из своих шести геномов (рис 25 5). Остальные три генома
В. arizonicus происходят, по-видимому, от какого-то аллогекса-
плоида, принадлежащего к родственной секции Neobromus
(рис 25.5) [795].
Двенадцатиплоидный вид Bromus arizonicus — обычный и
агрессивный сорняк на большей части своего ареала в Аризоне
и Калифорнии. Его диплоидные и тетраплоидные предковые
формы вымерли Его гексаплоидный предок, относившийся к
секции Ceratochloa, в Северной Америке вымер, возможно в пе-
риод неблагоприятных климатических изменений в плиоцене —
плейстоцене, по дожил до наших дней в более умеренных кли-
матических условиях Южной Америки Между тем 12-плоидное
производное этих вымерших диплоидов и низких полиплоидов —
агрессивный вид, процветающий в настоящее время и демонст-
рирующий, как непрерывное амфиплоидпое удвоение может
вдохнуть новую жизнь в старый полиплоидный комплекс [779,
781].
1 Некоторые систематики, в том числе Н Н Цвелев (loc cit, стр 207),
считают секцию Ceratochloa рода Bromus самостоятельным родом Cerato-
chloa — Прим ред
332
Часть шестая. Полиплоидия
Рис. 25.5. Филогенетические взаимоотношения высокополиплоидных видов
Bramas £781].
Диплоидные или тетраплоидные носители некоторых исходных геномов не-
известны.
Остатки древних полиплоидных комплексов
Стеббинс (787, 789] отметил наличие обратной корреляции меж-
ду степенью морфологической специализации и уровнем плоид-
ности в разных семействах покрытосеменных. Оказалось, что в
пределах данного семейства растений для морфологически более
примитивных родов характерно полиплоидное основное число
Гл 25. Полиплоидный комплекс
333
хромосом. Напротив, у родов с более специализированными
морфологическими признаками основные числа — диплоидные
Приведем несколько примеров [789]:
Liliaceae
"Wintcraceae
Polemoniaceae
Hydrophyllaceae
Compositae
Морфологически
примитивные виды, по-
липлоидные
основные числа
Tofieldia
Dritnys
Cantua, Cobaea
Wigandia
Heliantheae
Морфологически
специализированные ви-
ды, диплоидные основ-
ные числа
Trillium, Scilla
Tasmannia
Gilia, Phlox
Nemophila
Cichorieae
Эволюционно продвинутые роды в этих семействах сохра-
нили диплоидность своих предков Сохранение диплоидности
играло, возможно, важную роль в их способности достигнуть
высоких ступеней эволюционного прогресса. Вместе с тем при-
надлежащие к тем же семействам роды с высокими хромосом-
ными числами — давние полиплоиды, нередко остатки старых
полиплоидных комплексов, сохранившиеся в неизменном состоя-
нии [787, 789].
В некоторых семействах древесных двудольных все предста-
вители отличаются высоким основным числом хромосом, неко-
торые авторы [780, 787, 832] рассматривают это как сохранение
давнего полиплоидного состояния, что кажется правдоподобным.
Magnoliaceae x=19
Trochodendraceae x=19
Cercidiphyllaceae x=19
Salicaceae x=19
Hippocastanaceae x=20
Platanaceae x=21
У большинства главных групп Pteridophyta основные хро-
мосомные числа лежат в полиплоидном диапазоне, а у некото-
рых групп — в верхней его части [533, 554]. Например:
Polypodium, etc (Polypodiaceae)
Psilotum (Psilotaceae)
Tmesipterus (Psilotaceae)
Equisetum (Equisetaceae)
Ophioglossum (Ophioglossaceae)
x=37
x=50, 52
X=104
x=108
x=l20
334
Часть шестая. Полиплоидия
Наиболее вероятный вывод, к которому можно прийти на
основе этих данных, состоит в том, что многие виды современ-
ных Pteridophyta — дожившие до наших дней остатки геологи-
чески древних полиплоидных комплексов [554].
Необратимость
Эволюция, направленная от диплоида к тетраплоиду и поли-
плоиду более высокого уровня, в основном необратима. Она мо-
жет частично повернуть в обратную сторону в результате поли-
плоидного сброса (см. гл. 22 и 27). Так, у Hesperis, у которого
х=7, число хромосом в гаметах тетраплоидных видов п=14,
13 и 12.
Некоторые исследователи высказывали мнение о возможно-
сти резкого и полного возврата из тетраплоидного в диплоид-
ное состояние посредством образования полигаплоидов [160,
667]. Гаплоидные или полигаплоидные потомки от тетраплоид-
ных родительских форм, безусловно, будут диплоидными. Такие
полигаплоиды иногда действительно возникают в полиплоидных
популяциях растений. Однако полигаплоид, происходящий от
аллотетраплоида, и по определению, и по результатам наблю-
дений представляет собой диплоидный гибрид, характеризую-
щийся хромосомной стерильностью, и его главная возможность
размножаться половым путем — это вторичное удвоение. Поли-
гаплоиды вряд ли могут повернуть вспять эволюцию, направ-
ленную в сторону полиплоидизации.
Подавление изменчивости
Полиплоидное состояние по сравнению с диплоидным несколько
ограничивает возникнрвение мутационной и рекомбинационной
изменчивости, и эти ограничения становятся более жесткими с
повышением уровня плоидности. Дупликация генов и полисом-
ное наследование подавляют выражение новых мутаций. Эти
факторы, а также избирательное спаривание ограничивают об-
разование новых рекомбинантов.
Тышлер [831] отмечает, что мутантные проростки, обычные у
Triticum monococcum (2х), у Т. durum (4х) встречаются реже,
а у Г. aestivum (6х) крайне редки.
Полиплоидное состояние ограничивает возникновение измен-
чивости, но не исключает ее полностью. Сегментные аллополи-
плоиды могут расщепляться до некоторой степени по межвидо-
вым генным различиям У полиплоидов изредка происходит
выщепление мутантов в полисомных соотношениях. Тем не ме-
нее у полиплоидов по сравнению с диплоидами создающий из-
Гл. 25. Полиплоидный комплекс
335
менчивость механизм, связанный с половым процессом, подав-
лен.
Следует таким образом ожидать, что у полиплоидных видов
способность реагировать на изменяющиеся условия среды выра-
жена слабее, чем у диплоидных видов Безусловно, новые поли-
плоиды часто развиваются очень успешно, более успешно, чем
их диплоидные родичи, в местообитаниях разных типов (см.
гл. 24). Их успех объясняется наличием у них генетической сис-
темы, допускающей размножение определенного тибридного ге-
нотипа в чистоте. Но эта генетическая система одновременно
налагает ограничения на образование новых вариаций и тем са-
мым устанавливает пределы потенциальным эволюционным воз-
можностям данной полиплоидной линии.
По мере того как данная филетическая линия достигает (в
основном необратимо) высших уровней плоидности, эти ограни-
чения эволюционного потенциала возрастают. Полиплоидная
система, как утверждает Стеббинс [780], способна создавать
многочисленные родственные виды или даже роды, обладающие
в общем сходными признаками, однако она, очевидно, не прини-
мала никакого участия в возникновении новых семейств или
порядков.
Диплоидизация
Давно возникшие полиплоиды обычно больше похожи на дип-
лоидов, чем вновь образовавшиеся. Это явление известно под
названием диплоидизации. Диплоидизация влияет как на цито-
логические признаки полиплоидов, так и па их генетическую
конституцию.
Полагают, что для вновь образовавшегося, или «сырого»,
полиплоида, если это автополиплоид, должна быть характерна
высокая частота мультивалентов, а если это сегментный алло-
полиплоид, то образование некоторого их числа. Мультиваленты
снижают плодовитость полиплоида Естественный или искусст-
венный отбор на плодовитость будет благоприятствовать генам
(если такие гены имеются в генофонде), способствующим обра-
зованию бивалентов за счет мультивалентов
Джайлс и Рэндолф [243] сравнивали частоту мультивалеп-
тов в одной автотетраплоидной линии кукурузы (Zea mays,
х=10) в начале и в конце 10-летнего периода отбора на плодо-
витость У вновь образовавшихся автотетраплоидных растений
8—10 IV содержались в 89% материнских клеток пыльцы, а у
их потомков 10-го поколения — только в 52% этих клеток Час-
тота образования бивалентов возрастала по мере снижения час-
тоты квадривалентов. Искусственный отбор на плодовитость,
вызвавший это изменение, возможно, сопровождался бессозна-
336
Часть шестая. Полиплоидия
тельным отбором на генетические факторы, влияющие на способ
конъюгации хромосом при мейозе.
Отбор генов, оказывающих влияние на мейоз, вероятно, сыг-
рал важную роль в эволюции многих естественных амфиплои-
дов. У гексаплоидной пшеницы Triticum aestivum наблюдается
значительная гомология между разными геномами. Однако го-
меологичные хромосомы обычно не конъюгируют, а образуют
биваленты, состоящие из гомологичных хромосом. Такое пове-
дение в мейозе обусловлено одним геном, локализованным в
длинном плече пятой хромосомы пшеницы. При отсутствии этого
гена образуются мультиваленты. Если этот ген (ген 5В) имеет-
ся, то он подавляет образование мультивалентов и способствует
конъюгации гомологичных бивалентов [686].
Дупликатные факторы достаточно часто встречаются у новых
амфиплоидов и обычны у старых. Дупликатные локусы являют-
ся следствием сложения родственных геномов. Дупликатные
факторы могут оказаться ненужными для функционирования
организма; в таком случае функция одного из локусов может
отклониться от первоначальной функции. Если такой дивергиро-
вавший ген начнет выполнять какую-либо другую близкую
функцию, выгодную для данного организма, то он будет сохра-
няться отбором.
Интересный пример дивергенции между факторами, которые
прежде были дупликатными, обнаружен у аллотетраплоидных
хлопчатников Gossypium hirsutum и G. barbaden.se. Их геномы
имеют конституцию AADD. Диплоиды АА — хлопчатники, выра-
щиваемые в Старом Свете, а диплоиды DD — дикорастущие аме-
риканские виды.
Группа сцепления, в которую входит ген R, детерминирую-
щий пигмент антоциан, и ген С1, детерминирующий скученность
соцветия, содержится как в геномах А и D, так и в геноме АА
тетраплоидов. У диплоидов АА и DD ген R представлен в одном
локусе, но у тетраплоидов AADD —в двух локусах (7?t и /?2).
Подобным же образом ген С1 представлен в одном локусе у
диплоидов АА, а возможно, и у диплоидов DD, но у тетрапло-
идов AADD— в двух локусах (С1\ и С/2). В геноме тетрапло-
идов эти гены располагаются следующим образом: Я2—С12 —
в геноме Л, a — С1} — в геноме D [802, 803].
При возникновении тетраплоидного хлопчатника гены Rt и
R2 и Cli и С12 были, несомненно, дупликатными факторами.
Однако у современного тетраплоидного хлопчатника они уже не
являются дупликатными; гены Я1 и R2 взаимодействуют как па-
ра комплементарных факторов; то же самое можно сказать и
относительно генов Cli и С12 [802, 803].
Эволюция в направлении более высоких уровней плоидности
усложняет мейоз и создает более жесткие ограничения при рас-
Гл. 25. Полиплоидный комплекс
337
щеплении и рекомбинации. Процесс диплоидизация противодей-
ствует этим тенденциям, давая возможность формам с высокой
или средней плоидностыо вести себя в генетическом отношении
как более или менее настоящие диплоиды.
Можно только предполагать, что диплоидизация — это фак-
тор, дающий возможность некоторым полиплоидным комплек-
сам эволюционировать до высоких уровней плоидности. Кроме
того, этот фактор, вероятно, обусловливает способность некото-
рых родов с высокими основными числами (например, Epilo-
bium, х=18; Equisetum, х=108) к усиленному видообразованию
на старых уровнях плоидности. Короче говоря, диплоидизация:
может вдохнуть в давно возникшие полиплоиды новую жизнь.
22—45
26 Природные автополиплоиды
Основными критериями, по которым у растений в природе мож-
но отличать автополиплоидов от амфиплоидов, служат, как уже
было отмечено в гл. 23, следующие: образование мультивален-
тов, стерильность, тетрасомное наследование и морфологическое
сходство с родственными диплоидами. Эти критерии полезны,
но их нельзя считать диагностическими, и в отдельных случаях
они легко могут привести к ошибочным заключениям.
Было время, когда на основе этих критериев считали, что
автополиплоидия широко распространена у растений в природе
Критическая переоценка фактических данных, предпринятая
Клаузеном, Кеком и Хиси [102], заставила изменить это заклю-
чение. По сравнению с амфиплоидией автополиплоидия в при-
роде— явление, необычное у высших растений, однако оно
встречается в ряде групп.
Тем не менее в литературе продолжают появляться сообще-
ния об автополиплоидии. Слишком часто такие сообщения бы-
вают основаны на недостаточных данных. Например, утвержде-
ние об автополиплоидном происхождении тетраплоидных пред-
ставителей группы Epilobium angusiifolium основано на цитоге-
нетических данных, весьма обширных и наводящих на размыш-
ления, но не вполне убедительных [586, 587].
К наиболее хорошо документированным примерам природной
автополиплоидии у растений относятся те, в которых можно с
достаточной уверенностью назвать родительские виды полипло-
идов и в которых известны также плодовитость при скрещива-
ниях и взаимоотношения между диплоидными родительскими
популяциями. Для получения таких данных необходимо, конеч-
но, комбинировать полевые и экспериментальные методы, по
своей разрешающей способности значительно превосходящие
обычные цитотаксономические методы.
В этой главе мы рассмотрим три подлинных случая природ-
ной автополиплоидии и вкратце упомянем еще о двух примерах.
Недостаток места не дает нам возможности рассмотреть раз-
личные недавние сообщения, в которых заключение об автопо-
липлоидии делается без достаточных на то оснований.
Группа Biscutella laevigata
Biscutella laevigata и близкие к ней виды (Cruciferae) — это
группа многолетних травянистых растений, встречающихся в
гористых местностях Европы. Классическими исследованиями
Гл 26 Природные автополиплоиды
339
Ментон [5.52, 553] было установлено, что среди них есть формы
с числом хромосом 2х, 4х и 6х (х=9) Гексаплоиды известны
только из одной местности в Испании. То, что нами известно на
сегодняшний день об этой группе, относится главным образом
к диплоидам и тетраплоидам.
Тетраплоидная форма Biscutella laevigata sensu stricto имеет
непрерывный ареал, встречаясь в Альпах на всем их протяже-
нии. Диплоиды, известные под общим названием В coronopifo-
На, образуют ряд разобщенных популяций на низменностях к
северу и к востоку от Альп. В виде исключения один диплоид-
ный вид встречается в горах в окрестностях Вены.
Диплоиды из отдельных районов различаются морфологиче-
ски, и прежде их описывали как разные виды. Однако Ментон
[553], скрестив их между собой, обнаружила, что гибриды Fi
плодовиты и что у них происходит нормальная конъюгация хро-
мосом Следовательно, эти диплоиды, по-видимому, представ-
ляют собой расы, принадлежащие к одному виду, а их тетрапло-
идное производное, В. laevigata, — межрасовый автотетраплоид.
У Biscutella laevigata иногда встречаются квадриваленты,
что наводит на мысль об автотетраплоидии, но не может слу-
жить решающим доказательством Более убедительным доводом
представляется обнаружение тривалентов у искусственных три-
плоидных гибридов, полученных при скрещивании В laevigata
с различными диплоидными расами [553].
Географическое и экологическое распространение этих ди-
плоидов и тетраплоидов в сочетании с тем, что нам известно
о геологической истории области, где они встречаются, позво-
ляет считать, что диплоидные популяции низменностей являют-
ся реликтами межледниковой флоры. Что же касается альпий-
ского тетраплоида Biscutella laevigata, то он распространен в
области, которая во время последнего оледенения находилась
подо льдом, и поэтом} он, вероятно, нмигрировал сюда в пос-
леледниковый период 1552, 553]
Межрасовые комбинации генов оказались, по видимому, цен-
ными в адаптивном отношении для этого автотетраплоида
В связи с этим можно предположить, что диплоидная раса,
встречающаяся в горах вблизи Вены, участвовала в качестве
одной из родительских форм в межрасовом скрещивании или
скрещиваниях, давших начало удачному альпийскому автотетра-
плоиду [102, 552, 553].
Группа Solatium tuberosum
Сложная группа Solatium tuberosum очень тщательно изучена
рядом исследователей и представляет собой наиболее хорошо
проанализированный пример природной автополиплоидии
22*
•340
Часть шестая. Полиплоцдия
Начнем с того, что обрисуем таксономическое толожение
этой группы. Секция Tuberarium рода Solatium представляет со-
бой большую секцию, состоящую из 32 рядов, распространенных
в Северной и Южной Америке [123]. К ряду Тuberosa относят-
ся 6 видов, в том числе S tuberosum sensu lato, встречающийся
в Перу, Чили и Аргентине. Пять из этих видов — дикорастущие
растения, а шестой, S. tuberosum, содержит дикорастущие расы
из Патагонии и Чили, а также многочисленные культурные расы,
выращиваемые в районе Анд. (по [123]; другой таксономический
подход к ряду Tuberosa- см. [354]).
Это свободно скрещивающиеся многолетние травянистые рас-
тения, образующие клубни. Диплоидные формы самонесовмес-
тимы и перекрестно опыляются шмелями и другими пчелиными;
помимо размножения семенами у них происходит вегетативное
размножение при помощи клубней [175]. У полиплоидов, выра-
щиваемых в культуре, и пыльца, и семена обычно стерильны, и
их размножают вегетативно [354, 812, 814].
Solarium tuberosum sensu lato —сложный таксономический
вид. Он содержит не только ряд дикорастущих и культурных
форм, о чем уже упоминалось, но также полиплоидный ряд, со-
стоящий из форм 2х, Зх, 4х и 5х при х = 12 [814].
В соответствии с классификацией культурного картофеля,
созданной Доддсом [174], существует семь главных форм Sola-
num tuberosum sensu lato, различающихся по морфологии, рас-
пространению и плоидности. Для этих таксонов Доддс исполь-
зует неформальную категорию «группа»; его группы соответст-
вуют культурным расам по другим классификациям Ниже пе-
речислены эти семь групп с указанием их плоидности и некото-
рых других черт.
1. Stenotomum. 2х. Высокие плато в Андах.
2. Phureja. 2х. Долины в Андах.
3. Andigena. 4х. Картофель, выращиваемый в Андах.
4. Tuberosum (=S. tuberosum sensu stricto). 4x Обычный
культурный картофель, Европа и Северная Америка..
5. Chaucha. Зх. Стерильные семена.
6 Juzepczukii. Зх. Стерильные семена
7. Curtilobum. Зх. Стерильные семена.
Филогенетические взаимоотношения между этими таксонами
схематически изображены на рис. 26.1 (по данным [174, 175]).
Кроме них на этой схеме указан вид Solarium acaule, принадле-
жащий к ряду Acaulia. Выращиваемый в Андах картофель andi-
gena считается тетраплоидным производным межрасовых скре-
щиваний между stenotomum (2х) и phureja (2х). Тетраплоид,
очевидно, возник спонтанно в условиях первобытного земледе-
лия. Следовательно, обыкновенный культурный картофель, так-
Гл. 26. Природные автополиплоиды
341
Рис. 26 1. Филогения основных культурных рас картофеля Solarium tubero-
sum [174]
Объяснения см в тексте
же тетраплоидный, представлеят собой продукт селекции этого
более примитивного андийского картофеля.
Обыкновенный картофель, на основе достаточно веских ци-
тогенетических данных, давно считали автотетраплоидом. У не-
го наблюдается высокая частота квадривалентов при мейозе.
Наибольшей частоты — 9 IV на клетку —они достигают у под-
разновидности (subvariety) Chippewa [812]. У некоторых под-
разновидностей средняя частота квадривалентов составляет
4,4 IV [384]. Такому цитологическому признаку соответствует
тетрасомия по некоторым генам, наблюдаемая у культурных
рас картофеля [78].
Данные из других источников окончательно решают вопрос
в пользу автотетраплоидии. Искусственный триплоид, получен-
ный в результате гибридизации между тетраплоидной расой tu-
berosum и диплоидной stenotomum, содержит в среднем 8,2 III
на клетку. Такая высокая частота тривалентов свидетельствует
342
Часть шестая Полиплоидия
о том, что культурный картофель — автотетраплоид, содержащий
хромосомы stenotomum [384].
Дальнейшие доказательства дает редуцированная или поли-
гаплоидная форма Solanum tuberosum sensu stricto, у которой
2п=24 вместо нормальных 2п=48. У некоторых полигаплоидов
наблюдается 12 II, т. е., иными словами, полная конъюгация в
большинстве клеток и высокая фертильность пыльцы. Кроме
того, эти полигаплоиды при скрещивании с некоторыми дипло-
идными культиварами дают плодовитых гибридов [384, 812].
Наконец, диплоидные расы очень близки генетически. Между
ними нет никаких барьеров, препятствующих скрещиванию.
У межрасовых гибридов Fi происходит нормальная конъюгация
с образованием бивалентов. В одних сочетаниях возникают пло-
довитые гибриды, тогда как в других сочетаниях образующиеся
гибриды имеют стерильную пыльцу [175,815].
В этом разделе внимание было сосредоточен^ на автотетра-
плоидии в группе Solanum tuberosum. Чтобы завершить карти-
ну, следует отметить, что среди дикорастущих • видов секции
Tuberarium амфиплоидия довольно обычна. Так, Solanum acaule
(ряд Acaulia) — аллотетраплоид, a S. demissum (ряд Demis-
sa) — аллогексаплоид [384, 812].
Группа Dactylis glomerata
Ежа сборная — Dactylis glomerata (Gramineae), 2м =14 и 28,—
самонесовместимое свободноопыляющееся растение, способное
также к вегетативному размножению. Эта группа широко рас-
пространена в Евразии1, и к ней относится обширный ряд мор-
фологических и экологических вариантов на двух уровнях пло-
идности [797].
На диплоидном уровне Стеббинс и Цохари [797] различают
11 подвидов; 4 из них представляют собой крайние варианты в
отношении морфологии. Они встречаются среди растительности
старых типов и имеют прерывистые ареалы, тянущиеся от Ги-
малаев до Канарских островов и островов Зеленого Мыса. Эти
4 географические расы, по-видимому, образовались в результа-
те Давней первичной дивергенции на диплоидном уровне.
Остальные 7 подвидов занимают промежуточное положение
между четырьмя крайними типами и связаны с более совре-
менными типами растительности, в том числе с пастбищными
субклимаксами и сорными местами. Они, возможно, представля-
ют собой результат скрещиваний и интерградации между пер-
вичными расами [797].
1 А также в Макаронезии, Северной Африке и Азии. — Прил. ред.
Гл 26. Природные автополиплоиды
343
Диплоидные расы интерфертильны. Фертильность пыльцы у
гибридов Fi колеблется от полустерильной до полностью фер-
тильной, а семена фертильны [797].
Тетраплоиды по сравнению с диплоидами встречаются чаще,
распространены более широко и крайне изменчивы Морфологи-
ческие и экологические данные позволяют считать, что тетра-
плоиды возникли от различных межрасовых гибридов Таким
образом, они являются межрасовыми амфиплоидами [797].
Скрещивания между тетраплоидной и диплоидной расами
ежи сборной удаются с’трудом, а их гибриды Fi высокостериль-
ны [797]. Dactylis glotnerata в настоящее время представляет со-
бой, таким образом, таксономический вид, состоящий из двух
биологических видов.
Барьер стерильности между диплоидами и тстраплоидами
эффективен, по не является абсолютным. Триплондные гибриды
при опылении пыльцой тетраплоидов завязывают некоторое ко-
личество семян. Наиболее мощные потомки от возвратных скре-
щиваний — тетраплоиды. Поэтому возможна интрогрессия от
диплоидов к тетраплоидпым популяциям через промежуточных
триплоидов. Такая интрогрессия — еще один фактор, вносящий
вклад в изменчивость тетраплоидной популяционной системы
[797].
Другие примеры
Мы приведем здесь еще два примера природных авюполипло-
идов с указаниями па литературу, в которой можно найти даль-
нейшие подробности.
Medicago sativa, 2«=16 и 32 Диплоидная форма М sativa
( = М coerulea)—дикорастущее растение, распространенное от
Турции до Ирана и Каспийского моря. Тетраплоидная форма
(М. sativa sensu stneto) — культурное растение, возникшее, по-
видимому, спонтанно в области распространения диплоида, а
впоследствии распространившееся в качестве культурной формы
в Европе [465].
Galax aphylla (Diapensiaceae), 2п=12 и 24. Единственный
таксономический вид монотипического рода, распространенный
в юго-восточной части США. Во многих местах по всему ареалу
этого таксономического вида можно встретить растущие рядом
диплоидные и тетраплоидные индивидуумы [41]. Такая картина
сильно склоняет к предположению об автотетраплоидии [102,
778, 779, 789].
Часть седьмая
Анеуплоидия
27 Анеуплоидия
Различия между индивидуумами или популяциями (или клетка-
ми) по числу хромосом выражаются в трех основных формах:
полиплоидия, анеуплоидия и агматоплоидия. Если полиплоидия
представляет собой различия по числу целых хромосомных на-
боров, то анеуплоидией называют различия по числу отдельных
хромосом, а агматоплоидией — различия по числу хромосомных
участков.
К анеуплоидной изменчивости относится очень разнородная
группа явлений. Отклонения от стандартного числа хромосом
могут быть как в сторону увеличения, так и в сторону уменьше-
ния. Эти численные отклонения могут затрагивать одну хромо-
сому (полисомия, моносомия), одну пару хромосом или несколь-
ко их пар. Такие лишние хромосомы могут быть гомологами
членов нормального комплемента или не иметь к ним отношения
(добавочные, сверхкомплектные, или В-хромосомы). Кроме того,
анеуплоидия может возникать на диплоидном или полиплоид-
ном уровнях.
Анеуплоидия на полиплоидном уровне включает несколько»
различных явлений. Различные попарные комбинации разных
диплоидных основных чисел создают анеуплоидную изменчи-
вость среди аллотетраплоидных производных. Так, два вида с
8 парами хромосом каждый дают аллотетраплоид с 16 парами
хромосом, тогда как скрещивание между видом с 8 и видом с
9 парами дает аллотетраплоид с 17 парами хромосом. У многих
полиплоидов в результате нарушений мейоза возникают сверх-
комплектные пары хромосом. Кроме того, у полиплоидов возмо-
жен полиплоидный сброс, т. е. нисходящая (descending) анеу-
плоидия.
У одного и того же отдельного растения или в одной и той
же популяции можно встретить одновременно различные типы
анеуплоидной изменчивости. В данной группе растений может
наблюдаться анеуплоидия и на диплоидном, и на полиплоидном
уровнях. В одной и той же группе растений анеуплоидия может
также встречаться одновременно с агматополиплоидией.
Гл 27 Анеуплоидия
345
В этой главе мы рассмотрим несколько примеров обычной
анеуплоидии на полиплоидном и диплоидном уровнях. Полисе-
мии и В-хромосом мы здесь касаться не будем (см. [434]).
Нисходящая анеуплоидия на диплоидном уровне
Нисходящие ансуилоидные ряды на диплоидном уровне изве-
стны у довольно большого числа групп растений Стеббинс
(780] перечислил 25 таких групп. В гл. 14 мы привели несколько
примеров нисходящей анеуплоидии наряду с другими относя-
щимися к этому явлению сведениями Ниже представлено не-
сколько других примеров; цифры в этом списке соответствуют
гаплоидному числу хромосом (п), а скобками ( ) обозначены
необнаруженные промежуточные числа
Crepis (Compositae), 6-5-4-3 [33]
Youngia (Compositae), 8-( )-5 [36]
Eriophyllutn и Pseadobahia (Compositae), 8-7 ( )-4-3 [82]
Nicotiana (Solanaceae), 12-( )-10-9 [250]
Phacelia (Hydrophy llaceae), 11-10-9-8-7 [90, 91]
Clarkia (Onagraceae), 7-6-5 [492]
Knautia (Dipsacaceae), 10-( )-8 [191]
Cephalaria (Dipsacaceae) 9-( )-5 [191]
Lesquerella (Cruciferae), 8-7-6-5 [693]
Persoonia и родственные роды (Proteaceae) 7-( )-5 [413. 414]
Fritillaria (Liliaceac) 12-( )-9[144, 148]
Иногда нисходящие анеуплоидпые ряды бывают образованы
родами, принадлежащими к какому-либо одному семейству или
трибе Ограничимся двумя примерами В трибе Polemonieae сем
Polemoniaceae наблюдается ряд 9-8-7-6, а в сем Loranthaceae—
ряд 12-11-10-9-8 [42, 281].
Среди покрытосеменных у 783 видов диплоидное число хро-
мосом низкое — от л = 2 до л = 6 Распределение видов по числу
хромосом выглядит следующим образом [282]:
л = 6
л — 5
л = 4
л = 3
и-2
У » 408 видов
225 »
» 134 »
» 15 »
» 1 вида
Все эти виды представляют собой результат редукции основного
числа хромосом в процессе филогенеза
Механизм анеуплоидпой редукции на диплоидном уровне
связан с неравными реципрокными транслокациями [33, 610,
834]. В предковом хромосомном наборе хромосомы А и В обме-
346
Часть седьмая. Анеуплоидия
ниваются участками, очень неравными по длине, так что весь
генетически активный материал хромосомы А переносится в хро-
мосому В, тогда как центромера хромосомы А получает в об-
мен лишь небольшой инертный участок. Затем ставшая инерт-
ной хромосома А утрачивается в процессе одного из клеточных
делений. В результате получается хромосомный набор, содержа-
щий на одну пару хромосом меньше.
Классическое описание этого механизма дал Тогби [834] на
основе цитологического анализа Crepis neglecta (2п=8) и С. fu-
liginosa (2п=6). Последний вид с редуцированным числом хро-
мосом происходит от предкового вида, близкого к С. neglecta.
Отдельные хромосомы у этих видов можно идентифицировать в
соматических и в мейотических клетках по величине и морфоло-
гии. Четыре хромосомы в наборе С. neglecta ^обозначены А, В,
С, D, а три хромосомы С. fuliginosa — A, В, D (рис. 27.1).
А
Б
Рис. 27.1. Хромосомные наборы двух видов Crepis и гибрида между ни-
ми [834].
А. Гаплоидный набор соматических хромосом С. neglecta.
Б. Гаплоидный набор соматических хромосом С. fuliginosa.
В Диплоидный набор соматических хромосом гибрида Fi,
Г Конъюгация мейотических хромосом в метафазе I у гибрида Fi.
Гл 27. Анеуплоидия
347
Конфигурации пар, получающихся при конъюгации хромо-
сом у межвидовых гибридов, показывают, что эти два вида раз-
личаются по трем инверсиям, по одной транслокации в хромо-
сомах А и D и по другой транслокации в хромосомах В и С.
Особенно существенна эта вторая транслокация- одна часть
хромосомы В вида С. fuiiginosa конъюгирует с хромосомой В
вида С neglecta, а другая ее часть — с хромосомой С этого по-
следнего (рис 27 1) Это показывает, что хромосома В вида
С. fuiiginosa содержит участки, гомологичные хромосоме С ви-
да С neglecta.
В наборе С. fuiiginosa пет хромосомы С как таковой, и соот-
ветственно основное число хромосом у этого вида меньше; одна-
ко генетический материал, имевшийся у предковых форм хро-
мосомы С, содержится у С. fuiiginosa в хромосоме В Это про-
изошло в процессе филогенеза в результате неравной реципрок-
ной транслокации между предковыми хромосомами В и С, ко-
торая привела к образованию новой составной хромосомы В у
С. fuiiginosa [834].
Восходящая анеуплоидия па диплоидном уровне
У ряда групп растений обнаружена также восходящая анеупло-
идия на диплоидном уровне. Ниже перечислено несколько при-
меров; как и в предыдущем перечне, цифры соответствуют чис-
лам п, а недостающие числа обозначены скобками ( ).
Fritillaria (Liliaceae), 12-13 [144, 148]
Narcissus (Amaryllidaceae), 7-( )-10-ll [212]
Crocus (Iridaceae), 12-13-14-15 [69]
Phacelia (Hydrophyllaceae), 11-12-13 [90, 91, 119]
Clarkia (Onagraceae), 7-8-9 [492]
Cephalaria и Succisia (Dipsacaceae), 9-10 [191]
Неудивительно, что существуют анеуплоидные ряды, кото-
рые одни исследователи считают восходящими, а другие — нис-
ходящими. У Podocarpus (Podocarpaccae) с гаплоидными чис-
лами 10-11-12-13-( )-17-18-19, по мнению Кхошу [433], в про-
цессе филогенеза имела место восходящая анеуплоидия, а по
мнению Хейра и Бейценберга [327], — нисходящая анеуплоидия.
В трибе Astereae (Compositae) есть формы с гаплоидными чис-
лами 4-5-6-( )-8-9 Согласно одной школе исследователей, ос-
новное число хромосом у предковых форм х=5 (или 4) [844,
846]; согласно другой школе, исходное основное число х = 9
[720, 760].
Одним из способов возникновения анеуплоидного увеличения
числа хромосом может быть неправильное разделение центроме-
ры. Будучи сложной структурой, центромера редко делится по-
348
Часть седьмая. Анеуплоидия
I 2 3 4
С. biloba « I ..U I
I 2 3 4
С. lingulate t -ti i
4
Рис. 27.2. Гомологии некоторых хромосом Clarkia biloba и С. Ungulata и кон-
фигурация, образующаяся при их конъюгации [499].
перечно с образованием двух половинок, каждая из которых со-
храняет способность к координации с веретеном. В таких слу-
чаях одна метацентрическая (или субметацентрическая) хромо-
сома дает начало двум телоцентрическим (или субтелоцентри-
ческим) хромосомам [144, 148].
У Frttillaria основное число х=12 и хромосомный набор со-
стоит из 10 телоцентрических и 2 метацентрических хромосом.
Frttillaria pudica, у которой и ==13, содержит 12 телоцентриче-
ских и 1 метацентрическую хромосому. Следовательно, анеупло-
идное увеличение от п=12 до п=13 произошло, вероятно, в ре-
зультате превращения одной метацентрической хромосомы в две
телоцентрические [144, 148]. Подобным же увеличением числа
телоцентрических хромосом обусловлено анеуплоидное увеличе-
ние у Narcissus и Crocus [69, 212].
. Другой механизм анеуплоидного увеличения числа хромосом
сострит во включении в нормальный хромосомный набор сверх-
комплектной пары хромосом в результате отставания во время
клеточного деления; это приводит к образованию тетрасомика
(2п-|-2). Сверхкомплектные или тетрабомные хромосомы могут
затем дивергировать от своих гомологов путем взаимообменов
с другими негомологичными хромосомами и в результате дивер-
генций в функциях генов.
Исходное основное число у Clarkia (Onagraceae) х=7, а ви-
ды с 8 и 9 парами хромосом возникли позднее (492]. Анеуплоид-
ное увеличение у Clarkia, по крайней мере в некоторых случаях,
Гл. 27. Анеуплоидия
349“
произошло путем включения в набор тетрасомных хромосом,
претерпевших транслокации.
Люис и Робертс [499] провели подробное цитогенетическое
исследование пары близкородственных видов — Clarkia biloba
(п=8) и С. Ungulata (п=9). Эти авторы установили, что второйг
из указанных видов произошел от первого Конъюгация хромо-
сом у гибрида между этими видами свидетельствует о том, что
их геномы различаются по двум транслокациям и одной инвер-
сии, но в остальном гомологичны. Одна из транслокаций нахо-
дится в девятой хромосоме С Ungulata
Девятая хромосома С. Ungulata вместе с двумя другими хро-
мосомами этого вида и двумя хромосомами С biloba образует
у гибрида цепь из 5 хромосом Конфигурация этой цепи и ее
состав представлены на рис 27.2. Очевидно, девятая хромосома
возникла в результате обмена плечами между двумя хромосо-
мами С. biloba с терминальными участками 1-2 и 3-4, что при-
вело к образованию новой хромосомы 1-4 Затем две исходные
и одна новая хромосома закрепились у предка С. Ungulata Та-
ким образом, в наборе этого последнего на одну пару хромо-
сом больше, чем у предкового С. biloba [499].
Полиплоидный сброс
Анеуплоидная редукция па полиплоидном уровне была удачно
названа Дарлингтоном полиплоидным сбросом (polyploid drop
[144, 146, 148]). Полиплоид часто может выдержать утрату од-
ной или нескольких пар хромосом ввиду наличия у пего дупли-
катных факторов в гомологичных или гомеологичпых хромосо-
мах.
Дарлингтон [148] приводит примеры полиплоидного сброса
у представителей 20 групп растений. Некоторые из них перечис-
лены ниже; числа п даны в следующем порядке: диплоиды, по-
липлоиды и редуцированные апсуплоиды.
Iles peris (Cruciferae), 7-14-13-12
Linum (Linaceae), (8, постулировано)-16-15
Veronica (Scrophulariaceae), 7-21-20
Maydeae (Gramineae), 5-10-9
У Proteaceae анцестральное основное число x = 7; полипло-
идное удвоение до х=14 произошло в ранний период истории
этого семейства (в примитивном подсемействе Persoonioideae)
Это давнишнее тетраплоидное число хромосом встречается так-
же в производных подсемействах Protcoideac и Grevilleoideae,
В подсемействе Proteoideae имеется, кроме того, нисходящий
ряд х = 14-13-12-11-10, a v Grevilleoideae — ряд х= 14-13-11-10'
[414].
350
Часть седьмая. Анеуплоидия
Анеуплоидное увеличение у полиплоидных
гибридов
Межвидовые гибриды с унивалентными хромосомами, не имею-
щими гомолога при мейозе, представляют собой источник анеу-
плоидной изменчивости в экспериментальных условиях и в
меньшей степени — в природных популяциях. Анеуплоидия обыч-
но бывает возрастающей, поскольку приобретение сверхкомп-
лектной хромосомы переносится легче, чем утрата членов ос-
новного набора. Эту форму анеуплоидии чаще можно обнару-
жить на полиплоидном уровне, чем на диплоидном, вследствие
более высоких буферных свойств полиплоидных генотипов.
Хорошим примером служит скрещивание между гибридом и
•его потомками в группе Erophila verna (Cruciferae) [901]. Этот
пример уже был рассмотрен в гл. 20; здесь мы упомянем о нем
лишь вкратце в связи с анеуплоидией.
Гибрид был получен в результате скрещивания между фор-
мой с 15 парами и формой с 32 парами хромосом. У гибридов
F! наблюдались многочисленные неконъюгировавшие хромосо-
мы. В Fg произошло расщепление по числу хромосом и по дру-
гим признакам.
В последующих инбредных поколениях выщепенцы выдели-
лись в различные стабильные линии. Как показал цитологиче-
ский анализ восьми таких стабильных линий, они содержали
следующие анеуплоидные наборы хромосом: п=22, 23, 25, 29, 31
и 34. Анеуплоидия встречается также в природных популяциях
труппы Erophila verna в Европе, и вполне возможно, что она
возникла у них таким же образом (901].
Два тетраплоидных вида Gilia (G. malior и G. modocensis)
•с одинаковым числом хромосом (2п=36) дали при скрещивании
гибрид Fi, у которого при мейозе наблюдалось много неконъю-
тировавших хромосом. Этот гибрид в свою очередь дал несколь-
ко гомоплоидов, а также один стабильный анеуплоид с 2п=
= 50 (287].
Анеуплоидные числа хромосом гибридного происхождения у
полиплоидов иногда закрепляются благодаря агамоспермии. На-
пример, анеуплоидная изменчивость в группе Bouteloua curtipen-
dula (Gramineae) обусловлена сочетанием полиплоидии, гибри-
дизации и агамоспермии [261, 262]. Эти и другие сходные при-
меры будут рассмотрены в главах, посвященных агамоспермии
(см. часть восьмую).
Carex
Род Carex привлекает внимание своим длинным и почти непре-
рывным анеуплоидным рядом — от п=6 до и=56, — который со-
держит подряд все гаметные числа от п=12 до п=43 (156, 361,
Гл. 27. Анеуплоидия
351
380] Короткие последовательные анеуплоидные ряды (sensu la-
to) обнаружены в нескольких секциях Carex. Примерами слу-
жат Distantes (п = 28—37), Extensae (п = 30—35), Acutae
(п = 34—42) и группа Carex cariophyllea (п=31—34) [156].
У Carex известны основные числа х=6, 7, 8 и 9; предполага-
ется, что у этого рода встречается также х = 5, которое, возмож-
но, было первоначальным основным числом [371, 534, 867].
Кроме того, у Carex существуют полиплоиды с основным чи-
слом х = 5 и другими вторичными основными числами Так, в
группе С. siderosticta есть формы с п = 6 и п=12, в секции Capil-
lares (х=9) есть диплоидные, тетраплоидпые и гексаплоидные
виды, а в группе С stenaniha — формы с п=17 и п = 34. У Ca-
rex описаны полиплоидные ряды, основанные па х = 5, 6, 7, 8 и
9 [534, 867]. Таким образом, часть наблюдаемой у Carex апеу-
плоидпой изменчивости можно отнести за счет различных по-
бочных эффектов полиплоидии [534, 867]. Монобазическая поли-
плоидия с различными основными числами даст сходные, но
разные производные числа, а дибазическая полиплоидия запол-
нит разрывы Так, диплоиды с 7 и 8 парами хромосом в разных
комбинациях дадут ряд тетраплоидов с п=14, 15 и 16. Этот ряд
может продолжаться в обоих направлениях за счет полиплоид-
ных сбросов и приобретений
Кроме того, у рода Carex происходит естественная гибриди-
зация Друри [179] сообщает, что па Аляске почти все виды сек-
ции Vesicariae участвуют в гибридизации. В природе известны
гибриды между Carex rostrata, с одной стороны, и С. rotundata,
С. metnbranacea и С physocarpa — с другой Из скопления гиб-
ридов, возникшего в результате скрещиваний между С rostrata
и С. rotundata в одной из частей Аляски, выщеплястся новая
константная и плодовитая форма, численность которой возрас-
тает и которая развивается в новый вид гибридного происхож-
дения, получивший название С. paludivagans [179].
Большая часть анеуплоидной изменчивости у Carex обуслов-
лена гибридизацией и полиплоидией, однако у Carex, по-видимо-
му, встречается также агматоплоидия, о чем будет сказа-
но в гл. 28.
Claytonia virgirtica
Claytonia virginica (Portulacaceae) — небольшое многолетнее
травянистое клубнелуковичнос растение, широко распространен-
ное в центральных и восточных областях Северной Америки.
В этой группе имеется обширный ансуплоидный ряд. Ротуэлл
[698] и Люис [503] обнаружили в ней формы со следующими
значениями 2п: 12, 14, 16, 17, 18, 19, 20, 22, 24, 26, 28, 30, 31, 32,
352
Часть седьмая. Анеуплоидия
и:!): %.
2/7 = 12
2/7 =14
2л = 16
2п = 17 2п = 18 2л=19
2/7=26 2л=28
2л=20
2л = 30
2л=32
м.
Ш»"
2л=34
Рис. 27.3 Анеуплоидиая изменчивость у Claytonia virginica (по {698] с из-
менениями).
Хромосомы по большей части находятся в метафазе I мейоза Неконъюги-
ровавшие хромосомы белые, мультиваленты выделены точками.
.34, 36, 41, 48, 72. Некоторые из соответствующих этим числам
хромосомных наборов изображены на рис. 27.3.
Первоначальным основным числом в этой группе было, ве-
роятно, х=8 [698]. Изменчивость по числу хромосом можно све-
сти к нескольким компонентам диплоидной и полиплоидной
-анеуплоидии. Прежде всего в ней имеется нисходящая анеупло-
идия на диплоидной основе: от х = 8 к 7 и к 6. Затем в резуль-
тате разных попарных комбинаций этих диплоидов возникает
ряд тетраплоидов с числами п, равными 16, 15, 14 и, возможно,
12. Число и, равное 12, обнаружено также у триплоидов (х=8,
2п=3х=24), а форма с п=11—также триплоид. Большие чис-
Гл. 27. Анеуплоидия
353
ла соответствуют полиплоидам более высокого уровня, 48-хро-
мосомная форма — это 6х, а 72-хромосомная — это, вероятно, 9х
Наконец, наблюдается некоторая анеуплоидная изменчивость
вблизи четных полиплоидных уровней; примером служит п = 17
и 18 [698].
Анеуплоидная изменчивость иногда наблюдается в пределах
разных популяций В одной из популяций в Картидже (шт Те-
хас) число хромосом колебалось от 2/г = 24 до 2п=41; чаще все-
го встречались числа 2п=24—26 и 28—32. Оказалось, что в этой
популяции модальные числа различны в разные годы, а в по-
пуляции Септ-Луиса — ранней весной и летом [503, 504]. Иног-
да наблюдаются различия в числе хромосом в разных частях
одного и того же растения. Например, у одного индивидуума в
популяции Картиджа в клетках стебля и в материнских клетках
пыльцы 2п = 30, а в клетках корня 2п = 28 [506].
В восточной части Северной Америки широко распростране-
ны диплоиды (х=8), тетраплоиды (х = 14, 15, 16) и триплоиды
(х= 11, 12), причем они в значительной мере симпатричны Дип-
лоиды, у которых х — 7 и 6, имеют более ограниченное распрост-
ранение [507]. Эти формы рассматриваются как «цитотипы» и
«расы» одного вида [507]. С эволюционной точки зрения, одна-
ко, Claytonia virginica следует рассматривать как нспроанализи-
рованую полиплоидную группу видов, на которую наложилась
анеуплоидия.
Высокие основные числа у покрытосеменных
У многих родов и некоторых семейств древесных покрытосемен-
ных основные числа высокие и лежат в интервале от х=12 до
х = 21. Вопрос о происхождении таких высоких основных чисел
обсуждается чрезвычайно широко В качестве одного из воз-
можных объяснений нередко предлагали давно возникшую по-
липлоидию.
Чтобы облегчить анализ этой проблемы, удобно разбить вы-
сокие основные числа на две подгруппы Довольно высокие чис-
ла (х=19—21) встречаются у Magnoliaceae, Trochodendraceae,
Hippocastanaccae, Platanaceae и др Умеренно высокие числа
(х=12—16) обнаружены в некоторых других семействах древес-
ных растений, например у Fagaceac (х=12), Асегасеас (х=13),
Betulaceae (х=14), Ulmaceae (х=14), Juglandaceae
(х= 16) и др.
По мнению ряда авторов [144, 780, 832}, очень высокие основ-
ные числа первой подгруппы (х = 19—21) — результат древней
полиплоидии. Я согласен’ что в данном случае это наиболее ве-
роятное заключение (см. гл. 22 и 25).
23-45
354 Часть седьмая. Анеуплоидия
Рассмотрим далее подгруппу с умеренно высокими основны-
ми числами (х=12—16). Стеббинс [775, 778, 780, 787] считает,
что это также древние полиплоидные числа. Альтернативное
объяснение, которым пренебрегают, состоит в том, что такие
числа — результат восходящей анеуплоидии [293].
Это подводит нас к вопросу о первоначальном основном чис-
ле у покрытосеменных Высказывалось предположение [191, 282,
283], что оно лежит в пределах х=7—9. У многих семейств по-
крытосеменных, в том числе у некоторых примитивных, таких,,
как Аппопасеае [869], встречаются одновременно гаплоидные
числа п = 7, 8 и 9, которые, по-видимому, и представляют собой
основные числа для этого класса.
Многие исследователи [143, 150, 191, 194, 325, 659, 664, 665,
869] считают, что х=7 является первоначальным основным чис-
лом. Я, однако, не вижу никаких серьезных причин, по которым,
следовало бы выбрать именно х=7. Сколько-нибудь надежным
заключением, вытекающим из имеющихся данных, представля-
ется все же допущение о неком первоначальном основном числе
в пределах х=7—9*\
Приняв х=7—9 в качестве исходной точки, можно легко
представить себе, что такие умеренно высокие числа, как х= 12,
13 и 14, могли быть древнего тетраплоидного происхождения в
результате полиплоидного сброса или без него. Но их с равным
успехом можно считать и продуктами восходящей анеуплоидии.
Существует ряд причин, по которым следует ожидать, что
восходящая анеуплоидия имеет место во многих группах лесных
деревьев и кустарников. Высокое диплоидное число хромо-
сом— один из способов максимизировать генерацию рекомби-
национной изменчивости в популяции растений, а экологические
условия, существующие в стабильных лесных сообществах, бла-
гоприятствуют открытым рекомбинационным системам. Поэто-
му доминантные и субдоминантные лесные виды часто подвер-
гаются отбору на анеуплоидное увеличение. Следовательно,
можно ожидать, что многие древесные растения с высокими чис-
лами хромосом — настоящие диплоиды.
о А. Тахтаджяном (Происхождение и расселение цветковых растений,
сНаука», 1970, с 35) было высказано предположение, что наиболее вероят-
ное основное число хромосом у ранних цветковых растений было 6. По Стеб-
бинсу (Stebbins G. L., Flowering plants, evolution above the species level, 1974,
Harvard University Press, Cambridge, Massachusetts, p 206) первоначальные
основные числа могли быть 6, 7 или 8 — Прим. ред.
28 Агматоплоидия
Анеуплоидия (sensu stricto) означает изменение числа целых
хромосом. В отличие от этого агматоплоидией называют раз-
личия по числу независимо образующихся пар хромосомных
фрагментов в данной группе [534, 550].
Эти два типа изменения числа хромосом коррелируют с ти-
пом центромеры. У большинства растений и животных плечи
хромосом прикреплены к локализованным центромерам, вос-
производящим себя и свои хромосомы на протяжении ряда кле-
точных поколений. Приобретение или утрата одной или несколь-
ких пар хромосом данным набором зависит от приобретения или
утраты соответствующих центромер Точно так же плечи или
фрагменты хромосом могут сохраниться в данной клеточной ли-
нии или популяции только в том случае, если они прикреплены
к центромере Изменения числа хромосом бывают анеуплоид-
пыми (sensu stricto) или полиплоидными
У небольшого числа групп растений и животных центромеры
диффузные. Активность таких центромер не локализована в ка-
ком-то одном участке, а распространяется по всей длине данной
хромосомы Хромосомы, содержащие диффузные центромеры,
получили название полицентрических [61, 509]. Фрагменты, об-
разующиеся при поперечных делениях полицентрической хромо-
сомы, могут воспроизводиться на протяжении ряда клеточных
делений, если их разорванные концы заживляются; таким обра-
зом, они могут стать постоянными компонентами данной попу-
ляции. Диффузные центромеры создают возможность для воз-
никновения агматоплоидии [509, 534].
Агматоплоидныс ряды нельзя отличить от собственно анеу-
плоидных рядов, если располагать лишь простым перечнем хро-
мосомных чисел Как в том, так и в другом случае можно встре-
тить 2п = 10, 12, 14
Таким образом, термин анеуплоидия в его общем значении
все еще остается полезным в описательных целях, когда мы
имеем дело с изменчивостью числа хромосом, не связанной с
полиплоидией. Явление, известное ныне под названием агмато-
плоидии, прежде относили к анеуплоидии, однако после уста-
новления его природы и его особых черт оно было выделено как
самостоятельное явление [550]. В результате такого терминоло-
гического выделения анеуплоидия, сохраняя свое первоначаль-
ное широкое значение, приобрела вместе с тем и более узкий
смысл В этой главе нам придется использовать термин анеупло-
идия как в узком, ыкп в общем его значении, указывая всякий
раз. что именно имеется в виду
23*
356
Часть седьмая. Анеуплоидия
Из всего того, что будет сказано в этой главе, станет ясно,
что наши представления об агматоплоидии пока еще находятся
на стадии первых поисков. Цитогенетические данные во многих
случаях остаются спорными. Связь между агматоплоидией и ви-
дообразованием неясна. Мы не знаем, как достигается репродук-
тивная изоляция между разными агматоплоидными популяция-
ми. И нам непонятно, в чем заключается адаптивная ценность
агматоплоидных изменений.
Полицентрические хромосомы
В царстве животных хромосомы с диффузными центромерами
обнаружены у некоторых Hemiptera, или клопов, у Coccidae,
Aphidae и Heteroptera, а также у клещей [509]. Среди высших
растений диффузные центромеры встречаются у Juncaceae (Lu-
zula) и Сурегасеае (Eleocharis, Scirpus, Carex); предполагают,
что они имеются также у Zingiberaceae и Musaceae (обзор и ли-
тературу см. [534]).
Полицентрические хромосомы, содержащие диффузные цент-
ромеры, отличаются по морфологии и поведению от хромосом
обычного типа с локализованной центромерой. Вспомним, что
у обычных хромосом в метафазе митоза наблюдается ясно вы-
раженная центромерная перетяжка и что эти центромеры вы-
страиваются на метафазяой пластинке, а плечи хромосом сво-
бодно располагаются в разных положениях. У полицентрических
хромосом таких перетяжек нет, а вдоль метафазной пластинки
выстраиваются целые хромосомы. Когда в анафазе обычные
хромосомы перемещаются к полюсам, то впереди находится
центромера, которая тянет за собой плечи хромосомы. Поли-
центрические хромосомы, напротив, при перемещении к полюсу
бывают расположены примерно в одной плоскости, а иногда их
концы даже опережают среднюю часть |[534].
На рис. 28.1 можно видеть упомянутые выше особенности
полицентрических хромосом на примере делящихся клеток
Luzula campestris (Juncaceae) [72].
Возникал вопрос о том, распределяется ли активность цент-
ромеры по длине полицентрической хромосомы непрерывно или
прерывисто? Иными словами, является ли диффузность центро-
меры полной или неполной? Данные по Luzula purpurea свиде-
тельствуют в пользу второй возможности. Хромосомы этого ви-
да, по-видимому, содержат многочисленные центромерные уча-
стки [61].
Хорошо известно, что ацентрические фрагменты обычных
хромосом имеют тенденцию к отставанию во время клеточного
деления и рано или поздно утрачиваются жизнеспособными кле-
точными линиями. У организмов с диффузными центромерами
можно ожидать, однако, что фрагменты хромосом завершат нор-
Гл 28 Агматоплоидия
357
малыше метафазные и анафазные движения. Хоканссоп инду-
цировал у Eleocharis palustris (Сурсгасеае) разрывы хромосом
при помощи рентгеновских лучей. Он обнаружил, как и ожида-
лось, что в метафазе фрагменты хромосом, даже мелкие, рас-
полагались по экваториальной пластинке, а в анафазе направля-
лись к полюсам [330, 331].
Рис 28.1. Митоз у Luzula campestris (2n=12) [72]
Вил сбоку; видно расположение хромосом в метафазе и раппей анафазе.
Спонтанные разрывы хромосом у организмов с полицентриче-
скими хромосомами могут, следовательно, привести к возникно-
вению восходящих агматоплоидных рядов. У данной группы
видов может наблюдаться значительная изменчивость по числу
хромосомных пар, а наборы, состоящие из большого числа хро-
мосом, будут содержать по крайней мере несколько более мел-
ких членов [330, 534].
Это не означает, что агматоплоидия является неизбежным
результатом фрагментации полицентрических хромосом В ее
возникновении, несомненно, участвуют и другие факторы, как
цитологические, так и селективные. Уайт [889, 890J обратил вни-
мание на один, возможно важный, цитологический фактор, а
именно на роль теломеры, или терминального участка, хромосо-
мы. Если хромосомные фрагменты не имеют нормально зажив-
ленных концов, то они не могут долго оставаться жизнеспособ-
ными |[889, 890].
Фактически анеуплоидия (sensu lato) встречается не во всех
группах организмов, обладающих полицентрическими хромосо-
358
Часть седьмая. Анеуплоидия
мами. Среди Heteroptera с диффузными центромерами у неко-
торых семейств (Pefitatomidae и Corixidae) число хромосом от-
личается большим постоянством, тогда как в других семействах
оно колеблется в довольно широких пределах [889]. Разные ви-
ды Luzula, Eleocharis и Carex сохранили низкое основное число
хромосом [889]. У Eleocharis анеуплоидия (sensu lato) встреча-
ется относительно редко.
Carex
У рода Carex, как уже отмечалось в гл. 27, анеуплоидия (sensu
stricto) распространена достаточно широко, однако есть данные
о том, что у этого рода встречается также и агматоплоидия.
В хромосомах Carex нет ясно различимой локализованной
центромеры. Судя по морфологии хромосом, они, по-видимому,
обладают диффузной центромерой. Кроме* того, в анафазе при
движении к полюсам эти хромосомы часто бывают направлены
концами вперед. Все это указывает на полицентричность хромо-
сом и позволяет предполагать, что наблюдаемую изменчивость
по числу хромосом, возможно, частично обусловливает фрагмен-
тация (156, 534].
Хейльборн [360, 361] сравнивал Carex pilulifera и С. ericeto-
rum в отношении общей длины и числа соматических хромосом.
Он обнаружил, что у вида с более высоким числом хромосом —
С. ericetorum (п=15) —хромосомы короче, чем у вида с более
низким их числом — С. pilulifera (п=9). Число длинных хромо-
сом в наборе снижается по мере увеличения общего числа хро-
мосом [360, 361]. Более поздние исследователи подтвердили это
наблюдение на других видах группы Carex. У видов с более вы-
сокими числами хромосом хромосомы обычно короче, чем у
близких им видов с низким числом хромосом (156, 534].
Обратная зависимость между числом хромосом и их длиной
может быть обусловлена помимо фрагментации и рядом других
факторов. Следовательно, эта группа данных не может служить
доказательством того, что фрагментация и агматоплоидия уча-
ствовали в наблюдаемой у Carex изменчивости числа хромосом.
Однако зависимость между величиной и числом хромосом в со-
четании с наличием полицентрических хромосом и необычайно
широким развитием анеуплоидии (sensu lato) сильно склоняет
к мнению о том, что в видообразовании у рода Carex участвова-
ла агматоплоидия.
Luzula
Как показали Малейрос и др. [549], Норденшельд [619, 622]
и Браун (72], хромосомы Luzula (Juncaceae) обладают диффуз-
ными центромерами. Эти и другие авторы обсуждали возмож-
Гл 28 Агматоплоидия
359
ность того, что фрагментация хромосом участвовала в создании
различий по числу хромосом у этого рода
Число хромосом у Luzula колеблется от 2л = 6 у L purpurea
до 2п=66 и 72 у L. pilosa Число 2п=12 обнаружено у многих
видов и представляет собой основное число для данного рода.
У этого рода есть обыкновенный полиплоидный ряд, основанный
на х = 6, достигающий октоплоидпого уровня [619].
У Luzula наблюдается также анеуплоидия (sensu lato). Так,
у L. spicata обнаружены смежные числа п = 6 и 7, а у L orest-
га — п=10 и 11 В каждом случае набор с более высоким чис-
лом хромосом содержит две маленькие хромосомы, соответству-
ющие одной большой хромосоме в наборе с меньшим числом
хромосом Таким образом, анеуплоидпое увеличение, вероятно,
обусловлено фрагментацией и агматонлоидией [619, 620].
Кроме того, Норденшельд [619] обнаружил у этой группы
полиплоидные ряды особого типа, в которых общая длина всех
хромосом остается примерно постоянной Отдельные хромосомы
данного тетраплоида примерно вдвое меньше хромосом диплои-
дов, а хромосомы октоплоида примерно вдвое меньше, чем у те-
траплоидов. Такие соотношения наблюдаются в группах L cam-
pestris и spicaia. Норденшельд объясняет эти одновременные
изменения числа и величины фрагментацией всех хромосом
предкового набора. Численные соотношения между видами но-
сят полиплоидный характер, однако удвоение происходит, воз-
можно, путем фрагментации, а не умножения, и возникающие
при этом ряды представляют собой агматоплоидию особого ти-
па [618—621].
Эта гипотеза относительно агматоплоидии подтверждается
поведением крупных и мелких хромосом при их конъюгации
друг с другом у межвидовых гибридов, полученных при скрещи-
вании видов, различающихся по уровню плоидности. Luzula cam-
pestris и L pallescens содержат по 6 пар больших хромосом,
а у L. sudetica— 24 пары мелких хромосом; следовательно, по
числу хромосом этот вид формально можно считать октопло-
идом. При мейозе у межвидовых гибридов каждая из шести
больших хромосом, принадлежащих диплоидному родительскому
виду, конъюгирует с несколькими мелкими хромосомами, при-
надлежащими L sudetica (2п = 24) [619—621].
Сурсгасеаеи Juncaceae
Сем. Сурегасоае характеризуется несколькими необычными и
хорошо выраженными цитологическими и эмбриологическими
особенностями. На эти особенности указывали Хейльборп [360,
361] и многие более поздние исследователи.
360
Часть седьмая. Анеуплоидия
Во-первых, у некоторых родов Сурегасеае имеются полицент-
рические хромосомы. Как уже было сказано, у Eleocharis и Ca-
rex центромеры диффузные. В отличие от этого у Fimbristylis
и у некоторых видов Cyperus центромеры локализованные.
У Bulbostylis, по-видимому, встречаются центромеры как одно-
го, так и другого типа [534].
