




































































































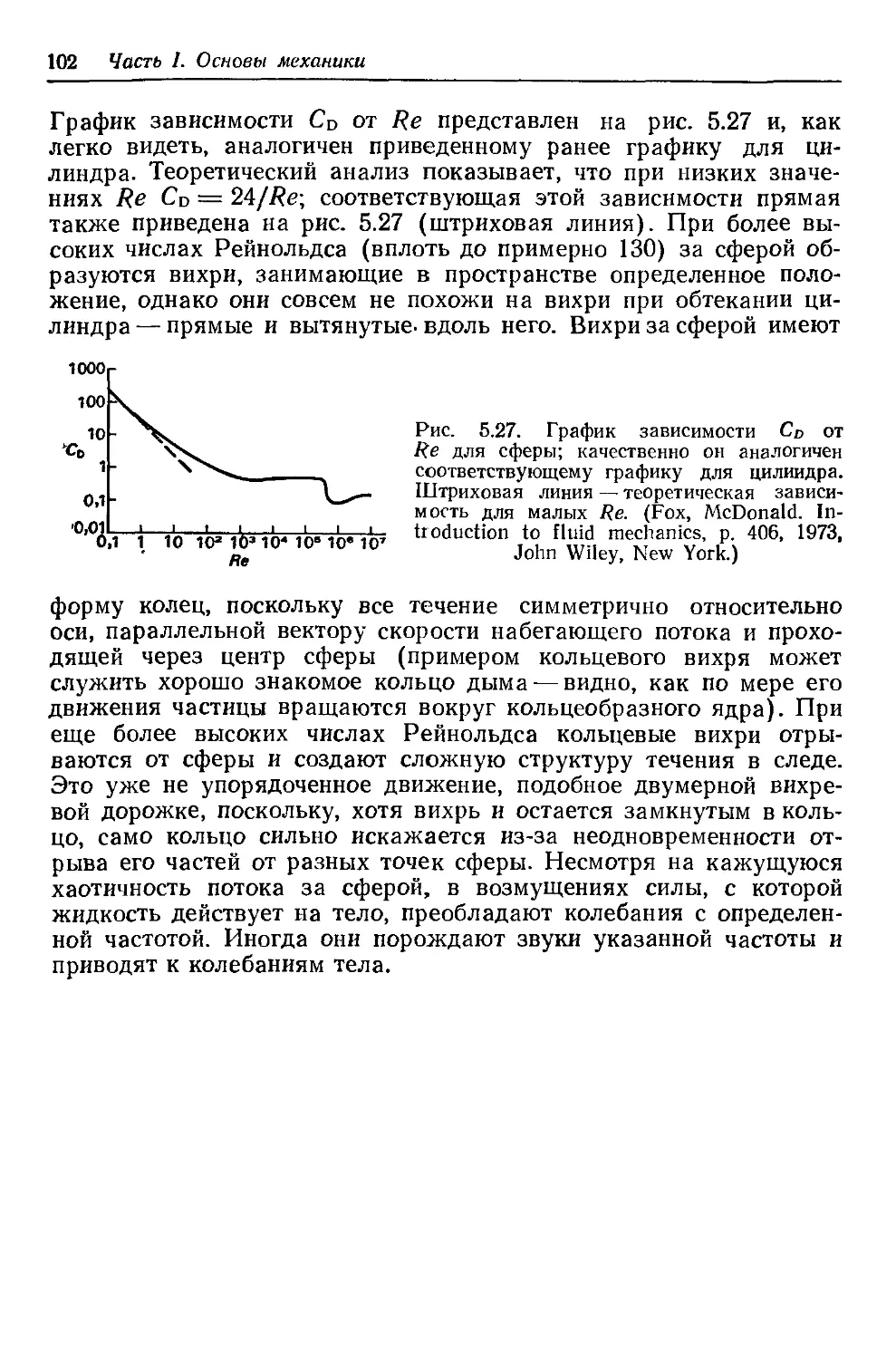














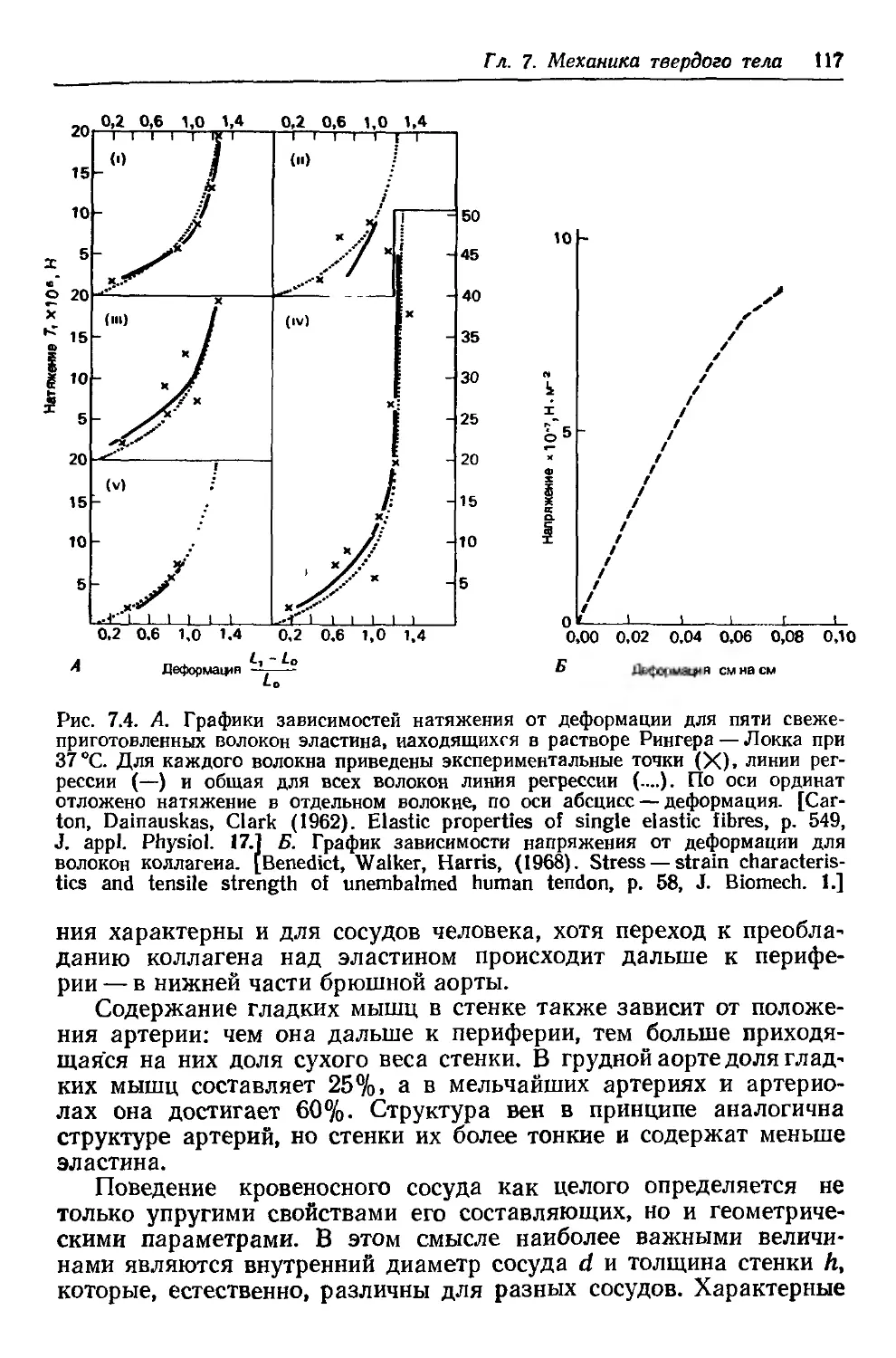





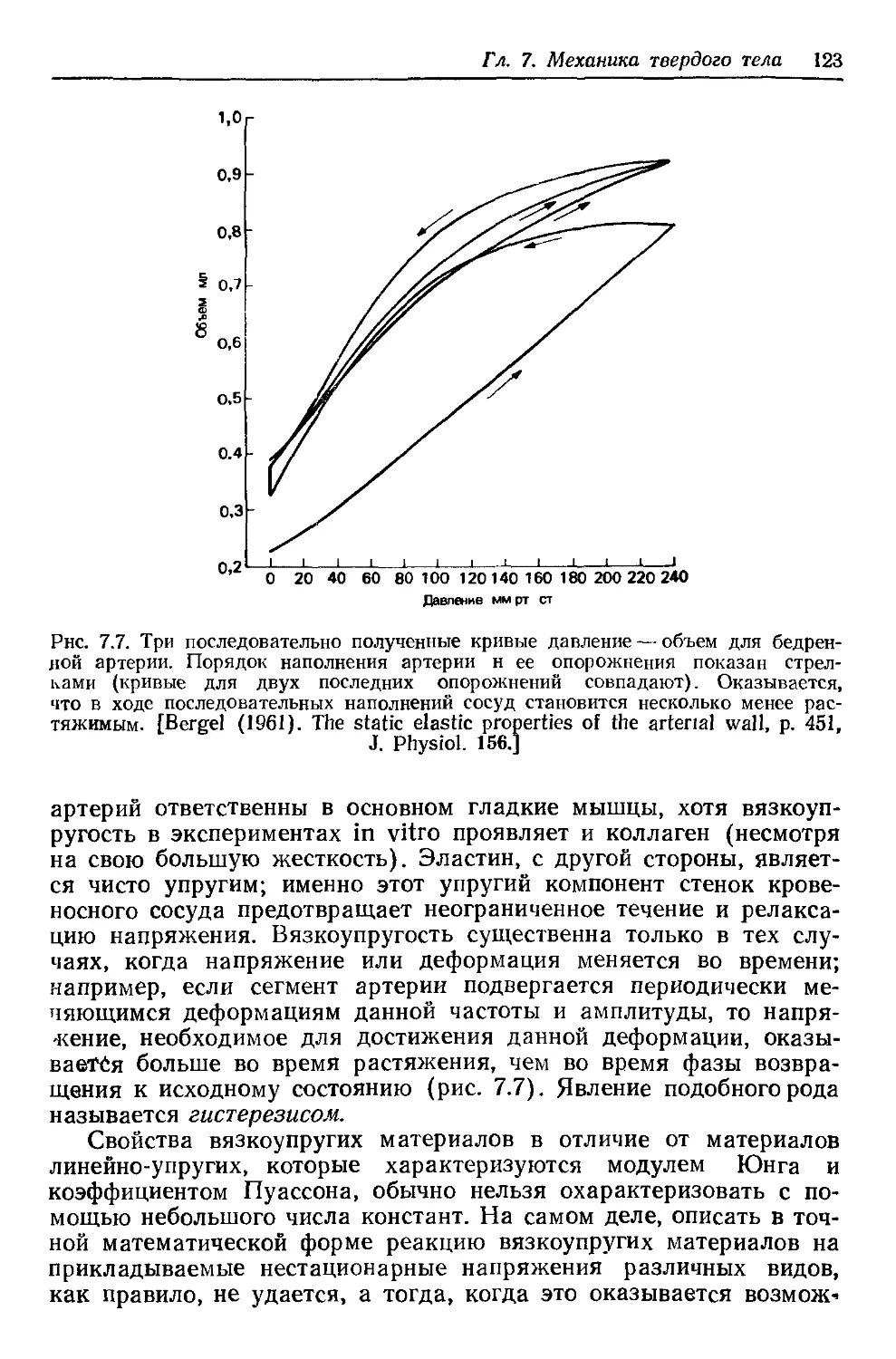



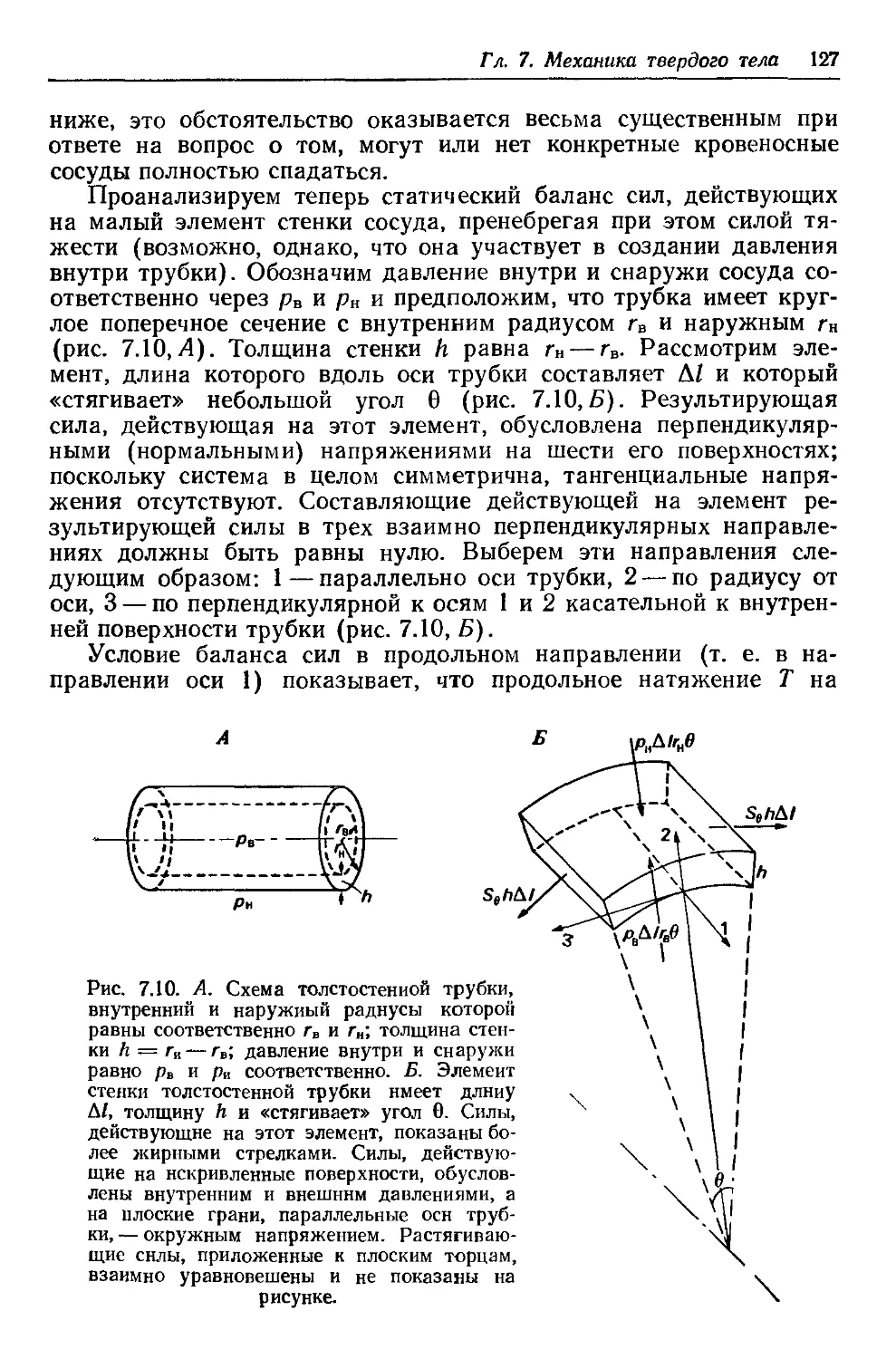

































































































































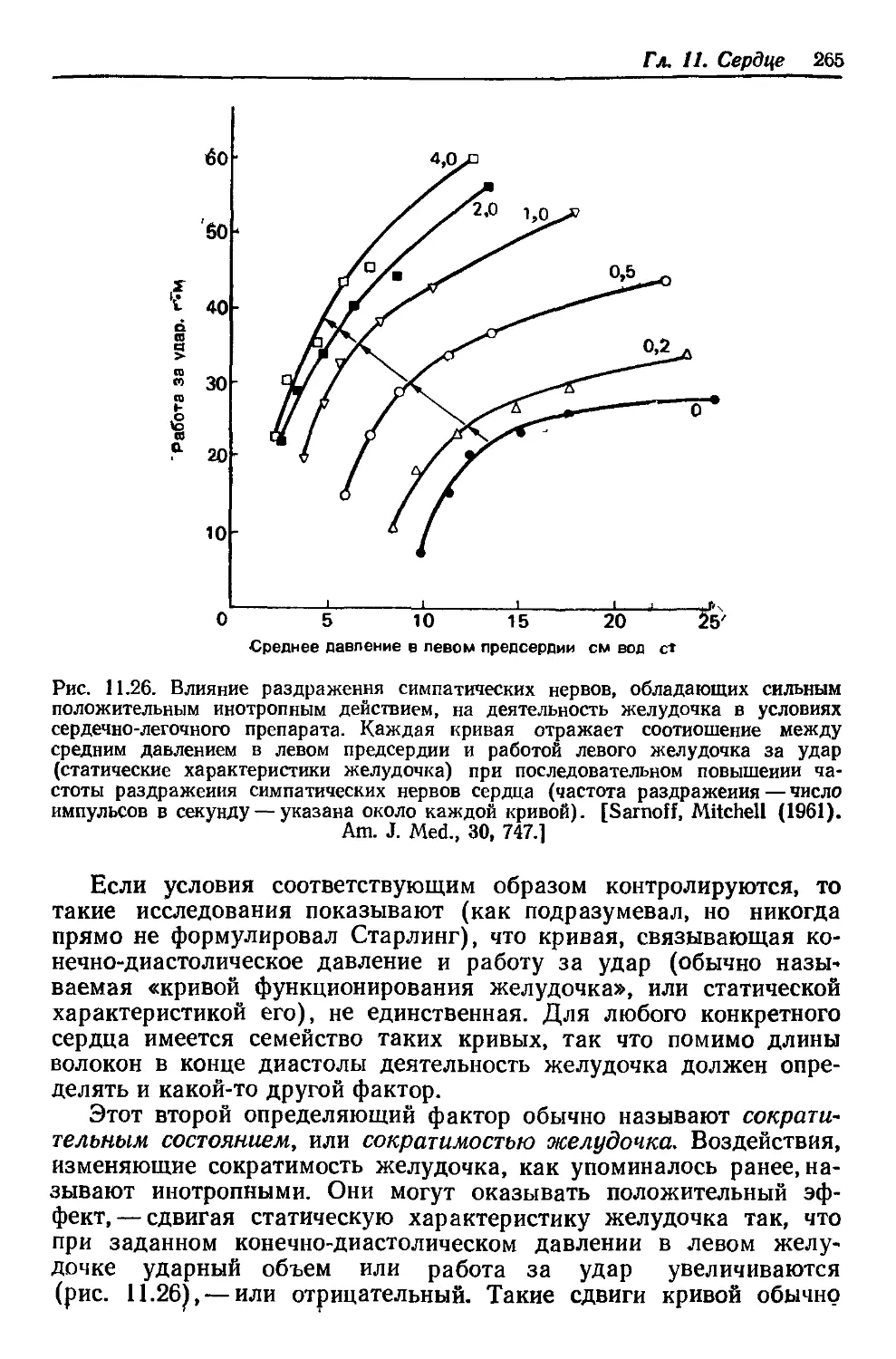



























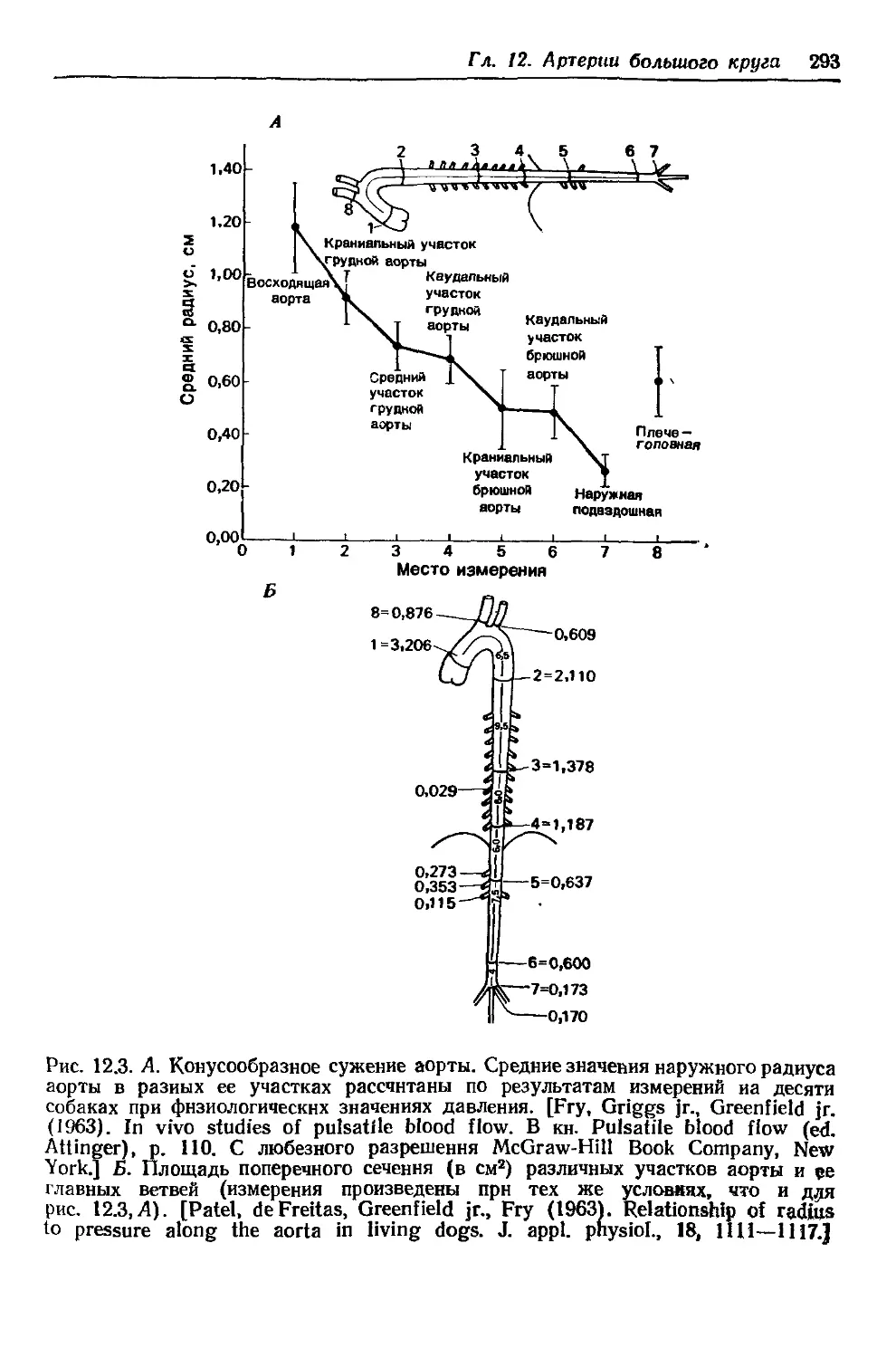










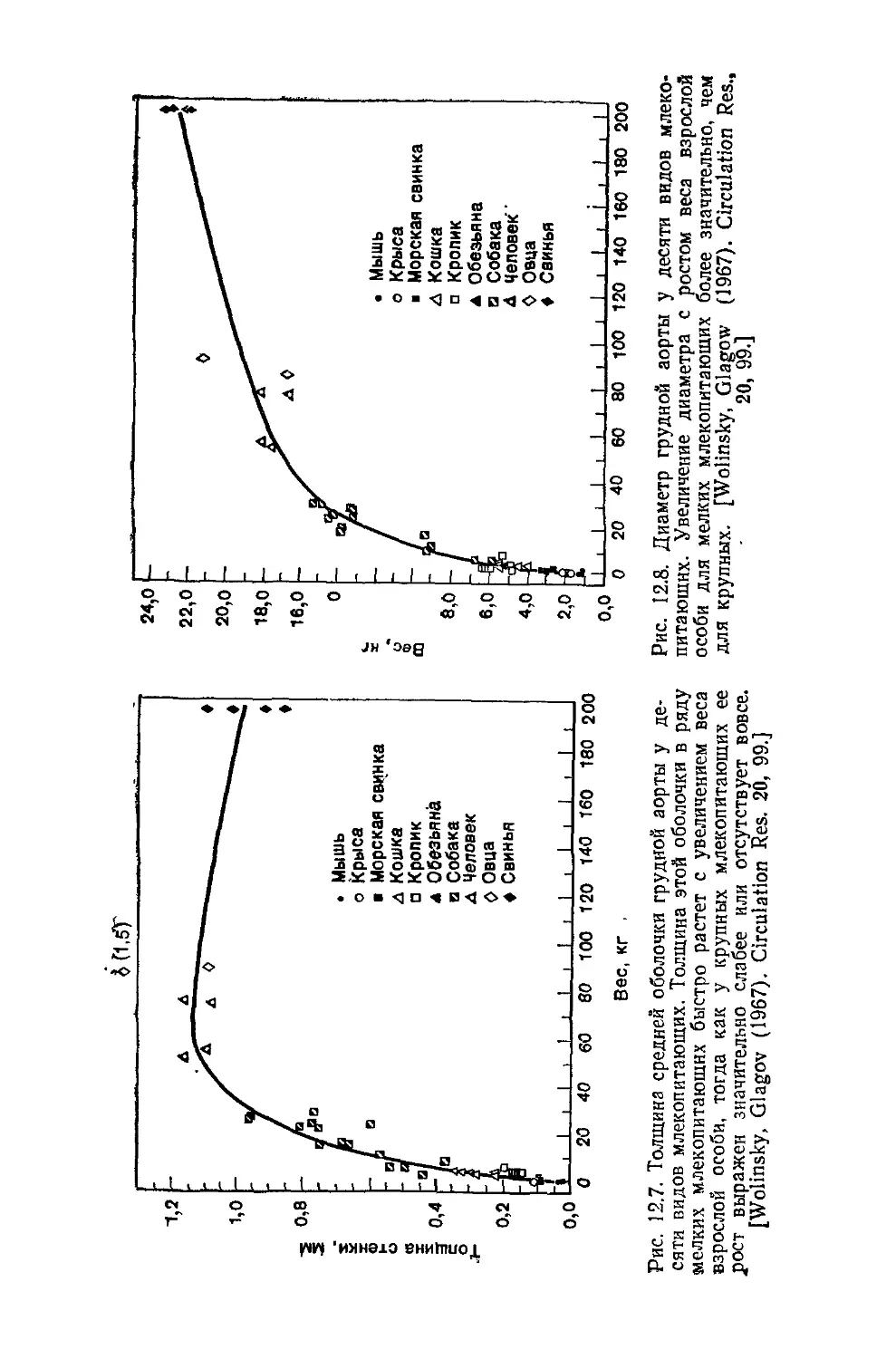



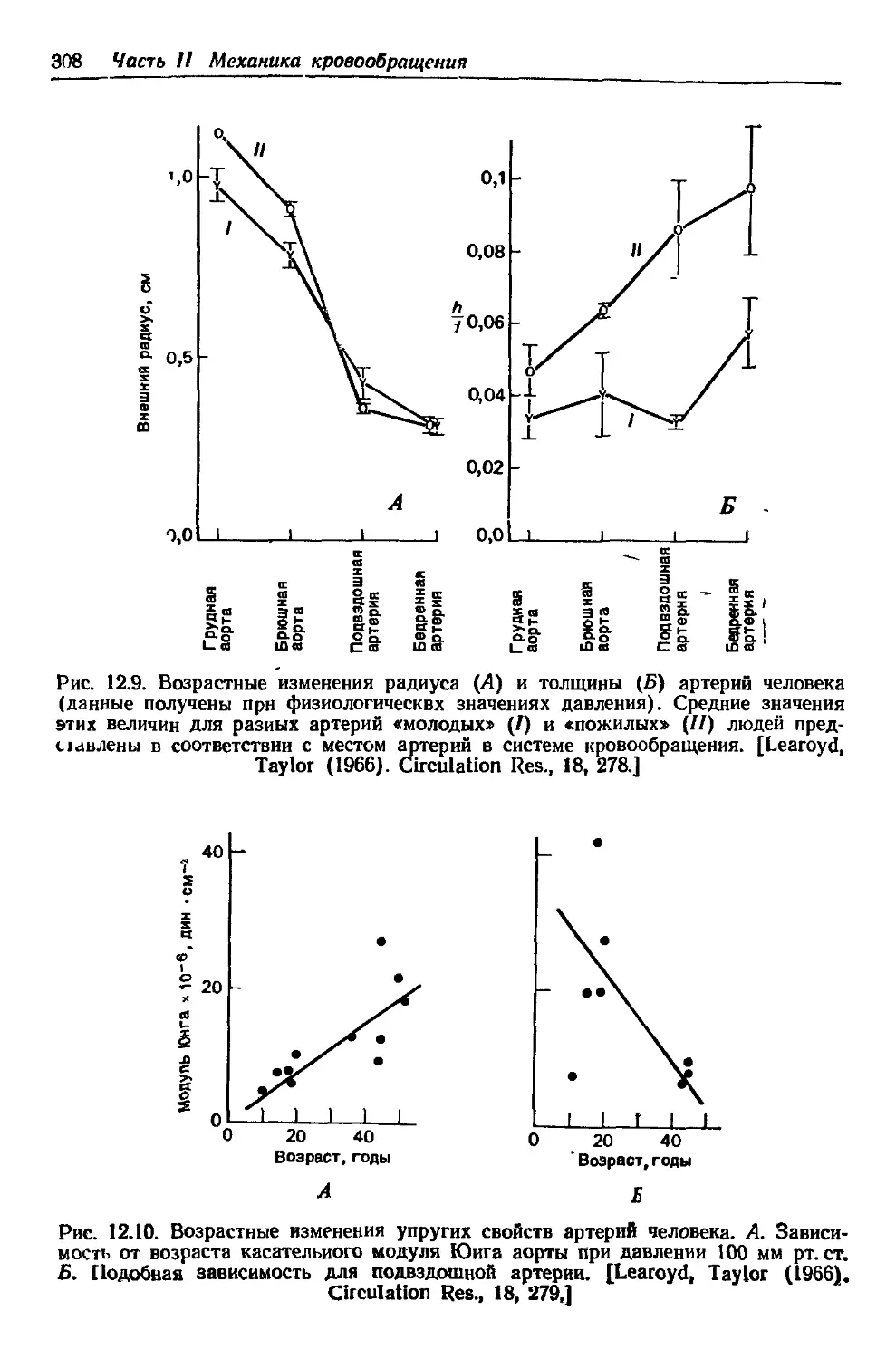


