Во-вторых, у Сурегасеае в качестве коррелированного состоя-
ния встречается также агматополиплоидия. Довольно обширные
и непрерывные анеуплоидные ряды (sensu lato) наблюдаются у
Carex. Некоторая анеуплоидия обнаружена у Eleocharis. Одна-
ко виды Fimbristylis образуют четный или почти четный поли-
плоидный ряд.
В-третьих, у различных представителей Сурегасеае два мейо-
тических деления происходят в порядке, противоположном тому,
в котором они происходят у других растений. Конъюгировавшие
хромосомы расходятся не при первом, а при втором делении
мейоза. В отличие от обычной последовательности первое деле-
ние эквационное, а второе — редукционное [156, 867].
Девис [156] отмечает, что у покрытосеменных и у насекомых
постредукционный мейоз тесно коррелирует с диффузным со-
стоянием центромер Причины такой корреляции неясны.
В-четвертых, для Сурегасеае характерен особый способ об-
разования пыльцы. В каждой тетраде микроспор три дегенери-
руют и только одна развивается в функциональное пыльцевое
зерно (156, 361, 867].
Интересно, что большинство этих необычных цитологических
и эмбриологических признаков проявляются вновь в сем. Jun-
сасеае.
Диффузные центромеры обнаружены у Luzula, как уже го-
ворилось ранее в этой главе, но они, по-видимому, не встреча-
ются у Juncus. В соответствии с этим у Luzula наблюдается зна-
чительная анеуплоидия (sensu lato), включая некоторую агма-
топлоидию, тогда как у Juncus анеуплоидия почти отсутствует.
Согласно Норденшельду [622], у Juncaceae происходит постре-
дукционный мейоз; у них образуются все четыре микроспоры, но
они остаются связанными в тетраде, что напоминает картину,
наблюдаемую у Сурегасеае [544, 914].
Перечисленные выше черты сходства по цитологическим и
цитогенетическим признакам между Juncaceae и Сурегасеае
вместе со сходством по различным морфологическим и анато-
мическим признакам указывают на значительно более тесное
родство между этими двумя семействами, чем допускалось в ста-
рых системах классификации, в которых Juncaceae считали род-
ственными лилиям, а Сурегасеае — злакам (130, 330, 544, 914].
Часть восьмая
Специализированные
генетические системы
29 Перманентная
гетерозиготность
по транслокациям
Перманентная гетерозиготность по транслокациям представляет
собой своеобразную и высокоспециализированную генетическую
систему, в основе которой лежат последовательные транслока-
ции и которая выражается в образовании больших хромосом-
ных колец. Во многих группах растений довольно обычны реци-
прокные транслокации. В природных популяциях таких групп
небольшие кольца (состоящие из 4 или 6 хромосом) встречают-
ся как полиморфные варианты наряду с соответствующими
структурными гомозиготами, образующими биваленты В неко-
торых случаях растения, для которых характерны небольшие
кольца, достигают в популяциях большой частоты вследствие
какого-либо селективного преимущества гетерозигот. В несколь-
ких случаях благодаря преимуществу гетерозигот в популяциях
сохраняется высокая частота растений, у которых образуются
большие кольца (обзор см. [294]). Эти случаи близки к состоя-
нию перманентной гетерозиготности по транслокациям, но не
соответствуют ему полностью
Перманентные гетерозиготы по транслокациям гетерозиготны
по последовательным транслокациям, в которых участвуют по
меньшей мере несколько, а обычно большая часть или все хро-
мосомы данного набора В соответствии с этим у них при мейозе
образуются большие или полные хромосомные кольца. Геноти-
пически это — диплоиды, состоящие из двух геномов особого
типа, известных под названием комплексов Реннера.
Перманентные гетерозиготы по транслокациям при половом
размножении воспроизводят в чистоте свою структурную гете-
розиготность с выщеплепием либо небольшого числа гомозигот-
ных индивидуумов, либо вообще без выщепления. Обычно это
достигается путем передачи одного комплекса Реннера через
яйцеклетки, а другого — через пыльцу Односторонняя передача
осуществляется при помощи системы сбалансированных дета-
лей. При оплодотворении (как правило, при самооплодотворе-
нии) два комплекса Реннера воссоединяются, и, таким образом,
362 Часть восьмая. Специализированные'генетические системы
данное гетерозиготное состояние передается от одного полового
поколения следующему.
Итак, парманентная гетерозиготность по транслокациям —
это генетическая система, для которой характерно* ' сочетание
следующих признаков: последовательные транслокации, хромо-
сомные кольца от средних до больших, системы сбалансирован-
ных деталей, половое размножение, обычно путем самооплодо-
творения, и воспроизведение в чистоте определенного состояния
структурной гетерозиготности. , п ,
Перманентная гетерозигота по транслокациям, Обладающая
ценной в адаптивном отношении генетической конституцией, мо-
жет создать популяцию идентичных индивидуумов и распростра-
ниться по территории с благоприятными условиями. Она может
превратиться в гетерогамный микровид.
В свою очередь гетерогамный микровид в результате даль-
нейшего развития может стать составной частью более крупной
совокупности — гетерогамного комплекса. Гетерогамный комп-
лекс представляет собой гибридный комплекс, состоящий из ге-
терогамных микровидов и их предковых видов или полувидов,
у которых мейоз протекает нормально с образованием бивален-
тов.
Генетическая система у Oenothera
Многочисленные гетерогамные микровиды, принадлежащие к
группе Oenothera biennis в Северной Америке и Европе, — тра-
вянистые дип л оиды (2п=14), размножающиеся преимуществен-
но при помощи самоопыления. Они являются перманентными
гетерозиготами по транслокациям, причем у многих микровидов
образуются полные кольца из всех 14 хромосом, а у некото-
рых—более мелкие кольца. Растение Oenothera, образующее
кольца, после Самооплодотворения дает исключительно кольце-
образующих потомков; классы структурных гомозигот, которых
Следовало бы ожидать, среди этих потомков отсутствуют. Гете-
розйготы по транслокациям воспроизводят свою гибридную кон-
ституцию в чистоте при помощи полового размножения.
Цитогенетика Oenothera разрабатывалась рядом исследовате-
лей. Классическими считаются работы де Фриза, открывшего
аномальное поведение этих растений при скрещиваниях, и ра-
боты Реннера, давшего объяснение такому поведению. Затем на
протяжении более 40 лет анализом этой проблемы усиленно
занимались Клеланд и его ученики. Хорошие обзоры ранних
Исследований составили Клеланд [109] и Дарлингтон [139].
Большое количество фактических данных собрано в моногра-
фии Клеланда с соавторами, опубликованной в 1950 г. Обзор
со ссылками на многочисленные литературные источники'дал
Гл 29 Перманентная гетерозиготность по транслокациям
363
Клеланд [112]. Краткий обзор современного состояния проблемы
принадлежит Гранту [294].
В целях удобства мы опишем здесь генетическую систему
перманентных гетерозигот по транслокациям, используя терми-
ны, относящиеся к классическому объекту, па котором было
изучено это явление, тек Oenothera latnarckiana (рис 29 1).
Этот вид широко распространен в Европе и Северной Америке
[604, 606], при конъюгации в мейозе у него образуются кольцо
из 12 хромосом и 1 бивалент (рис 29 2). Эгот вид, 1аким обра-
зом, гетерозиготен по последовательным транслокациям в 12 из
имеющихся у него 14 хромосом.
Рис 29 1 Oenothera lamarckiana ( = Ое erijfhrosepala. Ое glazioviana) [604].
364 Часть восьмая, Специализированные генетические системы
Взаиморасположение терминальных участков хромосом
Oenothera lamarckiana можно описать, сопоставляя его с их рас-
положением у близкого структурно-гомозиготного вида Ое. hoo-
ker I, произвольно принятого за стандартное. В табл. 29.1 указа-
ны последовательные номера, присвоенные 14 плечам 7 хромо-
сом стандартной расы Ое. hookeri. У большинства индивидуумов
Ое. hookeri два гаплоидных набора хромосом структурно гомо-
логичны, поэтому при мейозе они образуют нормальные бива-
ленты. Гаплоидные наборы Ое. lamarckiana отличаются от ана-
логичных наборов стандартной, или дефризовской, расы Ое. hoo-
keri по нескольким транслокациям (табл. 29.1). Кроме того, два
набора у Ое. lamarckiana отличаются один от другого по после-
довательным транслокациям во всех хромосомах, за исключе-
нием хромосом с терминальными участками 1—2 (табл. 29.1).
Таким образом, наблюдаемую кольцеобразную фигуру из 12
хромосом и 1 бивалент можно объяснить конъюгацией гомоло-
гичных плеч при мейозе (рис. 29.2, Б).
В метафазе I хромосомы, образующие кольцо, выстраиваются
на веретене зигзагообразно, чтобы затем разойтись к разным
полюсам (рис. 29.2, Б). В анафазе I соседние хромосомы кольца
отходят к противоположным полюсам; иными словами, хромо-
сомы, лежащие в кольце через одну, отходят к одному и тому
же полюсу. Таким образом, к концу мейоза два дифференциро-
ванных генома вновь воссоединяются в интактном виде [109,
112].
Геномы особого типа, обнаруженные у Oenothera, известны
под общим названием комплексов Реннера, и каждый из них
имеет собственное название. Два комплекса Реннера, имею-
щиеся у Ое. lamarckiana, носят названия velans и gaadens
(табл. 29.1). Следовательно, Ое. lamarckiana имеет гетерозигот-
ную геномную конституцию velans/gaudens. Описанный выше
тип расхождения в мейозе кольцеобразующих хромосом приво-
Таблица 29.1. Сравнение стандартного расположения терминаль-
ных участков хромосом в комплексах Реннера с расположением
этих участков у Oenothera lamarckiana [111, 114]
Гл 29 Перманентная гетерозиготность по транслокациям
365
Рис 29 2 Конъюгация и расхождение хромосом v Oenothera lamarcktana
(2n=14) [200]
Этот вид представляет собой гетерозиготу velansi gaudens 4 Дпакенез, вид-
но кольцо из 12 хромосом и 1 бивалент Б Метафаза 1, кольцо расположено
па веретене таким образом, чтобы п анафазе хромосомы могли расходиться
к рашым полюсам через одну (чередующееся расхождение)
дит к тому, что в одно дочернее ядро переходит целиком комп-
лекс velans, а в друюе — комплекс gaudens (рис. 29 2, Б).
Иными словами, гетерозигога velans!gaudens обычно образует
лишь два класса гамет velans и gaudens, если иметь в виду
только последовательность участков в хромосомах
Следовало бы ожидать, чго при самооплодотворении пли
при скрещивании сибсов Oenothera lamarcktana даст потомков
трех типов- velansivelans, v elans j gaudens и gaudens!gaudens в
мендслевском соотношении 1:2:1. Вместо этого, как уже гово-
рилось, образуются только гетерозиготы. Отсутствие ожидав-
шихся классов структурных гомозигот обусловлено действием
системы сбалансированных деталей [109, 112, 673].
Такая генная система спагается из комплементарных леталь-
ных факторов, сбалансированных в гетерозш отпои состоянии
Двойная гегерозигота жизнеспособна, гомозиготные ре-
комбинации и (тГ-) нежизнеспособны Каждый набор
комплементарных летальных факторов сцеплен с одним комп-
лексом Реннера. С учетом сбалансированных деталей геном
Oenothera lamarcktana можно записать в виде 'el<*ns+l2 уи.
gaudens lt+
пы velansivelans и gaudens/gaudens гомозиготны по летальным
факторам и гибнут на стадии зиготы
366 Часть восьмая. Специализированные генетические системы
С различными комплексами Реннера сцеплены не только ле-
тальные факторы, но также гены, детерминирующие те или иные
фенотипические признаки (см. (109, 112], а также сжатый об-
зор Гранта [294]). Кроме того, гены, находящиеся в разных хро-
мосомах, принадлежащих к одному и тому же комплексу Ренне-
ра, передаются в составе одной группы сцепления. Поэтому гиб-
риды по транслокациям воспроизводят в чистоте не только свое
особое состояние структурной гетерозиготности, но также, и
притом одновременно, особую совокупность фенотипических
признаков.
Несколько компонентов генетической системы перманентной
гетерозиготности по транслокациям варьируют в зависимости
от вида в пределах группы Oenothera lamarcklana. Один из та-
ких изменчивых компонентов — величина кольца. Кольцо из
12 хромосом, имеющееся у Ое. lamarckiana, не типично для груп-
пы в целом. У одной популяции Ое. irrigua ( = Ое. hookeri hewet-
tii) в Нью-Мексико постоянно образуется кольцо из 8 хромо-
сом -|- 3 бивалента. Чаще всего, однако, у микровидов, населяю-
щих всю центральную и восточную части США, образуется коль-
цо из 14 хромосом (111, 114].
Система сбалансированных деталей также подвержена из-
менчивости. У представителей рассматриваемой группы, оби-
тающих на западе США, эти летали отсутствуют или встреча-
ются спорадически. Сбалансированные летали характерны, од-
нако, для гетерогамных микровидов, распространенных в цент-
ральных и восточных областях Северной Америки и в Европе.
Существуют системы сбалансированных деталей двух типов
(рис. 29 3). В системах первого типа, примером которых служит
Oenothera lamarcklana, действие деталей проявляется на стадии
зиготы; в системах второго типа, широко распространенных в
Северной Америке, оно сказывается на гаметофитах. Соответ-
ственно растения энотеры с зиготными деталями производят
полустерильные семена, а растения с гаметофитными деталя-
ми— полустерильную пыльцу [109, 111, 112, 114]. Еще одно след-
Рис 29 3 Два типа систем сбалансированных деталей у Oenothera [113]
Слева. Система с участием зиготных деталей. Справа Система с участием
гаметофитных деталей [113].
Г л 29. Перманентная гетерозиготность по транслокациям
367
сгвие системы гаметофитных деталей состоит в том, что данный
гетерогамный микровид постоянно передает один из своих комп-
лексов Реннера через пыльцу, а другой —через яйцеклетки.
Источники изменчивости
Причины наследственной изменчивости у кольцеобразующих
энотер разнообразны. Одним из важных источников новых форм
служит происходящий время от времени ауткроссинг между
микровидами, у которых обычно наблюдается инбридинг. Дру-
гие процессы создают новые вариации внутри самой инбредной
линии Хотя Oenothera lamarckiana и родственные ей формы
после самоопыления обычно производят единообразное потомст-
во, тем не менее у них возникают с некоторой низкой частотой
новые формы, в том числе дефризовские мутации.
Прежде чем обсуждать частоту мутаций у энотеры, необхо-
димо уяснить себе, что некоторые гены сцеплены в кольцеобра-
зующей гетерозиготе с комплексом Реннера, а другие не сцеп-
лены с ним. Последние расщепляются обычным образом У та-
кой неполной гетерозиготы по транслокациям, как Oenothera
lamarckiana, расщепление по хромосомам, образующим бивален-
ты, происходит независимо от расщепления по хромосомам, об-
разующим кольцо, поэтому гены, локализованные в бивалентных
хромосомах, не входят в комплексы Реннера. Гены, находящиеся
в терминальных конъюгирующих участках кольцеобразующих
хромосом, могут более или менее свободно переходить у комп-
лексных гетерозигот из одного комплекса Реннера в другой. Та-
кие гены также не зависят от какого-то одного из этих комплек-
сов (см. [112]). Разумеется, эти свободно рекомбинирующиеся
гены не дают начала новым «мутантам» у Oenothera
Гены, которые детерминируют характерные фенотипические
признаки, наследуемые в виде единого блока через любой данный
комплекс Реннера и передаваемые своим потомкам любой дан-
ной комплексной гстсрозиютой, должны располагаться на не-
конъюгирующих дифференциальных участках хромосом с транс-
локациями [112, 139]. Эти участки хромосом являются прокси-
мальными и находятся по соседству с транслокационными раз-
рывами, участвующие в этом хромосомные механизмы рассмот-
рены Грантом [294]. Сбалансированные легальные факторы
должны быть также локализованы в этих дифференциальных
участках Таким образом, гетерозиготы по транслокациям вос-
производят в чистоте определенную гетерозиготную комбина-
цию генов в этой части генома при помощи генетической систе-
мы, описанной в пэсдыдущем разделе
Однако изредка, хотя и с равномерными интервалами, про-
исходят цитогенетические события, которые отделяют один или
368 Часть восьмая. Специализированные генетические системы
несколько генов, локализованных в дифференциальных участ-
ках, от соответствующих комплексов Реннера и дают им воз-
можность проявиться в новых гомозиготных сочетаниях. Одно
из таких событий — происходящий изредка кроссинговер и об-
мен генами между комплексами Реннера. При этом какой-либо
ген может иногда оказаться в противоположном комплексе Рен-
нера. Затем в результате самоопыления возникает несколько
потомков, которые гомозиготны по этому гену и появляются в
качестве новых стабильных мутантных типов.
Другое событие —возникновение новых реципрокных транс-
локаций в данном комплексе Реннера. Такие транслокации мо-
гут разорвать большое кольцо комплексной гетерозиготы на
меньшие кольца и биваленты. Затем после самоопыления целые
хромосомы и блоки генов могут образовать гомозиготных выще-
пенцев. Возникающие при помощи описанных выше и других
способов редкие гомозиготные выщепенцы представляют собой
диплоидных мутантов энотеры (см. [112, 114, 674]).
У хромосом, образующих кольца, при разделении в анафа-
зе I наблюдаются разного рода нарушения. Время от времени
возникает нерасхождение, в результате чего образуется некото-
рое число гамет с 8 хромосомами. Такие гаметы в свою очередь
дают несколько трисомиков с измененными фенотипами. Боль-
шинство мутантов у Oenothera lamarckiana оказались трисоми-
ками с 2п= 15 (112, 114].
Второй главный источник новых вариаций у энотеры —гиб-
ридизация между различными комплексными гетерозиготами.
Два гетерогамных микровида, каждый из которых при самооп-
лодотворении дает обычно потомков одного типа, нередко дают
в первом гибридном поколении четыре класса потомков. Так,
при скрещивании Oenothera lamarckiana (velansfgaudens) и
Ое. strigosa (deprimens/stringens) получаются все четыре воз-
можные диплоидные комбинации: velans/deprimens, velans/str in-
gens, gaudensjdeprimens, gaudens I str ingens [112, 139J.
Многие из этих новых комбинаций геномов, возникающих при
указанном выше и других гибридных скрещиваниях, содержат
больше бивалентов или мелких колец, чем любой из родитель-
ских видов [112, 139]. Некоторые пары генов, сцепленные у ро-
дительских микровидов с комплексами Реннера и сохраняющие-
ся у этих микровидов в состоянии перманентной гетерозиготно-
сти, у межмикровидовых гибридов будут распределяться неза-
висимо. В последующих поколениях такие гены могут начать
выщепляться в качестве новых гомозиготных типов. Если эти
гомозиготы окажутся жизнеспособными, то новые выщепенцы
станут постоянным добавлением к фонду изменчивости данного
гибридного комплекса.
Гл. 29. Перманентная гетерозиготность по транслокациям
369
У некоторых гибридов первого поколения, полученных от
скрещивания между двумя гетерогамными микровидами, име-
ются полные или почти полные кольца, ассоциированные с но-
вой комбинацией сбалансированных летальных факторов Та-
кие гибриды будут размножаться в чистоте. Они представляют
собой потенциальные зачатки новых гибридных микровидов,
происходящих непосредственно от скрещиваний между сущест-
вующими гибридными микровидами [112].
Гетерогамный комплекс в группе
Oenothera hookeri-biennis
Североамериканские виды подрода Oenothera рода Oenothe-
ra — высокие многолетние или однолетние травянистые расте-
ния, часто сорные, с желтыми цветками, распускающимися ве-
чером. Все они — диплоиды с 2п=14. Эта группа усиленно
изучалась с цитогенетической точки зрения Клеландом и его
школой [ПО—114] и в таксономическом плане — Мюнцем [604—
606]. Североамериканские энотеры — таксономически трудная
группа вследствие характерного для них широкого диапазона
изменчивости и отсутствия ясно выраженных границ между ви-
дами.
При ревизии этой группы Мюнц [606] выделил десять таксо-
номических видов, многие из которых политипические, с одним
или несколькими подвидами. Клеланд [112—114] различает
сходный ряд таксономических видов или «групп рас», как он их
называет Системы Мюнца и Клеланда, хотя они и не идентич-
ны, сходны и хорошо согласуются одна с другой (табл 29 2)
После того как была написана эта глава, Рсйвн, Дитрих и
Штуббе [662] опубликовали новую работу по таксономии подро-
да Oenothera. Принятая ими система существенно отличается от
системы Мюнца Все эти три системы представлены и сопостав-
лены в табл 29 2.
В большинстве генетических исследований используется но-
менклатура Клеланда Во избежание путаницы мы также будем
придерживаться этой номенклатуры в пашем обсуждении Так-
сономические эквиваленты единиц, различаемых Клеландом,
можно найти в табл 29 2
Вид Oenothera lamarcktana ролом из Европы был случайно
завезен в Северную Америку Нас здесь интересуют главным
образом остальные девять видов, возникшие в Северной Амери-
ке и составляющие группу Ое hookeri-biennis
Географическое распространение девяти местных видов по-
казано на рис. 29.4 и 29 5. Ни Клеланд, ни Мюнц не опублико-
вали хороших карт географического распространения этих ви-
24—45
370
Насть восьмая. Специализированные генетические системы
Таблица 29.2. Североамериканские таксоны Oenothera лоХрода
Oenothera, различаемые Мюнцем [606], Клеландом {114] и Рей-
вном и др. [662]
Система Мюнца Названия по Клеланду Названия по Рейвну и др.
Ое. hookeri (9 подвидов) Ое. hookeri Ое. elata {hookeri вклю- чен в elata)
Ое. longissitna (2 под- вида) Ое. jamesii Ое hookeri Ое longissima
Ое hookeri Ое jamesii'
Ое elata Ое elata Ое. elata (более широ- кая концепция)
— — Ое. wolfii (выделен из hookeri) Ое. argillicola
Ое. argillicola Ое. argillicola
Ое. grandiflora L’Heri- tier (не de Vries) Oe. parviflora Ое. grandiflora' , ’ Ое. grandiflora
subsp. parviflora Ое. parviflora II Ое oakeslana
subsp angustissima Oe. biennis Ое. parviflora I Ое. parviflora
subsp caeciarum Ое. biennis II Ое. biennis
subsp. centralis Ое. biennis I Ое biennis
subsp. austromon- tana Oe. strigosa (3 подвида) Ое biennis III Ое. austromontana
Ое. strigosa Ое. villosa
Oe. erythrosepala (аккли- матизирован в Север- ной Америке) Ое lamarckiana de Vries Ое. glazioviana
дов. Поэтому ареалы, изображенные на наших картах, лишь
приблизительные, хотя и основаны на опубликованных-этими
авторами сведениях. Как указывал Клеланд (111—114], рас-
сматриваемые таксономические виды распадаются на два ряда
по типу генетической системы. Одна подгруппа обладает нор-
мальной диплоидной генетической системой. У входящих в нее
растений крупные, свободно опыляющиеся цветки; при мейозе
у них образуются (за некоторыми исключениями) биваленты, и
они не содержат сбалансированных деталей. Другая подгруппа
состоит из перманентных гетерозигот по транслокациям. У вхо-
дящих в нее растений образуются крупные хромосомные кольца
(по большей части из 14 хромосом), они размножаются при по-
мощи самоопыления и содержат сбалансированные летали.
Биваленты образуются у следующих видов: Oenothera hoo-
keri, Ое. argillicola, Ое. grandiflora и Ое. elata. От них происхо-
дят кольцеобразующие виды: Ое. parviflora, Ое. biennis, Ое. stri-
gosa. и Ое. lamarckiana. В некоторых популяциях Ое. hookeri,
Ое. jamesii и Ое. longissitna образуются кольца небольших или
Рис 29 4. Географическое распространение в Северной Америке видов Oeno-
thera подрода Oenothera; образующих биваленты (по данным Мюнца [606]
и Клеланда [114]).
24*
Рис. 29.5. Географическое распространение в Северной Америке местных
кольцеобразующих видов Oenothera подрода Oenothera (по данным Мюнца
J606] и Клеланда [114]).
Гл 29 Перманентная гетерозиготность по транслокациям
373
средних размеров; иногда эти популяции несут также сбаланси-
рованные летали, занимая, таким образом, промежуточное по-
ложение между двумя основными подгруппами [112, 114, 604,
606].
Филогенетические корни видов, у которых образуются коль-
ца, явно следует искать среди видов, формирующих биваленты
Последние, несомненно, обладают нормальной половой генетиче-
ской системой. Они предпочитают умеренно влажный климат,
что считается в данной группе растений примитивным призна-
ком. Некоторые из видов, образующих биваленты, разобщены и
носят реликтовый характер, что указывает па их достаточно
древний возраст В противоположность этому кольцеобразующие
виды с их производной генетической системой агрессивны и час-
то являются сорняками Они заселяют распаханные поля на
Среднем Западе и проникают в северные области, недавно нахо-
дившиеся подо льдом
Очень знаменательно поэтому, что один комплекс Реннера,
известный как hookeri Johansen, обычен и широко распростра-
нен у Oenothera hookeri и встречается также у Ое grandiflora и
Ое. argillicola. В комплексе hookeri Johansen терминальные уча-
стки располагаются следующим образом 1—2 3—4 5—6 7—10
9—8 11 —12 13—14. Этот комплекс Реннера — не единственный,
обнаруженный у Ое hookeri, но он встречается чаще других
Кроме того, отдельные хромосомы из комплекса hookeri Johan-
sen часто встречаются в других комплексах По эшм причинам
комплекс hookeri Johansen считается комплексом с предковым
расположением терминальных участков в дайной группе [114,
606, 798, 799].
Три или четыре таксономических вида кольцеобразующих
Oenothera плохо различимы и очень изменчивы как фенотипи-
чески, так и генотипически. Фенотипически они варьируют от
мезофитов с широкими тонкими листьями до ксерофитов с уз-
кими толстыми листьями. Что касается геномов, то у Oenothera
parvijlora, Ое biennis и Ое strigosa известно примерно 130 комп-
лексов Реннера (это число получено в результате подсчета раз-
личных комплексов, перечисленных Клеландом [114; см прило-
жение]) Эти комплексы, соединяясь попарно, образуют бесчис-
ленное множество локальных или широко распространенных ге-
терогамных микровидов.
Вследствие этого филогенетические взаимоотношения между
производными кольцеобразующими видами и предковыми вида-
ми, образующими биваленты, весьма сложны и до сих пор не
выяснены. Некоторые филогенетические связи можно предста-
вить себе в общих чертах, однако еще остаются большие пробе-
лы. В целом можно сказать, что филогения этой группы носит
сетчатый характер, так как кольцеобразующне таксономические
374
Часть восьмая. Специализированные генетические системы
виды и микровиды представляют собой продукты гибридизации
между разными парами родительских видов.
Реконструкция филогении, созданная Клеландом [113, 114],
основана на данных по морфологии, географии, распределению
концевых участков хромосом, пластидам и S-факторам, или фак-
торам самонесовместимости, которые обусловливают появление
пыльцевых леталей в системах сбалансированных леталей. Дан-
ные, полученные в разных областях исследования, не всегда сов-
падают, и это вызывает сомнения в правильности некоторых час-
тей филогенетической схемы. Суть филогенетической гипотезы
Клеланда отражена в сжатом виде на рис. 29.6 и в нижеследую-
щих дополнительных замечаниях.
По мнению Клеланда, центр возникновения группы Ое. hoo-
keri-biennis находится в Мексике и Центральной Америке. Из
этого центра примитивные формы мигрировали в умеренные об-
ласти Северной Америки; пять последовательных волн миграции
дали начало пяти видам или полувидам, получившим соответ-
ственно названия популяций I, II, III, IV и V. Популяция I —
Непосредственный предок Ое. argillicola, популяция II —
Ое. grand if lor а, а популяции III и IV — Ое. hookeri. Популяции
III и IV — гипотетические. Популяции I—IV являются предками
кольцеобразующих Oenothera. Как показано на рис. 29.6, они.
гибридизировали в разных комбинациях, дав начало кольцеоб-
Гл. 29. Перманентная гетерозиготность по транслокациям
375
разующим типам. Кроме того, с течением времени в разных пи-
ниях накапливались транслокации.
Принятые в этой гипотезе допущения относительно центра
происхождения и волн миграции необязательны. Кроме того,
собственно Ое hookeri, согласно этим допущениям, не участво-
вала в создании производных типов, что представляется, однако,
маловероятным, если принять во внимание обширность области,
в которой опа симпатричпа с Ое. strigosa. Следовательно, было
бы, возможно, лучше отказаться от некоторых из этих допуще-
ний и просто постулировать, что в Северной Америке произошла
дивергенция на несколько предковых видов, из которых одни из-
вестны, а другие неизвестны. Теперь мы можем перейти к рас-
смотрению взаимоотношений кольцеобразующих типов, по кою-
рым имеется больше данных Верхняя часть филогенетической
сети (рис 29 6) известна лучше, чем ее основание
У комплекса Oenothera biennis, передаваемого через яйце-
клетку (или у так называемого альфа-комплекса), расположение
терминальных участков в двух известных популяциях относится
к типу hookeri Johanson; во многих других популяциях, зани-
мающих обширную территорию, наблюдается иной тип располо-
жения отличающийся от hookeri Johanson на одну транслока-
цию, наряду с другими типами, различающимися сильнее
Oenothera biennis I и II очень сходны фенотипически, их
предковые линии параллельны и происходят от одних и тех же
двух родительских видов Одним из этих видов был Ое grandi-
flora, от которого Ое. biennis 1 (ио не Ое biennis II) получила
комплекс hookeri Johansen. С другой стороны, Ое biennis I и II
имеют общих предков с Ое strigosa Но s/r/gosa-подобпыс приз-
наки локализованы в передаваемом через пыльцу (бета) комп-
лексе у Ое. biennis I и в женском (альфа) комплексе у Ое. bien-
nis II У Ое. biennis I и II альфа- и бета-комплексы ревертиро-
ваны. Ое biennis III происходит от скрещивания между Ое bien-
nis 19 и Ое biennis II d [114] (рис 29 6)
Комплексы Реньера у Oenothera parviflora, Ое strigosa и
Ое. biennis (за исключением упомянутого выше альфа-комплскса
Ое. biennis I) сильно отличаются ог предкового комплекса hooke-
ri Johansen, будучи удалены от него па три или более трансло-
кации. Два комплекса Реннера у Ое parviflora I, очень далеки
от комплексов их предполагаемых предков — Ое argillicola и
Ое. grandiflora. Ое. parviflora II также далек от своего предпо-
лагаемого предка— Ое argillicola — по своему геному
Oenothera parviflora I имеет общих предков с Ое. biennis по
своим альфа-комплексам Ое parviflora II в отличие от Ое par-
viflora I песет в своих альфа-комплексах srrt^osa-подобныс при-
знаки п, по-видимому, имеет общую родословную с Ое stiigosa
(рис. 29 6).
376 Часть восьмая. Специализированные генетические системы
Гетерогамные микровиды в группе
Oenothera biennis
У кольцеобразующих видов существует ряд разнообразных
комплексов Реннера, передаваемых как через яйцеклетку, так
и через пыльцу. Число альфа- и бета-комплексов, перечисленных
Клеландом [114, см. приложение], указано в табл. 29.3, а в
табл. 29.4 приведены некоторые примеры. Возможное число
диплоидных комбинаций этих альфа- и бета-комплексов в каж-
дом кольцеобразующем таксоне достигает нескольких десятков
или сотен.
Таблица 29.3. Число известных комплексов
Реннера у Oenothera parviflora и Ое. strigo-
sa (по данным о числе комплексов, приве-
денным Клеландом [114])
Таксон Число альфа- комплексов Число бета- КОМПД£КСОВ
Ое. parviflora I 9 2
Ое. parviflora II 6 4
Ое. biennis I 14 27
Ое. biennis II 12 7
Ое. biennis III 8 4
Ое. strigosa 27 11
Любая конкретная диплоидная комбинация двух комплексов
Реннера определяет конкретный гетерогамный микровид. Гете-
рогамные микровиды представляют собой основные популяци-
онные единицы гетерогамных комплексов, оправдывающие су-
ществование генетической системы энотеры, поскольку они явля-
ются размножающимися в чистоте гибридами, гетерозиготными
по группе последовательных транслокаций и по генам, локали-
зованным в хромосомах с транслокациями. Гетерозигота по то-
му или иному гену, если она обладает адаптивной ценностью,
может размножать свою гетерозиготную конституцию при помо-
щи генетической системы, свойственной энотере, и распростра-
ниться в качестве гетерогамного вида в локальном или регио-
нальном масштабе.
Клеланд не составил списка комплексных гетерозигот или
гетерогамных микровидов как таковых, однако их можно рекон-
струировать на основе его данных, скомбинировав альфа- и бе-
та-комплексы, зарегистрированные по отдельности в одной и
той же местности [114; см. приложение]. Каждый кольцеобра-
зующий вид насчитывает следующее число микровидов: Oenot-
Таблица 29.4. Избранные комплексы Реннера в группе Oenothe-
ra hookeri-biennis [114]
Вид Расположение терминальных участков Название комплекса или местность
Ое. hookeri 1—2 3—4 5-6 7-10 9-8 11—12 13-14 Hookeri Johansen
Ое. hookeri 1—2 3-4 5-6 7-8 9—10- 11—12 13—14 Hookeri de Vries
Ое. elata 1—4 3-2 5-6 7—10 9-14 11—12 13-8 Чапультепек, Мексика
Ое. paru iflora / 1—13 3-7 5-14 4-10 9-8 11—12 2—6 Клифтон-Форж, Виргиния
Ое. biennis I 1—2 3-4 5-14 7—10 9-8 t 11—12 13—G Средний Запад
Ое. biennis I 1—9 3—10 5-14 7-4 2-8 11—12 13—6 Ричмонд, Виргиния
Ое. slrigcsa 1—7 3-4 5-8 2—10 9—11 6—12 13-14 Колорадо-Спрингс, Колорадо
Ое. strigosa 1—7 3—4 5—9 6-8 10—2 11—12 13—14 Сатерленд, Небраска
Таблица 29.5. Хромосомная конституция пяти гетерогамных мик-
ровидов в группе Oenothera biennis (на основе данных Клеланда
[114])
Вид Местность Комплекс Расположение терминальных участков
Ое. strigosa Минтерн, Колорадо бета альфа 1—10 3—2 5-14 5-8 7—8 7—12 9—10 9-6 11—12 11—12 13-6 13-14
Ое. strigosa Кларкс, Небраска бета альфа 1—13 3—2 3—4 5—10 . 5-6 7—6 7—8 9-8 9—11 11—12 10-12 13—14 2-14
Ое. biennis I Варшава, Индиана бета альфа и 3—9 5-2 5-6 7-8 7—10 6—12 9—13 11-10 11-12 13—14 8—14
Ое. biennis II Северо-восток бета альфа 1-2 1—6 3—12 3—2 5-6 5-7 7—10 4—10 9-4 9—8 11—13 11-12 8—14 13—14
Ое. parviflora 1 Клифтон-Форж, Вирги- ния бета альфа 1-13 3-2 3-7 5—12 5-14 7—10 4—10 9—6 9—8 11—8 11—12 13—14 2—6
Гл 29 Перманентная гетерозиготность по транслокациям
379
hera parviflora — 4 микровида; Ое. biennis — 90 микровидов и
Ое strigosa — 41 микровид.
В табл. 29.5 показана в целях иллюстрации геномная консти-
туция нескольких гетерогамных микровидов Хотя протяжен-
ность ареалов этих и большинства других микровидов в данной
группе неизвестна, многие из них, вероятно, имеют локальное
распространение; некоторые же виды распространены широко.
Например, микровид Oenothera biennis II, обозначенный в
табл. 29 5 как северовосточный, обнаружен в ряде мест в
шт. Пенсильвания, Нью-Йорк, пров Онтарио, Нью-Брансуик и
Квебек; этот микровид, очевидно, представляет собой удачную
гетерозиготную комбинацию
За последние годы Левин и его сотрудники стали применять
для анализа микровидов группы Oenothera biennis методы элек-
трофореза
На одном из полей в шт. Коннектикут популяция Oenothera
biennis была исследована по 19 ферментным локусам Пять из
этих 19 локусов были гетерозиготными, т. е средний уровень
гетерозиютности был очень высок составляя 26% (5/19). Кроме
того, у всех индивидуумов в этой популяции оказался один и
тот же гетерозиготный генотип по этим ферментным локусам.
Иными словами, уровень полиморфизма у них низок Эту попу-
ляцию, а возможно, и ряд популяций на всем протяжении одной
обширной территории в шт. Коннектикут характеризует один
определенный гетерогамный микровид |[472].
Левин [472] исследовал 44 популяции Oenothera biennis I в
шт Иллинойс по 20 ферментным локусам Средняя гетерозигот-
ность составляла 4,5% локусов Уровни полиморфизма в этих
популяциях были следующими
1 генотип у 59% популяций
2 генотипа у 27% »
3—5 генотипов у 16% »
Основным типом изменчивости в данном случае является не
вн\трипоп\ляционная, а межпопуляционная изменчивость
Средняя гетерозиготность по ферментным локусам в более
обширном ряде популяций группы Oenothera biennis составляет-
9,5% у собственно Ое biennis, 14,9% у Ое parviflora и 2,8% у
Ое strigosa Эти данные сравнимы с данными по образующей
биваленты Ое argillicola, у которой уровень гетсрозиготности
составляет 8 0% [483, 487].
Утверждалось, что генная гетерозиготность в i руппе Ое. bien-
nis ниже, чем можно было бы ожидать у перманентных гетеро-
зигот [472]. Возможно, однако, что функция генетической систе-
мы Oenothera состоит не в поддержании высоких уровней геге-
розиготпости, а в том, чтобы обеспечивать репликацию опреде-
ленной гетерозиготной комбинации генов.
380
Часть восьмая. Специализированные генетические системы
Гетерогамные микровиды и комплексы
в других группах растений
Подрод Oenothera рода Oenothera не является уроженцем Евро-
пы; он был завезен сюда из Северной Америки, акклиматизи-
ровался и эволюционировал на протяжении двух или трех
веков. Различные линии энотеры, завезенные в Европу
(обычно на кораблях, в балласте), гибридизировали с образо-
ванием ряда новых местных гетерогамных микровидов. Реннер
[675] различал в Европе 18 кольцеобразующих микровидов под-
рода Oenothera.
Некоторые из них, например европейский вид Oenothera bien-
nis, по-видимому, возникли в Северной Америке, но большинство
из этих 18 европейских видов возникло de novo в результате гиб-
ридизации, происходившей в Европе. Самый известный из них —
Ое. lamarckiana (иelans/gaudens). Комлекс velans близок к
Ое. hookeri, а комплекс gaudens лежит в пределах диапазона
изменчивости Ое. biennis. Гибридизация между Ое. biennis и
Ое. hookeri происходила в Европе, и продукт этой гибридизации,
Ое. lamarckiana, распространился по всей Европе, а позднее
проник в Северную Америку. К другим хорошо известным евро-
пейским. видам относятся Ое. suaveolens (albicans/flavens),
Ое. mollis (simulans/planans) и Ое. rubricaulis (tingens/rubens)
[П4].
В Южной Америке Oenothera (подрод Oenothera) представ-
лен секцией или подсекцией Munzia, параллельной собственно
энотерам Северной Америки. Изучение Oenothera-Munzia, пред-
принятое недавно Дитрихом [164], показало, что эта группа со-
стоит из 57 таксонов, из которых 20 образуют биваленты, а
37 — кольца. Среди кольцеобразующих таксонов встречаются
растения с большими кольцами (из 14 и 12 хромосом) и расте-
ния с мелкими кольцами. Таким образом, Oenothera-Munzia
представляет собой еще один хорошо развитый гетерогамный
комплекс [164].
Кольца из .14 хромосом обнаружены у представителей дру-
гих подродов Oenothera и у других родов сем. Onagraceae, род-
ственных Oenothera. Ниже перечислены такие таксоны; все
они — обитатели Северной Америки.
Oenothera подрод Raimannia (356]
Oenothera подрод Hartmannia [322]
Oenothera подрод Lavauxia [322]
Oenothera подрод Rneiffia; Oe. perennis [807]
Gayophytum heterozygum [497, 500]
Gaura biennis и G. triangulata [663]
Calylophus serrulatus [835]
Гл 29 Перманентная гетерозиготность по транслокациям 38>1
Мелкие транслокационные кольца часто встречаются в более
обширной группе родов, родственных Oenothera, а именно в
большой трибе Onagreae, а также в маленькой трибе Наиуеае.
Кариотип у представителей этих родов характеризуется двумя
существенными особенностями — хромосомами более или менее
одинаковой длины с медианными центромерами и равными пле-
чами. Кроме того, по крайней мере у подрода Oenothera (рода
Oenothera) проксимальные участки вблизи центромеры гетеро-
хроматичпы Эти особенности облегчают возникновение и ус-
пешное функционирование реципрокных транслокаций. Затем
на основе полиморфизма по мелким кольцам могут образоваться
полные гетерозиготы по транслокациям [114, 459].
Центральноамериканский вид Rhoeo spathacea (2n=12, Com-
melinaceae), прежде известный под названием R discolor, яв-
ляется полной гетерозиготой по транслокациям с кольцом из 12
хромосом и, возможно, с зиготными деталями. У близкой к нему
формы в Британском Гондурасе, R. spathacea concolor (2n==12),
происходит нормальное образование бивалентов. Геном R. s. con-
color гомологичен одному из двух геномов кольцеобразующего
R spathacea spathacea:, источник другого генома этого послед-
него вида неизвестен [20, 711, 725, 897].
Некоторые из указанных выше гетерогамных групп имеют
простую (Rhoeo spathacea) или относительно простую (Gayop-
hytum heterozygum) таксономическую структуру, тогда как у
других она сложная (Oenothera-Munzia). Таксономическая
сложность той или иной гетерогамной группы, вероятно, частич-
но определяется числом способных гибридизировать предковых
видов, а частично — диапазоном доступных для заселения эколо-
гических местообитаний. Такая группа, как североамерикан-
ские виды подрода Oenothera рода Oenothera имеющая пример-
но четыре предковых вида и занимающая обширную и разнооб-
разную территорию, в процессе последовательных циклов гибри-
дизации может дать начало многочисленным и разнообразным:
микровидам.
30 Перманентная
нечетная полиплоидия
Термин перманентная нечетная полиплоидия (permanent odd
polyploidy) используется здесь для обозначения генетической
системы особого типа, а не в буквальном значении входящих в
него слов. Пентаплоидное агамоспермное растение Crepis или
-триплоидное живородящее растение Allium — это в буквальном
смысле «перманентные нечетные полиплоиды», однако они выхо-
дят за рамки этой концепции в принятом нами определении.
Под перманентной нечетной полиплоидией мы имеем в виду яв-
ление, когда растение воспроизводит в чистоте некое состояние
нечетной эуплоидности при помощи полового размножения. Это
осуществляется путем передачи одного генома через пыльцу и
двух или нескольких геномов через яйцеклетки.
Такая генетическая система обнаружена в группе Rosa cani-
na (Rosaceae) и Leucopogon juniperinus (Epacridaceae); к ней
приближается генетическая система группы Cardamine атага
(Cruciferae).
Таксономическая изменчивость в одной известной группе
перманентных нечетных полиплоидов принимает форму ряда ге-
терогамных микровидов, связанных в один гетерогамный комп-
лекс.
Генетическая система Rosa canina
Большая часть микровидов группы Rosa canina в Европе — пен-
таплоиды (х=7, 2п=35); в этой группе имеется также несколько
тетраплоидных и гексаплоидных форм. Мы рассмотрим репро-
дуктивный цикл типичной пентаплоидной формы.
При мейозе у пентаплоидной розы собачьей 14 из ее 35 хро-
мосом конъюгируют с образованием 7 бивалентов, а остальные
(21 хромосома) представляют собой униваленты. То же самое
наблюдается при конъюгации хромосом в материнских клетках
пыльцы и материнских клетках зародышевого мешка (711+211).
Но распределение хромосом в этих двух клеточных линиях про-
исходит совершенно по-разному (139, 819]. Схематически это
показано на рис. 30.1.
В метафазе I в материнских клетках зародышевого мешка
хромосомы 711+211 распределяются по двум группам: 7 бива-
лентов выстраиваются по экваториальной пластинке, а 21 не-
конъюгировавшая хромосома собирается на микропилярном
конце. Хромосомы, образующие биваленты, разделяются и во
время анафазы I расходятся к противоположным полюсам вере-
тена. В телофазе 7 конъюгировавших хромосом соединяются с
Гл 30 Перманентная нечетная полиплоидия
383=
Рис 30 1 Процесс мейоза и слияние гамет комплементарных типов в группе
Rosa cantna'[207].
Конъю! ирующис хромосомы черные, а унивалентныс — белые Показаны
только 2 бивалентные и 6 унпвалентных хромосом.
21 неконъюгировавшей хромосомой. Таким образом, к концу
первого мейотического деления 28 хромосом обычно бывают рас-
положены на микропилярном конце клетки, а 7 — на другом ее
конце. Второе мейотическое деление протекает нормально и
приводит к образованию линейной тетрады, состоящей из двух
больших и дв)х маленьких ядер. Одно из крупных 28-хромосом-
ных ядер, находящихся па микропилярном конце, превращается
в функциональный зародышевый мешок (рис. 30 1) [139, 390,
819].
В материнских клетках пыльцы униваленты и биваленты
располагаются в экваториальной плоскости. Во время первого
деления хромосомы, образующие биваленты, разделяются и пер-
выми направляются к полюсам, а затем делятся и следуют за
ними к полюсам унивалентные хромосомы. Во время второго де-
ления конъюгирующие хромосомы делятся и расходятся к новым
полюсам Некоиъюгирующие хромосомы при втором делении
также разделяются, хотя и с опозданием, и дочерние хромосомы
начинают перемещаться к полюсам^ но большая их часть до по-
люсов не доходит [139, 819].
384
Часть восьмая. Специализированные генетические системы
В результате мейоза в мужской линии образуются дочерние
ядра, содержащие основной комплемент из 7 конъюгирующих
хромосом и варьирующего числа неконъюгирующих хромосом.
В одной выборке из 63 микроспор, в которых подсчитывали чис-
ло хромосом, 9 клеток содержали по 7 хромосом, а остальные
54 — от 8 до 22 хромосом. Микроспоры различались по величине
и форме и в своем большинстве были абортивными. Функцио-
нальными были пыльцевые зерна с набором из 7 хромосом из
конъюгирующего генома (рис. 30.1) [139, 207, 819].
У тетраплоидов при конъюгации образуется 711+141, а у
гексаплоидов —711+281. Эти типы, таким образом, сходны с
пентаплоидами, содержащими 711+211, в том отношении, что
они содержат как конъюгирующие, так и неконъюгирующие ге-
номы Кроме того, подобно пентаплоидам, они передают унива-
лентные хромосомы через яйцеклетки [819].
Большая часть микровидов группы Rosa canina несет, как
отмечалось выше, 2п=5х=35 хромосом (149, 151, 819J. Сначала
полагали, что у розы собачьей это постоянное пентаплоидное
состояние поддерживается при помощи агамоспермии [390, 819],
однако позднее было показано, что они размножаются половым
путем [207, 316]. Ряд авторов утверждает, что эти растения в
значительной мере самоопыляются [139, 146, 502, 780], но поми-
мо этого у них в некоторой степени происходят также ауткрос-
синг и гибридизация. Желательно получить дополнительные ко-
личественные данные о частоте инбридинга и ауткроссинга у
этих растений.
При самооплодотворении у пентаплоидного растения Rosa
canina происходит слияние женской гаметы с 28 хромосомами
и мужской гаметы с 7 хромосомами. В результате этого события
восстанавливается пентаплоидное состояние (2п=35), а кроме
того, образуется потомство, сходное с родительским растением
по своему геному (711+211). Перекрестное опыление между
сестринскими пентаплоидными растениями приводит к таким же
результатам. Следовательно, роза собачья воспроизводит в чис-
тоте состояние нечетной полиплоидии при половом размножении.
Гетерогамный комплекс в группе Rosa canina
Группа Rosa canina составляет отдельную секцию (секция Cani-
пае) рода Rosa. К этой секции относятся дикорастущие колючие
кустарники с цветками от мелких до средних или, в редких слу-
чаях, крупных размеров (рис. 30.2); встречаются они в мелко-
лесье и на полях по всей Европе и в соседних с ней частях се-
верной Африки и на Ближнем Востоке.
К секции Caninae относятся многочисленные микровиды, из
которых одни распространены широко, а другие — локально,
Гл. 30 Перманентная нечетная полиплоидия
385
причем между ними происходит довольно сложная интерграда-
ция. Вследствие этого разные авторы представляют себе таксо-
номию этой группы по-разному, как это обычно бывает с гибрид-
ными комплексами Тэкхолм [819] различает 34 таксономичес-
ких вида, объединяя их в 5 групп видов. Хеги [359] низводит
некоторые из этих таксономических видов до ранга подвидов, но
различает в каждой видовой или подвидовой единице многочис-
25—45
386 Часть восьмая. Специализированные генетические системы
ленные разновидности и формы. Так, Rosa canina sensu stricto
и R. dumetorum он считает подвидами R. canina sensu lato. Ho
R. canina sensu stricto имеет при этом 7 разновидностей и
12 форм, a R. dumetorum —5 разновидностей и 10 форм [359].
В связи с этим принято считать, что рассматриваемая группа
таксономически разнообразна и сложна.
На основе морфологических признаков секцию Caninae мож-
но считать тесно спаянной единицей, хорошо отделенной от дру-
гих секций рода Rosa (рис. 30 2), поскольку, с одной стороны,.
Caninae морфологически достаточно ясно отличается от других
секций [819], а с другой — входящие в эту секцию микровиды
интерградируют в многочисленных комбинациях. Так, R. canina
sensu stricto интерградирует с R. dumetorum, R. stylosa, R. рои-
zini и R. chavini, R. dumetorum в свою очередь интерградирует с
R. stylosa, а кроме того, — с R. coriifolia и R. tomentella [359].
Основное число хромосом в группе Rosa canina х=7, как и
в других секциях этого рода. Однако в секции Caninae не изве-
стно ни одного диплоида. Большинство входящих в нее микро-
видов— пентаплоиды; есть также несколько тетраплоидов или
гексаплоидов [819].
Для рассмотрения частоты распределения разных уровней
плоидности в группе Rosa canina мы примем за основу состав-
ленный Тэкхолмом [819] список таксономических видов и снабдим
его данными о числе хромосом в соответствии со сведениями, опуб-
ликованными самим Тэкхолмом [819] и последующими автора-
ми (библиографию см. [149]). Из 34 таксономических видов 15 —
пентаплоиды, 3 — тетраплоиды, 3 — гексаплоиды, 4 содержат как
пентаплоидные, так и гексаплоидные формы, а по 9 видам под-
счетов не производилось.
К пеитаплоидам (2п=35) относятся: Rosa canina sensu stri-
cto, R. dumetorum, R. micrantha, R. tomentosa и R. stylosa (час-
тично). К тетраплоидам (2л=28)—R. rubrifolia, R. pomifera и
R. mollis. К гексаплоидам (2n=42)—R. junzillii, R. pouzini,
R. chavini и R. stylosa (частично).
Представления Тэкхолма о геномной конституции пентапло-
идного микровида, основанные на наблюдениях за конъюгацией
хромосом, можно выразить формулой AABCD. Исходя из ха-
рактера поведения при мейозе некоторых искусственных гибри-
дов между микровидами, он полагал, что у разных микровидов
из группы Rosa canina один геном общий, а по другому геному
они различаются. Так, три пентаплоидных микровида могут
иметь формулы AABCD, AACDE и AADEF соответственно.
Тэкхолм [819] считал пентаплоидные микровиды непосредствен-
ными продуктами гибридизации между одним предковым дип-
лоидным видом и различными октоплоидными видами. Поль-
Гл. 30. Перманентная нечетная полиплоидия
387
зуясь геномными формулами, микровид, образующийся в ре-
зультате скрещивания между А А и AABBCCDD, можно обозна-
чить AABCD; скрещивание AAxAACCDDEE даст микровид
AACDE и т. д.
Последующие цитогенетические исследования заставили мо-
дифицировать эти заключения в некоторых важных отношениях.
Густафсон и Хёканссон [319] скрещивали три микровида Rosa
canina (5х и 6х) с диплоидным видом R. rugosa (2л = 7П), при-
надлежащим к другой секции. У этих искусственных гибридов
при мейозе образовывалось по 14 бивалентов. Это свидетельст-
вует о том, что полиплоидные R. canina гомологичны не по двум,
а по трем геномам. Они автотриплоидны по одному из своих 5
или 6 хромосомных наборов [316, 319]. Фагерлинд {208] пошел
дальше, указывая на некоторую гомологию между всеми гено-
мами полиплоидных R. canina.
На основе цитогенетических данных, полученных Густафсо-
ном и Хёканссоном [319], Густафсон [316] предложил следующие
формулы геномов: для Rosa canina sensu stricto — AsAsAtCD,
а для родственного микровида R. rubiginosa — BBBCF. Геном C
обнаружен у R. rugosa, относящейся к другой секции, и, возмож-
но, распространен у данного рода более широко (139]. Основы-
ваясь на этих и ряде дополнительных данных, Фагерлинд [208]
низвел бы некоторые из обозначенных буквами геномов до суб-
геномов (см. также [62, 915]). Во всяком случае, микровиды
R. canina, по-видимому, представляют собой автоаллополипло-
иды, геномы которых в разной степени различаются по своей
структуре
Геномная конституция полиплоидных Rosa canina имеет важ-
ное значение для проблемы происхождения новых микровидов.
Классическая точка зрения Тэкхолма, считавшего, что различ-
ные микровиды R. canina — строгие аллополиплоиды и поэтому
размножаются в чистоте, привела к заключению, что эти мик-
ровиды— прямые продукты гибридизации между разными ком-
бинациями родительских видов [819]. Согласно этой точке зре-
ния, ряд потенциально возможных микровидов ограничивается
числом гибридизирующих предковых видов.
Если же исходить из того, что микровиды Rosa canina по
своей конституции являются автоаллополиплоидами, то тогда
один высокогетерозиготный .микровид может непосредственно
дать начало ряду новых микровидов в результате расщепления
по генным различиям между геномами. Так, у пентаплоида с
конституцией AsAsAtCD хромосомы At, которые обычно обра-
зуют униваленты, могут иногда включаться в конъюгирующие
наборы, и тогда они будут в дальнейшем передавать свои гены
следующему поколению как через мужские, так и через женские
гаметы [319].
25*
388 Часть восьмая Специализированные генетические системы
Короче говоря, один автоаллополиплоидный микровид, кото-
рый обычно размножается в чистоте, может иногда выщепить
один или несколько новых микровидов, размножающихся в чис-
тоте. Этот процесс позволяет объяснить возникновение очень
большого числа микровидов Rosa canina, реально существую-
щих в природе [319].
Правдоподобную картину филогенеза комплекса Rosa canina
воссоздал Тэкхолм [819]. Нам неизвестно ни одного диплоидного
вида и ни одного полиплоида с нормально протекающим мейо-
зом, который обладал бы морфологическими признаками, ха-
рактерными для секции Caninae. Все современные формы обла-
дают производной генетической системой. Следовательно, эта
группа, вероятно, довольно древняя. Кроме того, современные
производные микровиды занимают в Европе территории, на
которые оказал сильное воздействие ледниковый период. Дип-
лоидные предки группы Rosa canina, по-видимому, вымерли в
плейстоцене, а существующие в наши дни гибридные комплексы
возникли в плейстоценовое и постплейстоценовое время [819].
Leucopogon juniperinus
Перманентная нечетная полиплоидия была обнаружена также у
Leucopogon juniperinus (Epacridaceae) в Австралии [745]. Этот
вид — триплоид, размножающийся половым путем, с числом
хромосом 2п=3х=12. В процессе мейоза два гаплоидных набора
образуют биваленты, тогда как третий гаплоидный набор не
проявляет никакой тенденции к конъюгации Это растение ведет
Рис. 30.3 Формирование гамет комплементарных типов в мужской и жен-
ской линиях у Leucopogon juniperinus (4II+4I) >[747].
Зародышевый мешок содержит четыре унивалента, но в пыльцевых зернах
их нет.
Гл 30. Перманентная нечетная полиплоидия
389
себя, таким образом, как аллотриплоид с геномной конститу-
цией ААВ [745, 746].
В первой метафазе материнские клетки как зародышевого
мешка, так и пыльцы конъюгируют с образованием 4 1.
В анафазе Т в материнских клетках обоих типов все униваленты
отходят к одному полюсу В материнских клетках зародышево-
го мешка этот полюс — функциональный, а в материнских клет-
ках пыльцы — нефункциональный. В результате ядра зароды-
шевого мешка и яйцеклетки несут по 8 хромосом, принадлежа-
щих к геномам А и В, тогда как функциональные пыльцевые
зерна несут по четыре хромосомы генома А (рис. 30.3). При
оплодотворении восстанавливаются соматическое число хромо-
сом 2п=12 и геномная конституция АЛВ [745, 746].
Одно из условий, облегчающих возникновение сбалансиро-
ванной системы элиминации комплементарных гамет у Leucopo-
gon juniperinus, — своеобразный способ развития пыльцы у этих
растений У Leucopogon, как и у других представителей трибы
Styphelicae сем Epacridaceae, три микроспоры в каждой тетра-
де абортивные и только одна развивается в зрелое пыльцевое
зерно Это функционирующее пыльцевое зерно представляет со-
бой одно из двух 4-хромосомных продуктов мейоза у L. juniperi-
nus (рис. 30 3). У этого вида возможно существование системы
элиминации гамет с .мужской стороны без всякого сокращения
общего выхода фертильной пыльцы [746, 747].
Leucopogon juniperinus морфологически и цитологически еди-
нообразен па всем протяжении обширной территории, занимае-
мой им в Австралии. Это единый аллотриплоидный вид [746]
В этой группе растений не произошло образования гетерогамно-
го комплекса
Cardamine insueta
Между Cardamine amara и С rivularis (Cruciferae) в Швейца-
рии изредка происходит естественная гибридизация. Оба роди-
тельских вида — диплоиды (2/г=16), а их естественные гибри-
ды— триплоиды (2д=3х=24). Добавочный хромосомный набор
они получают от С rivularis и имеют геномную конституцию
RRA [852].
Гибриды высокостерильпы и дают всего 2—3% фертильной
пыльцы Тем не менее они хорошо воспроизводятся и распрост-
ранились локально по площади 16 га, где они преобладают над
родительскими видами. Размножаются они как вегетативным,
так и половым путем Гибридные растения, получившие назва-
ние С. insueta, размножаются половым путем, представляющим
собой некую модификацию схемы размножения Rosa canina
[852, 853].
&0 Часть восьмая. Специализированные генетические системы
Триплоидные растения Cardamine insueta испытывались по
потомству в трех сериях при разных условиях. Потомство три-
плоида, выращивавшегося в открытом поле, состояло из 21 рас-
тения; все они были триплоидами (Зл=24). Потомство трипло-
йда в контролируемых сестринских скрещиваниях состояло из
5 растений; 4 из них были триплоидами, а один — тетраплоидом
с геномом RRRA. Потомство от возвратных скрещиваний трипло-
ида с С. rivutaris состояло из форм 2х, Зх, 4х и 5х. Таким обра-
зом, насколько это известно, в природных условиях триплоидные
растения производят исключительно триплоидные растения, а
при искусственных близкородственных скрещиваниях дают пре-
имущественно, но не исключительно триплоидное потомство
[8521.
Обычная конфигурация, получающаяся при конъюгации в
метафазе I у природного триплоида, —8 П+8 I; предположи-
тельный геном этих форм RR 11+А I. Расхождение хромосом в
анафазе I варьирует в различных клетках. Часто встречается
соотношение 8:16, обнаруженное у 29% проанализированных
клеток. Таким образом, в этом случае выявляется тенденция к
поляризованному расхождению хромосом. У 7% клеток были
обнаружены реституционные ядра с 24 хромосомами. Нередко
наблюдается также неравномерное распределение хромосом,
что приводит к образованию анеуплоидных продуктов мейоза
[8521.
Cardamine insueta производит функциональные 8- и 16-хро-
мосомные яйцеклетки и 8- и 16-хромосомные пыльцевые зерна.
Он отличается в этом отношении от Rosa canina, у которой жен-
ские гаметы полиплоидные, а мужские гаплоидные.
Следовало ожидать, что у Cardamine insueta при соединении
8- и 16-хромосомных пыльцевых зерен с 8- и 16-хромосоиными
яйцеклетками образуется потомство, принадлежащее к полипло-
идным классам 2х, Зх и 4х. Кроме того, следовало бы ожидать,
что нередуцированные гаметы из 24-хромосомных реституцион-
ных ядер дадут начало некоторому числу потомков 5х (а также
индивидуумам других уровней плоидности). И в самом деле,
такие полиплоиды (2х—5х) были обнаружены в потомстве
С. insueta при искусственных возвратных скрещиваниях, о чем
уже говорилось выше. Однако потомки С. insueta, произрастав-
шие в открытом поле, не образовывали полный ряд таких поли-
плоидных форм; здесь обнаружены только триплоиды, о чем
уже говорилось ранее [852].