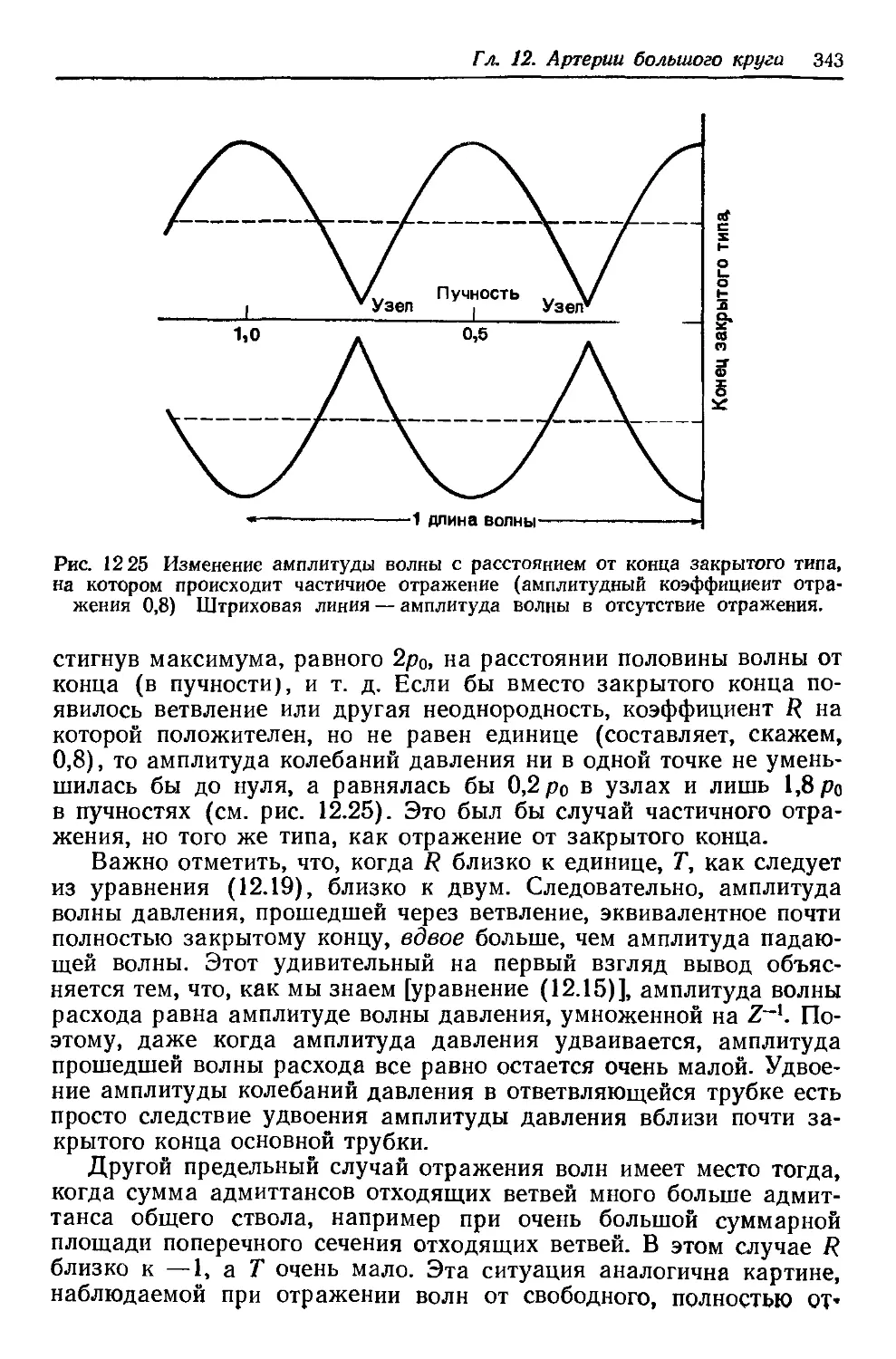























































































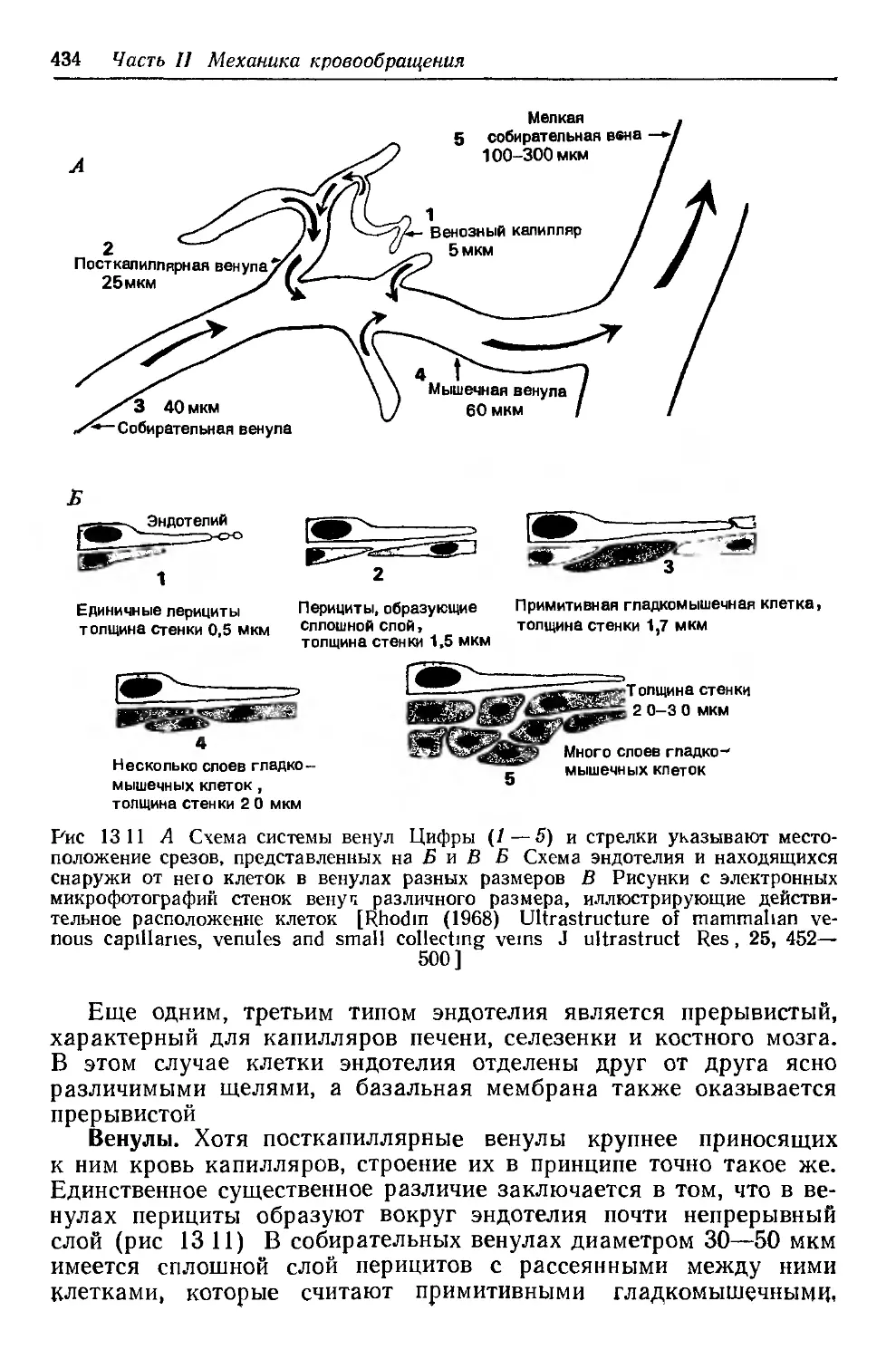



























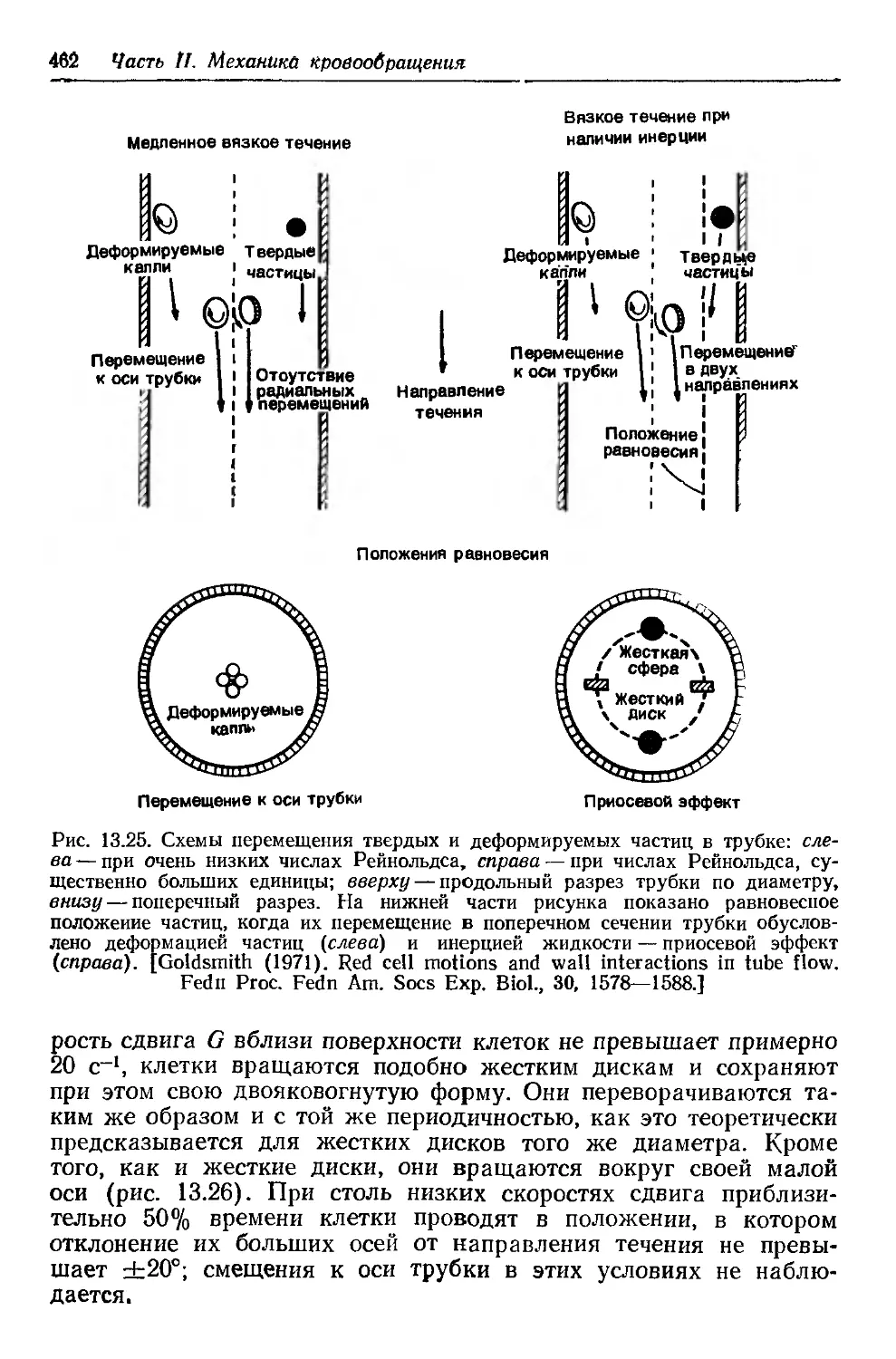

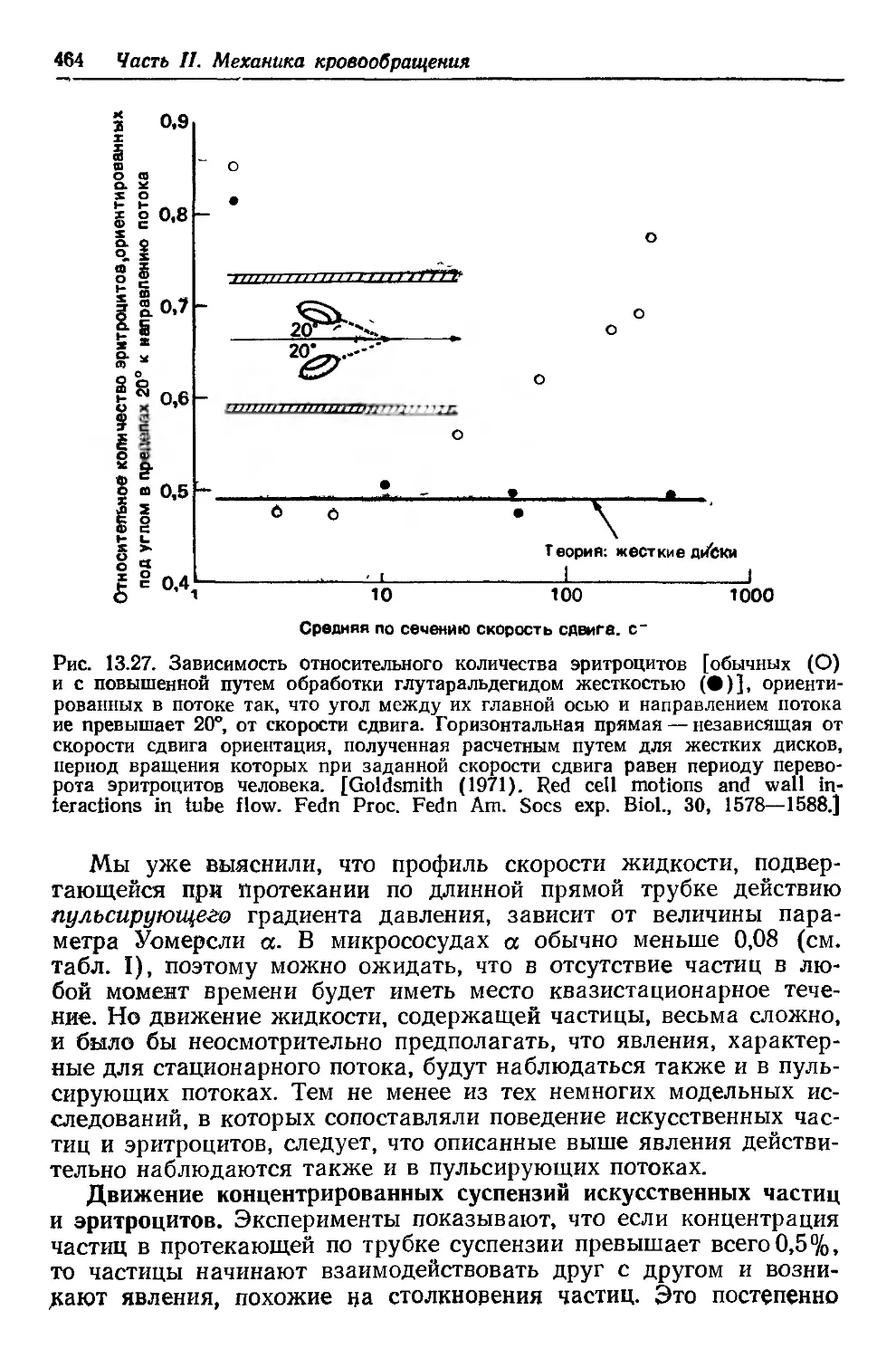










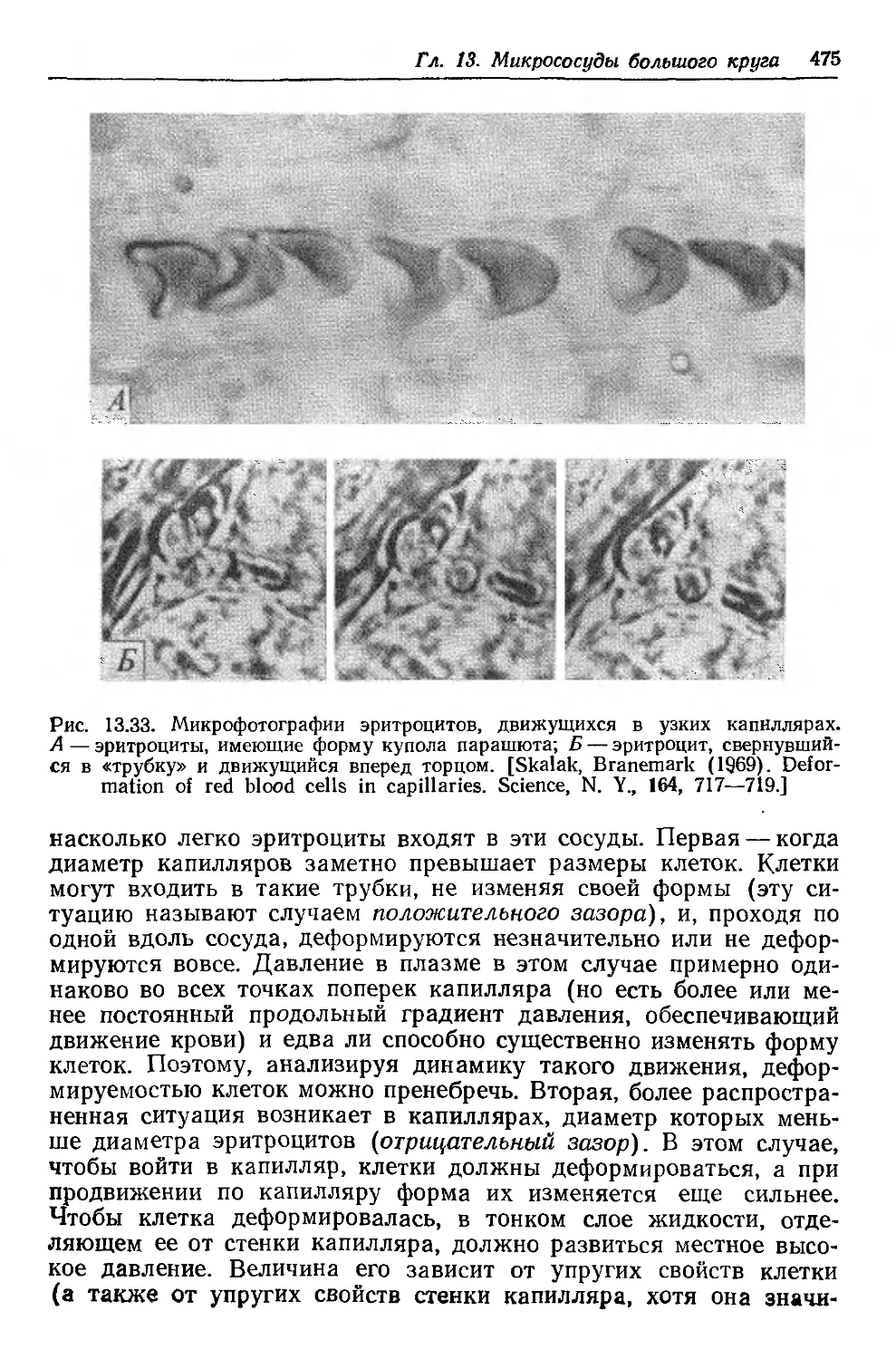


















































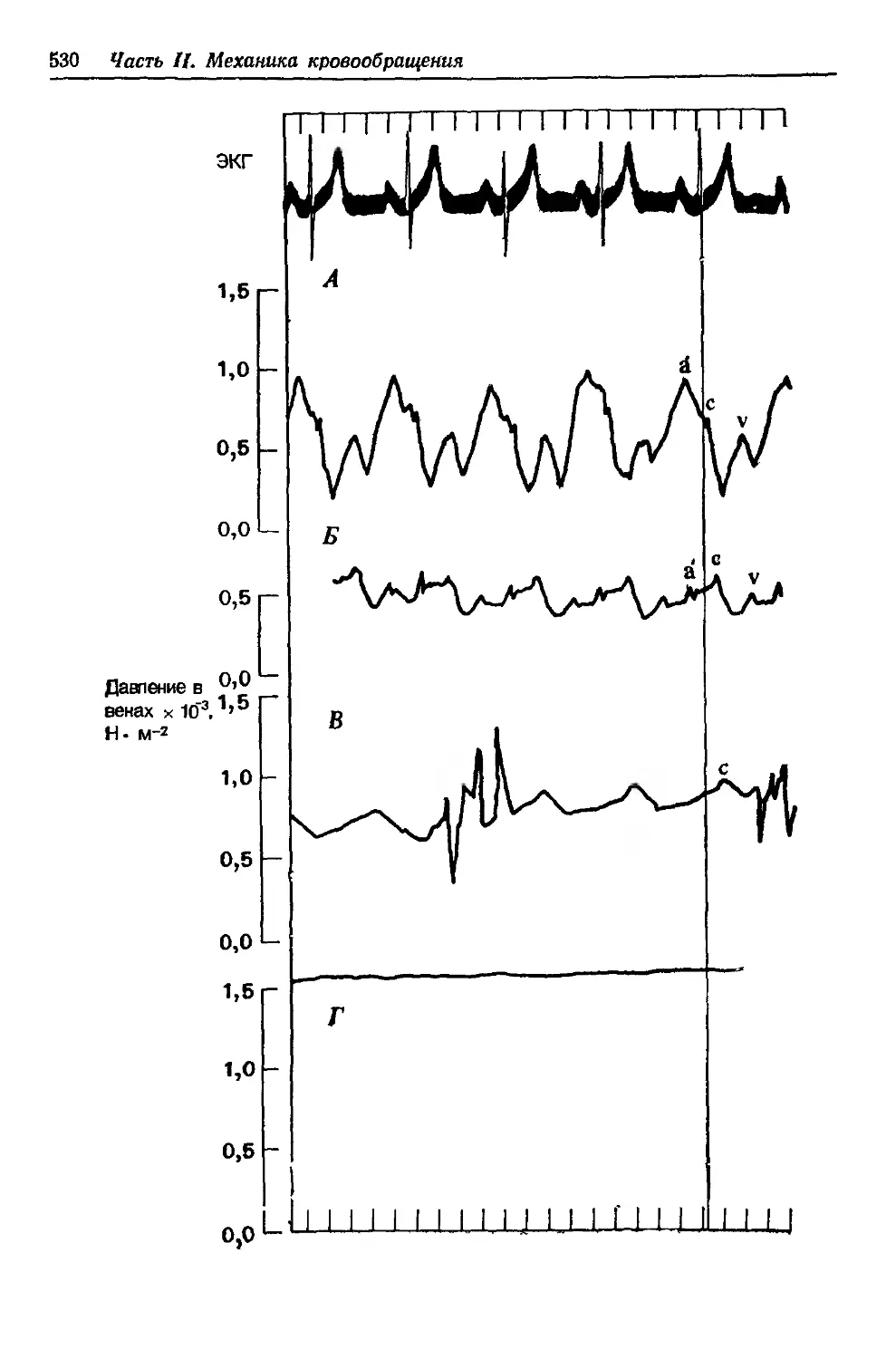



























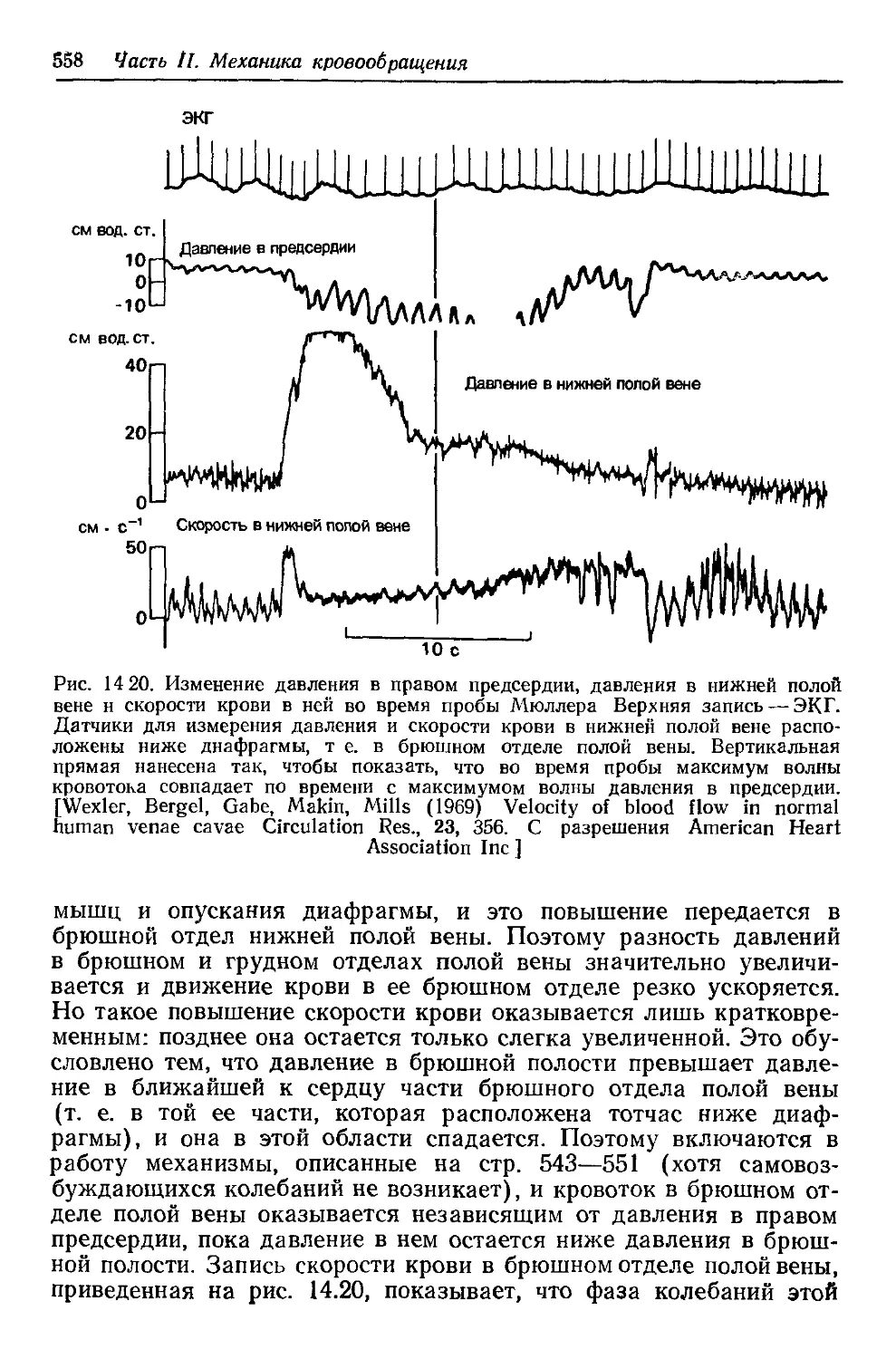





























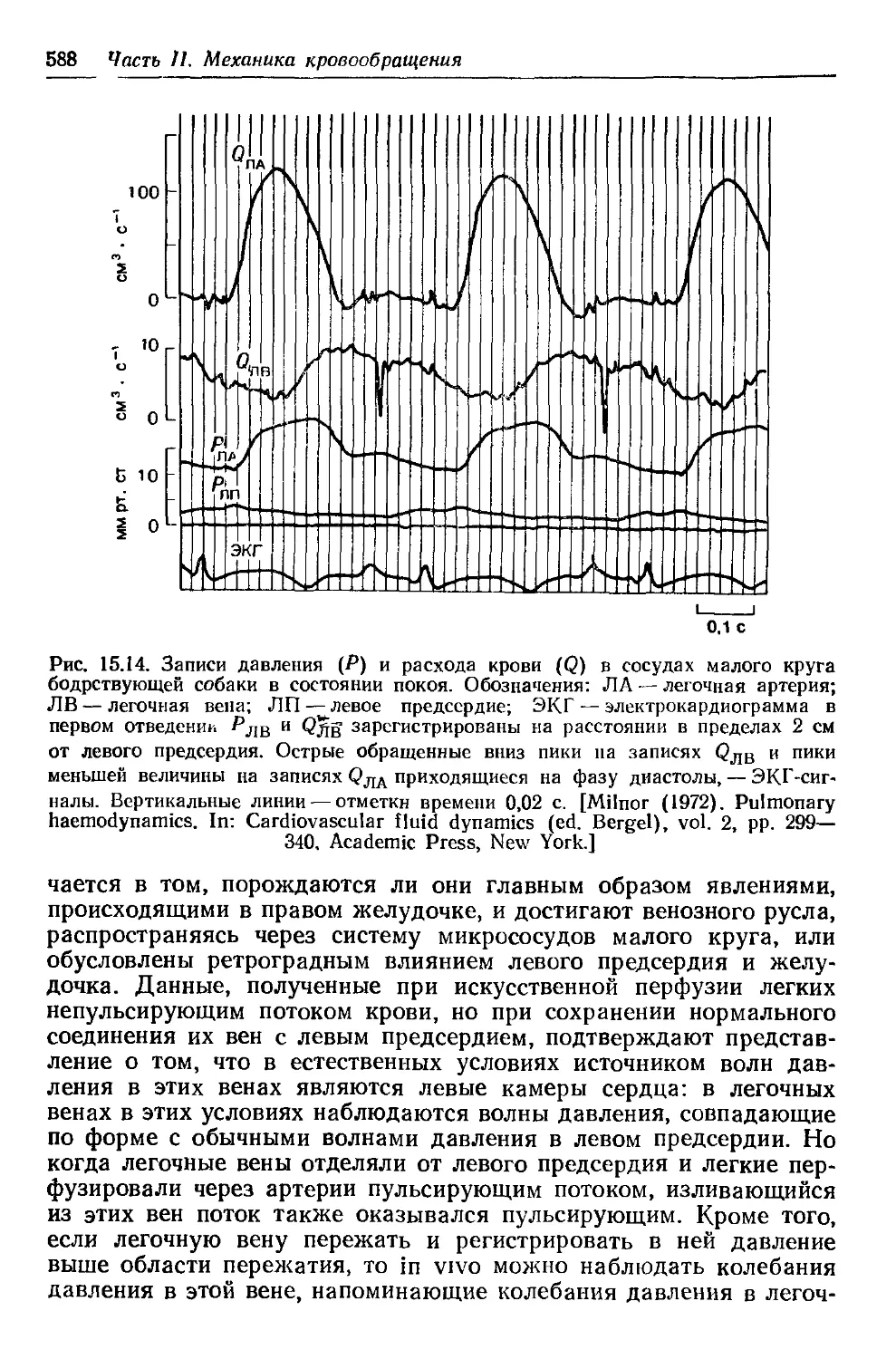




































Author: Каро К. Педли Т. Шротер Р. Сид У.
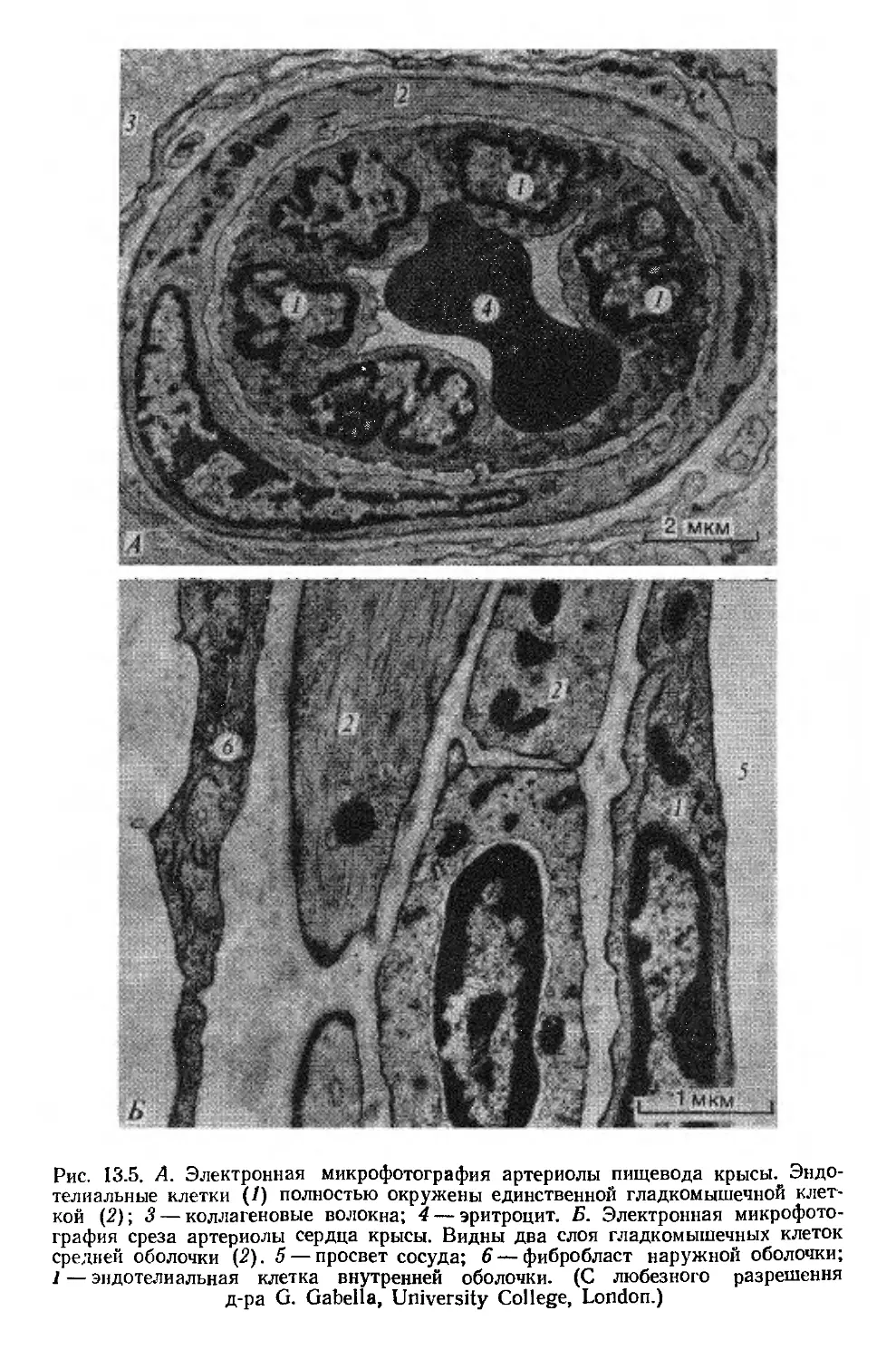
Tags: физиология животных медицина биология физиология биомеханика
Year: 1981




Text
НИЯ
МЕХИ НИМ =
КРОВО
ОБРИ
The mechanics of the
circulation
C. G. Caro
Professor of Physiological Mechanics;
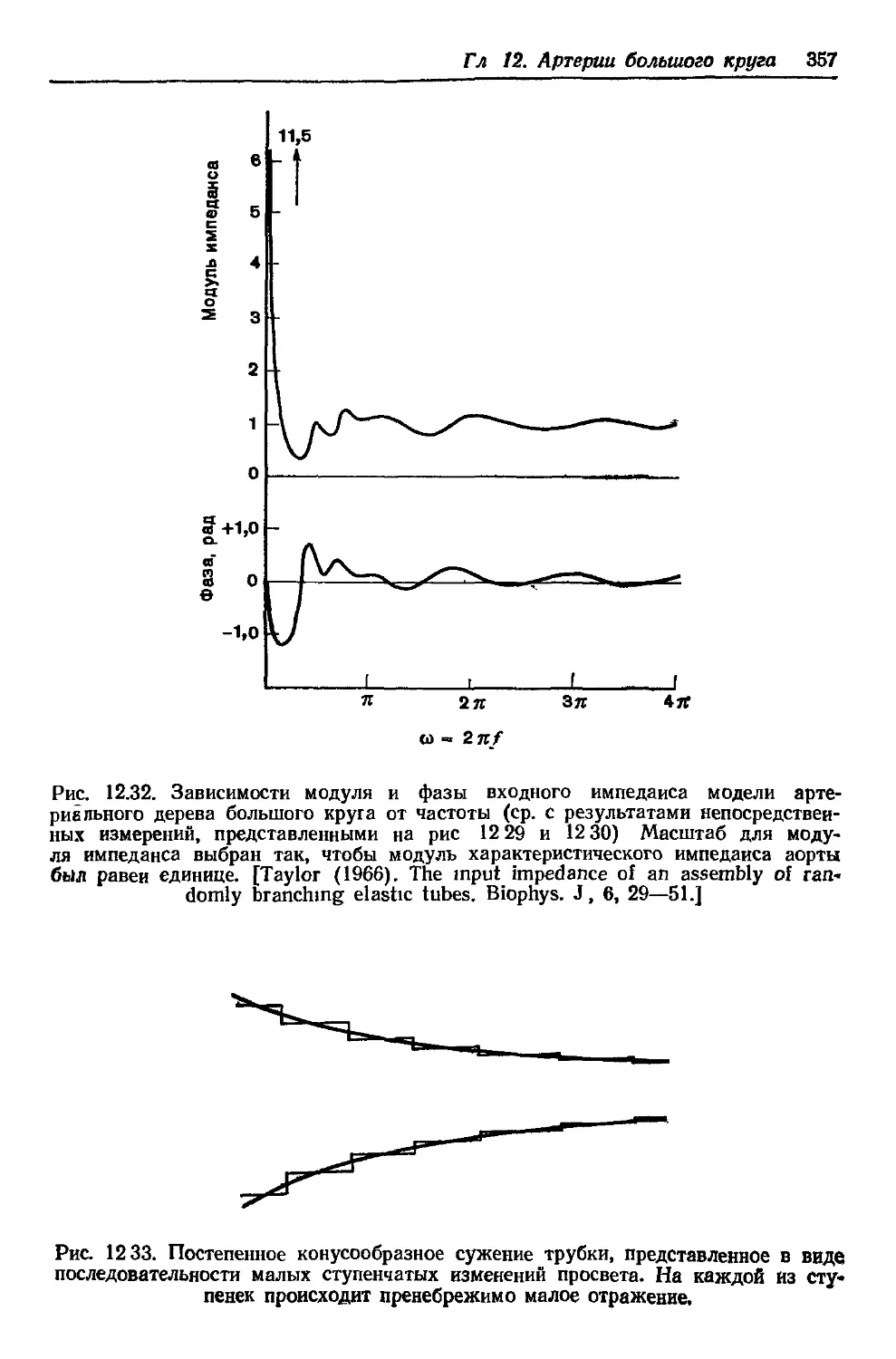
Director, Physiological Flow Studies Unit,
Imperial College London
T. J. Pedley
Assistant Director of Research in Physiological
Fluid Mechanics,
Department of Applied Mathematics and Theoretical
Physics,
University of Cambridge
R. C. Schroter
Lecturer, Physiological Flow Studies Unit,
Imperial College, London
W. A. Seed
Senior Lecturer in Medicine and Consultant
Physician,
Charing Cross Hospital, London
Oxford
Oxford University Press
New York Toronto
1978
,Х МЕХАНИКИ
' s КРОВООБРАЩЕНИЯ
Перевод с английского
канд. биол. наук Е. В. Лукошковой
и
А. Н. Рогозы
под редакцией
д-ра физ.-мат. наук С. А. Регирера
и
д-ра мед. наук В. М. Хаютина
ИЗДАТЕЛЬСТВО «МИР»
МОСКВА 1981
УДК 591.1 + 532.Б1
В коллективной монографии известных английских
ученых рассмотрены основные идеи механики упругих
тел и гидромеханнкн, механическое поведение крови,
статические н динамические свойства сердечной мышцы,
биомеханика и гидродинамика насосной функции серд-
ца, микроциркуляция, транскапиллярный перенос ве-
ществ, механические свойства сосудов и движение в
них крови, мышечный насос, механика легочного крово-
обращения.
Предназначена для физиологов, биофнзнков, пато-
физиологов, фармакологов, врачей, механиков, матема-
тиков и инженеров.
Редакция литературы по биологии
2001040000 © Oxford University Press 1978
М 2У>в4~~ 12Я. 120 —81 ч. 1 © Перевод на русский язык, «Мир», 1981
041 (01) •“ 81
ПРЕДИСЛОВИЕ РЕДАКТОРОВ ПЕРЕВОДА
Мы рады представить советскому читателю обстоятельный
труд по одному из наиболее важных для современной медицины
разделов физиологии. Эта книга коренным 'образом отличается
от существующих руководств по физиологии сердечно-сосудистой
системы.
С 40-х годов XIX в. идеи и представления механики стали си-
стематически использоваться в качестве основы для раскрытия
закономерностей функционирования системы кровообращения. Од-
нако до сих пор в большинстве учебников и книг по физиологии
механика сердечно-сосудистой системы излагается на уровне, не
сильно отличающемся от того, что позволяла сделать классиче-
ская механика XIX в. Между тем развитие как физиологии, так и
механики, а также измерительной и вычислительной техники при-
вело, начиная примерно с 50-х годов нашего столетия, к совер-
шенно новому положению вещей.
Появилась возможность подробно исследовать свойства крови
как жидкой ткани (которые вовсе не сводятся к одной только
вязкости), дать описание работы сердца на основе механических
моделей мышцы с привлечением понятий и аппарата механики де-
формируемых сред, построить завершенную линейную теорию рас-
пространения пульсовых волн и перейти к изучению нелинейных
эффектов в артериях, измерять не только средний расход в пуль-
сирующем потоке крови, но даже мгновенные профили скорости
в крупных артериях. Благодаря применению электронной микро-
скопии и изотопных методов начали раскрываться механизмы
транскапиллярного переноса веществ. Оказались осуществимыми
измерения скорости и перепада давления в отдельных микросо-
судах. По существу заново созданы представления о механике
венозного и легочного русла. Этот перечень можно было бы про-
должить— мы упомянули здесь только некоторые из рассмотрен-
ных в книге вопросов.
Уже в течение тридцати лет физиологи, механики, инженеры,
физики, математики, врачи возводят своего рода «вавилонскую
башню» — междисциплинарный подход к исследованию кровооб-
ращения. Усилиями участников этого процесса руководит, конечно,
не «гордыня», а потребности науки и практической медицины,
и эти усилия уже принесли ощутимые результаты, так что не
6 Предисловие редакторов перевода
приходится сомневаться в их полезности. Однако, поскольку и пер-
воначальный разрыв между науками был велик, «смешение языков»
усугубилось. В силу скромной физико-математической подготовки
большинству физиологов, биологов и врачей трудно постичь со-
временную механику, тогда как большинству инженеров и физи-
ков биологические дисциплины представляются недоступными
из-за их обширности и специальной терминологии. Вследствие та-
кой .разобщенности биологи часто не умеют четко поставить за-
дачу перед механиками, а механики, в свою очередь, нередко за-
нимаются второстепенными или вовсе надуманными задачами.
Четыре автора этой книги — физиолог, клиницист-кардиолог,
физик и специалист по прикладной математике — поставили своей
целью создать монографию, которая помогла бы представителям
различных наук лучше понимать друг друга. Сделать это чрезвы-
чайно трудно, и тем не менее попытка в целом оказалась успеш-
ной. Читателю с медицинским или биологическим образованием
предоставлена возможность изучить основы механики в той мере,
в какой это необходимо для понимания работы системы кровооб-
ращения. Хотя в книге содержится довольно много (непривычно
много для физиолога) формул, все они приведены в окончатель-
ном виде, так что чтение книги не требует активного владения
математическим аппаратом.
Основные сведения из механики занимают почти треть объема
книги, что на самом деле немного, если принять во внимание не-
обходимость начать изложение с азов и, как правило, не пользо-
ваться математическим языком. Опасность чрезмерного упроще-
ния, внушения читателю ложного представления о простоте про-
блем в таких случаях очень велика. Нам кажется, что авторы со-
знавали эту опасность и сделали все возможное для того, чтобы
сохранить логическую строгость и корректность при обсуждении
вопросов механики. Эти вопросы изложены на высоком профес-
сиональном уровне, без элементов «кустарщины», терминологиче-
ской неряшливости и многого другого, что часто встречается в со-
чинениях, трактующих применение механики в биологии, и порож-
дает у компетентного читателя справедливое раздражение, а у
неофита — устойчивые заблуждения, оправдываемые впоследствии
ссылками на авторитетный источник.
Столь же строгий и высоко профессиональный уровень отли-
чает вторую часть — изложение основ современной механики кро-
вообращения. Для читателя — физика или инженера — важно, что
каждую главу предваряют сжато написанные очерки морфологии
и физиологии, освещающие именно те аспекты, которые необхо-
димы для более подробного изложения вопросов механики крови,
сердца, артерий, вен, микрососудов большого круга и, наконец,
малого круга.
Отбор материала очень удачен — наряду с классическими дан-
ными в книгу вошли многие новейшие результаты. Избегая стиля
Предисловие редакторов перевода 7
критического обзора, авторы вместе с тем четко разграничивают
устоявшиеся воззрения и гипотезы и приводят небогатую, но тща-
тельно подобранную библиографию, в том числе все основные
источники, которые могут служить для углубленного изучения тех
или иных частных вопросов.
В силу самой структуры книги в ней рассматриваются отдель-
ные элементы системы кровообращения; читатель должен отдавать
себе отчет в том, что без этого невозможно дать описание меха-
ники системы в целом. Такого описания в книге нет, и это неуди-
вительно, так как одна из главных нерешенных проблем физиоло-
гии и состоит в создании механики сердечно-сосудистой системы
как целого и в постижении на этой основе закономерностей управ-
ления кровообращением. В конечном счете нервные и другие регу-
лирующие воздействия изменяют именно механические характери-
стики сердца и сосудов, приспосабливая их совокупную деятель-
ность к потребностям организма. Вот почему познание механики
сердечно-сосудистой системы есть предпосылка к пониманию про-
цессов управления этой системой. Такие же соображения можно
высказать и в отношении систем, определяющих кровоснабжение
различных органов.
Хочется думать, что появление этой книги будет способствовать
лучшему осознанию глубокой связи механики кровообращения и
традиционного для русской и советской физиологии направления —
изучения нервной регуляции кровообращения. Мы надеемся, что
перевод и издание данной книги явится заметным событием в объ-
единении усилий представителей различных дисциплин для даль-
нейшего познания физиологии кровообращения — основы столь
важного дела, каким является борьба с сердечно-сосудистыми за-
болеваниями.
С. Регирер
В. Хаютин.
ПРЕДИСЛОВИЕ АВТОРОВ К РУССКОМУ ИЗДАНИЮ
Наши коллеги — инициаторы и редакторы перевода этой книги,
просили нас написать небольшое предисловие и тем самым предо-
ставили нам возможность поделиться некоторыми соображениями
Наш опыт обучения студентов-медиков, а также врачей, биоло-
гов и физиков показал, что создание такой книги необходимо. То,
что читатели встретили ее благожелательно и она получила много
положительных отзывов в печати, была быстро переиздана и после
этого вышла массовым тиражом, а теперь появляется на русском
языке, еще раз подтверждает нашу уверенность в том, что потреб-
ность в руководстве такого типа насущна и что мы в какой-то
мере ее удовлетворили.
Применение физических знаний в физиологии возрастает, дока-
зывая со всей очевидностью, что врачи и биологи должны четко,
пусть и не всегда широко, понимать механику, а физики — вовле-
каться в исследования механики кровообращения. То, что наша
книга переведена на иностранный язык, имеет особо важное зна-
чение: это напоминает нам об истории развития представлений о
механике кровообращения и о постоянной необходимости сотруд-
ничества ученых разных стран. Перевод книги, имеющей междис-
циплинарный характер, а наша книга такова,—дело не легкое.
Мы выражаем свое восхищение тем, что проф. В. М. Хаютин и
д-р С. А. Регирер взяли на себя этот труд. Та тщательность,
с какой они отнеслись к оригиналу, позволила выявить некоторые
наши просчеты и исправить их в переводе.
Февраль, 1981.
ПРЕДИСЛОВИЕ
В 1808 г. Томас Юнг, читая в Королевском обществе Крунов-
скую лекцию о функциях сердца и сосудов, начал ее такими ело*
вами:
«Механические движения в теле животного подчиняются тем
же общим законам, что и движения тел неодушевленных, ... и по-
тому очевидно, что вопрос о том, каким образом и в какой сте-
пени движение крови по сосудам зависит от мышечных и упругих
сил сердца и артерий (если предположить, что природа этих сил
будет познана), сводится к проблемам, относящимся к узко спе-
циальным разделам теории гидравлики».
Для Юнга такой подход к физиологии был естественным. Как
и многие ученые девятнадцатого столетия, он не усматривал боль-
ших различий между биологическими и физическими науками.
В самом деле, всю свою жизнь Юнг был и практикующим врачом,
и профессором физики. И хотя ныне этого ученого помнят главным
образом как создателя волновой теории света, а также потому, что
его именем назван модуль упругости материалов, Юнг, кроме того,
является автором основополагающих работ по физиологической
оптике, цветовому зрению и кровообращению, в том числе и работ,
касающихся распространения волн по артериям.
Столь разносторонняя деятельность была, по-видимому, тради-
ционной для первых исследователей кровообращения. Об этом
свидетельствуют такие имена, как Борелли, Хейлз, Бернулли, Эй-
лер, Пуазейль, Гельмгольц, Фик, Франк. Однако развитие науки
предполагает все большую специализацию ученых, и исследова-
ние сердечно-сосудистой системы отделилось от физических наук.
Это отделение, конечно, не было полным: сотрудничество пред-
ставителей различных наук не прекращалось никогда. Тем не ме-
нее масштабы такого сотрудничества были весьма ограничены.
Многие врачи и физиологи избегали его в силу слабой подготовки
в области математики и механики, физиков же отталкивали слож-
ность и эмпиризм физиологических исследований, а также непри-
вычная терминология.
В наши дни вызванное специализацией разделение приняло но-
вую форму. Примерно на протяжении последних 20 лет физики и
инженеры внесли значительный вклад в понимание механики кро-
вообращения. Этот успех способствовал дальнейшему развитию
10 Предисловие
совместных исследований, но в то же время сделал данную об-
ласть еще более сложной для тех, кто не обладает глубокими зна-
ниями по физике и математике. О важности этой работы на стыке
наук свидетельствует ряд обзоров и монографий последнего вре-
мени, но они едва ли помогают читателю-врачу: в них неизменно
предполагается знание механики, а изложение нередко принимает
сугубо математический характер.
Данная книга призвана облегчить создавшееся положение. Ее
назначение — служить вводным курсом по механике кровообра-
щения, в котором использование математического аппарата све-
дено к минимуму, а механика излагается в доступной форме. Наш
опыт преподавания слушателям, изучающим физиологию и меди-
цину, а также кардиологам — терапевтам и хирургам, — показы-
вает, что такой подход весьма полезен, и данная книга адресована
в первую очередь именно этому кругу читателей. Кроме того, по-
скольку в ней изложены необходимые для анализа механики кро-
вообращения основы анатомии и физиологии и даны определения
различных терминов, а там, где это возможно, приведены числен-
ные значения соответствующих параметров, мы полагаем, что она
окажется полезной физикам, математикам и инженерам, интере-
сующимся данной областью знаний.
Книга состоит из двух частей. В первой части, названной «Ос-
новы механики», описаны в общих чертах (фактически без исполь-
зования математического аппарата) физические процессы и меха-
низмы, принципиально важные для понимания механики кровооб-
ращения. Эта часть представляет собой физическое введение к
дальнейшему изложению. Вместе с тем она в достаточной мере
самостоятельна. В ней рассматриваются преимущественно общие,
а не частные вопросы механики твердых тел и жидкостей, а также
массопереноса, и потому она может быть полезной как основа
при изучении не только кровообращения, но и других физиологи-
ческих систем.
Во второй части, названной «Механика кровообращения», до-
статочно подробно рассмотрены протекающие в сердечно-сосуди-
стой системе физиологические процессы и лежащие в их основе
физические механизмы. Сначала здесь описываются необходимые
для понимания механики кровообращения свойства крови, а затем
систематически, глава за главой, рассматриваются все отделы си-
стемы кровообращения, начиная с сердца. Активные физиологиче-
ские механизмы, а именно рефлекторные реакции, подробно не
анализируются, но изучаются обусловленные ими конечные изме-
нения физических свойств системы. В каждой главе сначала сооб-
щаются необходимые основные сведения по анатомии и физиоло-
гии, а затем обсуждаются вопросы механики. По ходу изложения
мы часто ссылаемся на приведенное в первой части книги описа-
ние основных физических процессов; более специальные физиче-
ские явления, относящиеся к механике определенных частей ед-
Предисловие 11
•темы кровообращения, рассматриваются в соответствующих раз-
делах.
Мы попытались охватить все механические свойства системы
кровообращения, которые в настоящее время считаются важными.
Однако эта книга была задумана не как обзор научных исследо-
ваний!, и потому мы в основном избегали ссылок в тексте на ори-
гинальные работы. Вместо этого каждая глава второй части
снабжена библиографическим списком, составленным так, чтобы
читатель, незнакомый со специальной литературой, смог найти со-
ответствующие обзоры и другие источники. Иллюстрации мы выби-
рали по возможности из наиболее солидных работ, во многих
случаях из оригинальных научных статей, так что ссылки в подпи-
сях к рисункам могут служить дополнением к библиографии.
При написании любой книги, находящейся на стыке наук, ве-
лико искушение чрезмерно упростить изложение, обычно — какой-
либо одной области; мы всеми силами старались избежать этого.
Мы также пытались всюду, где это уместно, приводить количе-
ственные данные. Для удобства наиболее важные параметры (из-
меренные и вычисленные), на которые приходится многократно
ссылаться на протяжении всей книги, собраны в одну таблицу
(она помещена в начале книги). Все величины приводятся в Меж-
дународной системе единиц (СИ), но для таких величин, как дав-
ление, мы указываем их значения и в традиционных единицах.
Для механики кровообращения очень важным параметром служит
размер тела, а единственным видом животных, для которого в
настоящее время проведены более или менее всесторонние иссле-
дования и получены достаточно достоверные данные, является со-
бака. Поэтому по всей книге приводятся сведения в основном
для этого животного, хотя при рассмотрении условий движения
крови в микрососудах мы были вынуждены обратиться к другим
видам (правда, межвидовые различия размеров в этой области си-
стемы кровообращения сравнительно невелики). Наконец, мы уде-
ляли специальное внимание сердечно-сосудистой системе человека
в тех случаях, когда механические явления в ней протекают осо-
бым образом или же, как мы предполагаем, имеют отношение
к заболеваниям.
К.К.
Т.П.
Р.Ш.
У.С.
Июль 1977
Нормальные значения параметров сердечно-сосудистой системы собак.
а ниже — диапаз
Восходящая аорта Нисходящая аорта Брюшная аорта Бедренная артерия
Внутренний диаметр h, см 1,6 1,0—2,4 1,3 0,8—1,8 0,9 0,5—1,2 0,4 0,2—0,8
Толщина стенкн d, см 0,065 0,05—0,08 0,05 0,04—0,06 0,04 0,02—0,06
Л/de 0,07 0,055—0,084 0,06 0,04—0,09 0,07 0,055-0,11
Длина, см 5 20 15 10
Примерная площадь по- веренного сечения, см4 2 1,8 0,6 0,2
Полная площадь поперечного сечения сосудов каж- дого уровня, см2 S 2 2 8
Скорость кровн (пиковое значение), см-с-' 120 40—290 105 25-250 55 50—60 100 100—120
Средняя скорость крови, см-с' 20 10-40 20 10—40 15 8—20 10 10-15
Число Рейнольдса (пнковое значение) 4500 3400 1250 1000
а (при частоте пульса 2 Гц) 13,2 11,5 8 3,5
Расчетная скорость волны со, см-с-' 580 770 840
Измеренная скорость волны с, см-с-' 500 400-600 700 600—750 900 800-1030
Модуль Юнга Е, Н-м-а 4,8-106 (3-6) - 10в 10 • 106 (9—11)- 105 10- 105 (9-12)- 10s
*) С. G. Саго, 1. J. Pedley, W. A. Seed (1974). Mechanics of the circulation, Chapter 1 of Car
Таблица I
Там, где это возможно, приведены приблизительные средние значения,
оны значений *)
Сонная артерия Артериолы Капилляры Венулы Каудальная полая вена Легочный ствол
0,5 0,2—0,8 0,005 0,001—0,008 0,0006 0,0004—0,0008 0,004 0,001—0,0075 l,o 0,6-1,5 1,7 1,0—2,0
0,03 0,02—0,04 0,002 0,0001 0,0002 0,015 0,01—0,02 0,02 0,01—0,03
0,08 0,053—0,095 0,4 0,17 0,05 0,015 0,01
15 10—20 0,15 0,1-0,2 0,06 0,02—0,1 0,15 0,1-0,2 30 20—40 3,5 3-4
0,2 2 • 10~8 3- IO-7 2 • IO-8 0,8 2,3
8 125 600 570 3,0 2,3
0,75 0,5-1,0 0,07 0,02—0,17 0,35 0,2-0,5 25 15-40 70
15 6-28
0,09 0,001 0,035 700 3000
4,4 0,04 0,005 0,035 8,8 15
850 100 350
800 600-1 юо 400 100-700 250 200-330
9- 105 (7-11). Ю6 0,7 - 106 6- 10е (0,4—1,0) • 106 (2—10) • 105
diovascular physiology (ed. А. С. Guyton), Medical and Technical Publishers, London.
БЛАГОДАРНОСТИ
Мы весьма обязаны многим коллегам за советы и помощь.
В особенности нам хотелось бы поблагодарить д-ра Л. Смайе,
участие которого в работе над физиологическими разделами главы
о микрососудах чрезвычайно велико; без его содействия мы не
справились бы с обзором этого предмета.
М-р П. Минтон из Imperial College, д-р Дж. Габелла из Uni-
versity College, а также д-р Гр. Миллер и д-р Д. Гибсон из Бромп-
тонской клиники предоставили в наше распоряжение новые дан-
ные и материалы для иллюстраций. Д-р Садлоу сделал многое
на первых этапах обсуждения содержания и формы изложения и
предоставил нам материалы для главы о венозных сосудах. Проф.
Ил. Ингрэму, д-ру Дж. Хоффману и д-ру М. Хьюзу мы обязаны за
ценные замечания и предложения по главам о крови, сердце и
кровообращении в малом круге (соответственно). Кроме того, мы
отдаем долг признательности всем тем авторам и редакциям жур-
налов, которые названы в подписях к рисункам, за разрешение вос-
произвести эти рисунки. ""
Наконец, мы приносим особую благодарность м-с Ив. Эдвердс:
ее четкость редактора помогала нам (и дисциплинировала нас) на
всем протяжении работы над этой книгой.
Часть I
ОСНОВЫ МЕХАНИКИ
Глава 1
ЧАСТИЦЫ И СПЛОШНЫЕ СРЕДЫ
Механика — это наука о движении (и о равновесии) тел, а
также о силах, вызывающих это движение. Кровь течет по крове-
носным сосудам, гонит ее по ним своеобразный насос — сердце;
упругие и растяжимые стенки сосудов также подвижны; между
кровью и стенками существует определенная сила взаимодействия,
что сказывается на движении как крови, так и стенок. Следова-
тельно, чтобы изучать механику кровообращения, мы должны сна-
чала понять основные законы движения жидкостей (к ним отно-
сится кровь), упругих тел (таковы стенки сосудов) и природу сил,
возникающих при взаимодействии двух перемещающихся относи-
тельно друг друга объектов (в частности, крови и стенок сосудов).
Законы механики применимы не только к движению объектов,
обладающих сравнительно большими размерами (кровь и стенки
сосудов как таковые), но и к движению на молекулярном уровне.
Таким образом, мы будем понимать под механикой науку, кото-
рая изучает все факторы, определяющие перенос вещества (вклю-
чая диффузию и конвективный перенос).
Начало развитию механики было положено в древней Греции,
когда предпринимались попытки сформулировать «законы», уп-
равляющие движением изолированных твердых тел. Древние греки
полагали, что, раз тело движется, значит, на него постоянно дей-
ствует какая-то сила, но физическая природа этой силы, дей-
ствующей, например, на летящую стрелу, представлялась таин-
ственной. Неизбежность существования подобной силы связывали
с одним из парадоксов греческого философа Зенона: в каждое
мгновение стрела занимает определенное положение, т. е. непод-
вижна, и в то же время движется, так как в последующее мгнове-
ние занимает другое положение.
Вопрос о силах, обусловливающих движение, не был оконча-
тельно решен вплоть до XVII в., когда Исаак Ньютон сформулиро-
вал свои три закона движения. Именно эти законы лежат в основе
механики, о которой пойдет речь в данной книге. Законы Ньютона
относятся к движению изолированных частиц, которые по определе-
нию представляют собой объекты, обладающие массой (и поэтому,
Например, Земля притягивает их), но не имеющие конечного
16 Часть I Основы механики
размера (точечные объекты). Разумеется, размеры каждого
реального тела, даже такого небольшого, как атом или электрон,
конечны, но тем не менее законы механики прямо применимы как
к изолированным реальным телам (будь то стрела в парадоксе
Зенона, Земля, движущаяся вокруг Солнца, или отдельная клетка
крови), так и к сплошным средам, которые могут деформировать-
ся самым разным образом. Такими деформируемыми средами яв-
ляются все упругие тела (сталь, резина, стенки кровеносных сосу-
дов и т. д.), любые жидкости (вода, патока, плазма крови) и газы
(воздух). Поскольку движение жидкостей и газов подчиняется од-
ним и тем же законам, мы будем использовать для них единый
термин — жидкость.
Законы Ньютона применимы к телам конечного размера, по-
тому что, как можно показать, любое тело перемещается таким
образом, будто вся его масса и все действующие на него силы
сосредоточены в одной точке. Эта точка называется центром масс *).
Так, траектория центра масс стрелы Зенона такая же, как и у
частицы с той же массой, на которую действуют такие же, как на
стрелу, силы тяжести и сопротивления воздуха. Движение Луны и
Земли и других планет в космическом пространстве также может
быть описано в рамках механики частиц. Это же относится и к
движению центра масс клетки крови, если только известны силы,
действующие на клетку со стороны плазмы. Однако «кувыркание»
клеток крови или вращение Земли вокруг ее оси, как и любое дру-
гое движение тела относительно его центра масс, зависит от осо-
бенностей формы тела, и такие движения нельзя описать, если
рассматривать тела просто как частицы.
Обосновать применимость законов Ньютона к движению сплош-
ных деформируемых сред труднее. Для этого сначала необходимо
объяснить, почему интересующие нас жидкости и упругие тела
можно считать сплошными — ведь давно известно, что все веще-
ства состоят из молекул, объединенных в разнообразные конфи-
гурации силами разной величины* 2 * * * * * *). Молекулы в свою очередь со-
стоят из атомов, а атом — из ядра, расположенного в центре, и
окружающего его облака электронов, которые движутся по орби-
*) Центр масс тела совпадает с центром тяжести. Если тело последовательно
подвешивать на нити, прикрепленной к разным его точкам, то окажется, что в
нем существует единственная точка, через которую проходят все прямые, пред-
ставляющие собой продолжение линии нити. Эта точка является центром тя-
жести.
2) В твердом теле межмолекулярные силы чрезвычайно велики и взаимное
расположение молекул изменяется незначительно; расстояния между молекулами
сравнимы с их размерами. В жидкости межмолекулярные силы слабее, молекулы
могут легко перемещаться (хотя межмолекулярные расстояния все еще сравнимы
с размерами молекул) и часто сталкиваются Друг с другом. В газе межмолеку-
лярные силы слабы и расстояния между молекулами горазю больше размеров
самих молекул, но все еще очень малы [примерно 3 10~9 м (30 А) для воздуха
при нормальных температуре и давлении].
Гл. 1. Частицы и сплошные среды 17
там с диаметрами, гораздо большими, чем диаметр ядра. Движе-
ние электронов вокруг ядра во многом аналогично движению пла-
нет вокруг Солнца, и так же, как в Солнечной системе, большая
часть атома (а следовательно, и вещества) представляет собой
ие занятое ничем пространство. Некоторые характерные размеры
объектов, о которых мы уже упоминали или о которых пойдет
речь ниже, представлены в табл. 1.1. Можно было бы считать, что
каждое ядро, каждый электрон, каждый атом и даже каждая мо-
лекула суть частицы, и попытаться определить движение каждой
из них под действием межмолекулярных сил из законов Ньюто-
на. Однако такая задача невыполнима, поскольку например, в
1 см3 воздуха при обычных температуре и давлении содержится
примерно 1О20 молекул и положение каждой из них нужно было
бы установить точно.
Таблица 1.1
Характерные размеры некоторых объектов
и расстояния между ними (в метрах)
Диаметр ядра атомов атома или молекулы газа молекулы полимера О КЗ ООО 1 1 L ® о СР
Расстояние между молекулами газа 3-10-9
Диаметр эритроцита капилляра артерии Земли Солнца Солнечной системы Галактики 8-10-6 (4 — 10)-10~6 ю-2 1,2-10’ 1,4 • 109 1,2 • 10*« Ю20
Расстояние между Галактиками 10м
Выход из положения можно найти, опираясь на тот факт, что
расстояние между молекулами обычно гораздо меньше, чем раз-
меры естественных или созданных в эксперименте областей, за-
нятых жидкостями, движения которых мы хотим описать (см.
табл. 1.1). Мы можем мысленно разбить объект на большое число
элементов, размеры которых очень малы по сравнению с разме-
рами объекта, но каждый из которых еще содержит очень боль-
шое число молекул. С точки зрения описания объекта подобный
элемент фактически является точечным, и поэтому его можно рас-
сматривать как частицу; однако с точки зрения движения на мо-
лекулярном уровне элемент весьма велик, и его обобщенные свой-
ства, такие, как скорость Движения или плотность содержащегося
18 Часть I. Основы механики
в нем вещества, могут быть получены путем усреднения по всем
входящим в него молекулам. Следовательно, мы можем пренеб-
речь хаотической природой молекулярного движения и считать
объект сплошным. Это позволяет нам применять законы Ньютона
к каждому элементу объекта (в случае жидких сред он называет-
ся элементарным объемом или частицей жидкости), получая при
этом точное и рациональное описание движения объекта как це-
лого.
В крови присутствуют довольно крупные молекулы [например,
липопротеиды, диаметр которых (3—5)-10~8 м], и хотя течет она
по очень узким трубочкам (диаметр некоторых капилляров со-
ставляет всего 4-10~6 м), диаметр последних гораздо больше раз-
мера молекул. Поэтому плазму крови можно считать сплошной
средой в указанном выше смысле. Цельную же кровь считать та-
ковой можно не всегда, поскольку она содержит не только плазму,
но и множество клеток (главным образом эритроцитов, см. гл. 10),
на долю которых в норме приходится около 45% всего объема
крови. С точки зрения описания движения крови было бы удобно,
если бы эти клетки были малы и содержались в крови в таком
количестве, чтобы индивидуальным поведением каждой из них
можно было пренебречь. Тогда их влияние на движение цельной
крови, рассматриваемой как сплошная среда, можно было бы
описывать усредненно. Именно так обстоит дело для больших ар-
терий (диаметр аорты, например, примерно в 2000 раз больше диа-
метра эритроцита). Но диаметр капилляра сравним с диаметром
эритроцита, и при описании потока в таких мелких сосудах плазму
и клетки крови необходимо рассматривать раздельно. Подводя
итог, можно сказать, что в крупных артериях цельная кровь яв-
ляется фактически сплошной средой, а в микрососудах — нет;
плазму можно считать сплошной средой в обоих случаях.
Далее мы рассмотрим основы механики сплошных сред более
глубоко, но сначала кратко изложим законы Ньютона примени-
тельно к отдельным частицам. При этом математические символы
будут использоваться нами исключительно для удобства — как
краткие обозначения, помогающие точно описать законы меха-
ники. Значения этих символов разъясняются по ходу изложения.
Читателю, хоть немного знакомому с высшей математикой, боль-
шая часть обозначений окажется известной.
Глава 2
МЕХАНИКА ЧАСТИЦ
2.1. Положение частицы
Описать движение частицы — это значит точно указать ее по-
ложение в пространстве, меняющееся в ходе движения. Чтобы ре-
шить эту задачу, представим себе, что в пространстве проведены
три фиксированные прямые, причем все они проходят через дан-
ную точку О и каждая из них перпендикулярна двум другим. На-
глядным примером таких прямых могут служить линии пересече-
ния двух стен и пола комнаты; точкой О в этом случае является
угол комнаты. Если по одной из стенок комнаты ползет муха
(рис. 2.1), то мы можем указать ее положение в некоторый мо-
мент времени, измерив расстояния от нее до пола (обозначим его
через z) и до перпендикулярной стенки (обозначим его через х).
Если муха летает по комнате, то ее положение можно указать
точно так же, измерив расстояние от нее по перпендикулярам до
трех взаимно перпендикулярных плоскостей (пола и двух стен).
Таким же образом можно задать положение любой точки Р. До-
пустим, что из точки Р опущены перпендикуляры на оси и что они
пересекают эти оси в точках X, Y, Z (рис. 2.1,Б). Тогда длины
трех отрезков, а именно ОХ (обозначим эту длину через х), OY (у)
и OZ(z), однозначно укажут нам положение точки Р. Длины от-
резков называются координатами точки Р по отношению к трем
осям, проходящим через точку О. Прямые OX, OY и OZ обычно
называют осями х, у и z соответственно. Используя теорему Пи-
фагора, можно показать, что расстояние между точками Р и О
равно V'х2 + у2 -f- z?; эта величина не зависит от направления
осей.
Важно не забывать два момента, неявно заложенных в подоб-
ном способе описания положения точек. Первый — выбор точки О
и трех координатных осей в принципе произволен, но если уж он
сделан, то должен оставаться неизменным. В противном случае
обсуждение, например, взаимодействия между двумя частицами,
координаты которых указаны в координатных системах, образо-
ванных разными углами комнаты, было бы затруднено. При вы-
боре осей обычно руководствуются соображениями удобства —
например, если частица перемещается по плоскости (муха на
стене), то разумно направить одну из осей (скажем, OY) перпен-
дикулярно этой плоскости, с тем чтобы координата у оставалась
постоянной и нужно было указывать длины только двух отрез-
20 Часть I. Основы механики
ков — х и г. Второй момент состоит в том, что единицы, в которых
измеряются длины отрезков х, у, г, должны указываться в явном
виде и быть одинаковыми для всех осей. Длина — это не просто
число; это физическая величина со своей размерностью, и для ее
измерения необходимы соответствующие единицы. В этой книге
для измерения длины мы, как правило, будем использовать метр
Рис. 2.1. А. Положение мухи на боковой стенке комнаты определено через рас-
стояние до нее от торцевой стенки (х) и от пола (z); х и z являются координа-
тами мухи относительно осей, образованных прямыми OX, OZ. Б. Положение
точки Р в трехмерном случае (например, положение мухи, летающей по комна-
те) можно определить, указав расстояния (х, у, г) до нее от трех взаимно пер-
пендикулярных плоскостей (две стенки и пол). Точка Р имеет координаты (х,
у, г). Началом координат в этом случае является угол комнаты О.
(м), сантиметр (1 см = 10~2 м) и микрон (1 мкм = 10~® м). Во,
лее полно вопрос о единицах измерения будет рассмотрен в гл. 3.
2.2. Скорость
Еще одной важной для описания движения частицы величиной
является скорость, или быстрота, с которой меняется положение
частицы. Рассмотрим частицу, движущуюся вдоль прямой ОХ
(рис. 2.2, Л). Ее положение полностью задается одной координа-
той-расстоянием х от точки О. Если координата точки в момент
времени t есть х, а в следующий момент, t’, — х’, то средняя ско-
рость движения частицы в интервале времени от t до Г составит
v = (х'— —t). Это определение справедливо для любого
сколь угодно малого интервала времени, даже если t'—t и, сле-
довательно, х'— х стремятся к нулю. Величину, к которой стре-
мится v, когда f—1->0, называют мгновенной скоростью час-
тицы в момент времени t\ эту величину записывают в виде
v =
dx
ЧТ-
(2.1)
Ясно, что скорость частицы, движущейся в обратном направлении,
т. е. к точке О, отрицательна, поскольку х' < х.
Гл. 2. Механика частиц 21
Положение
Время
Рис. 2.2. А. Частица, движущаяся по прямой ОХ, в момент времени t находится
на расстоянии х от неподвижной точки О, а в более поздний момент времени
t' — на расстоянии х'. Величина (х‘ — х)/ ((' — /) является средней скоростью ча-
стицы за интервал времени от t до t'. Если этот интервал делать все более ма-
лым, так что величина t' — t будет стремиться к нулю, то х' — х также станет
уменьшаться, но средняя скорость будет стремиться при этом к вполне опреде-
ленному пределу (ц). Эта величина и является скоростью частицы в момент
времени t. Б. На верхнем графике представлена зависимость пройденного части-
цей расстояния х от времени t. Величина (х'— x)/(t' — f) равна наклону пря-
мой РР' (и равна tg <р). Когда V — t стремится к нулю, прямая РР' прибли-
жается к касательной к кривой в точке Р (касательная изображена на рисунке
щтрихпунктирной линией), наклон которой равен t>( = tgi|>), т.е. скорости ча-
стицы в момент времени t. На нижнем графике представлена соответствующая
этому движению зависимость v от t.
Определение Ах/At можно пояснить графически (рис. 2.2,Б).
В верхней части этого рисунка представлено изменение х во вре-
мени. Рассмотрим две точки на графике — Р (х, t) и Р' (х', f). Ве-
личина (х' —x)/(f— t) равна тангенсу угла <р между прямой,
соединяющей эти две точки, и осью t. Эта величина называется
наклоном данной прямой. Когда t'—t стремится к нулю, точка Р'
приближается к точке Р, а соединяющая их прямая приближается
к касательной к кривой в точке Р (штрихпунктирная линия на
рис. 2.2). Из рисунка видно, что величина Ax/At (т. е. о) является
наклоном касательной к кривой в точке Р и равна по величине
tgty. На рис. 2.2, Б представлен также график изменения v во
времени.
Разрешение парадокса Зенона заключается в проведении опи-
санной выше процедуры предельного перехода; без него опреде-
22 Часть 1. Основы механики
лить мгновенную скорость частицы через ее положения в последо-
вательные моменты времени невозможно.
Идея предельного перехода была сформулирована лишь
в XVII в, когда Ньютон и Лейбниц создали основы теории диф-
ференциального исчисления. Символ d/d/, принятый в этой обла-
сти математики, обозначает скорость изменения некоторой вели-
чины во времени В рассмотренном выше примере все содержание,
которое мы вкладываем в выражение dx/dt,— это скорость, с ко-
торой х изменяется во времени. Единицы измерения скорости
цолжны соответствовать выбранным единицам измерения расстоя-
ния и времени. Если расстояние измерено в метрах, а время —
в секундах, то скорость необходимо измерять в метрах за секунду
(м-с- *).
Приведенное выше определение скорости легко обобщить на
случай трехмерного движения частицы. Если упоминавшаяся уже
муха летит из одного угла комнаты в Другой, то все ее коорди-
наты изменяются во времени. Чтобы полностью определить движе-
ние, необходимо указать, каким образом изменяются эти коорди-
наты. Точка X на рис. 2.1 перемещается вдоль оси х со скоростью
vx = dx/dt, точка Y движется вдоль оси у со скоростью vy =
= dy/dt, точка Z движется вдоль оси г со скоростью vz = dz/dt.
Скорость (трехмерная) точки Р, таким образом, полностью опре-
деляется тремя величинами (vx, vy, vz), которые называются со-
ставляющими скорости точки Р в направлениях х, у и z соответ-
ственно. Ясно, что значения составляющих зависят от ориентации
координатных осей, но не зависят от положения начала коорди-
нат— точки О. Полная скорость перемещения точки Р, т. е. со-
ставляющая ее скорости вдоль линии, строго параллельной на-
правлению движения, не зависит от ориентации осей. Можно по-
казать, что эта скорость (ее называют абсолютной величиной, или
модулем скорости) равна ^v2x + v2y + v2z ; она всегда положитель-
на, даже если одна или все составляющие отрицательны.
С таким же успехом мы можем задать скорость движения час-
тицы в трехмерном пространстве и другим способом, а именно
указав абсолютную величину скорости и направление движения
относительно любых двух координатных осей (например, направ-
ление движения мухи и абсолютная величина ее скорости пол-
ностью определяют скорость). Если абсолютная величина ско-
рости есть V, а углы, которые она составляет с осями х и у, равны
соответственно 6 и <р (рис. 2.3), то составляющие vx и vy задаются
выражениями