> Можно было также ожидать, что среди этих потомков будет
некоторое число анеуплоидных форм, однако их не было ни в
природных, ни в экспериментальных популяциях. У Cardamine
insueta проявляется определенная тенденция к воспроизведению
в чистоте своей триплоидной конституции в природных уело-
Гл. 30. Перманентная нечетная полиплоидия 391
виях. Механизмы, обусловливающие такой характер размноже-
ния, еще не вполне понятны [852].
Cardamine amara и С. rivularis дали также плодовитый авто-
аллогексаплоид (2n = 48, RRRRAA), известный под названием-
c. schulzii [852]. Здесь мы имеем дело со случаем, когда одна
пара диплоидных родительских видов дает начало формам, на-
ходящимся на двух различных уровнях плоидности и размно-
жающимся в чистоте. Урбанска продолжает свои интересные
исследования на С. insueta и С. schulzii, и, по-видимому, в буду-
щем нам станет более понятной биология размножения этих
видов.
Эволюционный потенциал гетерогамных
комплексов
Дарлингтон [139] обратил внимание на некоторые интересные
сходные черты в цитогенетике размножающихся в чистоте не-
четных полиплоидов группы Rosa canina и размножающихся в
чистоте гетерозигот по транслокациям группы Oenothera biennis.
У каждой группы есть несколько регулярно конъюгирующих
хромосом или участков хромосом: это 7 бивалентов у Rosa ca-
nina и терминальные конъюгирующие участки у Oenothera bien-
nis. У каждой группы есть также несколько неконъюгирующих
хромосом или участков — униваленты у Rosa и дифференциаль-
ные участки у Oenothera [139] (см. также [780]) В хромосомном
наборе каждой группы наблюдается разделение функций между
конъюгирующими и неконъюгирующими участками Некоторые
хромосомы или их участки, регулярно совершающие конъюга-
цию и подвергающиеся кроссипговеру, обеспечивают изменчи-
вость. Другие хромосомы или участки хромосом, которые не
конъюгируют, несут гены, детерминирующие постоянные приз-
наки разных микровидов [139].
Гибридизация между диплоидными видами, образующими
биваленты, дает в каждом случае ряд нумерических или струк-
турных гибридов. Эти гибриды передают одну часть своего хро-
мосомного набора через пыльцу, а другую — через яйцеклетки,
и гаметы двух комплементарных типов часто соединяются в
результате самооплодотворения. Таким образом, численность
гибридов увеличивается, и они развиваются в гетерогамные мик-
ровиды. Гибридные микровиды заполняют разрывы между пред-
ковыми видами и дают начало гетерогамному комплексу в его
первичной стадии. Затем микровиды гибридизируют друг с дру-
гом, давая начало новым вторичным микровидам, которые еще
больше обогащают изменчивость гетерогамного комплекса на
его поздних стадиях.
392 Часть восьмая. Специализированные генетические системы
Как у Rosa canina, так и у Oenothera biennis значительные
части генотипа «заперты» (locked up) в состоянии перманентной
гетерозиготности в неконъюгирующих участках хромосом, поэто-
му эти растения обычно размножаются в чистоте по таким ге-
терозиготным комбинациям генов. Однако происходящие в виде
исключений в этих же участках конъюгации или кроссинговеры
приводят время от времени к выщеплению новых константных
типов. Таких редких выщепенцев можно рассматривать как
запоздалые продукты гибридизации родительских видов, кото-
рые обычно размножаются в чистоте, но высокогетерозиготны.
Эти константные гибридные выщепенцы, если они окажутся
ценными в адаптивном отношении, могут превратиться в новые
микровиды и тем самым еще больше увеличить общую измен-
чивость гетерогамного комплекса.
Гетерогамный комплекс в стадии зрелости содержит десятки
или сотни микровидов, происхождение которых прямо или кос-
венно связано с гибридизацией и которые заселили территории,
сравнимые по площади с материками. Такие гетерогамные комп-
лексы, примером которых служат группы Rosa canina и Oenot-
hera biennis, несомненно, эволюционируют очень успешно.
И все же остается фактом, что у гетерогамных микровидов
значительные части генотипа «заперты». Рекомбинация у них
очень сильно ограничена тем же самым хромосомным механиз-
мом, который обеспечивает кратковременный эволюционный
успех. Можно ожидать, что в конечном итоге эти ограничения
сильно препятствуют появлению новых вариаций и загоняют
гетерогамный комплекс в эволюционный тупик [141, 780].
31 Агамоспермия
Агамоспермией называют образование семян без оплодотворе-
ния Это одна из основных форм апомиксиса, или размножения
без оплодотворения, у высших растений Само агамоспермное
размножение включает в себя очень разнообразные и сложные
явления, которые будут описаны в дальнейшем Агамоспермия
широко распространена у цветковых растений, о чем свидетель-
ствует перечень, представленный в табл. 31 1 В этой главе
дается обзор эмбриологических и цитогенетических явлений как
необходимой основы для понимания происхождения изменчиво-
сти у агамоспермных растений Опираясь на эти данные, мы
сможем затем перейти в следующей главе к рассмотрению эво-
люционного развития агамных комплексов.
Более подробное изложение этих проблем читатель найдет
в классической монографии Густафсона «Апомиксис у высших
растений» [317]. Многие дополнительные данные, полученные
позднее, содержатся в обзорах Стеббинса [780], Нюгрена
[625], Фрикселла [223], Батталья [48] и Рутисхаузера [701].
Эмбриологические пути
Различные пути эмбрионального развития, ведущие к агамо-
спермному образованию семян, показаны па рис 31.1 В этой
схеме использованы классификация и терминология Густафсона
[317] и сложные явления представлены в несколько упрощенной
форме, с тем чтобы сделать возможными широкие сопоставле-
ния.
Нормальный жизненный цикл семенного растения, являю-
щийся предковым для разного типа агамоспермных жизненных
циклов, слагается, как известно, из чередования диплоидного
спорофитного и гаплоидного гаметофитного поколений; эти по-
коления разделены такими событиями, как мейоз и оплодотво-
рение, которые соответственно сокращают и восстанавливают
соматическое число хромосом (рис. 31.1, А). При агамоспермных
жизненных циклах, которые обеспечивают сохранение более или
менее постоянного числа хромосом у ряда поколений растений,
развивающихся из семян, образование зародыша происходит
без мейоза и без оплодотворения.
Этот результат достигается двумя способами, соответствую-
щими двум основным типам агамоспермии — гаметофитному
апомиксису и адвентивной эмбрионии При гаметофнтном апо-
миксисе гаметофит как морфологическое образование сущест-
вует, но редукции хромосом не происходит (рис. 31.1,5). В от-
А НОРМАЛЬНЫЙ ЦИКЛ ПОЛОВОГО РАЗМНОЖЕНИЯ
Б, ГАМЕТОФИТНЫЙ АПОМИКСИС
В АДВЕНТИВНАЯ ЙМБРИОНИЯ
Рис. 31 1. Способы агамоспермного размножения (для сравнения приведен жизненный цикл растения с нормальным по-
ловым размножением).
Гл. 31. Агамоспермия
395
Рис 312 Апоспория у Crepis occidentalis (по [792] с изменениями).
А. Делящаяся материнская клетка зародышевого мешка (1), а под ней уве-
личивающаяся соматическая клетка (2) Б Линейная тетрада (3), а под ней
соматическая клетка, развивающаяся в апоспорический зародышевый ме-
шок (4). В Материнская клетка зародышевого мешка (1), дегенерирующая
при первом делении, а под ней одноклеточный апоспорический зародышевый
мешок (4).
Рис. 31 3 Диплоспория у Antennar.a porsildii, материнская клетка зароды-
шевого мешка из которой развивается нсредуцирсванный зародышевый ме-
шок [850].
личие от этого при адвентивной оvбрионии гаметофита нет, и
соответственно выпадает чередование поколений (рис 31.1, В).
Как показано на рис. 31.1, Б, при гаметофитном апомиксисе
возможны два альтернативных пути, ведущие от материнского
спорофита к нередупированному гаметофиту, и еще два пути,.
i396 Уасгв восьмая Специализированные генетические системы
ведущие от этого гаметофита к новому зародышу. При апоспо-
рии женский гаметофит образуется из соматических клеток ну-
целлуса или халазы семязачатка (рис. 31.2). При диплоспории
женский гаметофит образуется непосредственно из одной мате-
ринской клетки зародышевого мешка, не претерпевшей мейоза
(рис. 31.3).
В любом случае женский гаметофит содержит яйцеклетку с
нередуцированным числом хромосом и разного рода дополни-
тельные клетки. При диплоидном партеногенезе из такой яйце-
клетки без оплодотворения развивается новый зародыш. При
апогаметии какая-либо другая (не яйцеклетка) клетка женско-
го гаметофита образует зародыш (рис. 31.1,Б).
Апогамное развитие зародышей описано у ряда покрытосе-
менных. Густафсон приводит примеры развития зародышей из
синергид у Taraxacum, Hieracium, Alchemilla, Alnus и Роа и
примеры развития их из антиподов у Hieracium, Elatostema и
Allium [317]. Но, как отмечает Густафсон, в природе это явле-
ние, вероятно, не имеет большого значения для покрытосемен-
ных.
Остальные процессы, сочетаясь друг с другом, образуют две
полные линии развития, которые можно назвать диплоспория—
партеногенез и апоспория—партеногенез. Оба эти последова-
тельности встречаются в четко выраженной форме у цветковых
растений (табл. 31.1). Они встречаются также в смешанной
форме у некоторых покрытосеменных, у которых развиваются
и диплоспОровые, и апоспоровые зародышевые мешки
(табл. 31.1).
Наконец, при адвентивной эмбрионии новый зародыш разви-
вается непосредственно из нуцеллуса или интегумента семяза-
чатка на материнском растении и, таким образом, совершенно
обходит зародышевый мешок. Адвентивная эмбриония представ-
ляет собой тот тип агамоспермии, который в наибольшей степе-
ни приближается к живорождению как одной из форм вегета-
тивного размножения (или к ложному живорождению, согласно
ван дер Пэйлу [639]). В обоих случаях новый индивидуум обра-
зуется бесполым путем в соцветии материнского растения. Но
при адвентивной эмбрионии дочернее растение начинает разви-
ваться как зародыш, находящийся в семени, тогда как при
псевдоживорождении оно развивается из воздушной луковички,
возникающей внутри цветка или на его месте.
У некоторых агамоспермных покрытосеменных для апомик-
тического развития семени необходимо опыление. Такое стиму-
лирующее воздействие опыления, необходимое апомиктическому
цветковому растению в качестве предпосылки для образования
семени, известно под названием псевдогамии; при этом опыление
служит либо общим стимулом, инициирующим развитие заро-
Гл 31 Агамоспермия
397
дыша, либо, что более обычно, за опылением следует слияние
ядра спермия с ядрами эндосперма. Этот процесс оплодотворе-
ния инициирует развитие эндосперма, необходимого для нормаль-
ного роста зародыша. Сам зародыш, однако, не получает ника-
кого генетического материала от растения, поставляющего пыль-
цу, и по своему генотипу полностью сходен с материнской фор-
мой.
Агамоспермное растение может производить ссмеиа исклю-
чительно бесполым путем или же может время от времени да-
вать потомков в результате полового процесса В первом случае
имеет место облигатный, а во втором — факультативный апо-
миксис.
Одно и то же факультативно агамоспермное растение может
в разные годы давать в разных соотношениях семена, образую-
щиеся половым и бесполым путем, или даже давать те и другие
семена в урожае данного года Иными словами, половой путь
развития зародыша, схематически представленный на рис. 31 1,А,
может действовать в одной линии развития семян, тогда как в
другое время или в другом месте на том же самом растении
развитие семян может происходить альтернативным агамосперм-
ным путем (рис. 31.1, Б и В).
Hieracium aurantiacum в Карпатах (Польша) образует жиз-
неспособные семязачатки двух типов в одних и тех же соцветиях.
Некоторые семязачатки содержат редуцированные зародыше-
вые мешки, образовавшиеся в результате нормальною мейоза,
тогда как другие содержат нередуцированные зародышевые
мешки, возникшие в результате апоспории. В последних заро-
дыш и эндосперм развиваются автономно, тогда как в первых
им необходимо оплодотворение как предпосылка к дальнейше-
му развитию. Вследствие этого в одном соцветии семянки обра-
зуются как половым, так и бесполым путем. Соотношение семя-
нок этих двух типов различно у разных растений [73].
У папоротников, где агамоспермия, разумеется, отсутствует
по определению аналогичную роль играет апогаметия, пред-
ставляющая собой преобладающий способ бесполого образова-
ния зародышей у этой группы. У апогамных папоротников пос-
ледовательность событий начинается с образования в споран-
гиях материнского растения нерсдуцированпых спор. Эти споры
дают начало гаметофитам, несущим нсрсдуцироваппое число
хромосом. Затем из вегетативных клеток гаметофита без опло-
дотворения развивается новый спорофит [554]
Таксономическое распределение
В табл 31.1 представлен список агамоспермных цветковых рас-
тений Этот перечень составлен на основе работ Густафсона
<317] и Нюгрена [625], в которых можно найти ссылки на ориги-
398
Часть восьмая. Специализированные генетические системы
нальные иссйедования. Следует помнить, что многие таксономи-
ческие виды, перечисленные в табл. 31.1, содержат как половые,,
так И агамоспермные формы. Виды сгруппированы в этой таб-
лице, во-первых, по типу агамоспермии и, во-вторых, по принад-
лежности к тому или иному семейству растений.
Этот список неполон, но дает достаточно хорошее представ-
ление о распространении агамоспермии у покрытосеменных.
Можно видеть, что каждая из основных форм агамоспермии
встречается у представителей многих семейств. Так, дипдоспо-
рия — партеногенез встречается у многочисленных представите-
лей сложноцветных, а также во многих других семействах. Апо-
спория— партеногенез встречается у разных представителей ро-
зоцветных и в других семействах. Адвентивная эмбриония наб-
людается довольно часто у представителей семейств лилейных,,
рутовых и других.
Для Parthenium argentatum характерны одновременно и дип-
лоспория—партеногенез, и апоспория—партеногенез; такая
ситуация, когда обе системы агамоспермного размножения
встречаются вместе у одного и того же вида, обнаружена и у не-
которых других растений (табл. 31.1). Эти две близкие формы
гаметофитного апомиксиса наблюдаются у разных видов Роа.
и в разных породах Hieracium.
Псевдогамия широко распространена у агамоспермных по-
крытосеменных, в том числе у видов Роа, Rubus, Potentilla, Ra-
nunculus, Hypericum, Parthenium, Citrus и Allium. К группам*
у которых псевдогамия не наблюдается, относятся Hieracium,
Taraxacum, Antennaria, Crepis и Calamagrostis [317].
Если рассматривать представленный в табл. 31.1 список ви-
дов с точки зрения их географического и климатического рас-
пространения, то можно выявить некоторые тенденции. Многие
примеры адвентивной эмбрионии относятся к тропическим и
субтропическим растениям, тогда как гаметофитный апомиксис
чаще встречается у растений северных регионов с более холод-
ным климатом.
Что касается числа хромосом, то в выборке агамоспермных
цветковых растений очень велика доля полиплоидов [317]. Мно-
гие из них — высокие полиплоиды, как, например, Antennaria
alpina с 2л=84. Другие — нечетные полиплоиды, как триплоиды
Crepis и Malus. У третьих наблюдается широкий диапазон анеу-
плоидных чисел хромосом, варьирующих вблизи некоторой по-
липлоидной моды. Таким образом, полиплоидия в той или иной
форме обычно бывает связана с агамоспермией. Тем не менее
в родах Citrus, Nothoscordum, Potentilla и других имеется неко-
торое число агамоспермных диплоидов.
Густафсон [317] и Стеббинс [780] указывают, что бесполые-
диплоиды встречаются чаще в группах с адвентивной эмбрио-
Таблица 31.1. Избранные примеры агамоспермных покрытосе-
менных (по [317] и [625]; данные по Clusia [543])
I Диплоспория—партеногенез
Dicotyledoneae
Balanophoraceae
Balanophora japonica (2л=94—112); В. globosa (большое число)
Compositae
Antennaria alpina (2л=84) и 13 других видов
Chondrilla acantholepls (2л=15) и 8 других видов (2л =15, 20)
Erigeron annuus (2л=27) и 2 других вида (2л=27; 32—36)
Eupatorium glandulosum (2л=51)
Hieracium подрод Euhieracium, многочисленные виды (2л= 18,
27, 36)
Ixeris dentala (2n=21)
Rudbeckia californica (2n=76) и 5 других видов (2n=76)
Taraxacum, многочисленные виды (2л=24, 32, 40, 48)
Cruciferae
Arabis holboellii (2n=14, 21)
Plumbaginaceae
Statice olaeefolia confusa (2n=27)
Saururaceae
Houttuynia cordata (2n=94—104)
Thymeliaeaceae
Wikstroemia viridifolia (2n=27)
Urticaceae
Elatostema eurhynchum (2л =52) и 3 других вида (2л=60, 80)
Monocotyledoneae
Amaryllidaceae
Cooperia pedunculata (2 n=48)
Zephyranthes texana (2л=24)
Burmanniaceae
Burmannia coelestis (2n=64—72)
Gramineae
Calamagrostis canadensis (2n=42—68) и 6 других видов (2n=42—
П2)
Роа glauca (2n=42—70) и 4 других вида
И. Апоспория—партеногенез
Dicotyledoneae
Compositae
Crepis acuminata (2n=33, 44) и 8 других видов (2 л=22—88)
Hieracium подрод Pilosella, многочисленные виды (2л=27, 36,
42, 45)
Guttiferae
Hypericum perforatum. (2л = 32)
Polygonaceae
Atraphaxis frutescens (2n=88?)
RanuncLlaceae
Ranunculus auricomus (2л= 16, 32, 40, 48)
Rosaceae
Alchemilla arvensis (2л— 48) и 2 других вида
Malus hupehensis (2л=51)
Potentilla argentea (2n=14, 28, 35, 42, 56); P praecox (2и=42)
Rubus секция Moriferus, многочисленные виды (2л=21—49)
Sorbus chamaemespilus (2л=68); S hybrida (2л=68)
<Jrticaceae
Elatostema acuminatum (2л=50); E latifolium (2л=60)
Monocotyledoneae
Gramineae
Paspalum dilaiatum (2n=40)
Poa ampla (2л=63) и 5 других видов
III. Диплоспория—партеногенез и апоспория—партеногенез
Compositae
Antennaria carpatica (2л=40—42)
Parthenium argentaium (2n=54—144); P. incanum (2л=54—90)
Rosaceae
Potentilla verna (2n=42, 49, 84)
IV Адвентивная эмбриония
Dicotyledoneae
Anacardiaceae
Mangifera indica (2л=40, «52)
Betulaceae
Alnus rugosa (2л=28)
Buxaceae
Sarcococca pruniformls (2n=28) и другие виды
Cactaceae
Opuntia ficus-indica (2л=88) и другие виды
Capparidaceae
Capparis frondosa
Celastraceae
Celastrus scandens (2n=46)
Euonymus latifolius и другие виды
Clusiaceae
Clusia rosea; C. minor
Euphorbiaceae
Euphorbia dulcis (2n=12, 28)
Guttiferae
Garc ini a mangostana (2n«76)
Malphigiaceae
Hiptage madablota
Myrtaceae
Eugenia malaccensis (2л=22) и другие виды
Ochnaceae
Ochna serrulata (2 л=35)
Rutaceae
Citrus, многочисленные виды (2н= 18, 27, 36)
Triphasia aurantiola
Xanthoxylum bungei
Monocotyledoneae
Araceae
Spathiphyllum patina (2n=18)
Liliaceae
Allium odorutn (2л =16, 32)
Hosta coerulea (2л=60)
Nothoscordum fragrans (2n=16—19, 24)
Smilacina racemosa
Tulipa gesneriana (2n=24, 36)
Orchidaceae
Nigritella nigra (2л=64)
Zeuxine sulcata (2л=44)
Zygopetalum mackayi (2л=40)
Гл. 31. Агамоспермия
401
пией, подобно Nothoscordium fragrans и многим видам Citrus,
чем в группах с гаметофитным апомиксисом. Единственными из-
вестными диплоидами с гаметофитным апомиксисом являются,
по-видимому, некоторые члены комплексов Potentilla argentea,
Ranunculus auricomus, Hieracium umbellatum и Arabis holboel-
lii [317, 625, 780].
Мне не известно ни одного документированного случая ага-
моспермии у голосеменных. Не содержится таких примеров и в
сводном обзоре Фрикселла [223].
Ментон [554] перечисляет следующие примеры апогамных
папоротников (Polypodiaceae) с указанием в тех случаях, когда
это известно, уровней полиплоидии
Drtjopteris borreri (2х, Зх, 4х, 5х). D remota (Зх), D atrata (Зх)
Phcgopteris polijpoaioides
Cyrtomium falcatum. (Зх). C fortune! (Зх), С caryotideum (Зх)
Asplenium monanthes (Зх)
Pellaea atropurpurea (Зх)
Pteris cretica (2x, 3x, 4x)
У всех этих апогамных папоротников высокие хромосомные
числа — от 2п = 58 у одной формы Pteris cretica до 2п = 205 у
одной из форм Dryopteris borreri. Следует заметить, что боль-
шинство из них — нечетные полиплоиды, главным образом
триплоиды, хотя иногда встречаются и пентанлоиды [554]. О мно-
гих случаях апогаметии у Pteris сообщил также Уокер [871]
Клековски [441] отмечает, что у папоротников генетическая
система коррелирует с продолжительностью жизни гаметофитно-
го поколения. Большая продолжительность жизни гаметофитно-
го поколения обычно сочетается с половым размножением и
ауткроссингом. Апогаметия же часто наблюдается у растений
с короткой продолжительностью жизни гаметофитного поко-
ления.
Система размножения
Густафсон [317] исследовал 35 родов цветковых растений, у
которых имеются как агамоспермные виды, так и виды, размно-
жающиеся половым путем. Цель этого исследования состояла в
том, чтобы определить тип системы размножения у половых ро-
дичей агамоспермных растений. Полученные результаты можно
сгруппировать следующим образом В 10 из частично агамо-
спермных родов половые виды двудомные и(ийи) однодомные.
У 16 родов половые формы самонесовместимы. У 7 из этих ро-
дов механизм цветка способствует ауткроссингу. Лишь у одно-
го или, возможно, у двух из 35 родов, входящих в эту выборку,
обнаружены самооплодотворяющиеся цветки [317].
26-45
402 Часть восьмая. Специализированные генетические системы
Примеры агамоспермных растений с двудомными или одно-
домными родичами имеются в родах Elatostema, Balanophora,
Wikstroemia, Antennaria, Euphorbia и Alnus. Самонесовместимые
родичи агамоспермных растений встречаются у Calamagrostis,
Zephyranthes, Rubus, Parthenium, Taraxacum и Hieracium. Bur-
mannia — один из исключительных родов, содержащих самооп-
лодотворяющиеся виды, родственные агамоспермным видам
(]амонесовместимость оказалась наиболее обычной системой
размножения в выборке частично агамоспермных родов. Дву-
домность и однодомность, которые сравнительно редки у покры-
тосеменных в целом, у частично агамоспермных родов встреча-
ются с непропорционально высокой частотой. Из этого вытекает
чрезвычайно интересное общее заключение, что ауткроссинг,
осуществляемый при помощи той или иной системы размноже-
ния, преобладает у половых родичей почти всех агамоспермных
цветковых растений [317].
Гибридность и агамоспермия
Тесная связь между агамоспермией и гибридизацией отмечалась
многими исследователями, занимавшимися этой проблемой пос-
ле Эрнста [205]. Все агамоспермные цветковые растения, кото-
рые изучались генетически, оказались высокогетерозиготными.
Большинство из них — межвидовые гибриды или потомки гиб-
ридов. Более ранние обзоры относящихся к этой проблеме дан-
ных принадлежат Густафсону [317] и Стеббинсу [780].
Первая группа данных, приведшая к этому заключению, как
указывали Эрнст [205] и последующие авторы, вытекает из кар-
тины изменчивости большинства групп агамоспермных видов.
Такие группы часто содержат очень большое число слабо выра-
женных микровидов. Различные микровиды столь многочислен-
ны и столь взаимосвязаны морфологически, что удовлетвори-
тельная таксономическая трактовка этих групп становится фак-
тически невозможной. Подобная группа в целом по характеру
.изменчивости сходна с сильно увеличенным скоплением гибри-
дов.
Эта характерная картина изменчивости уже давно известна
у комплексов агамоспермных видов Hieracium, Taraxacum,
Antennaria, Crepis, Rubus, Crataegus, Poteniilla, Alchemilla, Poa,
^Calamagrostis и Citrus.
Для нескольких комплексов видов, в которые входят как
половые, так и агамоспермные популяции, было отмечено, что
агамоспермные формы содержат различные комбинации и ре-
комбинации морфологических признаков, имеющихся у двух или
нескольких половых видов. Такие данные, свидетельствующие
Гл. 31. Агамоспермия
403*
о гибридном происхождении агамоспермных видов, получены
для Antennaria [773, 774], для группы Crepis occidentalis [36],.
подрода Eubatus рода Rubus [315], Potentilla (см. [317]), Part-
henium [689] и Calamagrostis [626].
Прямые данные, свидетельствующие о естественной гибриди-
зации, получены для Antennaria [773, 774], Parthenium [689] и
Hypericum (см. [317]). Искусственные гибриды соответствуют
некоторым природным агамоспермным формам у Antennaria,
Potentilla и Rubus.
Как отмечалось ранее и как показывает табл. 31.1, преобла-
дающее большинство агамоспермных цветковых растений — по-
липлоиды В одной из предыдущих глав мы узнали, что у выс-
ших растений большая часть встречающихся в природе поли-
плоидов — амфиплоиды. Цитогенетические и морфологические
данные, свидетельствующие об амфиплоидности, получены для
следующих агамоспермных полиплоидов* Hieracium, Crepis, Ru-
bus, Potentilla, Роа и Calamagrostis (см. [317]).
Многие агамоспермные формы подобны межвидовым гибри-
дам: пыльца у них стерильная или полустерильиая, мейоз нару-
шен или пыльники абортивны Агамоспермные формы с муж-
ской стерильностью встречаются, например, у Taraxacum, Cre-
pis, Hieracium, Ranunculus, Hipericum, Crataegus, Potentilla, Ru-
bus. и Alnus [317].
Данные о мужской стерильности у пепсевдогампых апомик-
тов, подобных Taraxacum и Crepis, у которых пыльца не прини-
мает участия в агамоспсрмном образовании семени, можно ин-
терпретировать по-разному. Стабилизирующий отбор на муж-
скую фертильность у этих форм оолаблен, а следовательно, от-
сутствие фертильной пыльцы не обязательно свидетельствует о
гибридном происхождении [139, 317]. Однако у таких псевдогам-
ных апомиктов, как Potentilla, у которых для формирования се-
мян необходимо некоторое количество фертильной пыльцы, про-
дуцирование большого количества стерильной пыльцы после
нарушенного мейоза, несомненно, служит веским доказательст-
вом гибридности.
Факультативные апомикты, у которых эпизодически проис-
ходит половое размножение, иногда дают в Fi очень широкий
спектр вышепенцев (см. рис. 31.4). Масштабы расщепления в
потомстве Fi факультативного апомикта можно сравнить с рас-
щеплением в поколении F2 1ибрида, возникшего в результате
межвидового скрещивания в той же группе растений Такого
рода широкое расщепление, наблюдаемое у Potentilla, Rubus,
Sorbus и Citrus, вскрывает гибридную конституцию факульта-
тивно aiамоспсрмпою материнского растения Короче говоря
агамоспермпое растение ведет себя при размножении как меж-
видовой 1ибрид.
26*
404 Часть восьмая. Специализированные генетические системы
Образование зародыша бесполым путем связано с гибрид-
ностью также и у папоротников. Почти все апогамные папорот-
ники, изученные Ментон [554], имели гибридное происхождение.
Возникновение гаметофитного апомиксиса
Эмбриологические пути при гаметофитном апомиксисе проходят
вблизи тех мест, где обычно протекают мейоз и оплодотворение,
но обходят оба этих процесса. Всякая постоянная и устойчивая
агамоспермная линия должна обходиться и без мейоза, и без
оплодотворения. Между тем эти два процесса не зависят один
от другого. Интересно поэтому найти такие случаи, в которых
имеет место только одна из этих двух последовательных фаз
гаметофитного апомиксиса.
Примеры одной только апоспории обнаружены у Leontodon
hispiaus, Pieris hieracioides и у некоторых других видов сложно-
цветных, размножающихся половым путем. Зародышевые меш-
ки с ясно выраженными яйцеклетками или без них возникают
у этих видов из соматических клеток семязачатка. Однако эти
зародышевые мешки не дают начала зародышам. За апоспорией
не следует партеногенетического развития яйцеклетки или апо-
гамного развития ее клеток-спутников [60, 317]. Подобным же
образом в одной линии Parthenium argentatum нередуцирован-
ные яйцеклетки образуются в результате апоспории или дип-
лоспории, но затем эти клетки не претерпевают партеногенети-
ческого развития .[644].
Комплементарную ситуацию представляет собой гаплоидный
партеногенез, или развитие из редуцированного и неоп-лодотво-
ренного яйца гаплоидного зародыша. Это наблюдал Хагеруп
[324] у нескольких датских видов орхидей. Хагеруп [324] при-
водит иллюстрации партеногенетических гаплоидных зародышей
Listera ovata (n=17), Orchis strictifolius (n=20), Platanthera
chlorantha (л=21), Cephalanthera longifolia (n=16) и C. damn-
sonium (л =16). Эти гаплоидные индивидуумы должны быть
стерильными, и по этой, а также по другим причинам должны
представлять некое преходящее состояние в природе.
В царстве животных и у некоторых растений гаплоидный
партеногенез хорошо известен как лабораторное явление; но у
животных в отличие от высших растений он стал нормальным
процессом размножения в нескольких группах. Например, у всех
Hymenoptera и у некоторых Coleoptera, Hemiptera и Acarina
самцы развиваются в результате гаплоидного партеногенеза
[764, 887, 893].
Как указал Густафсон [317], наличия лишь одной фазы из
двухфазного процесса гаметофитного апомиксиса недостаточно
для того, чтобы агамоспермное размножение могло стать перма-
Гл. 31 Агамоспермия
405
нентным процессом. Однако существование каждой из этих изо-
лированных фаз по отдельности свидетельствует о тенденции в
направлении агамоспермии [317]. Агамоспермная система раз-
множения может возникнуть в том случае, когда две комплемен-
тарные тенденции — выпадение мейоза и диплоидный партено-
генез— встречаются одновременно у одною и того же орга-
низма
Что касается детерминации на генном уровне, то можно до-
пустить, что эти две тенденции — развитие в обход мейоза и
диплоидный партеногенез — контролируются отдельными гена-
ми или генными системами. В таком случае гамстофитный апо-
миксис можно рассматривать как выражение комплементарного
взаимодействия между этими отдельными генами
Аллели отдельных генов или генных систем, контролирую-
щих апомиксис, присутствовали, возможно, в предковых поло-
вых популяциях в виде рецессивных факторов Случайное скре-
щивание между носителями противоположных факторов могло
привести к образованию рецессивного комплементарного гено-
типа, который вел себя как агамоспермная форма. Сбалансиро-
ванная комбинация генов, детерминирующих агамоспермное
воспроизведение такого генотипа, можег, однако, легко разру-
шиться в потомстве этого нового агамоспермного растения в слу-
чае ауткроссинга [317, 644].
При адвентивной эмбрионии, когда новый зародыш развива-
ется из соматических клеток семязачатка, одного-единственпого
изменения в эмбриологическом пути оказывается достаточно,
чтобы обойти и мейоз, и оплодотворение. Следовательно, воз-
никновение адвентивной эмбрионии из предкового полового цик-
ла в эмбриологическом, а возможно, и в генетическом аспекте
проще, чем возникновение гаметофитного апомиксиса.
Поведение при скрещиваниях
Наследование гаметофитного апомиксиса можно изучать на по-
томках от скрещиваний между агамоспермными формами. Такие
скрещивания производились в ряде групп растений. Большая
часть опубликованных данных посит предварительный характер,
и они неубедительны. Результаты некоторых экспериментов по
гибридизации, проведенных на Potentilla, Роа, Calamagrostis и
Rubus, представляют, однако, значительный интерес и будут из-
ложены в этом разделе
У Potentilla можно сравнить результаты внутривидовых скре-
щиваний между агамоспермными формами с результатами меж-
видовых скрещиваний. Внутривидовые скрещивания Potentilla
vernaxP. verna и Р. tabernaemontanaXP. tabernaemontana дают
гибридов Fi, которые агамоспермны подобно двум своим роди-
406
Часть восьмая. Специализированные генетические системы
тельским формам. В отличие от этого при межвидовых скрещи-
ваниях между Р. recta и Р. adscharica, а также Р. canescens и
Р. verna поколение Fi состоит главным образом или целиком из;
половых гибридов. Следовательно, у Potentilla агамоспермное
состояние разрушается при межвидовой, но не при внутривидо-
вой гибридизации (ср. [317]).
Клаузен и др. [104] скрещивали шесть агамоспермных видов
Роа в восьми различных сочетаниях. Агамоспермными родитель-
скими формами были Р. pratensis, Р. ampla, Р. compressa*
Р. canbyi, Р. scabrella и Р. arida. Потомства разных индивиду-
умов из Fi выращивали по отдельности и получили семьи F2.
Некоторые из индивидуумов F2 были отобраны в качестве ро-
дительских особей для получения отдельных семей F3. Семьи
F2 и F3 обследовали для выяснения наличия или отсутствия рас-
щепления. Семьи, в которых наблюдалось расщепление, расце-
нивались как результат полового размножения родительских ин-
дивидуумов, тогда как семьи, все члены которых были одно-
родными и походили на материнское растение, расценивались
как результат агамоспермного размножения [104].
От скрещивания Роа ampla с Р. pratensis было получено
35 растений Fi, каждое из которых анализировали по потомст-
ву, получив 35 семей F2. Из этих семей F2 20 были признаны
агамоспермными, а 15 — половыми. Затем от ряда растений F2,
взятых из семей обеих категорий, было получено в Fs 13 агамо-
слермных и 9 половых семей. Другие межвидовые гибридные
комбинации агамоспермных Роа подобным же образом расщеп-
лялись в F2 и F3 на агамоспермные и половые потомства [104].
В сущности, многие из агамоспермных семей не были совер-
шенно однородными и матроклинными, но содержали некоторое
число половых выщепенцев. Доля половых индивидуумов в
семьях F2, в остальном агамоспермных, колебалась от 10 до
90%. Отсюда следует, что некоторые агамоспермные индивиду-
умы F] высокоагамоспермны, тогда как другие агамоспермии:
лишь в слабой степени [104].
Самый же главный момент состоит в том, что расщепление
по агамоспермии у гибридов между агамоспермными видами
Роа носит сложный характер. В F2 и в F3 иногда происходят ре-
версии от агамоспермии к половому размножению. И наоборот,
некоторые половые особи F2 дают начало агамоспермным F3.
Ряд скрещиваний между агамоспермами был описан у еже-
вики. Скрещивание между Rubus plicatus и R. caesius дало
крайне изменчивое поколение Fj. У гибрида Fi от скрещивания
Rubus polyanthemus с R. insularis пыльца была фертильной, и
от него было получено поколение F2, которое очень сильно рас-
щеплялось. Подобная же изменчивость наблюдалась в Fi и Fj
от скрещиваний между другими агамоспермными видами Rubus.
Гл. 31, Агамоспермия
407
И в данном случае гибридизация приводит к разрушению ага-
моспермии и частичному восстановлению полового размножения
£315]
У Calatnagrostis скрещивание между двумя половыми вида-
ми, С. epigeios и С. arundinacea, даст гибридов Fi и F2, которые
образуют некоторое число диплоспоровых зародышевых мешков
[624, 626]. Гены, детерминирующие агамоспермию, содержатся,
следовательно, у полового вида и могут соединиться при гибри-
дизации. Это важный факт, поскольку он показывает, каким
образом такой агамоспермный вид Calatnagrostis, как С purpu-
rea, о котором известно, что он представляет собой гибридное
производное С. epigeios и другого полового вида, может возник-
нуть в природе [624, 626].
Экспериментальные данные свидетельствуют о том, что га-
метофитный апомиксис наследуется, как и следовало ожидать,
сложным образом. Гибридизация между половыми видами мо-
жет иногда привести к сборке генной комбинации, детермини-
рующей агамоспермию. И наоборот, гибридизация между двумя
агамоспермными видами или же между одним агамоспермным
и одним половым родительским видом может разрушить эту же
самую генную комбинацию и восстановить половое размножение
у части потомства. Эти половые типы могут затем воспроизво-
дить новый набор агамоспермных форм в следующем поко-
лении.
Расщепление
В половом потомстве факультативных агамоспермных форм
обычно наблюдается высокая степень индивидуальной изменчи-
вости. Особенно заметно расщепление по морфологическим приз-
накам, оно затрагивает также число хромосом и физиологичес-
кую мощность.
На рис. 31.4 показана индивидуальная изменчивость призна-
ков плода у полового потомства Fi грейпфрута Citrus paradisi;
родительским растением, давшим пыльцу, был лимон (С. Limon).
Факультативно агамоспермные формы секции Moriferus рода
Rubus после ауткроссинга во многих гибридных комбинациях
давали очень изменчивое потомство. Некоторые из этих комби-
наций, такие, как Rubus plicatus^R. caesius, дающие расщеп-
ляющееся поколение Fi, упоминались в предыдущем разделе.
Ауткроссипги между R plicatus или другими агамоспермными
видами, с одной стороны, и половым диплоидом R. tomentosus —
с другой, дают поколения Ft, которые по морфологии и плодо-
витости сходны с расщепляющимся поколением F2 [315].
Гщппа Rubus acutus представляет собой сложную совокуп-
ное 1ь агамоспермных микровидов на Скандинавском полуостро-
408
Часть восьмая. Специализированные генетические системы
Рис 31.4 Расщепление по признакам плода в половом потомстве факульта-
тивно агамоспермного растения Citrus (из Frost по (717]).
А Женское родительское растение — грейпфрут С parodist Б. Мужское ро-
дительское растение — лимон С limon. В—Е. Четыре разных индивидуума Fj.
ве. Многие из этих микровидов — гибриды (либо гибриды Fb
либо выщепенцы Fa), образовавшиеся в результате скрещива-
ний между R. caesius (4х) и различными предсуществовавшими
видами группы R. acutus. Эти микровиды различаются по приз-
накам, характеризующим листья, стебель, колючки, опушенность
и т. п.
Лидфорсс (см. [315]) обнаружил в Скании (Швеция) гиб-
ридное растение, которое он счел гибридом Fi от скрещивания
Rubus gothicus и R. caesius (R. gothicus — один из микровидов
группы R. acutus). Пыльца этого растения была полностью фер-
тильной. Его пересадили в Ботанический сад в Лунде, где оно
дало многочисленное поколение F2, а затем несколько потомков
F3 и F4.
Поколение F2 от скрещивания Rubus gothicus и R. caesius
представляло большой интерес, поскольку оно содержало ряд
выщепенцев, сходных с различными, имеющими собственные
названия микровидами группы R. acutus. Так, некоторые инди-
видуумы были сходны с R. pruinosus— у них были такие же
листья с семью прилистниками и прямые колючки. Другие рас-
тения F2 походили на R. acutus, R. permixtus, R. wahlbergii,
R. gothicus, R. polycarpus, R. ruedensis или R. caesius. Как и сле-
довало ожидать, в F2 были и такие растения, которые не походи-
ли ни на один из именованных микровидов, встречающихся в
природе [315].
Гл 31 Агамоспермия
409
Нарушения мейоза у факультативно агамоспермпого вида
могут привести к образованию функциональных гамет с различ-
ным числом хромосом В таких случаях возникают зиготы с чис-
лами хромосом, образующими анеуплоидпый ряд. Такая анеу-
плоидиая изменчивость наблюдалась у потомков Роа и других
злаковых
Гран [310] определял число хромосом у отдельных растений
в поколениях Fi от нескольких межвидовых скрещиваний между
факультативно агамоспермными представителями Роа. Одно из
таких скрещиваний (Р. scabrellaXP pratensis), в котором число
хромосом у родительских растений было 2п = 83 и 68 соответст-
венно дало в Fi потомков, которые по числу хромосом образо-
вали следующий ряд. 2гг, = 65, 66, 67, 68, 68, 70, 70, 71, 71, 71, 74,
75, 75. 79 При другом скрещивании между Р ampla (2п = 64) и
Р pratensis (2п = 68) число хромосом у гибридов Fi варьирова-
ло от 2л = 60 до 2п— 117. Подобная анеуплоидная изменчивость
была обнаружена в поколениях Fi от скрещиваний разных форм
Роа примерно в 16 гибридных комбинациях [310]
Клаузен и его сотрудники изучали физиологическую мощность
потомков от межвидовых скрещиваний между агамоспермными
Роа. В общем гибриды Fi были жизнеспособными, так же как и
большая часть их агамоспермных потомков Fs. Однако среди
половых потомков Fo, полученных от тех же самых межвидовых
скрещиваний, наряду с мощными растениями было много сла-
бых и нежизнеспособных индивидуумов Такое расщепление и
рекомбинация по мощности среди половых потомков гибридов
Роа представляет собой обычное разрушение гибридов Агамо-
спермные потомки тех же самых гибридов в значительной мере
избегают этого [104]
Характер изменчивости
Основным источником новых вариаций в группе агамоспермных
растений служит гибридизация. Самыми обычными гибридными
комбинациями являются- половая форма X факультативно ага-
моспсрмная форма и факультативно агамоспермная форма X
Хфакультативно агамоспермная форма. В гибридизации в ка-
честве отцовской формы могут участвовать даже облигатные
агамоспермы. при условии, что они способны продуцировать хо-
тя бы незначительное количество фертильной пыльцы. Гибриди-
зирующие родительские растения могут находиться на любом
уровне генетической дифференциации — от малых биотипов до
видов, связанных лишь отдаленным родством.
Вторым источником изменчивости служит автосегрегация.
Автосегрегацией называют спонтанные изменения в генотипе
410
Часть восьмая. Специализированные генетические системы
яйцеклеток, развивающихся без оплодотворения. Так, у Taraxa-
cum мейоз в материнской клетке зародышевого мешка может
быть нарушен, что приводит к образованию яйцеклетки, у кото-
рой число хромосом больше или меньше, чем у материнского
растения. Или же гетерогенетическая конъюгация и кроссинго-
вер в материнской клетке зародышевого мешка могут привести
к образованию генотипически измененной яйцеклетки. Такие
яйцеклетки, развиваясь партеногенетически, могут дать расте-
ния, отклоняющиеся по фенотипу. Автосегрегация может проис-
ходить как у облигатно, так и у факультативно агамоспермных
форм [317].
Третьим источником изменчивости служат почковые мутации,
приводящие к развитию фенотипически измененных побегов. Та-
кие вегетативные спорты были обнаружены несколько раз у
агамоспермной ежевики [315, 317].
Отклоняющиеся индивидуумы, независимо от способа своего
возникновения, могут затем размножаться бесполым путем.
Агамоспермная система обеспечивает создание изменчивости
различными способами и чередующееся с этими вспышками из-
менчивости точное воспроизведение любых благоприятных новых
вариантов путем бесполого размножения.
В предыдущем разделе было описано, как одно-единствен-
ное гибридное растение из группы Rubus acutus произвело по-
ловых потомков F2, сходных с различными встречающимися в
природе агамоспермными микровидами, относящимися к той же
самой группе видов. Разных выщепенцев F2 можно было отнести
к R. acutus sensu stricto, R. pruinosus, R. polycarpus и т. д. При-
родные микровиды, вероятно, возникли таким же образом, т. е.
как половые выщепенцы гибридов, а затем достигли статуса
микровидов в результате увеличения численности путем агамо-
спермии и расширения занимаемой области [315].
В предыдущем разделе было также показано, что в группе
Роа pratensis у половых потомков гибридов существует анеупло-
идная изменчивость. И в этом случае было обнаружено анало-
гичное состояние анеуплоидии в природе.
Лёве [523] подсчитал число хромосом у отдельных растений
одного из членов группы Рао pratensis, известного под назва-
нием Р. irrigata и растущего на участке диаметром 10 м в
Исландии. На этой небольшой площади Лёве нашел растения
с 2п=82, 84, 87, 91, 95, 98, 105, 108, 111, 112, 113 и 119 [523].
Таксономический вид Р. pratensis в целом на протяжении об-
ширной занимаемой им циркумбореальной области образует
почти непрерывный анеупЛоидный ряд от 2п=22 до 2/1=147
[98, 310, 523].
Гл. 31. Агамоспермия
411
Преимущества агамоспермии
Агамоспермия — это одна из генетических систем, которая осно-
вана на гибридности и которая делает возможным воспроизве-
дение гибридного генотипа. Уместно спросить: в чем же состоят
особые адаптивные преимущества агамоспермной генетической
системы? В 30-х годах Дарлингтон высказал (но не развил) две
точки зрения, послужившие отправными в рассуждениях после-
дующих авторов. В первом издании «Современных успехов ци-
тологии» [138] Дарлингтон рассматривал агамоспермию под руб-
рикой «стабилизации гибридности». В последующих работах
Дарлингтона [140, 141, 145] концепция стабилизации гибридно-
сти уже не выдвигается. Вместо этого в «Эволюции генетиче-
ских систем» [141, 145] агамоспермия представляется как «из-
•бегание стерильности».
Необходимо заметить в скобках, что Дарлингтон в назван-
ных выше работах на самом деле пользуется термином апомик-
сис. Однако в данной книге термин апомиксис используется в
том смысле, какой в него вкладывал Густафсон [317], т. е. для
обозначения более широкого круга процессов бесполого разм-
ножения, чем апомиксис Дарлингтона. Апомиксис в понимании
Дарлингтона — это по существу то же самое, что в данной кни-
ге обозначается термином агамоспермия.
Представление об агамоспермии как о способе размножения,
позволяющем избежать стерильности, имеет широкое примене-
ние. Многие агамоспермные формы в различных родах расте-
ний— стерильные или полустерильные гибриды, как отмечалось
ранее в этой главе. Нечетные полиплоиды, в особенности три-
плоиды, обычно бывают стерильными и часто встречаются сре-
ди агамоспермных растений.
Существуют, однако, плодовитые агамоспермные гибриды.
К ним относятся гибриды между видами рота Citrus' мейоз у
них протекает нормально, и они плодовиты [677, 880]. Но при
этом многие гибридные формы Citrus размножаются путем
адвентивной эмбрионии. В данном случае агамоспермия не поз-
воляет избежать стерильности.
Таким образом, концепция стабилизации гибридности при-
ложима к известным случаям агамоспермии более широко и в
более общей форме, чем концепция избегания стерильности.
Представим себе, что в результате естественной гибридиза-
ции возникает некий высокогстерозш отный генотип, обладаю-
щий пенными в адаптивном отношении гетерозисными свойства-
ми Это1 lenoTiin может представлять собой гибрид Fi или отно-
ситься к одному из более поздних поколений, он может быть
стерильным или плодовитым. В любом случае данный гетеро-
зиготный генотип этого гибрида будет разрушен в результате
412
Часть восьмая Специализированные генетические системы
нормального полового размножения. Однако агамоспермия дает
возможность этому самому гибриду воспроизводиться в чистоте
при помощи семян. Агамоспермный гибрид может воспроизво-
дить точные копии своего ценного в адаптивном отношении ге-
нотипа в многочисленных растениях, развившихся из продуци-
руемых им семян.
Тот же самый процесс копирования, который мы только что
описали применительно к агамоспермии, может осуществляться,
и иногда осуществляется, путем вегетативного размножения.
Однако для семян обычно характерны некоторые адаптивные
свойства, а именно состояние покоя, устойчивость к различным
воздействиям и в особенности способность к распространению,
которыми не наделены в такой же степени органы вегетативного
размножения растений того же вида. Агамоспермия как генети-
ческая система унаследовала эти особые преимущества, прису-
щие размножению семенами.
Амфиплоидия — другая генетическая система, которая мо-
жет создавать и часто создает копии гибридного генотипа.
В природе возникают, однако, определенные типы нумерических
гибридов, которые не могут сохраняться при помощи амфиплои-
дии, но могут поддерживаться при помощи агамоспермии.
Нечетные полиплоиды — триплоиды, пентаплоиды и т. д.» —
если они обладают собственной адаптивной ценностью, могут
сохраняться при помощи последовательных поколений расте-
ний, развившихся из семян, которые образовались в результате
агамоспермии, но не в результате амфиплоидии. Естественные
гибриды между родительскими видами с высокими полиплоид-
ными числами хромосом могут оказаться не способными удваи-
вать число своих хромосом, не выходя за пределы, являющиеся
пороговыми для упорядоченного клеточного деления В этом
случае опять-таки агамоспермный механизм размножения поз-
воляет выйти из тупика. Наблюдаемую среди агамоспермных
растений высокую частоту полиплоидов двух типов — нечетных
полиплоидов и высоких полиплоидов — можно интерпретировать
как аргумент в пользу представления о том, что агамоспермия
способна привести к успеху в тех ситуациях, в которых амфи-
плоидия терпит поражение.
Как подчеркивают Густафсон [317] и другие авторы, агамо-
спермия не обязательно означает полный отказ от полового раз-
множения. В агамоспермных линиях время от времени может
происходить половое размножение, давая начало новым гибрид-
ным генотипам. Новые гибриды, образовавшиеся таким обра-
зом, могут затем размножаться в процессе еще одного цикла
агамоспермии. В результате внутри данной агамоспермной груп-
пы могут время от времени возникать новые адаптивные гетеро-
зиготы, которые затем воспроизводятся в свою очередь в массо-
вых количествах при помощи агамоспермного механизма.
Гл. 31. Агамоспермия 413
Агамоспермия, следовательно, обладает известными собст-
венными преимуществами по сравнению с другими конкретными
генетическими системами. Агамоспермия делает возможным
размножение редкого гомоплоидного гетерозиготного выщепен-
ца в гибридной популяции растений, размножающихся при по-
мощи ауткроссинга, тогда как половое размножение обеспечить
этого не может. Она сохраняет нечетные полиплоиды и высокие
вторичные анеуплоиды, что невозможно при амфиплоидии. Она
делает возможным размножение гибридов с высокими поли-
плоидными числами хромосом, когда дальнейшее удвоение числа
хромосом и амфиплоидия были бы безрезультатными. Она обес-
печивает бесполое размножение семенами, на что неспособно
вегетативное размножение.
•32 Агамный комплекс
-Сочетание процессов естественной гибридизации и агамосперм-
ного размножения привело с течением времени к возникновению
ряда микровидов, несущих в различных рекомбинациях призна-
ки родительских видов. Образующаяся в результате структура
представляет собой агамный комплекс, в котором агамосперм-
ные гибридные производные в процессе филогенеза наложились
на первоначальные половые виды.
Структура простого агамного комплекса схематически пока-
зана на рис. 32.1. На этом рисунке можно видеть многочислен-
ные агамоспермные производные, заполняющие разрывы и мас-
кирующие прерывистости в морфологических признаках между
диплоидными половыми видами. Изменчивость в агамных комп-
.лексах— это наиболее сложные картины изменчивости, извест-
ные у растений. Для форм, входящих в агамные комплексы
Rubus и Hieracium, были опубликованы тысячи латинских наз-
ваний видового или инфравидового ранга.
Проблема вида в том или ином агамном комплексе частично
разрешима. Диплоидные половые виды, если они сохранились
.до настоящего времени, можно обнаружить, дать им названия
и нанести на карту их ареалы. Отдельные агамоспермные произ-
водные, если они обладают достаточно однородной и ясно вы-
раженной комбинацией признаков в пределах данной географи-
ческой области, также следует признать таксономическими ви-
дами и дать им названия.
Однако в некоторых частях агамного комплекса проблема
вида неразрешима. Между диплоидными половыми видами и
достигшими наибольшего успеха и более широко распростра-
нившимися агамоспермными видами находится множество ага-
моспермных вариантов более низкого ранга — от малых биоти-
пов до локализованных агамоспермных микровидов. Какие из
них следует выделять таксономически и в каком таксономичес-
ком ранге, зависит от точки зрения и соображений удобства, а
на этот счет мнения исследователей часто расходятся.
Филогенетический анализ данного агамного комплекса со-
пряжен, кроме того, со многими трудностями, вытекающими из
большой сложности этой проблемы. Стадии анализа, обрисован-
ные выше применительно к полиплоидным комплексам, вполне
приложимы к агамным комплексам. Некоторые агамные комп-
лексы, такие, как группа Crepis occidentalis, подвергались де-
тальному цитотаксономическому анализу, а другие, подобно сек-
ции Moriferus рода Rubus, — детальному биосистематическому
-анализу, но ни один большой агамный комплекс не был исчерпы-
евающим образом изучен в генетико-таксономическом плане.
Гл. 32. Агамный комплекс
415
Комплекс Crepis occidentalis
Группа Crepis occidentalis (х=11)—совокупность однолетних
травянистых'растений, растущих в западных областях Северной
Америки. Эти растения встречаются на сухих склонах гор; центр
их распространения находится на Колумбийском плато, а ареал
тянется от Тихоокеанского побережья до Скалистых гор. В эту
группу входят половые диплоиды и многочисленные агамосперм-
ные полиплоиды [36].
Бэбкок и Стеббинс [36] проанализировали группу Crepis
occidentalis на основе гербарных материалов, полевых исследо-
ваний, цитотаксономических исследований и наблюдений за жи-
выми растениями, пересаженными из природных местообитаний.
Гибридизацию в экспериментальных условиях провести не уда-
лось, поэтому прямые данные относительно генетических взаи-
моотношений между видами отсутствуют.
Бэбкок и Стеббинс разбили вариации в группе Crepis occiden-
talis на 9 таксономических видов: С. acuminata, С. bakeri*
Рис 32 1 Структура простого агамного комплекса [36].
416 Часть восьмая. Специализированные генетические системы
С. barbigera, С. exilis, С. intermedia, С. modocensis, С. monticola,
С. occidentalis и С. pleurocarpa. В большинстве случаев эти так-
сономические виды содержат диплоидную половую популяцию
и различные агамоспермные формы, которые морфологически
более или менее сходны с диплоидом. Агамоспермные формы,
если они вообще имеют названия, рассматриваются как трино-
минальные категории forma apomictica. Таким образом, в группе
С. occidentalis именованные таксономические виды представляют
собой сложные и искусственные единицы.
В эту группу входят семь известных половых диплоидных
видов. У следующих таксономических видов — Crepis acuminata,
С. bokeri, С. exilis, С. modocensis, С. monticola, С. occidentalis
и С. pleurocarpa — они рассматриваются как инфравидовые еди-
ницы. В группе С. occiaentalis половые диплоидные популяции
составляют основные биологические виды. Эти семь диплоидных
видов хорошо различаются по морфологическим признакам
Лишь один из них (С. acuminata 2х) имеет широкое географиче-
ское распространение; ареалы остальных шести видов относи-
тельно ограничены и перекрываются симпатрически в различных
местах на северо-западе Тихоокеанского побережья [36].
Все агамоспермные члены группы Crepis occidentalis — поли-
плоиды (основное число х=11), достигающие октоплоидного
уровня. Среди них есть все возможные нечетные полиплоиды
(Зх, 5х, 7х). Морфологически некоторые из агамоспермных ти-
пов очень сходны с каким-либо одним диплоидным видом, но у
большинства из них комбинируются признаки двух или несколь-
ких диплоидных видов. Именно в этой суперструктуре полипло-
идных агамоспермных форм можно обнаружить интерградацию
между различными крайними комбинациями морфологических
|$изнаков [36].
Агамоспермные популяции обличаются друг от друга по
признакам листьев, стеблей, соцветий, цветков и семянок. Неко-
торые из них имеют локальное, а другие широкое географичес-
кое распространение. Те популяции, которые распространили
свою отличительную совокупность признаков по сколько-нибудь
значительной географической области, получили неформальное
таксономическое признание. Им присвоены триноминальные ла-
тинские названия неформальной категории forma apomictica
(apm), как уже упоминалось выше. На рис. 32.2 изображены
соцветия (корзинки) девяти получивших названия ага.мосперм-
ных микровидов из группы Crepis occidentalis. Примером хоро-
шо выраженного агамоспермпого микровида служит С. pleuro-
carpa apm. grayi, форма встречающаяся в северо-восточной час-
ти Калифорнии/ Это, возможно, пентаплоид. На основе морфо-
логических признаков его считают гибридным производным по-
ловых диплоидных форм С. pleurocarpa и С. occidentalis Другой
Л Б В
ГДЕ
Рис 32 2. Соцветия девяти агамоспермных микровидов Crepis occidentalis
(по 1(36]).
А —С.о. apm. humilior, Калифорния. Б — С. о. арт. nuttallu, Вашингтон.
В — С.о. арт. calyculata, Колорадо. Г —С.о. арт. grayi, Юта. Д — С.о. арт.
elliptica, северная часть Калифорнии. Е — С.о, арт. deltoidea, Альберта.
ЛС — Со. арт rydbergii, Монтана. З — С.о. арт. hamiltonensis, северная
часть Калифорнии. И — Со арт. crassa, Вашингтон.
27—45
418
Часть восьмая. Специализированные генетические системы
агамоспермный вид, распространенный в горах северо-восточной
части Калифорнии, — С. acuminata apm. sierrae, вероятно пред-
ставляющий собой триплоид.
Всего в монографии Бэбкока и Стеббинса приводится 113 та-
ких форм, получивших название апомиктических. Это лишь не-
большая доля всего множества обособленных агамоспермных
популяций Crepis в западной части Северной Америки.
Число агамоспермных микровидов относителньо велико в об-
ластях, находящихся вблизи центра распространения половых
популяций. Половые и факультативно апомиктические популя-
ции являются непрерывным источником возникновения новых
агамоспермных микровидов в центральных частях ареала. По
периферии области распространения всего комплекса, напротив,
число микровидов меньше, но каждый из них имеет более об-
ширный ареал. Здесь некоторым отдельным агамоспермным
формам, очевидно, благоприятствовал естественный отбор, по-
скольку они были лучше приспособлены к региональным усло-
виям среды, и эти формы размножились, заселив обширные тер-
ритории [36].
Филогенетическую историю комплекса Crepis occidentalis
можно воспроизвести в общих чертах по косвенным данным.
Современное географическое распространение каждого из по-
ловых диплоидных видов коррелирует с распространением опре-
деленных климатических и эдафических провинций, сложивших-
ся в плиоцене и в раннем плейстоцене. Следовательно, ареалы
дицлоидных видов, вероятно, установились в этот же период, от
раннего плиоцена до раннего плейстоцена, в ответ на установ-
ление более сухого климата и излияния лав [36].
Диплоидный вид, после того как он обосновался, гибридизи-
ровал в различных комбинациях, давая аллотетр аплой дных про-
изводных. Процесс гибридизации продолжался, возможно, до
недавнего времени и привел к возникновению агамоспермных
микровидов, представляющих собой вторичных и третичных гиб-
ридов, так как некоторые из этих микровидов имеют высокие
полиплоидные числа хромосом и связаны сложными родственны-
ми взаимоотношениями. Позднее, в течение плейстоцена, поли-
плоидные агамоспермные формы широко распространились, вы-
теснив диплоидные половые формы, которые оказались ограни-
ченными двумя центрами распространения в северо-западной
части тихоокеанского побережья Северной Америки.
Европейские ежевики
Обширный род Rubus (я=7) делится на четыре подрода; самый
сложный из них —подрод Rubus, к которому относятся ежевики.
Подрод Rubus образовал разнообразные совокупности видов и
Гл 32 Агамный комплекс
419
микровидов в ряде умеренных областей земного шара1 в Европе,
Азии, Средиземноморской области, в восточной части Северной
Америки, ла тихоокеанском побережье Северной Америки и в
районе Кордильер в Центральной и Южной Америке.
Ежевики Евразии и восточной части Северной Америки об-
разуют одну подгруппу Rubus, формально выделяемую как сек-
ция Moriferus Флора европейских ежевик — наиболее хорошо
изученная часть этой секции, и служит основным предметом
обсуждения в данном разделе Опа делится па две естественные
ветви, известные под названиями Moriferi Veri и Rubi Corylifo-
lii. Однако европейские ежевики не представляют собой само-
стоятельной естественной единицы, обособленной от членов сек-
ции Moriferus, произрастающих в восточной части Северной
Америки и в Азии и некоторые виды ежевики из двух послед-
них областей, несомненно, филогенетически связаны со своими
европейскими родичами [314, 315, 317]
Ежевики (подрод Rubus) и европейские ежевики в частно-
сти (часть секции Moriferus) изучались рядом систематиков
и I светиков — Фоккс, Лидфорссом, Густафсоном и другими
Побудительной причиной таких исследований, как таксономиче-
ских, так и генетических, была «почти неисчерпаемая изменчи-
вость этих ежевик» [317] Пытаясь отразить эту изменчивость в
таксономическом плане, различные авторы па протяжении ряда
лет опубликовали около 4—5 тыс. латинских названий Эти наз-
вания относятся к единицам самого разного уровня — от широко
распространенных видов до узко локализованных форм.