vx = V cos 0,
vy == V cos <p *).
*) Предполагается, что читатель знаком с элементарными свойствами сину-
сов и косинусов.
Гл. 2. Механика частиц 23
При этом третья составляющая равна
— v^-v2 = V д/(1 — cos2 0 — cos2<p)
и равна также V cos ip, где ip — угол между направлением скоро-
сти и осью 2.
Скорость частицы является примером физической величины,
характеризующейся определенной абсолютной величиной и опре-
деленным направлением. Скорость существует независимо оттого,
каким способом мы ее измеряли, хотя по абсолютной величине
Рис. 2 3 Стрелка с обозначением v пред-
ставляет скорость частицы, имеющую абсо-
лютную величину V и определенное на-
правление Составляющая скорости в на-
правлении произвольной координатной оси
равна произведению V на косинус угла меж-
ду осью и вектором скорости Так, vK =
= V COS 6, Vu = V COS <p, V? — V cos i|)
она зависит от выбранных единиц измерения. Величины, имею-
щие определенное направление, зависят от ориентации коорди-
натных осей; они называются векторами и обозначаются симво-
лами, выделенными полужирным шрифтом. Трехмерный вектор
скорости частицы, например, может быть записан одним симво-
лом v. Величины (vx, vy, vz) являются составляющими вектора v,
и последний можно рассматривать как эквивалент трех его со-
ставляющих, взятых в совокупности. В дальнейшем мы часто бу-
дем использовать запись v = (vx, vy, vz).
Другим примером векторной величины является вектор, задаю-
щий положение частицы Р (радиус-вектор) (рис. 2.1, Б). Его абсо-
лютная величина равна длине отрезка OP (V%2 + J/2 + z2), направ-
ление задается косинусом углов между ОР и любыми двумя (из
трех) координатных осей, а составляющими являются сами коор-
динаты х, у, г. Этот радиус-вектор (обозначим его через х) отли-
чается от рассмотренных нами ранее векторов, поскольку он за-
висит от положения начала координат О. Все другие векторы,
характеризующие физические величины, подобные скорости, от по-
ложения начала координат не зависят. Векторная форма записи,
как и запись d/d/ для обозначения «скорости изменения чего-
либо»,— это прежде всего удобная форма сокращения. Так как
по определению vx, vy, vz являются скоростями изменения х, у, г
(vx = dx/dt и т д), мы можем объединить обе формы записи, по-
лучив выражение
v = (2-2)
24 Часть I. Основы механики
Скорость (вектор) есть быстрота изменения положения (которое
также задается вектором).
Чтобы сложить векторные величины одного и того же типа
(например, две скорости или два радиус-вектора), нужно сложить
их составляющие. Так, если xt = (хь yit zt) и х2 = (х2, у2, z2)—
два таких вектора, то
Х1 + х2 = (*1 "Ь Х2г У\ + У2, 21 + z2). (2.3)
Действительно, рассмотрим двумерный случай (рис. 2.4) и сложим
вектор Xi=(xi,i/i), представляющий точку Рь с вектором х2 =
= (х2, Уъ), представляющим точку Р2. В рамках геометрической
Рис. 2 4. Геометрическая интерпретация операции сложения векторов, определен-
ной уравнением (2 3), для двумерного случая. Веекторы Х] и х2, представленные
отрезками ОР[ и ОР2, при сложении дают вектор Х| + х2, представленный отрез-
ком OQ. О и Q являются противоположными вершинами параллелограмма, боко-
вые стороны которого есть ОР| и ОР2.
интерпретации вектор суммы будет представлять точку Q с коор-
динатами (xi + х2, yi + У2). Это согласуется с выражением (2.3).
Рассмотрим, например, снаряд, который вылетает из неподвиж-
ного орудия со скоростью v = (vx, vy, vz). Предположим теперь,
что орудие движется по земле со скоростью U в направлении оси
х [вектор скорости V = (U, 0, 0)]. В этом случае скорость сна-
ряда относительно Земли возрастает на величину U в направле-
нии х, а в направлениях у и г останется неизменной, т. е. v -f- V =
= (Vx + U, Vy, Vz).
2.3. Ускорение
Подобно определению скорости частицы как быстроты измене-
ния ее положения, ускорение частицы определяется как быстрота
изменения скорости; это определение тоже может быть представ-
лено в виде математического выражения. При движении по пря-
мой ускорение есть Av/At и равно тангенсу угла наклона каса-
тельной к кривой, описывающей зависимость скорости v от вре-
мени t (рис. 2.2, Б). Ускорение, как и скорость, имеет три состав-
ляющие, равные быстроте изменения составляющих скорости, и
так же, как скорость, является вектором (обозначим его через а):
а <£t \ df ’ dt ’ dt )
Гл. 2. Механика частиц 25
В обозначениях, принятых в дифференциальном исчислении, если
и = dx/d(, то Au/At = d2x/d/2. Эта запись является кратким обо-
значением быстроты изменения быстроты изменений х. Таким обра-
зом, мы можем написать
а —— —Г— d2g —(2 51
а ~~ d/2 d/2 ’ ’ d/2 J ’
В соответствии с единицами измерения длины и времени ускоре-
ние должно измеряться в метрах за секунду в квадрате (м-с-2).
Четко уяснить смысл ускорения, когда оно является трехмер-
ной величиной, вероятно, не столь просто. Лучше обратиться к од-
номерному движению частицы по прямой ОХ: если в данный мо-
мент времени скорость v увеличивается, то ускорение а = Av/At
положительно; если же v уменьшается, то а отрицательно. Когда
частица движется по направлению к началу координат, ее ско-
рость v отрицательна; но если движение частицы в это время за-
медляется, то ее ускорение а положительно. Для того чтобы лучше
понять все сказанное выше, обратимся к рис. 2.5, где приведены
графики изменения во времени величин х, v и а для частицы, ко-
торая «стартует» из положения покоя в точке О и движется с ус-
корением до тех пор, пока не достигнет определенной скорости.
Последняя сохраняется неизменной в течение некоторого вре-
мени, а затем уменьшается с постоянной скоростью (частица дви-
жется с постоянным отрицательным ускорением). Далее скорость
меняет знак (частица начинает возвращаться к точке О) и дости-
гает постоянного отрицательного значения, равного по абсолют-
ной величине предшествующей положительной скорости. Через не-
которое время движение частицы начинает замедляться (с поло-
жительным ускорением), и она останавливается в исходной точ-
ке О. В приведенном примере направление ускорения (знак а) не
зависит от направления движения (т. е. от знака v).
При движении в двух- и трехмерном пространстве знак уско-
рения также не зависит от направления скорости. Частица приоб-
ретает ускорение при любом характере изменения вектора ско-
рости — будь то изменение его величины или направления. Пред-
положим, например, что частица движется по окружности с по-
стоянной скоростью, подобно шарику, привязанному к нити, или
спутнику, летящему по орбите вокруг Земли. В этом случае абсо-
лютная величина скорости постоянна, но частица испытывает уско-
рение, поскольку меняется направление скорости (рис. 2.6). Так
как абсолютная величина скорости постоянна, это ускорение долж-
но быть перпендикулярно направлению движения (в противном
случае имелась бы составляющая ускорения в направлении дви-
жения и абсолютная величина скорости изменялась бы), и, следо-
вательно, вектор ускорения проходит через центр окружности.
Скорость (ее абсолютная величина равна ц), имеющая в момент
времени t определенное направление, через небольшой промежу
26 Часть 1 Основы механики
Рис 2 5 Графики зависимости от времени t пройденного по прямой расстояния
х, скорости v и ускорения а для частицы, которая начала свое движение из со
стояния покоя в точке х = 0 в момент времени t = 0 До момента времени ti
частица движется с постоянным ускорением at и достигает скорости V] Эта
скорость поддерживается до момента /г, после чего частица с постоянным отри
цательным ускорением — а1 начинает перемещаться и движется так до тех пор,
пока не изменит направление своего движения и не достигнет в момент времени
t = t4 скорости, равной по абсолютной величине скорости, с которой она ранее
удалялась от начала координат (т е скорости —Vi) В момент времени f3 ско
рость частицы нулевая, и она наиболее удалена от начала координат Постоян
ная скорость —V, сохраняется до момента t5, после чего частица замедляется
с постоянным ускорением а1 и останавливается в начале координат в момент te
W
Рис 2 6 Частица, движущаяся по
окружности радиусом г с постоян-
ной скоростью v, имеет ускорение,
которое равно по абсолютной ве-
личине о2/г и направлено к центру
окружности Угол 6 равен углу, на
который поворачивается прямая,
соединяющая частицу с центром
окружности, за время от t до t'.
Гл 2 Механика частиц 2Л
ток времени (в момент t') приобретет составляющую в перпенди-
кулярном направлении, обусловленную ускорением. Вектор ско-
рости при этом несколько изменит свое направление. Составляю-
щая скорости, приобретенная за время t' — t, равна произведению
v иа удвоенный синус угла у 0 Выражение 2 sin О при очень
малых 6 равно расстоянию, которое частица проходит за данное
время, т.е. vXU'—/), деленному на радиус окружности г. Таким об-
разом, перпендикулярная составляющая скорости равна о2(/'—<)/г,
а скорость ее изменения (при С—/->0) составляет v2/r. Эта фор-
мула дает значение абсолютной величины ускорения, направлен-
ного к центру (центростремительного ускорения). Планеты, дви-
жущиеся вокруг Солнца, и спутники, летящие вокруг Земли,
имеют ускорение, направленное соответственно к Солнцу и к
Земле Точно так же и частица крови, движущаяся в потоке па-
раллельно стенкам дуги аорты, должна иметь ускорение, направ-
ленное к центру кривизны траектории.
2.4. Законы движения Ньютона: масса и сила
Итак, мы уже умеем описывать положение, скорость и уско-
рение частицы, движущейся по данной траектории в пространстве.
Но что приводит частицы или тела в движение и что останавли-
вает их? — Силы К сожалению, дать точное определение силы
довольно трудно, хотя все мы имеем интуитивное представление
о ней, основанное на нашем повседневном опыте С точки зрения
логики рассуждение, которое мы будем использовать для опре-
деления силы, образует порочный круг, так как мы говорим, что
сила вызывает движение, но саму силу определяем через движе-
ние, которое она вызывает. Тем не менее с научной точки зрения
это рассуждение приемлемо, так как оно основано на результа-
тах простых опытов, которые устраняют всякую двусмысленность
Когда мы поднимаем тяжелый груз, то прикладываем боль-
шую силу, чем при подъеме легкого груза. Если мы не хотим,
чтобы автомобиль, стоящий на склоне, покатился вниз, то должны
приложить силу, направленную вверх вдоль склона. Мы можем
остановить движение катящегося по столу шара, приложив к нему
силу, но, если никаких сил не прикладывать, шар будет продол-
жать двигаться дальше У нас нет необходимости привлекать
таинственную силу, которая, как полагали древние греки, необхо-
дима для поддержания движения тела, — мы можем просто ска-
зать, что тело продолжает движение с постоянной скоростью до
тех пор, пока к нему не будет приложена сила, которая изменит
эту скорость Чтобы удовлетворительно объяснить наши повсед-
невные наблюдения, следует, как показал Ньютон, связывать силы
не со скоростью движения тел, а с ее изменением, или, что то же
самое, с ускорением. Если сила равна нулю, то и ускорение равно
28 Часть I Основы механики
нулю т е скорость постоянна (она в свою очередь равна нулю,
если тело покоится).
Какова же в таком случае связь между силой и ускорением?
Если два шара, один из которых тяжелее другого, катятся с оди-
наковой скоростью по плоской горизонтальной поверхности, а мы
намерены остановить их за одно и то же время (т. е. сообщить
им одно и то же отрицательное ускорение), то к более тяжелому
шару следует приложить большую силу. Свойство, вследствие ко-
торого более тяжелый шар труднее как остановить, так и сооб-
щить ему положительное ускорение, называется инерцией. В этой
книге мы будем использовать слово «инерция» только как чисто
качественное понятие. Величиной, характеризующей инерцию тела,
является его масса, которая есть мера содержащегося в теле ко-
личества вещества. Тело с большей массой труднее заставить дви-
гаться с ускорением. Именно большая масса приводит к тому, что
один предмет оказывается тяжелее другого, т. е. обладает боль-
шим весом. Единица массы не зависит от единиц длины и времени,
а три эти единицы вместе образуют фундаментальную систему
единиц, при помощи которой измеряются все другие механиче-
ские величины. В данной книге мы применяем в качестве основ-
ной единицы массы килограмм (кг), хотя некоторые величины при-
водятся в граммах (1 г = 10~3 кг) (см. гл. 3). '
Точная связь между силой, массой и ускорением, из которой
смысл определения силы становится более ясным, сформулирована
в законах движения Ньютона. Изложим эти законы.
Первый закон. Каждая частица продолжает оставаться в со-
стоянии покоя или равномерного прямолинейного движения до тех
пор, пока на нее не действует внешняя сила или совокупность
сил. Другими словами, скорость v остается постоянной (или ну-
левой), если результирующая сила равна нулю (см. ниже).
Второй закон. Если на частицу массой m действует сила или
совокупность сил, которые сообщают ей ускорение а, то результи-
рующая сила равна произведению массы на ускорение. Другими
словами, результирующая сила — это вектор (обозначим его че-
рез F), задаваемый выражением
F = та. (2.6)
Это уравнение часто называют уравнением движения частицы.
Под результирующей силой понимается сумма всех сил, дей-
ствующих на частицу, причем природа этих сил может различать-
ся. Рассмотрим, например, тело, которое мы толкаем по горизон-
тальному столу (рис. 2.7). На него действуют следующие силы:
сила гравитационного притяжения Земли (т. е. вес тела W), сила
R, с которой стол действует на тело (в общем случае она состоит
из двух компонент — силы N, поддерживающей тело снизу, и силы
трения Т, действующей в направлении, противоположном движе-
нию), и, наконец, сила Р, направленная вперед (та самая, кото-
Гл 2 Механика частиц 29
рую мы приложили). Если при этом ускорение, с которым тело
движется вперед, равно а, то
Р + R + W = ma. (27)
Чтобы сложить силы, нужно сложить их составляющие [уравне-
ние (2.3)]. В соответствии с этим, если мы выберем оси х и у так,
как это показано на рисунке (предполагается, что ось z перпенди-
кулярна плоскости рисунка), и примем во внимание, что Р =
= (Р,0,0), R = (—T,N,0), W = (0,— №,0), а —(а,0,0), то урав-
нение (2.7) приобретет вид
(Р — Т, N-W, 0) = (иш, 0, 0).
Очевидно, два вектора могут быть равны только в том случае,
когда равны все их составляющие Отсюда следует, что N—W =
= 0 (т. е. направленная вверх сила, с которой стол действует на
предмет, равна весу предмета, и вертикальное ускорение отсут-
ствует), Р — Т = та. Последнее выражение связывает горизон-
тальную составляющую силы с ускорением тела.
Рис 2 7 На схеме представлены
все силы, которые действуют на
тело, перемещаемое вправо по го-
ризонтальной плоскости Р — при-
ложенная сила, W — вес тела, R —
сила, действующая на тело со сто-
роны стола Сила R имеет две со-
ставляющие N, направленную вер-
тикально вверх, которая присут-
ствовала бы, даже если бы тело
было неподвижно, и Т, обуслов-
ленную трением и направленную
против движения
Из уравнения (2.6) мы получаем единицу силы: так как масса
измеряется в кг, а ускорение — в м-с-2, то размерность силы
должна быть кг-м-с-2 (килограмм-метр в секунду за секунду).
Эта единица названа, как ей и подобает, ньютоном (между про-
чим, вес яблока среднего размера равен примерно одному ньюто-
ну). Подробнее вопрос о единицах мы рассмотрим в гл. 3 и 6, а
пока отметим, что все величины, входящие в любое уравнение,
подобное, например, уравнению (2.6), должны быть выражены
в правильных единицах. Более того, полезный способ проверки
непротиворечивости самого уравнения заключается именно в уста-
новлении идентичности размерностей его правой и левой частей.
Например, длина никогда не может быть равна площади, ско-
рость— ускорению, сила — массе; уравнения, в которых встре-
чаются подобные несоответствия, не просто ошибочны, но и бес-
смысленны.
30 Часть I. Основы механики
Третий закон. Каждому действию есть равное и противополож-
но направленное противодействие. Так, если одно тело действует
с силой F на другое, то второе тело действует на первое с равной
по величине, но противоположно направленной силой —F. Обра-
тимся к примеру, приведенному на рис. 2.7. Поскольку стол дей-
ствует на тело с силой, одна компонента которой направлена
вверх (N), а другая — против направления движения (—Т), то
и тело действует на стол с силой, направленной как вниз (—N),
так и по направлению движения (Т). Если вы задерживаете ка-
тящийся шар, приложив к нему силу, то ощущаете, что шар дей-
ствует на вас с противоположно направленной силой. Держа в
руках тяжелое тело, вы действуете на него с силой, направленной
вверх, потому что тело действует на вас с силой, направленной
вниз; вы действуете с силой, направленной вниз, и на Землю, а
она в свою очередь действует на вас с силой, направленной в про-
тивоположную сторону.
Рассматривая тело, находящееся на горизонтальной плоскости
(рис. 2.7), можно выделить силы двух видов — дальнодействую-
щие и близкодействующие.
Прежде всего сказывается вес тела, который есть проявление
гравитационного взаимного притяжения между любыми двумя те-
лами, обладающими массой. В данном случае одним из тел яв-
ляется Земля. Дальнодействующая сила, с которой Земля притя-
гивает частицу массы т, расположенную около ее поверхности
(она равна весу частицы W), направлена примерно к центру
Земли (т. е. вертикально вниз) и пропорциональна массе т. Ее
абсолютная величина в разных точках Земли несколько разли-
чается, но в любом локальном эксперименте может считаться по-
стоянной. Таким образом, мы можем записать, что
W — mg, (2.8)
где g— вектор, абсолютная величина которого постоянна и кото-
рый в каждой точке направлен вертикально вниз. Сопоставление
уравнений (2.8) и (2.6) показывает, что g имеет размерность уско-
рения, а, согласно уравнению (2.6), свободно падающее тело,
на которое действует единственная сила — его собственный вес,
будет испытывать направленное вниз ускорение g. g называется
ускорением силы тяжести; его абсолютная величина равна при-
мерно 9,8 м-с~2. Тот факт, что по абсолютной величине g при-
мерно постоянно, дает возможность очень просто измерить массу
тела, сравнив его вес с весом некоторого стандартного тела.
Гравитационное притяжение является дальнодействующей си-
лой в том смысле, что оно действует на любое тело со стороны
всех других тел независимо от того, соприкасаются они с ним или
нет. Другим примером силы такого типа является сила, которая
действует на заряженную частицу, движущуюся в электрическом
или магнитном полях.
Гл. 2. Механика частиц 31
Остальные силы, действующие на тело, изображенное на
рис. 2.7, являются близкодействующими. Они проявляются только
при соприкосновении тел. Р — сила, которую мы прикладываем к
телу, когда толкаем его вперед или тащим за собой. R — сила,
с которой стол действует на тело и которая не существовала бы,
если бы тела не соприкасались. Ее нормальная (перпендикуляр-
ная плоскости движения) составляющая N не равна нулю потому,
что тело и стол прижаты друг к другу. Подобная сила возникает
всякий раз, когда тела соприкасаются (когда вы дотрагиваетесь
до чего-либо, то ощущаете объект только потому, что он действует
на вас с силой, перпендикулярной поверхности вашего тела). Тан-
генциальная компонента Т противодействующей силы R— это сила
трения. Будь тело абсолютно гладким, она бы отсутствовала. Для
двух данных поверхностей, перемещающихся относительно друг
друга, Т обычно пропорциональна N(T — CN). Константу пропор-
циональности С называют коэффициентом трения. Если приложен-
ная сила Р меньше, чем CN, то тело не движется (а = 0) и реаль-
ная сила трения Т как раз такова, чтобы предотвратить движение,
т. е. равна Р. Дальше мы узнаем, что силы, действующие на «час-
тицы» сплошного деформируемого материала, тоже можно разде-
лить на дальнодействующие и близкодействующие.
Три закона Ньютона лежат в основе всей механики. Но прежде
чем распространить их на случай сплошных жидких и упругих
сред, полезно разобрать два примера применения этих законов
для анализа движения частиц (или центра масс больших тел, что
в принципе одно и то же).
1. Если шофер ведет автомашину (общей массой А1) со ско-
ростью V и желает остановить ее, то он обычно тормозит так,
чтобы создать приблизительно постоянную силу торможения F,
действующую в направлении, противоположном движению. Уско-
рение при этом равно F/7V1 [второй закон Ньютона, уравнение
(2.6)]. Составляющая F в направлении движения, конечно, отри-
цательна и замедляет движение. Водитель (массой tri) остается
неподвижным относительно машины, потому что он действует на
нее (нажимая на педали и упираясь руками в руль) с небольшой
силой, направленной по ходу ее движения, и испытывает со сто-
роны машины такую же по абсолютной величине противодей-
ствующую силу (третий закон Ньютона). Для того чтобы води-
тель двигался с таким же отрицательным ускорением, как и весь
автомобиль, абсолютная величина этой силы должна равняться
произведению абсолютной величины F на m/tA. Если сила тормо-
жения велика и прикладывается очень резко (например, при столк-
новении с деревом), то водитель может не суметь развить доста-
точно большую силу воздействия на автомобиль, чтобы тот в свою
очередь сообщил водителю такое же ускорение, как и у него са-
мого В этом случае водитель будет продолжать двигаться относи-
82 Часть 1 Основы механики
тельно машины вперед, пока не упрется, например, в руль или
пока его не остановят ремни безопасности.
2. Рассмотрим частицу массой tn, движущуюся по окружности
с постоянной скоростью (рис. 2.8, А). Как было показано, при та-
ком движении частица испытывает ускорение, направленное к
центру. Следовательно, согласно второму закону Ньютона, на нее
должна действовать сила в том же направлении. Если частица
представляет собой шарик, подвешенный на нити, то эта сила
Частица движется
по прямой со
скоростью и
Рис. 2.8. А. Частица, подвешенная на нити фиксированной длины, движется по
окружности радиусом г со скоростью v Натяжение нити должно быть равно
произведению массы частицы (т) на ее ускорение, которое направлено к центру
окружности (у2/г). Б. Если нить разрывается, на частицу перестают действовать
какие-либо силы и она летит по прямой со скоростью V.
прикладывается со стороны нити и называется ее натяжением1).
Если речь идет о планете, движущейся вокруг Солнца, эта сила
имеет гравитационную природу. Абсолютная величина силы равна
массе т, умноженной на абсолютную величину ускорения, т. е.
то2/г, где v — скорость частицы, а г — радиус окружности. Если
сила внезапно перестает действовать (например, вследствие раз-
рыва нити), то частица больше не испытывает ускорение и летит
по прямой, являющейся касательной к окружности. Скорость по-
лета частицы остается постоянной до тех пор, пока на нее не по-
действует какая-либо другая сила (рис. 2.8, Б). Наблюдая за по-
добным полетом, люди когда-то предполагали, что на частицу,
движущуюся по окружности, действует сила в направлении от
центра (ее называют центробежной силой), которая при движении
по кругу уравновешена натяжением нити. Считалось, что, когда
нить рвется, эта сила заставляет частицу сойти с круговой орбиты.
Такой взгляд на движение по кругу ошибочен: на самом деле
•) Натяжение равно суммарной силе, с которой нить действует на шарик,
и в механике слово «натяжение» всегда означает силу. Не следует путать натя-
жение с растягивающим напряжением, которое испытывает материал нити, так
как последняя величина представляет собой силу, приходящуюся на единицу
площади поперечного сечения нити (гл. 7). Надо также различать натяжение в
указанном смысле и поверхностное натяжение в пленках и мембранах, т. е. силу,
действующую на единицу длины границы. Не следует применять термин «натя-
жение» и к парциальному давлению газа в растворах.
Гл. 2. Механика частиц 88
после обрыва нити исчезает единственная действующая на частицу
сила. Другой пример тела, движущегося по окружности, дает по-
лет вокруг Земли искусственных спутников с экипажем на борту.
Как спутник, так и находящиеся в нем люди движутся по одной
и той же круговой орбите радиусом г и с одной и той же ско-
ростью (скажем, v). Поэтому и люди, и спутник испытывают одно
и то же ускорение о2/г, направленное к центру Земли. На каждое
движущееся таким образом тело должна действовать сила, имею-
щая такое же направление и равная произведению v2/r на массу
тела. Эта сила создается гравитационным полем Земли; она про-
порциональна массе тела и обратно пропорциональна квадрату
расстояния от тела до центра Земли (т. е. F = km/r2, где k — кон-
станта, m — масса тела). Таким образом, можно применить второй
закон Ньютона к любому объекту, движущемся по определенной
орбите вокруг Земли, и получить уравнение
km mv2
— = ~Г-
Отсюда следует, что при выполнении условия k = v2r любой
объект, на который не действуют никакие другие силы, кроме рас-
смотренных выше, будет двигаться по данной орбите. Например,
человек внутри космического корабля и сам корабль не будут
взаимодействовать до тех пор, пока этот человек не начнет пере-
мещаться относительно корабля. Космонавт оказывается в состоя-
нии невесомости в том смысле, что, «плавая» внутри корабля, он
не подвергается действию силы, стремящейся вернуть его на «пол»
корабля. Состояние невесомости распространяется и на кровь кос-
монавта. Для любой частицы крови существует соответствие между
силой ее притяжения к Земле и ускорением по направлению к
Земле, возникающим из-за движения этой частицы по окружно-
сти. Невесомость приводит к серьезным физиологическим пробле-
мам, поскольку в ходе эволюции человек приспособился к таким
условиям, в которых на него постоянно действует сила гравита-
ционного притяжения. Космонавт, вышедший из корабля в откры-
тый космос, находится в тех же условиях, что и в корабле, и не
перемещается относительно корабля, если на него не действуют
дополнительные силы. В открытом космосе нет ни сопротивления
воздуха, ни каких-либо других сил, стремящихся уменьшить ско-
рость полета человека по сравнению со скоростью корабля, так
что нет необходимости использовать для противодействия этим
силам натяжение фала, соединяющего человека с кораблем. По-
этому фал свободно провисает, а не натягивается, как было бы
в том случае, если бы человек вышел из корабля в атмосфере
Земли, где есть сопротивление воздуха.
Прежде чем рассматривать применение законов Ньютона к
сплошным средам, мы должны объяснить смысл еще нескольких
понятий механики.
84 Часть I. Основы механики
2.5. Количество движения
Второй закон Ньютона гласит, что сила равна произведению
массы на ускорение, т. е. на быстроту изменения скорости. Если
масса постоянна, можно сказать также, что сила равна скорости
изменения произведения массы на скорость, или в символической
записи
F = m^T = -^-(mv). (2.9)
Уравнение, записанное в такой форме, является более общим,
так как оно применимо к движению тел с переменной массой (на-
пример, к движению ракеты). Величина tnv называется количе-
ством движения частицы, так что закон Ньютона можно перефра-
зировать так: «Сила равна скорости изменения количества движе-
ния». В отсутствие внешних сил количество движения частицы,
тела или системы остается постоянным, или, как принято говорить,
сохраняется. Если, например, две частицы массой т.\, тч, движу-
щиеся со скоростями Vi, v2, сталкиваются и объединяются, то
вновь образовавшееся тело массой т\ + т2 должно иметь то же
самое количество движения, каким обладали в сумме исходные
частицы, так что его скорость должна быть равна величине
(mjVi + m2v2) / (mi + m2) • Когда столкновение происходит мгно-
венно, эта скорость приобретается тотчас после столкновения даже
в том случае, если на тела действуют внешние силы (например,
гравитация), так как для изменения количества движения под
влиянием таких сил необходимо некоторое конечное время. Если
легковая машина, обладающая массой М, движется со скоростью
V в определенном направлении (скажем, вдоль оси х), так что ее
вектор скорости равен (V, 0,0), а грузовик массой 10 М движется
1 т/ г
в противоположном направлении со скоростью V [вектор ско-
рости равен (—ру V, 0, о)], то их суммарное количество движе-
ния равно нулю, а общий центр масс неподвижен. Он будет по-
коиться и в том случае, если автомобили столкнутся, хотя движе-
ние каждого из них относительно общего центра масс катастрофи-
ческим образом изменится.
2.6. Работа и энергия
Если под действием силы F, приложенной к телу, последнее пе-
ремещается на расстояние d в направлении действия силы, то
говорят, что сила совершает работу, равную Fd. Это положение
остается справедливым и в том случае, когда сила и направление
движения не параллельны друг другу (рис. 2.9), но теперь d равно
не самому расстоянию, а его проекции на направление действия
Гл. 2. Механика частиц 35
силы. Можно сказать и так: совершенная работа равна произве-
дению составляющей силы в направлении движения (Feos6, см.
рис. 2.9) на пройденное расстояние d\, выражения Fd и Fcos6-d!
равны, поскольку d = di cos 0. Если F = (Fi, F2, F3) есть сила, a
положение конечной точки пройденного пути относительно точки,
откуда «стартовало» тело, задается вектором х = (ху х2, х3), то
можно показать, что совершенная работа равна Fi%i -]- F2x -f- F3x3.
Это выражение принято записывать в виде F-x. Когда F не яв-
ляется постоянной величиной или когда направление движения ме-
няется, траекторию частицы необходимо разбить на небольшие
Рис. 2.9. Работа, совершаемая силой абсолютной величины F, действующей иа
частицу, которая перемещается по прямой на расстояние di, равна FdiCOS0,
где 6 — угол межд}' вектором силы и направлением движения. Эта работа равна
также Fd, где d(=d1cos6) является проекцией di на направление действия
силы. Это выражение часто записывают в виде F-x, где F — вектор силы, ах —
вектор, соединяющий конечную точку пройденного пути с начальной точкой.
Направлений
движения
прямолинейные участки, на которых F можно с достаточной точ-
ностью считать постоянной. Складывая величины работы, совер-
шенной на каждом участке, можно найти суммарную работу.
Когда тело движется по окружности с постоянной скоростью
подобно шарику, раскручиваемому на конце нити (разд. 2.4), то
действующая на него сила направлена к центру окружности, т. е.
всегда перпендикулярна направлению движения и потому не со-
вершает никакой работы.
Исходя из второго закона Ньютона, можно показать, что пол-
ная работа, совершенная над частицей за некоторое время всеми
действующими на нее силами, равна изменению величины — tnv2,
где m — масса частицы, a v — ее скорость. Эту величину назы-
вают кинетической энергией частицы. Если мы применим только
что сформулированное правило к движению тела за очень корот-
кий промежуток времени, то получим дополнительный результат:
в каждый момент времени скорость, с которой совершается ра-
бота над частицей, равна скорости увеличения ее кинетической
энергии.
Когда частица движется в гравитационном поле Земли, дей-
ствующая на нее гравитационная сила равна по абсолютной вели-
36 Часть I Основы механики
чине mg и направлена вертикально вниз. Если ось z направлена
вертикально вверх, то составляющие этой силы есть (0,0 — tng).
Введем величину mgz, называемую потенциальной энергией час-
тицы. Работа, совершаемая в единицу времени гравитационной
силой, равна (—mg)&z/&t, или, что то же самое, —&(mgz)/dt,
т. е. взятой со знаком минус скорости изменения потенциальной
энергии. В отсутствие других сил работа в единицу времени долж-
на быть равна скорости изменения кинетической энергии. Другими
словами, скорость изменения полной энергии (кинетической и по-
тенциальной) равна нулю; этот результат называют принципом
сохранения энергии:
Кинетическая энергия + потенциальная энергия =
= у mv2 + mgz — Е = const. (2.10)
Величина Е, вообще говоря, произвольна, так как она зависит от
выбора начала координат (т. е. уровня, на котором z принимается
равным 0), но если этот выбор сделан, то Е задается однозначно.
Тело обладает кинетической энергией вследствие движения, в то
время как его потенциальная энергия определяется его положе-
нием. Предположим, что во время движения в отсутствие других
сил, кроме гравитационной, тело теряет высоту, т. е. его потен-
циальная энергия уменьшается. Согласно принципу сохранения
энергии, кинетическая энергия тела (а следовательно, и его ско-
рость) должна при этом возрастать. Рассуждая аналогичным об-
разом, можно показать, что при наборе высоты скорость тела
уменьшается.
Под потенциальной энергией понимается запасенная телом
энергия, которая при высвобождении может перейти в кинетиче-
скую энергию (т. е. в движение). Энергию можно запасать, не
только поднимая тело в гравитационном поле, но и многими дру-
гими способами. С точки зрения механики сердечно-сосудистой си-
стемы наиболее важным из них является растяжение (или, напро-
тив, сжатие) упругого материала. Если прикрепить тело к пру-
жине, растянуть ее, а затем отпустить, то запасенная потенциаль-
ная энергия перейдет в кинетическую и тело придет в движение.
Точно так же, если создать давление в резиновом шарике, надув
его, а затем открыть выпускное отверстие, то воздух устремится
наружу, т. е. потенциальная энергия, запасенная в растянутой ре-
зине, перейдет в кинетическую энергию воздуха. То же самое
справедливо и для кровеносных сосудов, стенки которых растяги-
ваются при локальном повышении давления крови, а затем сжи-
маются после падения давления (гл. 12).
Упругие и гравитационные силы обладают тем свойством, что,
когда тело вынуждают двигаться против них, совершаемая работа
запасается в виде потенциальной энергии, которая впоследствии
Гл. 2. Механика частиц 37
может высвободиться в виде кинетической энергии. Поскольку в
этом случае выполняется принцип сохранения энергии, подобные
силы называют консервативными. Консервативными являются да-
леко не все силы. В частности, силы, природа которых связана
с процессами трения [подобные силе трения между двумя сколь-
зящими друг по другу твердыми телами (см. разд. 2.4) или сопро-
тивлению воздуха], обладают тем свойством, что любая работа,
совершенная против них, сопровождается потерей механической
энергии. В этом случае механическая энергия не сохраняется, а
диссипирует (рассеивается; фактически она переходит в тепло), и
поэтому подобные силы называют диссипативными. При наличии
диссипативных сил принцип сохранения механической энергии не
выполняется. Тем не менее при расчетах рассеянием энергии мож-
но пренебречь, мало теряя в точности, если удается показать, что
потери энергии за счет рассеяния малы по сравнению с ожидае-
мыми для данного случая изменениями других видов энергии, ко-
торые подчиняются закону сохранения.
В качестве примера использования энергетического подхода
рассмотрим шарик, подброшенный в воздух. Предположим, что
единственной действующей на него силой является вес (сопротив-
лением воздуха мы пренебрегаем). Если с определенного уровня,
который мы можем принять за уровень z = 0, шарик бросают
вверх с начальной скоростью V, то его скорость v в следующие
моменты будет связана с высотой подъема z относительно исход-
ного уровня в соответствии с уравнением (2.10). Значение по-
стоянной Е определяется условиями в момент бросания, когда
2 = 0 и v= V. Таким образом, E = ymV2 и, согласно уравнению
(2.10),
g2 + 4u2 = |v?. (2.11)
Заметим, что горизонтальная составляющая движения в дан-
ном случае отсутствует, поскольку в начальный момент она равна
нулю, а никакие горизонтальные силы не прикладываются. Из
уравнения (2.11) мы можем вычислить максимальную достижи-
мую шариком высоту. Так как на этой высоте (скажем, z = h) его
скорость нулевая (о = 0), должно выполняться равенство
ft = V2/2g. (2.12)
Аналогичным образом можно показать, что направленная вниз
скорость шарика в момент возвращения его в точку бросания
(г = 0) снова будет равна V- если при движении вверх вектор
скорости имел составляющие (0, 0, V), то при движении вниз —
составляющие (0,0, — И).
38 Часть I. Основы механики
Легко проверить, что уравнение (2.12) корректно с точки зре-
ния размерностей: V измеряется в м-с-1, g — в м-с-2; следова-
тельно, величина V2/2g измеряется в
( М \2 - у с2
(т) Х-5- = м>
т. е. имеет размерность длины. Единицей измерения энергии (рав-
ной половине произведения массы на квадрат скорости) является
кг-м2-с-2, что, конечно, совпадает с единицей измерения работы
(произведение силы на расстояние). Единицей измерения количе-
ства движения (произведения массы на скорость) является
кг-м-с-1.
Глава 3
ЕДИНИЦЫ ИЗМЕРЕНИЯ
Каждому, кто приступает к изучению физиологии, очень скоро
становится ясно, что в ней используются самые разные системы
единиц измерения и разнообразные термины. Например, физио-
логи, изучающие дыхание, измеряют давление в сантиметрах вод-
ного столба, а физиологи, занимающиеся сердечно-сосудистой си-
стемой,— в миллиметрах ртутного столба. По мере усложнения
различных областей физиологии к их исследованию все более ши-
роко привлекаются другие научные дисциплины. В результате
этого число применяемых единиц измерения увеличивается до та-
кой степени, что перевод из одной системы единиц измерения в
другую становится весьма трудоемким занятием и может служить
источником разного рода недоразумений и ошибок. Нередко мы
оказываемся свидетелями неправильного использования терминов,
которое приводит к путанице; например, парциальное давление
кислорода в крови в англоязычной литературе часто называют
«натяжением кислорода» *), тогда как на самом деле натяжение
означает силу, и это слово в данном контексте неуместно.
Чтобы исправить такое положение дел, которое со временем
все более усугубляется, делаются упорные попытки преобразовать
и объединить применяемые в физиологии системы наименований