Для того ч-обы проиллюстрировать масштабы изменчивости
у европейских ежевик Гус1афсоп изложил вкратце систему* их
классификации созданную более ранним исследователем этой
I руппы, Судре. Судре различает в группе Moriferi Veri 691
микровид; все эти микровиды он объединяет в 109 таксономиче-
ских видов В одной лишь Европе существует около 1930 имено-
ванных вариететов Moriferi Veri [317] Другая основная ветвь
европейских ежевик, Rubi Corylifolii, содержит один таксономи-
ческий вид (Rubus caesius), по бесчисленное множество микро-
видов [315]
Европейские ежевики образуют полиплоидный ряд (х = 7).
В нем представлены следующие уровни плоидности- 2%, Зх, 4х,
5х, 6х и, возможно, 7х. Кроме того, в этой группе имеется не-
сколько вторичных анеуплоидов с числом хромосом, отклоняю-
щимся от полиплоидного. Большая часть этих микровидов —
тетраплоиды. Несколько триплоидных и пентаплоидных микро-
видов занимают обширные ареалы [315, 317]
Ежевики восточной части Северной Америки также образуют
полиплоидный ряд с основным числом хромосом х=7; среди них
есть трийлОиды,' пентаплоиды и гексаплоиды, а также несколько
420
Часть восьмая. Специализированные генетические системы
диплоидных видов [315]. В отличие от этого все формы, произ-
растающие на тихоокеанском побережье, — высокие полиплоиды
С уровнем плоидности от 6х до 12х [71, 102].
Полиплоидные европейские ежевики являются по большей
части факультативно агамоспермными формами. Установлено,
что многие из них — гибриды или гибридные производные, о чем
свидетельствуют в разных случаях морфологические признаки,
стерильность пыльцы, состояние нечетной полиплоидное™ и (или)
высокий уровень расщепления при возврате к половому 'размно-
жению. Было также обнаружено, что и у восточноамериканских
ежевик существует агамоспермия. Помимо образования агамо-
спермных семян ежевика распространяется, как это хорошо из-
вестно, при помощи вегетативного размножения [315, 317].
В Европе большинство агамоспермных форм образуют ге-
ографически локализованные микровиды. Некоторые из этих
локальных микровидов достаточно обособлены; другие же гиб-
ридизируют и интерградиру ют с родственными микровидами.
Среди европейских агамоспермных форм некоторые образовали
географически широко распространенные популяции, превра-
тившись, таким образом, в агамоспермные виды.
Филогенетическую основу агамного комплекса у европейских
ежевик образуют диплоидные половые виды. В западной части
Евразии известно пять диплоидных видов; Rubus bollei, R. inca-
nescens, R. moschus, R. iomentosus и R. ulmifolius (2n=14). По-
лагают, что и некоторые другие виды, такие, как R. alnicola, —
диплоиды. Возможно также, что в эволюционном развитии евро-
пейского агамного комплекса сыграли некую роль и другие по-
ловые диплоиды, произрастающие в восточной части Северной
Америки, например R. allegheniensis [315, 317].
Все диплоидные половые виды европейских ежевик принад-
лежат к группе Moriferi Vert. Они хорошо отличаются от агамо-
спермных микровидов и видов, а также друг от друга. Некото-
рые из них имеют широкое, а другие — ограниченное распростра-
нение.
Rubus tomentosus и R. ulmifoliusхарактеризуются обширными
ареалами, тянущимися от южной части Европы до Малой Азии;
R. moschus встречается в горах Кавказа; R. incanescens — на
западе Средиземноморской области, a R. bollei — на Канарских
островах. Возможный диплоид R. alnicola распространен в Пи-
ренеях и горах Юры. В тех местах, где эти виды встречаются в
природе, они часто гибридизируют с образованием стерильных
гибридов. Многие из агамоспермных микровидов Moriferi Veri—
давнишние или недавние производные таких гибридов [315, 317].
В целом диапазон изменчивости агамоспермных Moriferi Ve-
ri в Европе сильно выходит за пределы крайних отклонений в
морфологических признаках, наблюдаемых у европейских ди-
Гл. 32. Агамный комплекс
421
плоидов. Но некоторые европейские агамоспермы морфологиче-
ски очень близки к некоторым восточноамерикапским дипло-
идам, таким, как Rubus allegheniensis. Из этого можно сделать
вывод, что среди диплоидных половых предков агамного комп-
лекса Европы были виды, распространение которых в настоящее
время ограничено восточной частью Северной Америки, а также
различными хребтами в западной части Евразии (315, 317]
В группе Rubi Corylifolii диплоиды неизвестны. В эту группу
входит, однако, сизая ежевика (Rubus caesius, 2n = 28)—алло-
тетраплоид, происходящий от диплоидных видов, которые, ве-
роятно, уже вымерли. R caesius представляет собой факульта-
тивно агамоспермный вид. Он свободно гибридизирует с други-
ми видами и микровидами ежевики, но изолирован от них таки-
ми барьерами, как стерильность и нежизнеспособность гибридов.
Подобно диплоидам в группе Moriferi Veri, он образует обособ-
ленный естественный вид [315, 317].
Rubus caesius 1ибридизирует с диплоидами R. ulmifolius и
R tomentosus, а также с другими диплоидными и полиплоидны-
ми видами ежевики Mhoi очисленные агамоспермные микровиды
Rubi Corylifolii образовались в результате расщепления среди
потомков от происходивших ранее гибридизаций между Rubus
caesius и различными представителями Moriferi Veri. Другие
агамоспермные Rubi Corylifolii — продукты вторичных возврат-
ных скрещиваний между Rubus caesius и другими членами груп-
пы Rubi Corylifolii Так, естественные гибриды между R caesius
и R gothicus (Corylifolii) в результате расщепления дают нача-
ло формам, сходным с другими именованными микровидами
Rubi Corylifolii, такими, как R acutus, R wahlbergii и R pruino-
sus [314, 315, 317].
Обыкновенная малина Rubus idaeus (2п= 14), принадлежа-
щая к другому подроду, Idaeobatus, гибридизирует с R caesius
(2п = 28). Триплоидные гибриды стерильны. Однако иногда из
нередуцированных гамет образуются плодовитые или пол}пло-
довитые тетраплоидные и гексаплоидные гибриды Эти гибриды
в результате возвратных скрещиваний с R caesius вносят в
группу R caesius отдельные гены малины Некоторые агамо-
спермные формы Rubi Corylifolii, по-видимому, представляют со-
бой продукты гибридизации R idaeus и R caesius Таким обра-
зом. обогащению изменчивости в комплексе европейских ежовик
способствовал также один диплоидный вид, не принадлежащий
к подроду Rubus [317]
Диплоидные половые виды ежевики в Европе в настоящее
время встречаются в виде реликтов на юге Средиземноморской
области. Некоторые из них изолированы географически. В наши
дни американские диплоиды, конечно, изолированы от европей-
ских диплоидов. Возможно, однако, что в доледниковое или
422 Специализированные генетические системы
межледниковое время (или же как в то, так и в другое) эти
диплоидные виды имели более обширные ареалы и сильнее пе-
рекрывались друг с другом. Быть может, именно тогда и возник-
ла первая волна аллотетраплоидов и агамоспермных производ-
ных [314].
Эта межледниковая флора ежевик вынуждена была отступить
во время последнего оледенения, но в послеледниковое время с
улучшением климатических условий она, вероятно, вновь рас-
пространилась и дала новые поколения агамоспермных гибрид-
ных производных. В настоящее время агамоспермные ежевики
в отличие от своих диплоидных половых предков занимают об-
ширный ареал по всей северной и восточной Европе. Агамо-
спермные микровиды гибридного происхождения успешно засе-
лили холодные области этого материка, которые еще недавно
находились подо льдом [314, 315].
Taraxacum
Широко распространенный род Taraxacum встречается как в
Старом, так и в Новом Свете, как в Северном, так и в Южном
полушариях. Центр его распространения находится в Евразии,
где он на востоке доходит до Японии, а на севере проникает в
Заполярье. Многочисленные варианты сгруппированы в 32 сек-
ции. Основное число хромосом х=8. Известны формы 2х, Зх, 4х,
5х и 6х, а также несколько анеуплоидов.
У этого рода различают 28 половых диплоидных видов. Хо-
рошо известным примером служит Taraxacum kok-saghyz (2п=
= 16) из Средней Азии. В Австрии обнаружена диплоидная по-
ловая форма Т. officinale (224]. Большинство половых дипло-
идов самонесовместимы. Многие из них относятся к нескольким
примитивным секциям этого рода [681]. Некоторые — узко энде-
мичные или реликтовые формы. Так, Т. pieninicum— реликт
ледникового времени, встречающийся в горах в Польше, а
Т. cylleneum эндемичен для одной горы в Греции [225, 227, 545].
Искусственные гибриды между диплоидными пойовыми ви-
дами высокостерильны при одних комбинациях родительских
форм [224, 317, 547] и плодовиты при других [681].
Полиплоидные представители Taraxacum являются агамо-
спермами, в большинстве случаев облигатными, но иногда фа-
культативными. Агамоспермные таксоны заходят в своем рас-
пространении в арктическую область, а в других флорах неред-
ко растут как сорняки [148, 317, 681, 763]. Фюрнкранц [226] об-
наружил, что в Австрии полиплоиды обычно встречаются в сме-
шанных популяциях с диплоидами.
Гл 32 Агамный комплекс
423
Наиболее обычный уровень плоидности у этого рода — три-
плоидия, наблюдаемая в 114 из 218 таксонов, в которых произ-
водились подсчеты [681] Например, одуванчик лекарственный
{Taraxacum officinale}—это, как правило, агамоспермный три-
плоид (2« = 24). хотя у этого вида известна и диплоидная фор-
ма Часто встречаются тетраплоидные формы Taraxacum
(60 таксонов); известны также пентаплоидные формы (15 так-
сонов) и одна гсксаплондная форма [681].
Для полиплоидных агамоспермных форм характерен нару-
шенный мейоз с низким уровнем конъюгации. Некоторые из них
дают стерильную пыльц}, а другие ие образуют ее вовсе. При
ai амоспермном размножении материнская клетка зародышевого
мешка делится с образованием диады Нередуцированная хала-
зальная клетка этой диады образует зародышевый мешок. За-
тем из нередуцированной яйцеклетки партеногенетически разви-
вается зародыш [546, 681].
У агамоспермных Taraxacum имеется множество микровидов.
По данным Густафсона [317], в Скандинавии насчитывается
450 микровидов. Олгард [630] недавно описал 26 новых «видов»
(читай «микровидов») в одной только Дании. Согласно Ричард-
су [681], у этого рода в целом описано почти 2000 агамосперм-
ных микровидов. Микровиды распадаются на скопления, кото-
рые соответствуют примерно 26 таксономическим секциям [681].
Эти микровиды довольно однородны, хотя даже в отдельных
популяциях существует некоторая индивидуальная генетическая
изменчивость, как это было показано для Taraxacum officinale
[757]
Taraxacum произрастает в основном в умеренном поясе,
с центром распространения в центральных и западных районах
Евразии Ричардс [681] считает, что это — древний род, проис-
ходящий из западных частей Гималаев; оттуда наиболее про-
двинутые формы распространились в Европу, в арктические и
дручпе регионы Изменения климата в плейстоцене способство-
вали контактам между разными видами, гибридизации и обра-
зованию в этот период многочисленных агамоспермных микро-
видов.
Филогения этою рода сложна и не вполне понятна Ричардс
[681] полаюет, что при гибридизациях, в результате которых
возникли многие из нынешних агамоспермных микровидов,
одним из родительских видов были представители секций Сега-
tophora и Arclica Так, секция Bulgaria представляет собой ре-
зультат скрещивания CeratophoraX (q), а секция Alpina — ре-
зультат скрещивания ArcticaX Taraxacum ( = секция Vulgaria}.
Для того чтобы подтвердить это предположение, необходимо,
однако, провести более подробные исследования.
424
Часть восьмая. Специализированные генетические системы
Агамные комплексы типа Rubus Moriferus
Агамные комплексы Rubus, Crepis и Taraxacum принадлежат к
определенному типу, наблюдаемому также у ряда других групп
растений северной умеренной области. Этот достаточно часто
встречающийся тип удобно назвать типом Rubus Moriferus,
поскольку он наиболее подробно изучен на примере этой
группы.
Наиболее существенные черты агамных комплексов типа Ru-
bus Moriferus сводятся к следующим. Эти комплексы насчиты-
вают сотни или тысячи взаимосвязанных агамоспермных микро-
видов. Все эти микровиды гибридного происхождения. Они про-
дуцируют семена путем гаметофитного апомиксиса и нередко
также до некоторой степени — половым путем. Агамоспермные
формы являются полиплоидами, четными или нечетными, причем
нечетные более обычны. Полиплоидные агамоспермные формы
филогенетически и таксономически представляют собой над-
стройку над рядом ясно выраженнйх половых диплоидных ви-
дов, существующих в настоящее время. Половые диплоиды, од-
нако, в большинстве случаев имеют ограниченные или даже ре-
ликтовые ареалы, тогда как их агамоспермные производные бо-
лее агрессивны и имеют более широкое распространение.
Помимо комплексов Rubus, Crepis и Taraxacum к типу Rubus
Moriferus относятся некоторые другие агамные комплексы, пере-
численные ниже; обзоры по большей части этих комплексов
можно найти у Густафсона [317], а комплексы Cotoneaster,
Townsendia и Hieracium pilosella рассмотрены Саксом [710], Би-
меном [52] и Скалинска [735].
Hieracium подрод Pilosella (х=7) в Евразии
Hieracium подрод Hieracium (х=7) в Северном полуша-
рии
Alchemilla подрод Alchemilla (х=8) в Северном полу-
шарии
Alchemilla подрод Alchemilla (я=8) в Евразии, Новом
Свете и Австралии
Townsendia (х=9) в западных районах Северной Аме-
рики
Hypericum (х=8) в Евразии
Parthenium (х=9) в южных районах Северной Америки
Sorbus (х=17) в Евразии
Cotoneaster (х=17) в Евразии
Antennaria (х=7) в Северном полушарии
Catamagrostis (х=7) в Евразии
Все эти примеры, разумеется, различаются в деталях. Так,
подрод Hieracium содержит много диплоидных половых видов,
Гл 32 Агамный комплекс
425
Рис 32 3. Распространение Bouieloua curtipendula [262]
Три приведенные здесь разновидности содержат следующее число хромо-
сом Вс curtipendula— 2п=40, 41—64; В. с caespitosa— 2л=58—103;
В с tenuis — 2п = 20, 40—42.
а у Alchemilla они редки. У Sorbus и у Cotoneaster также хоро-
шо представлены половые диплоиды, но здесь это старые дип-
лоидизированные аллотетраплоиды (х2=17). У Antennaria
(х = 7), Parthenium (х = 9) и Calamagrostis (х=7) предковые
половые виды были, по-видимому, тетраплоидами
Интересно, что большинство перечисленных здесь агамных
комплексов типа Rubus Moriferus относится к сем. Compositae
и Rosaceae, очень успешно развивающимся в Северном полуша-
рии. К сложноцветным принадлежат Crepis, Taraxacum, Hiera-
cium, Townsendia, Parthenium и Antennaria; к розоцветным —
Rubus, Alchemilla, Sorbus и Cotoneaster Кроме того, среди при-
веденных примеров есть один представитель сем. Gramineae —
Calamagrostis и один представитель сем Hypericaceae — Hype-
ricum.
426
Часть восьмая. Специализированные генетические системы
Комплекс Bouteloua curtipendula
Грамовы травы, образующие комплекс Bouteloua curtipendula,
широко распространены в грасслендах Северной Америки—от
Канады до Мексики, а также встречаются в других разобщен-
ных областях Центральной и Южной Америки. Гулд и Кападиа
[262], произведя ревизию этой группы, выделили в ней 12 видов.
Один из них, собственно В. curtipendula, имеет очень обширный
ареал, пересекающий всю центральную часть США и Мексики
в виде широкого пояса (рис. 32.3). Остальные 11 видов имеют
более ограниченное распространение (рис. 32.4) [262].
Число хромосом у этих видов варьирует от 2п=20 до 2га=103
(табл. 32.1) при основном числе х=10. Среди них есть дипло-
иды, тетраплоиды, гексаплоиды и более высокие полиплоиды
вплоть до 10х, а также многочисленные анеуплоиды [261, 262].
Таблица 32.1. Число хромосом у разных членов комплекса
Bouteloua curtipendula (х=10) 1[260—262]
Вид Ареал Число 2л
В. triaena В. media Южная Мексика От центральной части Мексики до Гондураса От Юты до северных районов центральной части Мексики От западного Техаса до Нью- Мексико От Калифорнийского залива до Соноры Центральная Мексика От тропической Мексики до Перу Острова Карибекого бассейна То же Оахака Центральная Мексика Мексика От -южной части Канады до Техаса От юго-западной части США до южной части Мексики, а также в Южной Америке 20 20
В. uniflora В. warnockil В. reflexa 20 (30 у одного гибри- да) 21, 22, 23, 25, 28, 38, 40 20
В. pedicillata В. disticha Вероятно, диплоид (су- дя по величине пыль- цевых зерен) То же
В. vaneedenii В. juncea В. distans В. purpurea В. curtipendula var. tenuis nn. curtipendula var. caespitosa > » Не известно; возможно, днплоид То же 40 20, 40, 42 40, 41, 42, 43, 44, 45, 46, 48, 50, 51, 52, 53, 54, 55, 56, 58, 59, 60, 62, 64 58, 60, 62, 64, 69, 72, 77, 78, 80, 82, 84, 85. 86, 87, 88, 90, 91, 92, 94, 96, 98, 100, 102, ЮЗ
Гл 32 Агамный комплекс
427
Высокие полиплоиды и апсуплоиды группы Bouteloua curti-
pendula представляют собой агамоспермные формы, размножаю-
щиеся путем апоспории и псевдогамии; у них часто наблюда-
ются нарушенный мейоз и низкая степень конъюгации хромо-
сом. Половые гексаплоиды или половые анеуплоиды близкие к
гексаплоидному уровню, в этой группе не известны. Диплоиды,
тетраплоиды и анеуплоиды невысокого уровня размножаются,
насколько это известно, половым путем. Мейоз протекает нор-
мально у диплоидов и некоторых тетраплоидов, а у других те-
траплоидов и анеуплоидов с низким числом хромосом — с уме-
ренными нарушениями [220, 259, 346, 581].
Произведя подсчет хромосом в одной из выборок, Фрстср и
Браун [220] обнаружили, что граница между половым размно-
жением и агамоспермией проходит в этой iруппе па уровне
2д = 52 Все растения, у которых число хромосом выше 52, раз-
множаются при помощи агамоспермии, а те, у которых опо равно
52 или ниже, во всех известных случаях размножаются половым
путем. Возможно, однако, что в дальнейшем будут обнаружены
агамоспермные формы с низким числом хромосом [220, 259].
Диплоидные половые виды, такие, как Bouteloua uniflora,
В triaena, В. media и другие, встречаются главным образом от
субтропических до тропических широт и имеют относительно ог-
раниченные ареалы. То же самое относится к половым тетра-
плондам В purpurea и частично —к В warnockii (рис. 32.4).
Половые диплоидные и тетраплоидные формы В. curtipendula
tenuis также встречаются в Мексике (рис. 32.3).
В отличие от диплоидов представители этой группы с высо-
кими хромосомными числами, а именно Bouteloua curtipendula
curtipendula (2^ = 40—64) и В, с. caespitosa (2п = 58—103), име-
ют очень обширные ареалы, расположенные севернее п тяну-
щиеся через равнины до южных частей Канады (рис. 32 3) Как
отмечалось ранее, растения с более высокими числами хромо-
сом— в Д1 апазоне примерно 5х—10д — являются агамосперм-
ными. На карте, представленной на рис 32.5, можно видеть,
что эти агамоспермные формы В curtipendula занимают об-
ширную область на юго-западе США и на севере Мексики
[259].
Еще одна черта, характерная для видов Bouteloua с высоки-
ми хромосомными числами, — наличие у пих обширных апсупло-
идных рядов. В табл. 32.1 можно видеть почти непрерывные
анеуплоидные ряды лишь с несколькими очень небольшими раз-
рывами, как, например, у В curtipendula curtipendula (2п = от40
до 64) и В с caespitosa (2п = от 58 до 103) При подсчетах хро-
мосом в выборках эуплоидные числа встречаются чаще всего,
а различные анеуплоидные числа представляют собой отклоне-
ния от этих эуплоидпых узлов [261, 262]
28*
Рис. 32.4. Распространение 9 половых видов (диплоидных и тетраплоидных) комплекса Bouteloua curtipendula [262].
На карте показаны только южная часть ареала В. uniflora и северная часть ареала В. disticha.
Гл. 32. Агамный комплекс
429
Рис. 32 5 Распространение половых и агамоспермных форм Bouteloua cur-
tipendula [259].
Такой обширный анеуплоидный ряд возникает в результате
комбинации процессов естественной гибридизации, нарушенного
мейоза и агамоспсрмного размножения.
Естественная гибридизация между популяциями с разным
числом хромосом ведет к выщеплению анеуплоидных форм в
последующих поколениях. Вероятно, в этом — причина возникно-
вения простых анеуплоидных рядов, наблюдаемых у полового
вида Bouteloua warnockii (см. табл. 32.1). В. warnockii (2х, 4х)
гибридизирует в западной части Техаса с В. curtipendula caespi-
tosa — видом, характеризующимся высоким хромосомным чис-
лом {262]. Полагают, что наиболее обширный анеуплоидный ряд
в пределах самого вида В. curtipendula возник в результате бо-
лее сложных гибридизаций внутри этого таксономического вида,
а также между ним и различными другими видами данной груп-
пы [261, 426].
430
Часть восьмая. Специализированные генетические системы
Рис 32 6. Первая анафаза в делящейся материнской клетке пыльцы одного
растения Bouteloua curtipendula caespitosa с высоким числом хромосом;
обратите внимание на очень неравномерную сегрегацию хромосом '[262]
У растений вида Bouteloua curtipendula с высокими хромо-
сомными числами мейоз часто бывает нарушен, о чем уже гово-
рилось выше. Наиболее интересная для нас особенность мейоза
заключается в очень неравномерном расщеплении в анафазе I:
большая часть хромосом направляется к одному полюсу и лишь
несколько — к другому (рис. 32.6). Клетки, получившие боль-
шое число хромосом, по-видимому, дают жизнеспособные пыль-
цевые зерна, которые в свою очередь могут дать анеуплоидных
потомков. Различные анеуплоидные формы, после того как они
образовались, могут затем воспроизводиться при помощи агамо-
спермии {261, 262].
Подводя итоги, можно сказать, что группа Bouteloua curti-
pendula представляет собой половой полиплоидный комплекс на
тетраплоидном уровне, происходящий от В. uniflora, В. curtipen-
dula 2х и различных других диплоидных видов. От этого полово-
го полиплоидного комплекса возник агамный комплекс с диапа-
зоном плоидности от до 10х А эти эуплоидные агамосперм-
ные формы в свою очередь дали начало обширному и почти не-
прерывному ряду вторичных анеуплоидов.
Циркумбореальные группы Роа pratensis и Р. palustris —
агамные комплексы, сходные в общих чертах с группой Boute-
loua curtipendula. В группе Роа половые формы — низкие полип-
Гл 32. Агамный комплекс
431
лоиды или, что реже, они диплоидны (х = 7). Формы бо-
лее высокой плоидности являются агамоспермными или живо-
родящими Эуплоидные узлы соединены почти непрерыв-
ным рядом вторичных анеуплоидов. В гл. 31, посвященной
агамоспермии, уже говорилось о том, что у таксономического
вида Р. pratensis число хромосом варьирует от 2п=22 до-
гм =147.
У агамоснермного комплекса Potentilla gracilis (х = 7) на за-
паде Северной Америки также преобладает вторичная анеупло-
идия. Число хромосом у этого комплекса варьирует от 2п«56
до 2и«109 [100].
Тип агамоспермных комплексов, характерный для Bouteloua,
Роа и Potentilla, можно рассматривать как один из вариантов
более обычного типа Rubus Moriferus, у которого вторичная
анеуплоидия развивалась в широких масштабах.
Citrus
Род Citrus принадлежит к подтрибе Citrinae трибы Citreae под-
семейства Aurantioideae, сем. Rutaceae. Подтриба Citrinae —
группа из 13 родов вечнозеленых или — в редких случаях —
листопадных деревьев и кустарников, приносящих плоды с соч-
ной мякотью. Большинство представителей Citrinae — уроженцы
Юго-Восточной Азии Малайзии и Австралии; некоторые члены
группы встречаются в Африке [818].
Citrus принадлежит к группе из шести родов Citrinae. свя-
занных близким родством; все они приносят плоды типа апель-
синов или лимонов. Помимо самого Citrus в эту группу входят ро-
ды Fortunella, Eremocitrus, Poncirus, Clymenia и Microcitrus.
Представители всех этих родов могут скрещиваться между со-
бой. Они произрастают в Индии и Китае и по цепи островов в
Тихом океане доходят до Австралии [818].
Самый большой и наиболее широко распространенный род в
этой группе — Citrus с 16 видами и двумя подродами.
В табл. 32.2 представлена в сжатом виде система рода Citrus
по Свинглю [818]
К подроду Papeda относятся дикорастущие виды с мелкими
цветками и несъедобными плодами. Соковые мешочки мякоти
плодов у Papeda содержат капельки едкого масла. Для подрода
Citrus, напротив, характерны крупные душистые цветки и
съедобные плоды, соковые мешочки мякоти которых не содер-
жат едкого масла. С. indica и С. tachibana, по-видимому, дико-
растущие виды. Остальные виды подрода Citrus (виды 9—16 в
табл. 32.2) культивируются, и большинство из них, в частности
С. sinensis и С. paradisi, в дикорастущем состоянии неизвестны.
432
Часть восьмая. Специализированные генетические системы
Таблица 32.2 Система рода Citrus (по [818])
I. Подрод Papeda 1. С. ichangensis 2 С. latipes 3. С micrantha 4. С. celebica 5. С macroptera — ичанг папеда. Китай — кхаси папеда. Индия и Бирма — мелкоцветковая папеда Филиппины — Целебесская папеда. Сулавеси и Филиппины — меланезийская папеда Юго-Восточная Азия и Поли- незия
6 С. hystrlx II Подрод Citrus 7. С indicus — маврикийская папеда. Юго-Восточная Азия и Ма- лайзия — индийский дикий апельсин. Восточная часть Гима-
8. С. tachibana 9. С. medica лаев — тачибанский апельсин. Япония — цитрон. Восточная часть Азии; в настоящее время
10. С. И топ широко выращивается в культуре — лимон. Юго-Восточная Азия; широко выращивается
11. С. aurantifolia в культуре — лайм Юго-Восточная Азия и Малайзия; широко вы-
12. С auranlium ращивается в культуре
— померанец. Юго-Восточная Азия, широко выращи- вается в культуре
13. С. sinensis — сладкий апельсин. Китай н Индокитай; широко вы- ращивается в культуре
14 С. reticulata — мандарин Юго-Восточная Азия и Филиппины; широ- ко выращивается в культуре
15. С. grandis — помело, или шеддок Юго-Восточная Азия; широко выращивается в культуре
16 О parodist — грейпфрут. Острова Карибскопо моря; широко вы- ращивается в культуре
Вероятная первоначальная родина каждого из выращиваемых
видов указана в табл. 32.2.
Важно понять, что таксономия рода Citrus, и в особенности
его культурных форм, отнюдь не так проста, как это может
показаться из табл. 32.2. Признание культурных форм отдельны-
ми видами (виды 9—16) неизбежно делает эту классификацию
типологической. Тем не менее такая система полезна в качестве
предварительной схемы, и поэтому мы ее здесь приводим, одна-
ко она не дает точной картины характера изменчивости в этой
группе.
Для изменчивости, с которой столкнулись исследователи
культурных древесных форм Citrus с начала XIX в. и которая
озадачила их, характерно отсутствие четко выраженных видов.
Вместо этого в данной группе можно видеть ряд различных раз-
новидностей, многие из которых — гибриды, происходящие от
той или иной комбинации родительских форм и соединяющие
именованные виды в один обширный сингамеон. Именованные
виды 9—16 из табл. 32.2 служат удобными узловыми точками в
этом переплетении форм.
Гл. 32. Агамный комплекс
433
Ряд именованных видов, перечисленных в табл. 32.2, — не
единственная система, которую можно создать для Citrus
В сущности, уже на протяжении полутора веков в равной мере
компетентные исследователи плодовых деревьев рода Citrus
делятся на крайних «видообъединителсй» и крайних «видодроби-
телей»
Эти две противоположные тенденции иллюстрируются систе-
мами Танаки [822, 823] и Свингля [818] Танака давал видовые
названия каждой морфологической форме и каждому гибрид-
ному типу. В результате в роде Citrus в целом Танака [823] раз-
личает 157 видов, тогда как Свингль — только 16 [818]. В свою
очередь Свингль различает только наиболее ярко выраженные
разновидности и гибриды и рассматривает их как инфравидовые
единицы, относящиеся к различным таксономическим видам
[818]
При бо!лее детальном сопоставлении альтернативных систем
Свингля и Танаки можно обнаружить, что эти две системы дос-
таточно хорошо совпадают в отношении дикорастущих видов
подрода Papeda. Расхождения между системами наиболее вели-
ки в том, что касается культивируемых древесных форм Citrus
Знаменательно что именно в этой части рода таксономическая
путаница достигает максимума
Основное число хромосом у Citrinae, так же как и у других
Aurantioidae. обычно равно х = 9. Почти все виды Citrus и род-
ственных родов — диплоиды (2н=18). Тетраплоиды и трипло-
иды встречаются как отклоняющиеся типы у ряда культурных
древесных форм Citrus [151, 677].
Виды рода Citrus можно скрещивать искусственно, и они
спонтанно гибридизируют во многих комбинациях. Виды Citrus
скрещиваются также с представителями Poncirus, Fortunella,
Microcitrus и Eremocitrus. Многие межвидовые и даже некото-
рые межродовые гибриды плодовиты при половых скрещива-
ниях. Другие гибриды Citrus стерильны при половых скрещива-
ниях, но продуцируют семена бесполым путем [677, 818].
В смысле взаимоотношений по плодовитости группа Citrus при-
надлежит к модели Ceanothus.
Большинство культивируемых разновидностей Citrus пред-
ставляет собой факультативно агамоспермные или псевдогам-
ные формы. Наблюдаемый у них тип агамоспермии — это адвен-
тивная эмбриония с развитием зародышей из клеток нуцеллуса.
После опыления наряду с зародышами, развивающимися из зи-
готы, могут развиваться и нуцеллярные зародыши. В результа-
те семя содержит два или несколько зародышей — явление, из-
вестное под названием полиэмбрионии; зародыши могут быть
половыми и (или) бесполыми в различных соотношениях. Опы-
ление может приводить к расщеплению среди потомков или к
434 Часть восьмая. Специализированные генетические системы
образованию потомков только материнского типа в зависимости
от родительских форм [317, 677, 880].
В одной серии испытаний от лимона сорта Эврика после опы-
ления было получено 233 растения, из которых 33% были ага-
моспермными и 67%—половыми. При испытаниях, проведен-
ных на других цитрусовых, процент всех растений, возникших
путем нуцеллярной эмбрионии, составил: у апельсинов сорта
Навель Вашингтонский 97%; у апельсинов сорта Валенсия 85%;
у грейпфрута сорта Марш 96%; у американских лаймов 78% и
у мандаринов сорта Денси 100% |[677].
Культурный Poncirus trifoliata также яв1ляется факультатив-
но агамоспермным видом. Нуцеллярная эмбриония обнаружена
и у выращиваемых в культуре Fortuneila. Наряду с этим при
Citrus sinensis
Citrus reticulata
Citrus aurantium
Citrus grandis
Citrus tachibana
Citrus limon
Citrus celabica
Citrus hystnx
Poncirus
trifoliata
Citrus
aurantitolia
Citrus
ichangensis
Citrus
macroptera
Fortuneila
marganta
Fortuneila
apomca
Рис. 32.7. Спонтанные гибриды плодовых деревьев Citrus, возникшие в са-
дах на Дальнем Востоке (по [818]).
Родственные связи установлены на основе морфологических данных и поле-
вых исследований.
Гл. 32. Агамный комплекс
435
испытаниях по потомству одной линии дикорастущего вида Cit-
rus ichagensis и нескольких культиваров С. grandis до сих пор
не выявлено никаких признаков агамоспермии [677].
Для рассматриваемых нами проблем весьма важно, что не-
которые искусственные межвидовые гибриды Citrus размножа-
ются исключительно при помощи нуцслляркой эмбрионии. Так
обстоит дело у гибридов Fj от скрещивания С. paradisi и С. reti-
culata (танжело), а также С. sinensis и Poncirus trifoliata (цпт-
ранж) Эти гибриды размножаются в чистоте при помощи семян,
и иногда для получения больших насаждений их разводят семе-
нами [818].
Гибридные древесные формы Citrus обычны в приусадебных
садах и в деревенских рощах по всему Дальнему Востоку. Сре-
ди них есть самые разнообразные гибриды (рис 32 7). Некото-
рые из них встречаются лишь спорадически и локально, а дру-
гие широко распространились благодаря культивированию. Эти
последние, по-видимому, возникли спонтанно в смешанных на-
саждениях старых разновидностей Citrus, а затем их сохра-
няли, а иногда и разводили, если они обладали желаемыми при-
знаками [818].
На рис. 32.7, созданном по данным Свингля [818], показана
многочисленность спонтанных гибридных комбинаций, обнару-
женных в восточных районах Азии и Тихоокеанском регионе.
Заключения о родительских формах этих гибридов, возникших
в садах, делались в большинстве случаев на основе морфологи-
ческих данных и полевых наблюдений, а в нескольких случаях
подтверждены результатами искусственных скрещиваний, про-
водившихся в контролируемых условиях Спонтанное возникнове-
ние некоторых из этих гибридов, а также нескольких новых, как,
например, Eremocitrus glauca^Citrus sinensis, наблюдалось так-
же в цитрусовых насаждениях в Северной Америке [818].
Юзу — гибрид, возникший, вероятно, в результате скрещива-
ния между Citrus reticulata и С. ichangensis, он выращивается
повсеместно в Китае и Японии. Как свидетельствуют китайские
травники, юзу выращивали еще в древнем Китае начиная по
меньшей мере с 1108 г. п. э., а возможно, и с 237 г. до н. э. [818]
Юзут представляет собой высокоагамоспермное растение, до
95% потомков которого возникает из семян, развивающихся в
результате нуцеллярной эмбрионии [677]. Таким образом, он мо-
жет воспроизводить свою гибридную конституцию бесполым пу-
тем в ряде последовательных поколений.
Грейпфрут (Citrus paradisi) неизвестен в естественном или
пол у естественном состоянии на Дальнем Востоке, где находится
центр распространения рода Citrus, он возник, по-видимому, в
Вест-Индии. Судя по морфологическим признакам, грейпфрут,
возможно, представляет собой гибрид от скрещивания С. gran-
436 Часть восьмая. Специализированные генетические системы
dis и С. sinensis [818]. Если это правдоподобное, но недоказан-
ное предположение справедливо, то грейпфрут мог возникнуть
в результате спонтанной гибридизации между помело и сладким
апельсином, которые выращивались на этих островах в ранний
период заселения их европейцами. Как уже отмечалось выше,
грейпфрут — высокоагамоспермная форма и поэтому, очевидно,
воспроизводит свой гибридный генотип, размножаясь семенами.
Источники изменчивости групп Citrus многообразны. О гиб-
ридизации между ранее существовавшими видами, полувидами
или разновидностями уже говорилось. Гибриды при половом
размножении могут давать расщепляющихся потомков
(рис. 31.4). Часто возникают почковые мутации; у апельсинов
Навель Вашингтонский и других форм они привели к появлению
новых разновидностей. Большая часть культивируемых разно-
видностей Citrus дает некоторую определенную долю автотетра-
плоидных растений [677].
Новые варианты, независимо от их природы и происхожде-
ния, могут затем размножаться в чистоте при помощи семян,
образующихся в результате нуцеллярной эмбрионии, и тем са-
мым увеличивать общее число агамоспермных микровидов в
группе Citrus. Бессознательный отбор, издавна производившийся
человеком, а в более недавнее время — целенаправленный искус-
ственный отбор превратили некоторые из мелких микровидов в
широко распространенные разновидности и агамоспермные ви-
ды. В результате этих процессов дикорастущие виды Citrus с
несъедобными плодами дали начало разнообразным культурным
плодовым цитрусовым деревьям, которые теперь выращивают
во всех областях земного шара с субтропическим и теплым уме-
ренным климатом.
Citrus служит примером агамных комплексов третьего типа
У этого типа в отличие от типов Rubus Moriferus и Bouteloua
агамоспермные формы размножаются при помощи адвентивной
эмбрионии и остаются преимущественно на диплоидном уровне.
Pteris ,
Апогаметия широко распространена среди папоротников, она
встречается у Adiantum, Asplenium, Dryopteris, Polystichum,
Pteris и у других родов. Особенно часто она наблюдается у об-
ширного, распространенного по всему земному шару рода Pteris,
у которого 34% таксономических видов апогамные [871, 872].
У рода Pteris имеется несколько различных агамных комп-
лексов. Некоторые из них состоят из смеси апогамных и поло-
вых форм, к другим относятся преимущественно или исключи-
тельно облигатно агамоспермные папоротники. Наличие или
отсутствие гибридизирующих половых форм коррелирует, как и
Гл 32 Лгамный комплекс
437
следовало ожидать, с величиной и разнообразием агамного
комплекса [871].
Группа Pteris quadriaurita— очень большая и сложная груп-
па, содержащая как половые, так и апогамные формы. Две по-
ловые формы, Pteris quadriaurita sensu stricto (2n = 58) и
P. multiaurita (2n = 58), гибридизируют на Шри Ланке; кроме
того, эти виды или их гибриды гибридизируют также с третьим
членом этой группы — апогампым Р. confusa (2п = 58). Все это
переплетение гибридизаций выходит за пределы центральной
группы Р. quadriaurita, так что в него вовлекаются члены дру-
гих групп видов, относящихся к роду Pteris i[870, 871].
Группа Pteris quadriaurita, распространенная в восточной
Азии, состоит, насколько это известно, целиком из облигатных
апогамных форм. Большинство из них — «диплоиды» (2п = 58),
а некоторые — триплоиды (2п = 87) Они образуют ряд размно-
жающихся в чистоте микровидов, отличающихся один от друго-
го по второстепенным признакам строения спор и молодых
листьев. Эти морфологические вариации сгруппированы в один
обособленный таксономический вид. Простота таксономической
структуры агамного комплекса Р. biaurita связана с отсутствием
в этой группе полового размножения и естественной гибридиза-
ции или их редкостью [871]
Pteris служит примером агамных комплексов четвертого ти-
па, довольно широко распространенных среди Polypodiaceae.
Для этого типа характерно бесполое размножение при помо-
щи апогаметии, обычно облигатной. Апогамные папоротники мо-
гут быть диплоидными или полиплоидными, но число хромосом
у них всегда высокое. В их агамные комплексы часто входят
как половые, так и апогамные члены, а иногда они состоят иск-
лючительно из апогамных форм.
Стадии развития
Относительную стадию развития данного агамного комплекса
можно определить по степени его таксономической сложности,
широте географического распространения и наличию или отсутст-
вию диплоидных половых предковых форм. На основе этих четы-
рех критериев различают четыре главные стадии развития комп-
лекса. юность, раннюю зрелость, позднюю зрелость и старость.
Parthenium служит примером агамного комплекса, находя-
щегося на относительно юной стадии развития Наблюдаемые
у него морфологические вариации можно сгруппировать всего в
два таксономических вида — Р. argentatum и Р incanum Этот
комплекс встречается только в одной географической провин-
ции, занимающей область от Техаса до Аризоны и заходящей в
438 Часть восьмая. Специализированные генетические системы
прилежащие части Мексики. Половые предковые формы этого
комплекса — тетрапЛоидные формы Р. argentatum и Р. inca-
num — существуют в настоящее время, хотя и имеют несколько
ограниченное распространение [689—691].
Агамный комплекс, содержащий сотни или тысячи агамо-
спермных микровидов, распространенных по обширной геогра-
фической области, находится на стадии развития, которую мож-
но назвать зрелостью. Зрелые агамные комплексы можно под-
разделить на комплексы, у которых сохранились многие или
бблыная часть диплоидных предковых видов, и комплексы, у
которых многие или большинство диплоидных половых предко-
вых видов вымерло. Считается, что первые находятся на стадии
ранней зрелости, а вторые —на стадии поздней зрелости.
В предыдущих разделах этой главы мы рассмотрели несколь-
ко примеров агамных комплексов, находящихся на стадии ран-
ней зрелости. У родов Crepis, Rubus, Taraxacum, Hieracium и
Citrus достаточно хорошо представлены диплоидные половые
виды вместе с очень разнообразным рядом агамоспермных про-
изводных.
У агамного комплекса, находящегося На стадии поздней зре-
лости, число диплоидов и половых полиплоидов сокращается, и
они исчезают, но агамоспермная суперструктура достигает пол-
ного развития. Примеры этой стадии развития можно видеть у
Alchemilla, Potentilla и Роа.
Агамный комплекс подрода Aphanes рода Alchemilla содер-
жит один диплоидный половой вид A. microcarpa (2п=16) и
много агамоспермных полиплоидов. У подрода Alchemilla ро-
да Alchemilla половое размножение встречается очень редко, а
агамоспермные полиплоиды многочисленны [317]. В группах
Роа pratensis и Р. polustris половые диплоиды неизвестны; эти
группы состоят главным образом из агамоспермных и живоро-
дящих форм с полиплоидным и вторично анеуплоидным числом
хромосом [317]. Точно так же Potentilla gracilis (х=7) в запад-
ной части Северной Америки содержит по крайней мере один
половой полиплоид, Р. pectinisecta (6х), а возможно, и другие;
однако в этой группе не известно ни одного полового диплоида,
но встречается много агамоспермных полиплоидных форм [101].
Старый агамный комплекс обеднен не только половыми ди-
плоидами и половыми полиплоидами, но также и числом агамо-
спермных микровидов. В таксономическом аспекте он представ-
ляется нам изолированным агамоспермным видом, не имеющим
известных половых родичей.
Houttuynia cordata (Saururaceae) в Восточной Азии —един-
ственный вид данного рода. Он является высоким полиплоидом
(2п=94—98), размножающимся при помощи агамоспермии
и не имеющим известных половых родичей. Вполне возможно,
Гл. 32. Агамный комплекс
439
что он представляет собой остатки какого-то агамного комплек-
са, который в прошлом был более многочисленным и разнооб-
разным, а в настоящее время оказался крайне обедненным [36].
Эволюционный потенциал
В число агамных комплексов входят некоторые из наиболее про-
цветающих в настоящее время групп растений. Комплексы Ru-
bus, Hieracium, Роа и подобные им роды заселили обширные и
разнообразные континентальные области с неблагоприятными
климатическими условиями; они сумели осуществить это благо-
даря необычайно широкой изменчивости, воплощенной во мно-
гих десятках видов и сотнях или тысячах микровидов. Следует,
однако, отчетливо представлять себе, что агамные комплексы,
находящиеся на поздних стадиях развития, утратили большую
часть своего прежнего набора видов и микровидов, в значитель-
ной степени утратили способность порождать новые вариации,
а следовательно, почти полностью или полностью лишились спо-
собности успешно справляться с разного рода изменениями, ко-
торые могут возникнуть в среде в будущем.
Из этих противоположных аспектов агамных комплексов вы-
текают противоположные суждения об их эволюционном потен-
циале Дарлингтон и Стеббинс считают, что эволюционный по-
тенциал агамных комплексов сильно ограничен, тогда как Гус-
тафсон подчеркивает, сколь высокого эволюционного успеха они
могут достигнуть и достигают в благоприятных случаях.
В «Эволюции генетических систем» [141] Дарлингтон пишет:
«Таким образом, мы видим, что апомиксис спасает то, что можно спасти
в тех случаях, когда плодовитость при половом разм.-.ожении оказывается
утраченной Половое размножение благодаря сопровождающей его
рекомбинации обеспечивает всем последующим поколениям основу для
адаптации. Апомиксис обеспечивает в этом смысле лишь поколение непосред-
ственных .ютомков. Он сохраняет у некоторых видов кое какие остатки по
ловоч системы, что дает им возможность отсрочить вымирание и даже при-
водит к кратковременному расцвету новых форм Однако с утратой половой
рекомбинации для апомикта так же как и для перманентного гибрида, путь
к длительном} существова.-ию отрезан Апомиксис дает возможность избе-
жать стерильности, но при этом он заводит в эволюционный тупик».
Итак, по мнению Дарлингтона, агамоспермия — это «эволю-
ционный тупик», Стеббинс же считает [780], что это «эволюцион-
ный оппортунизм, доведенный до предела». Кроме того (Бэбкок
и Стеббинс [36]):
«Конечную судьбу агам-ого комглсчса половые преткп которого оказа-
лись ограниченными в своем распространении или вымерли, можно предска-
зать до тех пор пока преобладают хслочия, существовавшие в период его
формирования, он будет процветать, ио он окажется неспособным справ-
440
Часть восьмая. Специализированные генетические системы
литься с какими-либо новыми изменениями среды! и поэтому с течением вре-
мени будет все больше и больше сокращаться, пока в конце концов не ис-
чезнет».
Но, как считает Густафсон [317], мнение о том, что агамо-
спермия кладет конец изменчивости, справедливо лишь отчасти.
А если агамный комплекс обречен на истощение изменчивости
и вымирание, то «не типично ли это для всех видов и родов?
Подобно отдельным индивидуумам, виды и роды рождаются,
достигают расцвета и умирают» [317].
Рассмотрим эти противоположные точки зрения, связав их с
основной биологической функцией полового размножения. Пол—
это генетическая система, обеспечивающая рекомбинацию генов,
которая в свою очередь служит главным источником наследст-
венной изменчивости в популяции (см. гл. 1). По|лный отказ от
полового процесса и переход к облигатному апомиксису лишает
данную популяционную систему ее главного источника новых
вариаций. А промежуточные формы бесполого размножения,
как при факультативном апомиксисе, ограничивают возникнове-
ние изменчивости до некоторого промежуточного объема.
В сущности, механизм полового размножения представляет
собой главный источник новых вариаций как в группе, размно-
жающейся исключительно половым путем, так и в агамном
комплексе. Агамный комплекс, состоящий, подобно Rubus Мой-
ferus, из факультативных агамоспермных форм и половых ди-
плоидов, может обладать высоким уровнем изменчивости и
большим эволюционным потенциалом. Однако не все агамные
комплексы обладают этими чертами. У такого изолированного
комплекса облигатно апогамных папоротников, как Pteris biau-
г На, наблюдается относительно стереотипная картина изменчи-
вости [871]. У комплекса Crepis occidentalis, который в областях,
занимаемых половыми популяциями, и вблизи от этих областей
содержит, помимо факультативно агамоспермных форм и поло-
вых диплоидов, облигатных агамоспермов, изменчивость очень
велика; однако в тех' областях, где обитают исключительно об-
лигатные агамоспермы, она сводится к одному или нескольким
типам [36].
Судя по данным фитогеографии, некоторые агамоспермные
типы остаются неизменными на протяжении длительных перио-
дов времени. Taraxacum reichenbachii и Т. dovrense — два почти
идентичных агамоспермных микровида, встречающихся как ре-
ликты ледникового времени в Альпах и Норвегии соответствен-
но. Со времени последнего межледниковья они дивергировали
очень незначительно. Древние агамоспермные микровиды обна-
ружены также у Hieracium и Alchemilla. Возраст некоторых
агамоспермных форм, судя по данным фитогеографии, должен
достигать по крайней мере 75 или 100 тыс. лет [37].
Гл. 32. Агамный комплекс
441
Как уже говорилось, многие „агамоспермные растения явля-
ются полиплоидами. В таких случаях к ограниченности реком-
бинаций, обусловленной агамоспермией как таковой, следует
добавить ограничения, налагаемые полиплоидностью.
Сохранение данной группой растений способности к полово-
му размножению не гарантирует ей длительного эволюционного
успеха. Как верно отмечает Густафсон, группы видов, размно-
жающиеся половым путем, приходят в упадок и гибнут подобно
апомиктическим группам, тогда как частично бесполые группы
могут успешно развиваться в течение долгого времени. Кроме
того, полная утрата данной группой способности к половому
размножению не обязательно означает, что она погибнет в бли-
жайшем будущем. Между тем переход к облигатному апомикси-
су, очевидно, влечет за собой в конечном счете вымирание.
Приведем в заключение точку зрения Стеббинса [780] по
этому вопросу:
«Нет никаких данных о том, что апомикты когда-либо были способны
создать в процессе эволюции новый род или хотя бы подрод. В этом смысле
вес агамные комплексы — закрытые системы и эволюционные «тупики»...
И если половые виды могут в период своего существования давать начало
совершенно новым типам при помощи мутационного процесса и рекомбина-
ции генов, то агамным комплексам суждено создавать только новые вариа-
ции на старую тему».
29-45
33 Вегетативное размножение
гибридов
Среди многолетних растений широко распространены разнооб-
разные способы вегетативного размножения. Многим или, воз-
можно, большинству групп многолетних растений, способных к
естественной гибридизации, присущ тот или иной способ веге-
тативного размножения.
Вегетативное размножение — свойство, выгодное для гибри-
дизирующей группы растений. Оно обеспечивае'Д сохранение и
размножение гибридного генотипа, обладающего адаптивной
ценностью. Данный гибрид может быть плодовитым, и в этом
случае его ценная в адаптивном отношении комбинация генов
будет утрачена его половыми потомками, или же он может быть
стерильным, и в таком случае вегетативное размножение —
главная надежда на выживание.
Гибрид, размножающийся вегетативным путем, если он хо-
рошо адаптирован, может увеличивать свою численность и рас-
ширять занимаемую им область, проходя через последователь-
ные стадии гибридного клона и клонального микровида. Ника-
ких объективных различий между этими стадиями не сущест-
вует, они различаются только по относительным размерам попу-
ляций. В этой главе будут приведены примеры каждой из этих
стадий.
В тех случаях, когда в разных линиях данной группы расте-
ний независимо проходит образование гибридов, так что от трех
или большего числа исходных родительских видов возникают
два или большее число вегетативно размножающихся гибрид-
ных видов, эта группа развивается как клональный комплекс.
Гибридные комплексы такого типа менее известны, чем агамные
и полиплоидные комплексы. Тем не менее в этой главе в даль-
нейшем можно будет привести несколько примеров таких комп-
лексов.
Гибридные клоны
Многие культурные разновидности цветковых растений про-
изводят стерильные семена, и поэтому их размножают вегета-
тивным путем. Хорошо известными примерами служат десерт-
ный банан, картофель и батат — среди плодов и овощей, а сре-
ди декоративных растений — некоторые розы и тигровые лилии.
Так, большая часть выращиваемых съедобных бананов — бессе-
мянные триплоидные (2п=3х=33) разновидности Musa acumi-
nata, а также гибриды М. acuminata и М. balbisiana, которые
разводят корневищами и луковицами [727, 728].
Гл. 33. Вегетативное размножение гибридов
443
Гибриды, размножающиеся вегетативным способом, обнару-
жены во многих группах дикорастущих растений, хотя у них это
явление встречается несколько реже, чем у культурных расте-
ний.
Elymus triticoid.es и Е. condensatus (Gramineae) — хорошо
дифференцированные виды дикорастущего многолетнего злака,
распространенного в Калифорнии. Искусственный гибрид меж-
ду этими видами жизнеспособен, но высокостерилен. В некото-
рых местах встречаются также естественные гибриды между ни-
ми; гибриды эти также высокостерильпы, но благодаря образо-
ванию корневищ они смогли распространиться локально. Неко-
торые из таких клонов морфологически сходны с искусственно
полученными Fi и представляют собой естественные Fb однако
другие, судя по их морфологическим признакам, — выщепенцы
из более поздних поколений, возникших из тех редких семян,
которые образуются у стерильных индивидуумов Fi 1171, 786]
Holcus mollis в Польше — другое высокостерильное травяни-
стое растение (возможно, гибрид), размножающееся корневи-
щами [877]. Естественные гибриды IrisfulvayJ. hexagona gigan-
ticaerulea в дельте Миссисипи размножаются вегетативно при
помощи корневищ и образуют локальные клоны [686, 861].
Интересный пример гибридных клонов, часть которых, веро-
ятно, возникла очень давно, обнаружен у Quercus в Юте и
Аризоне Гибриды между Q gambelii и Q. turbinella встречают-
ся в местах, разбросанных по северной части Аризоны, южной
части Юты и северной ее части, вблизи Соленого озера. Эти рас-
тения пе образуют желудей. Они образуют большие гибридные
колонии шириной от 25 до 60 м. Кроме того, эти колонии мор-
фологически однородны, что дает основание считать их клопа-
ми, образованными при помощи подземных побегов. Такое пред-
ставление подтверждается результатами эксперимента с рас-
пространением радиоактивного индикатора в одной из этих ко-
лоний. Клоны эти достаточно обширны, кроме того, они возник-
ли давно, по крайней мере некоторые из них [125]
Quercus gambelii — дерево, распространенное в южной части
Скалистых гор, тогда как Q turbinella — кустарник, растущий в
юго-западных районах США. Гибридные клопы на севере Ари-
зоны и на юге Юты находятся в пределах зоны симпатричсскою
перекрывания ареалов этих двух родительских видов, однако
местность на севере Юты, где они также встречаются, лежит
более чем на 400 км севернее современной северной границы
Q turbinella-. следовательно, в прошлом юго-западный вид
Q turbinella заходил дальше на север. По всей вероятности,
Q- turbinella доходил до северных районов Юты в период между
7500 и 4000 гг. до н. э., когда климат был теплее и суше, чем
впоследствии Если эта хронология верна, то возраст северных
29*
444 Часть восьмая. Специализированные генетические системы
гибридных клонов равен 4000 лет, а может быть, и больше
£1251.
Осины группы Populus tretnuloid.es размножаются в природе
вегетативно — корневыми побегами, отходящими от корневой
шейки, и образуют хорошо известные клоны; эту свою способ-
ность они передали и естественным гибридам. Гибрид между
Populus tremuloides и Р. grandidentata ( = Р.Xbarnesii) зани-
мает обширную территорию от Миннесоты до Новой Англии и
часто образует большие клоны [45, 865]. Гибридные клоны с та-
кими же признаками встречаются также в северной части Айда-
хо. Р. grandidentata в настоящее время в данной области не
встречается, но этот вид или его близкий родич был, вероятно,
распространен здесь прежде [44].
Клональные микровиды
Гибридизация между Iris fulva и I. hexagona giganiicaerulea1
в районе дельты Миссисипи дает не только гибридов Fi, но так-
же ряд выщепенцев из последующих поколений и продукты воз-
вратных скрещиваний. Все они часто образуют Илоны. Некото-
рые из них достигли уровня клональных микровидов, занимаю-
щих различные микроместообитания в районе дельты (15, 861].
Вариации среди популяций Iris настолько резко выделяются,
что многие из них были описаны прежде в качестве новых так-
сономических видов. Смолл и Александер [738] различали в
группе Iris fulva—hexagona в этой области 84 вида. Как пока-
зал Виоска [861], все они, за исключением трех, представляют
собой различные продукты гибридизации между I. fulva и I. he-
xagona giganiicaerulea. Число получивших названия таксономи-
ческих видов — хороший показатель разнообразия клональных
микровидов в этой группе.
Межклональная изменчивость хорошо выражена также в
группе Populus tremuloides—grandidentata. В этом случае она
привела к выделению нескольких новых именованных видов
(например, Р. аигеа), а также многочисленных разновидностей
и форм Р. tremuloides и Р. grandidentata (45].
Solanum chacoense подвид chacoense — дикорастущий клуб-
необразующий представитель секции Tuberarium рода Solanum,
распространенный на юге Южной Америки. Эта группа очень
изменчива, и многие ее члены были описаны в литературе в ран-
ге видов, подвидов или форм с соответствующими латинскими
названиями. Хоке и Иертинг [354] свели 32 таких названия в
1 Iris fulva и I. giganiicaerulea обычно рассматриваются как два само-
стоятельных вида. — Прим. ред.
Гл. 33. Вегетативное размножение гибридов
445
синонимику подвида S. chacoense chacoense\ в этой группе они
различают также 13 микровидов.
Solanum chacoense часто гибридизирует с S. microdontum и
реже — с некоторыми другими видами. В результате этой гиб-
ридизации у S. chacoense создается высокий уровень изменчиво-
сти, причем некоторые вариации становятся клональными мик-
ровидами [354].
Известно, что кустарниковые кактусы Opuntla fulgida и
О. spinosior гибридизируют в двух местностях в Аризоне с об-
разованием высоко, но не полностью стерильных гибридов.
У этих растений имеет место вегетативное размножение при по-
мощи отчленяющихся частей стебля, которые укореняются и
дают побеги. В начальной стадии гибридизации образуются гиб-
ридные клоны. Затем в результате ограниченного полового раз-
множения гибридов в более поздних поколениях появляются
различные выщепенцы, которые также размножаются вегета-
тивно и образуют клональные микровиды. Индивидуумы, состав-
ляющие эти микровиды, морфологически очень однородны [302].
Микровиды, как правило, характеризуются небольшими раз-
мерами и локальным распространением, но один обширный мик-
ровид гибридного происхождения \микровид W) занимает уча-
сток пустыни длиной 32 и шириной около 42 км. Гибриды от
скрещивания между Opuntla fulgida и О. spinosior образуют от-
личительный элемент флоры Аризоны; поэтому они были выде-
лены в таксономический вид О. kelvinensis [302].
В Техасе Opuntla lindheimeri гибридизирует с О. phaeacantha
major и О. edwardsii. Гибриды Fi и выщепенцы из последующих
поколений размножаются вегетативно при помощи отчленяю-
щихся частей стебля, как это свойственно опунциям вообще.
Гибридные клоны весьма обычны. Для изменчивости в этой
группе характерно также наличие клональных микровидов со
своими отличительными комбинациями признаков и микрогеог-
рафическими областями распространения. У О. lindheimeri име-
ются десятки или сотни таких микровидов. Как и в других рас-
смотренных выше примерах, систематики прежних лет описыва-
ли многие из этих микровидов как новые таксономические виды
[304, 305].
Клональные комплексы, размножающиеся
ответвлениями стебля
Очень немногие клональные комплексы проанализированы фи-
логенетически. В главе по автополиплоидии был описан кло-
нальный комплекс в группе Solanum tuberosum. Это, однако, не
просто клональный комплекс; данная группа характеризуется
также сочетанием некоторой степени межрасовой автополипло-
446 Часть восьмая. Специализированные генетические системы
идии и вегетативного размножения. Кроме того, исходные роди-
тельские формы связаны очень близким родством.
Клональный комплекс существует также у кустарниковых
кактусов подрода Cylindropuntia рода Opuntia, распространен-
ных на юго-западе США. У этих растений происходит половое
размножение семенами и вегетативное размножение отчленяю-
щимися частями стебля. Соотношение полового и вегетативного
размножения различно у разных видов. Половое размножение
преобладает у О. spinosior, О. versicolor, О. acanthocarpa и
О. echinocarpa, а вегетативное размножение— у О. fulgida и
О. bigelovii ({302, 303], а также неопубликованные данные).
Пинкава и его сотрудники [641—643] показали, что все эти виды
являются диплоидами с числом хромосом 2п=22.
В пустынях Аризоны и Калифорнии эти виды гибридизируют
в природе в различных комбинациях и образующиеся гибриды
размножаются вегетативным путем. Структура возникающего
при этом клонального комплекса изображена на рис. 33.1. Гиб-
рид Opuntia spinosior и О. fulgida дал начало О. kelvinensis со
стерильными семенами и вегетативным размножением, о чем
уже говорилось в предыдущем разделе. Однако в результате
гибридизации между О. spinosior и О. versicolor образуются
скопления гибридов, размножающихся половым путем [302, 303].
Opuntia bigelovii — очень близкий родич О. fulgida, — воз-
можно, гибридизирует с О. acanthocarpa и О. echinocarpa на юге
Калифорнии. Бенсон [58] сообщает о наличии в этой области
двух различных таксонов, промежуточных по морфологическим
признакам между О. bigelovii и либо О. acanthocarpa, либо
О. echinocarpa- эти таксоны продуцируют стерильные семена и
размножаются вегетативно. Вполне возможно, что они представ-
ляют собой гибриды, происходящие от родительских форм, ука-
занных на рис. 33.1. Такие предполагаемые гибридные формы
требуют, по мнению Бенсона [58], дальнейшего изучения.