и единиц измерения. Однако любые согласованные меры по наве-
дению порядка ценны лишь в том случае, когда их результаты
внутренне непротиворечивы и могут широко использоваться. А для
этого они должны быть основаны на правильном понимании мате-
матических принципов и законов физики.
Система единиц, которая принята во всем мире и применяется
сейчас почти во всех областях науки, называется Международной
Системой Единиц, или системой СИ (разд. 3.8).
Эта самосогласованная система основана на метрической си-
стеме мер и таких единицах, как килограмм массы, метр и се-
кунда. Она создает фундамент для унификации единиц, применяе-
мых в настоящее время в различных разделах физиологии. Кроме
того, перевод из ранее принятой системы СГС в СИ не составляет
особого труда.
*) В отечественной литературе используется термин «напряжение кислоро-
да». — Прим, перев.
40 Часть 1. Основы механики
3.1. Единицы измерения и размерности
И в повседневной жизни, и р научных исследованиях признано
необходимым, говоря о какой-то конкретной физической величине,
точно указывать единицы ее измерения. Так, расстояние обычно
измеряют в километрах, сантиметрах, ангстремах и т. д., а ско-
рость— в километрах в час или сантиметрах в секунду. Число, ко-
торое указывает «количество» данной величины в определенной
системе единиц измерения, называют мерой; оно обратно пропор-
ционально «размеру» используемых единиц измерения. Так, 1 ки-
лометр содержит 103 метров или 105 сантиметров.
Говоря, например, что размер листа бумаги равен 10X8 дюй-
мам, а коробка имеет размеры см, неявно пользуются
представлениями о площади и об объеме, единицы измерения ко-
торых равны соответственно единице длины в квадрате или кубе.
Размерности этих двух величин записываются в виде [L2] и [L3].
Аналогично размерность скорости равна [L] / [Т], или [LT-1], а
размерность плотности — [ML-3].
Размерности уже упомянутых физических величин или оче-
видны в силу самой природы этих величин, или вытекают из их
определения. Однако нередко связь размерности одной физической
величины с размерностью других может быть выведена только из
физического закона. Примером такого рода служит вывод размер-
ности силы нз размерностей массы, длины и времени. Согласно
второму закону Ньютона,
Сила — Масса X Ускорение,
или
F = М-а,
где а — ускорение, размерность которого есть [£Г~2]. Следова-
тельно,
[F] = [M • a]^[MLT-2].
С другой стороны, мы можем определить размерность массы как
[М] в [FL-1T2].
В конечном счете размерности всех величин в механике могут
быть выражены через [М], [L] и [Т] или через [F], [L] и [Т].
Эти альтернативные методы получения размерностных выра-
жений широко известны как «основанные на массе» и «основанные
на силе», и каждому из них соответствует своя система единиц.
3.2. Масса, длина и время как основные величины
Таким образом, размерности всех физических величин, не оп-
ределенных через температуру, могут быть представлены через
размерности основных величин — массы, длины и времени, кото-
рые обычно используют в науке в качестве основных единиц. Их
Гл. 3. Единицы измерения 41
рассматривают как независимые друг от друга и определяют в со-
ответствии с эталонами, принятыми просто по соглашению.
Например, при измерении длины пользуются метром. Раньше
эталоном метра служил хранящийся в Париже металлический
стержень; на нем были нанесены поперечные штрихи, расстояние
между которыми (при строго определенной температуре) и было
принято в качестве эталона метра. В настоящее время эталоном
является «световой метр»; он определяется как длина, равная
1 650 763,73 длины волны в вакууме оранжевой линии, излучаемой
разрядной лампой, которая заполнена газом 86Кг. Хотя этот способ
определения метра может показаться искусственным и сложным,
он обладает двумя важными преимуществами. Во-первых, новый
эталон имеет фактически такую же длину, что и старый, а, во-вто-
рых, он доступен любой солидной лаборатории, так что нет необхо-
димости ездить в Париж.
Из основных единиц измерения массы, длины и времени могут
быть получены единицы измерения других используемых в меха-
нике величин. Так, в системе СГС единицей площади является
квадратный сантиметр (см2), а в системе СИ — квадратный метр
(м2). Единицей измерения плотности в СИ становится килограмм
на кубический метр (кг-м-3).
Согласно второму закону Ньютона, единицей силы является та-
кая сила, которая при действии на единичную массу сообщает ей
единичное ускорение. Определенным таким образом единицам
силы в зависимости от того, в какой системе единиц они выведены,
присвоены разные наименования: в системе СГС — дина (1 дина =
= 1 г-см-с-2), в системе СИ — ньютон (1 ньютон = 1 кг-м-с-2).
Один ньютон равен 10Б дин. Отсюда напряжение и давление (см.
разд. 4.1), которые равны силе, приходящейся на единицу пло-
щади, в системе СИ измеряются в Н-м-2, или в кг-м-1-с-2.
Выбор массы, длины и времени в качестве основных величин
не единственная возможность. При решении некоторых частных
задач иногда оказываются более удобными другие системы неза-
висимых величин. Так, в ряде технических систем в качестве ос-
новных величин часто используют силу, длину и время. Тогда еди-
ница массы становится производной единицей. Однако в точных
науках в качестве основных величин почти повсеместно исполь-
зуются масса, длина и время; эта система удобна и хорошо обос-
нованна.
3.3. Неудобство использования силы в качестве основной величины
В системе единиц, основанной на силе, единица измерения
силы первоначально была определена как вес единичной массы.
Однако мы знаем, что вес тела представляет собой результат дей-
ствия на него силы тяжести; выше было введено понятие ускоре-
42 Часть 1 Основы механики
ния силы гравитационного притяжения, которое немного разли-
чается для разных точек Земли, а на Луне, например, значительно
меньше. Поэтому система единиц, основанная на силе, обладает
серьезным недостатком, обусловленным тем, что сила гравита-
ционного притяжения зависит от места, где проводятся измерения.
Масса тела является просто показателем количества содержа-
щегося в нем вещества. Ее можно определить, измерив сопротив-
ление тела изменению движения, с помощью второго закона Нью-
тона:
Сила == Масса X Ускорение.
Так, если сообщить телу известное ускорение, то его массу
можно вычислить, если измерить силу, которая необходима для
поддержания этого ускорения. Вместо этого можно сопоставить
массы двух тел, сравнив силы, необходимые для поддержания
одинакового ускорения у этих тел.
Понятно, что единицы, основанные на силе, в отличие от абсо-
лютных единиц измерения массы, длины и времени относятся к
единицам, зависящим от гравитационного притяжения. Причаст-
ность силы гравитации к определению понятия «вес» и небрежное
использование слов «вес» и «масса» часто приводят к несообраз-
ностям в расчетах. Одна из важных особенностей системы СИ
именно в том и состоит, что в ее основу положена масса, а не
сила, и этим преодолеваются трудности, связанные с использова-
нием веса вещества.
3.4. Энергия и теплота
В механике нам очень часто приходится иметь дело с такой
величиной, как энергия системы. Размерность энергии — [ML2T-2],
и в системе СИ энергия измеряется в единицах, названных джоу-
лями. В физиологии, как и в других областях науки, часто бывает
необходимо определить количество тепла, которое на самом деле
является одной из форм энергии, а потому имеет указанную выше
размерность и измеряется в тех же единицах. Температура, ко-
торая связана с теплом, также считается основной величиной и
имеет размерность [0], в качестве единицы измерения использует-
ся градус Кельвина.
3.5. Понятие о количестве вещества
Используя в качестве одной из характеристик тела его массу,
мы не в состоянии указать, сколько молекул оно содержит. Од-
нако при различных химических превращениях нам необходимо
как-то охарактеризовать количество молекул, с которыми мы
имеем дело. Мы хотим знать количество вещества, а не его массу.
Гл. 3. Единицы измерения 43
Это количество измеряется числом молей вещества, а моль в свою
очередь определяется как молекулярный вес, выраженный обычно
в граммах.
3.6. Размерностная однородность и совместимость единиц
Как мы уже отмечали, уравнения, имеющие физический смысл,
связывают величины одной и той же физической природы и по-
этому являются однородными с точки зрения размерностей. Это
правило известно как принцип размерностной однородности; оно
молчаливо принималось выше при выводе производных размер-
ностей и единиц измерения из основных. Именно применяя этот
принцип, мы можем обеспечить совместимость всех получаемых
производных единиц измерения.
3.7. Использование понятий объема и расхода в физиологии
В прошлом понятия объема и расхода трактовали слишком
вольно. В точных научных описаниях это недопустимо и потому
имеет смысл рассмотреть эти величины более подробно.
Объем — это некая область пространства и имеет размерность
[L8], так что в системе СИ он измеряется в кубических метрах
(м3). Рассматриваемый объем пространства может содержать в
себе то или иное количество некоторого вещества или же быть
совершенно пустым. Неправильно считать, что «объем» — это что-
то вроде количества вещества внутри него. Конечно, когда из-
вестна плотность вещества в пространстве, мы можем определить
и массу внутри объема.
Расход — это скорость перемещения данного количества веще-
ства из одной области пространства в другую. Таким образом, мы
имеем дело с передвижением данной массы за данное время, но
не с перемещением объема. Размерность расхода — [М/Т], а не
[L3/T], как часто полагают (см. гл. 4)1). В системе СИ расход
измеряется в кг-с_|.
3.8. Международная система единиц (СИ)
СИ представляет собой результат рационального выбора еди-
ниц измерения из метрической системы, поэтому некоторые ее еди-
ницы не являются новыми. Существует семь основных единиц
(табл. 3.1.) и несколько производных; отдельным из них при-
своены специальные наименования (табл. 3.2). Производные еди-
*) В отечественной литературе применяются термины «массовый расход» и
«объемный расход» Если плотность жидкости постоянна, то эти две величины
отличаются постоянным множителем Далее авторы почти всюду под расходом
крови подразумевают именно объемный расход, размерность которого [1ЛТ-1].—
Прим ред.
44 Часть I. Основы механики
ницы введены только для удобства, на самом деле все они могут
быть выражены через основные единицы.
Таблица 3.1
Основные единицы системы СИ
Величина Наименование единицы измерения Обозначение
Длина метр м
Масса килограмм кг
бремя секунда с
Термодинамическая температура градус Кельвина к
Электрический ток ампер А
Сила света кандела кд
Количество вещества моль МОЛЬ
Таблица 3.2
Распространенные производные единицы СИ
Физическая величина Единица измерения Обозначение
Сида, натяжение ньютон Н = кг • м • с-2
Работа, энергия, количество джоуль Дж = Н • м
тепла
Мощность ватт Вт = Дж • с~’
Частота герц Гц = с-1
Площадь квадратный метр м2
Объем кубический метр м*
Плотность (массовая плот- килограмм на кубический кг • М“3
ность) метр
Скорость метр в секунду м • с-*
Угловая частота радиан в секунду рад • с_|
Ускорение метр в секунду в минус вто м • с-2
Давление, напряжение рой степени ньютон на квадратный метр Н-м-2 = Па
(называется также паска- лем)
Поверхностные натяжение ньютон на метр Нм-1
Динамическая вязкость ньютон-секунда на квадрат- ный метр Н • с - м~2
Кинематическая вязкость |
Коэффициент диффузии J метр квадратный в секунду м2-с 1
Хотя система СИ — это просто развитие существующей си-
стемы метр — килограмм — секунда, она совершеннее ее, потому
что является самосогласованной. Это означает, что в результате
перемножения единичных величин получается единичная вели-
чина. Например:
1 Н X 1 м — 1 Дж,
кг X 1 м: 1с2«= IН.
Гл. 3. Единицы измерения 45
Отсутствие в этих соотношениях безразмерных числовых мно-
жителей значительно упрощает расчеты и исключает трудоемкую
процедуру применения переводных коэффициентов. В табл. 3.3
приведены коэффициенты для перевода некоторых распространен-
ных единиц измерения в соответствующие единицы СИ.
Таблица 3.3
Коэффициенты перевода некоторых распространенных
единиц измерения в единицы системы СИ
Величина Единица измерения си
Объем 1 фут3 0,02832 м3
Масса 1 фунт 0,4536 кг
Сила 1 дииа ю-в Н
Работа 1 эрг 10“7 Н-м
Давление 1 см водного столба 98,1 Н-м"2
1 мм ртутного столба 133,3 Н • м-2
Вязкость 1 пуаз 0,1 Н-с-м-2
Система СИ дает также четко определенную структуру записи
единиц, кратных основным и производным единицам (табл. 3.4).
Таблица 3.4
Множители и приставки для образования десятичных кратных
и дольных единиц
Множитель Приставка Обозначение Пример
10>2 тера т
ю9 гига г
10е мега м мегаватт (МВт)
103 кило к километр (км)
ю2 гекто г
10> дека да декаграмм (даг)
10“i деци д дециметр (дм)
Ю 3 санти С сантиметр (см)
10 . милли м миллиграмм (мг)
10 ’ микро мк микросекунда (мкс)
}о’2 иано н нанометр (нм)
10 ПИКО п пикограмм (пг)
Указанными приставками следует пользоваться с осторожностью.
Приставка всегда пишется слитно с названием дайной единицы
измерения, например меганьютон (МН), килоджоуль (кДж), мик-
росекунда (мкс). Перед данной единицей может стоять только
одна приставка, например 1000 килограммов равна 1 мегаграмму
(Мг), а не 1 кило-килограмму. Буква «м» означает основную еди-
ницу измерения «метр», а также приставку «милли», и поэтому во
46 Часть I. Основы механики
избежание недоразумений следует соблюдать определенные пра-
вила записи. Например, мН означает миллиньютон, тогда как
м-Н означает метр-ньютон, или единицу измерения работы. Для
простоты, однако, метр лучше писать на втором месте, т. е. так:
ньютон-метр (Н-м).
Следует заметить также, что когда единица, кратная основной,
возводится в степень, показатель степени относится ко всей вели-
чине в целом, а не только к основной единице; например, 1 км2
означает 1 (км)2 — 106 м2, а не 1 к (м)2 = 103 м2 ’).
*) Система СИ используется далее не везде: данные из других работ вос-
производятся без пересчета в систему СИ. Читатель должен помнить об этом
во избежание недоразумений. — Прим. ред.
Глава 4
ОСНОВЫ МЕХАНИКИ ЖИДКОСТЕЙ
В гл. 1 мы установили, в каких случаях реальное вещество,
в частности жидкость, можно рассматривать как сплошную среду:
для этого необходимо, чтобы расстояния, на которых изменяются
усредненные свойства среды (например, плотность), были гораздо
больше, чем межмолекулярные расстояния. В таком случае среду
можно мысленно разбить на ряд элементарных объемов и к каж-
дому из них применить законы механики частиц. Эти законы мы
также изложили. Однако прежде, чем использовать их, необхо-
димо установить, какие силы действуют на каждый элемент. Как
и в случае тела, скользящего по столу (рис. 2.7), на мысленно вы-
деленный элемент жидкости в общем случае действуют силы двух
видов — дальнодействующие и близкодействующие.
Влиянию сил, действующих на больших расстояниях (объем-
ных и массовых сил}, подвергаются все элементы жидкости; двумя
наиболее распространенными из них являются силы гравитацион-
ного и электромагнитного происхождения. Электромагнитная сила,
действующая на элемент, зависит от таких величин, как, напри-
мер, его электрический заряд, а гравитационная сила, т. е. вес эле-
мента, зависит только от его массы. Дальше мы будем рассмат-
ривать лишь один вид массовых сил — гравитационную силу. Если
элемент жидкости Р, находящийся в момент времени t в точке х,
занимает объем V и если жидкость в окрестности точки х имеет
плотность р *), то гравитационная сила, действующая на указан-
ный элемент, будет равна pVg.
4.1. Напряжение
Близкодействующие силы действуют иа элемент Р со стороны
только соприкасающихся с ним элементов. Эти силы склады-
ваются из всех межмолекулярных сил, с которыми молекулы, при-
легающие к поверхности элемента снаружи, действуют на моле-
кулы внутри элемента, также прилегающие к поверхности. Рас-
смотрим небольшую часть поверхности элемента (рис. 4.1, Л) и
) Плотность равна массе (кг), приходящейся на единицу объема (м3), и
измеряется в кг-м~3; для обозначения плотности принято использовать грече-
скую букву р. г
48 Часть I. Основы механики
Рис. 4.1. A. S представляет собой часть поверхности элемента жидкости, мыс-
ленно выделенного внутри среды. Сила F есть сумма всех сил, с которыми мо-
лекулы, находящиеся со стороны I, действуют на молекулы, находящиеся со сто-
роны II (линии действия этих сил пересекают поверхность S). Сила F имеет
тангенциальную (Ft) и нормальную (Fn) составляющие. Если площадь S равна
А, то напряжение на этой поверхности равно F/Л. Б. Вид сбоку на часть поверх-
ности S. Напряжение F'M', действующее на элемент поверхности S' (площадью
А'), параллельный поверхности S, но смещенный относительно нее, в общем слу-
чае отлично от напряжения F/А Точно так же отлично от F/А и напряжение
F"/A", действующее на элемент поверхности S" (площадью А"), находящийся
там же, где и S, но иначе ориентированный.
сложим все силы, с которыми молекулы, находящиеся со стороны
I, действуют на молекулы, находящиеся со стороны II, причем
такие силы, что линии нх действия пересекают рассматриваемую
часть поверхности. В результате мы получим некоторую силу F
(векторная величина, характеризующаяся определенной абсолют-
ной величиной и направлением). Согласно третьему закону Нью-
тона, сила, с которой молекулы со стороны II действуют на мо-
лекулы, расположенные со стороны I, равна —F. Абсолютная ве-
личина F пропорциональна площади А рассматриваемой части
поверхности; величиной же, не зависящей от А, является сила
F/А, которая приходится на единичную площадь, Эту силу назы*
Гл. 4. Основы механики жидкостей 49
ваюг напряжением * *)• Обычно напряжение зависит как от поло-
жения, так и от ориентации выделенной части поверхности2).
Рассмотрим, например, поверхность S' площадью А', параллель-
ную поверхности S, но смещенную относительно нее (рис. 4.1,5).
Напряжение F'/A', характерное для поверхности S', в общем слу-
чае отлично от напряжения F/А. Более того, напряжение, кото-
рое действует на поверхности S" (площадью А") и приложено в
той же точке, что и на поверхности S, но поверхность S" имеет
другую ориентацию (штриховая линия на рис. 4.1,5), также в об-
щем случае отлично от F/А. Легко видеть, что, как и сила, с ко-
торой стол действует на перемещающееся по нему тело (рис. 2.7.),
сила F (и напряжение), вообще говоря, имеет две составляющие —
нормальную Fn и тангенциальную Ft (см. рис. 4.1,Л).
Для большинства течений, которые нам предстоит рассмотреть,
существенный вклад в нормальную составляющую напряжения
вносит давление-, оно одинаково во всех направлениях и не зави-
сит от ориентации выбранного элемента поверхности. Тангенциаль-
ное напряжение во многом аналогично силе трения Т (рис. 2.7).
Оно противодействует относительному перемещению соседних
слоев жидкости. Поэтому, когда жидкость течет над плоской по-
верхностью (рис. 4.4) и ее более удаленные от стенки слои дви-
жутся быстрее, чем близлежащие, тангенциальная составляющая
напряжения, действующая между соседними слоями, стремится
замедлить движение более быстрых из них и ускорить движение
более медленных. Тангенциальное напряжение обязано своим про-
исхождением наличию у Жидкостей свойства, которое Ньютон на-
звал «дефектом скольжения»; теперь это свойство называют вяз-
костью.
4.2. Гидростатическое давление
Выясним сначала, какие напряжения существуют в совершенно
неподвижной жидкости, например в покоящейся воде, налитой в
неподвижный сосуд. Предположим, что мы опускаем в эту жид-
кость приборы, способные измерять напряжения, которые дей-
ствуют на небольшой элемент поверхности. Приборы покажут, что
тангенциальные напряжения в среде отсутствуют и что нормаль-
ные напряжения в данной точке не зависят от ориентации эле-
мента поверхности. Другими словами, в покоящейся жидкости
напряжения сводятся к одному только давлению. Мы увидим
также, что при любом перемещении точки измерения в горизон-
*) Подобная «молекулярная» интерпретация понятия напряжения является
лишь удобным поясняющим соображением, ио отнюдь не строгим физическим
рассуждением. — Прим. ред.
*) Через данную точку можно провести множество по-разному ориентиро-
ванных площадок, каждой из которых соответствует свое напряжение. Однако,
зцая напряжения, действующие на каких-либо трех площадках, можно вычис-
лить напряжения на любой другой площадке. — Прим, ряд.
50 Часть I. Основы механики
тальком направлении давление не меняется; оно зависит лишь от
расположения этой точки относительно свободной поверхности
жидкости в сосуде. Разность между давлением, измеренным на
глубине z, и давлением у поверхности прямо пропорциональна z.
Этот результат можно получить теоретически из второго за-
кона Ньютона [уравнение (2.6)]. Рассмотрим условия равновесия
(баланса) сил, действующих на небольшой кубический элемент
среды (рис. 4.2, Л). Так как внешняя массовая сила строго верти-
Рис. 4.2. А. Малый элемент жидкости в виде параллелепипеда, имеющего вы-
соту г' и площадь горизонтальных граней А, находится в равновесии. Силы дав-
ления на его боковые грани должны уравновешивать друг друга, и поэтому дав-
ление в горизонтальной плоскости не должно меняться. Вертикальные силы,
действующие на этот элемент (они показаны стрелками), также должны быть
уравновешены, и это требование приводит к уравнению (4.1). Б. Разность дав-
лений р\ — р2 между двумя изображенными областями измеряется по разности
уровней жидкости плотностью р в двух ветвях U-образной трубки: щ — pz—gph.
калька, горизонтальные силы давления должны быть взаимно
уравновешены, что в свою очередь означает постоянство давле-
ния вдоль данной горизонтальной плоскости, которое мы и наблю-
дали. Однако давление должно меняться с высотой (по коорди-
нате z). Действительно, движение жидкости отсутствует, поэтому
вес элемента должен быть уравновешен так, что направленная
вверх сила давления, действующая на нижнюю грань кубика,
должна быть больше, чем направленная вниз аналогичная сила,
действующая на верхнюю грань. Пусть А — площадь горизонталь-
ного поперечного сечения элемента, a pi и р2— давление соответ-
ственно на нижней (1) и верхней (2) гранях. Тогда действующая
вверх результирующая сила давления равна (pi — р2)А Она
должна быть равна силе веса элемента, которая направлена вниз
и равна произведению плотности жидкости р на объем элемента
Az' (где z' — высота элемента) и на g.
Следовательно, уравнение баланса сил имеет вид
(Pi — Pz) А = gpAz',
цлц
pl — p2^gpz'. (4.1)
Гл. 4. Основы механики жидкостей 61
Таким образом, с увеличением высоты на величину z' давление
хменьшается на величину gpz'. Если мы примем, что на поверх-
ности сосуда давление равно атмосферному (рл) и выберем уро-
вень z = 0 на этой поверхности (при этом z внутри жидкости от-
рицательно), то давление в любой точке на уровне z определится
выражением
Р = РА~ ^Pz- (4.2)
По мере увеличения глубины z становится все «более отрица-
1ельным» и давление растет. Давление, определяемое уравнением
(4.2), называют гидростатическим, поскольку оно вычислено для
покоящейся жидкости. Легко видеть, что единицы измерения дав-
ления (сила на единичную площадь, или кг-м_1-с~2) идентичны
единицам измерения величины gpz. Если плотность среды ме-
няется с z, как это происходит в море, где вода по мере увели-
чения глубины становится более холодной и соленой (и, следова-
тельно, более плотной), или в земной атмосфере, которая с уве-
личением высоты становится более разреженной, то уравнение
(4.2) перестает выполняться, но уравнение (4.1) остается в силе.
Для того чтобы в этом случае вычислить давление на некоторой
глубине при условии, что оно известно на определенном уровне,
необходимо мысленно разбить среду на горизонтальные слои,
внутри каждого из которых величина р практически постоянна, и
просуммировать величины gpz' (z'— толщина слоя) для всех
слоев *).
Уравнение (4.2) служит основой наиболее широко применяе-
мого метода измерения давления. Если два сосуда (наполненные,
скажем, газом), давления в которых различны, соединить с двумя
ветвями (7-образной трубки, заполненной жидкостью известной
плотности (рис. 4.2,Б), то уровни жидкости в этих ветвях будут
различаться на высоту h. Тогда из уравнения (4.2) мы прямо по-
1учим, что разность давлений в этих сосудах равна gph, где р —
плотность жидкости.
Для измерения давления широко используют две жидкости —
воду (плотность 1000 кг-м-3) и ртуть (плотность 13600 кг-м~3).
В результате давление обычно измеряют в таких единицах, как
см вод. ст. или мм рт. ст., имея в виду соответствующую высоту
водного или ртутного столба. Так как применение разных единиц
чрезвычайно затрудняет сравнение различных давлений, мы ис-
*) Если разделить обе части уравнения (4.1) на z' и перейти к пределу при
«'-►О, то мы получим уравнение
в котором левая часть представляет собой скорость изменения давления с высо-
той. Сравните эту запись с символическим обозначением «скорости изменения во
времени», определенном в уравнении (2.1).
52 Часть 1. Основы механики
пользуем международную единицу — ньютон на квадратный метр,
или кг-м-1 -с-2. Соотношения между ней и другими единицами
1 см вод. ст. = 98,1 Н • м~2,
I мм рт. ст. = 133,3 Н • м-2 (см. табл. 3.3).
В гравитационном поле Земли помещенное в покоящуюся жид-
кость тело испытывает по всей своей поверхности направленные
внутрь силы давления. Они возрастают сверху вниз вдоль поверх-
ности тела, так как давление растет с глубиной, в результате чего
возникает направленная вверх сила, названная выталкивающей
силой. Если тело представляет собой цилиндр с вертикальными
стенками и горизонтальными верхней и нижней поверхностями с
одинаковой площадью А, то выталкивающая сила равна произве-
дению разности давлений снизу и сверху на площадь А. Пусть
высота цилиндра равна h; тогда мы получаем, что
Выталкивающая сила — gphA. (4.3)
Но произведение hA равно объему цилиндра, a phA — массе
вытесненной им жидкости, поэтому правая часть уравнения (4.3)
равна весу вытесненной жидкости. Можно показать, что правило
«выталкивающая сила равна весу вытесненной жидкости» спра-
ведливо для тел какой угодно формы. Впервые оно было сформу-
лировано древнегреческим ученым Архимедом и известно под на-
званием закона Архимеда. Если тело состоит из однородного ма-
териала плотностью р', то суммарная действующая на него сила,
обусловленная силой гравитационного притяжения (вес + вытал-
кивающая сила), равна произведению объема тела на g и на раз-
ность плотностей (р — р'). Она направлена вниз, если р < р', и
вверх, если р > р'. Только при р = р' результирующая вертикаль-
ная сила равна нулю и тело остается в покое.
4.3. Напряжение в движущейся жидкости; вязкость
Обратим теперь внимание на напряжения в движущихся жид-
костях. Для этой цели удобно рассмотреть силы, действующие на
небольшой кубический элемент жидкости, и предположить, что су-
ществует такое направление, в котором компоненты всех дей-
ствующих на элемент напряжений взаимно уравновешены. В этом
случае все интересующие нас силы лежат в плоскости, перпенди-
кулярной этому направлению (плоскость рис. 4.3), и картина ста-
новится двумерной. Пусть элемент находится в покое и массовые
силы, подобные силе тяжести, отсутствуют. Ввиду того что дав-
ление не зависит от ориентации, а в данном случае — и от рас-
положения поверхности, на которую оно действует, направленные
внутрь напряжения на каждой грани кубика будут одинаковы
(рис. 4.3,А). Эти напряжения взаимно уравновешены и не дефор-
Гл. 4. Основы механики жидкостей 63
мируют кубический элемент никаким иным способом, кроме все-
стороннего сжатия, которое, однако, произойдет только в том слу-
чае, если жидкость сжимаема. Мы же в дальнейшем будем счи-
тать ее практически несжимаемой (относительно условий, при ко-
торых такое предположение справедливо, см. разд. 4.6). Давле-
ние, приложенное равномерно по поверхности небольшого эле-
мента жидкости, не сможет ни изменить его форму, ни препятство-
вать этим изменениям. Однако если нормальные напряжения на
гранях куба не равны, то они несомненно приведут к деформа-
циям.
Рис. 4.3. Двумерные схемы деформаций, которые испытывает элементарный кубик
жидкости под действием напряжений, показанных стрелками. А. Когда действует
только давление, деформации не возникают. Б. Равные по величине и противо-
положно направленные растягивающие напряжения на гранях 1 н 2 совместно
с равными и противоположно направленными сжимающими напряжениями на
гранях 3 н 4 вызывают деформацию, изображенную справа. В. Если кубик, пред-
ставленный на схеме Б, повернуть на 45°, то система напряжений преобразуется
в постоянное во всех точках давление, наложенное на тангенциальные напряже-
ния, направления которых указаны на рисунке; такая система напряжений при-
водит к изображенной справа деформации.
Допустим теперь, что силы, приложенные к одной паре проти-
волежащих граней кубика (грани 1 и 2 на рис. 4.3,5), равны и
направлены наружу (т. е. напряжения растягивающие), а силы,
приложенные к двум другим противолежащим граням (грани 3
и 4), равны им по величине, но направлены внутрь (т. е. напря-
жения сжимающие). Так как силы на противоположных гранях
равны, элемент в целом не приобретает ускорения и его центр
масс остается неподвижным. Тем не менее система напряжения
подобного рода будет деформировать кубик таким образом, как
это показано на рис. 4.3, Б. Такие же деформации будут иметь
место и в том случае, когда напряжения для всех плоскостей на-
правлены внутрь, но на гранях 1 и 2 они меньше, чем на гранях 3
64 Часть 1. Основы механики
и 4. Картина качественно не изменится, если систему, подобную
изображенной на рис. 4.3, Б, дополнительно подвергнуть всесто-
роннему сжатию. Если мысленно выделить кубик, наклоненный на
45° (см. рис. 4.3, В), то он будет деформироваться так, как это
показано на рисунке, а напряжения будут состоять из системы
равных между собой тангенциальных напряжений на гранях, на
которую наложено всестороннее давление. Таким образом, как
неуравновешенные нормальные напряжения, так и любые танген-
циальные напряжения на гранях элемента жидкости приводят к
г
/xdu
dz
Рис. 4.4. Жидкость течет в направ-
лении х над плоской стенкой
(z = 0), причем скорость и прямо
пропорциональна расстоянию г от
стенки (т. е. градиент скорости
du/dz постоянен). На верхнюю по-
верхность Л] элемента среды дейст-
вует тангенциальное напряжение
ydu/dz, направленное вперед по по-
току; иижняя поверхность А2 испыты-
вает такое же по величине танген-
циальное напряжение, направленное
назад. Напряжение, действующее на
стенку (оно направлено вперед),
имеет ту же величину.
его деформации и, следовательно, к движению. В общем случае
элемент будет двигаться и вращаться как целое и деформиро-
ваться; однако ясно, что деформации могут быть связаны только
с системой напряжений, отличных от давления.
Как уже отмечалось, все напряжения, отличные от давления,
обязаны своим происхождением вязкости Мы видели (разд. 2.4),
что при перемещении соприкасающихся твердых поверхностей
друг относительно друга возникает противодействующая движе-
нию сила трения. Аналогично при движении жидкости, сопровож-
дающемся местной деформацией ее элементарных объемов, воз-
никают вязкие напряжения, которые стремятся воспрепятствовать
деформации. Абсолютная величина вязких напряжений зависит
от скорости деформации. Например, тело, перемещающееся с
большой скоростью в жидкости, вызывает более быструю дефор-
мацию ее элементов, чем тело, движущееся медленно. Поэтому
и возникающие вязкие напряжения в первом случае больше, и та-
кое тело испытывает большее сопротивление. Почти во всех обыч-
ных жидкостях напряжения прямо пропорциональны скорости де-
формации. Полное математическое описание подобной взаимо-
связи для любых течений было бы здесь неуместно, но суще-
ствует один тип течения, представленный на рис. 4.4, для которого
эта зависимость чрезвычайно проста. При таком течении все эле-
менты жидкости движутся в одном направлении (параллельно
оси х) со скоростью и, пропорциональной координате z (ось г
Гл. 4. Основы механики жидкостей 55
перпендикулярна направлению движения среды). В этом случае
вязкое напряжение на поверхности элемента, лежащей в плоско-
сти с данным z (поверхности Л1 или А2 на рис. 4.4), является
тангенциальным, направлено вдоль оси х и абсолютная величина
его пропорциональна градиенту скорости (скорости изменения и
вдоль оси z) в данной точке. Другими словами, напряжение на
поверхности Ль направленное вдоль оси х, равно
5 = (4.4)
где р. — константа, называемая коэффициентом вязкости (или
часто просто вязкостью) данной жидкости. Напряжение на поверх-
ности А2 направлено в противоположную сторону и такое же по
абсолютной величине. Поскольку напряжение равно силе на еди-
ницу поверхности, а градиент скорости — это величина, обратная
времени и измеряемая в с~*, то из уравнения (4.4) следует, что
вязкость р измеряется в кг-м-1-с-1. Тангенциальное напряжение
подобного типа обычно называют сдвиговым напряжением, или
напряжением сдвига, так как оно связано с движением, при ко-
тором соседние слои жидкости скользят (сдвигаются) один отно-
сительно другого. Градиент скорости в жидкости иногда называют
скоростью сдвига ’).
Жидкости, в которых напряжение всюду прямо пропорцио-
нально местной скорости сдвига, т. е. жидкости, обладающие
постоянной вязкостью, называются ньютоновскими. Сюда могут
быть отнесены многие обычные среды, как то: воздух, вода, гли-
церин, ртуть и т. д. Однако нередко встречаются жидкости, в ча-
стности такие, в которых растворены крупные молекулы (напри-
мер, белки), с вязкостью, зависящей от скорости сдвига. Имеются
также вещества (подобные крему), которые начинают течь, толь-
ко если напряжение сдвига становится выше определенной крити-
ческой величины (предельного напряжения сдвига), а при мень-
ших напряжениях ведут себя как упругие тела. Есть и вещества
(подобные простой оконной замазке), которые при длительно под-
держиваемых невысоких тангенциальных напряжениях обладают
текучестью (примером может служить деформация замазки, ле-
*) Рассматривая элемент жидкости (рнс. 4.4) в два близких момента вре-
мени t, t', нетрудно подсчитать, что за малый интервал Д/ — Г — t первоначаль-
но вертикальная грань наклонится на малый угол в« (и2—где
и2, «1 — значения скорости на поверхностях А2, Л], Az — расстояние между ними.
Отсюда, переходя к пределу при Д/ н Дг, стремящихся к нулю, видим, что ско-
рость сдвиговой деформации (т. е. скорость изменения 6 во времени) в точно-
сти равна (для данного течения!) градиенту скорости du/dz. Это замечание свя-
зывает формулу (4.4) с ранее высказанным утверждением о пропорциональности
вязких напряжений скоростям сдвига. В общем случае градиент скорости и ско-
рость сдвига — разные величины, но для некоторых течений их численные значе-
ния совпадают. Применительно к формулам типа (4.4) необходимо пользоваться
для du/dz термином «скорости сдвига». — Прим. ред.
66 Часть 1. Основы механики
жащей на ровной поверхности, под действием собственного веса)
и вместе с тем при кратковременных больших напряжениях прояв-
ляют свойства упругого тела (шарик из такого материала может
подпрыгивать и даже разбиваться при ударе молотком). Все эти
жидкости являются неньютоновскими-, в гл. 10 мы узнаем, что в
определенном диапазоне напряжений и скоростей сдвига кровь
также является неньютоновской жидкостью: она обладает пре-
дельным напряжением сдвига, а ее вязкость зависит от скорости
сдвига. В крупных артериях следствия этих факторов столь незна-
чительны, что кровь можно считать ньютоновской жидкостью и
описание ее движения может быть упрощено; при описании дви-
жения крови в микрососудах подобно упрощение невозможно.
Вязкость не только противодействует относительному движе-
нию соседних слоев жидкости, но и предотвращает ее скольже-
ние по поверхности любого твердого тела. Элемент жидкости, со-
прикасающийся с поверхностью твердого тела, «прилипает» к ней
и приобретает, таким образом, такую же скорость, что и поверх-
ность тела. Это условие, называемое условием прилипания, яв-
ляется эмпирическим законом. Строгого доказательства этого по-
ложения, которое основывалось бы на физических законах меж-
молекулярного взаимодействия, не существует, но, с другой сто-
роны, до настоящего времени не поставлен ни один опыт, кото-
рый опроверг бы его. Условие прилипания на границе является
ограничением, которому должны удовлетворять все теоретические
суждения о картинах течения, и, как мы узнаем в дальнейшем, это
условие имеет важные следствия для движения крови. Оно под-
разумевает, в частности, что жидкость, движущаяся вдоль непод-
вижной границы твердого тела, всегда создает на ней напряже-
ние сдвига (как показано на рис. 4.4). Это происходит потому, что
жидкость вблизи границы движется быстрее, чем непосредственно
на самой границе. Таким образом, вплоть до границы стенки ско-
рость сдвига не нулевая, и, как следует из уравнения (4.1), на-
пряжение сдвига на границе также отлично от нуля.
4.4. Уравнение движения жидкости
Обратимся теперь к приложению законов Ньютона к движу-
щимся элементам жидкости. Согласно второму закону, произве-
дение ускорения элемента жидкости на его массу равно сумме
действующих на него напряжений и массовых сил. Пусть, напри-
мер, элемент жидкости движется в направлении х в прямолиней-
ном потоке, скорость в котором изменяется с координатой г (ось?
направлена вертикально вверх), но не зависит от х, у и от вре-
мени (см. рис. 4.5). Сила тяжести направлена вниз (в сторону
отрицательных z), и поэтому давление, как и скорость, вдоль
третьей оси (у) не меняется. Из условия баланса сил в верти-
кальном направлении следует, что при данном х давление изме-
Гл. 4. Основы механики жидкостей 57
няется с высотой таким же образом, как и в покоящейся жидко-
сти, и величина его задается уравнением (4.1). Рассмотрим теперь
баланс горизонтальных сил (в расчете на единицу длины в на-
правлении у). В отсутствие движения вязкие напряжения были бы
равны нулю и давление на правой и левой гранях элемента (при
х = Хо и х = х0 4- х' соответственно) было бы одинаковым. Но,
как только появляется движение, возникают вязкие силы; в при-
веденном примере они стремятся замедлить элемент, так как на-
правленное против движения напряжение сдвига на нижней по-
верхности элемента (при z — Zo) больше (потому что скорость
Рнс. 4.5. Силы, действующие на прямоугольный элемент жидкости в потоке,
скорость которого u(z) имеет составляющую только в направлении х и меняется
только с координатой z. Из условия баланса вертикальных сил следует, что дав-
ление при определенном значении х меняется с высотой (z) так же, как и в по-
коящейся среде [уравнение (4.1.)]. Направленное вперед напряжение сдвига
(pdu/dz) на верхней поверхности (z = z0 + г') меньше по абсолютной величине,
чем направленное назад напряжение сдвига на нижней поверхности (z = z0),
поскольку скорость и меняется с высотой таким образом, как это показано на
рисунке. Поэтому давление на левой грани (х = Хо) должно быть больше, чем
на правой (х = хо + х'), т.е. на жидкость действует отрицательный (способ-
ствующий течению) градиент давления.
сдвига du/dz больше), чем направленное по ходу движения на-
пряжение сдвига на верхней поверхности (при z = Zo-]-2<). При
равенстве давлений на вертикальных гранях элемент среды в
этих условиях должен был бы замедлиться. Движение без замед-
ления возможно только в том случае, если давление, действую-
щее на левую грань, больше, чем на правую, т. е.
р(х0) — р(х0 + х')>0.
Другими словами, в жидкости существует отрицательный гра-
диент давления в направлении х1)- Отметим, что отрицательный
*) Градиент давления здесь определен как наклон кривой зависимости р
от х и, таким образом, равен пределу отношения {р(х0 + х') — р(х0))/х' при х',
стремящемся к нулю. Другими словами, он равев dp/dx (см. разд. 2.2, где об-
суждался предельный переход).
58 Часть J. Основы механики
градиент давления способствует движению, тогда как положи-
тельный противодействовал бы ему. Если нет внешних массовых
сил, обеспечивающих движение, то для ускорения жидкости или
поддержания ее равномерного движения всегда требуется подоб-
ный «способствующий» градиент давления, который преодолевает
действие вязкости. Например, в горизонтальной трубе составляю-
щая внешней массовой силы (веса) в направлении движения жид-
кости равна нулю, и потому равномерный поток может существо-
вать только в том случае, когда давление на одном конце трубки
больше, чем на другом.
Для любого течения уравнение движения каждого элемента
жидкости (т. е. второй закон Ньютона) может быть представлено
в следующей форме:
Масса X Ускорение = Массовая сила + Сила, обусловленная
градиентом давления 4- Вязкая сила. (4.5)
Так как связь между вязкими силами и местными изменениями
скорости жидкости известна (напряжения сдвига пропорциональ-
ны скоростям сдвига), это уравнение связывает градиент давле-
ния и массовую силу в любой точке с местной скоростью среды и
скоростями ее изменения как во времени (ускорение), так и в про-
странстве (скорость сдвига). В теоретической гидромеханике ста-
вится задача определить движение каждого элемента жидкости
исходя из этого уравнения и ряда дополнительных ограничений,
характерных для данного потока, — таких, как условие прилипа-
ния на твердой стенке или постоянство давления на свободной по-
верхности жидкости. Обычно такая задача может быть решена
только приближенно, но в некотором сравнительно небольшом
числе случаев удается найти и точное теоретическое решение. Все
члены в правой части уравнения (4.5) представляют силы, тогда
как в левой части стоит произведение массы на ускорение, пред-
ставляющее инерцию элемента среды. Если этот член перенести
с противоположным знаком в другую часть уравнения, то его
можно будет рассматривать как еще одну силу. И действительно,
в механике жидкостей этот член принято называть «силой инер-
ции». Однако, как и в случае «центробежной силы», важно всегда
помнить, что в действительности это не реальная сила, а произве-
дение массы на ускорение.
4.5. Конвективное и местное ускорение
Чтобы измерить составляющие скорости или давление в дви-
жущейся жидкости, погружают в определенную точку потока из-
мерительный прибор, фиксируют его положение и определяют, как
в ходе эксперимента меняются во времени показания прибора. Эту
процедуру можно повторить, помещая прибор в различные точки
Гл. 4. Основы механики жидкостей 59
и наблюдая изменение скорости и давление при переходе из од-
ной области в другую, условия в которых могут различаться. Этот
метод отличается от подхода, используемого в механике изолиро-
ванных частиц (гл. 2), когда следят за состоянием отдельных час-
тиц по мере их движения от точки к точке. Если такой подход
применить для описания элементов жидкости, которые непрерывно
деформируются, то связь между напряжением и скоростью сдвига
станет чрезвычайно сложной. Поэтому при теоретических исследо-
ваниях гораздо проще использовать подход, изложенный вначале;
к тому же его легче осуществить экспериментально. Этому под-
ходу, однако, присущ один недостаток: в этом случае гораздо
труднее получить выражение для ускорения элемента среды, так
как быстрота изменения скорости в какой-либо точке отражает
различие в скоростях двух разных элементов, занимающих ее в
последовательные моменты времени. При этом ускорение отдель-
ного элемента среды должно зависеть не только от того, как ме-
няется во времени скорость в данной точке, но и от того, как ско-
рость меняется от точки к точке, поскольку элемент последова-
тельно занимает разные точки. Частицы среды могут иметь уско-
рение, даже когда скорость в каждой точке не меняется во вре-
мени (тогда поток называется стационарным), так как при этом
частицы могут двигаться между точками, скорости в которых раз-
личаются. Хорошим примером такого рода может служить поток
воды, перетекающий через плотину. Элементы жидкости, находя-
щиеся выше плотины, движутся чрезвычайно медленно, по мере
приближения к плотине ускоряются, а над ней проносятся очень
быстро. Тем не менее поток в этом случае стационарный, так как
скорость течения в каждой точке постоянна. Подобный вклад в
ускорение частицы называется конвективным ускорением-, час-
тица испытывает его, когда перемещается с потоком из точки в
точку. Вклад же в ускорение от изменения во времени скорости
в данной точке называется местным ускорением. Примером неста-
ционарного потока, в котором главную роль играют местные уско-
рения, является течение крови в крупных артериях (гл. 12). Отме-
тим, что именно наличие члена, описывающего конвективное уско-
рение, чрезвычайно затрудняет решение в общем виде уравнения
движения жидкости [уравнение (4.5)].
4.6. Сохранение массы
Прежде чем переходить к подробному обсуждению свойств по-
тока жидкости в конкретных обстоятельствах и, в частности, по-
тока крови в сердечно-сосудистой системе, необходимо рассмот-
реть еще два общих принципа механики сплошных сред. Первый
из них гласит, что масса не может исчезать или появляться (ис-
ключение составляют только ядерные реакции, в ходе которых
60 Часть I. Основы механики
масса может переходить в энергию, и наоборот1)) и называется
принципом сохранения массы. Этот очевидный физический прин-
цип накладывает строгие ограничения на виды движения, которые
могут осуществляться в данном потоке. Он требует, например,
чтобы масса жидкости, вытекающей из системы трубок, была рав-
на массе втекающей жидкости, если только не увеличиваются плот-
ность жидкости в трубках (тогда тот же объем может занимать
большая масса) или объем трубок (из-за растяжения их стенок).
Все реальные жидкие среды до некоторой степени сжимаемы (т. е.
плотность их может меняться), но такие жидкости, как вода и
Рис. 4.6. Трубка, по которой течет несжимаемая жидкость, имеет перемен-
ную площадь поперечного сечення А. Вследствие этого продольная скорость и
(усредненная по поперечному сечению) также изменяется по длине. В жесткой
трубке в любой момент времени расход Q(= иА) в каждом сеченнн одинаков,
даже если величина Q меняется во времени. Аналитически это правило записы-
вается в виде UiAi = U2A2.B растяжимой трубке А может меняться во времени,
и в каждый момент величина (ujAi — и2А2) равна скорости увеличения объема
заштрихованной области.
кровь, сжимаемы в гораздо меньшей степени, чем газы, и для мно-
гих случаев с высокой точностью можно считать их несжимае-
мыми. Это приближение становится несправедливым, когда ско-
рость потока жидкости приближается к скорости звука в ней. По-
следняя равна скорости распространения небольших изменений
давления в жидкости (но не скорости распространения возмуще-
ния по стенкам сосуда, содержащего жидкость; распространение
волн в стенках, как показано в гл. 12, имеет большое значение для
кровообращения). Поскольку максимальные наблюдаемые ско-
рости движения крови в сердечно-сосудистой системе составляют
менее 1% от скорости звука в крови, мы можем далее считать
кровь несжимаемой жидкостью и пренебречь всеми явлениями,
связанными с ее сжимаемостью. Поэтому когда кровь течет в си-
стеме жестких трубок, то, сколько ее втекает, столько должно и
') Принцип сохранения массы, как, впрочем, и многие другие утверждения
физики в их привычной формулировке, справедливы лишь для движений со ско-
ростями, много меньшими скорости света. — Прим. ред.
Гл. 4. Основы механики жидкостей 61
вытекать; в частности, если по ходу некоторой жесткой трубки
изменяется площадь ее поперечного сечения А (рис. 4.6), то и
скорость потока по длине трубки будет неодинаковой. Однако про-
изведение площади А на среднюю по данному поперечному сече-
нию продольную скорость и является величиной постоянной (не-
зависимо от сечения). Эта постоянная величина называется объ-
емным расходом Q через трубку: иА — Q.
Таким образом, когда широкая трубка сужается, скорость дви-
жения жидкости возрастает; когда же трубка становится шире,
течение замедляется. Если трубка не жесткая, то объемный рас-
ход не обязан быть одинаков во всех сечениях, поскольку в этом
случае может меняться во времени площадь поперечного сечения
трубки. Если, например, площадь поперечного участка сегмента
трубки в некоторый момент времени возрастает, приток жидкости
в него превышает ее отток, так что величины иА на концах уча-
стка должны различаться (рис. 4.6). Объемный расход будет оди-
наков во всех точках растяжимой трубки только в случае ста-
ционарного потока.
4.7. Теорема Бернулли
Второй общий принцип механики сплошных сред касается со-
хранения энергии. В отсутствие ядерных реакций полная энергия
системы складывается из механической энергии (кинетической и
потенциальной; см. разд. 2.6) и тепловой (тепла). Согласно за-
конам термодинамики, скорость изменения полной энергии си-
стемы равна скорости, с которой совершают работу приложенные
к системе силы. Когда внешние силы отсутствуют, полная энергия
остается постоянной, т. е. сохраняется, хотя в принципе при этом
возможно превращение механической энергии в тепло (и наобо-
рот), например, благодаря действию вязких сил (разд. 4.3). Прин-
цип сохранения энергии можно использовать для получения важ-
ного результата, касающегося стационарного потока жидкости в
условиях, когда роль вязкости в каждой точке чрезвычайно мала
и ею можно пренебречь. Эти условия обычно выполняются вдали
от границ твердых тел. Отсутствие вязкости означает, что механи-
ческая энергия не переходит в тепло и сохраняется неизменной,
пока нет внешних сил.
Здесь удобно ввести понятие линии тока; это воображаемая
кривая, проведенная в данный момент в пространстве так, что в
каждой своей точке она параллельна направлению движения (век-
тору скорости) среды в этой точке. При стационарном потоке,
когда скорость в каждой точке не меняется во времени, линии
тока совпадают с траекториями движения отдельных частиц среды.
При нестационарном же потоке, хотя частица среды и движется
всегда в направлении локального вектора скорости, в следующий
момент времени это направление может измениться и линия тока,
62 Часть I. Основы механики
на которой теперь окажется частица, может не проходить через
точку, соответствующую ее положению в предыдущий момент.
Представим себе узкую трубку, выделенную в стационарном по-
токе жидкости таким образом, что границы трубки образованы це-
ликом из линий тока. В таком случае у элементов среды не будет
составляющей скорости, пересекающей эту границу, и среда, теку-
щая в трубке, будет всегда оставаться в ней (рис. 4.7). Такая
«трубка тока» напоминает жесткую трубку с переменной пло-
щадью поперечного сечения, конечно, с тем отличием, что ее
«стенки» не являются поверхностью твердого тела, и поэтому нет
Рис 4.7. Схема трубки тока при стационарном течении жидкости. Боковые
«границы» трубки образованы линиями тока, и жидкость не пересекает их Пло-
щадь поперечного сечения А, уровень г, скорость среды и и давление р на кон-
цах трубки различны. Принцип сохранения энергии указывает, что в тех случаях,
когда влияние вязкости несущественно, величина Н [определяемая уравнением
(4.6)] в трубке тока остается постоянной (теорема Бернулли).
необходимости соблюдать условия прилипания на границе. Пред-
положим теперь, что концы такой трубки конечной длины распо-
ложены на разных уровнях и что среда от уровня z — Zj на ниж-
нем конце трубки (где скорость движения равна иь а площадь
поперечного сечения Л1) поднимается к верхнему концу на уро-
вень z = z2 (здесь скорость движения и2, площадь Л2). Применим
к объему среды, который в данный момент заполняет этот конеч-
ный отрезок трубки и в течение короткого времени поднимается
по нему, принцип сохранения энергии (механической). В словес-
ной формулировке он будет выглядеть так: увеличение кинетиче-
ской энергии (в случае, изображенном на рис. 4.7, она именно
увеличивается, так как среда течет к более узкой части трубки
тока, где, согласно принципу сохранения массы, скорость движе-
ния возрастает) плюс увеличение потенциальной энергии (среда
течет вверх, и потому эта энергия повышается) равны работе, со-
вершенной силами давления на концах трубки (давление р — pi
на конце 1, р2 на конце 2). Результат применения принципа со-
хранения энергии, представленный в математической форме, та-
Гл. 4. Основы механики жидкостей 63
ков: величина Hi, задаваемая как Hl = pY + ри2 + gpzt (где р —
плотность среды), равна величине Н%, имеющей тот же вид, за тем
исключением, что индекс 1 надо заменить на 2.
Если мы введем величину Н, называемую полным давлением
(или полным напором), в виде
Н = Р + y pw2 + gpz, (4.6)
где р, и и z— соответственно давление, скорость и уровень в лю-
бой точке среды, то сможем сделать вывод, что при стационарном
потоке величина Н, определяемая (4.6), остается постоянной
вдоль линии тока (если только влияние вязкости жидкости на те-
чение пренебрежимо мало). Постоянная вдоль линии тока вели-
чина Н, вообще говоря, для каждой линии тока имеет свое значе-
ние. Приведенный выше результат называется теоремой Бернулли.
Его можно вывести непосредственно из уравнения движения (4.5)
таким же способом, каким уравнение сохранения энергии час-
тицы выводится из уравнения ее движения (разд. 2.6). Позднее
мы подробно изучим следствия теоремы Бернулли, имеющие зна-
чение для движения крови. Пока же отметим, что при малом влия-
нии вязкости (что обычно справедливо для крупных артерий; см.
гл. 12), когда жидкость движется в горизонтальной плоскости
(т. е. z постоянна), давление р падает в тех участках, где абсо-
лютная величина скорости и возрастает вдоль линии тока, и, на-
оборот, увеличивается на участках замедления потока. При дан-
ном уровне самое низкое давление наблюдается на участках с са-
мыми большими скоростями, тогда как в покоящейся жидкости —
в самых верхних слоях. Так как изменения величин ри2 и gpz
вызывают противоположные изменения давления, они могут счи-
таться эквивалентными последнему. Величина gpz эквивалентна
гидростатическому давлению (и ее часто так и называют), а ве-
личину у ри2 называют динамическим эквивалентом давления
(или просто динамическим давлением), поскольку она связана с
движением.
Все реальные жидкости обладают вязкостью, которая может
обусловливать необратимое превращение механической энергии
в тепло, т. е. диссипацию энергии. Поэтому скорость увеличения
механической энергии элемента жидкости равна скорости, с ко-
торой совершают работу приложенные к нему силы за вычетом
скорости диссипации механической энергии (и преобразования ее
в тепловую). Вдали от границ твердых тел влияние вязкости
обычно мало, и полное давление Н вцоль небольшого отрезка ли-
нии тока примерно постоянно. Однако диссипация энергии на са-
мом деле происходит и движущийся элемент жидкости теряет ме-
ханическую энергию Поэтому свойственная элементу величина Н
64 Часть I. Основы механики
по мере его движения вдоль линии гока постепенно, но неуклонно
уменьшается. На больших расстояниях такая потеря давления мо-
жет стать заметной. Диссипация механической энергии происходит
при любом движении, когда возникают силы трения или вязкие
силы. Одним из примеров такого рода является диссипация энер-
гии при движении тела по шероховатой поверхности (гл. 2,
разд. 2.6): тело совершает работу против силы трения, и потеря
механической энергии здесь также необратима. Тепло, выделяю-
щееся вследствие диссипации энергии в вязкой жидкости, может
сказаться на ее движении: с повышением температуры среды плот-
ность некоторых ее элементов понижается и они перемещаются
вверх (всплывают) относительно других элементов. Это явление,
называемое свободной конвекцией, не имеет большого значения
для движения крови: температурные перепады в ней настолько
малы, что их даже не удается измерить. *)
*) Изменения плотности крови вследствие изменения температуры и иных
факторов (давления, состава), несущественные для физиологических процессов,
оказываются важными для некоторых экспериментальных методов (например,
ультразвуковых) изучения свойств и движения крови. — Прим. ред.
Глава 5
ДВИЖЕНИЕ ЖИДКОСТИ В ТРУБКАХ И ПРИ ОБТЕКАНИИ ТЕЛ
5.1. Пуазейлевское течение в трубке
Чтобы понять характер течения биологических жидкостей, в
частности крови, которая может проявлять неньютоновские свой-
ства, необходимо сначала рассмотреть поведение обычных, т. е.
ньютоновских жидкостей, например воды. Представим себе, что
вода течет по очень длинной горизонтальной трубке и что эта
трубка имеет круглое поперечное сечение диаметром d (рис. 5.1).
Начало и конец трубки соединены с достаточно большими резер-
вуарами, так что перепад давления на концах можно поддержи-
вать постоянным и обеспечить тем самым стационарность потока.
В стенке трубки через небольшие интервалы проделаны малень-
кие отверстия, служащие отводом для измерения давления с по-
мощью манометров. При этом можно измерять перепад давления
на единичной длине, т. е. продольный градиент давления в трубке.
Пусть давление на входе в трубку равно /?Вх, а на выходе рВых-
Повышая уровень в напорном резервуаре, т. е. увеличивая рвх—рВЬ1Х
(или Ар)1) мы увидим, что объемный расход Q тоже растет. При
этом давление по длине трубки (слева направо) распределяется
следующим образом: на некотором сравнительно большом участке,
который может составлять 100d, оно очень быстро падает, а за-
тем начинает уменьшаться с расстоянием линейно (рис. 5.2,Л).
Участок трубки, на протяжении которого давление падает срав-
нительно быстро, называется начальным участком или областью
входа. Поток за начальным участком называют полностью разви-
тым. В части трубки с полностью развитым потоком давление па-
дает с расстоянием линейно, т. е. градиент давления &p/L (раз-
ность давлений в двух точках, находящихся на расстоянии L, де-
ленная на L) постоянен. Кроме этого, в этой области градиент
давления линейно растет с увеличением объемного расхода
(рис. 5.2, Б).
Проведя подобные эксперименты с трубками различных диа-
метров d, мы увидим, что в области полностью развитого потока
при данном градиенте давления объемный расход Q резко увели-
чивается с ростом диаметра, а именно — он пропорционален d*
*) Символ Др означает разность давлений между двумя произвольными точ-
ками 1 и 2; таким образом, это просто сокращенная форма записи выражения
Pi — Р2. В общем случае ДЛ1 означает разность между значениями величины в
состояниях 1 и 2.
66 Часть I. Основы механики
(рис. 5.2,В). Поэтому, если мы хотим, чтобы объемный расход
через трубки с последовательно уменьшающимися диаметрами
был постоянным, мы должны будем прикладывать к ним гра-
диент давления, увеличивающийся как величина, обратная четвер-
той степени диаметра.
Можно поставить весьма наглядный эксперимент, который про-
демонстрирует, как меняется скорость потока в каком-либо попе-
речном сечении трубки на участке с полностью развитым потоком,
т.е. покажет нам профиль скорости в потоке. Предположим, что
мы быстро ввели в трубку (поперек нее) узкую полоску краски,
Рис. 5.1. Схематическое изображение длинной прямой горизонтальной трубки,
в которой поддерживается стационарный поток. Показаны напорный и сливиой
резервуары и боковые отводы для измерения давления.
как это показано на рис. 5.3; поскольку вода движется, эта по-
лоска будет перемещаться с ней и постепенно растягиваться та-
ким образом, как это показано на рисунке. Каждый небольшой
отрезок полоски будет перемещаться вниз по течению со ско-
ростью, равной местной скорости жидкости. Регистрируя положе-
ние полоски краски в различные моменты времени, мы смо-
жем рассчитать профиль скорости. Этот опыт можно поставить
и по-другому, вводя небольшие капли краски на разных расстоя-
ниях от оси трубки и следя за их движением. Как и ранее, мы
увидим, что каждый отдельный элемент перемещается со ско-
ростью, равной скорости движения жидкости в данной точке. При
этом в процессе перемещения капля сохраняет радиальное поло-
жение: она не смещается к оси трубки или стенке и не совершает
винтового движения. Это означает, что течение прямолинейное
(осевое). Кроме того, оно симметрично, т. е. скорость жидкости
на любом данном расстоянии от оси одна и та же независимо от
того, какой радиальный луч мы-выбираем. Подобное течение на-
зывается осесимметричным. Ниже будет показано теоретически,
что профиль скорости в действительности имеет форму параболы,
причем скорость достигает максимума на оси и постепенно умень-
шается до нуля у стенки.
В 1840 г. Пуазейль сделал первый шаг в изучении механики
кровообращения, опубликовав результаты количественных иссле-
Гл. Б. Движение жидкости в трубках и при обтекании тел 67
Рис. 5.2. А. Распределение давления вдоль трубки. Сначала (на начальном
участке) давление резко падает, а затем на достаточно большом расстоянии от
входа линейно уменьшается с расстоянием. Б. Вдали от входа градиент давления
(Ap/L) является линейной функцией объемного расхода Q через трубку. В. Вда-
ли от входа при постоянном градиенте давления объемный расход Q растет про-
порционально четвертой степени диаметра d4.
дований течения в трубках для участков, удаленных от входного
сечения; течения в этой области трубки носят теперь его имя.
В своих исследованиях Пуазейль не только менял расход и раз-
Рис. 5.3. Узкая полоска краски, имевшая вначале вид поперечной прямой линии,
под влиянием потока жидкости все больше и больше растягивается.
Стациона^иь'Й
ЛОТОК
меры трубки, но и изучил влияние вязкости на характеристики
течения. Он установил, что при увеличении вязкости для поддер-
жания заданного постоянного расхода необходимо увеличить гра-
диент давления. На основании своих экспериментов Пуазейль про-
вел теоретический анализ течения и вывел знаменитый закон для
68 Часть 1. Основы механики
стационарного потока в части трубки, достаточно удаленной от ее
начала:
Ар = 128^-.
(5.1)
Здесь ц — вязкость жидкости.
Рассматривая физическую картину течения жидкости в этой
части трубки, можно показать, как был выведен закон Пуазейля.
Для этой цели применим условие баланса сил к среде в некото-
ром сечении трубки. В гл. 4 уже было приведено в общем виде
соотношение для совокупности сил, которые необходимо рассмат-
ривать при любом балансе [уравнение (4.5)]. При пуазейлевском
Рис. 5.4. При пуазейлевском течении наблюдается параболическое распреде-
ление скорости по диаметру трубки.
течении мы имеем дело с прямолинейным стационарным потоком,
в котором жидкость движется без всякого ускорения, и поэтому
левая часть уравнения (4.5) равна нулю. Кроме того, если мы
рассматриваем поток в горизонтальной трубке, то гравитационные
силы не существенны и член, представляющий массовые силы,
также равен нулю.
Таким образом, условие баланса сил записывается в нашем
случае в следующем виде:
Вязкие силы = —Сила, обусловленная градиентом давления.
Мысленно выделим в потоке жидкости ряд концентрических
слоев, скользящих друг относительно друга так, как это показано
на рис. 5.4, и проведем анализ баланса сил, действующих на от-
дельный слой с внутренним радиусом г и толщиной dr. Если паде-
ние давления на небольшой длине слоя L равно Ар, то обусловлен-
ная давлением результирующая сила, действующая на слой, равна
произведению разности давлений на площадь поперечного сечения
слоя:
Результирующая сила = Ар X л {(г + dr)2 — г2}-
Так как dr мало, можно записать:
Результирующая сила = Арл2г dr.
Гл. 5. Движение жидкости в трубках и при обтекании тел 69
Результирующую вязкую силу, действующую на слой, можно
получить как разность вязких сил для двух поверхностей слоя.
Напряжение сдвига в произвольной радиальной точке дается вы-
ражением (разд. 4.3)
Тогда скорость изменения этого напряжения с радиусом имеет
вид
d / ди\
dT^-cd-
Таким образом, при малых приращениях радиуса dr разность на-
пряжений сдвига в точках с радиусами г и (г + dr) задается вы-
ражением
А (рА-) dr
dr dr 7
и результирующая вязкая сила, действующая на слой, есть
Сила = -А (гр, -А-) dr X 2лЛ.
Следовательно, уравнение баланса двух сил, действующих на слой,
имеет вид
— Ар X 2л rdr = (гр. А-) dr X 2 л/,.
Отсюда
d (г^ = — ‘ rdr‘
Если мы теперь проинтегрируем это уравнение (т. е. просумми-
руем подобные равенства по всем слоям в сечении и устремим dr
к нулю), то получим закон Пуазейля [уравнение (5.1)]. Мы мо-
жем также показать, что
« = 1/о(1 —£). (5.2а)
где и есть скорость среды в точке, отстоящей от оси трубки на
расстояние г, Uo — скорость потока на оси трубки, а = 4zd — ра-
диус трубки. Выражение для средней скорости среды можно запи-
сать в виде
U = 4Q/mi2 = Q/na2t
и уравнение (5.2а) при этом будет иметь вид
(5.26)
70 Часть I. Основы механики
Таким образом, распределение скорости по сечению трубки яв-
ляется параболическим, и это объясняет результаты описанного
выше эксперимента с введением в поток краски. Заметим также,
что скорость на оси трубки вдвое выше средней скорости жид-
кости.
Из параболичности профиля скорости по сечению трубки сле-
дует, что скорость сдвига является линейной функцией радиуса:
Скорость сдвига = — — 2U0r/a2.
На оси (г = 0) скорость сдвига равна нулю, по мере приближе-
ния к стенке она линейно возрастает и при г = а достигает мак-
симальной величины, равной —2(/0/ci. Таким образом, вязкое на-
пряжение сдвига на поверхности стенки равно —2t4p/a, и это есть
максимальное напряжение сдвига в потоке.
Можно получить закон Пуазейля и другим путем, основываясь
на рассмотрении баланса всех сил, которые действуют на участок
трубки длиной L и диаметром d. Если разность давлений на двух
концах участка равна Ар, то суммарная приложенная к нему сила,
обусловленная давлением, есть
Суммарная сила = \pnd2!^.
Сила, противодействующая ей, равна суммарной вязкой силе тор-
можения, действующей по всей поверхности образованного жид-
костью цилиндра. Таким образом,
Вязкая сила = Вязкое напряжение сдвига на стенке X
X Площадь внутренней поверхности трубки ------X л dL.
Так как другие силы отсутствуют,
— Apnd2/4 = — inUoLii.
Подставив Uo = 2CJ и О = 4Q/jtd2 в это уравнение и перегруппи-
ровав члены, получим
Ар= 128
r nd4
Легко видеть, что это и есть закон Пуазейля, представленный
уравнением (5.1).
Исследование полного баланса сил применительно к потоку
весьма поучительно, так как оно позволяет продемонстрировать,
что потоку противодействуют именно вязкие силы торможения со
стороны стенок трубки *).
’) Формула Пуазейля (5.1) и теорема Бернулли (разд. 4 7) практически ни-
когда не используются одновременно, так как относятся к противоположным идеа-
лизированным случаям течения: первая — к вязким течениям без инерционных
эффектов, вторая — к течениям невязкой жидкости. — Прим, ред.
Гл. 5. Движение жидкости в трубках и при обтекании тел 71
5.2. Течение на начальном участке трубки
Необходимо подчеркнуть, что условия пуазейлевского течения
реализуются только в прямых трубках, причем на достаточном
удалении от входа, а также от изгибов, сужений и других источ-
ников возмущений. Распределение скорости элементов жидкости,
проходящих через поперечное сечение ближе к входу в трубку,
меняется вдоль трубки. У самого входа все элементы жидкости
движутся с одинаковой скоростью, и профиль скорости однород-
ный, или плоский. Однако та часть жидкости, которая соприка-
сается со стенкой трубки, полностью останавливается из-за прили-
пания на стенке (разд. 4.3). Немедленно устанавливается гра-
диент скорости между неподвижной жидкостью у стенки и сосед-
ними элементами жидкости в ядре потока. По мере продвижения
Рис. 5.5. При стационарном течении изменение профиля скорости с расстоя-
нием вдоль трубки отражает увеличение толщины пограничного слоя, которая на
расстоянии к от входа равна 6. Профиль скорости, вначале плоский, постепен-
но изменяется и на большом расстоянии от входа становится параболическим.
жидкости вдоль трубки вязкость все в большей степени видоизме-
няет начальный плоский профиль скорости, как это показано на
рис. 5.5. Исходно высокий градиент скорости у стенки становится
меньше, а сдвиг охватывает все большую часть ядра потока. Сле-
дует иметь в виду, что в то же самое время центральная часть
потока ускоряется, с тем чтобы объемный расход через любое се-
чение оставался постоянным. Градиент скорости, который сначала
был заметен только около стенки, начинает проявляться на все
больших расстояниях от нее. В конечном счете скорость становит-
ся неодинаковой по всему поперечному сечению трубки: она мак-
симальна на оси и постепенно уменьшается по направлению к
стенкам. Профиль скорости принимает вид параболы, устанавли-
вается пуазейлевское течение. Когда этот полностью развитый
профиль достигнут, он уже не меняется дальше вниз по течению.
Подобное последовательное развитие профиля скорости в труб-
ке соответствует картине изменения градиента давления по мере
удаления от -входа.
На начальном участке, где профиль скорости меняется с рас-
стоянием, градиент давления исходно очень высок; однако ниже
по течению, где уже произошла существенная перестройка про-
"t2 Часть I. Основы механики
филя, градиент меньше. В конечном счете градиент давления при-
нимает минимальное значение и становится постоянным в области
с полностью развитым течением. Если вычислить вязкую силу тор-
можения в каком-либо поперечном сечении на начальном участке,
то она окажется меньше, чем движущая сила, обусловленная гра-
диентом давления. Это превышение движущей силы «расходуется»
на ускорение потока.
Основное различие между течением на начальном участке и
полностью развитым течением в трубке заключается в том, что
в первом случае элементы жидкости испытывают ускорение (по-
ложительное около оси трубки и отрицательное около стенки).
Вследствие того что поток стационарен, скорость в каждой точке
не меняется во^ времени, т. е. местное ускорение отсутствует. Тем
не менее отдельные элементы жидкости ускоряются или замед-
ляются по мере их движения по трубке. Это один из примеров
конвективного ускорения.
В общем случае при течении жидкости существуют как мест-
ные, так и конвективные ускорения и следует принимать во вни-
мание силы, необходимые для преодоления инерции текучей среды
и создания обоих этих ускорений (инерционные силы). Именно
указанным силам отвечает левая часть уравнения (4.5).
5.3. Представление о пограничном слое
Как мы уже знаем, на начальном участке профиль скорости
около стенки непрерывно меняется. Вязкая сила торможения, с
которой стенка трубки действует на прилегающие к ней слои жид-
кости, последовательно передается все более удаленным слоям.
Это обусловлено наличием вязкости.
Непосредственно около входа в трубку толщина слоя, в кото-
ром существенную роль играет вязкость, чрезвычайно мала
(рис. 5.5); за пределами этого слоя профиль скорости практически
плоский и влияние вязкости незначительно. Вниз по течению тол-
щина слоя, в котором проявляется влияние вязкости, возрастает.
Мы назовем подобный слой пограничным *). Этот термин отно-
сится к области, в которой вязкость играет определяющую роль
в формировании свойств потока.
Пограничные слои существуют не только в трубках; они есть
везде, где реальная жидкость обтекает какую-либо твердую по-
верхность, например у дна реки или вокруг корпуса корабля. Эти
’) Определение толщины пограничного слоя. Пограничным слоем называют
область жидкости, где скорость (относительно стенки) увеличивается с удале-
нием от стеики. Непосредственно около стеики скорость изменяется с расстоя-
нием чрезвычайно быстро, а в отдаленных слоях она Приближается к скорости
свободного потока Uo очень медленно Для практических целей будем принимать
за толщину пограничного слоя расстояние от стеики, на котором местная ско-
рость достигает значения 0,99 Uq.
Гл. 5. Движение жидкости в трубках и при обтекании тел 73
слои создаются и в том случае, если вместо того, чтобы застав-
лять жидкость обтекать тело, мы будем перемещать его в жидко-
сти. Так, изучая распределение скорости вокруг тела, опускающе-
гося с постоянной скоростью в воде, мы обнаружим точно такой
же пограничный слой, как и при обтекании неподвижного тела по-
током воды, движущимся с той же скоростью. Здесь существенно
только относительное движение тела и жидкости. Сказанное спра-
ведливо для всех реальных жидкостей, но детальная картина про-
Рис. 5.6. Баланс сил, действующих на элемент жидкости в пограничном слое.
филя скорости в пограничном слое зависит от вязких свойств жид-
кости; не следует думать, что профиль будет одинаков для ньюто-
новских и неньютоновских сред.
Рассмотрим теперь скорость увеличения толщины погранич-
ного слоя по длине трубки; она определяется балансом между
инерционными силами и вязкими силами торможения в слоях,
прилегающих к стенке. Пусть мы имеем малый элемент жидкости
в области с изменяющейся скоростью (рис. 5.6). Тангенциальное
напряжение (напряжение сдвига) на поверхности A i (пло-
щадью Л) равно |x(dw/dx/)i. Аналогично напряжение сдвига на по-
верхности А2 (площадь ее также равна Л) есть |x(du/dx/)2- Два
этих напряжения немного различаются по абсолютной величине,
поскольку в рассматриваемой области градиент скорости не по-
стоянен (рис. 4.7). Таким образом, результирующая вязкая сила,
действующая на элемент, определяется через зависимость напря-
жения от расстояния до стенки у:
Вязкие силы', действующие на элемент, должны быть уравнове-
шены инерционными силами. Выражение, отвечающее инерцион-
ной силе, имеет вид
р X Конвективное ускорение X Объем элемента.
74 Часть I. Основы механики
Здесь р — плотность жидкости, а объем элемента равен А (у2— ух).
Если приравнять эти две силы, действующие на элемент, и по-
пытаться разрешить получившееся уравнение, то окажется, что
методов нахождения аналитического решения подобных уравне-
ний не существует. Для частных задач можно получить решение
численным методом.
В гидромеханике обычно используется подход, в рамках кото-
рого для получения представления о масштабе или порядке вели-
чины интересующей нас физической величины задают масштабы
или порядки величин определяющих переменных (из гл. 6 мы
узнаем, что это частный случай метода анализа размерностей).
Так, чтобы получить оценку вязких сил, мы можем связать их с
локальной толщиной пограничного слоя б и скоростью свободного
потока U. Тогда вязкая сила будет пропорциональна
^~А(у2 — r/О.
С другой стороны, мы можем получить порядок величины силы
инерции. Пусть рассматривается толщина пограничного слоя на
расстоянии X от входа в трубку; время прохождения среды от
входа до сечения X (время переноса) имеет порядок величины
Х/U. Тогда конвективное ускорение среды на расстоянии X имеет
масштаб U/(X/U), или U* 2/X.
Приравняв порядки величин сил, мы получим соотношение
где k — численная постоянная, величину которой следует опреде-
лить экспериментально. Таким образом, мы приходим к выводу,
что __ __________________
6oc/\/pU'’ или (5.3) *)
Из уравнения (5.3) видно, что толщина пограничного сдоя растет
вдоль трубки2) как корень квадратный из расстояния от входа
(см. рис. 5.5), т. е. как Х1/2. С увеличением расхода или скорости
свободного потока толщина пограничного слоя во всех точках
уменьшается. При данном расходе толщина слоя увеличивается
с ростом вязкости жидкости.
Проведенный анализ дает очень хорошую оценку толщины по-
граничного слоя для области, примыкающей к входу в трубку,
т. е. там, где величина б мала по сравнению с радиусом трубки.
Однако когда толщина пограничного слоя становится довольно
*) Здесь v = ц/р — кинематическая вязкость жидкости. — Прим. ред.
2) Применение подобных рассуждений для пограничного слоя на стенке труб-
ки требует, вообще говоря, учета градиента давления в балансе сил, однако
можно доказать, что это не скажется на окончательных выводах. — Прим. ред.