Группа Potentilla anserina (х=7) содержит таксоны тетра-
плоидного и гексаплоидного уровней. Тетраплоидные популяции
этой группы размножаются половым путем и распадаются по
крайней мере на два интерстерильных вида. Но у гексаплоид-
ных форм мейоз нарушен, и они"образуют стерильную пыльцу и
стерильные семена. Эти гексаплоиды усиленно размножаются
ползучими побегами. При помощи такого способа размножения
они образовали ряд клональных микровидов, происходящих от
различных тетраплоидных популяций и соответственно родствен-
ных последним [699].
Группа Stellaria longipes (Caryophyllaceae), к которой отно-
сится несколько именованных видов, широко распространена по
всему Северному полушарию. Собственно Stellaria longipes
(2п=52) образует фертильные семена. Но S. crassipes — пред-
Гл. 33. Вегетативное размножение гибридов
44Z
ПРЕДПОЛАГАЕМЫЕ
Рис 33.1. Филогения клонального комплекса у кустарниковых кактусов
Opuntia (подрод Cytindropuntia).
ставитель этой группы, произрастающий в Арктике и содержа-
щий большое число хромосом (2я= 104), — образует высоко-
стерильные семена и размножается при помощи ползучих под-
земных побегов. Высокая плоидность представителей вида
S. crassipes позволяет предполагать, что он произошел от како-
го-то низ коллоидного вида данной группы и, возможно, пред-
ставляет собой амфиплоид. Современное широкое его распрост-
ранение в арктической области наводит на мысль, что этот вид
возник довольно давно и распространился при помощи вегета-
тивного размножения [317, 388].
448
Часть восьмая. Специализированные генетические системы
Клональные комплексы, размножающиеся
при помощи ложного живорождения
Большинство видов Allium размножаются половым путем и
производят фертильные семена. Однако у этого большого рода
широко распространено живорождение или, вернее, ложное жи-
ворождение [639]. Разные виды Allium размножаются при по-
мощи луковичек, образующихся в соцветиях вместо семян. Сте-
пень ложного живорождения варьирует в разных случаях от
частичной до полной, и соответственно варьирует редукция об-
разования семян, вплоть до полного их отсутствия у таких ви-
дов, как A. sativum и A. paradoxum. Мы рассмотрим здесь вкрат-
це ситуацию в двух группах видов — в группах A. paniculatum
и A. canadense.
Группа Allium paniculatum (х=8) в Евразии содержит виды
A. paniculatum, A. pulchellum, A. flavum, A. oleraceum и A. cari-
natum. Из них A. paniculatum, A. pulchellum и A. flavum — ди-
плоидные половые виды (2п=16). Они распространены на юге
Европы [466]. Ложноживородящие виды — A. carinatum и
A. oleraceum — встречаются в северной части Европы, и их
плоидность варьирует от 2х до 5х. Ложноживородящий вид
A. carinatum, произрастающий в Альпах, диплоиден, а произ-
растающий на севере Европы — триплоиден. Он морфологиче-
ски близок к половым видам A. pulchellum и A. flavum. Ложно-
живородящий вид A. oleraceum с тетраплоидным или пентапло-
идным числом хромосом (2/1=32, 40) морфологически близок к
половому виду A. paniculatum [151, 317, 466].
В восточных областях Северной Америки распространен дру-
гой клональный комплекс — группа Allium canadense (х=7),—
содержащий несколько половых форм (2х и 4х) и несколько
ложноживородящих производных (Зх и 4х). Родственный комп-
лекс A. geyeri (х=7), распространенный в западных областях
Северной Америки, также содержит несколько половых дипло-
идов и половых тетраплоидов, а кроме того, — ряд живородящих
полиплоидных производных (4х, 5х, 6х) [632].
Группа Saxifraga stellaris — другой большой комплекс, рас-
пространенный в Северном полушарии. S. stellaris sensu stricto
(2п=28) и S. ferruginea — половые виды, произрастающие в зо-
не с умеренно холодным климатом. S. foliolosa (2п=56, «64)—
ложноживородящий вид. Это бесполое полиплоидное производ-
ное имеет циркумполярный ареал, который расположен север-
нее, чем ареалы его половых родичей, и заходит в арктическую
область [317].
Густафсон [317] приводит несколько примеров ложноживоро-
дящих видов, которые обладают высокой плоидностью и рас-
пространены в северных или арктических областях, и близкие по-
Гл 33 Вегетативное размножение гибридов
449
ловые родичи которых неизвестны. К числу таких таксономиче-
ски изолированных ложпоживородящих видов принадлежат.
Polygonum viviparum (2rz = 83—ПО), Cardamine butbijera
(2n^96) и Saxifraga cernua (2rt«66)
Ложное живорождение обнаружено у нескольких родов зла-
ков, в частности, у Agrostis, Deschampsia, Eleucine, Festuca и
Poa В одной из предыдущих глав уже говорилось о том, чю
агамоспермия часто встречается также у Gramineac. У Роа в
некоторых группах видов одновременно наблюдается и а<амо-
спермия, и ложное живорождение.
Циркумбореальная группа Роа alpina (х=7) содержит поло-
вые, агамоспермные и ложноживородящие формы. Формы всех
этих трех типов существуют у самого таксономического вида
Роа alpina, у которого имеются диплоидные половые популяции
(2п=14), полиплоиды вплоть до гексаплоидного уровня и раз-
личные происходящие от них ложноживородящие и агамосперм-
ные популяции. Р. bulbosa, принадлежащий к этой группе, пред-
ставлен на юге Европы половыми формами с низкой плоид-
ностью, а на севере Европы — производными Ложноживородящи-
ми формами с тетраплоидным или гсксаплоидным числом хро-
мосом [317].
Группа Deschampsia caespitosa (х = 7) представляет собой еще
один обширный клональный комплекс, распространенный по все-
му Северному полушарию и доходящий в некоторых местах до
Южного полушария Половые тетраплоидные виды или полуви-
ды— D caespitosa, D. arctica, D. boitnica и другие — широко
распространены и связаны между собой гибридизацией. Места-
ми среди половых популяций встречаются ложиоживородящие.
Близкий к ним вид D alpina. обитающий в арктической зоне,
содержит несколько высоких полиплоидов и вторичных апеуп-
лоидов (2п = 39, 41, 49 56), и все его популяции почти исключи-
тельно ложноживородящие [317]
Эволюционный потенциал
Комбинация полового и вегетативного размножения выгодна
естественным гибридам и их потомкам. Она создает возмож-
ность дня возникновения целого ряда новых типов и сохранения
тех из них, которые обладают ценными адаптивными признака-
ми. Комбинация этих двух способов размножения характеона
в числе прочих для гибридов Iris fulvaXl hexagona giganticae-
rulea Populus tremuloidesXP grandidentata, Solanum chacoen-
sexS microdontum и Opuntia fulgidaxO. spinosior
Гибриды или потомки гибридов, обладающие адаптивной
ценностью, развиваясь, превращаются в мональные микрсвиды
и распространяются по всему местообитанию, к которому они
450
Часть восьмая. Специализированные генетические системы.
адаптированы. Ряд стадий — от мелких локальных до больших
широко распространенных микровидов — обнаружен в двух
группах видов опунции, изученных в этом плане [302, 305].
Если подобный процесс повторяется в различных линиях од-
ной и той же группы видов, то возникает клональный комплекс,
и нам известно несколько примеров таких комплексов. Но это
не «чистые» клональные комплексы. В большинстве случаев ве-
гетативное размножение сочетается у них с другими генетиче-
скими системами: с межрасовой автополиплоидией — в группе
Solatium tuberosum, с половым размножением — в группе Opun-
tia fulgida — spinosior, с амфиплоидией —в группе Potentilla
anserina и с агамоспермией — в группе Роа alpina.
У агамоспермных групп растений и у групп с вегетативным
размножением существует некоторое равновесие между половым
и бесполым размножением. Уместно сравнить эти две системы.
Преимущество вегетативного размножения состоит в том, что
оно обычно легко доступно гибридным растениям во многих
группах. В отличие от этого агамоспермия — высокоспециализи-
рованная генетическая система, которая обычно должна возник-
нуть de novo у гибридов или гибридных производных.
Вместе с тем семена в общем обеспечивают более эффектив-
ное распространение, чем вегетативные органы. Бесполые фор-
мы данного агамного комплекса используют для распростране-
ния адаптации, присущие семенам и плоидам, и благодаря этому
их микровидам удается во многих случаях заселить области
средних или больших размеров. Потенциальные возможности
сильно варьируют: так, воздушные луковички, или детки, могут
обеспечить распространение почти на такие же далекие расстоя-
ния, как желуди, а ползучие побеги пригодны для расселения
лишь на небольшие расстояния. Таким образом, следует ожи-
дать, что некоторые клональные микровиды, обладающие эффек-
тивными способами расселения, достигнут широкого распростра-
нения; однако существуют и многие другие клональные комп-
лексы, состоящие из растений, лишенных возможности распро-
страняться на далекие расстояния при помощи вегетативных
органов расселения. У таких комплексов варианты, возникшие
в результате гибридизации, могут никогда не выйти за преде-
лы той локальной местности, в которой они возникли.
Другим фактором, способным ограничить развитие многих
клональных комплексов, служит старение клонов. Старение кло-
нов обсуждалось с разных точек зрения Молишем [582], Брин-
ком [70] и Камероном и Фростом [80]. Это явление все еще оста-
ется не вполне понятным. Несомненно, многие виды растений
могут на протяжении сотен и тысяч лет размножаться вегета-
тивно без заметного понижения жизнеспособности. Несколько
примеров этого было приведено в гл. 1, и к ним можно доба-
Гл 33 Вегетативное размножение гибридов
451
вить много других. Но так бывает не всегда [582]. Некоторые
группы растений (например, Citrus, картофель, сахарный трост-
ник) после длительного вегетативного размножения проявляют
симптомы старения или дегенерации [80, 562]. Наблюдаемое
старение клонов, возможно, обусловлено главным образом на-
коплением в цитоплазме клеток растений патогенных вирусов
[80, 562. 625]. Возможно также, что одним из факторов такого
старения служат регулярные направленные изменения генов в
соматических клетках—некая форма парамутирования.
У Citrus агамоспермные растения, развившиеся из семян,
бывают более мощными, чем растения, выращенные из отвод-
ков. Образовавшиеся бесполым путем семена, по-видимому, сво-
бодны от вирусов, которыми заражены ткани стебля того же
самого растения [80, 625]. Это наблюдение представляет боль-
шой интерес для вопроса, который мы сейчас обсуждаем. Оно
позволяет предполагать, что агамные комплексы, избегая старе-
ния клонов благодаря размножению семенами, часто способны
развиваться дольше во времени и шире в пространстве, чем
клональные комплексы.
Клональные комплексы имеют, в общем, более простую так-
сономическую структуру, чем агамные комплексы. Они обычно
содержат меньше микровидов Число микровидов в данном гиб-
ридном комплексе, находящемся в стадии зрелости, может слу-
жить мерой его эволюционного потенциала. Более низкий эво-
люционный потенциал клональных комплексов по сравнению с
эволюционным потенциалом агамных комплексов может быть
частично обусловлен такими факторами, как старение клонов,
а нередко также пониженной способностью к распространению
органов вегетативного размножения
В предшествующих плавах было показано, что эволюционный
потенциал агамных комплексов, хотя он, несомненно, велик,
ограничен по сравнению с эволюционным потенциалом групп,
размножающихся половым путем. Теперь мы видим, что эволю-
ционный потенциал клональных комплексов еще более ограни-
чен.
34 Естественные триплоиды
В очень многих группах растений встречаются спонтанные три-
плоидные индивидуумы и спонтанные триплоидные клоны, при-
чем как среди дикорастущих (например, Зх-формы Populus tre-
inula и Claytonia virginica), так и среди культурных видов
(Зх-формы Pyrus communis и Juniperus virginiana). Как и сле-
довало ожидать, спонтанные триплоиды редки среди дикорас-
тущих, но не слишком редки среди культурных растений. Неко-
торые примеры приведены в табл. 34.1.
Нас здесь интересуют два вопроса относительно спонтанных
триплоидных растений: как они возникают и как они воспроиз-
водятся? В этой главе мы попытаемся рассмотреть эти вопросы
на основе данных, имеющихся как по дикорастущим, так и по
культурным видам.
В природе пентаплоидные и гептаплоидные растения встреча-
ются редко. Естественные пентаплоидные формы обнаружены у
Lemna minor и L. gibba (2n=50) (851] и у Holcus mollis
(2л=35) [877], а также в группах Rosa canina и Solanum tube-
rosum, рассматривавшихся выше. Естественные гептаплонды об-
наружены у Lemna gibba (2n=70) [851] и у гибридов Opuntia
ficus-indica (536].
Проблемы образования и сохранения этих форм с более вы-
соким нечетным уровнем плоидности мало чем отличаются от
тех же проблем у триплоидов, поэтому эти формы рассматрива-
ются здесь вместе.
Проблема сохранения решается у триплоидов и у других
нечетных полиплоидов несколькими различными способами.
В этом участвуют несколько генетических систем, каждая из
которых рассматривалась в одной из глав части седьмой.
Возникновение
Существует два основных способа возникновения триплоидных
растений. Один из них — гибридизация между тетраплоидом и
диплоидом. Другой — отсутствие редукции у одной из диплоид-
ных родительских форм: при этом обычно происходит образо-
вание нередуцированных яйцеклеток и последующее их слияние
с нормальными редуцированными пы!льцевыми зернами.
Как показали эксперименты, проведенные с 121 диплоидной
линией Zea mays, в 18 из этих линий нередуцированные (2л)
яйцеклетки возникают спонтанно с определенной низкой часто-
той. Средняя частота нередуцированных яйцеклеток в этих
18 линиях составляла 59 на 10000 яйцеклеток (0,59%), а общая
Таблица 34.1. Некоторые неагамоспермные виды1*, у которых
имеются спонтанные триплоидные индивидуумы и триплоидные
клоны (по данным Дарлингтона и Уайли [151], Готтшока [258)
и другим источникам)
Семейство Вид Число хромосом з триплоидной фор мы Состояние
I Хвойные
Cupressaceae ’Juniperus virginiana л=11, 2/г=33 Культ.
И. Двудольные Thuja plicata excelsa х=Н, 2/г=33 »
Ranunculaceae Delphinium moerheimii х=8, 2/1 = 24 »
Cruciferae Nasturtium officinale х=8, 2/2=24
Cardamine insueta х=8, 2«=24 Дикор
Portulacaceae Clayionia virginica х=8, 2/г=24 »
Cactaceae Opuntla basilaris treleasei х=11, 2л = 33 х=8, 2п=24
Rosaceae Prunus serrulata Культ.
Matus sylvestris и дру- гие виды х=17, 2/г=51
Pyrus communis х=17, 2п=51
Sabcaceae Populus tremula х=19, 2/г =57 Дикор
Epacridaceae Leucopogon juniperinus х=4, 2/2=12 Дикор.
Compositae Melianthus multiflorus х=17, 2/г=51 Культ.
Inula brittanica /=8, 2/г=24
Primulaceae Primula sieboldii х=12, 2/1 = 36 »
Solanaceae Solanum juzepczukii и другие виды х=12, 2п=36 Ди кор и культ.
Scrophularia- ceae Limosella subulata X X aquatica х=10, 2/1=30 Дикор.
III Однодольные
Musaceae Musa acuminata х—11, 2п=33 Культ.
Cannaceae Саппа indica х=9, 2п=27
Liliaceae Hyacinthus orientalis х=8, 2п=24 »
Urginea indica х = 5, 2/1=15 Дикор
Allium сера tiuiparum х = 8, 2/г=24 Культ.
Allium rubellum и A ca- rinatum х=8, 2/г=24 Дикор.
I .ilium lancifolium х=12, 2п=36 Культ и дикор.
Tulipa gesneriana и дру- гие виды х=12, 2/г=36 Культ.
Paris verticillata х=5, 2п=15 Ди кор.
Amaryllidaceae Lycoris radiata х = 11, 2п=33 Культ.
Narcissus tazetta Hemerocallis fulva и H disticha х=10, 2д=30 х —11, 2/2=33
Iridaceae Crocus sativus и другие виды х=8, 2/1-24 Культ.
Freesta hybrida х=11, 2/1 = 33
Gladiolus formosus и другие виды х=15, 2/г=45
Araceae A corus calamus х=12, 2п=36
Colocasia antiquorum х=12, 2/1=36
Lemnaceae Lemna minor х=10, 2п=30 Дикор
Gramineae Phleum nodosum х=7, 2/1=21
О У всех перечисленных видов (за исключением Leucopogon) также имеются дип-
лоидные формы, у большинства из них имеются близко родственные тетраплоидные-
формы, а у некоторых видов или групп видов — анеуплоиды или высокие полиплоиды
454 Часть восьмая. Специализированные генетические системы
средняя частота для всех 121 линий составила 8,78 на 1000Q
яйцеклеток, т. е. 0,088% [3].
Сандфер [706] получил оценки частоты спонтанных триплоид-
ных проростков среди потомков диплоидных растений д£ля двух
культиваров ячменя; она составила 0,04% для одного сорта и
2,6% —для другого.
Частоты, лодобные обнаруженным у кукурузы и ячменя, дос-
таточны для того, чтобы обеспечить спонтанное возникновение
триплоидных растений во многих больших природных популя-
циях.
Естественным триплоидом, возникшим, вероятно, таким об-
разом, а именно в результате одностороннего отсутствия редук-
ции, является форма gigas вида Populus iremula в Швеции [575,
600]. Если говорить о культурных растениях, то некоторые три-
плоидные формы Musa acuminata также представляют собой
автотриплоиды, непосредственно происходящие от диплоидов
[726, 727].
Во многих других случаях Как среди дикорастущих, так и
среди культурных видов триплоидные формы возникли, по-види-
мому, в результате гибридизации между диплоидами и тетра-
плоидами. Большинство триплоидных растений встречается у
видов или у групп видов, содержащих и диплоидные, и тетра-
плоидные формы (см. табл. 34.1). Во многих реальных случаях,
как мы увидим далее, на основе ‘морфологических, цитологиче-
ских и другого рода данных установлено, что триплоиды пред-
ставляют собой гибриды.
Сохранение
При половом размножении триплоиды стерильны и нестабиль-
ны. Как же в таком случае они сохраняются?
Первое условие, необходимое триплоиду для достижения ус-
пеха,— наличие какого-либо способа вегетативного размноже-
ния. Все растения, перечисленные в табл. 34.1, —многолетники,
в большинстве своем мно"голетние травы, но среди них есть и
несколько древесных форм; многие из этих растений усиленно
размножаются вегетативным способом. Надежные способы ве-
гетативного размножения лежат в основе комбинированного
клонально-полиплоидного комплекса в группе Solanum tubero-
sum, которая содержит несколько триплоидных и пентаплоидных
микровидов, о чем уже говорилось в одной из предыдущих глав.
Второй хороший способ поддержания триплоидного состоя-
ния или нечетной полиплоидии более высокого уровня — это
агамоспермия. Среди агамоспермных членов группы Crepis occi-
dental's имеются микровиды с числом хромосом Зх, 5х и 7х (см.
Гл. 34, Естественные триплоиды
455
гл 32). Триплоиды встречаются также у агамоспермного вида
Taraxacum officinale
Генетическая система Rosa canina представляет собой спе-
циализированный способ воспроизведения состояния нечетной
полиплоидии. В группе Rosa canina эта система связана с псн-
таплоидией, у Leucopogon juniperinus — с триплоидией, а у
Cardamine insueta— также с триплоидией, но в модифицирован-
ной форме (гл. 30).
Избранные примеры
Обычная форма Populus tremula — диплоид (2п = 38). В отдель-
ных местностях как в южной, так и в северной частях Швеции
встречается форма gigas с 2п=±57, представляющая собой ав-
тотриплоид, у которого при мейозе образуются многочисленные
тривалснты Он продуцирует полустерильную пыльцу и в своих
природных местообитаниях распространяется при помощи веге-
тативного размножения. Эти триплоиды, вероятно, возникли в
результате соединения нередуцировапной и редуцированной га-
мет [575, 600].
Opuntia basilaris в пустынях на юго-западе Северной Амери-
ки представлена диплоидной формой (х=11). В одной из мест-
ностей в Калифорнии недавно было обнаружено триплоидное
растение. Этот триплоид возник, по-видимому, в результате от-
сутствия редукции у одной из родительских форм, поскольку в
остальном — это диплоидный вид и ни одной тетраплоидной фор-
мы у него не известно [643]
Культурные съедобные бананы в большинстве случаев явля-
ются триплоидами (х=11). Они происходят от дикорастущих
диплоидных видов Musa acuminata и М. balbisiana, плоды кото-
рых содержат семена. Бессемянные триплоидные сорта размно-
жаются вегетативно клубнелуковицами и корневищами. Неко-
торые триплоидные культивары, такие, как Грос Михель,—авто-
трип лоилы, происходящие от М acuminata, тогда как другие —
аллотриплоиды, возникшие в результате скрещивания между
Af. acuminata и М. balbisiana [726, 727]
Lilium lancifolium (х=12) встречается в Китае и Японии1.
Диплоидная форма, по-видимому, широко распространена Не-
которые культурные клоны — триплоиды; в отдельных местах в
Японии триплоиды встречаются также в природе. Триплоидные
растения продуцируют стерильную пыльцу и размножаются ве-
гетативно воздушными луковичками и луковицами. Прежде по-
лагали, что они представляют собой автотриплоиды, но теперь
1 Этот вид лилии встречается также у нас на Дальнем Востоке —Прин
ред
456 Часть восьмая. Специализированные генетические системы
их считают сегментными аллотриплоидами. Эти триплоиды, ве-
роятно, происходят от гибридов между L. lancifolium flaviflorum
(2х) и L. maximowiczii (2х). Один из родительских видов, по-
видимому, внес нередуцированную гамету, что и привело к воз-
никновению триплоидности. Триплоиды, очевидно, независимо
возникали в Японии несколько раз и распространились локально
в разных местах вегетативными способами [93, 617J.
Helianthus multiflorus — декоративное многолетнее травяни-
стое растение, выращиваемое в Европе. Это триплоид (2п=51)
со стерильными семенами, и его размножают вегетативными
органами. Он был получен в результате гибридизации между
тетраплоидом Н. decapetalus (2п—ЗА: II) и диплоидом И. annuus
(2n=17 II). Н. multiflorus является аллотетраплоидом, образую-
щим при мейозе 17 11+17 I [573, 575].
Представители Juniperus — преимущественно диплоиды
(х=11). У видов J. virginiana и J. chinensis имеются триплоид-
ные культивары [333, 805]. У J. chinensis и других видов встре-
чаются также тетраплоидные культивары [333]. Установлено,
что триплоидные культивары — это гибриды между J. chinensis
(4х) и /. virginiana (2х) [333].
Lemna minor, L. gibba и L. perpusilla (x=10) содержат фор-
мы, находящиеся на различных уровнях плоидности, в том чис-
ле и нечетные полиплоиды. У L. minor известны полиплоидные
формы 2х, Зх, 4х, 5х, 8х; у L. gibba — 4х, 5х, 7х, 8х, у L. регри-
silla— 2х, 4х, 5х, 6х, 8х [851]. Lemna размножается вегетатив-
ным путем и образует клоны, так что нечетные полиплоиды мо-
гут поддерживаться именно таким способом.
Триплоидные формы папоротников отличаются, как и следо-
вало ожидать, высокими хромосомными числами. Например,
группа Pityrogramma triangularis (Pteridaceae) (х=30) в Ка-
лифорнии состоит главным образом из диплоидов и тетрапло-
идов. В нескольких местностях обнаружены естественные три-
плоиды с 2плг 90. Триплоидные растения стерильны, и мейоз у
них нарушен. Установлено, что они являются гибридами, воз-
никшими в результате различных скрещиваний между тетра-
плоидами и диплоидами [12, 739]. Сохранение и распростране-
ние этих стерильных гибридных растений обеспечивается пол-
зучими подземными побегами.
Часть девятая
Заключение
35 Вид и макроэволюция
Сетчатая эволюция
Филогения многих или большинства групп высших растений,
которые были тщательно исследованы в природе и в лаборато-
рии, представляется сетчатой, а не исключительно дихотомиче-
ской и ветвящейся. Модель филогенетического древа нельзя пе-
реносить из зоологии в ботанику, так как во многих случаях это
приведет к искажению истинных взаимоотношений между гру п-
пами. Во всех крупных группах высших растений нам приходит-
ся иметь дело не с филогенетическими деревьями, а с филогене-
тическими сетями.
Формулировкой и разработкой концепции сетчатой эволюции
в царстве растений занималось несколько поколений ботаников.
Первыми авторами, высказавшими мнение о важной роли гиб-
ридизации в эволюции растений и сформулировавшими этот те-
зис в виде научно обоснованного обобщения, были Кернер [431]
и Лотси [519]. Их ранние формулировки страдали недостатками,
присущими всем первым шагам в любой области. Так, напри-
мер, Лотси вновь определил виды как чистые линии и поэтому
был склонен приравнивать гибридизацию главным образом, хо-
тя по исключительно, к обычному скрещиванию Работы Керне-
ра относятся, разумеется, к доменделевской эпохе, а кроме то-
10, он воздерживался от слишком катеюричных или слишком
определенных высказываний относительно эволюционных изме-
нений более крупных масштабов, чем видообразование Тем не
менее Кернер и Лотси поставили эту проблему достаточно
четко.
В первой половине нашего века ряд специалистов по гене-
тике растений— Винге, Тэкхолм, Реннер. Мюнтцинг, Густаф-
сон, Ментон и другие, — подойдя к изучению проблемы гибриди-
зации и эволюции с применением экспериментальных и индук-
тивных методов, стали получать для вес большего числа групп
растений данные в пользу сетчатой филогении Среди система-
тиков этого периода, также разделявших мнение о сетчатом ха-
рактере филогении у растений, были Дильс, Хайата и Дю Рие.
458
Часть девятая. Заключение
Несколько позднее в результате слияния генетических и таксо-
номических подходов возникла современная школа биосистема-
тики. К числу представителей этой школы, внесших важный
вклад в понимание сетчатой эволюции, принадлежат Андерсон,
'Стеббинс, Хейзер,'X. Люис, Эрендорфер, Л. Бенсон, Рейвн и
многие другие.
Для описания одного из аспектов сетчатой эволюции Эрен-
дорфер предложил полезную концепцию циклов дифференциа-
ции-гибридизации. Он подчеркивает, что во многих группах
растений процесс эволюции представляет собой циклическое
'чередование двух фаз — дивергенции и гибридизации [187, 188].
Эволюционный потенциал гибридных комплексов
В предшествующих главах было показано, что для гибридных
комплексов большинства типов — полиплоидных, агамных, кло-
яальных и гетерогамных — характерен период расцвета, за ко-
торым следует период упадка. Эти последовательные стадии
эволюционного продвижения и эволюционного отступления кор-
релируют с генетическими системами, в которых гены замкнуты
в определенных гетерозиготных комбинациях.
Амфиплоидия, агамоспермия, вегетативное размножение,
перманентная гетерозиготность по транслокациям и перманент-
ная нечетная полиплоидия — способы воспроизведения опреде-
ленных благоприятных гетерозиготных комбинаций генов, воз-
никших в результате гибридизации. Такое воспроизведение дос-
тигается путем строгого ограничения рекомбинации генов, а его
результаты выгодны перманентным гибридам в период расцвета
гибридного комплекса. Но в конечном счете перманентным гиб-
ридам приходится расплачиваться за свой оппортунизм, ибо,
когда в среде наступают, наконец, глубокие изменения, гибриды
порой оказываются не способными создавать новые генные ком-
бинации, адаптированные к новым условиям среды. Гибридный
комплекс вступает в период упадка.
Гомогамные комплексы отличаются от гибридных комплексов
всех других типов тем, что входящие в них гибридные произ-
водные обладают такой же неограниченной способностью к ре-
комбинации, как и исходные родительские виды. Гомогамный
комплекс может пользоваться всеми преимуществами, предостав-
ляемыми гибридизацией, не жертвуя при этом не менее выгод-
ной способностью к свободной рекомбинации генов. Отсюда
следует, что эволюционный потенциал гомогамного комплекса
не имеет никаких особых ограничений, а может, напротив, про-
грессивно развиваться на протяжении очень длительного време-
ни, давая начало новым крупным ветвям (272].
Гл 35 Вид и макроэволюция
459
Когда в агамном, клональном или гетерогамном и — в мень-
шей степени — полиплоидном комплексе предковые диплоидные
половые виды вымирают, их гибридные производные лишаются
важного источника новых вариаций. Весьма вероятно, что такая
утрата нанесет урон последующей эволюции данного 1ибридного
комплекса. В отличие от этого в гомо!ампом комплексе исход-
ные виды могут вымереть, не подвергая опасности эволюцион-
ный потенциал производных видов. В сущности, вполне возмож-
но, что предковые виды одного гомогамного комплекса пред-
ставляют собой конечные продукты какого-либо предыдущего
гомогамною комплекса [272]
На рис. 21.1 показаны различные «слои» (strata) гибридного
видообразования в гомогамном комплексе. Нет никаких осно-
ваний полагать, что в филетической истории данной группы су-
ществовали юлько эти известные нам «слои» гибридизации На-
против, по всей вероятности, эволюция этой и многих других
групп растений представляла собой последовательную смену го-
могамных комплексов [272]
Короче говоря, если для агамных, клональных и гетерогам-
ных комплексов типична закрытая система эволюции, а для
полиплоидных комплексов — ограниченная система, то можно
сказать, что гомогамные комплексы обладают открытой эволю-
ционной системой, т. е. системой, которая может существовать
в течение неопределенно долгого времени
Эволюция покрытосеменных — это макроэволюция: кроме
того, это эволюция сетчатая. Происходившие на протяжении
длительного времени прогрессивные эволюционные изменения в
различных основных филстических линиях покрытосеменных
могли сочетаться с сетчатым характером филогенетических свя-
зей в том и только в том случае, если эти филетическис линии
развивались как последовательные гомогамные комплексы [272],
потому что гомогамный комплекс — единственный тип гибрид-
ного комплекса с открытой эволюционной системой
Значение сетчатого характера эволюции
для филогении покрытосеменных
В этой кнше рассматривается эволюция растений па уровне
видов и групп видов Эволюция растений на этом уровне в зна-
чительной степени определяется относительно простой органи-
зацией растения, открытой системой его роста, относительной
простотой генных систем, контролирующих многие признаки, лег-
костью естественной гибридизации и тенденцией к унипаренталь-
ному размножению. Факторы эволюции и ее различные типы,
выявляемые в результате генетического и таксономического изу-
чения групп видов, позволят, быть может, пролить свет на про-
460
Часть девятая. Заключение
блемы, связанные с филогенетической классификацией высших
категорий.
Филогенетическая система классификации многих порядков
и надпорядков (или подклассов) покрытосеменных после таксо-
номического изучения на протяжении нескольких столетий все
еще остается неясной. В настоящее время в этой области ведет-
ся много успешных исследований представителями нескольких
научных школ, которые понимают трудности, связанные с соз-
данием такой системы, и не скрывают этого [130, 135, 788, 821,
828].
Трудность и неопределенность классификации покрытосемен-
ных на уровне порядков проистекает из сетчатого характера фо-
нетического сходства между разными группами. Рассмотрим че-
тыре родственные группы, обозначив их А, В, С и D. Группы А
и В имеют несколько общих морфологических признаков; другой
набор признаков является общим для групп В и С; и еще один
набор — для групп С и D. Для всей этой совокупности групп раз-
ные ученые могут и будут предлагать различные системы клас-
сификации— в зависимости от того, каким признакам каждый
из них придает первостепенное значение. Так, в соответствии с
одной системой, таксонами одного ранга могут считаться группы
А, В—С и D, в соответствии с другой — группы А—В и С—D.
Такого рода трудности и частично удовлетворительное их
разрешение характерны для поисков естественной системы по-
рядков в различных крупных группах покрытосеменных. Труд-
ности, возникающие в реальных случаях, больше, чем в нашей
простой гипотетической модели, так как переплетение фенети-
ческих взаимосвязей обычно захватывает дополнительные груп-
пы Е, F, G и т. д. Эта картина хорошо описана Кронквистом
для ряда реально существующих надпорядков и порядков в его
«Эволюции и классификации цветковых растений» [130].
Весьма важно, что описанная выше переплетающаяся таксо-
номическая структура, несмотря на ее широкое распростране-
ние у покрытосеменных, не универсальна. Некоторые крупные
группы покрытосеменных представляют собой ясно выраженные
филетические единицы и уже давно признаны таковыми. Хоро-
шим примером служат очень обширное семейство Compositae
и порядок Asterales, к которому оно относится. Деление покры-
тосеменных на две основные группы —на однодольных и дву-
дольных— признается всеми со времен Джона Рэя. Дихотоми-
ческая филогения высших животных — другой пример системы,
резко контрастирующей с сетчатой системой, характерной для
многих групп покрытосеменных.
Итак, во многих обширных группах покрытосеменных мы
наблюдаем сетчатую модель сходства по морфологическим при-
знакам; это подлинный факт, а не артефакт, обусловленный
Гл 35. Вид и макроэволюция
461
недостатками наших методов исследования, поскольку эти же
самые методы позволяют выявить дихотомическую в своей ос-
нове филогению там, где опа действительно существует, даже
в больших и разнообразных группах
Сетчатый характер морфологических взаимосвязей, наблю-
даемый на уровне порядков у покрытосеменных, можно также
наблюдать во mhoihx семействах тех же покрытосеменных на
промежуточном уровне родов и триб. Эта сетчатая модель фено-
типического сходства на всех уровнях заставляет вспомнить сет-
чатую модель генетических взаимосвязей, обнаруженную в комп-
лексах видов у растений. Такая общая модель побуждает нас
начать поиски лежащих в ее основе общих причин, присущих
как самим растениям, так и формам их эволюции
Одной из причин того, что как макро-, так и микроэволюция
растений носит сетчатый характер, является конвергенция, т. с.
приближение одной филетической линии к другой по несколь-
ким пли многим признакам
Вернемся к нашему гипотетическому примеру. Вспомним,
что у групп .4, В, С. D обнаружено сходство по морфологическим
признакам в комбинациях А и В, В и С, С и D. Л1ы часто обна-
руживаем то или иное приближение к следующей общей ситуа-
ции Родственные группы А и В различаются по признакам, ко-
юрые у группы А находятся на более примитивной, а у группы
В — на более продвинутой стадиях эволюции Сходным образом
у группы С наблюдается прогресс по ряду признаков по срав
ненпю с родственной, но более примитивной группой D В отли-
чие от этого сходство между группами В и С касается продви-
нутых или специализированных признаков. Иными словами, эти
сходные признаки — результат конвергентной эволюции.
Конвергентная эволюция может быть обусловлена отбором
без гибридизации или же совместным действием гибридизации
и отбора В любом случае полагают, что отдельные линии под-
вергаются отбору, благоприятствующему сходным адаптивным
реакциям па одни и те же или сходные условия среды. Оба ти-
па конвергентной эволюции часто встречаются у растений.
Легкость, с которой разные группы растений переходят из
одной экологической ниши в другую, что приводит к конвер-
генциям по целым наборам признаков, можно продемонстриро-
вать на сотнях примеров. Многие отдельные роды многолетних
трав в Средиземноморской области дали начало однолетникам;
различные группы мезофитов в пустыне конвергентно дали на-
чало ксерофитам; там, где живут колибри, у родов, опыляющих-
ся пчелами, неоднократно возникали виды, цветки которых при-
способлены к опылению колибри, и т д
Тот факт, что линии растений сравнительно свободно перехо-
дят из одной адаптивной зоны в другую, коррелирует с отно-
462
Часть девятая. Заключение
сительной простотой участвующих в этом генных систем. На ра-
совом уровне дивергенции в некоторых случаях можно обнару-
жить комбинации контрастирующих признаков, адаптированные
к различным экологическим нишам. Поэтому переключения, или
квантовые сдвиги, которые велики на экологическом уровне, но
относительно малы в генетическом плане, могут возникать не-
однократно в разных филетических линиях под направляющим
влиянием одного лишь параллельного отбора.
В пределах скрещиваемости, которые во многих группах
растений достаточно широки, дальнейшей конвергенции способ-
ствует гибридизация; в этом заключается вторая важная при-
чина сетчатого характера фенетических взаимосвязей у покры-
тосеменных.
Вид как единица макроэволюции
Некоторые молекулярные биологи, пытавшиеся изучать эволю-
цию органического мира, по сути дела, сводили* это явление к
равнодействующей между всего лишь двумя силами — мутаци-
онным процессом и отбором (см., например, [22, 51] и в мень-
шей степени [23]). К тому же представители этой школы не при-
нимали во внимание существование видов как неких единиц и
существование видообразования как эволюционного процесса.
Между тем, поскольку множество видов действительно возник-
ло в процессе эволюции, любое рассмотрение этого предмета,
не учитывающее формирование видов, не может внушать до-
верия.
Комбинацией мутационного процесса и отбора можно объяс-
нить биохимическую эволюцию, самые первые этапы биологиче-
ской эволюции и некоторые очень простые эволюционные изме-
нения у высших организмов. Но мутационно-селективная теория
не дает удовлетворительного объяснения эволюции более слож-
ных органических структур и функций. В результате одних лишь
мутаций и отбора на нашей планете вряд ли могло произойти
развитие растений и животных в течение того времени, которое
для этого имелось.
Признаки, при помощи которых организмы адаптируются к
условиям окружающей их среды, во всех случаях, за исключе-
нием самых простых, детерминируются не отдельными генами,,
а их комбинациями, обычно довольно сложными. На сравни-
тельно ранних этапах эволюционной истории, еще у однокле-
точных форм, возник половой процесс, представляющий собой
механизм, который создает комбинации и рекомбинации раз-
личных генов. В то же самое время в ответ на различные комп-
лексы условий среды в пределах одной общей области обитания
стали возникать дивергентные специализации. Воспользоваться
Гл. 35. Вид и макроэволюция
463:
преимуществами, которые предоставляет половое размножение,
сохраняя при этом дивергентные специализации, можно было’
только путем создания барьеров, препятствующих гибридизации,
как было показано ранее в этой книге.
Короче юворя, в поисках адекватного объяснения органи-
ческой эволюции мы никак не можем ограничиваться одними
лишь мутациями и отбором. Для развития сложных адаптаций
необходимо было половое размножение, а это в свою очередь
потребовало формирования репродуктивно изолированных ви-
дов
С тех пор как Дарвин дал своей книге название «О проис-
хождении видов», профессиональные эволюционисты признают,
что вид занимает центральное место в процессе эволюции Боль-
шинство биологов-эволюционистов разделяет представление о
том, что разнообразие видов на земном шаре — это система,
обеспечивающая непрерывное сохранение различных специали-
зированных образов жизни. Чго же касается соотношения меж-
ду видами и филетической эволюцией, то по этому вопросу бы-
ло высказано несколько точек зрения
Хаксли [392] рассматривал видообразование как процесс,
порождающий разнообразие, но не имеющий никакого отноше-
ния к прогрессивным эволюционным направлениям. Позднее
Майр [569] пришел к выводу, что если большинство видов — это
в сущности эволюционные тупики, то небольшому, но все же
достаточному их числу досталась такая комбинация благоприят-
ных признаков и экологических возможностей, которая позво-
лила им стать основателями новых доминантных групп. Пра-
вильность такой точки зрения отрицать невозможно. В дополне-
ние к ней я высказал предположение [282, 296], что видообра-
зование неразрывно связано с некоторыми случаями филетиче-
ской эволюции, в которых прогрессивное развитие происходит
путем ряда последовательных этапов видообразования.
За последние годы получает все большее признание так на-
зываемое прерывистое представление о макроэволюции. Соглас-
но этому представлению, виды и видообразование играют глав-
ную роль в эволюционных взрывах, характеризующих модель
прерывистой макроэволюции. Недавно Стенли [771, 772] разра-
ботал и очень хорошо сформулировал взгляд на видообразова-
ние как на основной процесс в макроэволюции.
Литература
1 Adams R. Р.» Turner В. L. Chemosystematic and numerical studies of natu-
ral populations of Juniperus ashei Buch, Taxon, Id, 728—751 (1970).
2. Addison G.f Tavares R. Hybridization and grafting in species of Theobroma
which occur in Amazonia, Evolution, 6,380—386 (1952).
3. Alexander D. E., Beckett J. B. Spontaneous triploidy and tetraploidy in
maize, J. Hered., 54, 103—106 (1963).
4. Allan H. H. Wild species-hybrids in the phanerogams, Bot. Rev, 15, 77—
105 (1969).
5. Allard R. W., Jain S. K. Population studies in predominantly self-pollinated
species. II. Analysis of quantitative genetic changes in a bulk-hybrid popu-
lation of barley, Evolution, 16, 90—101 (1962).
6. Allard R. W., Kahler A. L. Allozyme polymorphisms in plant populations,
Stadler Symposia (Univ. Missouri), 3, 9—24 (1971).
7. Allard R. W., Workman P. L. Population studies in predominantly self-
pollinated species. IV. Seasonal fluctuations in estimated values of genetic
parameters in lima bean populations, Evolution, 17, 470—480 (1963).
8. Alston R. E., Mabry T. J., Turner B. L. Perspectives in chematoxonomy,
Science, 142, 545—552 (1963).
9. Alston R. E., Turner B. L. New techniques in analysis of complex natural
hybridization, Proc. Nat. Acad. Sci., 48, 130—137 (1962).
10. Alston R. E., Turner B. L Biochemical Systematics, Prentice-Hall, Engle-
wood Cliffs, N J., 1963.
11 Alston R. E„ Turner B. L. Natural hybridization among four species of Bap-
tisia (Leguminosae), Amer. J. Bot., 50, 159—173 (1963).
12. Alt K. A, Grant V. Cytotaxonomic observations on the goldback fern, Brit-
tonia, 12, 153—170 (1960)
13. Anderson E Recombination in species crosses, Genetics, 24, 668—698
(1939).
14. Anderson E. Hybridization of the habitat, Evolution, 2, 1—9 (1948).
15. Anderson E, Introgressive Hybridization, Wiley, New York, 1949.
16. Anderson E. Introgressive hybridization, Biol. Reviews, 28, 280—307
(1953).
17. Anderson E., Anderson B. R. Introgression of Salvia apiana and Salvia
mellifera, Ann. Missouri Bot. Gard., 41, 329—338 (1954).
18. Anderson E., Gage A. Introgressive hybridization in Phlox bifida, Amer.
J. Bot., 39, 399—404 (1952).
19. Anderson E., Hubricht L. Hybridization in Tradescantia. III. The evidence
for introgressive hybridization, Amer. J. Bot., 25, 396—402 (1938).
20. Anderson E., Sax К A cytological monograph of the American species of
Tradescantia, Bot. Gaz., 97, 433—476 (1936).
21. Anderson E., Schafer B. Species hybrids in Aquilegia, Ann. Bot., 45, 639>—
646 (1931).
22 Anfinsen С. B. The Molecular Basis of Evolution, Wiley, New York, 1959.
[Имеется перевод: Анфинсен К. Молекулярные основы эволюции. — М.:
ЙЛ, 1962.]
23. Antonovics J. The effects of a heterogeneous environment on the genetics
of natural populations, Amer. Sci., 59, 593—599 (1971).
24. Antonovics J., Bradshaw A. D. Evolution in closely adjacent plant popula-
tions. VIII. Cllnal patterns at a mine boundary, Heredity, 25, 349—362
(1970).
25. Avery A. G., Satina S., Rletsema J. Blakeslee: The Genus Datura, Ronald
Press, New York, 1959.
26
27
28
29
30
31
32.
33
34
35
36
37.
38
39.
40
41.
42
43
44
45
46
47
48
49
50
Литература 465
Avery Р Cytogenetic evidences of Nicotiana phylesis in the a/afa-group,
Univ Calif Publ. Bot, 18, 153—194 (1938).
A vtshai Ai, Zohar и D Genetic affinities among the Oncocyclus Irises, Bot.
Gaz. 141, 107—115 (1980)
Ayala F J, ed Molecular Evolution, Sinauer Associates, Sunderland, Mass,
1976
Ayala F J, Valentine J W Evolving The Theory and Processes of Orga-
nic Evolution, Benjamin-Cummings, Menlo Park, Calif, 1979
Axelrod D I Evolution of the California closed cone pine forest, Proc
Symp. Biol Calif Islands (Santa Barbara, Botanical Garden), 93—149
(1967)
Axelrod D I History of the coniferous forests, California and Nevada, Univ
Calif Publ Bot. 70, 1—62 (1976)
Babbel G R . Wain R P Genetic structure of llordeum jubalum I Outcros-
sing rates and heterozygosity levels, Canad J Genet Cytol, 19, 143—
152 (1977)
Bobcock E В The genus Crepis, Univ. Calif Publ Bot Vols, 21, 22 (1947)
Babcock E B, Jenkins J. A Chromosomes and phylogeny in Crepis III
The relationships of one hundred and thirteen species, Univ. Calif. Publ
Bot, 18, 241—292 (1943)
Babcock E В, Stebbins G L The genus Youngia, Carnegie Inst Washing-
ton Publ no. 484, 1937.
Babcock E В, Stebbins G. L The American species of Crepis. Their inter-
relationships and distribution as affected by polyploidy and apomixis, Car
negie Inst Washington Publ no 504, 1938
Babcock E B, Stebbins G. L, Jenkins J A Chromosomes and phylogenv
in some genera of the Crepidinae, Cytologia, Fujii Jubilee Vol 188—210
(1937)
Baetcke К P, Alston R E. The composition of a hybridizing population of
Baptista sphaerocarpa and Baptisia leucophaea, Evolution, 22, 157—165
(1968)
Baker H. G Hybridization and natural gene-flow between higher plants,
Biol Rev, 26, 302—337 (1951)
Baldwin J T. Kalanchoe. the genus and its chromosomes, Amer J Bot,
25, 572—579 (1938).
Baldwin J T Galax- the genus and its chromosomes, J Hered, 32, 249—
254 (1941)
Barlow В A, Wiens D The cytogeography of the loranthaceous mistletoes,
Taxon, 20. 291—312 (1971)
Barnes В. V. The clonal growth habit of American aspens, Ecology, 47,
439—447 (1967).
Barnes В V. Indications of possible mid-Cenozoic hybridization in the
aspens of the Columbia plateau, Rhodora, 69, 70—81 (1967)
Barnes В. V Natural variation and delineation of clones of Populatus tre-
muloides and P grandidentata in northern Lower Mich'gan. Silvae Gene-
tica, 18, 130—142 (1969)
Bateman .4 J Genetical aspects of seed-growing. Nature, 157, 752—755
(1946)
Bateman A. J Cryptic self-incompatibility in the wallflower Cheiranthus
cheiri L„ Heredity, 10, 257—261 (1956)
Battaglia E Apomixis In Recent Advances in the Embryology of Angio-
sperms, ed P. Maheshwari, Internal Soc Plant Morph, Delhi, 1963
Baur E Einfuhrung in die expen’mentelle Ve-erbungslehre, 2nd ed , Born-
traeger, Berlin. 1914.
Baur E Artumgrenzung und Artbildung in de" Gattung Antirrhinum, Sck-
tion Antirrhinasirum, Zeitschr ind Abstammungs- u Vererbungslehre, 63,
256—302 (1932)
30-45
466
Литература
51. Beadle G. W. Genetics and Modern Biology, American Philosophical Society,
Philadelphia, Pa, 1963.
52. Beaman, J. H The systematics and evolution of Townsendia (Compositae),
Contrib. Gray Herbarium (Harvard), No. 133, 1957.
53. Beaudry J. R. The species concept: its evolution and present status, Revue
Canadienne de Biologie, 19, 219—240 (1960).
* 54. Beeks R. M. Variation and hybridization in southern California populations
of Diplacus (Scrophulariaceae), Aliso, 5, 83—122 (1962).
55 . Bell C. R. The Sanicula crassicaulis complex (Umbelliferae); a study of
variation and polyploidy, Univ. Calif. Publ. Bot., 27, 133—230 (1954).
56 . Bennett E. Historical perspectives in genecology Scottish Plant Breeding
Station Record, 49—115 (1964).
57 . Benson L. Plant Taxonomy, Ronald Press, New York, 1962.
58 . Benson L. The Native Cacti of California, Stanford University Press, Stan-
ford, 1969.
59 . Benson L., Phillips E. A., Wilder P. A Evolutionary sorting of characters in
a hybrid swarm. I. Direction of slope, Amer. J. Bot., 54, 1017—1026 (1967).
60 . Bergman B. Zytologische Studien fiber die Fortpflanzung bei den Gattun-
gen Leontodon und Pieris. Svensk Bot. Tidskrift, 29, 155—301 (1935).
61 Bernardini J. V„ Lima-de-Faria A. Asynchrony of DNA replication in the
chromosomes of Luzula, Chromosoma, 22, 91—100 (1967).
62 Blackhurst H T. Cytogenetic studies on Rosa rubiginosa L. and its hybrids,
Proc. Amer. Hort. Sci, 52, 510—516 (1948).
63 . Bogart J. P. Evolutionary implications of polyploidy in amphibians and
reptiles In* Polyploidy; Biological Relevance, ed. W. H. Lewis, Plenum
Press, New York and London, 1980
64 . Bradner N. R., Frakes R. V, Stephen W. P. Effects of bee species and iso-
lation distance on possible varietal contamination in alfalfa, Agron J., 57,
247—248 (1965).
65 . Bradshaw A. D. Population differentiation in Agrostis tenuis Sibth. I. Mor-
phological differentiation, New Phytol., 58, 208—227 (1959).
66 . Brainerd E Some natural violet hybrids of North America. Vermont Agric.,
Exp. Sta. Bull. No. 239, 1924.
67 . Breedlove D E. The systematics of Fuchsia section Encliandra (Onagra-
ceae), Univ. Calif Publ Bot., 53, 1—69 (1969).
68 . Bridges С. B. The origin of variations in sexual and sex-limited characters,
Amer. Nat, 56, 51—63 (1922)
69 . Bright C. A. Telocentric chromosomes in Corsican Crocus L. (Iridaceae),
Plant Syst. Evol., 129, 299—314 (1978).
70 . Brink R. A Physe change in higher plants and somatic cell heredity, Quart.
Rev. Biol., 37, 1—22 (1962).
71 . Brown S. The origin and nature of variability in the Pacific coast black-
berries (Rubus ursinus Cham, and Schlecht. and R. lemurum sp. nov.),
Amer. J. Bot, 30, 686—697 (1943).
72 . Brown S Mitosis and meiosis in Luzula campestris DC, Univ. Calif Publ.
Bot, 27, 231—278 (1954)
73 . Brown W. L., Wilson E. O. Character displacement, Systematic Zool, 5,
49—64 (1956).
74 . Buffon C. de. Histoire Naturelie, vol. 2, Paris, 1749.
75 . Bungenberg de Jong С. M. Polyploidy in animals, Bibliographia Genetica,
17, 111—228 (1957).
76 . Burma В. H. Reality, existence, and classification: a discussion of the spe-
cies problem, Madrono, 12, 193—209 (1954).
77 Bush G L Modes of animal speciation, Ann. Rev. Ecol. Syst., 6, 339—364
(1975).
78 . Cadman С H Autotetraploid inheritance in the potato; some new evidence,
J. Genetics, 44, 33—52 (1942).
Литература
467
79 Cain A J Animal Species and Their Evolution. Hutchinson, London and
Harper and Row, New York, 1954
80 Cameron J W, Frost H В Genetics, breeding, and nucellar embryony
In The Citrus Industry, 2d ed , vol 2, eds W Reuther, L D Batchelor and
Webber, University of California Press, Berkeley and Los Angeles,
81 Camp 117 H, Gilly C L The structure and origin of species, Brittonia, 4,
323-385 (1943).
82 Carlquist S On the genetic limits of Eriophyllum (Compositae) and rela-
ted genet a, Madrono, 13, 226—239 (1956)
83 Carson H L The genetic characteristics of marginal populations of Dro-
sophila, Cold Spring Harbor Symp Quant Biol, 20, 276—287 (1955)
84 Carson H L Genetic conditions which promote or retared the formation
of species, Cold Spring Harbor Symp Quant Biol, 24, 87—105 (1959)
85 Carson H / Chromosome tracers of the origin of species, Science, 168,
1414—1418 (1970)
86 Carson H L Speciation and the founder principle, Stadler Genet Sym-
posia (Univ Missouri), 3, 51—70 (1971).
87 Carson H L The genetics of speciation at the diploid level, Amer. Nat,
109. 83—92 (1975)
88 Carson H L, Heed W В St-uctural homozygosity in marginal populations
of nearclic and neotropical species of Drosophila in Florida, Proc. Nat
Acad Sci, 52, 427—430 (1964)
89 Caspari E Cvtoplasmic inheritance, Advances in Genetics, 2, 1—66
(1948)
90 Cave Л1. S, Constance L Chromosome numbers in the Hvdrophyllaceae
III, Univ. Calif Publ. Bot, 18, 449-465 (1947)
91 Cave Л4 S, Constance L Chromosome numbers in the Hydrophyllaceae
IV, Univ Calif Publ Bot, 23, 363—382 (1950).
92 . Chambers К L. A biosvstematic study of the annual species of Microseris
Contrib Dudley Herbarium (Stanford), 4, 207—312 (1955).
93 Chandler C. Porterfield W M, Stout A В Microsporogenesis in diploid
and triploid types of Liliutn tigrinum with special reference to abortions,
Cytologia, Fujii Jub, vol 2, 756—784 (1937)
94 Chase V C, Raven P H Evolutionary and ecological relationships between
Aquilegia formosa and A pubescens (Ranunculaceae), two perennial plants,
Evolution, 29, 474—486 (1975)
95 Clausen J. Genetical and cytological investigations on Vto/a tricolor L. and
V arvensis Murr, Hereditas, 8. 1—156 (1926)
96 Clausen J Stages in the Evolution of Plant Species, Cornell University
Press, Ithaca, N Y, 1951.
97 . Clausen J Population studies of alpine and subalpine races of conifers and
willows in the California high Sierra Nevada, Evolution, 19, 56—68
(1965)
98 Clausen J, Hieseu W M Experimental studies on the nature of species
IV Genetic structure of ecological races. Carnigie Inst Washington Publ,
615, 1958
99 Clausen 1, Hiesey U7 M The balance between coherence and variation in
evolution, Proc Nat Acad Sci, 46, 494—506 (1960)
100 Clausen J, Keck D D, Hiesey P7 M The concept of species based on expe-
riment, Amer. J Bot, 26, 103—106 (1939)
101 . Clausen J, Keck D D. Hiesey W M Experimental studies on the nature
of species I Effec* of varied environments on western American plants,
Carnegie Inst. Washington Publ, 520, 1940
102 Clausen J, Keck D D. Hiesey W M Experimental studies on the nature of
species II Plan: evolution through amphioloidy and autoplo'dy, with exam-
ples from the Madiinae, Carnegie Inst Washington Publ., 564, 1945
30’
468
Литература
ЮЗ. Clausen. J., Keck D. D., Hiesey W. M. Experimental studies on the nature of
species. III. Environmental responses of climatic races of Achillea, Carne-
gie Inst. Washington Publ., 581, 1948.
104 . Clausen J.., Keck D. D., Hiesey W. M., Grun P. Experimental taxonomy,
Carnegie Inst. Washington Yearbook, 48, 95—106 (1949).
105 . Clausen R. E., Goodspeed T. H. Interspecific hybridization in Nicotiana.
II. A tetrapioid glutinosa-tabacum hybrid, and experimental verification of
Winge’s hypothesis, Genetics, 10, 279—284 (1925).
106 . Clausen R. T. Sedutn of the Trans-Mexican Volcanic Belt, Cornell Univer-
sity Press, Ithaca, N. Y., 1959.
107 . Clayton E. E Male-sterile tobacco, J Heredity, 41, 171—175 (1950)
108 . Clegg M. T, Allard R. W. Viability versus fecundity selection in the slen-
der wild oat, Avena barbata L., Science, 181, 667—668 (1973).
109 . Cleland R E Some aspects of the cyto-genetics of Oenothera, Bot. Rev.,
2,316—348 (1936).
110 . Cleland R E. Phylogenetic relationships in Oenothera, Hereditas, Suppl.
Vol., 173-188, 1949.
111 . Cleland R. E. Studies in Oenothera cytogenetics and phylogeny, Indiana
Univ. Publ., Science Series, No. 16 (1950).
112 . Cleland R. E. The cytogenetics of Oenothera, Advances in Genetics, 11,
147—237 (1962).
113 . Cleland R. E. The evolutionary history of the North American evening pri-
mroses of the «biennis group>, Proc. Amer. Phil. Soc., 108, 88—98
(1964).
114 . Cleland R. E. Oenothera: Cytogenetics and Evolution, Academic Press, New
York, 1972.
115 . Clifford H. T. Analysis of suspected hybrid swarms in the genus Eucalyp-
tus, Heredity, 8, 259—269 (1954).
116 . Cockrum E. L. A check-list and bibliography of hybrid birds in North Ame-
rica north pf Mexico, Wilson Bull., 64, 140—159 (1952).
117 . Colwell R. N. The use of radioactive isotopes in determining spore distri-
bution patterns, Amer. J. Bot., 38, 511—523 (1951).
118 Conant D, S., Cooper-Driver G Autogamous allohomoploidy in Alsophila
and Nephelea (Cyatheaceae): a new hypothesis for speciation in homoploid
homosporous ferns, Amer J. Bot., 67, 1269—1288 (1980).
119 . Constance L. Chromosome number and classification in Hydrophyllaceae,
Brittonia, 15, 273—285 (1963).
120 . Cook R. E. Patterns of juvenile mortality and recruitment in plants In:
Topics in Plant Population Biology, ed. O. Solbrig, Columbia University
Press, New York, 1979.
121 . Cook S. A. Genetic system, variation and adaptation in Eschscholzia cali-
fornica, Evolution, 16, 278—299 (1962).
122 . Cooperrider M. Introgressive hybridization in Quercus marilandlca, Amer.
J Bot., 44, 804-810 (1957).
123 . Correll D S The Potato and its Wild Relatives, Section Tuberarium of the
Genus Solanum, Texas Research Foundation, Renner, Texas, 1962.
124 . Cottam W P Prevernal leafing of aspen in Utah mountains, J. Arnold
Arboretum, 35, 239—248 (1954).
125 Cottam W P, Tucker J. №.. Drobnick R. Some clues to Great Basin post-
pluvial climates provided by oak destributions, Ecology, 40, 361—377
(1959).
126 . Cousens J. E. Variation of some diagnostic characters of the sessile and
pedunculate oaks and thetr hybrids in Scotland, Watsonia, 5, 273—286
(1963).
127 . Cousens J. E. The status of the pedunculate and sessile oaks in Britain,
Watsonia, 6, 161—176 (1965).
Литература
469
128 Crane М В, Mather К The natural cross-pollination oi crop plants with
particular reference to the radish, Ann Appl Biol, 30, 301—308 (1943).
129 Critchjield U7 В Interspecific hybridization in Pinus a summary review,
Proc 14th Meeting Canadian Tree Improvement Assoc, 2, 99—105 (1975).
130 Cronquist A The Evolution and Classification oi Flowering Plants, Hough-
ton, Boston, Mass, 1968
131 Cronquist A Once again, what is a species’ Beltsville Symp Agric Re-
search, 2, 3—20 (1978)
132 Cruden ft W Pollen-ovule ratios' a conservative indicator of breeding sys-
tems in flowering plants, Evolution, 31, 32—46 (1977)
133 . Crumpacker D. W7, Williams ! S Density, dispersion, and population struc-
tuie in Drosophila pseudoobscura, Ecol. Monogr, 43, 4991—538 (1973).
134 Cuenot L L'Evolution biologique, Masson, Paris, 1951
135 Dahlgren ft. A system of classification of the angiosperms to be used to
demonstrate the distribution of characters, Botaniska Notiser, 128, ПЭ-
147 (1975)
136 . Danser В H Ober die Begnffe Komparium, Kommiskuum und Konvivium
und uber die Entstehungswcise der Konvivicn, Genetica, II, 399—450
(1929)
137 Dansereau P M Etudes sur les hybrides de Cistes VI. Introgression dans
la section Ladanum, Canad. J. Res, C, 19, 59—67 (1941).