Гл. б. Движение жидкости в трубках и при обтекании тел 75
большой, характер изменения ее роста меняется, так как в каж-
дом поперечном сечении начинает сказываться взаимозависимость
скоростей роста толщины пристеночного слоя в соседних точках
периметра сечения. В результате взаимодействия соседних обла-
стей пограничного слоя его формирование несколько замедляется.
Тем не менее из уравнения (5.3) можно оценить расстояние, при
котором пограничный слой заполняет всю трубку и устанавли-
вается течение Пуазейля. В этом случае его толщина становится
равной радиусу трубки d/2. Следовательно,
Выражение Ud/v называется числом Рейнольдса (Ре) в честь
английского ученого, который первым выполнил многие важные
исследования потоков в трубах; роль числа Рейнольдса в гидро-
механике более подробно будет рассмотрена ниже. Константа k',
определенная экспериментально, составляет примерно 0,03. Таким
образом, длина начального участка для стационарного потока в
прямой трубке задается выражением
X = 0,03d (Ре). (5.4)
Для трубки диаметром 2 см длина начального участка составит
при числе Рейнольдса 1000 около 60 см — именно эти значения
были бы характерны для аорты человека, если бы она была пря-
мой трубкой, а поток в ней — стационарным и имел скорость
0,2 м-с-1.
Уравнение (5.4) удовлетворительно предсказывает длину на-
чального участка только при значениях числа Рейнольдса в ин-
тервале от 10 до 2500. Когда Ре очень мало (меньше единицы),
влияние инерции несущественно, а длина начального участка чуть
больше одного диаметра трубки. Когда же Ре превышает 2500.
характеристики потока резко отличаются от описанных выше:
плавные линии тока отсутствуют, наблюдается турбулентность
(разд. 5.5).
Ниже мы увидим, что течение на уровне микрососудов можно
считать полностью развитым, а в крупных артериях оно близко
к течению на начальном участке и к тому же усложнено из-за
особенностей геометрии сосудов.
5.4. Число Рейнольдса
При анализе баланса сил, действующих на элемент жидкости,
мы уже обращали внимание на отношение величин инерционных
и вязких сил. В действительности это отношение является важной
характеристикой всех течений жидкости.
Для потока в трубке характерными величинами скорости и
размера системы являются U и d. Поэтому можно сказать, что
76 Часть I. Основы механики
порядок величины (масштаб) вязких сил такой же, как у произ-
ведения вязкости на масштаб скорости сдвига (=p.t//d). С дру-
гой стороны, инерционная сила пропорциональна кинетической
энергии единичного объема жидкости (р£72). Тогда относительная
значимость этих двух величин может быть оценена как
Число Рейнольдса =—еРционные силы (5.5)
Вязкие силы ц v ' '
где v — кинематическая вязкость жидкости (ц/р). Число Рей-
нольдса Re—безразмерная величина, и потому оно не зависит от
единиц, в которых измеряются различные входящие в него пара-
метры, при условии, конечно, что эти единицы согласованы друг
с другом. В этой книге мы всегда будем характеризовать рассмат-
риваемый поток числом Рейнольдса. Оно применимо для описа-
ния потоков не только в длинной прямой трубке, но и вообще лю-
бого потока и рассчитывается из характерных величин скорости
и размера системы. Например, рассматривая обтекание шара, ис-
пользуют в качестве таких величин диаметр шара и скорость жид-
кости вдали от него. Когда в предыдущем разделе мы вычисляли
скорость увеличения толщины гидродинамического пограничного
слоя в трубке, то получили уравнение (5.3), которое может быть
представлено в виде
б » k ("77’) •
или ______
6 “ kX •
Легко видеть, что член UX/v— это число Рейнольдса Rex для по-
тока, в котором роль характерного размера играет расстояние X
от начала трубки до данного сечения. Поэтому можно переписать
уравнение (5.3) в виде
f) = kXRe~il2_ (5.6)
Когда мы можем считать, что вязкие силы преобла-
дают, а инерционные пренебрежимо малы. Например, на уровне
микрососудов (сюда мы будем относить сосуды диаметром по-
рядка 100 мкм и менее) типичные числа Рейнольдса меньше еди-
ницы и течение может рассматриваться как чисто вязкое. Когда
же Re^> 1, напротив, преобладают инерционные силы, а вязкость
(во всех областях, кроме пограничных, где она по-прежнему су-
щественна) лишь незначительно изменяет характер течения. При-
мерами такой ситуации служат течения в крупных артериях и
венах.
Гл. 5. Движение жидкости в трубках и при обтекании тел 77
5.5. Турбулентность при течении в трубке
В 1883 г. Рейнольдс опубликовал результаты важных исследо-
ваний, касающихся течения в длинных трубках. Измеряя градиент
давления вдоль трубки при увеличении расхода, он обнаружил,
что при относительно низких скоростях градиент давления прямо
пропорционален расходу, что и следовало ожидать из закона
Пуазейля [уравнение (5.1)], а при более высоких становится при-
мерно пропорционален квадрату расхода (рис. 5.7). При некото-
рых промежуточных значениях расхода связь между градиентом
Рис. 5 7. Построенный в
логарифмическом масштабе
график зависимости объем-
ного расхода Q от градиен-
та давления kp/L при ста-
ционарном течении, иллюст-
рирующий изменение харак-
тера этой зависимости по
мере увеличения расхода.
давления и расходом менялась во времени: в один момент наклон
кривой на рис. 5.7 мог быть равен 1, а в другой — увеличиться
примерно до 2. Если в ходе эксперимента принимались специаль-
ные меры для обеспечения полной стационарности потока на входе
в трубку, а сама трубка была прямой и чрезвычайно гладкой, то
удавалось добиться того, что критическое значение расхода, при
котором наблюдалась смена одного режима течения другим, ста-
новилось гораздо выше.
Для потока воды в трубке диаметром 1 см критический рас-
ход, при котором происходит изменение характера указанной за-
висимости, составляет примерно 1 л-мин-1. Эта величина мала по
сравнению с теми расходами, которые встречаются в повседнев-
ной практике (скажем, в водопроводных трубах или шлангах для
полива).
Рейнольдс поставил еще одну серию классических экспери-
ментов, где ему удалось визуализировать процессы внутри по-
тока. Он изучал течение воды в стеклянной трубке, вводя в ту ее
часть, которая достаточно удалена от начала, тонкую струйку
подкрашенной воды по оси трубки (т. е. в основном потоке), а
затем наблюдая за ее поведением. При низких расходах струйка
оставалась сплошной и двигалась вдоль оси трубки в виде от-
78 Часть I. Основы механики
Рис. 5.8. Движение струйки подкрашенной воды в длинной прямой трубке.
Вверху, стационарное осевое движение струйки в потоке при низких Re. Посе-
редине: при Re, немного превышающих критическое значение, ламинарное движе-
ние нарушается и наблюдаются всплески турбулентности. Внизу: при более вы-
соких Re поток становится полностью турбулентным, краска движется хаоти-
чески.
дельной нити. При более высоких расходах непрерывность нару-
шалась и струйка начинала перемешиваться со всем объемом
воды (рис. 5.8); скорость перемешивания увеличивалась с ростом
расхода. При расходах, лишь немного превышающих критическое
значение, наблюдалось чередование областей плавного движения
струйки и нарушенного ее течения.
Хаотичность движения подкрашенной воды означает, что и в
самом потоке происходят интенсивные хаотические движения. Если
бы мы провели скоростную киносъемку движения струйки, а за-
тем медленно просмотрели пленку, то увидели бы, что отдельные
элементы струйки движутся беспорядочно, так что их скорости
Гл. 5. Движение жидкости в трубках и при обтекании тел 79
хаотично изменяются во времени и в трех направлениях. Однако,
рассчитав среднюю по времени скорость небольших элементов
струйки, проходящих через некоторую точку данного поперечного
сечения трубки, а затем повторив этот расчет для другой точки
того же сечения, мы увидели бы, что существует четко выражен-
ный профиль средней по времени скорости. При этом средняя ско-
рость в любой точке сравнима по величине с мгновенными скоро-
стями хаотического движения элементов. Увеличение градиента
давления, наблюдавшееся в первой серии экспериментов Рей-
нольдса, происходит как раз в тот момент, когда начинается тур-
булентное движение. Турбулентность приводит к увеличению дис-
сипации энергии и, как следствие этого, к более быстрому паде-
нию давления.
Расчет числа Рейнольдса для течений в описанных выше опы-
тах показывает, что при Re < 2300 поток всегда ламинарный,
когда же Re превышает 2500, в потоке постоянно наблюдается
турбулентность, которая становится все более интенсивной с рос-
том Re. В диапазоне между 2300 и 2500 происходит смена режи-
мов и турбулентность то появляется, то исчезает ')•
Метод визуализации течения с помощью струйки красителя
можно использовать также для наблюдения за развитием турбу-
лентности в трубке по мере удаления от входа. Для этого в си-
стему, изображенную на рис. 5.1, когда расход достаточно высок
и поток турбулентен, вводят струйки красителя на разных рас-
стояниях от начала трубки и в разных точках данного сечения,
наблюдая за их поведением по мере того, как они сносятся вниз
по течению. Непосредственно на входе профиль скорости плоский,
все элементы жидкости движутся с одинаковой скоростью. На не-
большом расстоянии от входа под действием вязких сил происхо-
дят определенные изменения профиля (их характер рассматри-
вался нами выше), но струйки еще сохраняются и турбулентность
отсутствует. Ниже по течению непрерывность струек внезапно на-
рушается, т. е. поток становится турбулентным. Первыми разру-
шаются струйки, проходящие по краю пограничного слоя, а через
небольшое расстояние —струйки внутри пограничного слоя и в
ядре потока. Сформировавшийся далеко вниз по течению профиль
средней по времени скорости отличается по форме от параболи-
ческого, свойственного ламинарному потоку. Ядро потока имеет
*) Подчеркнем, что авторы не ставят здесь целью ни дать полные опреде-
ления понятий ламинарного и турбулентного режимов, ни тем более разъяснить
физические механизмы турбулентности. Утверждается лишь, что при течении
жидкости в трубах (как и в других типах течений) по достижении определен-
ных (для каждого типа течений) значений числа Рейнольдса резко изменяется
характер течения. Эти значения Re принято называть критическими (7?екР). При
переходе Re через критическое значение в потоке всегда возникают беспорядоч-
ные возмущения. Последние по определению свидетельствуют о переходе течения
ИЗ ламииариого режима в турбулентный. — Прим. ред.
80 Часть I. Основы механики
гораздо более плоский профиль, а градиент скорости у стенки бо-
лее крутой; максимальная скорость наблюдается у оси и при-
мерно в 1,2 раза выше средней по сечению скорости.
Длина начального участка для турбулентного течения меньше,
чем для ламинарного; ее можно оценить из следующего соотно-
шения:
(д.7)
5.6. Нестационарное течение в очень длинной трубке
Обратимся вновь к ламинарному течению в длинной прямой
трубке. Если к жидкости приложен медленно колеблющийся гра-
диент давления, то течение будет замедляться, останавливаться,
изменять свое направление, ускоряться в новом направлении,
снова замедляться и т. д. Когда все это происходит достаточно
медленно, то течение в каждый момент будет характеризоваться
параболическим профилем скорости с максимумом, пропорцио-
нальным мгновенному значению расхода. Однако по мере увели-
чения частоты колебаний градиента давления профиль скорости
постепенно искажается (рис. 5.9). Инерция не позволяет жидко-
сти в центральной части потока двигаться синхронно с приложен-
ным градиентом давления, наблюдается запаздывание, величина
которого растет при увеличении частоты колебаний градиента
давления. В то же время амплитуда движений жидкости в ядре
уменьшается. При очень высоких частотах движения в ядре чрез-
вычайно малы и течение происходит только в очень тонком слое
около стенки, где преобладают вязкие силы и течение менее чув-
ствительно к влиянию инерции.
Простейшая форма колебаний градиента давления — синусои-
дальная. В этом случае мгновенный градиент давления Др« свя-
зан с максимальным (пиковым) градиентом Дро зависимостью
Др# = Др0 sin &t. (5.8)
Здесь t — время, ® — угловая частота (® = 2л/, где f — частота
колебаний). Мгновенный градиент давления изменяется во вре-
мени следующим образом: в момент t = 0 он равен нулю; далее
Др« растет до максимальной величины Дро, затем падает до нуля,
становится отрицательным и уменьшается до величины —Др0,
после чего вновь начинает расти и проходит через нуль; далее
цикл повторяется. Период времени Т, на протяжении которого
происходит один цикл колебаний, равен 2 л/<о. Более полно сину»
соидальные функции рассмотрены в гл. 8.
Весьма полезным безразмерным параметром является число
Уомерсли а, показывающее, как сильно отличается профиль ско-
рости при ламинарном течении в длинной трубке от пуазейлев-
Гл. 5. Движение жидкости в трубках и при обтекании тел 8t
Рис. 5.9. Построенные с интервалом 16° профили скорости для жидкости, те-
кущей в длинной прямой трубке под действием синусоидального градиента
давления- (McDonald. Blood flow in arteries, 1974, p. 104, Edward Arnold,
Ьопдоп.)А.Значение а достаточно велико для возникновения около стенок воз-
вратного” потока, в ядре же жидкость движется вперед. Б, В н Г. Нарастание
искажения профиля скорости при значениях а, соответствующих второй, третьей
и четвертой гармоникам основной частоты колебаний, которую представляет
график А,
ского, когда жидкость подвергается воздействию синусоидально
меняющегося с угловой частотой © градиента давления:
(5-9)
Здесь d — диаметр трубки, v — кинематическая вязкость (v =
= р/р). Параметр а может рассматриваться как число Рейнольд-
са для нестационарного потока, поскольку он характеризует отно-
шение инерционных и вязких сил, определяющих движение за
время порядка одного периода колебаний ’)
При низких значениях а (меньше единицы) течение «квазиста-
ционарное»; в каждый момент времени профиль скорости парабо-
лический, расход определяется мгновенным значением градиента
давления, в течении преобладают вязкие силы, а инерционными
можно пренебречь. При несколько более высоких значениях а
мгновенный профиль скорости искажается (рис. 5.9) вследствие
*) Наряду с а для нестационарного потока существенно и число Рейнольдса
в обычном смысле: Re где U — средняя за период илн максимальная ско-
v
рость. Число Re в отличие от а отражает отношение сил в среднем за цвкл. -«
Прим, ред,
82 Часть I. Основы механики
влияния инерционных сил. Более того, мгновенный расход уже не
определяется мгновенным градиентом давления: поток запазды-
вает по отношению к приложенному градиенту давления. Можно
описать эту ситуацию аналитически следующим образом. При
низких значениях а приложенное давление, изменяющееся со-
гласно уравнению (5.8), порождает расход Qt, величина которого
Рис. 5.10. Изменение амплитуды (ввер-
ху) и фазы (внизу) колебательного
потока, возникающего прн синусоидально
меняющемся градиенте давления, с уве-
личением параметра а. Амплитуда ха-
рактеризуется отношением максимально-
го мгновенного расхода Qmax к стацио-
нарному расходу <2стац> который устано-
вился бы при максимальном (соответ-
ствующем амплитудному значению гра-
диента давления) перепаде давления на
трубке. (McDonald. Blood flow in arte-
ries, 1974, p 125, Edward Arnold, Lon-
don.)
определяется мгновенным градиентом давления в соответ-
ствии с законом Пуазейля. Следовательно,
Qt — Qct sin (<о/), (5.10)
где QCT — расход, полученный из закона Пуазейля для градиента
давления Дро- При более высоких а (в диапазоне 1—3) мгновен-
ный расход отстает от мгновенного градиента давления на время
<р/и и уравнение (5.10) приобретает вид
Qt = Qct sin {и/ — <р},
т. е. расход в данный момент определяется через градиент давле-
ния в предшествующий момент t — <р/<о. При еще более высоких
а (больших, чем примерно 4) в потоке начинают преобладать
инерционные силы, причем в такой степени, что не только увели-
чивается отставание по фазе, но и существенно уменьшается пико-
вое значение расхода Qa по сравнению с величиной QCT, предска-
зываемой законом Пуазейля для градиента давления Др0- Изме-
нения запаздывания по фазе <р и пикового расхода при увеличе-
нии а представлены на рис. 5.10.
В системе кровообращения величины а (рассчитанные для час-
тоты сокращений сердца) изменяются в широком диапазоне: в
аорте а может быть больше 10, тогда как в капиллярах эта вели-
чина составляет 10-3. Более того, в системе кровообращения и во
многих других представляющих интерес системах встречаются
гораздо более сложные профили скорости, чем пуазейлевский,—
например профили на начальном участке трубки или профили,
искаженные из-за геометрических факторов. В этих случаях пра-
Гл. 5. Движение жидкости в трубках и при обтекании тел 88
вильнее было бы определять а не через диаметр трубки или со-
суда, а через толщину б гидродинамического пограничного слоя,
который существовал бы в этих условиях при стационарном тече-
нии, так как в действительности а характеризует искажения ста-
ционарного течения в данной системе при колебаниях. Таким об-
разом, следовало бы положить а = б д/ ~ • Поскольку толщина
пограничного слоя всегда меньше радиуса трубки (d/2), то, оче-
видно, и а всегда меньше, чем его значение, полученное исходя
из параметров развитого течения. Для системы кровообращения,
однако, не удается точно определить величину б, и параметр а
всегда рассчитывается исходя из диаметра сосуда.
5.7. Влияние сужений на характер течения в трубке
До сих пор мы рассматривали течения в трубках с постоянным
диаметром. В этом разделе будут проанализированы эффекты,
возникающие при течении через сужения, а в следующих — свой-
ства течений вокруг плохо обтекаемых тел и в изогнутых трубках.
Рис. 5 11. Поток в круг-
лой трубке, площадь попе-
речного сечения которой уве-
личивается от Л! до Л2; ско-
рость при этом уменьшается
от f7i до U2.
Рассмотрим длинную горизонтальную трубку, площадь попе-
речного сечения которой резко увеличивается от At до At
(рис. 5.11). Будем считать, что жидкость в трубке несжимаемая,
а течение стационарное и ламинарное; кроме того, предположим
на время, что вязкостью жидкости можно пренебречь. Ясно, что
при переходе жидкости в более широкую часть трубки скорость ее
должна уменьшиться, поскольку объемный расход в обоих сече-
ниях одинаков:
UXAX = U^. (5.11)
Так как вязкость отсутствует и диссипации энергии не происхо-
дит, можно применить теорему Бернулли и записать соотношение
баланса энергии потока в двух сечениях:
уР^ + Р1 = уР^ + р2,
или
<е‘2)
84 Часть 1. Основы механики
Поскольку, как мы знаем, {72 меньше, чем Ub статическое давле-
ние в расположенном ниже по потоку сечении с большей пло-
щадью будет больше, чем в сечении выше по потоку. Избыток ки-
нетической энергии, связанной с потоком, «преобразуется» при
этом в статическое давление. Если бы поток имел противополож-
ное направление, то произошло бы падение статического давления
на входе в более узкую часть трубки. Эти рассуждения справед-
ливы и в случае реальной жидкости с низкой вязкостью, по-
скольку падение давления из-за наличия вязкости на коротком от-
резке трубки в области изменения сечения мало по сравнению с
давлениями, входящими в уравнение (5.12).
Исходя из этого положения была сконструирована так назы-
ваемая трубка Вентури — прибор для измерения скорости потока.
р, ₽>
Рис. 5.12. Поток в трубке Вентури, где площадь поперечного сечения сна-
чала уменьшается от А до Л2, а затем медленно увеличивается до Д. Давле-
ние измеряется до сужения (pt) в наиболее узкой части (р3) и по выходе нз
трубки (р2).
Как показано схематически на рис. 5.12, этот прибор состоит из
трубки, которая сначала сужается от исходной площади попереч-
ного сечения At до горловины площадью А2, а затем опять расши-
ряется до первоначальной площади сечения. Чтобы избежать осо-
бого рода потерь энергии, связанных с отрывом потока, расши-
ряющаяся часть выполнена таким образом, что половина угла
раствора стенок не превышает пяти градусов (явление отрыва мы
обсудим позднее).
В предположении отсутствия потерь на трение на основании
сказанного выше можно заключить, что pi больше, чем р3, и что
вниз по течению давление снова увеличивается, достигая к концу
трубки значения р2, которое должно быть равно plt хотя в дей-
ствительности небольшие потери на трение существуют и р2 ока-
зывается несколько ниже, чем pi. В правильно сконструирован-
ном приборе эта разница мала и ею можно пренебречь. Объемный
расход Q можно рассчитать следующим образом (существенно,
что здесь мы говорим не о массовом расходе, а об объемном;
перепутав эти две величины, мы придем к неоднородным уравне-
ниям с несовместимыми единицами). Используя уравнение (5.12)
для баланса энергии в сужающейся части, получим
Р!-Рз = |р(^-^). (5-13)
Гл. б. Движение жидкости в трубках и при обтекании тел 86
Согласно уравнению (5.11),
171Д1 — 172Л2,
т. е.
Подставив это выражение в уравнение (5.13), будем иметь
Но поскольку Q = UiA ь
откуда
Q =
Таким образом, если геометрия системы известна, то можно рас-
считать объемный расход по измеренному уменьшению давления
в сужающейся части трубки.
Обычный лабораторный водоструйный насос для откачки, ко-
торый подсоединяется к водопроводному крану, представляет со-
бой устройство, во многом похожее на трубку Вентури. Вода, те-
кущая через сужение, ускоряется, что вызывает локальное умень-
шение статического давления и обусловливает отсасывающий эф-
фект.
Когда после сужения трубка расширяется слишком быстро, на-
блюдается «отрыв потока». Если раньше имело место простое дви-
жение жидкости вдоль линий тока, заполняющих все поперечное
сечение, то теперь в ядре потока жидкость движется по линиям
тока вперед, а у стенок поток затормаживается и наблюдается
возвратное течение жидкости (рис. 5.13). Подобный отрыв обу-
словлен увеличением давления в этой области (см. выше).
В прямой трубке давление в направлении движения падает, и
это обеспечивает безотрывность течения; в сужающейся части из-
мерителя давление тоже падает, причем даже быстрее, чем в пер-
вом случае. Напротив, в расширяющейся части измерителя дав-
ление в направлении движения растет. Эта направленная назад
сила давления стремится замедлить поток. В ядре инерция жид-
кости противодействует замедлению, однако у стенок скорость
жидкости вследствие прилипания мала и влияние инерции на-
столько низко, что им можно пренебречь по сравнению с вязкими
силами. В результате если в ядре направленный назад градиент
давления только замедляет жидкость, то у стенки он в конечном
счете изменяет направление потока. Когда это происходит, на-
блюдается резкое изменение.свойств пограничного слоя, который
86 Часть I. Основы механики
Рис. 5.13. А Отрыв потока при течении жидкости через сужение в трубке,
за которым следует быстрое расширение. Б Профили скорости в сечениях А—А'
н В—В'. До сужения поток всюду направлен вперед; за сужением жидкость
около стенки движется в обратном направлении.
перестает быть тонким и, как говорят, отрывается. Характер изме-
нения течения в ядре при отрыве потока схематически изображен
на рис. 5.13. Пока течение ламинарно, степень отрыва или размер
зоны отрыва, как и интенсивность возвратного течения, растут
с увеличением расхода. Тенденция к отрыву у турбулентных пото-
ков менее сильна, чем у ламинарных: турбулентный поток может
оставаться безотрывным при более резких изменениях геомет-
рии ’).
В трубке можно создать гораздо более сильное сужение, чем
в описанном выше приборе Вентури. К резкому изменению в по-
токе, например, приводит вставленная в трубку диафрагма (за-
слонка с отверстием). Рассмотрим здесь явления в трубке с диаф-
рагмой, имеющей острую кромку, ограничиваясь вначале стацио-
нарным ламинарным течением реальной жидкости (см. рис. 5.14).
При подходе к сужению линии тока изгибаются, а скорость уве-
личивается. Скорость жидкости в области, ограниченной стенкой
*) Отрыв потока, образование застойных зон и т. п. поясняют обычно кар-
тинами лнннй тока илн траекториями частиц жидкости. Когда речь идет о подоб-
ных явлениях в турбулентном режиме, имеют в виду линии тока н траектории
для усредненного течения. — Прим. ред.
Гл. 5. Движение жидкости в трубках и при обтекании тел 87
трубки и диафрагмой, чрезвычайно мала; эта область фактически
является застойной зоной. При истечении через диафрагму струя
продолжает еще какое-то время сжиматься, пока не достигнет
точки «сжатого сечения», где эффективная площадь поперечного
сечения потока минимальна. Затем поток начинает расширяться
и в конечном счете снова заполняет всю трубку. Непосредственно
за диафрагмой всегда образуется зона отрыва, для которой ха-
рактерно вихреобразное возвратное течение. Оно является резуль-
татом резкого увеличения поперечного сечения сразу за отвер-
стием, что создает очень большой градиент давления, противо-
действующий потоку. При выходе из отверстия поток фактически
Зона отрыва
Поток
"Сжатое сечение"
Застойная зона
Рис. 5.14. При истечении через диафрагму струя продолжает какое-то время
сжиматься, достигая минимальной площади поперечного сечения в точке «сжа-
того сечения». Непосредственно за диафрагмой происходит отрыв потока.
ведет себя как струя, и скорость ее расширения сравнительно
мала; в результате зона отрыва потока распространяется на за-
метное расстояние вниз по течению.
Если мы измерим статическое давление перед отверстием, где
поток еще мало возмущен, и сравним его со статическим давле-
нием на уровне «сжатого сечения», то окажется, что разность ме-
жду ними несколько больше, чем можно ожидать исходя из ба-
ланса энергии, предполагающего отсутствие вязких эффектов.
Ниже по течению соответствие теореме Бернулли также не на-
столько полное, как для трубки Вентури, что обусловлено вязкими
потерями, связанными с отрывом потока. Сравнивая статические
давления достаточно далеко от отверстия выше и ниже по тече-
нию, мы обнаружим значительные уменьшения давления (или
энергии)—результат вязкой диссипации энергии (перехода ее в
тепло) в области возмущенного потока за отверстием. При низ-
ких расходах диссипация происходит в основном в отрывной зоне,
а при более высоких сама выходящая из отверстия струя стано-
вится возмущенной и турбулентной, и это вносит значительный
дополнительный вклад в потери энергии.
88 Часть I. Основы механики
Исследование устойчивости течения *) через диафрагму пока-
зывает, что при низких расходах истекающая струя ламинарна, а
при более высоких в ней появляются возмущения и турбулент-
ность. Подсчитав с помощью уравнения (5.5) число Рейнольдса
для потока через отверстие (используем в качестве параметров
диаметр отверстия и среднюю скорость проходящей через него
жидкости), получим, что изменение характера истекающей струи
происходит при числах Рейнольдса 300—400. Когда в трубке име-
ется некое препятствие с отверстием, очень часто именно так и
происходит — в большей части трубки течение ламинарно, но
струя, истекающая из отверстия, турбулентна.
В системе кровообращения сужения потока возникают при
коарктации и стенозе. Коарктация, по-видимому, сходна с суже-
нием в трубке Вентури, а стеноз — с диафрагмой. Стеноз, который
встречается чаще, сопровождается четко выраженными звуковыми
колебаниями; их объясняют шумами, вызванными турбулент-
ностью в струе за отверстием (гл. 11 и 12).
5.8. Течение в изогнутых трубках
До сих пор мы рассматривали свойства потока в прямой труб-
ке; течениям в изогнутой трубке присущи некоторые специфиче-
ские особенности, представляющие для нас интерес.
Когда жидкость течет стационарно и без турбулентности по
изогнутой части трубки (рис. 5.15), то каждый ее элемент изме-
няет направление движения. Это означает, что он имеет состав-
ляющую ускорения, перпендикулярную направлению движения.
Последнее в свою очередь означает, что, как и в случае отдельной
частицы, движущейся по окружности, на каждый элемент среды
действует сила, перпендикулярная скорости. В данном случае та-
кая сила создается «боковым» градиентом давления (в плоскости
рис. 5.15), направленным от наружной стенки изгиба к внутрен-
ней. Этот градиент более или менее одинаков по всему попереч-
ному сечению трубки, и ко всем элементам с одинаковой массой
приложена, таким образом, примерно одинаковая боковая сила,
позволяющая им совершить поворот. Поэтому и боковое ускорение
у\ всех элементов одинаково. Но это означает, что элементы, дви-
жущиеся с большой скоростью, будут как более инерционные мед-
леннее изменять направление своего движения, нежели элементы,
*) Понятие устойчивости течения (как н вообще механического движения)
возникает в связи с изучением действия внешних возмущающих факторов. Если
после кратковременного возмущения система самопроизвольно возвращается к
исходному состоянию, то она устойчива. В реальных течениях всегда существуют
возмущения, и ламинарный режим возможен лишь при условии устойчивости
Течения. — Прим, ред,
Гл. б. Движение жидкости в трубках и при обтекании тел 89
Рис 5.15 Продольное осевое сечение изогнутой круглой трубки радиусом а.
На входе в изогнутый участок поток пуазейлевский. Радиус кривизны осевой ли-
нии трубки равен R. Элемент жидкости, движущийся со скоростью и по траек-
тории с радиусом кривизны г, испытывает направленное к центру ускорение, рав-
ное и!/г.
Рис. 5.16. А. Искажение профиля продольной скорости вследствие изгиба труб-
ки. Б. Проекция линий тока на плоскость поперечного сечения. Хорошо видна
форма вторичного течения.
движущиеся с малой скоростью1). Таким образом, жидкость,
имеющая большую скорость (т. е. жидкость из центральной части
трубки), будет стремиться переместиться к наружной стенке из-
гиба, а на смену ей придет жидкость из пристеночной области, где
скорость меньше. В результате возникнет поперечная циркуляция,
или вторичное течение (рис. 5.16). Кроме того, из-за смещения
более быстро движущихся элементов жидкости от осевой линии
’) С другой стороны, абсолютная величина бокового ускорения произвольной
частицы равна и2/г, где и есть скорость частицы, а г — радиус кривизны ее
траектории; таким образом, если боковое ускорение одно и то же, то траекто-
рии частиц, движущихся медленнее, должны иметь меньший радиус кривизны.
90 Часть I. Основы механики
профиль скорости потеряет симметричность и максимальная ско-
рость будет достигнута ближе к наружной стенке. Такая картина
течения наблюдается в длинной трубке постоянной кривизны
вдали от входного сечения, т. е. в области с полностью развитым
течением.
Важное следствие искажения пуазейлёвского профиля скорости
и наличия вторичных течений состоит в появлении областей (в ча-
стности, около наружной стенки изгиба, рис. 5.16), где скорости
сдвига значительно выше, чем в пуазейлевском потоке. Высокие
Рис. 517 Искажение плоского про-
филя скорости при входе в изогну-
тую часть трубки
скорости сдвига приводят к высокой скорости диссипации механи-
ческой энергии из-за наличия вязкости. Таким образом, при про-
движении среды через изогнутую трубку необходимо совершать
большую работу, чем когда трубка прямая. Другими словами,
градиент давления, необходимый для поддержания заданного объ-
емного расхода, в изогнутой трубке больше, чем в прямой Если
изогнутая трубка затем переходит в прямую, то в потоке за изги-
бом на некотором расстоянии еще будут представлены описан-
ные выше вторичные течения. Постепенно затухая, они исчезнут
совсем, и на достаточно большом расстоянии от изгиба вновь уста-
новится пуазейлевский поток. Длина трубки, на которой сущест-
вуют вторичные течения и возмущения, сравнима с определяемой
уравнением (5.4) длиной начального участка для пуазейлевского
течения в прямой трубке.
В изогнутой трубке, как и в прямой, у стенок образуется тон-
кий пограничный слой. Однако профиль скорости в ядре потока,
где роль вязкости несущественна, далек от плоского, каким он
был на входе в изгиб. Эксперимент показывает, что скорость жид-
кости у внутренней стенки больше, чем у наружной (рис. 5.17).
Этот эффект можно объяснить с помощью теоремы Бернулли
[уравнение (4.6)], которая для потока в горизонтальной плоско-
сти (когда сила гравитационного притяжения не сказывается)
гласит, что величина
Я = р + 4р«2 (5.14)
Гл. 5. Движение жидкости в трубках и при обтекании тел 9!
постоянна вдоль каждой линии тока; как и ранее, здесь р, р и и
соответственно плотность, давление и скорость. Ввиду того что
каждая линия тока начинается из области, где р и и одинаковы
во всех точках, константа Н имеет одно и то же значение для всех
линий тока. Далее, хотя на начальном участке явные вторичные
движения в ядре потока отсутствуют, потому что все элементы
жидкости имеют примерно одинаковую скорость, должен суще-
ствовать боковой градиент давления, заставляющий жидкость дви-
гаться по изогнутым траекториям. Следовательно, у наружной
стенки р больше, чем у внутренней, и чтобы величина Н остава-
лась постоянной во всех точках сечения, скорость среды и у на-
ружной стенки должна быть меньше, чем у внутренней.
Вторичные течения будут малы до тех пор, пока толщина по-
граничного слоя не станет достаточно большой и не установится
полностью развитый поток. Однако недавние эксперименты и тео-
ретические исследования показали, что детальная картина тече-
ния внутри пограничного слоя, где слабые вторичные течения воз-
никают сразу, зависит от кривизны стенок. Это приводит к инте-
ресным последствиям, для объяснения которых удобнее всего вос-
пользоваться понятием скорости сдвига. Если бы пограничный
слой был одинаков по всему периметру поперечного сечения, то
скошенность профиля скорости в ядре создавала бы условия, при
которых скорость сдвига у внутренней стенки была бы больше,
чем у наружной. Однако вторичные течения, зарождающиеся в по-
граничном слое, приводят к тому, что уже на расстоянии от входа
порядка одного диаметра трубки картина изменяется на противо-
положную: как и в полностью развитом потоке, наибольшая ско-
рость сдвига достигается ближе к наружной стенке.
5.9. Обтекание тел
До сих пор мы говорили о течении жидкости по трубкам или
по системе трубок и при этом установили, что наиболее сложные
картины течения возникают при одновременном действии вязких
и инерционных сил, например в начальном участке трубки. Еще
одним примером ситуации, когда имеет место подобное сочетание
сил, служит обтекание жидкостью твердого тела. Мы рассмотрим
этот пример подробно по двум причинам: во-первых, он помогает
глубже понять общие свойства движущейся жидкости, а во-вто-
рых, многие из возникающих при обтекании тел явлений знакомы
нам из повседневного опыта. 4
Пусть тело погружено в жидкость полностью, так что поверх-
ностные волны не возникают — наличие таких волн исказило бы
всю картину. Для удобства представим также, что тело является
очень длинным цилиндром и жидкость течет перпендикулярно его
оси, поэтому вблизи средней части цилиндра влияние его торцов
пренебрежимо мало, а составляющие скорости, параллельные оси
92 Часть I. Основы механики
цилиндра, отсутствуют. Это означает, что течение в этой области
по своему характеру таково, как будто цилиндр бесконечно длин-
ный; вектор скорости любого элемента жидкости всюду лежит
в плоскости, перпендикулярной оси цилиндра, и зависит от поло-
жения элемента в данной плоскости и от времени, но не от рас-
стояния воль оси цилиндра. В этом случае о потоке говорят, что
он двумерный-, его гораздо легче представить наглядно, чем трех-
мерный поток. Пусть, кроме того, все другие Границы жидкой
среды настолько удалены от цилиндра, что не оказывают влияния
на течение; тогда среду можно считать бесконечной. Наконец, бу-
дем сначала рассматривать только цилиндр с круглым попереч-
ным сечением, так как эта форма удобна и именно для нее вы-
полнены многие теоретические и экспериментальные исследова-
ния. Поскольку важно лишь относительное движение среды и
тела, безразлично, что именно движется — среда (скажем, ветер,
обдувающий здание) или само тело (например, самолет, движу-
щийся в воздухе, или песчинка, опускающаяся на дно лужи).
Обычно легче наблюдать поток, когда тело неподвижно, хотя
иногда более полезным оказывается противоположный подход.
Мы будем изучать такие системы, когда тело имеет постоянную
скорость относительно удаленных от него областей среды. Это
означает, что поток в целом стационарен, если рассматривается
неподвижное тело, и нестационарен, когда тело движется в среде,
неподвижной на достаточно большом удалении от него.
Рассмотрим сначала гипотетическую жидкость, вязкость кото-
рой равна нулю, и временно будем считать, что прилипание на по-
верхности тела отсутствует. Это означает, что пограничный слой
на твердых границах (разд. 5.3) в данном случае не образуется.
Влияние пограничного слоя будет учтено ниже. Предположим, что
цилиндр неподвижен и жидкость на достаточном расстоянии от
него движется вперед с постоянной во всех точках скоростью U
(рис. 5.18). Вблизи тела на пути некоторой части жидкости возни-
кает препятствие, жидкость замедляет свое движение и обтекает
контур тела.
Элементы жидкости, находящиеся в начальный момент доста-
точно далеко от тела, по мере приближения к цилиндру откло-
няются вверх или вниз, как это показано на рис. 5.18, а затем,
оставив тело далеко позади, возвращаются на свои невозмущен-
ные траектории. При стационарном течении элементы жидкости
движутся вдоль линий тока (линий, всюду параллельных вектору
скорости; разд. 4.7), и поэтому рис. 5.18 является также схемати-
ческим изображением картины линий тока (движущийся цилиндр
будет толкать жидкость впереди себя и подтягивать ее за собой;
картина формирующегося при этом потока изображена на
рис. 5.19). Для жидкости «легче» обтекать подобным образом
контур цилиндра (чтобы привести жидкость в движение из состоя-
ния покоя, требуется меньше энергии), чем, например, образовать
Гл. 5. Движение жидкости в трубках и при обтекании тел 93