138 Darlington C D Recent Advances in Cytology, 1st ed, Churchill, London,
1932
139 Darlington C. D Recent Advances in Cytology, 2nd ed. Churchill, London,
1937
140 Darlington C D The earlv hybridisers and the origins of genetics, Herber-
tia 4 63—69 (1937)
141 Darlington C D The Evolution of Genetic Systems, 1st ed, Cambridge
University Press. Cambridge, 1939
142 Darlington C D Taxonomic species and genetic systems In The New Sys-
tematics, ed J Huxley, Clarendon Press, Oxford, 1940
143 Darlington C D Natural populations and the breakdown of classical gene-
tics, P-oc. Roy Soc. В 145, 350—364 (1956)
144 Darlington C D. Chromosome Botany, 1st ed , George Allen and Unwin,
London 1956
145 Darlington C D The Evolution of Genetic Systems. 2d ed , Basic Books,
New York 1958
146 Darlington C D Chromosome Botany and the Origins of Cultivated Plants,
2d ed , Hafner, New York, 1963
147 Darlington C D Cytology, Churchill, London. 1965
148 Darlington C D Chromosome Botany and the Origin of Cultivated Plants,
3d cd, Hafner, New York, 1973
149 Darlington C D, Janaki Ammal E ft Chromosome Atlas of Cultivated
Plants, 1st cd, George Allen and Unwin, London 1945
150 Darlington C D, Mather ft The Elements of Genetics George Allen and
Unwin, London, 1949
151 . Darlington C D, Wylie A P Chromosome Atlas of Flowering Plants, 2nd
ed . George Allen and Unwin, London, 1955
152 Darwin C. On the Origin of Species, 1st ed., John Murray, London, 1859
Reprinted by Watts and Co, London, 1950; and by Harvard University
Press, Cambridge, Mass. 1964
153 Darwin C The Variation of Animals and Plants under Domestication,
2 vols, John Murray, London, 1868
154 Darwin C On the Orig.n of Species, 6th ed, John Murray, London, 1862,
various modern reprints
155 . Davidson J F A dephlogisticated species concept, Madrono, 12, 246—251
(1954).
470
156.
157.
158.
159.
160.
161.
162.
163.
164.
165.
166.
161.
168.
169.
170.
171
172.
173.
174
175.
176.
177.
178.
179.
180
Дитература
Davies Е. Cytology, evolution and origin of the aneuploid series in the
genus Carex, Hereditas, 42, 349—365 (1956).
Davis P. H„ Heywood V H Principles of Angiosperm Taxonomy Van Nost-
rand, Princeton, N. J., 1963,
Day A The evolution of a pair of sibling allotetraploid species of Cobwebby
Gilias (Polemoniaceae), Aliso, 6, 25—75 (1965).
Delevoryas T. Polyploidy in gymnosperms. In: Polyploidy; Biological Re-
levance, ed. W. H. Lewis, Plenum Press, New York and London, 1980
De Wet J. M. J. Diploid-tetraploid-haploid cycles and the origin of variabi-
lity in Dichanthium agamospecies, Evolution, 22, 394—397 (1968).
Dempster L T„ Ehrendorfer F. Evolution of the Galium multiflorum complex
in western North America. II. Critical taxonomic revision, Brittonia, 17,
289-334 (1965).
Dhillon T. S., Garber E. D. The genus Collinsia XIX. «New» genomes
from two fertile interspecific hybrids, Cytologia, 27, 189—203 (1962).
Diels L. Die Methoden der Phytographie und der Systematik der Pflanzen.
In: Abderhaldens Handbuch der biologischen Arbeitsmethoden, Abt. 11, Ber-
lin, 1921.
Dietrich W. The South American species of Oenothera sect, Oenothera
(Raimannia, Renneria; Onagraceae), Annals Missouri Bot. Gard., 64, 425—
626 (1977).
Dobzhansky Th. Genetic nature of species differences, Amer. Nat., 71, 404—
420 (1937).
Dobzhansky Th. Genetics and the Origin of Species, 1st, 2d and 3d eds.,
Columbia University Press, New York, 1937, 1941, 1951:
Dobzhansky Th. Speciation as a stage in evolutionary divergence, Amer.
Nat., 74, 312—321 (1940).
Dobzhansky Th. Mendelian populations and their evolution, Amer. Nat.,
84, 401—418 (1950)
Dobzhansky Th. Species after Darwin. In. A Century of Darwin, ed.
S. A. Barnett, Heinemann, London, 1958.
Dobzhansky Th. Genetics of the Evolutionary Process, Columbia University
Press, New York, 1970.
Dobzhansky Th, Ayala F J. Stebbins G. L„ Valentine J. W. Evolution,
W H. Freeman, San Francisco,
Dobzhansky Th, Ehrman L, Pavlovsky О Drosophila insularis, a new
sibling species of the willistoni group, Studies in the genetics of Drosophila
(Univ, of Texas), 9, 39-47 (1957).
Dobzhansky Th., Epling C. Contributions to the genetics, taxonomy, and
ecology of Drosophila pseudoobscura and its relatives, Carnegie Inst. Wa-
shington Publ, 554, 1944.
Dodds К S. Classification of cultivated potatoes. In- The Potato and Its
Wild Relatives, by D. S. Correll, Texas Research Foundation, Renner, Te-
xas, 1962.
Dodds K. S., Paxman G. J. The genetic system of cultivated diploid pota-
toes, Evolution, 16, 154—167 (1962)
Dodson С. H. Cytological studies in the genus Oncidium, Ph. D. thesis, Cla-
remont Graduate School, Claremont, Calif, 1956.
Dodson С. H. Frymire G. P. Preliminary studies in the genus Stanhopea
(Orchidaceae), Ann. Missouri Bot. Gard, 48, 137—172 (1961).
Doll R. Revision der sect Erythrosperma Dahlst emend Lindb f der Gat-
tung Taraxacum Zinn, Feddes Repertorium, 83, 673—740; 84, 1—180
(1973).
Drury W H The ecology of the natural origin of a species of Carex by
hybridization, Rhodora, 58, 51—72 (1956).
Duffield J. W Relationships and species hybridization in the genus Pinus,
Zeitschr. Forstgenetik u. Forstpflanzenzucht, 1, 93—97 (1952).
Литература
47Г
181 . DuRielz G E The fundamental units of biological taxonomy, Svensk Bot.
Tidskr, 24, 333-428 (1930)
182 Easi E M Studies on size inheritance in Nicotiana, Genetics, 1, 164—176-
(1916)
183 . Edwardson / R. Cytoplasmic male-sterility, Bot. Rev, 22, 696—738
(1956)
184 . Ehrendor^er F. Zur Phylogenie der Gattung Galium L, I Polyploidie und
geographisch-okologische Einheiten in der Gruppe des Galium pumilum
Mur-ay (Sekt Lcptogalium Lange sensu Rouy) im osterreichischcn Alpen-
raum, Osterr Bot Zeitschr, 96, 109-138 (1949).
185 Ehrendorfer F Zur Phylogenie der Gattung Galium II Rassengliederung,
Variab’lLatszentrcn und gcographische Merkmalsprogressionen als Aus-
druck der raum-zeitlichen Entfaltung des Formenkreises Galium incanum
S S Osterr Bot Zeilschr , 98, 427—490 (1951)
186 Ehrendorfer F Okologisch-geographische Mikro-Differenzierung einer Popu-
lation von Galium pumilum Mrr s str Osterr, Bot Zeitschr, 100, 616—
638 (1953)
187 Ehrendorfer F Zur Phylogenie der Gattung Galium VI En Variabilitas-
zentrum als «fossiler» Hybrid Komplex* dor ost-mediterranc Galium grae-
cum L —G canum Req Formenkreis, Osterr, Bot Zeitschr, 105, 229—279
(1958).
188 Ehrendorfer F Differentiation-hybridization cycles and polyploidy in Achil-
lea, Cold Spring Harbor Symp. Quant Biol, 24, 141—152 (1959)
189 . Ehrendorier F Evolution of the Galium multtflorum complex in western
North Amer ca I. Diploids and polvploids in this dioecious group, Madrono,
16, 109—140 (1961)
190 Ehrendorier F Beitrage zur Phylogenie der Gattung Rnautm (Dipsacaceae).
I Cytologische Grundlagen und allgemeine Hinweise, Osterr, Bot Zeitschr,
109, 276—343 (1962)
191 Ehrendorfer F Cytologic, Taxonomie and Evolution bei Samenpflanzen,
Vistas in Botany, 4, 99—186 (1964)
192 Ehrendorfer F Dispersal mechanisms, genetic systems and colonizing abili-
ties in some 'lowering plant families In The Genetics of Colonizing Spe-
cies, eds H G. Baker and G L Stebbins, Academic Press, New York,
1965
193 Ehrendcrfer F Polyploidy and distribution In Polyploidy, Biological Re-
levance, ed W H. Lewis, Plenum Press, New York and London, 1980.
194 Ehrendorjer F, Krendl F, Habeler E, Sauer W Chromosome numbers and
evolution in orimitive angiosperms, Taxon, 17, 337—353 (1968)
195 Ehrlich P R, Holm R W Patterns and populations, Science, 137, 652—
657 (1962)
196 Ehriich P R, Raven P H Differentiation of populat ons, Science, 165,
1228—1232 (1969)
197 Ehrlich. P R, Wnite R R Colorado checkerspot butterflies isolation, neutra-
lity, and the biospccies. Amer. Nat, 115, 328—341 (1980)
198 Ehrman L Hybrid sterility as an isolating mechanism in the genus Dro-
sophila, Quart Rev Biol, 37, 279—302 (1962)
199 Ehrman L Direct observation of sexual isolation between allopatric and
between sympatric strains of the different Drosophila paulistorum races,
Evolution. 19. 459-464 (1965)
200 Emerson. S H The genetic nature of De Vries’s mutations in Oenothera
lamarckiana, Amer Nat, 69, 545—559 (1935).
201 Endler J A Geographic variation Speciation, and Clines, Princeton Uni-
versity Press, Princeton, N. J, 1977
202 . Epling C Natural hybridization of Salvia apiana and S mellifera, Evolu-
tion, 1, 69—78 (1947).
472
Литература
203 . Epling С., Dobzhansky Th. Genetics of natural populations, VI. Microgeo-
graphic races in Linanthus parryae, Genetics, 27, 317—332 (1942).
204 . Epling C., Lewis H., Ball F. M. The breeding group and seed storage:
a study in population dynamics, Evolution, 14, 238—255 (1960).
205 . Ernst A. Bastardierung als Ursache der Apogamie im Pflanzenreich, Gustav
Fischer, Jena, 1918.
206 Faegri K, van der Pijl L. The Principles of Pollination Ecology, 1st and 3d
eds., Pergamon Press, Oxford, 1966; 1979.
207 . Fagerling F. Sind die canina-Rosen agamospermische Bastarde? Svensk
Bot. Tidskr., 34, 334—354 (1940).
208 . Fagerlind F. Die Bastarde der canina-Rosen, ihre Syndese- und Formbil-
dungsverhaltnisse, Acta Horti Bergiani (Uppsala), 14, 7—37 (1945).
209 . Fassett N. C. Juniperus virginiana, J. horizontalis and J. scopulorum II.
Hybrid swarms of J. virginiana and J. scopulorum, Bull. Torrey Bot. Club.,
71, 475-483 (1944)
210 . Федоров А. А. ред. Хромосомные числа цветковых растений, Изд-во АН
СССР, Л, 1969.
211 . Feldman М. Wheats. In: Evolution of Crop Plants, ed. N. W. Simmonds,
Longman, London and New York, 1976.
212 . Fernandes A Sur la phylogfcnie des especes du genre Narcissus L., Both
Soc. Broteriana, 25, 113—190 (1951).
213 . Fisher fi. A The Genetical Theory of Natural Selection, Clarendon Press,
Oxford, 1930
214 . Fisher fi A Retrospect of the criticisms of the theory of natural selection.
In Evolution as a Process, ed. J. Huxley, George Allen and Unvin, London,
1954.
215 . Flake fi. H., von fiudloff E., Turner B. L. Quantitative study of ciinal va-
riation in Juniperus virginiana using terpenoid data, Proc. Nat. Acad. Sci.,
64, 487—494 (1969).
216 Flake fi. H., Urbatsch L„ Turner В L. Chemical documentation of allopat-
ric introgression in Juniperus, Syst. Bot, 8, 129—144 (1978).
217 . Focke W. О Die Pflanzenmischlinge, Borntraeger, Berlin, 1881
218 . Ford E B. Ecological Genetics, 1st and 3d ed, Methuen and Co., and
Chapman and Hall, London, 1964; 1971.
219 . Frankel O., Munday A The evolution of wheat. In: The Evolution of Living
Organisms, Symposium, Royal Society, Victoria, Australia, 1962.
220 Freter L. E.. Brown W V. A cytotaxonomic study of Bouteloua curtipendula
and В uniflora, Bull. Torrey Bot. Club, 82, 121—130 (1955).
221 . Frisch fi Der Farbensinn und Formensinn der Biene, Zool. Jahrb., 35, 1—
182 (1914).
222 . Frisch fi Uber den Geruchslnn der Biene und seine blutenbiologische Be-
deutung, Zool. Jahrb., 37, 1—238 (1919)
223 . Fryxell P. A. Mode of reproduction in higher plants, Bot. Rev, 23, 135—
233 (1957).
224 FUrnkranz D. Cytogenetische Untersuchungen an Taraxacum in Raume von
Wien, Osterr, Bot. Zeitschr., 107, 310-350 (I960).
225 . FUrnkranz D. Taraxacum apenninum — ein altes Element mediterraner Ge-
birge, Osterr. Bot. Zeitschr., Ill, 231—239 (1964).
226 . FUrnkranz D. Untersuchungen an Populationen des Taraxacum-officinale
Komplexes im Kontakgebiet der diploiden und polyploiden Biotypen, Osterr.
Bot Zeitschr, 113, 427—447 (1966)
227 . FUrnkranz D Eine neue mediterrane Reliktart der Gattung Taraxacum,
Osterr. Bot. Zeitschr, 116, 149—156 (1969).
228 . Gaiewski W Cytogenetic investigations on the genus Geum L, Hereditas,
Suppl. Vol., 578—579 (1949)
229 . Gaiewski W. A fertile amphipolyploid hybrid of Geum rivale with G macro-
phyllum, Acta Soc. Bot. Polon, 22, 411-439 (1953).
Литература
473
230 Gajewski W A cytogenetic study on the genus Geum L. Monographiae
Botanicue (Warsaw), vol. 4 (1957).
231 Gajewski №. Evolution in the genus Geum, Evolution, 13, 378—388
(1959)
232 Garber E D. The genus Collinsia 111 The significance of chiasmata fre-
quencies as a cy totaxonomic tool, Madrono, 14, 172- 176 (1957).
233 Garbe' e d The genus Collinsia IX Speciation and chromosome repatter-
ning, Cyrologia, 25, 233—243 (1960).
234 . Garber E D Collinsia Handbook of Genetics, 2, 333—361, ed R C. King,
Plenum Press, New York, 1974
235 Garber E D, Gorsic J The genus Collinsia II. Interspecific hybrids invol-
ving C heterophylla, C concolor and C sparsiflora, Bot Gaz, 118, 73—77
(1956)
236 Gates R R The taxonomic units in relation to cytogenetics and gene eco-
logy, Amer. Nat, 85, 31—50 (1951)
237 Gerassitnova H Chromosome alterations as a factor of divergence of forms.
1 New experimentally produced strains of C tectorum which are physiolo-
g cally isolated from the original foims owing to reciprocal translocation,
Compt Rend Acad Sci URSS, 25, 148-244 (1939)
238 Gerstel D U Chromosomal translocations in interspecific hybrids of the
genus Gossypium, Evolution, 7, 234—244 (1953).
239 . Gersiel D U A new lethal combination in interspecific cotton hybrids, Ge-
netics, 39, 628—639 (1954).
240 Gersiel D U, Phillips L L Segregation of synthetic amphiploids in Gossy-
pium and Nicotiana. Cold Spring Harbor Symp. Quant. Biol., 23, 225—237
(1958)
241 Gersiel D U, Sarvella P A Additional observations on chromosomal trans-
locations in cotton hybrids, Evolution, 10 408—414 (1956)
242 Gibson J B, Thoday J M Effects of disruptive selection, IX Low selection
intensity, Heredity, 19, 125-130 (1964).
243 Gilles A , Randolph L F Reduction in quadrivalent frequency in autotetra-
ploid maize during a period of 10 years, Amer J Bot, 38, 12—16 (1951).
244 Gilmour J S L, Heslop-Harrison J The deme terminology and the units
of micro-evolutionary change, Gcnctica, 27, 147—161 (1954).
245 Glass В The germination of the idea of biological species In Forerunners
of Darwin, 1745—1859, eds В Glass, О Temkin and W L Straus, John
Hopkins University Press. Baltimore, 1959
246 Goldblatt P Polyploidy in angiosperms monocotyledons In Polyploidy;
Biological Relevance, ed W H Lewis, Plenum Piess, New York and Lon-
don, 1980
247 . Goldschmidt R В Physiological Genetics, McGraw-Hill, New York, 1938
248 Goldschmidt R В The Material Basis of Evolution, ’Yale University Press,
New Haven Conn, 1940
249 Goldschmidt R В Theoretical Genetics, University of California Press,
Berkeley, Calif, 1955
250 Goodspeed T H The Genus Nicotiana, Chronica Botanica, Waltham, Mass,
1954
251 Gottlieb L D Evolutionary relationships in the outcrossing diploid annual
species of Stephanomeria "(Composilae), Evolution, 25, 312—329 (1971).
252 Gottlieb L D Genetic differentiation, sympatric speciation and the origin
of a diploid species of Stephanomeria, Amer J Bot, 60, 545—553 (1973)
253 Gottlieb L D Enzyme differentiation and phylogeny in Clarkia franciscana,
C rubicunda and C amoena. Evolution, 27, 205—214 (1973)
254 Gottlieb L D Genetic confirmation of the origin of Clarkia Ungulata, Evo-
lubon 28, 244—250 (1974)
255 Gottlieb L D Electrophoretic evidence and plant systematics, Annals Mis-
souri Bot Gard, 64, 161 —180 (1977)
474
Литература
256 . Gottlieb L. D. Phenotypic variation in Stephanomeria exigua ssp. coronaria
(Compositae) and its recent derivative species «malheurensis», Amer. J. Bot.,
64, 873—880 (1977).
257 Gottlieb L. D. Stephanomeria malheurensis (Compositae), a new species
from Oregon, Madrono, 25, 44—46 (1978).
258 . Gottschalk W Die Bedeutung der Polyploidie fflr die Evolution der Pflan-
zen, Gustav Fischer, Stuttgart, 1976.
259 . Gould F. W Notes on apomixis in sideoats grama, J. Range Management,
12, 25—28 (1959).
260 . Gould F. №. Chromosome numbers of some Mexican grasses, Canad. J. Bot.»
44, 1683—1696 (1966).
261 . Gould F. W., Kapadia Z. J. Biosystematic studies in the Bouteloua curti-
pendula complex. I. The aneuploid rhizomatous B. curtipendula of Texas,
Amer. J. Bot., 49, 887—892 (1962).
262 . Gould F. №.. Kapadia Z. J. Biosystematic studies in the Bouteloua curti-
pendula complex. II Taxonomy, Brittonia, 16, 182—207 (1964). \
263 . Grant A., Grant V. Genetic and taxonomic studies in Gilia. VIII. The Cob-
webby Gilias, Aliso, 3, 203—287 (1956).
264 Grant K. A.. Grant V. Mechanical isolation of Saliva apiano and Salvia
mellifera (Labiatae), Evolution, 18, 196—212 (1964).
265 . Grant V. Pollination systems as isolating mechanisms in flowering plants,
Evolution, 3, 82—97 (1949).
266 . Grant V Genetic and taxonomic studies in Gilia. I. Gilia capitata, Aliso,
2, 239—316 (1950).
267 . Grant V. The flower constancy of bees, Bot. Rev., 16, 379—398 (1950).
268 . Grant V. Genetic and taxonomic studies in Gilia. II. Gilia capitata abro-
tanifolia, Aliso, 2, 361—373 (1952).
269 . Grant V. Genetic and taxonomic studies in Gilia. III. The Gilia tricolor
complex, Aliso, 2, 375—388 (1952).
270 . Grant V. Isolation and hybridization between Aquilegia formosa and
A pubescens, Aliso, 2, 341—360 (1952).
271 . Grant V. Cytogenetics of the hybrid Gilia millefoliataXachilleafolia. I. Va-
riations in meiosis and polyploidy rate as affected by nutritional and gene-
tic conditions, Chromosoma, 5. 372—390 (1952).
272 . Grant V. The role of hybridization in the evolution of the Leafystemmed
Gilias, Evolution, 7, 51—64 (1953).
273 . Grant V. Genetic and taxonomic studies in Gilia. IV. Gilia achillea folia,
Aliso, 3, 1—18 (1954).
274 . Grant V. Genetic and taxonomic studies in Gilia. VI. Interspecific relation-
ships in the Leafy-stemmed Gilias, Aliso, 3, 35—49 (1954).
275 . Grant V. Genetic and taxonomic studies in Gilia. V. Gilia clivorum, Aliso,
3, 19—34 (1954).
276 . Grant V. The genetic structure of races and species in Gilia, Advances in
Genetics, 8, 55-87 (1956).
277 . Grant V. Chromosome repatterning and adaptation, Advances in Genetics,
8, 89—107 (1956).
278 . Grant V The influence of breeding habit on the outcome of natural hybri-
dization in plants, Amer. Nat, 90, 319—322 (1956).
279 . Grant V. The plant species in theory and practice. In: The Species Problem,
ed E. Mayr, Amer. Assoc. Adv. Sci., Washington, D. C., 1957.
280 . Grant V. The regulation of recombination in plants, Cold Spring Harbor
Symposia Quant. Biol., 23, 337—363 (1958).
281 . Grant V. Natural History of the Phlox Family, M. Nijhoff, The Hauge,
1959.
282 . Grant V. The Origin of Adaptation, Columbia University Press, New York,
1963.
Литература
475
283 Grant V The biological composition of a taxonomic species in Gilia, Advan-
ces in Genetics» 12, 281—328 (1964).
284 Grant V The Architecture of the Germplasm, John Wiley, New York,
1964
'185 Grant V Genetic and taxonomic studies in Gilia XII. Fertility relation-
ships of the polyploid Cobwebby Gilias, Aliso, 5, 479—507 (1964).
286 Grant К Species hybrids and spontaneous amphiploids in the Gilia laci-
niata group, Heredity, 20, 537—550 (1965)
287 . Grant V Block inheritance oi viability genes in plant species, Amer. Nat.,
100, 591—601 (1966).
288 Grant V Linkage between viability and fertility in a species cross in Gilia,
Genetics, 54, 867—880 (1966)
289 Grant V Selection for vigor and fertility in the progeny of a highly sterile
species hybrid in Gilia, Genetics, 53, 757—775 (1966)
290 Grant к The selective origin oi incompatibility barriers in the plant genus
Gilia. Amer Nat, 100, 99—118 (19b6)
291 Grant V. lhe origin of a new species oi Gilia in a hybridization experiment,
Genetics, 54, 1189—1199 (1966)
292 Grant V. Linkage between morphology and viability in plant species, Amer.
Nat, 101, 125—139 (1967)
293 Grant V Plant Speciation, 1st ed, Columbia University Press, New York,
1971
294 Grant V. Genetics of Flowering Plants, Columbia Univ Press, New York,
1975
295 . Grant V Isolation between Aquilegia formosa and A pubescens. a reply
and reconsideration, Evolution, 30, 625—628 (1976)
296 Grant V Population structure in relation to macroevolution, Biol. Zentral-
_ blat\ 96, 129-139 (1977)
29/ Grant V Character coherence in natuial hybrid population in plants, Bot.
Gaz , 140, 443—448 (1979)
298 . Grant V Gene flow and the homogeneity of species populations, Biol Zen-
tralblatt, 99, 157—169 (1980)
299 . Grant V, Grant .4 Genetic and taxonomic species in Gilia VII The Wood-
land Ginas, Aliso, 3, 59—91 (1954).
300 Grant к , Grant A Genetic and taxonomic studies in Gilia. XI. Fertility
relationships of the diploid Cobwebby Gilias, Aliso, 4, 435—481 (1960)
301 Grant к , Grant К A Flower Pollination in the Phlox Family, Columbia
University Press, New York, 1965
302 Grant V, Grant К A Dynamics of clonal microspecies in cholla cactus,
Evolution, 25, 144—155 (1971)
303 . Grant V, Grant K. A Natural hybridization between the cholla cactus
species Opuntia spinosior and О versicolor, Proc Nat Acad Sci, 68,
1993—1995 (1971)
304 Grant V, Grant К A Hybridization and \ arialion in the Opuntia phaeacan-
tha group in Central Texas, Bot. Gaz, 140, 208—215 (1979)
305 Grant V , Grant К A Clonal microspecies of hybrid origin in the Opuntia
lindhetnieri group, Bot Gaz 141, 101—106 (1980)
306 Grant IF F Bullen Af 7?, Nettancourt D The cytogenetics of Lotus.
—' I Embryo-cultured interspecific diploid hybrids closely related to L corni-
culatus L, Canad J Genetics and Cytol 4, 105—128 (1962)
307 Gray A P. Mammalian Hybrids Commonwealth Agricultural Bureaux,
Farnham Royal, England, 1954
308 Gray A P. Bird Hybrids Commonwealth Abricultural Bureaux, Farnham
Royal England, 1958
309 . Greenleaf tt7. H. Sterile and fertile amphidiploids their possible relation to
the origin of Nicotiana tabacum, Genetics, 26, 301—324 (1941).
476 Литература
310 Grun Р. Cytogenetics studies of Poa. I. Chromosome numbers and morpho-
logy of interspecific hybrids, Amer. J. Bot4 41, 671—678 (1954).
ЗП. Grun P Early stages jn the formation of internal barriers to gene exchange
between diploid species of Solanum, Amer. J. Bot., 48, 79—89 (1961).
312. Grun P, Aubertin M, Radlow A. Multiple differentiation of plasmons of
diploid species of Solanum, Genetics, 47, 1321—1333 (1962).
313. Gulick J. T. Evolution, racial and habitudinal, Carnegie Inst, Washington
Publ, 25 (1905).
314. Gustafsson A. The origin and properties of the European (blackberry flora,
Hereditas, 28, 249—277 (1942) I
315. Gustafsson A. The genesis of the European blackberry flora. Lunds Univer-
sitets Arsskrilt 39(6), 1—200 (1943).
316. Gustafsson A. The constitution of the Rosa canina complex, Hereditas, 30,
405—428 (1944).
317. Gustafsson A Apomixis in higher plants. Lunds Universitets Arsskrift,
42—43, 1—370 (1946, 1947).
3L8. Gustafsson A. Polyploidy, life form and vegetative reproduction, Hereditas,
34, 1—22 (1948)
319. Gustafsson A., Hakansson A. Meiosis in some Rosa-hybrids, Botaniska No-
tiser, 1942, 331—343 (1942).
320. Gustafsson M. Evolutionary trends in the Atriplex. triangularis group of
Scandinavia. I. Hybrid sterility and chromosomal differentiation, Botaniska
Notiser, 126, 345—392 (1973)
321. Haartman J. Plantae Hybridae, Uppsala, 1751; Amoenitates Academicae,
Holm, 1764
322. Hagen C. W. A contribution to the cytogenetics of the genus Oenothera
with special reference to certain forms from South America, Indiana Univ.
Publ, Science Series, 16, 305—348 (1950).
323. Hagerup O. Uber Polyploidie in Beziehung zu Klima, Okologie und Phylo-
genie, Hereditas, 16,19—40 (1932).
324. Hagerup O. The spontaneous formation of haploid, polyploid and aneuploid
embryos in some orchids, Biologiske Meddelelser, 20, 1—22 (1947).
325. Hair J. В Biosystematics of the New Zealand flora 1945—1964, New. Zea-,
land J. Bot, 4, 559—595 (1966).
326. Hair J. B. The chromosomes of the Cupressaceae. I. Tetraclineae and Acti-
nostrobeae (Callitroideae), New Zealand J. Bot., 6, 277—284 (1968).
327. Hair J. В, Beuzenberg E J. Chromosomal evolution in the Podocarpaceae,
Nature, 181, 1584—1586 (1958).
328. Hair J. В, Beuzenberg E J High polyploidy in a New Zealand Poa, Natu-
re, 189, 160 (1961).
329. H&kansson A. Contributions to a cytological analysis of the species diffe-
rences of Godetia amoena and G. whitneyi, Hereditas, 33, 235—260 (1947).
330. H&kansson A. Meiosis and pollen mitosis in X-rayed and untreated spikelets
of Eleocharis palustris, Hereditas, 46, 325—345 (1954).
331. H&kansson A. Holocentric chromosomes in Eleocharis, Hereditas, 44, 531—
540 (1958).
332 Hall В M Genetic analysis of interspecific hybrids in the genus Bromus,
section Ceratochloa, Genetics, 40, 175—192 (1955)
333. Hall M T., Mukherjee A., Crowley IF. R. Chromosome numbers of culti-
vated junipers, Bot. Gaz., 140, 364—370 (1979)
334. Haller J R. Variation and hybridization in ponderosa and Jeffrey pines,
Univ. Calif. Publ. Bot., 34, 123—166 (1962).
335. Hamrick J L. Genetic variation and longevity. In: Topics in Plant Popula-
tion Biology, ed. O. Solbrig, Columbia University Press, New York, 1979.
336. Hanelt P. Polyploidie-Frequenz und geographische Verbreitung bei hoheren
Pflanzen, Biol. Rundschau, 4, 183—196 (1966).
Литература
477
337 Hanelt Р„ Mettin D. Cytosystematische Untersuchungen in der Arten-
gruppe um Vicia sativa L II Die Kulturpflanze (Berlin), 14, 137—161
(1966)
338. Harberd D J Observations on population structure and longevity of Festu-
ca rubra L, New Phytol., 60, 184—206 (1961)
339 Harberd D J. Some observations on natural clones in Festuca ovina. New
Phytol, 61, 85—100 (1962)
340 Hardin G The competitive exclusion principle, Science, 131, 1292—1297
(1960).
341. Hardin J. W. Studies in the Hippocastanaceae III. A hybrid swarm in the
buckeyes, Rhodora, 59, 45—51 (1957).
342 Hardin J W Studies in the Hippocastanaceae. IV. Hybridization in Aescu-
lus, Rhodora, 59, 185—203 (1957)
343. Hardin J U/. Hybridization and introgression in Quercus alba, J. Arnold
Arb, 56, 336—363 (1975).
344 Harding J., Allard R. W, Smeltzer D G. Population studies in predominan-
tly self pollinated species IX Frequency-dependent selection in Phaseolus
lunatus. Proc Nat. Acad Sci, 56, 99—104 (1966)
345. Harlan J R Natural breeding structure in the Bromus carinatus complex
as determined by population analysis, Amer. J. Bot, 32, 142—148 (1945)
346. НаНап J R Apomixis in side-oats grama, Amer. J. Bot, 36, 495—499
(1949).
347 Harlan J ft, de Wet J M J Toward a rational classification of cultivated
plants, Taxon 20, 509—517 (1971)
348 Harlan J ft, Zohary D Distribution of wild wheats and barley, Science,
153, 1074—1080 (1966).
349. Harland S C The genetical conception of the species, Biol Rev, 11, 83—
112 (1936)
350. Harper J L Biological flora of the British Isles Ranunculus acris L, Ra-
nunculus repens L„ Ranunculdus bulbosus L, J Ecol, 45, 289—342
(1957)
351 Harper J L. Population Biology of Plants, Academic Press, New York,
1977
352. Harper J L, White J. The demography of plants, Ann Rev Ecol Syst, 5,
419—463 (1974).
353 Haselton S C. Ep phyllum Handbook, Abbey Garden Press, Pasadena, Ca-
lif. 1951.
354 Hawkes J G, H;erting J P The Potatoes of Argentina, Brazil, Paraguay
and Uruguay. A Biosystematic Study, Clarendon Press, Oxford, 1969
355. Hayata В The natural classification of plants according to the dynamic
system, Icon. Plant. Formos., 10, 97—234 (1921)
356 Hayman В 1, Mather K, The progress of inbreeding when homozygotes
are at a disadvantage, Heredity, 7, 165—183 (1953)
357. Hecht A Cytogenetic studies of Oenothera subgenus Raimannia, Indiana
Univ Publ, Science Series, 16, 255—304 (1950)
358 Heckard L R Taxonomic studies in the Phacelia tnagellanica polyploid
complex with special reference to the California members, Univ. Calif.
Publ Bot., 32, 1—126 (1960)
359 Hegi G Illustrierte Flora von Mittel-Europa, 7 vols Carl Hanser, Munich,
1909-1931.
360 Heilborn О Chromosome numbers and dimensions, species formation and
phylogeny in the genus Carex, He,editas, 5, 129—216 (1924)
361. Heilborn О Aneuploidy and polyploidy in Carex, Svensk Bot Tidskr, 26,
137—146 (1932).
362 Heilborn О On the origin and preservation of polyploidy, Hereditas, 19,
233—242 (1934)
478
Литература.
363. Heilborn О. Chromosome studies in Cyperaceae. Ill—IV, Hereditas, 25,
224—240 (1939).
364. Heiser С. B. Hybridization between the sunflower species Helianthus an-
nuus and H. petiolaris, Evolution, 1, 249—262 (1947).
365. Heiser С. B. Natural hybridization with particular reference to introgression,
Bot. Rev., 16, 645—687 (1949).
366. Heiser С. B. Study in the evolution of the sunflower species Helianthus
annuus and H. bolanderi, Univ Calif. Publ. Bot., 23, 157—208 (1949).
367. Heiser С. B. A comparison of the flora as a whole and the weed flora of
Indiana as to polyploidy and growth habits, Proc. Indiana Acad. Sci., 59.
64-70 (1950).
368. Heiser С. B. Hybridization in the annual sunflowers* Helianthus annuusx
XH. debilis var. cucutnerifolius, Evolution, 5, 42—51 (1951).
369. Heiser С. B. Hybridization in the annual sunflowers. Helianthus annuusX
XH. argophyllus, Amer. Nat., 85, 65—72 (1951).
370 Heiser С В Variation and subspeciation in the common sunflower, Helian-
thus annuus, Amer. Midland Nat., 51, 287—305 (1954).
371 Heiser С. В The North American sunflowers (Helianthus), Memoirs Torrey
Bot. Club, 22(3), 1—218 (1969).
372. Heiser С. B. Introgression re-examined, Bot. Rev, 39, 347—366 (1973).
373. Heiser С. B. The Sunflower, University of Oklahoma Press, Norman, Okla,
1976.
374. Heiser С. B, Martin W. C., Smith D M Species crosses in Helianthus.
I. Diploid Species, Brittonia, 14, 137—147 (1962).
375. Heiser С. B., Smith D M. The origin of Helianthus multiflorus, Amer. J.
Bot, 47, 860—865 (1960).
376. Herbert W. On the production of hybrid vegetables, with the result of many
experiments made in the investigation of the subject, Trans. Hort. Soc. Lon-
don, 4, 15—50 (1820).
377. Hiesey W. M., Nobs M. A., Bjorkman O. Experimental studies on the na-
ture of species. V. Biosystematics, genetics and physiological ecology of the
Erythranthe section of Mimulus, Carnegie Inst. Wash. Publ., 628, 1971.
378. Hiorth G. Eine Serie multipled Allele fur Blutenzeichnungen bei Godetia
amoena, Hereditas, 26, 441—453 (1940)
379. Hiorth G Zur Genetik und Systematik der amoena-Grnppe der Gattung
Godetia. Zeitschr. ind. Abslammungs- u Vererbengslehre, 80, 289—349
(1942).
380. Holler H. C„ Abrahamson IF. G. Seed and vegetative reproduction in re51
lation to density in Fragaria virginiana (Rosapeae), Amer. J. Bot., 64,
1003—1007 (1977).
381. Hollingshead L. A lethal factor in Crepis effective only in an interspecific
hybrid, Genetics, 15, 114—140 (1930).
382. Hopper S. D., Burbidge A. H. Assortative pollination by red wattlebirds in
a hybrid population of Anigozanthos Labill. (Haemodoraceae), Austral. J.
Bot, 26, 335—350 (1978).
383. Hovanitz W. Increased variability in populations following natural hybri-
dization. In: Genetics, Paleontology and Evolution, ed. G. L. Jepsen, Price-
ton University Press, Princeton, N. J., 1949.
384. Howard H. W. Potato cytology and genetics, Bibliographia Genetica, 19,
87—216 (1952—1959).
385. Hubbs C., Delco E A. Mate preference in males of four species of gambu-
siine fishes. Evolution, 14, 145—152 (1960).
386. Hubbs C. L. Hybridization between fish species in nature, Systematic Zool.,
4, 1—20 (1955).
387. Hubbs C. L., Miller R. R Mass hybridization between two genera of cypri-
nid fishes in the Mohave Desert, California, Papers Michigan Acad. Sci.
Arts and Letters, 28, 343—378 (1943).
-Литература
479
388 Hulten Е Stellarla longipes Goldie and its allies, Botaniska Notiser, 251—
270 (1943)
389 Hulten E The Cerastium alpinum complex A case of world-wide in tro-
gressive hybrid zation, Svensk Bot. Tidskr., 50, 411—495 (1956)
390 Hurst С C Embryo sac formation in diploid and plvploid species of Ro-
seae, Proc. Roy. Soc., B, 109, 126—148 (1931)
391. Hutchinson J. B, Stephens S G. The Evolution of Gossypium, Oxford
University Press, London, 1947.
392. Huxley J S Evolution. The Modern Synthesis, George Allen and Unwin,
London, 1942
393. Huziwara Y. Chromosomal evolution in the subtribe Asterinae, Evolution,
13, 188—193 (1959).
394. Iman A. G, Allard R. W Population studies in predominantly self-pollina-
ted species. VI. Genetic variability between and within natural populations
of wild oats from differing habitats in California, Genetics, 51, 49—62
(1965).
395. Jackson R C New low chromosome number for plants, Science, 128,
1115—1116 (1957).
396 Jain S K. Male sterility in flowering plants, Bibliographia Genetica, 18,
101—166 (1959)
397. Jain S. K. Cytogenetics of rye (Secale spp ) Bibliographia Genetica, 19,
1—86 (1960).
398 Jain S К Population structure and the effects of breeding system In Crop
Genetic Resources for Today and Tomorrow, ed О. H Frankel and
J G Hawkes, Cambridge University Press, New York, 1975
399 Jain S К Adaptive strategies polymorphism, plasticity and homeostasis.
In Topics in Plant Population Biology, ed О Solbrig, Columbia Univer-
sity Press, New York, 1979
400 Jain S R, Allard R. IF. Population studies in predominantly self-pollina-
ted species. I Evidence for hctcrozygote advantage in a closed population
of barley, Proc. Nat. Acad. Sci. 46, 1371—1377 (1960)
401 Jain S R, Allard R W The nature and stability of equilibria under opti-
mizing selection. Proc. Nat Acad Sci, 54, 1436—1443 (1965)
402. Jain S R, Bradshaw A D Evolutionary divergence among adjacent plant
populations I The evidence and its theoretical analysis, Heredity, 21,
407-441 (1966).
403 Jain S R, Jain К В L The progress of inbreeding in a pedigree-bred po-
pulation of barley, Euphytica, 11, 229—232 (1962)
404 Jain S R, Marshall D. R Population studies in predominantly self-pollina-
ted species. X. Variation in natural populations of Avena fatua and A bar-
bata, Amer. Nat, 101, 19—33 (1967).
405 Jepson U7 L The Trees of California. Associated Students Store, Berkeley,
Calif., 1923
406. John В, Lewis К R. Genetic speciation in the grasshopper Eyprepocnemis
plorans, Chromosome. 16. 308—344 (1965)
407 John В , Lewis R. R The Meiotic System, Protoplasmatologia, Springer-
Verlag. New York, 1965
408 John В, Lewis R. R. Chromosome variability and geographic distribution
in insects, Science, 152, 711—721 (1966)
409. Johnson A W, Packer J G Polyploidy and environment in arctic Alaska,
Science, 148, 237—239 (1965).
410. Johnson A IF, Packer J G, Reese G. Polyploidy, distribution and environ-
ment In* The Quaternary of the United States, ed H E. Wright and
D. G. Frey, Princeton University Press, Princeton, N. J, 1965.
411. Johnson B. L. Cyto-taxonomic studies in Oryzopsis, Bot Gaz., 107, 1—32
(1945).
412 Johnson G. В Enzyme polymorphism- genetic variation in the physiological
480
Литература
phenotype In- Topics-in Plant Population Biology, ed. O. Solbrig, Colum-
bia University Press, New York, 1979.
413. Johnson L. A S., Briggs B. G. Evolution in the Proteaceae, Australian J.
Bot., 11,21—61 (1963).
414. Johnson £. A. S. On the Proteaeceae — the evolution and classification of
a southern family, Bot. J. Linnean Soc. (London), 70, 83—182 (1975).
415. Johnston J. S., Heed W. B. Dispersal of desert-adapted Drosophila', the
saguaro-breeding D mgrospiracula, Amer. Nat., 110, 629—651 (1976).
416. Jones D. F. Genic and cytoplasmic control of pollen abortion in maize,
Brookhaven Symp. Biol., 9, 101—112 (1956).
417. Jones E. E. Pollinator constancy as a pre-pollination isolating mechanism
between sympatric species of Cercidium, Evolution, 32, 189—198 (1978).
418. Jones S В Cytogenetics and affinities of Ver onia (Compositae) from the
Mexican highlands and eastern North America, Evolution, 30, 455—462
(1976).
419. Jonsell B. Linnaeus’s views on plant classification and evolution. Botaniska
Notiser, 131, 523—530 (1978).
420. Jordan D. S. The origin of species through isolation, Science, 22, 545—562
(1905).
421. Jordan R. On mechanical selection and other problems, Novit. Zool., 3,
426-525 (1896).
422. Jowett D Population studies on lead-tolerant Agrostis tenuis, Evolution,
18, 70—81 (1964).
423. Jukes T. H. Molecules and Evolution, Columbia University Press, New York,
1966.
424. Ramemoto H., Shlndo R. Meiosis in interspecific and intergeneric hybrids
of Vanda, Bot. Gaz., 125, 132—138 (1964)
425. Rannenberg L. W.. Allard R W Population studies in predominantly self-
pollinated species. VIII. Genetic variability in the Festuca microstachys
complex, Evolution, 21, 227—240 (1967).
426 Rapadia Z J, Gould F. W. Biosystematic studies in the Bouteloua curti-
pendula complex IV. Dynamics of variation in B. curtipendula var. caespi-
tosa, Bull. Torrey Bot. Club, 91, 465-478 (1964).
427. Карпиченко Г. Д. Полиплоидные гибриды Raphamus L.X Brassica olera-
cea L. Труды бюро прикл. бот., генет. и селекции, 17, 305—410 (1927).
428. Reck D. D. Trends in systematic botany, Survey of Biol Progress, 3, 47—
107 (1957).
429. Reep E. Interspecific hybridization in Ribes, Genetica, 33, 1—23 (1962).
430. Remperman J. A., Barnes В. V. Clone size in american aspens, Canadian
J. Bot., 54, 2603—2607 (1976).
431. Rerner A. The Natural History of Plants, 2 vols. Translation, Blackie and
Son, London, 1894—1895.
•[Имеется перевод: Кернер А. «Форма и жизнь растения», т. I, т-во «Про-
свещение», Санкт-Петербург, 1899; «История растений», т II, 1902]
432. Rerster Н. W., Levin D. A. Neighborhood size in Lithospermum caroliniense,
Genetica, 60, 577—587 (1968)
433. Rhoshoo T. N. Polyploidy in gymnosperms, Evolution, 13, 24—39 (1959).
434. Rhush G. S. Cytogenetics of Aneuploidy, Academic Press, New York, 1973.
435. Riang Y. T. Floral structure, hybridization and evolutionary relationships of
two species of Mimulus, Rhodora, 75, 225—238 (1973).
436 Rihara H Cytologische und gcnetische Studien bei wichigen Getreidearten
mit besonderer Rucksicht auf das Verhalten der Chromosomen und die Ste-
rilitat in den Bastarden, Mem Coll Sci. Kyoto Univ, B. 1, 1—200 (1924).
437 Rihara H Considerations on the evolution and distribution of Aegilops
species based on the analyzer-method, Cytologia, 19, 336—357 (1954).
438 Rihara H, Ono T. Chromosomenzahlen und systematische Gruppierung der
j?«?nex-Arten, Zeitschr. Zellforsch. u mikr. Anatomie, 4, 475—481 (1926).
439
440.
441
442
443
444.
445
446
447.
448
449.
450
451
452
453
454
455
456
457
458
459
460
461
462
463
464.
465.
Литература 484
King J. C Interspecific relationships within the guarani group oi Droso-
phila, Evolution, 1, 143—153 (1947).
King M, Wilson A. C Evolution al two levels in humans and chimpanzees.
Science, 188, 107—116 (1975).
Klekowski E J Reproductive biology of the Pteridophyta II Theoretical
considerations, Bot. J. Linnean Soc , 62, 347—359 (1969)
Klekowski E J, Baker H G Evolutionary significance oi polyploidy in
the Plenodophyta, Science, 153, 305—307 (1966).
Knight G R, Robertson A , Waddington С H Selection for sexual isola-
tion within a species, Evolution, 10, 14—22 (1956).
Kblreuter J G Voriaufige Nachricht von einigen das Geschlecht der Pflan-
zen betreffenden Versuchen und Bcobachtungen. Reprint, 1893 In Ost-
wald’s Klassiker, Leipzig, 1761—1766.
Koopman K. F Natural selection for reproductive isolation between Dro-
sophila pseudoobscura and Drosophila persimilis, Evolution, 4, 135—148
(1950).
Kruckeberg A R Intraspecific variability in the response oi certain nati\e
plant species to serpentine soil, Amer. J. Bot, 38, 408—419 (1951)
Kruckeberg A. R. Chromosome numbers in Silene (Caryophvllaceae) I.
Madrono, 12, 238—246 (1954).
Kruckeberg A. R. Interspecific hybridization of Silene, Amer J Bot, 42,
373—378 (1955).
Kruckeberg A R Variation in fertility of hybrids between populations of
the serpentine species, Streptanthus glandulosus Hook, Evolution, 11, 185—
211 (1957)
Kruckeberg A. R Chromosome numbers in Silene (Caryophylaccae) II Mad-
rono, 15, 205—215 (1960)
Kruckeberg .4 R Art ficial crosses of western North American Silenes, Brit-
tonia, 13, 305—333 (1961).
Kruckeberg .4 R. The implications of ecology for plant systematics, Taxon,
18, 92—120 (1969)
Kugler H Bliitcnokologic, 2nd ed, Gustav Fischer, Stuttgart, 1970
Kurabayas'm M Evolution and variation in Japanese species of Trillium,
Evolution, 12, 286—310 (1958)
Kurabayashi M, Lewis II, Raven P H A comparative study of mitosis in
the Onag-aceae Amer J Bot, 49, 1003—1026 (1962).
Kwak M Af Poll.nation, hybridization and ethological isolation of Rhinan-
thus minor and R serotinus (Rhinanthoidcae Scrophulariaceae) by bum-
blebees (Bombus Latr), Taxon, 27, 145—158 (1978).
Ladiges P. У. Variation in drought resistance in adjacent edaphic popula-
tions of Eucalyptus vlminalis Labill, Austral J Ecol, 1, 67—76 (1976)
Lamprecht H Die Artgrenze zuischen Phaseolus vulgaris L und muliiflo-
rus Lam, Hc-cditas, 27. 51 —175 (1941)
Laniprecht H Die genisch plasmatische Grtindlage der Artbarriere, Agr
Hort Genet, 2, 75—142 (1944)
Latimer H A study of the breeding barrier between Gilia australis and
Gilia splendens, Ph D thesis, Claremont Graduate School, Claremont,
Calif, 1958
Lenz L W A revision of the Pacific coast irises, Aliso, 4. 1—72 (1958).
I enz L U7 Hybridization and speciation in the Pacific coast irises, Aliso,
4. 237—309 (1959).
Lenz L W Iris tenuis S Wats, a new transfer to the subsection Evansia,
Aliso, 4, 311—319 (1959)
Lenz L W, Wimber D E Hybridization and inheritance in orchids In-
The Orchids, ed C Withne”, Ronald Press, New York, 1959
Lesins К Alfalfa, lucerne In Evolution of Crop Plants, ed. N W Simmonds,
Longman, London and New York, 1976
31—45
482
Литература
466. Levan A. Cytological studies in the Allium pardculatum group, Hereditas,
23, 317—370 (1937).
467. Levan A. Polyploidy in flax, sugar beets and timothy, Hereditas, Suppl.
Vol, 46—47 (1949).
468. Levin D. A. Chromatographic evidence of hybridization and evolution in
Phlox maculata, Amer. J. Bot., 53, 238—245 (1966).
469. Levin D A Hybridization between annual species of Phlox', population
structure, Amer. J. Bot., 54, 1122—1130 (1967).
470 Levin D A. The genome constitutions of eastern North American Phlox
amphiploids, Evolution, 22, 612—632 (1968).
471. Levin D. A. Assortative pollination in Lythrum. Amer. J. Bot, 57, 1—5
(1970)
472 Levin D. A. Pest pressure and recombination system in plants, Amer. Nat,
109, 437—451 (1975)
473 Levin D. A. Genic heterozygosity and protein polymorphism among local
populations of Oenothera biennis, Genetics, 79, 477—491 (1975).
474 Levin D. A. The origin of isolating mechanisms in flowering plants, Evol.
Biol., 11, 185—317 (1978).
475. Levin D A The nature of plant species. Science, 204, 381—384 (1979).
476 Levin D. A, ed. Hybridization. An Evolutionary Perspective, Dowden, Hut-
chinson and Ross, Stroudsburg, Pa, 1979.
477. Levin D A, Howland G., Steiner E Protein polymorphism and genic hete-
rozygosity in a population of the permanent translocation heterozygote,
Oenothera biennis, Proc. Nat Acad. Sci, 69, 1475—1477 (1972).
478 Levin D. A., Kerster H. W. An analysis of interspecific pollen exchange in
Phlox, Amer. Nat., 101, 387—400 (1967).
479. Levin D. A., Kerster H. W. Natural selection for reproductive isolation in
Phlox, Evolution, 21, 679—687 (1967).
480. Levin D. A., Kerster H. W. Local gene dispersal in Phlox, Evolution, 22,
130—139 (1968).
481 Levin D A, Kerster H. W. Density-dependent gene dispersal in Liairis,
Amer. Nat, 103, 61—74 (1969).
482. Levin D. A, Kerster H W. Gene flow in seed plants, Evol Biol., 7, 139»—
220 (1974).
483. Levin D. A., Ritter K-> Ellslrand N. C. Protein polymorghism in the narrow
endemic Oenothera organensis, Evolution, 33, 534—542 (1979).
484. Levin D. A., Schaal B. A. Corolla color as an inhibitor of interspecific hyb-
ridization in Phlox, Amer. Nat., 104, 273—283 (1970).
485. Levin D. A., Wilson A. C. Rates of evolution in seed plants: net increase
in diversity of chromosome numbers and species numbers through time,
Proc. Natl. Acad. Sci, 73, 2086—2090 (1976).
486. Levins R. Evolution in Changing Environments. Princeton University
Press, Princeton, N J., 1968.
487. Levy M, Levin D. A. Genic heterozygosity and variation in permanent
translocation heterozygotes of the Oenothera biennis complex, Genetics, 79,
493—512 (1975).
488 Lewis D. Physiology of incompatibility in plants. III. Autopolyploids, J.
Genetics, 45, 171—185 (1943).
489. Lewis D. The incompatibility sieve for producing polyploid, J. Genetics, 45,
261—264 (1943).
490. Lewis D. The genetic integration of the breeding system. In: Reproductive
Biology and Taxonomy of Vascular Plants, ed. J. G. Hawkes, Pergamon
Press, Oxford, 1966.
491. Lewis H. The mechanism of evolution in the genus Clarkia, Evolution, 7,
1—20 (1953).
492. Lewis H. Chromosome phylogeny and habitat preference of Clarkia, Evolu-
tion, 7, 102—109 (1953).
Литература
483
493 Lewis Н Catastophic selection as a factor in speciation, Evolution, 16_
257—271 (1962).
494 Lewis H Speciation in flowering plants, Science, 152, 167—172 (1966).
495. Lewis H. Epling C Delphinium gypsophilum, a diploid species of hybrid
origin, Evolution, 13, 311—525 (1959).
496 Lewis H, Lewis M. The genus Clarkia, Univ Calif Publ Bot, 20, 241 —
392 (1955).
497. Lewis H, Raven P, H. Rapid evolution in Clarkia, Evolution, 12, 319—
336 (1958).
498 Lewis H, Raven P H, Venkatesh C S, Wedberg H L Obseivations of
meiotic chromosomes in the Onagraceae, Aliso, 4, 73—86 (1958)
499 Lewis H, Roberts Al. R. The origin oi Clarkia Ungulata, Evolution, 10,.
126—138 (1956).
500 Lewis H, Szweykowski J The genus Gayophylum (Onagraceae), Brittonia,
16, 343—391 (1964).
501. Lewis K. R. John B. Hybridisation in a wild population of Eleocharis pa-
lustris, Chromosoma. 12, 433—448 (1961).
502 Lewis К R, John В Chromosome Marker, Little, Brown and Co, Boston,
Mass., 1963
503 Lewis IV H Chromosomal drift, a new phenomenon in plants, Science, 168,
1115—1116 (1970).
504 Lewis W. H Temporal adaptation correlated with ploidy in Claytonia vir-
ginica. Systematic Bot, 1 340—347 (1976)
505 Lewis TV. H. ed Polyploidy; Biological Relevance, Plenum Press, New York,
and London, 1980
506 Lewis TV. H, Oliver R L, Luikart T J. Multiple genotypes in individuals
of Claytonia uirginica, Science. 172, 564—565 (1971)
507. Lewis TV. H, Semple J. C Geography of Claytonia virginica cvtotvpes,
Amer J Bot, 64, 1078—1082 (1977).
508 Lewontin R C The Genetic Bas s of Evolutionary Change, Columbia Uni-
versity Press, New York, 1974 [Имеется перевод Левонтии P. Гепетиче
ские основы эволюции. —М «Мир», 1978 ]
509 Lima-de-Faria A Genetics, origin and evolution of kinetochores, Hereditas,
35, 422—444 (1949)
510 Lindsay D TV. Vickery R К Comparatne evolution in Mimulus guttaius
of the Bonneville basin? Evolution, 21, 439—456 (1967)
511 Lindsirom E W Genetics oi polyploidy, Bot Rev, 2, 197—215 (1936)
512 Linnaeus C Critica Botanica, Leiden, 1737
513 Linnaeus C Philosophia Botamca Stockholm. 1751
514 Linnaeus C Disquisitio de Scxu Plantarum St Petersburg, 1760, Amoenita-
tes Academicae. Erlangen, 1790
515 Unnaeus C Genera Plantarum. 6th ed . Vienna 1767
516 Little E L , Pauley S S A natural hybrid between black and white spruce
in Minnesota, Amer Midland Nat, 60, 202—211 (1958)
517. Lokki J., Saura A Polyploidy in insect evolution In- Polyploidy, Biologi-
cal Relevance, ed W H Lewis, Plenum Press New York, London, 1980
518 Lorkotic Z Some peculiarities of spa dally and sexually restricted gene
exchange in the Erebta tyndarus g'oup, Cold Spring Harbor Svmp Quant
Biol. 23. 319—325 (1958)
519 Lotsy J P. Evolution bv Means of Habridization, M Nijhoff, The Hague,
1916
520 Lotsy J P Species or Linneon, Genetica, 6, 487—506 (1925)
521 Lotsy J P On the species of the taxonomist in its relation to evolution,
Genetica, 13, 1—16 (1931)
522 Love A Cvtogenetic studies on Rumex, subg Acetosella, Hereditas, 30, 1 —
136 (1944)
523 Love A Preparatory studies lor breeding Icelandic Poa irrigata, Hereditas,
38, 11—32 (1952).
31*
484 Литература
524. Love A. Subarctic polyploidy, Hereditas, 39, 113—124 (1953).
525 Love A. The biological species concept and its evolutionary structure, Ta-
xon, 13, 33—45 (1964).
526 Love A., Kapoor В. M. An alloploid Ophioglossum, The Nucleus, 9, 132—
138 (1966)
527. Ldve A, Kapoor В. M The highest plant chromosome number in Europe
Svensk Bot, Tidskrift, 61, 29—32 (1967).
528. Love A, Love D. The geobotanical significance of polyploidy. I. Polyploidy
and latitude, Portugalae Acta Biologica, Suppl, vol. 273—352 (1949).
529. Love A, Love D The geo-botanical significance of polyploidy Proc. 6th In-
ternal. Grasslands Congress, 240—246 (1953).
530 Love A, Love D. Arctic polyploidy, Proc. Genetics Soc. Canada, 2, 23—27
(1957).
531. Love A., Love D. Polyploidy and altitude: Mt Washington, Biol. Zentral-
blatt. Suppl. vol, 307—312 (1967).
532. Love A., Love D. Cytotaxonomical Atlas of the Arctic Flora, J. Cramer,
Vaduz, 1975.
533. Ldve A., Ldve D, Pichi Sermolli R. E. G. Cytotaxonomical Atlas of the
Pteriodophyta, J. Cramer, Vaduz, 1977.
534 Love A., Ldve D, Raymond M. Cytotaxonomy of Carex section Capillares,
Canad. J. Bot., 35, 715—761 (1957).
535 MacKey J Neutron and X-ray experiments in wheat and a revision of the
speltoid problem, Hereditas, 40, 65—180 (1954)
536. McLeod M Q. A new hybrid fleshy-fruited prickly-pear in California, Mad-
rono, 23, 96-98 (1975).
537. McMinn H. E. The importance of field hybrids in determining species in
the genus Ceanothus, Proc. Calif. Acad. Sci., 25, 323—356 (1944).
538. McNaughton I. H., Harper J. L. The comparative biology of closely related
species living in the same area II. Aberrant morphology and a virus-like
syndrome in hybrids between Papaver rhoeas L and P. dubium L., New
Phytol., 59, 27—41 (1960).
539. McNaughton I. H, Harper J. L The comparative biology of closely related
species living in the same area. III. The nature of barriers isolating sym-
patric populations of Papaver dibium and P lecoquii, New Phytol., 59,
129-137 (1960).
540. McNeilly T., Antonovics J. Evolution in closely adjacent plant populations.
IV. Barriers to gene flow, Heredity, 23, 205—218 (1968).
541. Macior L W The pollination ecology of Pedicularis (Scrophulariaceae) in
the Sierra Nevada of California, Bull. Torrey Bot. Club, 104, 148—154
(1977).
542 Magoon M L, Cooper D. C., Hougas R. W. Cytogenetic studies of some
diploid Solanums section Tuber arium, Amer. J. Bot, 45, 207—221 (1958).
543. Maguire В Apomixis in the genus Clusia (Clusiaceae) — a preliminary
report, Taxon, 25, 241—244 (1976).
544. Maheshwari P. The male gametophyte of angiosperms, Bot. Rev., 15, 1—
75 (1949).
545 Malecka J. Studies in the mode of reproduction of the diploid endemic spe-
cies, Taraxacum pieninicum Pawl, Acta Biological Cracoviensia, 4, 25—41
(1961).
546. Malecka J. Processes of degeneration in the anthers’ tapetum of two male-
sterile species of Taraxacum, Acta Biol. Cracoviensia, Bot., 14, 1—10
(1971).
547. Malecka J. Cyto-taxonomical and embryological investigations on a natu-
ral hybrid between Taraxacum kok-saghz Robin and T. officinale Web. and
their putative parent species, Acta Biol. Cracoviensia, Bot., 14, 179—197
(1971).
548 Malecka J Problems of the mode of reproduction in microspecies of Tara-
Литература
485
549
550
551
552.
553
554.
555
556
557
558
559
560
561.
562.
563
564
565
566.
567
568
569
570
571
572
573
574
xacum palusiria Dahlstedt Acta Biol Cracovicnsia, scr Bot, 16, 37—84
(1973)
Malhetros N. De Castro D, Camara A Chromosomas sem centromere lo-
calizado О caso da Luzula purpurea Link, Agronomid Lusitana, 9, 51—71
(1947)
Malhetros Garde N, Garde A Agmatoploidia no genero Luzula DC, Gene-
tica Iberica, 3, 155—176 (1951)
Manglesdorf P C The mutagenic effect oi hybridizing ma ze and teosinte,
Cold Spring Harbor Symp Quant Biol, 23, 409—421 (1958)
Manton 1 The problem of Biscutella laevigata L. Zeitschr ind Abstamm,
Vererbungslehre, 67, 41—57 (1934)
Manton I The problem of Biscutella lue.ugaia L II The evidence from
meiosis, Ann Bot, n s, 1, 439—462 (1937)
Manton 7 Problems of Cytology and Evolution in the Pteridophyta, Cam-
bridge University Press. Cambridge, 1950
Manton 7 Cyto ogy of Polypodium in America, Nature, 167, 37 (1951)
Manton I The concept oi the aggregate species In Systemahcs of Today,
ed О Hedberg, Uppsala, 1958
Marsden-Jones E M Th; genetics of Geum intermedium Willd hand Ehih
and its back-crosses J Genetics, 23, 377—395 (1930)
Mason H L Evidence for the genetic submergence of Pinus remoru'a In
Genetics, Paleontology and Evolut on, ed G L Jepsen, Princeton Univer-
sity Press, Princeton, N J , 1949
Mason H L Taxonomy, svstematics botanv and biosystematics, Madrono,
10, 193—208 (1950)
Mather К Species crosses in Antirrhinum I Genetic isolation of the species
majus, glutinosum. and orontium. Heredity, 1, 175—186 (1947)
Mather Polvmorph’sni as an outcome of disruptive selection, Evolution,
9,52—61 (1955)
Mather К B-eeding systens and response to selection In Reproductive
Biology and Taxonomy o' Vasctlar Plants, ed J G Hawkes, Pcrgamon
Press, Oxford 1966
Matti eld J Uber hvbndogene Sippen der Tannen, Bibliotheca Botanica,
vol 25, 1960
Mayr E Systematics and the Origin of Species, Columbia University
Press New York, 1942
Mayr E Change oi genetic environment and evolution In Evolution as a
Process, ed J Huxlev, George Allen and Unvin, London, 1954
Mayr E The species as a systematic and as a biological problem In:
Biology Colloquium iOregon State College, Corvallis, Oregon), 1955
Mayr E Karl Jordan’s contribution to current concepts in systematics and
evolution, T-ans Royal Entmol. Soc (London), 1955, 45—66 (1955).
Mayr F Species concepts and definitions In The Species Problem, ed
E Mayr, Amer Assoc Adv Sci, Washington, 1957
Maur E Animal Species and Evolution, Harvard University Press, Cambrid-
ge, Mass , 1963
Mayr E Populations, Species and Evolution, Harvard Umv Press, Cam-
bridge, Mass, 1970. [Имеется перевод Майр Э. Популяции, вилы и эво-
люция — М «Мир» 1974 ]
Mayr Е Evolution and the Diversity of Life Selected Essays, Harvard
University Pres< Cambridge, Mass , 1976
Maynard Smith J Sympatric speciation, Amer Nat, 100, 637—650 (1966).
Maze J Past hybridization between Quercus macrocarpa and Quercus gam-
belii, Brittonia, 20, 321—333 (1968)
Meghtsch P A On the nature of the species, Systematic Zool, 3, 49—65
(1954).
486
Литература
575. Melander У. A new giant Populas tremula in Norrbotten, Hereditas, 24,
189—194 (1938).
576. Mettin D., tianelt P. Ober Speziationsvorgange in der Gatting Vida L.,
Kulturpflanze, 21, 25—54 (1973).
577. Michaelis P. Entwicklungsgeschichtlich-genetische Untersuchungen an Epi-
lobium II. Die Bedeutung des Plasmas ftir die Pollenfertilitat des Epilo-
bium luteum-hirsutum Bastardes, Zeitschr. ind. Abstammungs- u. Verer-
bungslehre, 65, 1—71; 353—411 (1933).
578. MUlicent E., Thoday J. M. Effects of disruptive selection. IV. Gene flow
and divergence, Heredity, 16,199—217 (1961).
579. Mirov N. T. The Genus Pinus, Ronald Press, New York, 1967.
580 Moens P. В Chiasma distribution and the segregation of markers in chro-
mosome 2 of the tetrapioid tomato, Genetics, 49, 123—133 (1964).
581. Mohamed A. H., Gould F. W Biosystematic studies in the Bouteloua curti-
pendula complex. V. Megasporogenesis and embryo sac development, Amer.
J. Bot., 53, 166—169 (1966).
582 Molisch H. The Longevity of Plants, Translation, E. H. Fulling, New York,
1938
583. Moore R. J., ed. Index to Plant Chromosome Numbers 1967—1971. Inter-
net. Bureau Plant Taxon, Utrecht, 1973.
584. Moore W. S. An evaluation of narrow hybrid zones in vertebrates, Quart
Rev. Biol., 52, 263—277 (1977).
585. Mosquin T. Toward a more useful taxonomy for chromosomal races, Brit-
tonia, 18, 203—214 (1966).
586. Mosquin T Evidence for autopolyploidy in Epilobium angustlfolium (Ona-
graceae), Evolution, 21, 713—719 (1967).
587. Mosquin T„ Small E. An example of parallel evolution in Epilobiutn (Ona-
graceae), Evolution, 25, 678-682 (1971).
588. Muldal S. The chromosomes of the earthworms I The evolution of poly-
ploidy, Heredity, 6, 55—76 (1952).
589. Muller С. H. The oaks of Texas, Contrib. Texas Research Foundation, 1(3),
21—323 (1951)
590 Muller С. И. The significance of vegetative reproduction in Quercus, Mad-
rono, 11, 120—137 (1951).
591. Muller С H. Ecological control of hybridization in Quercus'. a factor in
the mechanism of evolution, Evolution, 6, 147—161 (1952).
592. Muller С. H. The role of chemical inhibition (allelopathy in vegetational
composition, Bull. Torrey Bot Club, 93, 332—351 (1966).
593. Muller С. H., Quercus L In Manual of the Vascular Plants of Texas.
D. S. Correll and M. C. Johnston. Texas Research Foundation, Renner, Te-
xas, 1970.
594. Muller H. J. Why polyploidy is rarer in animals than in plants, Amer. Nat.,
59, 346—353 (1925).
595. MUntzing A. Outlines to a genetic monograph of the genus Galeopsis, He-
reditas, 13, 185—341 (1930).
596. MUntzing A. Uber Chromosomenvermehrung in Ga/eopsZs-Kreuzungen und
ihre phylogenetische Bedeutung, Hereditas, 14, 153—172 (1930).
597. MUntzing A. Cyto-genetic investigations on synthetic Galeopsis tetrahit,
Hereditas, 16, 105—154 (1932).
598. MUntzing A. Chromosome fragmentation in a Crepis hybrid, Hereditas,
19,284—301 (1934).
599. MUntzing A. The evolutionary significance of autopolyploidy, Hereditas, 21,
263-378 (1936).
600 MUntzing A The chromosomes of a giant Popidus tremula, Hereditas, 21,
383-393 (1936).
601. MUntzing A Sterility and chromosome pairing in intraspecific Galeopsis.
hybrids, Hereditas, 24, 117—188 (1938).
Л итература
487
€02. Munlzing A Genetics Basic and Applied, 2nd ed , LTs Forlag, Stockholm,
1961
603 Mu.tuZi.ng A, Prakken R Chromosomal aberrations in rje populations, He-
reditas, 27, 273—308 (1941)
604. Munz P A The Oenothera hookeri group, Aliso, 2, 1—47 (1949)
605. Munz PAA. California Flora, Unhcisity of California Press, Berkeley,
Caln , 1959
606 Munz P 4 Onagraceae Noith Ainei , Flora, 5, 1—278 (1965)
607 Munz PAA. Flora of Southern California, University of California Press,
Berkeley, 1974
608 Namkoong G Statistical analysis of mti egression, Biometrics, 22, 488—
502 (1986)
609. Naudin C Nouvelles recherches sur 1'hy bridite dans les vegetaux, Ann Sci
Nat, Ser Botan (Paris), 19 180—203 (1863)
610 Navashin M S The d slocation hypothesis of evolution of chromosome num-
bers, Zeitschr ind Abstammimgs- u Vererbungslehie, 63, 224—231 (1932).
611 Neva E Adaptive convergence and divergence of subterranean mammals,
Ann Rev Ecol Sjst, 10,269-308 (1979)
612 Newton W C F, PeUew C Primula kewensis and its denvatives, J Ge-
netics. 20, 405—467 (1929).
613. Ninan C A Studies on the cytology and phylogeny of the Pteridophytes
VI Observations on the Ophioglossaceae, Cytologia, 23, 291—316 (1958)
614 Nobs iM A Experimental studies on species relationships in Ceanothus,
Carnegie Inst Wash Publ, 623 (1963)
615. Nobs M A , Hiesey \X7 M Studies on differential selection in Mimulus
Carnegie Inst Washington Yearbook, 56, 291—292 (1957)
616 Nobs M A, Hiesey № M Perioimance of Mimulus races and their hyb
r’ds in contrasting~environments Carnegie Inst, Washington Yearbook, 57,
270—272 (1958)
617. Noda S Cytogenetic relationships between Liliutn iigrinurn and related
species and the hybrid origin of Liliutn tigrinum, Lilv Yearbook, 27, 98—
111 (1974).
618 Nordenskiold H The somatic chromosomes of some Luzula species, Botani-
ska Notise-, 1949, 81—92 (1949)
619 Nordenskiod H Cvtot'xonomical studies in the genus Luzula I. Heredi-
tas, 37, 325—355 (1951)
620 Nordenskiold H Cyto taxonomical studies in the genus Luzula II Hybridi-
zation experiments in the campestris-mulliflora complex, Hereddas, 42, 7—
73 (1956)
621 Nordenskiold H Tetrad analysis and the course of meiosis in three hybrids
of Luzula canipestris, Hereditas, 47, 203—238 (1961)
622 Nordenskiold H Studies oi meiosis in Luzula purpurea, Hereditas, 48, 503—
519 (1962)
623 Nygren A Tl.e genesis of some Scandinavian species of Calamagrostis,
Hereditas, 32, 131—262 (1946)
624. Nygren A Fo'm and biotype formation in Calatnagrostis purpurea. Here
ditas, 37, 519—532 (1951)
625 Nygren A Apomixis in the angiosperms, II Bot Rev, 20, 577—649
(1954).
626 Nygren A Artificial and natural hybridization in European Calamagrostis,
Symbolae Botanicae Upsalienses, 17, 1—105 (1962)
627 Nygren 4 On Poa badensis, Poa concinna, Poa pumila, Poa xerophila and
the possible origin of Poa alpina, Annales Academiae Regiae Scicntiarum
Upsaliensis, 1962(6), 1—29 (1962).
628 Oehlkers F Bastardierungsversuche in der Gattung Streptocarpus Lindley.
Ill Neue Ergebnisse liber die Gcnetik von Wuchsgestalt und Geschlechts-
bestimmung, Ber Deutsch Bot Ges , 58, 76—91 (1940).
488 ' Литература
629. Olivieri А. М, Jain S К Variation in the Helianthus exilis—bolanderi
complex: a reexamination, Madrono, 24, 177—189 (1977).
630. Ollgaard H. New species of Taraxacum from Denmark, Bot Not, 131,
497—521 (1978).
63 L Ornduff R. Concepts of species. (Book review.), Syst. Bot., 1, 347 (1976).
632. Ownbey M., Aase H. C. Cytotaxonomic studies in Allum. I. The Allium
canadense alliance, Research, Studies State College of Washington, Mono-
graph Suppl., 1, 1—106, 1955.
633. Palmer E J. Hybrid oaks of North America, J. Arnold Arboretum. 29, 1—
48 (1948).
634. Paterniani E. Selection for reproductive isolation between two populations
of maize, Zea mays L. Evolution, 23, 534—547 (1969)
635. Paterniani E., Stort A. C. Effective maize pollen dispersal in the field, Euphy-
tica, 23, 129—134 (1974).
636. Pazy B., Zohary D The process of introgression between Aegilops poly-
5>loids: natural hybridization between A. variabilis, A. ovata, and A. biuncia-
is, Evolution, 19, 385—394 (1965).
637. Peterson P. A. Cytoplasmically inherited male sterility in Capsicum, Amer.
Nat, 92, 111—119 (1958).
638. Pianka E R. On r- and К-selection, Amer. Nat, 104, 592—597 (1970).
639 Pijl L. van der. Principles of Dispersal in Higher Plants, 2d ed, Springer-
Verlag, Berlin and New York, 1972
640. Pimentel D., Smith G. J., Soans J. A population model of sympatric specia-
tion, Amer. Nat., 101, 493—504 (1967).
641. Pinkava D. J., McLeod M. G. Chromosome numbers in some cacti of wes-
tern North America, Brittonia, 23, 171—176 (1971).
642. Pinkava D. J., McLeod M. G., McGill L. A., Brown R. C. Chromosome num-
bers in some cacti of western North America. II Brittonia, 25, 2—9 (1973).
643 Pinkava D. J., McGill L. A., Reeves T, McLeod M. G Chromosome num-
bers in some cacti of western North America. III. Bull. Torrey Bot. Club.,
104, 105—110 (1977)
644 Powers L. Fertilization without reduction in guayule (Parthenium argenta-
tum Gray) and a hypothesis as to the evolution of apomixis and polyploidy,
Genetics, 30, 323—346 (1945).
645. Prager E. M., Fowler D. P., Wilson A. C. Rates of evolution in conifers
(Pinaceae), Evolution, 30, 637—649 (1976).
646. Prazmo W. Genetic studies on the genus Aquilegia L. I. Crosses between
Aquilegia vulgaris L. and Aquilegia ecalcarata Maxim, Acta Soc. Bot, Po-
lon., 29, 57—77 (1960).
647 Prazmo W. Genetic studies on the genus Aquilegia L II Crosses between
Aquilegia ecalcarata Maxim, and Aquilegia chrysantha Gray,* Acta Soc.
Bot. Polon, 30, 423—442 (1961).
648. Prazmo W Cytogenetic studies on the genus Aquilegia III Inheritance of
the traits distinguishing different complexes in the genus Aquilegia, Acta
Soc Botan Polon, 34, 403-437 (1965)
649. Prazmo W. Cytogenetic studies on the genus Aquilegia. IV/Fertility rela-
tionships among the Aquilegia species, Acta Soc. Bot. Polon., 34, 667—
685 (1965).
650. Pryor L. D. Species distribution and association in Eucalyptus. In: Biogeo-
graphy and Ecology in Australia, Monographiae Biologicae, 8, 461—471
(1959).
651. Puirament A. Some observations on male sterility in Geranium sylvaticum
L. var alpestre Schur. Acta Soc. Bot. Polon., 31, 723—736 (1962)
652. Rajhathy T., Thomas H. Cytogenetics of oats (Avena L.), Genetics Soc.
Canada Misc. Publ., 2 (1974).
653. Ramsbottom J. Linnaeus and the species concept, Proc. Linnean Soc. Lon-
don, 150, 192—219 (1938).
Литература
489
654 Randolph L. F Cytogenetics of tetrapioid maize, J Agr Research, 50, 591 —
605 (1935)
655 Randolph L F, Nelson I S, Plaisted R L Negative evidence of introgres-
sion affecting the stability of Louisiana Iris species, Cornell Univ. Agr. Exp.
Station Memoir, 398 (1967)
656 Ratter J A Cytogenetics studies in Spergularia IX Summary and conclu-
sions, Notes from Royal Bot Gard (Edinburgh), 34 411—428 (1976).
657 Raven P H Interspecific hybridization as an evolutionary stimulus in
Oenothera, Proc Linnean Soc London, 173, 92—98 (1962).
658. Raven P H A revision of the genus C amis sonia (Onagraceae), Contrib
US Nat Herbarium, 37, 159—396 (1969)
659 Raven P H The bases of angiosperm phvlogeny cytology, Ann Missouri
Bot Gard, 62, 724—764 (1975)
660. Raven P H Systematics and plant population biology, Syst Bot, 1, 284—
316 (1976).
661. Raven P. II Future directions in plant population biology In Topics in
Plant Population Biology, ed О T Solbrig, Columbia University Press,
New York, 1979
662 Raven P H, Dietrich W, Stubbe W An outline of the systematics of
Oenothera subsect Euoenothera (On'agraceac), Svst Bot, 4, 242—252
(1979—1980). (Fall 1979 issue, publ April, 1980)
663 Raven P H, Gregory D P Observations of meiotic chromosomes in Gaura
(Onagraceae). Brittonia, 24, 71 -86 (1972)
664. Raven P H., Ryhos D U? New evidence concerning the original basic num-
ber of angiosperms, Evolution, 19, 244—248 (1965)
665 Raven P H , Kyhos D IV, Cave M S Chromosome numbers and relation-
ships in Annoniflorae Taxon 20, 479—483 (1971)
666 Raven P H, Raven T E The Genus Fpilobium in Australasia. Dept Sci
Indust Research. Christchurch, New Zealand, 1976
667 Raven P H Thompson II J Haploidy and angiosperm evolution, Amer
Nat, 98, 251—252 (1964)
668 Ray J Historia Plan*arum, London, 1686
669 Reddy M Al. Garber E D Genetic studies of variant enzymes III Compa-
rative electrophoretic studies of esterases and peroxidases for species, hyb
rids and amphiploids in the genus Nicotiana, Bot Gaz, 132, 158—166
(1971)
670 Rees H Genotypic control of chromosome foim and behavior, Bot Rev,
27, 288-318 (1961)
671. Rehfeldt G E Ecotypic diffei entiation in populations of Pinus monticola
in north Idaho-myth’or reality? Amer Nat, 114, 627—636 (1979)
672 Remington C L Suture-zones of hybrid interaction between recently joined
biotas, Evolutionary Biol. 2, 321—428 (1968)
673. Renner О Untersuchungen uber die faktoricllc Konstitution einiger komple-
xheterozygotischer Oeno^heren, Bibliotheca Genetica, 9, 1—168 (1925)
674 Renner О Uber die Entstehung homozvgotische- Formen aus komplcxhctcro-
zygotischen Oenotheren, F ora, 35, 201—238 (1941)
675. Renner О Europaischc Wildarten von Oenothera, Ber Deutsch Bot Ges,
60 448—466 (19421
676 Rensch В Das Pi inzip geographischer Rassenkieise und das Problem der
Artbildung, Bo ntraeger, Berlin, 1929
677 Reuther U7, Batchelor L D, Webber II I. eds The C'trus Industry,
vol II Anatomy Physiology, Genetics and Reproduction 2d ed Univer-
sity of California Pt ess, Berkeley and Los Angeles, Calif, 1968
678 Reuther IF, Webber J, Batchelor L D. eds The Citrus Industry, vol I.
History, World Distribution Botany and Varieties 2d ed University of Ca-
lifornia Press, Berkeley and Los Angeles, Calif, 1967
490 Литература
679. Rhoades М. М, Dempsey Е. Induction of chromosome doubling at meiosis
by the elongate gene in maize, Genetics, 54, 505—522 (1966).
680 Rhyne C. L Linkage studies in Gossypium. I. Altered recombination in allo-
tetraploid G. hirsutum L. following linkage group transference from rela-
ted diploid species,Genetics, 43, 822—834 (1958).
681. Richards A. J. The origin of Taraxacum agamospecies, Bot. J. Linn. Soc.„
66, 189—211 (1973).
682. Richardson M M. Structural hybridity in LiUum martagon albumxL. han-
sonii, J. Genetics, 32, 411—450 (1936).
683. Rick С. M. Differential zygotic lethality in a tomato species hybrid, Gene-
tics, 48, 1497—1507 (1963).
684. Rieger R. Die Genommutationen, Veb Gustav Fischer, Jena, 1963.
685. Righter F. L, Duffield J. IF. Interspecies hybrids in pines, J. Heredity, 42„
75—80 (1951).
686. Riley H. P. A character analysis of colonies of Iris fulva, Iris hexagona var_
giganiicaerulea and natural hybrids, Amer. J. Bot., 25, 727—738 (1938).
687. Riley R. The diploidisation of polyploid wheat, Heredity, 15, 407—429
(1960).
688. Roberts H. F. Plant Hybridization before Mendel, Reprint, 1965, Hafner,.
New York, 1929, 1965.
689. Rollins R. C. Evidence for natural hybridity between guayule (Parthenium.
argentatum) and mariola (Parthenium incanum), Amer. J. Bot., 31, 93—
99 (1944)
690. Rollins R. C. Interspecific hybridization in Parthenium I, Crosses between
guayule (P. argentatum) and mariola (P. incanum), Amer. J. Bot., 32, 395—
404 (1945).
691. Rollins R. C. Interspecific hybridization in Parthenium. II. Crosses invol-
ving P. argentatum, P. incanum, P. stramonium, P. tomentosum and
P. hysterophorus, Amer. J. Bot., 33, 21—30 (1946).
692. Rollins R. C. The evolution and systematics of Leavenworthia (Cruciferae) r
Contrib. Gray Herbarium (Harvard), No. 192 (1963).
693. Rollins R C., Shaw E A The Genus Lesquerella (Cruciferae) in North
America, Harvard University Press, Cambridge, Mass., 1973.
694. Roose M. L., Gottlieb L. D Genetic and biochemical consequences of poly-
ploidy in Tragopogon, Evolution, 30, 818—830 (1976).
695. Ross H. H. Principles of natural coexistence indicated by leafhopper popu-
lations, Evolution, 11, 113—129 (1957).
696. Ross H. H. Evidence suggesting a hybrid origin for certain leafhopper spe-
cies, Evolution, 12,337—346 (1958).
697 Ross J. G., Duncan R. E. Cytological evidences of hybridization between
Juniperus virginiana and /.. horizontalis. Bull. Torrey Bot. Club., 76, 414—
429 (1949).
698. Rothwell N. V. Aneuploidy in Claytonia virginica, Amer. J. Bot, 46, 353—
360 (1959).
699 Rousi A. Biosystematic studies on the species aggregate Potentilla ansert-
na L., Annales Botanici Fennici, 2, 47—112 (1965).
700. Russell N. H. Three field studies of hybridization in the stemless white vio-
lets, Amer. J. Bot., 41, 679—686 (1954)
701. Rutishauser A.. Embryologie und Fortpflanzungsbiologie der Angiospermen,
Springer Verlag, Vienna, 1969.
702. Sachs J. History of Botany, Translation, Oxford University Press, Oxford,
1906.
703. Salisbury E. J. Reproductive Capacity of Plants, G. Bell and Sons, Lon-
don, 1942.
704. Salisbury E J. Weeds and Aliens, Collins, London, 1961.
705. Salisbury E. J A note on shade tolerance and vegetative propagation of
woodland species, Proc. Royal Soc. London, B, 192,257—258 (1976)
Литература
491
706. Sandfaer J High frequency oi spontaneous triploids in barlev, Hereditas,
64, 131—134 (1970)
707. Sana Y, Rita F Genes for reproductive isolation located on rearranged
chromosomes, Heredity, 41, 377—383 (1978)
708 Sana Y, Chu Y, Oka H. Genetic studies of speciation in cultivated rice
II Character variations in backross derivaties between Oryza sativa and
O. glaberrima M V linkage and key characters, J. J. Genetics, 55, 19—39
(1980)
709 Satina S, Blakeslee A F Cytological effects of a gene in Datura which
causes dyad formation in sporogenesis Bot Gaz, 96, 521—523 (1935).
710 Sax H J Polyploidy and apomixis in Cotoneaster, J Arnold Arboretum,
35, 334—365 (1954).
711 Sax К Chromosome ring format.on in Rhoeo discolor, Cytologia, 3, 36—
53 (1931)
712 Schaal В A, Levin D A Morphological differentiation and neighborhood
size in Liatris cylindracea, Amer J Bot, 65, 923—928 (1978)
713. Schultz R. J Hybridization, unisexuality and polyploidy in the teleost
Poectliopsis (Poeciliidae) and other vertebrates, Amer Nat, 103, 605—619
(1969).
714. Schultz R J Role of polyploidy in the evolution of fishes In Polyploidy,
Biological Relevance, ed W H Lewis, Plenum Press, New York and Lon-
don, 1980
715 Schwanitz F Spornbildung bei cincm Bastard zwischen drei Digitalis-Ar-
ten, Biol Zeniralblatt, 76, 226—231 (1957).
716. Sears E R Chromosome pairing and fertility in hybrids and amphiploids in
the Triticinae, Univ Missouri Agr Exp Station Bull, No 337 (1941)
717. Sears E R Cytogenetic studies with polyploid species of wheat. II Addi-
tional chromosomal aberrations in Triticum vulgare, Genetics, 29, 232—
246 (1944)
718 Sears E R The cytology and genetics of the wheats and their relatives,
Advances in Genetics, 2, 239—270 (1948)
719 Sears E R The systematics, cytology and genetics of wheat, Handbuch
der Pflanzenziichtung, 2, 164—187 (1959)
720 Semple J C, Brouillet L Chromosome numbers and satellite chromosome
morphology in Aster and Lasallea, Amer J Bot, 67, 1027—1039 (1980)
721 Shindo R, Ramemoto H Chiomosorne numbers and genome relationships
of some species in the Nigrohirsutae Section of Dendrobium, Cytologia, 28,
68—75 (1963)
722 Sibley C G Species formation in the red-eyed towhees of Mexico, Univ.
Calif Publ Zool. 50, 109 (1950)
723 Sibley C G Hybridization in the red-eyed towhees of Mexico, Evolution,
8, 252—290 (1954)
724 Silander J A. Microevolution and clone structure in Spartina patens,
Science, 203, 658—660 (1979)
725. Simmonds N. IF Meiosis in tropical Rhoeo discolor, Natuic, 155, 731
(1945)
726 Simmonds N W The Evolution of the Banana Longmans Green, London,
1962
727 Simmonds N W Bananas, 2d ed, Longmans Green. London, 1966
728 Simmonds N IF, cd Evolution of Crop Plants, Longman Group Ltd, Lon-
don, 1976
729. Simpson В В, ed Mesquke Its Biology in Two Desert Scrub Ecosystems,
Dowden, Hutchinson and Ross, Stroudsburg, Pennsylvania, 1977
730. Simpson G G Tempo and Mode in Evolution, Columbia University Press,
New York, 1944 [Имеется перевод Симпсон Дж Темпы и формы эволю-
ции — М ИЛ, 1948 ]
731 Simpson G G The species concept, Evolution, 5, 285—298 (1951)
492
Литература
732. Simpson G G. Principles of Animal Taxonomy, Columbia University Press,
New York, 1961.
733. - Sinskaja E. N. The study of species in their dynamics and interrelation
with different types of vegetation, Bull. Appl. Bot. Genetics Plant Breeding
(Leningrad), 25, 1-97 (1931)
734. Skalinska M. Seed development after crosses of Aquilegia with Isopyrum,
Studies in Plant Physiol (Prague), 213—221, 1968
735. Skalinska M. Cytological analysis of some Hieracium species, subg. Pilo-
sella from mountains of southern Poland, Acta Biologica Cracoviensia, Bo-
tanica, 10, 127—141 (1967).
736. Скворцов А R. Ивы СССР. Таксономический и географический обзор. —
М.: <Наука», 1968.
737. Slobodchikoff С N., ed. Concepts of Species, Dowden, Hutchinson and Ross,
Stroudsnurg, Pennsylvania, 1976.
738. Small J. K„ Alexander E. J Botanical interpretation of the iridaceous plants
of the Gulf states, Contrib. New York Bot. Card., No. 327 (1931).
739. Smith D. M„ Craig S„ Santarosa I Cytological and chemical variation in
Pityrogramma triangularis, Amer. J. Bot., 58, 292—299 (1971).
740. Smith D. M„ Levin D. A. A chromatographic study of reticulate evolution
in the Appalachian Asplenium complex, Amer. J. Bot, 50, 952—958 (1963).
741. Smith F. H, Clarkson Q D Cytological studies of interspecific hybridiza-
tion in Iris, subsection Californicae, Amer. J. Bot., 43, 582—588 (1956)
742 Smith H. H. Development of morphologically distinct and genetically iso-
lated populations by interspecific hybridization and selection, Caryologia,
Suppl Vol, 867—870, 1954.
743. Smith H. И Genetic plant tumors in Nicotiana, Annals New York Acad.
Sci, 71, 1163—1177 (1958).
744. Smith H. H, Daly K. Discrete populations derived by interspecific hybri-
dization and selection in Nicotiana, Evolution, 13, 476—487 (1959).
745. Smith-While S Polarised segregation in the pollen mother cells of a stable
triploid. Heredity, 2, 119—129 (1948)
746. Smith-White S. The life history and genetic system of Leucopogon junipe-
rinus, Heredity, 9 79—91 (1955).
747. Smith-White S Cytological evolution in the Australian flora, Cold Spring
Harbor Symp Quant BioL, 24, 273—289 (1959).
748. Smith-White S. Pollen development patterns in the Epacridaceae, Proc.
Linnean Soc. New South Wales, 84, 8—35 (1959).
749. Snogerup S. Studies in the Aegean flora. IX. Erysimum sect. Cheiranthus.
B. Variation and evolution in the small-population system, Opera Botani-
ca, 14, 1—86 (1967).
750. Snyder L. A. Morphological variability and hybrid development in Elymus
glaucus, Amer. J. Bot., 37, 628—636 (1950).
751. Snyder L A Cytology of inter-strain hybrids and the probable origin of
variability in Elymus glaucus, Amer. J. Bot., 38, 195—202 (1951).
752. Soans A. B„ Pimentel D., Soans J. S. Evolution of reproductive isolation
in allopatric and sympatric populations, Amer. Nat., 108, 117—124 (1974)
753. Sokal R. R. The species problem reconsidered, Syst. Zool., 22, 360—374
(1973)
754 Sokal R R, Crovello T J The biological species concept: a critical eva-
luation, Amer. Nat, 104, 127—153 (1970).
755. Solbrig О. T. Cytotaxonomic and evolutionary studies in the North Ameri-
can species of Guttierrezia (Compositae), Contrib Gray Herbarium (Har-
vard Univ), 188, 1-63 (1960).
756. Solbrig О T. Principles and Methods of Plant Biosystematics, MacMillan,
New York, 1970.
757. Solbrig О. T. The population biology of dandelions, Amer. Sci., 59, 686—
694 (1971)
Литература
495
758
759
760
761
762
763
764
765
766
767
768
769
770
771
772
773
774
775
776.
777
778
779
780
781
782
783
Solbrig О Т On the ielative advantages of cross and self-fertilization,
Ann Missouri Bot Gard, 63, 262—276 (1976)
Solbrig О T A cost benefit analysis of recombination in plants In Topics
in Plant Population Biology, ed О. T. Solbrig, Columbia University Press,
Neu \ork, 1979
Solbrig О T., Anderson I. C, Kyhos D W. Raven P H, Ruedenberg L
Chromosome numbers in Compositae V Astereae II, Amer J Bot, 51,
513-519 (1964).
Solbrig О T, Jain S, Johnson G В, Raven P II., eds Topics in Plant
Population Biology, Columbia University Press, New York, 1979
Sonneborn T M Breeding systems, reproductive methods, and species prob-
lems in Protozoa In The Species Problem, ed E Mayr, Amer Assoc Adv
Sci, V»ashington, D C , 1957
Serensen T, Gudjonsson G Spontaneous chromosomc-abberrants in apo-
mictic Taraxaca Morphological and cytogenetical investigations, К Dan
skeVidensk Selskab Biol, 4, 1—48 (1946)
Soumalainen E Partbcnogencs s in animals, Advances in Genetics. 3, 193—
253 (1950)
Sprague E Pollination and evolution in Pedicularis (Scrophulanaceae),
Aliso, 5. 181—209 (1962)
Squillace A E, Bingham R T Localized ecotypic variation in western uhite
pine, Forest Sci, 4, 20—34 (1958)
Stace C A Hybridization and the Flora of the British Isles, Academic
Prose, London, 1975
Stafleu F A Linnaeus and the Linnaeans A Oosthoek, Utrecht, 1971
Stalker Ff D A case of polyploidv in Diptera Proc Nat Acad Sci, 42,
194-199 (1956).
Stalker H D The phvtogcnetic relationships of the species in the Droso-
phila melanica group, Genetics 53 327—342 (1966)
Stanleu S M A theory of evolution above the species level, Proc Nat.
Acad Sci. 72, 646—650 ‘(1975)
Stanleu S M Macroevo'ution, Pattern and Process, W H Freeman, San
Francisco, 1979
Stebbins G L Cytologv of Antennaria I No-mal species, Bot Gaz, 94,
134—151 (1932)
Stebbins G L A new species of Antennaria from the Appalachian region,
Rhodora, 37, 229—237 (1935)
Stebbins G L Cytological characteristics associated with the different
growth habits in the dicotyledons Amer J Bot, 25, 189—198 (1938)
Stebbins G L The role of isolation in the diffcrention of plant species,
Biol Symp , 6, 217—233 (1942)
Stebbins G L The cvtological analysis of species hybrids, II, Bot. Rev,
11,463-486 (1945)
Stebbins G L Types of polyploids their classification and significance,
Advances in Genetics, 1, 403—429 (1947)
Stebbins G L The origin ot the complex of Bromus carinatus and its phy-
togeographic implications, Contrib Grav Hcrbaiium (Harvard), 165, 42—
55 (1947)
Stebbins G L Variation and Evolution in Plants, Calumbia University
Press, New Yor.-c, 1950
Stebbins G L Cvtogcnctics and evolution of the grass family, Amer J
Bot, 43, 890—905'(1956)
Stebbins G L Self fertilizat.on and population variability in the higher
plants, Amer Nat, 91, 337—354 (1957)
Stebbins G L The hybrid origin of microspecies in the Elymus glaucus
complex, Cytologia, Suppl vol, 336—340, 1957.
-494 Литература
784. Stebbins G. L The inviability, weakness and sterility of interspecific hyb-
rids, Advances in Genetics, 9, 147—215 (1958).
785. Stebbins G. L. Longevity, habitat, and release of genetic variability in the
higher plants, Cold Spring Harbor Symp Quant. Biol, 23, 365—378 (1958)
786. Stebbins G. L. The role of hybridization in evolution, Proc. Amer. Phil.
Soc., 103, 231—251 (1959).
787. Stebbins G L Chromosomal Evolution in Higher Plants, Edward Arnold,
London, 1971.
788. Stebbins G. L. Flowering Plants. Evolution Above the Species Level, Har-
vard University Press, Cambridge, Mass., 1974.
789. Stebbins G. L Polyploidy in plants- unsolved problems and prospects. In-
Polyploidy:Biological Relevance, ed. W. H. Lewis, Plenum Press, New York
and London, 1980.
790. Stebbins G. L., Daly K. Changes in the variation pattern of a hybrid popula-
tion over an eight-year period, Evolution, 15, 60—71 (1961).
791. Stebbins G. L, Ferlan L Population variability, hybridization, and intro-
gression in some species of Ophrys, Evolution, 10, 32—46 (1956)
792 Stebbins G L, Jenkins J A Aposporic development in the North American
species of Crepis, Genetica, 21, 191—224 (1939).
793. Stebbins G L., Jenkins J A, Walters M. S Chromosomes and phylogeny
in the Compositae, tribe Cichorieae, Univ. Calif. Publ. Bot., 26, 401—429
(1953).
794. Stebbins G L., Tobgy H. A. The cytogenetics of hybrids in Bromus I Hyb-
rids within the section Ceratochloa, Amer. J. Bot., 31, 1—11 (1944).
795. Stebbins G. L, Tobgy H A, Harlan J R The cytogenetics of hybrids in
Bromus. II. Bromus carinatus and Bromus arizonicus, Proc. Calif. Acad.
Sci,25, 307—322 (1944).
796. Stebbins G. L., Vaarama A Artificial and natural hybrids in the Gramineae
tribe Hordeae. VII. Hybrids and alloploids between Elymus glaucus and
Sitanion spp. Genetics, 39, 378—395 (1954).
797. Stebbins G. L., Zohary D Cytogenetic and evolutionary studies in the ge-
nus Dactylis. I Morphology, distribution and interrelationships of the di-
ploid subspecies, Univ. Calif. Publ Bot, 31, 1—40 (1959).
798. Steiner E Phylogenetic relationship of certain races of Euoenothera from
Mexico and Guatemala, Evolution, 5, 265—272 (1951).
799. Steiner E Phytogenetic studies in Euoenothera, -Evolution, 6, 69—80
(1952).
•800. Stephens S. G The cytogenetics of speciation in Gossypium. I. Selective
elimination of the donor parent genotype in interspecific backcrosses, Gene-
tics, 34,627—637 (1949).
'801. Stephens S. G. The internal mechanism of speciation In Gossypium, Bot
Rev., 16, 115—149 (1950).
•802. Stephens S. G. Possible significance of duplication in evolution, Advances
in Genetics, 4, 247—265 (1951).
803. Stephens S. G. «Homologous» genetic loci in Gossypium, Cold Spring
Harbor Symposia Quant. Biol., 16, 131—141 (1951).
804. Stephens S. G, Finkner M. D. Natural crossing in cotton, Economic Bot.,
7, 257—269 (1953).
805. Stiff M. L. A naturally-occurring triploid juniper, Virginia J. Sci. N. S, 2,
317 (1951).
806. Stinson H. T. Cytogenetics and phylogeny of Oenothera argillicola
Mackenz, Genetics, 38, 389—406 (1953).
807. Straley G B. Systematics of Oenothera sect, Rneiffia (Onagraceae), Annals
Missouri Bot. Gard., 64, 381—424 (1977).
808. Straw R M. Hybridization, homogamy, and sympatric speciation, Evolu-
tion, 9, 441—444 (1955).
809. Straw R M. Floral isolation in Penstemon, Affier. Nat., 90, 47—53 (1956).
Литература
495
810 Streams F. A, Pimentel D Effects of immigration on the evolution of po-
pulations, Amer Nat., 95, 201—210 (1961)
811 Strid A Studies in the Aegean flora XVI Biosystematics of the Nigella
arvensts complex with special reference to the problem of nonadaptive ra-
diation, Opera Botanica, 28, 1—169 (1970)
812 Swaminathan M S Microsporogenesis in some commercial potato varieties,
J Heredity, 45, 265-272 (1954)
813 Swaminathan M S, Hougas R W Cytogenetic studies in Solanum verruco-
sum variety spectabilis, Amer. J Bot, 41, 645—651 (1954)
814 Swaminathan M S, Howard II W The cytology and genetics of the po-
tato (Solatium tuberosum) and related species, Bibliographia Genetica,
16, 1-192 (1953)
815 Swaminathan M S, Magoon M L Origin and cytogenetics oi the commer-
cial potato, Advances in Genetics, 10, 217—256 (1961).
816 Switlinska Z Spontaneous polvploidization in Rumex hybrids, Acta Soc.
Botan Polon, 29, 79—98 (I960)’
817 Swiethnska Z, Zuk J Further observations on spontaneous polyploidization
in Rumex hybrids, Acta Soc Botan Polon , 34, 439—450 (1965)
818 Swingle W T The botany of Citrus and its wild relatives In The Citrus
Industry, 2d ed , vol 1, eds W Reuther, H J Webber and L D Batchelor,
University of California Press, Berkeley and Los Angeles, Calif, 1967
819 Tatkholm G Zytologische Studien uber die Gattung Rosa. Acta Horti Ber-
giani (Uppsala), 7 97—381 (1922).
820 lahara Л1 Cvtological studies on Chrysanthemum II Bot Mag Tokyo, 29,
48—50 (1915)
821 Takhtajan .4 Flowering Plants Origin and Dispersal Translation, Oliver
and Boyd, Edinburgh, 1969
822 Tanaka T Species Problem in Citrus Japanese Society for the Promotion
of Science, Leno, Tokvo. Japan, 1954
823 Tanaka T Citrologia Citrologia Supporting Foundation, Osaka, Japan
(Summarized in Sw.ngle, 1967), 1961
824 Tedin О Ve'crbung, Variation und Svstematik in der Gattung Camelina,
Hereditas, 6, 275—386 (1^25)
825 Thoday J .W, Boom T В Effects of disruptive selection II. Polymorphism
and divergence without isolation, Heredity, 13 205—218 (1959)
826 Thodai/ J M , Gibson J В Isolation by disruptive selection, Nature, 193,
1164—1166 (1962)
827 Thoday J .4 , Gibson J В The probability of isolation by disruptive selec-
tion, Amer Nat 104,219—230 (1970)
828 Thorne R T Synopsis of a putatively phylogenetic classification of the
flowering plants, Allso, 6, 57—66 (1968)
829 Ting Y C Common inversions in maize and teosinte, Amer Nat, 101, 87—
89 (1967).
830 Tischler G D e Bedeutung der Polyploidie fur die Verbrcitung der Angio-
spermen, Bot Jahrb, 67, 1—36 (1935)
831 Tischler G Allgcmcinc Pilanzenkaryologie III Angewandte Pflanzenkaryo-
logie, Handbuch dor Pflanzenanatomie, Berlin, 1953
832 Tischler G Das Problem der Basis-Chromosomenzahlen bei den Angiosper-
men-Gattungen und -Familien, Cytologia, 19, 1 — 10 (1954)
833 Tischler G Der Grad der Polyploidie bei den Angiospermen in verschiede-
nen Grossarealen, Cyto’og.a, 20 101—118 (1955)
834 Tobgy HAN cytological study of Crepis fuliginosa, C neglecta, and
their Fi hybrid, and its bearing on the mechanism of phylogenetic reduc-
tion in chromosome number, J Genetics, 45, 67—111 (1943)
835. Towner H F The biosystematics of Calylophus (Onagraceae), Annals
Missouri Bot Gard, 64, 48—120 (1977)
496
Литература
836. Tucker J. M. Taxonomic interrelationships in the Quercus dumosa complex,
Madrono, 11, 234—251 (1952).
837. Tucker J. M. Two new oak hybrids from California, Madrono, 12, 119—
127 (1953).
838. Tucker J. M. Identity of the oak tree Live Oak Tanks Joshua Tree National
Monument, California, Madrono, 19, 256—266 (1968).
£39. Tucker J M , Sauer J Aberrant Amaranthus populations of the Sacramento-
San Joaquin delta, California, Madrono, 14, 252—261 (1958).
840. Turesson G The genotypical response of the plant species to the habitat,
Hereditas, 3, 211—350 (1922).
•841. Turesson G. The plant species in relation to habitat and climate, Hereditas,
6, 147—236 (1925)
842. Turesson G. Zur Natur und Begrenzung der Arteinheiten, Hereditas, 12,
323—334 (1929)
£4 3. Turesson G. The selective effect of climate upon the plant species, Heredi-
tas, 14, 99—152 (1930).
£4 4. Turner В L Summary of the biology of the Compositae. In’ The Biology
and Chemistry of the Compositae, ed. V. H. Heywood, J. B. Harborne and
В L. Turner, Academic Press, London and New York, 1978.
•8 45. Turner B. L, Alston R. Segregation and recombination of chemical consti-
tuents in a hybrid swarm of Baptisia laevicaulisXB. virldis and their taxo-
nomic implications, Amer. J. Bot, 46, 678—686 (1959).
£4 6. Turner В L, Horne D. Taxonomy of Machaeranthera sect. Psllactis (Com-
positae-Asterae), Brittonia, 16, 316—331 (1964)
£4 7. Turner B. L., Powell A. M. Deserts, gypsum and endemism. In: Arid Land
Plant Resources, ed. J R. Goodin and D. K. Northington, Texas Tech Uni-
versity, Lubbock, 1979.
848 U N Genome-analysis in Brassica with special reference to the experimen-
tal formation of B. napus and peculiar mode of fertilization, Japanese J
Bot., 7, 389—452 (1935).
£4 9 Upcott M. The nature of tetraploidy in Primula kewensis, J. Genetics, 39,
79—100 (1939).
850 Urbanska-Worytkiewicz K. L’agamospermie, systeme de reproduction im-
portant dans la speciation des angiospermes, Bull. Soc. Bot. France, 121,
329—346 (1974).
851. Urbanska-Worytkiewicz К Cytological variation within Lemna L. Aquatic
Botany, 1, 377—394 (1975).
852. Urbanska-Worytkiewicz K. Reproduction in natural triploid hybrids (2n=
=24) between Cardamine rivularis Schur and C amara L. Ser. Geobot.
Inst ETH, Stiftung Rubel (Zurich), 44, 42—85 (1977)
853. Urbanska-Worytkiewicz K. An autoallohexaploid in Cardamine L., new to
the Swiss flora, Ber Geobot Inst ETH, Stiftung RQbel (Zflrich), 44, 86-
ЮЗ (1977).
854. Urbanska-Worytkiewicz K. Segregation polarisee chez les hybrides naturels
triploides (2n=24) entre Cardamine rivularis Schur (2n= 16) et C amara
L. (2n=16), Bull. Soc. Botan. France, 125, 91—93 (1978).
855 Valentine D. H, Love A Taxonomic and biosystematic categories, Britto-
nia, 10, 153—166 (1958).
856 Vasek F. C Outcrossing in natural populations. I. The Breckenridge Moun-
tain population of Clarkia exilis, Evolution, 18, 213—218 (1964).
857. Vasek F C Outcrossing in natural populations II. Clarkia unguiculala,
Evolution, 19, 152—156 (1965).
858. Vasek F C. Outcrossing in natural populations III. The Deer Creek popu-
lation of Clarkia exilis, Evolution, 21, 241—248 (1967).
859. Vasek F. C. The relationships of two ecologically marginal, sympatric
Clarkia populations, Amer. Nat., 102, 25—40 (1968).
Литература
497
860 Vickery R К, Case studies in the evolution of species complexes in Mitu-
lus. Evol Biol, 11,405—507 (1978)
861 Viosca P The ir ses of southeastern Louisiana, Bull Amer. Iris Soc, No 57
(1935)
862 Waddington К D Divergence in inflorescence height, an evolutionary res-
ponse to pollinator, Oecologia 40, 43—50 (1979)
863 Wagner Л1 Die Entstehung der Arten durch raumliche Sonderung, Benno
Schwalbe, Basel, 1889 ,
864 Wagner W H Reticulate evolution in Appalachian Aspleniums, Evolution,
8, 103-118 (1954).
865 Wagner W. H The Barnes hybrid aspen, PopulusXbarnesii, hyb nov. A
nomenclatural case n point, Mich Botanist, 9, 53—54 (1970).
866 Wagner W H, Schoen D J Shingle oak {Quercus imbricaria) and its hyb-
rids in Michigan. Michigan Bot, 15, 141—155 (1976).
867 Wahl H A Chromosome numbers and meiosis in the genus Carex Amer
J Bot, 27, 458—470 (1940)
868 Waines J G A model for the origin of diploidizing mechanisms in poly-
ploid species, Amer. Nat, 110 415—430 (1976)
869 Walker J W Chromosome numbers, phylogeny, phytogeography of the
Annonaceae and their bearing on the (original) basic chromosome number
of angiosperms, Taxon, 21, 57—65 (1972)
870 Walker T G Hybridization in some species oi Pieris L. Evolution, 12, 82—
92 (1958)
871 Walker T G Cvtology and evolution in the fern genus Pteris L , Evolution,
16, 27-43 (1962)
872 Walker T G Apomix.s and vegetative reproduction in ferns In Repro-
ductixe Biology and Taxonomy of Vascular Plants, ed J G Hawkes, Per-
gamon Press Oxford, 1966
873 Wallace 4 R Darwinism An Exposition of the Thco-y of Natural Selection,
Macmillan, London, 1889
874 Wallace В Genetic divergence of isolated populations of Drosophila mela-
nogaster, Caryologia, Supp1 vol, 761—764 (1954)
875 Wallace В Influence of genetic systems on geographical distribution, Cold
Spring Harbor Symp Quant Biol ,'24, 193—204 (1959)
876 Waller D M The relative costs of se’f- and cross-fertilized seeds in Impa-
tients capensis (Balsaminaceael, Amer J Bot, 66, 313—320 (1979)
877 Walter R Holcus mollis L in Poland II Studies in the origin of polyploid
types. Acta Biol Cracovicnsia, 20, 113—131 (1977)
878 Wafers M S Studies oi spontaneous chromosome brekage in interspecific
hybrids of Bromus, Univ. Calif Publ Bot, 28, 335—447 (1957).
879 Walters M S Rates of meiosis, spindle irregularities and microsporocyte di-
vision in Bromus trimiXB carinatus Chromosoma, 11, 167—204 (1960)
880 Webber II J, Batchelor L D The Citrus Industry, vol I History, Botany
and Breeding, 1st ed University of California Press, Berkeley and Los An
gcles, Calif, 1943
881 Weil J , Allard R W The mating system and genetic variability in natural
populations of Collinsia heterophylla, Evolution. 18, 515—525 (1964)
882 Wet smarm A Vortrage ube- Deszendenztheorie, 3rd ed, Fischer, Jena,
1913
883 Wells H A distance coefficient as a hybridization index an example using
Mimulus longiftorus and Af jlemingii (Scrophulariaceae) from Santa Cruz
Island California, Taxon, 29, 53—65 (1980).
884 tfW Л’ В. Garber E Г) Genet c studies of variant enzymes IT The inhe-
ritance of esterases and leucine aminopeptidases in Phaseolus vulgarisX
XP coccineus Canad J Genet Cytol, 9, 646—655 (1967)
885. Westergaard M Studies on cytology and sex determination in polyploid
forms of Molandrtum album Dansk Bot Arkiv, 10, 1 — 131 (1940)
32-45
498
Литература
886. Whalen М. D. Reproductive character displacement and floral diversity in
Solanum section Androceras, Syst. Bot., 3, 77—86 (1978).
887. White M. J. D. Evolution of cytogenetic mechanisms in animals. In: Gene-
tics in the Twentieth Century, ed. L. C. Dunn, Macmillan, New York, 1951.
888. White M. J. D. Animal Cytology and Evolution, 2d ed. Cambridge Univer-
sity Press, Cambridge, 1954.
889. White M. J. D. Cytogenetics and systematic entomology, Ann. Rev. Ento-
mol„ 2, 71—90 (1957).
890,-W7ufe M. J. D Some general problems of chromosomal evolution and spe-
ciation in animals, Survey of Biol. Progress, 3, 109—147 (1957).
891. White M. J. D. Speciation in animals, Australian J. Sci„ 22, 32—39 (1957).
892. White M. J. D. Principles of karyotype evolution in animals. In: Genetics
Today, Pergamon Press, Oxford, 1964.
893. White M. J. D. Animal Cytology and Evolution, 3rd ed., Cambridge Univer-
sity Press, Cambridge, 1973.
894. White M. J. D. Modes of Speciation, W. H. Freeman, San Francisco, 1978.
895. Wiegand K. M., 1935. A taxonomist’s experience with hybrids in the wild,
Science, 81, 161—166 (1935).
896. Wilson A. C Evolutionary importance of gene regulation, Stadler Sympo-
sia (Univ. Missouri), 7, 117—134 1975.
897. Wirnber D. E. The nuclear cytology of bivalent and ring-forming Rhoeos
and their hybrids, Amer. J. Bot., 55, 572—574 (1968).
898. Winge 0. The chromosome: their number and general importance, Compt.
Rend. Trav. Lab. Carlsherg, 13, 131—275 (1917).
899. Winge 0. Contributions to the knowledge of chromosome numbers in plants,
La Cellule, 35, 303—324 (1924).
900. Winge O. Inheritance of species characters in Tragopogon, Compt. Rend.
Trav. Lab. Carlsberg, Series Physiol., 22, 155—194 (1938).
901. Winge O. Taxonomic and evolutionary studies in Erophila based on cytoge-
netic investigations, Compt Rend. Trav. Lab. Carlsberg, Series Physiol.,
23, 41—74 (1940).
902. Winkler H. Uber die experimentelle Erzeugung von Pflanzen mit abwei-
chenden Chromosomenzahlen, Zeitschr. f. Bot., 8, 417—531 (1916)
903. Wolf С. B. The Gander oak, a new hybrid oak from San Diego County, Ca-
lifornia, Proc. Calif. Acad. Sci., 25, 117—188 (1944).
904. Wolf С. B. Taxonomic and distributional studies of the New World cypres-
ses, Aliso, 1, 1—250 (1948).
905. Wright J. W. Species crossability in spruce in relation to distribution and
taxonomy, Forest Sci, 1, 319-349 (1955).
906. Wright J. W. Genetics of eastern white pine. Forest Service Research Pa-
per US. Rept. Agriculture, 1970.
907. Wright J W.. Gabriel W. J. Species hybridization in the hard pines, series
Sylvesttres, Silvae Genetica, 7, 109—115 (1958).
908 Wright S. Evolution m Mendelian populations, Genetics, 16, 97—159
(1931).
909. Wright S Isolation by distance, Genetics, 28, 114—138 (1943).
910 Wright S. An analysis of local variability of flower color in Linanthus par-
ry ae, Genetics, 28, 139—156 (1943).
911. Wright S. Isolation by distance under diverse systems of mating Genetics,
31, 39—59 (1946).
912. Wright S Adaptation and selection. In: Genetics, Paleontology and Evolu-
tion, ed. G. L. Jepsen, Princeton University and Evolution, ed. G. L. Jep-
sen, Princeton University Press, Princeton, N. J., 1949.
913. Wright S. Evolution and the Genetics of Population, vol 4. Variability Wi-
thin and Among Natural Populations, University of Chicago Press, Chicago,
1978.
914. Wulff H. D. Die Pollenentwicklung der Juncaceen nebst einer Auswertung
Литература 499
der enbryologischen Befunde hinsichtlich der Verwandtschaft der Jtfflca<-
ceen und Cyperaceen, Jahrb. Wiss. Botanik, 87,533—556 (1939).
915. Wulff H. D. Uber das spontane Auftreten einer Caninae-Meiosis bei der
Mikrosporogenese der diploiden, Rosa ruga Lindt Osterr. Bot. Zeitschr.,
101, 539—557 (1954).
916. Yarnell S. H. Cytogenetics of the vegetable crops. II. Crucifers, Bot Rev.,
22, 81—166 (1956).
917. Zobel B. The natural hybrid between Coulter and Jeffrey pines, Evolution,
5, 405—413 (1951).
918. Zohar if D., Feldman M. Hybridization between amphidiploids and the evo-
lution of polyploids in the wheat (Aegilops-Triticum) group, Evolution, 16,
44-61 (1962).
919. Zohary D., Harlan /. R., Vardi A. The wild diploid progenitors of wheat
and their breeding value, Euphytica, 18, 58—65 (1969).
32*
Указатель авторов
Айала (Ayala) 153
Александер (Alexander) 444
Аллен (Allan) 191
Андерсон (Anderson) 128, 189, 194—
196, 201, 205, 213, 214, 220
Батталья (Battaglia) 393
Бейкер (Baker) 189
Бейценберг (Beuzenberg) 284, 347
Беннет (Bennett) 94
Бенсон (Benson) 216, 446, 458
Бимен (Beaman) 424
Бинхем (Bingham) 36
Бодри (Beaudry) 85
Браун (Brown) 186, 427
Бреднер (Bradner) 30
Бринк (Brink) 450
Бунгенберг (Bungenberg) 280
Бурма (Burma) 94, 95
Буш (Bush) 148
Бэбкок (Babcock) 415
Бэтке (Baetcke) 200
Бюффон (Buffon) 91
Вагнер (Wagner) 150
Вален (Whalen) 186
Валентайн (Valentine) 153
Вет де (de Wet) 89, 90
Виганд (Wiegand) 194
Викери (Vickery) 135
Винге (Winge) 141, 239, 254, 256,
274, 275
Винклер (Winkler) 274
Виоска (Viosca) 444
Гольдшмидт (Goldschmidt) 168, 169
Готлиб (Gottlieb) 160
Готтшалк (Gottschalk) 276, 453
Гран (Grun) 409
Грант (Grant) 24, 106, 109, 114, 151,
153, 167, 170, 243, 250, 363, 366,
367
Грэй (Gray) 197
Гулд (Gould) 426
Густафсон (Gustafsson) 17, 79, 81,
301—303, 309, 393, 396—398, 401,
404, 412, 419, 423, 424, 440, 441,
448
Дарвин (Darvin) 84, 163, 184, 185,
238
Дарлингтон (Darlington) 391, 411,
439, 453
Девис (Davis) 360
Джайлс (Gilles) 355
Джейн (Jain) 210
Джон (John) 167
Джонсон (Johnson) 17, 305, 306
Джордан (Jordan) 150
Дильс (Diels) 457
Дитрих (Dietrich) 369, 380
Добржанский (Dobzhansky) 51, 55,
56, 150, 153, 160, 167, 170, 184, 247,
276, 278, 286, 289, 292, 296, 301,
302, 310, 349, 362
Доддс (Dodds) 340
ДРУРИ (Drury) 351
Дю Рие (Du Rietz) 51, 199, 227, 237,
457
Дэвидсон (Davidson) 94, 95
Дэй (Day) 65, 66, 325
Гарбер (Garber) 258
Гейтс (Gates) 94, 95
Герасимова (Gerassimova) 243, 254,
255
Герберт (Gerbert) 237
Герстель (Gerstel) 294
Голдблат (Goldblatt) 287
Йертинг (Hierting) 444
Иордан (Jordan) 150
Камерон (Cameron) 450
Кападиа (Kapadia) 426
Указатель авторов
501
Карсон (Carson) 155
Кек (Keck) 48, 276, 289, 290, 295,
296, 338
Кернер (Kerner) 190, 191. 193—195,
237, 238, 244, 457
Ксрстер (Kerster) 183
Кёльрейтер (Kolreuter) 188, 189,
193, 238
Кип (Keep) 127
Кихара (Kihara) 275, 289, 327
Кларксон (Clarkson) 134, 135
Клаузен (Clausen) 48, 107, 134, 135,
138, 276, 289, 290, 295, 296, 338,
406
Клеланд (Cleland) 362, 364, 369-374,
376, 378
Кокрсм (Cockrum) 197
Кровелло (Crovello) 94 95
Кронквист (Cronquist) 84, 94
Крукеберг (Kruckeberg) 48, 135, 165,
193
Кхошу (Khoshoo) 303, 347
Кэйн (Cain) 55, 82
Лампрехт (Lariprecht) 239, 254
Латимер (Latimer) 281
Левин (Levin) 27, 94, 115, 183, 189,
285, 312, 379
Леве (Levy) 276, 280, 304, 305, 410
Лени (Lenz) 134
Линней (Linnaeus) 50, 51, 91, 92,
188, 189, 237, 238, 408, 419
Лотси (Lotsv) 227, 237, 457
Люис (Lewis) 155 156, 167, 169, 276,
280,309,349,351 458
Мазер (Maser) 160, 170
Майр (Мауг) 51, 60, 151, 152, 154,
155, 160, 161, 189, 197, 463
Малсйрос (Malheiros) 358
Марсден Джонс (Marsden Jones) 199
Меглич (Mcgliisch) 70
Мейсон (Mason) 94, 95, 211
Мендель (Mendel) 57, 189, 238
Ментон (Manton) 280, 283, 283, 285,
288, 296, 301, 339, 401, 404
Мёллер (Muller) 309
Молиш (Molisch) 450
Моор (Moore) 197
Москуин (Mosquin) 94
Мур (Moore) 281, 287
Мюнц (Muhz) 369—372
Мюнтцинг (Muntzing) 242, 243, 276,
286, 289, 296, 301, 303
Нобс (Nobs) 108, 123, 125, 126
Нодэн (Naudin) 237, 238
Норденшельд (Nordenskiold) 358,
359, 360
Нюгрен (Nygren) 393, 397
Олгард (011gard) 423
Оливьери (Olivieri) 210
Олстон (Alston) 200
Оно (Ono) 275, 289
Пази (Pazy) 220
Пальмер (Palmer) 229
Патерпиапи (Paterniani) 177
Пинкава (Pinkava) 446
Празмо (Prazmo) 129
Прайор (Pryor) 128
Пэйл ван дер (Pijl) 396
Пэкер (Packer) 306
Радлоф (Radloff) 39
Райли (Riley) 204, 205
Райт (Wright) 33, 148, 154, 164
Рейвн (Raven) 60, 94, 95, 369, 458
Ремингтон (Remington) 189
Роннер (Renner) 361, 364, 367, 368,
376, 377, 380
Реши (Rensch) 51, 150
Риган (Regan) 84
Ригер (Rieger) 276
Ризе (Reese) 305
Ричардс (Richards) 423
Ричардсон (Richardson) 104
Робертс (Roberts) 156, 189, 349
Рудберг (Rudberg) 237
Рутисхаузер (Rutishauser) 393
Рэй (Ray) 50, 91, 460
Рэндолф (Randolph) 205, 335
Сакс (Sachs) 424
Сандфер (Sandfaer) 454
Сауэр (Sauer) 245
Свипгл (Swingle) 431, 433, 435
Симпсон (Simpson) 82, 83, 87, 154
Сннская (Sinskaja) 48
Сирс (Sears) 293
Скалинска (Skalinska) 129, 424
Скуиллас (Squillace) 36
Смит (Smith) 134, 135, 257
Смолл (Small) 444
Сокал (Sokal) 94, 95
Солбриг (Solbrig) 24, 25, 27, 157
Солсбери (Salsbury) 28
Стеббинс (Stebbins) 167, 170, 171,
189, 190, 191, 194, 220, 243, 245,
250, 258, 276, 278, 286, 290, 296,
301, 305, 332, 335, 342, 393, 398,
402, 415, 439, 441, 458
502
Указатель авторов
Стейс (Stace) 189,191
Стенли (Stanley) 463
Такер (Tucker) 245
Танака (Tanaka) 433
Тахара (Tahara) 274
Тернер (Turner) 39
Тогби (Togby) 346
Тодэй (Thoday) 160, 161
Турессон (Turesson) 47
Тышлер (Tischler) 276, 304, 334
Тэкхолм (Tackholm) 385—388
Уайлдер (Wilder) 216
Уайли (Wylie) 453
Уайт (White) 148, 153, 167, 280, 357
Уилсон (Wilson) 169,186
Уокер (Walker) 192, 401
Уоллес (Wallace) 184—186
Фассет (Fassett) 211
Федоров (Fedorov) 281, 287
Фельдман (Feldman) 324, 327
Ферлан (Ferlan) 245
Филлипс (Phillips) 216, 294
Фишер (Fischer) 184, 185
Флейк (Flake) 39
Фокке (Focke) 238, 419
Форд (Ford) 161, 162
Фретер (Freter) 427
Фрикселл (Fryxell) 393
Фриш (Frisch) 182
Фрост (Frost) 450
Фюрнкранц (Fiimkranz) 422
Хаартман (Haartman) 188, 190
Хагеруп (Hagerup) 304, 404
ХайаТа (Hayata) 237, 457
Хаксли (Huxley) 184, 463
Ханельт (Hanelt) 304, 305
Хардин (Hardin) 233
Харлан (Harlan) 89, 90, 327, 329
Хеги (Hegi) 244, 385
Хейзер (Heiser) 189, 191, 194, 210,
303, 458
Хейльборн (Heilborn) 358, 359
Хейр (Heir) 284, 347
Хекард (Heckard) 321, 322
Хёканссон (Hakansson) 387
Хиси (Hiesey) 48, 107, 108, 276, 289,
290, 295, 296, 338
Хоке (Hawkes) 444
Холм (Xolm) 94, 95
Хэббс (Hubbs) 197
Цохари (Zohary) 324, 327, 342
Чезальпино (Cesalpino) 50
Шааль (Schaal) 183
Шафер (Shafer) 128, 129
Штуббе (Stubbe) 369
Эвери (Avery) 104
Эплинг (Epling) 194
Эрендорфер (Ehrendorfer) 307, 319,
458
Эрлих (Ehrlich) 60, 94, 95
Эрман (Ehrman) 176
Эрнст (Ernst) 402
Указатель латинских
названий
Abies 225, 226
— alvordensis 87
— amabilis 87
— concolor 87
— concoloroides 87
— grandls 87
— klamathensis 87
— laiicarpus 87
— magnifica 87
— recurvata 87
— shastensis 87
— sonomensis 87
— toxirious 87
Aceraceae 353
Achillea 47, 192, 323
— asplenifolla 322, 324
— borealis 48, 322
— collina 322, 324
— lanulosa 322
— millefolium 48, 322, 324
— setacea 322, 324
— virescens 322, 324
Acor us calamus 453
Adiantum 436
Aegilops 293, 318, 324, 326, 329
— bicornis 329
— caudata 329
— columnaris 330
— comosa 329
— crassa 330
— cylindrica 330
— juvenalis 330
— kotschyi 330
— mutica 329
— ovata 330
— sharonensis 329
— speltoides 328, 329
— squarrosa 327—329
— triaristata 330
— triuncialis 330
— umbellata 329
— uniaristata 329
— variabilis 219, 220, 330
— ventricosa 330
Agropuron 90
Agrostis 38
— tenuis 36, 37, 48
Alchemilla 396, 402, 425
— arvensis 399
— microcarpa 438
Allium 79, 382, 448
— canadense 448
— carinatum 448, 453
— сера viviparum 453
— flavum 448
— odorum 400
— oleraceum 448
— paniculatum 448
— paradoxum 448
— pulchellum 448
— rubellum 453
— sativum 448
Alnus 279, 396, 402
— rugosa 400
Alpina 423
Alsophila bryophila 246
— dryopteroides 246
Amaranthus 245
— caudatus 245
— powellii 245
— retroflexus 245
Amaryllidaceae 347, 399
Amelanchier 194
Amsinckia 308
Anacardiaceae 400
Anigozanthos 114
Annonaceae 354
Antennaria 398, 402, 403, 425
— alpina 399
— carpatica 400
— porsildii 395
Anthophora 130, 163
Anthoxanthum odoratum 36
Anlirrhinastrum 130
Antirrhinum 101, 130, 141, 166
— glutinosum 101
— majus 101
— orontium 130
Apogon 134, 166, 228
504
Указатель латинских названий
Aquilegia 98, 99, 128, 141, 166, 173,
192, 214, 216, 228, 279
— aipina 99, 128
— canadensis 128
— chrysaniha П, 99, 128, 129
— eccdcarata 99, 128, 129
— formosa 78
— pubescens 78
— vulgaris 99, 128, 129
Arabis holboellii 399, 401
Araceae 400, 453
Arachnion 136, 151, 164, 181, 22%
308, 324
Arbutus arizonica 76
— menziesii 76
Artemia salina 280, 309
Asarina 131
Asclepiadaceae 114
Asclepius 114
Asplenium 285, 319, 436
— monanthes 401
— montanum 319
— pinnatifldum 319
— platyneuron 319
— rhyzophillum 319
Astereae 460
Atraphaxis frutescens 399
Atriplex 139, 142
— litorale 47
— praecox 47
— sarcophyllum 47
— triangularis 139
Aurantioideae 431
Avena 139, 142
— barbata 22, 23
— faiua 22, 23
— disticha 426
— iuncea 426
— media 426, 427
— pedicillata 426
— purpurea 426, 427
— reflexa 426
— triaena 426, 427
— uniflora 426
— vaneedemi 426
— warnockii 426, 427
Brassica 106, 138, 142, 278
— campestris 48, 278
— napus 278
— nigra 48
— oleracea 241, 278
— rapa 31
Bromopsis 331
Bromus 116, 139, 142, 318, 331
— anomalus 332
— arizonicus 330, 331, 332
— carinatus 330, 331, 332
— catharticus 332
— ciliatus 332
— haenkeanus 332
— marglnatus 331, 332
— mollis 22
— orcuttianus 332
— pltensis 332
— stamineus 332
— trinii 332
Buddleja 285
— colvilei 280
Bulbosiylis 360
Burmannia 402
— coelestis 399
Burmanniaceae 399
Buxaceae 400
Balanophora 402
— globosa 399
— japonica 399
Balanophoraceae 399
Baptisia 78, 203
— leucophaea 200
— sphaerocarpa 200
Bembix 30
Berberi daceae 279
Betula 227, 279
Betulaceae 353, 400
Biscutella coronopifolia 296, 297, 339
— laevigata 307, 338, 339
Bombus 30, 130
Bombylius 49
Boraginaceae 33
Bouteloua 426
— curtipendula 321, 350, 426—431
— distans 426
Cactaceae 400, 453
Caesalpinaceae 279
Calamagrostis 398, 402, 403, 424, 425
— arundinacea 407
— canadensis 399
— epigeios 407
— purpurea 407
Californica 228
Calochortus 121
Caltha 47
— palusiris 48
Calylophus serrulatus 380
Camelina saliva 48
Camissonia micrantha 141, 142
Campanula rotundifolia 48
Саппа Indica 453
Cannaceae 453
Cantua 333
Указатель латинских названий
505
Capparidaceae 400
Capparis frondosa 400
Capsicum 116
Cardamine amara 382, 389, 391
— bulbifera 449
— insueta 389—391, 452
— rivularis 389, 391
— schulzii 391
Cai ex 192, 350, 358, 360
— caryophyllea 351
— ericetorum 358
— membranacea 351
— paludivagans 351
— physocarpa 351
— pilulifera 245, 358
— rostrata 245, 351
— r of undata 245, 351
— siderosticta 351
— stenantha 351
Caryophyllaceae 135, 446
Catalpa bignonioides 6, 76
— ovata 6, 76
Catlleya 129
Ceanothus 79, 123, 125, 141, 165, 166,
178, 191, 302, 433
— cuneaius 124
— jepsonii 124
— masonii 124
— pumilus 124
Cclastraceae 400
Celastrus scandens 400
Cephalanthera damasonium 404
— longifolia 404
Cephalaria 345, 347
Cerastes 123—125
Ceratochloa 331
Ceratophora 139, 423
Cercidiphyllaceae 333
Cercidium 114
Chamaemyiidae 280
Cheiranthus cheiri 21, 38
Chenopodiaceae 139
Chenopodium 274, 275
Chloralictus 163
Chondrilla acantholepsis 399
Chrysanthemum 274, 275, 277, 283
— indicum 277
— japonense 277
— lacustre 283
— mankinoi 277
— ornatum 277
— pacificum 277
— yezoensis 277
Cichorieae 333
Cistus 225, 226
Citrus 21,' 398, 400, 411
— aurantifolia 432
— aurantium 432
— celebica 432
— grandis 432, 435
— hystrix 432
— ichangensis 432
— indica 431
— latipes 432
— limon 432
— macroptera 432
— medica 432
— micrantha 432
— paradisi 431, 432, 435
— reticulata 432, 435
— sinensis 431, 432, 435
— tachibana 431, 432, 435
Clarkia 106, 109, 122, 138, 142, 143,
155, 156, 172, 278, 322, 345, 348
— biloba 102, 155—158, 349
— davyi 278
— exilis 155
— franciscana 102, 155, 159
— Ungulata 102, 155—158, 349
— mildrediae 155
— prostrata 278
— rubicunda 102, 155, 159
— speciosa 278
— stollata 155
— unguiculata 22, 155
— virgala 155
— xantiana 36
Claytonia virginica 351, 352, 452, 453
Clusiaceae 400
Clusia minor 400
— rosea 400
Clymenia 431
Cnemidophoras 286
Cobaea 333
Cochliodeae 129
Coleoptera 280
Collinsia 139, 142, 255—257
— corymbosa 258
— heterophylla 36
— multicolor 258
— tincioria 258
Colocasia antiquorum 453
Coluria 132
Comparetticae 129
Compositae 33, 136, 139, 144, 157,
160, 279, 301, 322, 333, 346, 399
Cooperia pedunculata 399
Coprosma 199, 225, 226
Cordia 114
Corixidae 358
Corylaceae 174
Cotoneaster 424, 425
Crassulaceae 285 ' *
Crataegus 402
506
Указатель латинских названий
Crepis 21, 79, 106, 144, 243, 255, 382, 398 — acuminata 399, 415 — baceri 415 — barbigera 416 — capillaris 115 — exilis 416 — foetida 290 — fulginosa 346, 347 — intermedia 416 — modocensis 416 — monticola 416 — neglecta 346, 347 — occidentalis 277, 395, 403, 415— 417, 439 — pleurocarpa 416 — rubra 290 — rubra-foetida 289 — tectorum 115, 255 Crocus 347, 348 — sativus 453 Cruciferae 21, 48, 138, 165, 301, 307, 345, 349, 350, 382, 389 Crustacea 280 Cupressaceae 279, 453 Cupressus 38 Curculionidae 280 Cyatheaceae 246 Cylindropuntia 446, 447 Cyperaceae 356, 357, 359 Cyrtomium caryotideum 401 — falcatum 401 — fortune! 401 Cystopteris 307 Diploxilon 134 Dipsacaceae 345, 347 Diptera 280 Draba verna 140 Dracophyllum 199, 225, 226 Drimys 333 Drosera 325 Drosophila 80 — guarani 179 — melanogaster 161 — nigrospiracula 32 — persimilis 64, 65, 177 — pseudoobscura 32, 61, 64, 65, 177 — willistoni 179 Dryopteris 307, 436 — atrata 401 — borreri 401 — remota 401 Elatostema 396, 402 — acuminatum 399 — eurhynchum 399 — latifolium Eleocharis 356, 358, 360 — palusiris 357 Eleucine 449 Elodea canadensis 20 Elymus 90, 106, 191, 255, 256 — condensatus 443 — glaucus 141, 142, 219, 257, 259 — triticoides 443 Epacridaceae 382, 453 Ephedra 279 Epidendrum 129 Epilobium 116, 143, 192, 245, 337
Dactylis glomerate 342 Dahlia 276 — variabilis 276 Datura 115 — meteloides 115, 311 — stramonium 244 Delphinium gypsophilum 244 — hesperium 244 — moerheimii 453 — recurvatum 244 Dendroblum 130 Deschampsia 449 — alpina 449 — arctica 449 — bottnica 449 — caespitosa 449 Diapensiaceae 343 Dicrurus hottentottus 155 Digitalis 101 Diplacus 40, 79, 192, 216, 228 — longiftorus 36 — angustifolium 338 — crassum 246 — porphyrium 246 — purpuratum 246 Epiphyllum 130, 131, 141 Equisetaceae 333 Equisetum 280, 283, 333, 337 Eragrostis pallescens 296 Erebia tyndarus 179 Eremocitrus 431, 433 — glauca 434 Eriastrum densifolium 49 Erigeron annuus 399 Eriophyllum 345 Erophila 72, 140, 239, 255 — confertifolia 256 — verna 71, 140—142, 256, 350 — violacea-petiolata 256 Erysimum 38 Erythrobalanus 126, 127, 129 Erythroneura 122
Указатель латинских названий
507
Eschscholtzia 308
— californica 36
Eucalyptus 127, 141, 192
— viminalis 36
Euchridium 322
Euchlaena 301
Eugenia malaccensis 400
Eulonchus 49
Euonymus lalifolius 400
Eupatorium glandulosum 399
Euphorbia 401
— dulcis 400
Euphorbiaceae 400
Fagaceae 174, 353
Fagus grandiflora 26
Festuca 79, 449
— microstachys 22
— ovina 20
— rubra 19, 20
Fimbristylis 360
Fitzroya cuprssoides 279
Fortuneila 431, 433, 434
- japonica 434
— margarita 434
Fragaria vesca 48
Freesia 453
— hybrida 453
Fritillaria 310, 345, 347, 348
— pudica 348
Fuchsia encliandra 187
— parviflora 187
Galax aphylla 296, 343
Galeopsis 106, 239, 275, 308
— pubescens 242
— speciosa 242
— tetrahit 241, 242, 300
Galium 300, 310, 318
— anisophyllum 319, 320
— austriacum 319
— incanum 319
— mollugo 296
— multiflorum 319
— pumilum 36, 319
— verum 196
Gambusia affinis 179
Garcinia mangostana 400
Gaura biennis 380
— triangulata 380
Gayophytum heterozlgum 380, 381
Geum 47, 79, 116, 131, 141, 142, 166,
173, 192
— aleppicum 133
— canadense 132
— coccineum 132
— hispidum 132, 133
— intermedium 199
— macrophyllum 132, 133
— molle 132, 133
— montanum 132, 133
— reptans 132, 133
— rivale 48, 78, 132, 133, 199
— rivularis 132, 133
— urbanum 78, 132, 133, 199
Gilia 106, 1009, 115, 116, 122, 142,
143, 151, 173, 179, 180, 214, 216,
226, 228, 243, 255
— achilleifolia 38, 45, 46, 49, 264.
265, 272, 273, 312
— aliquanta 65—68, 140
— angelensis 264, 265, 272, 273
— austrooccidentalls 140, 158
— brecciarum 140
— сапа 225
— capitata 21, 36, 48, 52—55, 136,
180, 182, 264, 265, 271, 273
— clivorum 271, 273
- clokeyi 65—68, 140, 325
- crassifolia 140
- diegensis 140
— inconspicua 139, 140, 142, 173, 218,
259, 307, 324
— interior 140
— jacens 140, 158, 159
— latiflora 44, 45, 151, 153, 225, 325
— leptantha 44, 45, 151, 153, 225
— lomensis 180
- malior 67, 68, 109, 111, 117, 140,
218, 219, 259, 260, 262, 325, 350
— mexicana 140, 153
— millefoliata 180, 272, 273, 312
— minor 65—67, 140
— modocensis 109—111, 117, 140,
218, 219, 259—261, 325, 326, 350
— nevinii 181
— ochroleuca 140, 153, 164, 225
— ophthcdmoides 140, 325, 326
— sinuata 140, 325, 326
— splendens 49
— tenuiflora 153, 158, 159
— tetrabreccia 140
- transmontana 65, 67, 68, 140, 325
— tricolor 53—55, 136, 180, 192, 193,
271, 272
— tweedyi 140
— valdiviensis 312
Gladiolus formosus 453
Gnetum 279
Godetia 322
Gossypium 106, 109, 115, 261, 276,
279, 294
— arboreum 29, 111, 276, 294
508
Указатель латинских названий
— barbadense 101, 276, 294, 336
— herbaceum 276
Gramineae 22, 279, 301, 330, 349, 350,
399, 400
Grevilleoideae 349
Gutierrezia 157
Guttiferae 157
Haemodoraceae 114
Halpopappus gracilis 280
Haploxylon 127
Hartmannia 380
Haynaldia 90, 293
Heliantheae 333
Helianthus 139, 142, 144, 192, 202,
206,-225, 226, 301
— annuus 36, 144, 206, 208, 209, 456
— argophyllus 210
— bolanderi 36, 206, 208—210
— debilis 210
— decapetalus 456
— giganteus 144
— multiflorus 453, 455
— petiolaris 210
Hetnerocallis disticha 453
— fulva 453
Hesperis 334, 349
Heteroptera 358
Hieracium 8, 21, 37, 70, 79, 396, 398
— aurantiacum 397
— umbellatum 36, 401
Hippocastanaceae 333
Hiptage madablota 400
Holcus mollis 443, 452
Hordeum jubatum 22
— vulgare 22
Hosta coerulea 400
Hostia 144
Houttuynia cordata 399, 438
Hyacinthus orientals 453
Hydrophyllaceae 333, 345, 347
Hydrocharitaceae 20
Hypericaceae 425
Hypericum 398, 425
— perforatum 399
Impatiens capensis 25
Inula britianica 453
Ipomopsis 79, 216
— aggregata 77
Iridaceae 279, 347, 353
Iris 115, 130, 134, 141, 166, 202, 203,
214, 220, 228
— bracteata 228
— chrysophylla 228
— douglasiana 184, 228, 229
— fernaldii 228
— fulva 78, 204, 205, 207, 443, 449
— giganiicaerulea 204, 205
— hartwegii 228
— hexagona giganiicaerulea 78, 204,
207, 443, 449
— innominate 228
— macrosiphon 134, 228, 229
— munzii 228, 229
— purdyi 228
— ienax 115, 134, 135, 228
— tenuis 115, 134, 135
— tennuissima 228
— thompsonii 228
Isoetes 280
Isopoda 280
Isopyrum 128, 129
— thaliclroides 129
Ixeris dentaia 399
Juglandaceae 353
Juncaceae 356, 358—360
Juncus 360
Juniperus 192, 203, 213, 220, 228, 279
— ashei 192
— chinensis 456
— scopulorum 211, 213
— virginiana 39, 192, 211—213, 452,
453
Kalanchoe 280, 285
Knautia 144, 345
Kneiffia 380
Labiatae 114
Laelia 129
Laelleae 129
Lavauxia 380
Layia 106, 136, 142
— gaillardioides 117, 138
— glandulosa 138
— heterotricha 138
— hieracioides 117, 138
— pentacheta 138
— platy glossa 138
Leavenworthia 192, 193
Leguminosae 78, 128, 141, 159, 203,
301
Lemna gibba 452, 456
— minor 452, 453, 456
— perpusilla 456
Lemnaceae 453
Leontodon hispldus 404
Lepidobalanus 231
Lepidoptera 229, 280
Lepidoserls 144
Указатель латинских названий
509
Lesquerella 345
Leucopogon juniper us 382, 388, 463
Liatris aspera 33, 61
— cylidracea 33, 34
Liliaceae 321, 333, 345, 347, 453
Lilium 279, 310
— hansonii 104
— lancifolium 453, 455
— mart agon 104
— maximowiczii 456
Limosella 453
Linaceae 349, 400
Linanthus parryae 34
Linaria 237
Linum 349
Listera ovaia 300, 403
Lithospermum caroliniense 33, 34
Loasaceae 141
Loganiaceae 285
Lotus corniculatus 192, 193
Lumbricius 280
Luzula 358
— campesiris 356, 357
— orestra 359
— pallescens 359
— pilosa 359
— purpurea 356, 359
— spicata 359
— sudetica 359
Lycopersicon 109
Lycopodiales 280
Lycopodium 280
Lycoris radiata 453
Lythrum 114
Madia 106, 138, 142, 143, 166, 173,
271, 303, 308
Magnoliaceae 333, 353
Malphigiaceae 400
Malus 398
— hupehensis 399
— sylvestris 453
Malvaceae 279
Mangifera indica 400
Maydeae 349
Medicago coerulea 343
— sativa 30, 31, 343
Melandrium 227, 310
Melilotus 109
Meliphagidae 114
Mentzelia 308
— albicaulis 141, 142
Metasequoia 279
Microcitrus 431, 433
Microseris 139, 142, 308
Mimulus 114, 135, 142, 159, 192, 216,
228 i
— cardinalls 107
— glabratus 135
— guttatus 49, 135, 160
— lewisii 107
— longiftorus 40—44, 77
— nasutus 160
— puniceus 40—43, 77
Могуferas 71, 72, 247,419
Musa acuminata 442, 453, 454
— balbisiana 442, 454
Musaceae 356, 453
Musca domestica 161
Myrtaceae 127, 400
Narcissus 347, 348
— tazetta 453
Nasturtium officinale 453
Nemauchenes 144
Nemophila 333
Nephelea portoricensis 246
Nicotiana 104, 106, 115, 116, 139, 142,
167, 239, 256, 257, 275, 294, 345
— alata 104
— bonariensis 104
— gluiinosa 276
— langsdorffii 104, 115, 257
— longiflora 99
— sanderae 257
— labacum 276
Nigella degenii 38
Nigritella nigra 400
Nothofagus 227
Nothoscordum 398
— fragrans 400, 401
Nuphar 194, 238
Ochnaceae 400
Ochna serrulata 400
Ochthiphila 309
— polystigma 280
Oenothera 270, 362, 369—381
— argillicola 370, 374
— austromontana 370
— biennis 72, 236, 270, 362, 369, 370,
373, 375—377
— brevipes 113
— clavaeformis 113
— elata 370
— erythrosepala 370
— glazioviana 370
— grandiflora 370, 373, 374
— hookeri 364, 369, 370, 373, 375,
376
— irrigua 366
510
Указатель латинских названий
— jamesii 370
— lamarckiana 363—369, 380
— longissima 370
— oakesiana 370
— parviflora 370, 376
— perennis 380
— strigosa 368, 370, 373, 375, 376
— villosa 370
— wolfii 370
Onagraceae 113, 141, 143, 172, 322,
345, 347, 348
Onagreae 138, 381
Oncidieae 129
Oncocyclus 130, 141
Ophioglossaceae 285, 333
Ophioglossum 281, 285, 333
— azoricum 281
— nipponicum 281
— reticulatum 281
— thermale 281
— vulgatum 281
Ophrys 130
— fusca 245
— lutea 245
— murbeckii 245
Opuntia 192, 214, 446
— acanthecarpa 446
— basilar is 453,455
— bigelovii 446
— echinocarpa 446
— edwardsii 445
— ficus-indica 400, 452
— fulgida 247, 445, 446, 449
— kelvinensis 446
— lindheimeri 445
— munzii 446
— phaecantha major 445
— spinosior 247, 445—448
— versicolor 446
Orchidaceae 114, 400
Orchis strictifolius 404
Oreogeum 131, 133
Oryza 109, 260
— glaberrima 260
— sativa 260
Orysopsls 321
Otiorrhynchus 280
Oxitropis 214
Papaver 115
— dublum 192, 193
— lecoquii 192, 193
Papaveraceae 310
Papilionaceae 114
Paris verticillata 453
Parthenium 402, 403, 424, 425, 437
— argentatum 398, 400, 404, 437
— incanum 437, 438
Paspalum dilatatum 400
Passifloraceae 279
Pedicularis 114
— attollens 114
— groenlandica 114
Peloria 237
Peltanthera 130
Penstemon 114, 130, 131, 141, 244
— centranthifolius 131, 244, 245
— grinnellii 131, 244, 245
— spectabilis 131, 244
Pentatomidae 358
Persoonia 345
Persoonioideae 349
Phacelia 345, 347
— californica 322
— heterophylla 321
— nemoralis 322
Phaseolus 109, 239, 243
— coccineus 102, 310
— lunatus 23
— multiflor us 310
— vulgaris 102
Phleum nodosum 453
— pratense 285
Phlox 113, 114, 312
— glaberrima 183
— pilosa 33, 183, 312
Phytodesia 144
Picea 164, 165
— breweriana 87
— engelmannii 87
— glauca 191
— lahontense 87
— mariana 191
— sonomensis 87
— hieracioides 404
Pilosella 399
Pinus 113, 121, 127, 143, 165, 211, 225,
228, 273
— albicaulis 36, 37
— alvordensis 87
— contorta 87
— cotdteri 191
— florissanti 87
— jeffreyi 191
— monticola 36, 87
— muricata 211
— murrayana 36
— ponderosa 77, 87, 127
— quinquifolia 87
— radiata 127
— remorata 211
Pityrogramma triangularis 456
Указатель латинских названий
511
Platanaceae 333
Platanthera chlorantha 404
Platanus occidentalis 6, 112
— orientalis 6, 112
— racemosa 76
— wrightii 76
Plumbaginaceae 399
Poa 21, 79, 284, 396, 398, 438
— aplina 321, 449
— ampla 400, 406, 409
— arida 406
— canbyi 406
— compressa 406
— glauca 399
— irrigata 410
— liiorosa 284
— palustris 438
— scabrella 406, 409
Podocarpaceae 347
Podocarpus 142, 347
Polemoniaceae 22, 44, 45, 52, 53, 135,
273
Polemonieae 33, 307, 333
Polemonium 121
— micranthum 22
Polygonaceae 399
Polygonum viviparum 449
Polypodiaceae 280, 319, 333
Polypodium 285, 307, 333
— vulgare 278, 285, 321
Polystichum 278, 285, 436
Pomoideae 278
Poncirus 431, 433
— trifoliate 433
Populus 279
— aurea 444 t
— barnesii 444
— grandidentata 17, 444, 449
— tremula 452—454
— tremuloides 17, 19, 444, 449
Portulacaceae 351, 453
Potentilla 79, 277, 284, 398, 438
— adscharica 406
— anserina 446, 450
— ar gentea 399, 401
— canescens 406
— glandulosa 36, 48, 101, 109
— gracilis 438
— haematochroa 284
— praecox 399
— recta 406
— tabernaemontana 405
— verna 400
Primula florlbunda 241
— ke we ns is 241, 242, 289—291, 300,
302
— sieboldii 453
— verticillate 241
Primulaceae 453
Prosopis 127, 128, 141
Proteaceae 142
Proteoideae 349
Prunoideae 279
Prunes serrulate 453
Pseudobahia 345
Pseudotsuga menziesii 87
— sonomensis 87
Psilotaceae 333
Psilotales 333
Psilotum 280, 296, 333 '
Psychidae 280
Pteridaceae 456
Pteridophita 279, 280, 288, 296, 301
Pteris 436
— biaurita 437
— confusa 437
— cretica 401
— multtaurita 437
— quadriaurita 437
Purshia 225, 226
Pyrus communis 452, 453
Quercus 79, 121, 126, 141, 143, 165,
192, 193, 226-229, 273, 301
— ajoensis 231
— alba 232—234
— arizonica 231
— austrina 233
— bicolor 233
— chapmanii 233
— Cornelius mulleri 231
— douglasii 126, 216, 230, 231
— dumosa 126, 127, 230, 231
— durandii 233
— durata 126, 231
— engelmannii 230
— gambelii 232, 443
— garryana 126, 127, 230
— grisea 126
— havardii 126, 232
— lobata 126, 231
— lyrata 232, 233
— macrocarpa 232, 233
— margaretta 232, 233
— michouxii 233
— minima 232
— mohriana 126
— muehlenbergil 233
— oglethorpensis 233
— oleoid.es 232
— petraea 234
— prinoides 232, 233
— prinus 232, 233
512
Указатель латинских названий
— robur 233, 234
— rugosa 231
— sinuata 232
— stellata 232, 233
— turbinella 216, 230—232, 443
— virginiana 17, 18, 232
Raimannia 380
Ranunculaceae 128, 399, 453
Ranunculus 79, 398
— acris 192, 193
— auricomus 399, 401
— bulbosus 192
— repens 192
Raphanobrassica 241, 242, 275, 289,
290, 300
Raphanus sativus 31, 241
Rhamnaceae 123
Rhinanthus 101, 114
Rhododendron 194, 238, 244
— ferrugineum 238, 244
— hirsutum 238, 244
— intermedium 238, 244
Rhoeo discolor 381
— spathacea 381
Ribes 127, 141, 303
Rosa 384
— agrestis 385
— canina 7, 236, 249, 270, 277, 382,
384
— chavini 386
— coriifolia 386
— dumetorum 385, 386
— junzillii 386
— micrantha 385, 386
— mollis 386
— pomifera 386
— pouzini 386
— rubiginosa 387
— rubrifolia 385, 386
— stylosa 386
— tomentella 385, 386
— tomentosa 386
Rosaceae 131, 279, 382, 399, 453
Rubi Corilifolii 420, 421
Rubiaceae 279, 319
Rubus 70—72, 79, 194, 308, 399, 402,
418
— acutus 407, 408, 410
— allegheniensis 420, 421
— alnicola 420
— bollei 420
— caesius 406—408, 421
— gothicus 408 *
— idaeus 421
— incanescens 420
— insularis 406
— moschus 420
— plicatus 406, 407
— polyanthemus 406
— polycarpus 408, 410
— prulnosus 410
— ruedensis 408
— tomentosus 420
— ulmifolius 420
— wahlbergii 421
Rudbeckia californica 399
Rumex 310
— acetosa 311
— thyrsiflorus 311
Rutaceae 400, 431
Salicaceae 279
Salix 199, 227, 279, 310
— daphnoides 191
— incana 191
Salmonidae 280
Salvia 113, 114, 194, 238
— apiana 112, 117, 163, 194
— melllfera 112, 117, 163, 194
Sanicula bipinnatifida 319
— crassicaulis 319
— hoffmannii 319
— laciniata 319
Sarcococca pruniformis 400
Saururaceae 399, 438
Saxifraga cernua 449
— ferruginea 448
— foliolosa 448
— stellaris 448
Saxifragaceae 127
Scilla 333
Scirpus 356
Scrophulariaceae 40, 77, 114, 131, 135,
160, 349, 453
Secale 90
— cereale 23, 291, 329
Sedum 192
Selaginella 288
Senecio 284
— jacobaea 28
— robertifriesii 284
Sequoia sempervirens 279, 301—302
Silene 135, 141, 192, 193
Slmlolus 135, 142
Sitanion hystrix. 258 ,
— jubatum 258, 259
Smilacina racenwsa 400
Solanaceae 133,309, 345, 453
Solanum 116, 133,-142, 186, 274, 309
— acaule 340, 341, 342 ,
— chacoense 444, 445, 449
Указатель латинских названий
513-
— citrullifolium 186 — demissum 342 — grayi 186 — lumholtzianum 186 — microdontum 449 — nigrum 274 — roslratum 186 — stenotomum 341, 342 — tuberosum 297, 339—342, 445, 450 Solenobia triquetrella 280 Sorbus 403 — chamaemespilus 399 — hybrida 399 Sorghum 301 Spartina patens 20 Spathiphyllum patinii 400 Spergularia 142 Spiraeoidea 279 Stanhopea 114 Statice olaeefotia confusa 399 Stellaria crassipes 446 — longipes 446 Stephanomeria 139, 142, 160 — exigua coronaria 160 — malheurensis 160 Sterculiaceae 142 Stipa 321 Streptanthus glandulosus 48, 165 Streptocarpus 116 Styphelieae 389 Succisia 347 — pratensis 102 Tricher a 144 Tricheroides 144 Trichocentreae 129 Trichoniscus elizabethae 280 frifolium pratense 190 — repens 190 Trillium 333 — kamtschatcense 321 Triphasia aurantiola 400 Triticale 329 Triticum 109, 167, 277, 293, 318, 32b, 327 — aestivum 90, 327—329, 336 — durum 334 — monococcum 90, 327, 329, 334 — speltoides 328 — tauschii 328 — titnopheevii 90, 327, 329 — turgidum 90, 327, 328 — vulgare 327 Trichodendraceae 333 Tsuga heterophyIla 87 — mertensiana 87 — mertensioides 87 — sonomensis 87 Tulipa 279 — gesneriana 400, 453 Ulmaccac 353 Umbelliferae 319 Urginea indica 453
Tanysiptera galatea 60, 155 — hydrocharis , Taraxacum 8, 21, 79, 396, 398, 402, 410, 422 — dovrense 439, 440 — kok-saghyz 422 — officinale 422, 423, 455 — pieninicum 422 — reichenbachii 439, 440 Tasmannia 333 Taxodiaceae 279 Thalictrum 279 Theobroma 142 Thuja pile at a excelsa 453 Thymeliaeceae 399 Tmesipteris 280, 333 — tannensis 281 Tofieldia 333 Townsendia 425 Tradescantia 225, 226 Tragopogon 239 — dubius 102 — porrijolius 102 Urticaceae 399 Vaccinium uliginosum 296 Vanda 130 Verbascum thapsus 28 Vernonia 135, 142 Veronica 349 Vicia 159 — sativa 141, 142 Vinca major 14 Viola 100, 167, 192 — arvensis 100 — tricolor 100 Welwitschia 279 Wigandia 333 Wikstroemia 402 — viridifolia 399 Winteraceae 333 Woronowia 132
33-45
514 Указатель латинских названий
Xanlhoxylum bungoi 400
Xylocopa 130, 131. 163
Yaungia 345
Zausrhneria 300
Zea 109, 110,
— mays 177. 311, 335
— perennis 296
Zepkyranihes 402
— texana 399
Zeuxine sulcata 400
Zingiberaceae 356
Z>ruua 192
Zygcpelalum mackayl 400
Предметный указатель
Аберрантные признаки в перифериче-
ских популяциях 154
Автогамия 15, 16, 21—24, 139—141,
159-160,270 п _
Автополиплоидия 275, 299, 338—343
Автосегрегацня 410
Агамный комплекс 267, 415—441
---- стадии развития 437—439
----филогенетический анализ 414,
418
----эволюционный потенциал 439—
441
Агамоспермия 15—16, 20—21, 247,
248, 269, 392—413, 454, 455, 458
— облигатная 21
— факультативная 21
Агматоплондия 344, 354—360
Адаптивные генные комбинации, рас-
пад 117
Адвентивная эмбриония 20, 393—
396, 405, 433—436
Аддитивность 102, 103
Аллопатрия 118, 119
Аллополиплоид геномный 290, 291
— сегментный 290, 291, 295, 296
Аллоплоидия см Амфиплоидия
Амфибии 280, 286
Амфиплоидия 236, 240—242, 267,
275, 298-299
Анеуплоидия 344—354 См. также
Агматоплондия
— восходящая 347—349
— вторичная 431
— нисходящая 344—347
— полиплоидный уровень 347—349,
410, 413, 427—431
Анеуплоидный ряд 287
Апельсин 431—436
Апогаметия 395, 397, 436, 437
Апомиксис 247, 248 См также Ага-
моспермия, Вегетативное размно-
жение
— геметофнтный 393—397, 404—405,
424
— облигатный 397
— факультативный 397
Апоспория 394—397, 404
Асинапсис 311
Ассортативное скрещивание 147, 148,.
161, 162
Ауткроссннг облигатный 308
Банан 455, 456
Барвинок 14
Барьеры несовместимости 177, 179—
181
— этологические 177
Биотипы малые 414
Биосистематические категории 82, 88,
89
Блоки генов 262, 367, 368
Бук 26
В-хромосомы 344
Вегетативные спорты 410
— химеры 274
Взаимоотношения по плодовитости
123, 141—145
--------модели 141—143
Видовая популяция, гомогенность
59—63, 73—74
Видообразование 146—149, 150—
166, 268—269
— аллопатрическое 157, 160
— апомиктическое 247—248
— биотически-симпатрическое 160—
163
— географическое 147, 150—153
— гибридное 235—240, 244—246, 248,
249
— квантовое 147—149, 155—157
— парапатрическое 148
— первичное 151—155
— рекомбинационное 242—244, 248
250-265
— симпатрическое 147, 148, 160—163
— способы 146—149
— стасипатрическое 148—149
33’
516
Предметный указатель
Виды аллопатрические 85, 86
— биологические 45, 50—68, 82, 85,
86, 91—97
— генофонд 59
— двойники 64—68, 92—94, 315, 316
— морфологические 82
— последовательные 82, 86—87
— симпатрические 85, 86, 119, 121,
122
— таксономические 82—86, 91—94
— эволюционные 83, 87, 88
Вицинизм 15
Водоросли 280
Выщепенцы гомологичные 246
Гаметный фонд 200
Гаметофитный апомиксис 393, 397,
404, 405, 424
Генетическая идентичность, показа-
тель 102
Генетические системы, специализиро-
ванные 361—364
---Oenothera, 362—367
Генетический дрейф 38
---сочетание с отбором 38
Генетическое равновесие ПО, 111
Генная регуляция 169
Генные системы 98—102, 106, ПО—
111
— — сериальные 98—102
Генотип интегрированный ПО, 111
Генофонд 59, 89—90
— вторичный 90
— первичный 89
— третичный 90
Гены, блоки 367, 368
— жизнеспособности 17—110
— множественные 98, 99
— модификаторы 98—101
— морфологические 107—ПО
— мутаторы 169
— оппозиционные 98—102, 103
— поток 28—30, 33—36, 59—63, 74,
156—157
— регуляторы 103, 104, 169
Гетерогамный комплекс 268, 362,
369—375, 380—382, 384—388, 391,
392
---эволюционный потенциал 391,
392
Гетерозиготность структурная 253,
254, 267, 268
Гетерозис 298—300, 411, 412
Гибридизация 72—79, 188—198, 225—
226
— ^естественная 188—190, 227
— и агамоспермия 402—404
— и микровиды 72, 73
— интрогрессивная 225, 226
— и полувиды 77—79
— местообитания 195, 196
Гибридная мощность 298—300
— несовместимость 179—181
Гибридность нумерическая 237, 249
— структурная 237, 249
Гибридные полувиды 79—80
Гибридный индекс 201—203
— комплекс 267—273
Гибрид-сипгамеоны 227
Гибриды, нежизнеспособность 116
— разрушение 263—265
— скопления 78, 190—192
— спонтанные 190—192
— стабилизация воспроизведения
235—237
— стерильность 116, 117
Гинкго 279
Гомеологичные хромосомы 240, 292,
336, 349
Гомеостаз физиологический 298
Гомогамный комплекс 267, 268, 270—
273, 458—459
Гомозиготпость структурная 253, 254
Гомоплоидные выщепенцы 246
— гибриды 262
— линии 250, 262
Грейпфрут 407—408, 432—436
Грибы 280
Делеция 104
— интерстициальная 105
Дивергенция 85, 86, 163—165, 458,
462
— эволюционная 117
— экологическая 122
Диплоидизация 335—337
Диплоспория 394—397, 404
Дисгармония генная 138
Дисперсность (зернистость) среды 35
Дифференциальное сродство 240, 241
Дифференциация расовая 33, 34
— микрогеографическая 36—38
— экологическая 119, 120
Дождевые черви 280
Дрейф генетический 38
Дронго 155
Дубы белые 229—234
— черные 229
Дупликация 105, 240, 299
Ежевика 247, 418—422
— микровиды 418, 419
Предметный указатель
517
— уровни плоидности 419, 420
Естественная гибридизация 77—79
— система классификации 93
Живорождение ложное 16, 396, 448,
449
Жизненная форма 143—145, 301. 302
Жорданон 71
Жужжала 49
Закрытое сообщество 26, 27
Замещение видов Г21
Зимородок ракетохвостый 155
Зона вторичной интерградацни 190
Идентичность генетическая 102
Иерархия таксономических катего-
рий 83—86
Изменчивость анеуплоидная 351, 352
— географическая 39—44
— клинальная 39
— подавление 334, 335
— электрофоретическая 102, 103,
203, 379
Изолирх тощие механизмы 51, 89,
112—117, 163—166, 204—210
---- классификация 113
---- пространственные 112—113
----репродуктивные 112—117
----средовые 112—ИЗ
Изоляция пространственная 151
— репродуктивная 69, 151, 163—166
— сезонная 163
Изоферменты 20, 102, 103, 203
Инверсии 104—105, 170, 347, 349
Индекс гибридный 201—203, 208
Интерградация 43
Интрогрессивная гибридизация 199
Интрогрессия 199—226
Ирисы 134-135, 194, 204—206,
228-229
Искусственная система классифика-
ции 93
Искусственный отбор 177—178
Кактусы 247, 445
— кустарниковые 446—447
Картофель 340—342, 451. См так-
же Solanum
Классификация полиплоидов 289—
291
Клещи 356
Клины 39—44
Клональные комплексы 267, 442,
445—449
Клоны 17—20, 442—444
Клопы 356
Колибри 129, 131
Кольца хромосомные 361—369
Комары 122
Компариум 89
Комплексы агамные 393
— гетерогамные 268, 362, 369—375,
380—388, 391—392
— гибридные 267—273, 442—444,
458—459
— гомогамные 267—268, 270—273,
458—459
— клональные 267, 442, 445—449
— полиплоидные 315—344
Конвергенция 461—462
Конкурентное исключение 121
Конкуренция межвидовая 119—121
Концепция вида, биологическая 94—
97
--- дуализм 93
---номиналистическая 95
— генетических революций 154
— стабилизации гибридности 411—
412
Конъюгация гетрогенетическая 292
— юмогенетическая 292
— межтеномная 392
— нарушения 311
Критерий ассоциации признаков 201
Кроссинговер 223—224
Кукуруза 31, 311, 454
Культивары 90, 327, 342, 435, 454—
_ 456
Культурные растения, единицы клас-
сификации 89—90
К-отбор 27
Лайм 432—434
Летали гаметофитные 366
— зиготные 366
— сбалансированные 362, 365—366
370
Лимоны 407—408, 431—436
Локальные расы 28—38, 150
Люперна 30—31
Макрорекомбинация 101—102
Макроэволюция 457—463
Малина 421
Мандарин 432—436
Межвидовая конкуренция 119—121
Межсекционные гибриды 125
518
Предметный указатель
Мейоз постредукционный 360
Местообитание открытое 25—27, 196,
200
— гибридизация 195—196
Микровиды 71—73, 418—419
— автогамные 71—73
— агамоспермные 21, 72, 79, 237,
411-412, 421, 424
— гетерогамные 71—72, 362, 376—
382
— гибридные 72—73
— клональные 71—72, 237, 442—444
— локальные 420
Моносомия 345
Мутации почковые 410
Мхи 280
Наследование блочное 261—263
— тетрасомное 293—294
Нежизнеспособность гибридов 116
Нехватки 240
Ниша пищевая 122
— экологическая 118, 461—462
Обмен генами, блокирование 112
Оплодотворение перекрестное 15—16
Опознавательные знаки цветков 182—
184
Опунция 445, 450
Опыление 29—31, 49, 113—115, 128—
131, 185-187
Орхидеи 129—130, 166, 404
Отбор дизруптивный 37, 148, 161,
162
— катастрофический 156
— межвидовой 119—123
— стабилизирующий 69, 195
— типы г- и К- 27
Открытая система роста 80—81
Открытое местообитание 25—27
— сообщество 26—27
Осина 17—19, 444
Осы-псевдомазориды 131
Палеонтологическая летопись 86—87
Панмиксия 15—16
Панмиктическая единица 31—33
Папоротники 192, 246, 269, 285, 296,
307, 319, 397—401, 404, 436—437,
440
— апогамные 401, 437, 440
Парапатрия 118—119
Партеногенез 20, 394—400, 404—405
Перекрестное оплодотворение 15—16
Перепончатокрылые 30
Перестройки независимые 218
Перетяжка центромерная 356
Перманентная гетер ознготность по
транслокациям 249, 267—270, 361—
381
— гибридность 249, 267
— нечетная полиплоидия 249, 267,.
383—392, 456
Печеночники 280
Подсолнечник 206—210
Полигаплоиды 334, 342
Полиморфизм 39—44, 56, 150, 160—
163
— по ферментам 102—103
Полиплоидизация непрерывная 315
Полиплоидия 271—276„ 298, 301, 396
— дибазическая 351
— и жизненная форма 301—302
— монобазическая 351
— негибридная 275
— нечетная 458
— распространение 279, 280
— частота 286—288
Полиплоидный ряд 340
-----пшеницы 328
— сброс 278, 284-285, 334, 344
Полиплоидные комплексы 267, 315,
430
-----генетико-таксономический ана-
лиз 317—318
-----идентификация 315—316
-----методы анализа 315—318
-----остатки древних 332—334
-----старые 330—332
----- структура 316
-----филогенетический анализ 317
-----цитотаксономический анализ
316—321
Полиплоиды дибазические 278
— подавление изменчивости 334—335
— способы образования 299—300
— типы 289—297
Полисомия 344
Полиспермия 300
Полицентрические хромосомы 356—
358, 360
Полиэмбриония 433
Половое размножение 13—15, 55—
58
Полувиды 75—78, 151—152, 227—228
Померанец 432—434
Популяции дизъюнктивные 76
— периферические 153—155
- природные 178—179
— производные 236—237
— степени интеграции 70—71
Предметный указатель
519
Поток генов 28—31, 33—36, 59—63,
74, 156—157
Почковые мутации 410, 436
Предпочтительная конъюгация 241
Прерывистая эволюция 463
Прививки 274—275
Признаки, связность 201—203
— смещение 120, 186—187, 225—226
Принцип основателя 154
Природные автополнплоиды 338—343
Псевдогамия 20, 396
Пчелиные 130
Пчелы 113, 117, 163, 182-183
— каменщицы 131, 163
Пшеницы 326—330
— геномная конституция 327
— геномный анализ 327—328
— система классификации 327
— филогения 328—329
Равновесие генетическое 110—111
Размножение бипаренталь ое 16, 69-
72, 75
— вегетативное 14—20, 247, 267—
268, 442—443
— половое 13—15, 55—58
— унипарентальное 16, 69—72
Разрушение гибридов 112—113
Расообразование 225—226
— гибридное 235
Расселение 28—31, 61—63
Расы биотические 48
— географические 38—49, 76
— дизъюнктивные 44—45, 189—190
— интрогрессивные 225
— локальные 28—38, 86, }50
— перекрывающиеся 77
— эдафические 37
— экогеографические 76
— экологические 45—47
Регуляция 103—104
Редукция числа хромосом 345—317
отсутствие 445
Редька 31
Рсинверсии 252—253
Рекомбинационная система 24—27
— - закрытая 25—26
----открытая 26
---- экология 25—27
Реннера комплекс 361—369
Репродуктивная изоляция 89
----внешняя 113—115
----внутренняя 115—117
Ресинтез полиплоидов 317, 325
Реципрокные транслокации 361
Рожь 291
Роза собачья 382—388
Рыбы 280
Ряд анеуплоидный 350—353
— полиплоидный 276—279, 359
г-Отбор 27
Саговники 279
Самонесовместимость 308—309
Самоопыление 158
Сахарный тростник 451
Сбалансированные летали 362, 365—
366, 370
Связность генетическая 106—107,
213—215
— признаков 201—203, 213—215
Сетчатая филогения 457—458, 459—
461
— эволюция 457—459
Симпатрия 118—119, 121—122
— биотическая 119, 122
— смежная 119
Сингамеон 79, 86, 153, 227—234, 432
Синтетические амфиплоиды 294
Система(ы)генная 98—102, 106—107
— классификации 93
---- искусственная 93
----филогенетическая 460
— размножения 13—27, 308—310
— рекомбинационная 24—27
— роста открытая 80—86, 459, 460
— скрещивания 15—16, 21, 143—147,
223—224, 235—236, 268—270, 401—
402
,— эволюции закрытая 459
----открытая 459
Системные мутации 168—169
Скопления гибридов 78, 190—192,
402
Скосарь турецкий 309
Скрещивание ассортативное 147—148,
161—162
— случайное 15—16
Смещение признаков 120, 186—187»
225—226
Соматическое удвоение 299—300
Сообщество закрытое 26—27
— открытое 26—27
Соседства 31—34
Сосна 29, 211 См также Pinus
Сосуществование 118—119, 120—121
Спаривание избирательное 334
Специфичный кариотип 167
Среда, дисперсность 35
Стабилизация воспроизведения гибри-
дов 235—237
520
Предметный указатель
— гибридности 411—412
Старение клонов 450, 451
Стерильность гибридов 115—117
Структурные гомозиготы 171—172
Сцепление 106—110, 170—171, 201—
203, 213—215, 217—220, 262—263
— многофакторное 106—107
— M—V 107—110, 217—220, 260—
262
Тетрасомия 341
Тетрасомные хромосомы 348
Тимофеевка 285—286
Трансгрессия барьеров стерильности
220—224
Транслокации 104—105, 170, 250—
252, 258, 347—349
— плеча хромосомы 105
— последовательные 362
— реципрокные
Транспозиции 105, 252—253
Триплоиды 389—391, 452—456
— возникновение 452—454
Трисомики 368
Удвоение вторичное 334
— соматическое 299—300
Уоллеса эффект 185—187
Уровни плоидности 281—286
Факторы отбора 47—49
Фил ада 143
Филетическая эволюция 461—462
Филогенетическая сеть 374—375
— схема 273
Флоксы 183
Фрагменты ацентрические 356—357
Хвойные 279
Хемотаксономические методы 321
Химеры 274
Хлопчатник 336
Хроматография на бумаге 321
Хромосомные кольца 361—369
— перестройки 104—106, 116—117,
167—173, 220—224, 240—242, 250—
254
— числа, диапазон 280—281
----- направления 170, 360
-----основные 276—279, 280—281,
333, 345—349
-----у агамоспермных растений
399—401, 415—418
Хромосомы, величина 310—311
— гомологичные 240, 292, 336, 349
— добавочные 344
— основное число высокое 353—354
-------низкое 357—358
------- первоначальное 354
-------редукция 345—347
— полицентрические 356—358, 366
-----фрагментация 356—360
— структурные изменения 167—174
— тетрасомные 348
— терминальные участки 364, 373
Ценовиды 82, 88—89
Центромера диффузная 355—358,
359—360
Цнкадки 122
Цикл дифференциации — гибридиза-
ции 458
Цитоплазматические факторы 98
Цитрон 432
Чехлоноски 309
Чешуекрылые 122
Число хромосом основное 276—281,
333, 345—349
-----удвоение 274—275
-------соматическое 275
------- спонтанное
Шмели 129, 340
Эволюционная дивергенция 117
Эволюция конвергентная 461—462
— филетическая 461—462
Эгилопс 326
Эковиды 82, 88—89
Экология рекомбинационных систем
25—27
Экологическая ниша 118, 461—462
Экологические адаптации 328—329
Экотип 88, 328—329
Электрофорез 203
Электрофоретическая изменчивость
102—103, 203, 379 ч
Эмбриония адвентивная 433—436
— нуцеллярная 435—436
Эндомитоз 310
Эффект положения 167—170
— последовательности участков 168—
174
— Уоллеса>Л85—187
Ячмень 454
Ящерицы 280
Оглавление
Предисловие редактора перевода..................5
Предисловие к русскому изданию..................10
Предисловие.....................................П
Часть первая. Популяции и расы
Глава 1. Размножение растений..................13
Отклонения от случайного перекрестного оплодотво-
рения .........................................15
Вегетативное размножение.......................16
Агамоспермия...................................20
Автогамия......................................21
Рекомбинационные системы.......................21
Экология рекомбинационных систем ... 25
Глава 2. Локальные расы........................28
Расселение и поток генов.......................28
Панмиктические единицы и соседства .... 31
Поток генов и расовая дифференциация ... 33
Взаимодействие между Потоком генов и отбором 34
Микрогеографическая расовая дифференциация 36
Сочетание отбора с генетическим дрейфом . . 38
Глава 3. Географические расы...................39
Непрерывная географическая изменчивость . , 39
Дизъюнктивные географические расы ... 44
Экологические расы.............................45
Факторы отбора, участвующие в формировании рас 47
Часть вторая. Природа видов
Глава 4. Биологический вид.....................50
Пример.........................................53
Биологический вид как естественное следствие по-
лового размножения в биотическом сообществе . 55
522
Оглавление
Концепция генофонда вида......................
Поток генов и гомогенность видовых популяций
Виды-двойники.................................
Глава 5. Видовые единицы у унипарентальных
растений..................'...................
Системы унипарентальных популяций ....
Степени интеграции............................
Микровиды.....................................
Гибридность и микровиды.......................
Пересмотр вопроса о гомогенности вида
Глава 6. Вопрос о виде у растений.............
Полувиды как результат постепенного видообразо-
вания ........................................
Полувиды как результат естественной гибридизации
Масштабы проблемы вида........................
Причины и следствия, вытекающие из концепции
вида у растений...............................
Глава 7. Разные типы видов....................
Типы видов ...................................
Таксономический вид...........................
Биологический вид как одна из стадий процесса ди-
вергенции ....................................
Последовательные виды......................
Эволюционный вид..............................
Биосистематические категории..................
Единицы классификации культурных растений
Биологический вид или таксономический вид?
Критика биологической концепции вида
Часть третья. Генетические и экологические аспекты вида
Глава 8. Генетическая основа различий между
видами .......................................
Генные системы................................
Гены, кодирующие ферменты.....................
Замечания относительно регуляции ....
Хромосомные перестройки.......................
Системы сцепления.............................
Многофакторное сцепление.....................
Сцепление М—V............................. .
Интегрированный генотип.......................
Оглавление
523
Глава 9. Изолирующие механизмы...............112
Основные формы изоляции......................112
Внешняя репродуктивная изоляция . . . .113
Внутренняя репродуктивная изоляция . . .115
Выводы...................... .... 117
Глава 10. Экологические взаимодействия . . .118
Сосуществование...............................ИЗ
Эффекты межвидового отбора .... .119
Сосуществование без межвидового отбора или за-
мещение видов.............................. .120
Эффекты экологических требований ... 121
Глава 11. Формы взаимоотношений по плодовитости 123
Древесные растения...........................123
Многолетние травянистые растения с резко выра-
женными различиями в механизме цветка . .128
Многолетние травянистые растения с ярко выражен-
ными межвидовыми различиями в механизме
цветка.......................................131
Однолетние травянистые растения..............135
Автогамные однолетники.......................139
Общие модели.................................141
Общие корреляции.............................143
Выводы..................................... 145
Часть четвертая. Эволюционная дивергенция
Глава 12. Способы видообразования 146
Силы и поля...................................146
Способы видообразования .... 147
Обсуждение....................................148
Глава 13. Первичное видообразование 150
Географическое видообразование .... 150
Квантовое видообразование . 153
Квантовое видообразование у Clarkia . .155
Аллопатрическое видообразование, связанное с са-
мооплодотворением ........................... 157
Биотически-симпатрическое видообразование . . 160
Репродуктивная изоляция как побочный продукт
дивергенции...................................163
Выводы....................................... 165
524
Оглавление
Глава 14. Структурные изменения хромосом при
видообразовании................................167
Теория эффекта последовательности участков в хро-
мосомах .-.............................168
Теория, основанная на сцеплении генов . . . 170
Закрепление новых структурных гомозигот . .171
Выводы.........................................173
Глава 15. Отбор на репродуктивную изоляцию
Особые условия.................................175
Эксперименты по искусственному отбору . .177
Данные по природным популяциям .... 178
Барьеры несовместимости у Gilia................179
Опознавательные знаки цветков..................182
Исторические аспекты...........................184
Эффект Уоллеса и смещение признаков . . .186
Часть пятая. Естественная гибридизация и ее результаты
Глава 16. Естественная гибридизация . . . .188
Гибридизация как процесс, обратный дивергенции 189
Гибриды и скопления гибридов .................. 190
Внутренний контроль над гибридизацией . . .192
Внешний контроль над гибридизацией . . .193
* Гибридизация местообитания.........................195
* Сопоставление высших растений с высшими живот-
ными .......................................197
Глава 17. Интрогрессия........................199
Факторы, способствующие интрогрессии . . .199
Методы .... 201
Примеры.......................................204
Связность признаков как критерий гибридности 213
Экологическая сегрегация морфологических типов 215
Сцепление М—V в связи с интрогрессией . .217
Трансгрессия барьеров хромосомной стерильности 220
Интрогрессия и расообразование................225
Выводы................'.......................226
Глава 18. Сингамеон...........................227
Тихоокеанские ирисы....................- . . 228
Белые дубы....................................229
Глава 19. Гибридное видообразование . . . 235
Стабилизация воспроизведения гибридов . . . 235
История вопроса...............................237
' Оглавление 525
Введение в амфиплоидию...................240
Введение в рекомбинационное видообразование 242
Гибридное видообразование при наличии внешних
барьеров.................................244
Введение в апомиктическое видообразование . 247
Выводы...................................248
Глава 20. Рекомбинационное видообразование 250
Гипотеза........................... . 250
Экспериментальный подход......................254
Ранние эксперименты . 255
Последующие эксперименты при существовании
слабых барьеров стерильности..................255
Более поздние эксперименты при существовании
мощных барьеров стерильности..................258
Роль блочного наследования в рекомбинационном
видообразовании.......................... . .261
О роли рекомбинационного видообразования в при-
роде .........................................263
Глава 21. Гибридные комплексы.................266
Классификация.................................267
Роль системы размножения......................268
Гомогамный комплекс.......................... 270
Гомогамный комплекс облиственностебельных
гилий.........................................271
Часть шестая. Полиплоидия
Глава 22. Полиплоидия: распространение и частота 274
Исторический очерк............................274
Масштабы проблемы и ее суть...................275
Полиплоидные ряды........................... 276
Распространение полиплоидии в разных таксономи-
ческих группах................................270
Диапазон хромосомных чисел....................280
Диапазон уровней плоидности...................281
Частота.......................................286
Глава 23. Типы полиплоидов....................289
Разработка классификации......................289
Цитологические критерии........................291
Генетические критерии..........................292
Морфологические критерии.......................294
526
Оглавление
Относительная частота автополиплоидов и амфи-
плоидов в природе ..........................
Глава 24 Факторы, способствующие возникновению
полиплоидии ................................
Преимущества, внутренне присущие амфиплоидно-
му состоянию....................... . . •
Способы образования .......................
Полиплоидия и жизненная форма..............
Факторы, имеющие первостепенное значение .
Широта и высота местности..................
Экологические факторы......................
Полиплоидия и система размножения ....
Влияние величины хромосом...................
Влияние генотипа...........................
Общие выводы................................
Глава 25. Полиплоидный комплекс ....
Методы анализа..............................
Цитотаксоном ические примеры................
Биосистематические примеры..................
Комплекс Gilia inconspicua..................
Triticum и Aegilops .........
Старые полиплоидные комплексы...............
Остатки древних полиплоидных комплексов
Необратимость...............................
Подавление изменчивости.....................
Диплоидизация ..............................
Глава 26. Природные автополиплоиды
Группа Biscutella laevigata.................
Группа Solanum tuberosum....................
Группа Dactylis glomerata...................
Другие примеры..............................
Часть седьмая. Анеуплоидия
Глава 27. Анеуплоидия.......................
Нисходящая анеуплоидия на диплоидном уровне
Восходящая анеуплоидия на диплоидном уровне
Полиплоидный сброс .........................
Анеуплоидное увеличение у полиплоидных гибридов
Carex ...........
Claytonia virginica .........
Высокие основные числа у покрытосеменных
Оглавление
527
Глава 28. Агматоплоидия.......................’ 355
Полицентрические хромосомы . ... 355
Carex..........................................358
Luzula.........................................358
Сурегасеае и Juncaceae ....................... 359
Часть восьмая. Специализированные генетические системы
Глава 29. Перманентная гетерозиготность по
транслокациям ................................361’
Генетическая система у Oenothera............. 362'
Источники изменчивости.........................367
Гетерогамный комплекс в группе Oenothera hookeri-
biennis........................................369
Гетерогамные микровиды в группе Oenothera bien-
nis ........................................ 376>
Гетерогамные микровиды и комплексы в других
группах растений............................. 380s
Глава 30. Перманентная нечетная полиплоидия 382
Генетическая система Rosa canina..............382
Гетерогамный комплекс в группе Rosa canina 384
Leucopogon Iuniperinus........................388
Cardamine insueta............................. 389
Эволюционный потенциал гетерогамных комплексов 391
Глава 31. Агамоспермия........................398
Эмбриологические путы.........................393
Таксономическое распределение.................397
Система размножения...........................401
Гибридность и агамоспермия ... . 402
Возникновение гаметофитного апомиксиса . . 404
Поведение при скрещиваниях v..................405
Расщепление...................................407
Характер изменчивости.........................409
Преимущества агамоспермии.....................411
Глава 32. Агамный комплекс .... . 414
Комплекс Crepis occidentalis.....................415
Европейские ежевики..............................418
Taraxacum........................................422
Агамные комплексы типа Rubus Moriferus . . 424
Комплекс Bouteloua curtipendula..................426
Citrus......................................... 431
4528 Оглавление
Pieris.......................................436
Стадии развития..............................437
Эволюционный потенциал.......................439
Глава 33. Вегетативное размножение гибридов 442
Гибридные клоны..............................442
Клональные микровиды.........................444
Клональные комплексы, размножающиеся ответ-
влениями стебля . 445
Клональные комплексы, размножающиеся при по-
мощи ложного живорождения....................448
Эволюционный потенциал.......................449
Глава 34. Естественные триплоиды .... 452
Возникновение .............................. 452
Сохранение...................................454
Избранные примеры............................455
Часть девятая. Заключение
Глава 35. Вид и макроэволюция...............457
Сетчатая эволюция...........................457
Эволюционный потенциал гибридных комплексов 458
Значение сетчатого характера эволюции для фило-
гении покрытосеменных.......................459
Вид как единица макроэволюции...............462
Литература..................................464
Указатель авторов ......................... 500
Указатель латинских названий................503
Предметный указатель........................515