простирающуюся вдоль оси х бесконечно длинную область, в ко-
торой жидкость полностью неподвижна.
Одна из линий тока разделяет часть жидкости, обтекающую
цилиндр сверху, от той ее части, которая обтекает его снизу.
В точке, где эта линия встречается с цилиндром, скорость должна
быть нулевой, хотя мы не принимали условие прилипания на гра-
нице. Нормальная составляющая скорости равна нулю, так как
Рис. 5.18. Картина течения в пло-
скости, перпендикулярной оси цилинд-
ра. Показаны линии тока (траекто-
рии частиц) для стационарного тече-
ния невязкой жидкости, обтекающей
цилиндр. Скорость жидкости вдали от
цилиндра одинакова во всех точках и
параллельна линии Х'Х. А и D — кри-
тические точки; В и С — точки, где
в отсутствие вязкости скорость дости-
гает максимального значения.
Рис. 5.19. Линии тока при движе-
нии цилиндра в жидкости, неподвиж-
ной на большом расстоянии от него
(напомним, что это течение нестацио-
нарное). При движении жидкость
расталкивается в стороны перед ци-
линдром и смыкается за ним.
жидкость не проходит сквозь твердое тело, а тангенциальная со-
ставляющая нулевая, потому что иначе течение вокруг цилиндра
было бы не симметричным. Указанная точка (на рис. 5.18 она
обозначена через А) называется критической точкой, или точкой
торможения.
Из теоремы Бернулли [уравнение (4.6)] известно, что давле-
ние в точке А больше давления р0 в отдаленных областях жид-
кости, где поток однороден, на величину, равную pU2; это превы-
шение давления может рассматриваться как фактор, порождаю-
щий силу, которая отклоняет элементы жидкости, приближаю-
щиеся к телу. Скорость элемента, который проходит вблизи точки
94 Часть I. Основы механики
А и потому имеет здесь скорость, близкую к нулю, обязательно
должна возрастать, так как вследствие избытка давления он под-
вергается воздействию боковой силы и с ускорением движется к
точке В (или С) цилиндра. Линии тока в этой области располо-
жены теснее, чем вдали от цилиндра, указывая, что скорость здесь
больше, чем U. Действительно, скорость жидкости в точках В и С
(рис. 5.18) равна 2U; все элементы среды ускоряются при обте-
кании передней (носовой) половины цилиндра, пока не достигнут
его верхней (или нижней) части. Согласно теореме Бернулли,
О
давление в точках В и С меньше, чем р0, на величину — р!72.
Когда вязкость отсутствует, картины обтекания задней (кормо-
вой) и передней половин цилиндра симметричны. После прохож-
дения линии, соединяющей точки В и С, элементы жидкости за-
медляются, давление вдоль поверхности цилиндра растет (в точке
D оно вновь превышает р0 на величину рС/2); далеко за ци-
линдром снова устанавливается однородный поток.
Симметричность потока позволяет думать, что результирую-
щая сила, действующая на цилиндр, равна нулю. Можно показать
аналитически, что действительно ни одно тело в однородном по-
токе невязкой жидкости не подвергается действию силы, увлекаю-
щей его в направлении движения потока. Это утверждение, впер-
вые высказанное французским ученым Д’Аламбером в 1752 г., не
согласуется с нашими повседневными наблюдениями: мы знаем,
что все тела, движущиеся в жидкости, испытывают некоторое
противодействие, или сопротивление. Это сопротивление имеется
даже тогда, когда коэффициент вязкости чрезвычайно мал (при
20°С для воздуха р= 0,00181 Н-с-м~2, а для воды 0,1 Н-с-м~2).
Данное противоречие, называемое парадоксом Д'Аламбера, очень
занимало ученых XIX в. Его разрешение состоит в том, что абсо-
лютно все жидкости обладают вязкостью, и, как бы ни было
мало р, вязкость искажает картину потока настолько, что сопро-
тивление становится заметным.
Рассмотрим теперь реальную жидкость, скорость которой
вдали от неподвижного цилиндра диаметром d равна U. В этом
случае число Рейнольдса есть Ud/v, где v = р/р — кинематиче-
ская вязкость. Когда это число велико, инерционные силы преоб-
ладают над вязкими всюду, кроме прилегающих к границам об-
ластей, в которых должно сказываться прилипание. Следователь-
но, можно полагать, что везде, кроме этих областей, поток будет
очень похож на поток невязкой жидкости. Элементы жидкости
будут замедляться по мере приближения к критической точке А
(см. рис. 5.18) и ускоряться при обтекании передней половины ци-
линдра. Около стенки будет существовать область, в которой ско-
рость среды падает до нуля, как это диктуется условием прили-
Гл. 6. Движение жидкости в трубках и при обтекании тел 05
пания, и мы сможем распознать в этой области уже известный
пограничный слой. Он имеет наименьшую толщину непосредствен-
но перед цилиндром (около точки А) и остается довольно тонким
на всем своем протяжении до точек В и С, так как скорость среды
вне слоя увеличивается с расстоянием вдоль поверхности цилинд-
ра. [Согласно уравнению (5.3), толщина пограничного слоя рас-
тет как корень квадратный из расстояния вдоль границы, но
уменьшается как корень квадратный из скорости среды на гра-
нице слоя.] После того как жидкость проходит верхнюю часть
цилиндра, ее скорость начинает падать, павление растет так, как
Рис. 5.20. Профили скорости поперек пограничного слоя для различных то-
чек иа поверхности тела. Показано образование области возвратного течения,
начинающейся за точкой отрыва пограничного слоя.
это описано выше для невязкой жидкости. Это создает градиент
давления, противодействующий потоку в пограничном слое, в ре-
зультате чего поток около стенки, как и в быстро расширяющейся
трубке, останавливается и изменяет направление на противопо-
ложное (рис. 5.20). Начиная с точки, где поток впервые становит-
ся возвратным, область течения, не похожего на течение невязкой
жидкости вследствие существенной роли вязких сил, более не ог-
раничивается тонким пограничным слоем. Наступает отрыв этого
слоя (разд. 5.7). В результате давление, упавшее до значения,
равного давлению в точке отрыва, полностью не восстанавливает-
ся и за цилиндром оказывается гораздо ниже, чем перед ним. Та-
ким образом, тело должно оказывать значительное сопротивле-
ние, сколь бы ни было велико число Рейнольдса1)- Распределение
давления на поверхности цилиндра при числе Рейнольдса 1,1-105
приведено на рис. 5.21. Для сравнения на том же рисунке изобра-
жено соответствующее распределение для невязкой среды. Суще-
ственные различия между этими распределениями давления на
задней половине цилиндра и ответственны за значительную часть
сопротивления. Вклад в сопротивление дает и прямое действие на
стенку тангенциальных вязких напряжений, связанных с высокими
’) Т. е. как бы ни были малы вязкие силы. — Прим перев.
96 Часть 1. Основы механики
скоростями сдвига, соответствующими профилю скорости внутри
тонкого пограничного слоя (рис. 5.20). Однако для течения вокруг
плохо обтекаемого тела типа круглого цилиндра это непосредствен-
ная вязкая компонента сопротивления составляет при числах Рей-
нольдса, превышающих 100, всего около 1% от общего сопротив-
ления.
Рис. 5.21. Изменение дав-
ления, действующего на ци-
линдр, с расстоянием х
вдоль поверхности цилиндра
от критической точки на
передней стенке. Штриховая
линия — результаты измере-
ний при /?е=1,1-105,
сплошная — теоретическая
кривая для невязкой среды.
Различия между этими рас-
пределениями давления на
задней части цилиндра от-
ветственны за большую
часть лобового сопротивле-
ния. [Бэтчелор Дж. Введе-
ние в динамику жидкости
(рис. 5.11.5). Пер. с англ.—
М.: Мир, 1973.]
Характер потока, обтекающего тела, очень сильно зависит от
числа Рейнольдса Re. Для чисел Re, меньших примерно 10, кон-
цепция пограничного слоя — даже для передней половины ци-
линдра— неприменима. Пока мы исключим такие потоки из рас-
смотрения. При значениях Re примерно между 10 и 50 непосред-
Рис. 5.22. Схематическое изображение замкнутых вихрей- за круглым цилинд-
ром и профиль скорости в вязком следе далеко вниз по течению. А и С — кри-
тические точки.
ствеино за цилиндром образуется область, в которой поток разби-
вается на два вихря-, здесь течение похоже на водовороты
(рис. 5.22). Элементы жидкости находятся как бы в ловушке, не-
прерывно кружась, в то время как основной поток проходит мимо.
Движение в этих вихрях, где скорость среды значительно ниже,
чем в основном потоке, поддерживается за счет вязких напряже-
ний, с которыми основной поток действует на границы вихрей.
Чем больше Re (в диапазоне 10—50), тем дальше вихрй вытяги-
ваются вниз по течению,
Гл. 5. Движение жидкости в трубках и при обтекании тел 97
Ниже по потоку от точки, где основной поток вн®вь смыкается,
существует длинная, довольно узкая зона, в которой скорость
жидкости остается более низкой, чем в невозмущенном потоке
(характерный профиль скорости показан на рис. 5.22). Эта зона
носит название спутной струи, или вязкого следа тела. Вязкие
следы существуют независимо от того, есть ли у жидкости свобод-
ная поверхность, но в обычных условиях их удается наблюдать
только на свободной поверхности, хотя там они часто искажаются
из-за движения волн. Например, след корабля виден в основном
благодаря создаваемым им волнам на поверхности моря, хотя эта
поверхность вспенивается также и вследствие сильных возмуще-
ний, генерируемых винтом и существующих далеко за кораблем
в зоне, аналогичной вязкому следу. Когда река обтекает опору
моста, неизбежно возникают волны, но при этом видно, что плы-
вущие по поверхности воды объекты движутся за опорой мед-
леннее, чем основной поток. Другой знакомый пример — след, воз-
никающий за быстро движущимся грузовым автомобилем. Дви-
жение воздуха в следе вперед (связанное с наличием области низ-
кого давления непосредственно за машиной) стремится увлечь за
собой любые небольшие предметы. Такое движение воздуха в сто-
рону грузового автомобиля ощущал каждый, кто стоял на обо-
чине автострады; оно связано с увеличением относительной ско-
рости воздуха на боковых сторонах машины (ср. с рис. 5.18 или
5.19). Конечно, число Рейнольдса в каждом из этих примеров ко-
лоссальное [около 5-10® для опоры шириной 2 м и реки, текущей
не очень быстро — со скоростью 2,5 м-с-1, или для грузового ав-
томобиля шириной 4 м, движущегося со скоростью 15 м-с-1
(около 55 км в час)], но названные основные черты потока присут-
ствуют всегда.
При значениях Це выше примерно 40 симметричный стацио-
нарный поток, изображенный на рис. 5.22, становится неустой-
чивым, наблюдаются медленные колебания следа, причем ампли-
туда колебаний растет с удалением от тела (рис. 5.23). С ростом
Це колебания следа смещаются ближе к цилиндру, и когда Це
приближается к 60, они начинают влиять на вихри за цилиндром.
Оба вихря колеблются вместе из стороны в сторону, в конце каж-
дого полупериода колебаний от них отделяются небольшие вихри,
причем попеременно с разных сторон цилиндра. Поведение следа
на этой стадии весьма примечательно (рис. 5.24). В жидкости
вблизи цилиндра самопроизвольно возникают отдельные вихри,
организованные в виде двух регулярных, расположенных в шах-
матном порядке последовательностей по обе стороны от централь-
ной линии. На рис. 5.24 вихри показаны стрелками. Такая струк-
тура, называемая вихревой дорожкой, перемещается вниз по те-
чению со скоростью, меньшей U, и сохраняется на расстояниях, во
много раз превосходящих диаметр цилиндра. Периодическая кар-
тина следа наблюдается в очень широком диапазоне чисел Рей-
98 Часть I. Основы механики
Рис. 5.23. Фотография колебаний, возникающих в следе за цилиндром. [Бэт-
челор Дж. Введение в динамику жидкости (фото 4.12.6). Пер. с англ. — М.:
Мир, 1973.]
Рис. 5.24. Фотография вихревой дорожки (Ре =102). Стрелками показано
направление вращения вихрей. [Бэтчелор Дж. Введение в динамику жидкости
(фото 4 12.6). Пер. с англ. — М.: Мир, 1973.]
нольдса, примерно до 2500. При больших Re след становится тур-
булентным, в нем трудно различить отдельные вихри, но и тогда
составляющая основу следа периодическая структура сохраняет-
ся— вплоть до чрезвычайно больших значений Re (по меньшей
мере до 105).
С периодическим сходом вихрей связаны пульсации поля дав-
ления в окрестности тела. Это вызывает пульсации силы, дей-
ствующей на тело, которые могут вызвать его вибрацию с часто-
той, равной частоте схода вихрей, если тело закреплено не же-
стко. Вибрации в свою очередь могут увеличить интенсивность ко-
лебаний в жидкости и т. д. Если частота этих колебаний равна
собственной частоте, с которой тело колеблется после слабого воз-
мущения, то может возникнуть резонанс (разд. 8.4), сопровож-
дающийся прогрессивным увеличением размаха колебаний. Такой
резонанс был причиной ряда инженерных катастроф, в которых
строения обрушивались под действием ветра (известным приме-
ром является мост через пролив около Сиэтла, США, рухнувший
вскоре после того, как он был построен в 1940 г.).
Периодический сход вихрей ответствен и за явления меньшего
масштаба, в частности за гудение на ветру телефонных проводов;
он составляет основу механизма создания звуков в ряде музы-
кальных инструментов и органах речи человека. В крови сход вих-
рей, возникающих за препятствиями, такими, как аортальный кла-
пан или стеноз, также создает пульсации давления, которые при
определенных обстоятельствах имеют большое значение (гл. 12).
Частота f схода вихрей за цилиндром может быть связана
только с параметрами, которые определяют картину потока: ско-
Гл. 5. Движение жидкости в трубках и при обтекании тел 99
ростью U, диаметром d, вязкостью р и плотностью р. Простейшая
величина с размерностью частоты, составленная из данных пара-
метров,— это U/d, а более сложные можно сконструировать пу-
тем умножения ее на произвольную функцию, зависящую только
от безразмерного числа Рейнольдса Re = pUd/ц. Отсюда можно
было бы заключить, что безразмерная величина fd/U для тела
данной формы зависит только от Re. На самом же деле эта вели-
чина, называемая числом Струхаля, в широком диапазоне чисел
Рейнольдса, вплоть до значений порядка 103, практически по-
стоянна (0,2 для круглого цилиндра). В этом диапазоне частота
схода вихрей для данного тела, а следовательно, и высота созда-
ваемого звука, увеличивается пропорционально скорости потока.
Именно поэтому в шуме ветра с увеличением его скорости появ-
ляется более высокий тон.
Силу сопротивления, с которой жидкость действует на тело,
например на круглый цилиндр, можно измерить эксперименталь-
но. Чтобы получить формулу, применимую к цилиндрам различ-
ного диаметра при различных скоростях потока, обычно делят
силу сопротивления на величину, которая также имеет размер-
ность силы и может рассматриваться как масштаб для типичных
сил динамического давления, действующих на тело. Эта величина
равна
±-pU2dL, (6.15)
где L — длина цилиндра, которая должна быть достаточно боль-
шой, чтобы поток был близок к двумерному. Отношение суммар-
ной силы сопротивления к величине (5.15) называется коэффи-
циентом лобового сопротивления-.
Сила сопротивления
С D — i • (5.1b)
-i-pt/ML
Для цилиндра с некруглым поперечным сечением коэффициент ло-
бового сопротивления определяется по аналогичной формуле, но
вместо диаметра d используется размер цилиндра в направлении,
перпендикулярном набегающему потоку (рис. 5.25,А). Для ци-
линдра данной формы коэффициент лобового сопротивления, как
и число Струхаля, может зависеть только от Re. Связь между CD
и Re для круглого цилиндра представлена в логарифмическом
масштабе на рис. 5.25, Б.
При очень низких значениях Re коэффициент CD быстро падает
с увеличением Re. Это обусловлено тем, что в диапазоне низких Re
инерционные силы пренебрежимо малы по сравнению с вязкими,
и «типичная» инерционная сила или сила динамического давле-
ния, к которой отнесена сила сопротивления в уравнении (5.16),
на самом деле типична не для всех видов движения. В рассмат-
100 Часть 1. Основы механики
риваемом случае низких Re существует равновесие между вяз-
кими силами и силами, обусловленными давлением; пограничный
слой выделить нельзя, так как присутствие тела искажает свобод-
ное течение в значительной области и во всех направлениях. На са-
мом деле линии тока перед телом и за ним примерно симмет-
ричны, как и при обтекании невязкой жидкостью. Конечно, по-
ток с низкими числами Рейнольдса в отличие от потока невязкой
жидкости сопровождается не нулевым, а, как можно видеть из
рис. 5.25, Б, весьма большим вязким сопротивлением. В предполо-
жении, что инерционные силы очень малы, характер потока при
Рис. 5 25. А. Определение эффективного диаметра d для цилиндра с иекруг-
лым поперечным сечением, величина d входит в уравнение (5 16), задающее коэф-
фициент лобового сопротивления Со Б График зависимости Ср от Re для
круглого цилиндра, построенный в логарифмическом масштабе. Штриховая ли-
ния — теоретическая зависимость для малых Re До Re = 0,2 она совпадает с
экспериментальной кривой. Для значений Re от 200 до 200 000 коэффициент Ср
примерно постоянен. (Fox, McDonald Introduction to fluid mechanics, p. 408,
1973, John Wiley, New York.)
низких числах Рейнольдса может быть детально проанализирован
теоретически. Полученная в результате этого анализа зависимость
коэффициента лобового сопротивления от числа Re изображена
на рис. 5.25, Б штриховой линией. Видно, что для значений Re,
меньших 0,2, теория дает достаточно точные значения коэффи-
циента сопротивления.
Из рис. 5.25, Б можно заключить также, что для значений Re
примерно от 200 до 200 000 коэффициент Cd на удивление постоя-
нен и составляет около 1,2. Следовательно, сила динамического
давления [выражение (5.15)] действительно «типична» для таких
потоков. Тем самым подкрепляется сделанное ранее утверждение
о том, что сопротивление преимущественно порождается именно
изменением распределения давления, обусловленным отрывом по-
граничного слоя, само же вязкое сопротивление чрезвычайно мало.
Резкое уменьшение CD при значениях Re, слегка превышающих
200 000, связано с турбулентностью, развивающейся в пограничном
слое у передней части цилиндра (след становится турбулентным
при гораздо меньших Re). Таким образом,турбулентность, вместо
того чтобы увеличить сопротивление, чего естественно было бы
ожидать, зная о неизбежном увеличении вязкой диссипации в тур-
булентном потоке, приводит к его падению. Причина этого явле-
Гл. 5 Движение жидкости в трубках и при обтекании тел 101
ния состоит в том, что турбулентность в пограничном слое неяс-
ным пока образом затягивает его отрыв. В результате точка от-
рыва пограничного слоя смещается по поверхности цилиндра на-
зад (рис. 5.26), что в свою очередь приводит к уменьшению пло-
щади, иа которую действует пониженное давление в области от-
рыва, и результирующая разность сил давления перед телом и за
ним также уменьшается. Хотя этот круг вопросов динамики жид-
кости достаточно интересен, мы не останавливаемся на нем далее,
так как он не имеет отношения к кровообращению, где макси-
мальные числа Рейнольдса, рассчитанные так, как их вычисляют
для течения в трубках, не превышают 10.000.
Ламинарный Т урбулентный
Рис. 5 26. Схема, показывающая, насколько турбулентность в пограничном слое
затягивает его отрыв, уменьшая таким образом лобовое сопротивление тела.
До сих пор мы рассматривали только двумерные тела, по-
скольку картину их обтекания сравнительно легко наблюдать. Во
многих же повседневных случаях обтекания как само тело, так и
поток вокруг него трехмерны. Оказывается, однако, что основные
свойства потоков вокруг трехмерных тел аналогичны описанным
выше, хотя необходимо принимать во внимание и некоторые отли-
чия. Из трехмерных систем наиболее изучено обтекание шара,
поскольку поток вокруг него удобнее создавать в эксперименте и
легче анализировать, чем поток вокруг тела любой иной формы.
Приведем несколько примеров часто встречающихся в повседнев-
ной жизни тел примерно сферической формы: большой шар, под-
нимающийся в воздухе (большое Re), песчинка, погружающаяся
на дно водоема (малое Re), шарик подшипника в масле (малое
Re), мяч, летящий в воздухе (довольно большое Re), и т. д. Во
всех этих случаях число Рейнольдса, как и прежде, подсчиты-
вается по диаметру сферы d, относительной скорости U жидкости
вдали от шара и кинематической вязкости v (т. е. Re=Ud/v).
Коэффициент лобового сопротивления по-прежнему определяется
как суммарное сопротивление, деленное на «типичную» силу дина-
мического давления, которая для трехмерного тела обычно берет-
ся в виде произведения ypf/2 на площадь сечения тела f-j-nd8
в случае шара). Таким образом,
„ Сила сопротивления
Ср— ; j
2- pU2 4- nd»
2 4
102 Часть I. Основы механики
График зависимости CD от Re представлен на рис. 5.27 и, как
легко видеть, аналогичен приведенному ранее графику для ци-
линдра. Теоретический анализ показывает, что при низких значе-
ниях Re Со = 2^/Re\ соответствующая этой зависимости прямая
также приведена на рис. 5.27 (штриховая линия). При более вы-
соких числах Рейнольдса (вплоть до примерно 130) за сферой об-
разуются вихри, занимающие в пространстве определенное поло-
жение, однако они совсем не похожи на вихри при обтекании ци-
линдра — прямые и вытянутые, вдоль него. Вихри за сферой имеют
Рис. 5.27. График зависимости Сс от
Re для сферы; качественно он аналогичен
соответствующему графику для цилиндра.
Штриховая линия — теоретическая зависи-
мость для малых Re. (Fox, McDonald. In-
troduction to fluid mechanics, p. 406, 1973,
John Wiley, New York.)
форму колец, поскольку все течение симметрично относительно
оси, параллельной вектору скорости набегающего потока и прохо-
дящей через центр сферы (примером кольцевого вихря может
служить хорошо знакомое кольцо дыма — видно, как по мере его
движения частицы вращаются вокруг кольцеобразного ядра). При
еще более высоких числах Рейнольдса кольцевые вихри отры-
ваются от сферы и создают сложную структуру течения в следе.
Это уже не упорядоченное движение, подобное двумерной вихре-
вой дорожке, поскольку, хотя вихрь и остается замкнутым в коль-
цо, само кольцо сильно искажается из-за неодновременности от-
рыва его частей от разных точек сферы. Несмотря на кажущуюся
хаотичность потока за сферой, в возмущениях силы, с которой
жидкость действует на тело, преобладают колебания с определен-
ной частотой. Иногда они порождают звуки указанной частоты и
приводят к колебаниям тела.
Глава 6
АНАЛИЗ РАЗМЕРНОСТЕЙ
Выше мы уже не раз использовали принцип размерностной од-
нородности для проверки правильности математического представ-
ления того или иного физического закона. В данной главе этот
принцип будет рассмотрен более подробно, что позволит выявить
всю его мощь. Кроме того, мы покажем, как анализ размерностей
может быть использован при выводе физических законов.
В большинстве случаев не составляет особого труда преобразо-
вать математическое выражение закона к безразмерному виду;
даже такая перегруппировка сомножителей часто может кое-что
сказать нам о соотношениях между физическими величинами, вхо-
дящими в закон. Вернемся, например, к закону Пуазейля (гл. 5),
который в размерном виде записывается так:
' nd4
Если определить среднюю скорость О с помощью равенства
q = 2 nd2U,
подставить Q в выражение для Др и разделить обе его части на
произведение pU2, то мы получим аналитическую запись закона
Пуазейля в безразмерном виде
или, воспользовавшись выражением для числа Рейнольдса Re,
^=32(А)/<ад.
Запись закона Пуазейля в такой форме подводит нас к поня-
тию подобия. Сравним, например, условия течения жидкостей раз-
личающейся вязкости в двух трубках разного диаметра при раз-
личающихся расходах. Условия течения в них будут «подобными»,
если каждая из комбинаций величин — (Др/рС2), (L/d) и Re —
окажется для обоих случаев одинаковой. Если хотя бы одна из
этих комбинаций для разных потоков различается, течения не по-
добны.
104 Часть 1. Основы механики
6.1. Подобие и моделирование
Концепция подобия открывает большие возможности для экс-
периментальных исследований, так как обосновывает подход, при
котором ставятся и анализируются эксперименты на построенных
в определенном масштабе (т. е. уменьшенных или увеличенных)
Моделях изучаемой системы.
Если реальная система слишком велика или мала, мы можем,
Ьыбрай соответствующие масштабы величин, изучать динамически
подобное состояние на модели приемлемых размеров. При анализе
потока в трубах, например, можно, основываясь на принципе по-
добия, использовать вместо очень больших труб трубы меньшего
размера, а жидкости, которые вызывают сильную коррозию, заме-
нить более инертными средами соответствующей вязкости и плот-
ности— воздухом, водой или маслом.
Аэродинамические свойства новой марки самолета не обяза-
тельно испытывать в полете: можно провести испытания неболь-
ших моделей в аэродинамических трубах. Регулируя скорость воз-
духа, обдувающего модель в трубе, и ориентацию модели, можно
предсказать, как самолет будет реагировать на подобные дина-
мические изменения в условиях полета. С другой стороны, микро-
сосуды имеют такие небольшие размеры, что подлежащие изме-
рению величины (например, перепад давления) слишком малы, и
вследствие этого возникают технические трудности при их изме-
рении. Поэтому часто строят модели микроциркуляции в увели-
ченном масштабе. Большая часть данных о механике микроцир-
куляции получена при изучении именно таких моделей (гл. 13).
Однако, применяя такой модельный подход для решения ка-
кой-либо задачи, мы должны помнить, что он окажется несостоя-
тельным, если при изменении масштаба одного из параметров
сверх приемлемых пределов для описания состояния модели пона-
добятся дополнительные законы. Например, легко представить си-
туацию, когда в ходе модельного эксперимента градиент давления
в потоке жидкости увеличивается до такой степени, что ее следует
рассматривать как сжимаемую, и упрощения, связанные с несжи-
маемостью и верные для реального объекта, становятся недействи-
тельными.
Анализ размерностей и безразмерные комбинации величин
можно использовать и при изучении сложных систем, чтобы выяс-
нить, какие комбинации будут оставаться постоянными на протя-
жении всего времени и в каких пределах будут меняться другие
переменные. Это существенно облегчит представление результа-
тов, поскольку последние могут быть выражены наилучшим обра-
зом в виде графиков зависимостей параметров, сгруппированных
именно так, чтобы получились безразмерные величины. При этом
можно выявить реакцию данной системы на изменения тех или
иных комбинаций переменных безотносительно к единицам изме-
Гл. 6. Анализ размерностей 105
рения, в которых эти переменные выражены. Подобный способ
представления ценен, в частности, при сравнении результатов, по-
лученных разными исследователями.
6.2. Некоторые примеры выбора масштабов в биологических
системах
Описанные выше примеры моделирования касались только ме-
ханических процессов; однако во многих случаях можно приме-
нить указанный подход к физиологическим свойствам животных
в целом. Например, можно сопоставить такие свойства, как
Рис. 6.1. Зависимость между площадью поверхности и массой тела у позвоноч-
ных. Сплошная прямая соответствует зависимости между площадью поверхности
и массой для шара плотностью 1 г-см-3. (Лайтфут Э. Явления переноса в жи-
вых системах. Пер. с англ. — М: Мир, 1977.)
интенсивность обмена веществ, анатомические параметры и хими-
ческие процессы, протекающие в организме, в зависимости, ска-
жем, от размера тела. Подобные корреляции иногда выявляют
удивительно тесную взаимозависимость между различными свой-
ствами, а зачастую и исключения из общих правил, позволяющие
в свою очередь высказать новые соображения.
Простой и до некоторой степени неожиданной является связь
между массой тела и его поверхностью и формой. Оказывается,
что для позвоночных (в очень большом диапазоне масс) площадь
поверхности тела близка к удвоенной площади сферы, имеющей тот
же объем (рис. 6.1). У млекопитающих отношение объема отдель-
ных органов к объему тела также оказывается удивительно по-
106 Часть I. Основы механики
Рис. 6.2. Влияние pH на уменьшение содержания кислорода в крови (эффект
Бора) у мелких животных больше, чем у крупных.
стоянным. Так, объем сердца составляет примерно от 0,5 до 0,6%
объема тела. Сердце скаковых лошадей и борзых собак, однако,
оказывается исключительно большим, что, впрочем, и неудиви-
тельно, если учесть те требования, которые предъявляются к сер-
дечно-сосудистой системе этих животных.
У млекопитающих проявляются две формы зависимости свойств
крови от размера тела. Во-первых, гемоглобин мелких животных
при данном pH быстрее высвобождает кислород, что способствует
поддержанию у таких животных высокой скорости обменных про-
цессов. Во-вторых, эффект высвобождения кислорода при умень-
шении pH крови [или увеличении р(СОг)] — эффект Бора — выра-
жен тем сильнее, чем мельче животное (смотри рис. 6.2). Из
этого правила есть два интересных исключения. Из рис. 6.2 видно,
что эффект Бора чревычайно сильно выражен у лошади — живот-
ного, которому приходится преодолевать большие расстояния.
С другой стороны, исключение составляет и чихуа-хуа — сравни-
тельно новая порода собак, выведенная исключительно по экс-
терьеру для декоративных целей; у нее эффект Бора выражен
слабо. Это следствие быстрого селективного выведения породы,
по-видимому, является определенным недостатком.
Г л. 6 Анализ размерностей 107
6.3. Метод получения размерностно однородных зависимостей
между переменными
Хотя во многих случаях не составляет особого труда устано-
вить, какие именно переменные нужны для описания того или
иного процесса, до тех пор, пока неизвестны законы, управляющие
самим процессом, связь между переменными остается неясной.
Отыскать ее часто помогает анализ размерностей. Мы уже знаем,
что любое уравнение, адекватно описывающее некоторый процесс,
должно быть размерностно однородным, а потому необходимо
сгруппировать выделенные нами переменные таким образом, чтобы
выполнялся указанный принцип. Способ такой группировки иногда
называется методом показателей-, его применение иллюстрируется
ниже на двух простых примерах.
В качестве первого примера рассмотрим еще раз задачу о те-
чении в прямой трубке. Мы знаем, что продольный градиент дав-
ления (&p/L) зависит от диаметра трубки d, объемного расхода Q
и вязкости жидкости ц. Если ограничиться течением вдали от на-
чала трубки, то, как нам известно, ускорение элементов жидкости
будет равно нулю. Таким образом, в этом случае мы не учиты-
ваем инерционность среды, и ее плотность р не играет роли.
Итак, можно сказать, что градиент давления есть функция
вязкости, объемного расхода и диаметра трубки:
= Ж Q, d).
Предположим, что градиент давления является однозначной
функцией этих переменных и что, кроме того, переменные связаны
между собой следующим образом:
где tn, п и р — показатели степени, в которую должны быть воз-
ведены эти переменные, a k — безразмерная постоянная. В тер-
минах размерностей из этой связи получим
[^]МиГт<.
Но [p]^[M/LT], [QWL7T], ИМЦ, [ Ьр/L] = [M/L2T2].
Сравнивая показатели степени при каждой из размерностей
(М, L и Т) в обеих частях уравнения, приходим к выводу, что
для однородности требуется
для массы 1 = т,
для длины — 2 — — т -j- Зп -j- р,
для времени — 2 = — т — п.
108 Часть I. Основы механики
Решая систему трех этих уравнений, получаем m = 1, п = 1,
р = —4, или
[Др/L] = [р] [Q]/[rf]4-
Таким образом, единственная размерностно однородная связь
между переменными имеет вид
Ар и<2
L
ИЛИ
Др___ь и<2
L ~R d* '
Именно таким и должен быть вид реальной зависимости. Легко
опознать в этой записи закон Пуазейля, для которого, согласно
теоретическим расчетам, k = 128/л.
В качестве второго простого примера рассмотрим обтекание
шара потоком с очень низкой скоростью (гл. 5). Как и раньше,
мы можем пренебречь инерцией и рассматривать только вязкие
эффекты. Сила F, с которой среда действует на шар, зависит оз
размера шара (его радиуса а), скорости и потока, обтекающегс
шар, и вязкости жидкости р.
Таким образом,
F — f(a, и, р).
Учитывая размерности переменных, приходим к зависимости
[F] [а]т [«]" [р]р.
Но
[F] = [ML/T2],
[«ML/T],
[p]^[M/LTJ.
Сравнивая показатели степени при каждой из размерностей (М, L
и Т), получаем
1 = Р>
1 = m + п — р,
—2 = — п — р.
Таким образом, m = 1, п = 1, р = 1, а потому
F ос иар.
Рассмотрим теперь частный случай шара, медленно опускаю
щегося в вязкой жидкости под действием силы тяжести (иапри
мер, мелкую песчинку, медленно оседающую на дно). Действую
Щая на шар сила пропорциональна его объему и разности плот
Гл. 6. Анализ размерностей 109
ностей (рш — р») шара и жидкости
F ос (рш — рж) ga3.
Таким образом,
k (Рш — Рж) ga3 = нац,
или
и = k (рш — рж) gra2/p. (6.1)
В действительности это не что иное, как закон Стокса для мед-
ленного обтекания тела сферической формы, падающего в вязкой
жидкости. Можно показать, что значение константы k в этом
2 1)
уравнении равно -д '.
') Дальнейшие сведения о теории размерностей и примеры ее использования
читатель может найти в книге: Седов Л. И. Методы подобия и размерности в ме-
ханике.— М.: Наука, 1977. — Прим. ред.
Глава 7
МЕХАНИКА ТВЕРДОГО ТЕЛА
И СВОЙСТВА СТЕНОК КРОВЕНОСНЫХ СОСУДОВ
Стенки кровеносных сосудов упруги и могут изменять свои
размеры и форму под влиянием различных приложенных к ним
сил. К этим силам относятся как давление и напряжение сдвига,
с которыми действует на сосуды кровь, так и воздействия со сто-
роны окружающей ткани («эффект прикрепления»). В этой главе
мы в общих чертах изложим основные принципы механики дефор-
мируемых твердых тел и рассмотрим их применимость к стенкам
кровеносных сосудов. Разумеется, основу механики твердых тел
составляют законы Ньютона движения частиц. Как и жидкость,
твердое тело можно рассматривать как совокупность большого
числа малых элементов, к каждому из которых применимы эти
законы. Силы, действующие на подобные элементы, состоят из
дальнодействующих массовых (или объемных) сил и близкодейст-
вующих сил напряжения; механика твердых тел и жидкостей раз-
личается именно характером связи между напряжениями дефор-
мациями.
7.1. Свойство упругости
Начнем с нескольких определений. Упругим, является такой ма-
териал, который деформируется под действием приложенной к
нему силы, но принимает исходную форму (причем без какой-
либо диссипации энергии), когда снимается нагрузка. Это озна-
чает, что его молекулы возвращаются к своим исходным положе-
ниям. Первый шаг в изучении явления упругости сделал в 1678 г.
Роберт Гук (английский астроном и физик). Подвешивая к одно-
родной цилиндрической проволоке с исходной длиной /0 и пло-
щадью поперечного сечения Ао, закрепленной одним концом не-
подвижно, различные грузы W (рис. 7.1, А) и измеряя растяже-
ние проволоки Гук обнаружил, что в широком диапазоне при-
кладываемых нагрузок оно пропорционально весу груза («ut ten-
sio sic vis») (рис. 7.1,5). Более поздние эксперименты, выполнен-
ные, в частности, Томасом Юнгом (1773—1829 гг.; английский
врач, физик и египтолог), позволили уточнить формулировку этого
закона (называемого поныне законом Гука), и он стал применим
к изготовленным из одного материала проволокам с различной
площадью поперечного сечения. Было найдено, что удлинение,
Гл. 7. Механика твердого тела 111
Рис. 7.1. А. Эксперимент Гука с растяжением проволоки. Растягивается метал-
лическая проволока, один конец которой неподвижно закреплен. Без груза ее
длина равна 10, а когда подвешивается груз W, проволока растягивается на ве-
личину С — мысленно проведенное сечение, используемое для определения на-
тяжения Т в проволоке, равного силе, с которой верхняя часть проволоки дейст-
вует на нижнюю (или наоборот). Условие баланса сил, действующих на ниж-
нюю часть, показывает, что Т должно быть равно И7. Б. Результаты эксперимен-
та, подобного представленному на рис. А: растяжение пропорционально весу И7
до тех пор, пока последний не слишком велик, б. Результаты полного экспери-
мента с растяжением проволоки, представленные в виде графика зависимости
напряжения Т/Ao (До — начальная площадь поперечного сечения проволоки) от
деформации Г/l. Пока напряжение меньше Sa, деформация прямо пропорциональ-
на напряжению. При больших напряжениях наблюдается пластическое течение,
т. е. деформация не уменьшается до нуля при снятии нагрузки (штриховая пря-
мая). По достижении максимально допустимого напряжения SB течение продол-
жается, даже если напряжение падает ниже Sb. В точке С происходит разрыв
проволоки. Г. Схематическое изображение проволоки, в которой началось обра-
зование «шеек».
приходящееся на единицу длины Г/10 (эта величина обычно назы-
вается продольной деформацией е), пропорционально натяжению Т
в проволоке, деленному на начальную площадь поперечного сече-
ния Ло- Чтобы определить натяжение в проволоке, мысленно ра-
зобьем ее сечением С на две части, как это показано на рис. 7.1,4,
112 Часть I. Основы механики
Суммарная направленная вниз сила, с которой часть проволоки,
лежащая ниже сечения, действует на часть проволоки, располо-
женную выше С, и называется натяжением Т. Согласно третьему
закону Ньютона, направленная вверх сила, с которой верхняя
часть проволоки действует на ее нижнюю часть, по абсолютной
величине также равна Т. Кроме того, если рассмотреть баланс
сил, действующих на нижнюю часть проволоки, то мы увидим, что
Т должно быть равно сумме веса груза W, подвешенного к ниж-
нему концу проволоки, и собственного веса нижней части про-
волоки. В большинстве подобных экспериментов можно прене-
бречь весом проволоки по сравнению с W, так что натяжение Т
оказывается равным W. Этот результат остается справедливым
при любом положении сечения С, и поэтому мы приходим к вы-
воду, что натяжение Т одинаково во всех точках по длине про-
волоки. Как можно видеть, натяжение, приходящееся на единич-
ную площадь, Т/Ао, имеет смысл напряжения (разд. 4.1); в дей-
ствительности оно и является продольным нормальным напряже-
нием, существующим в любом поперечном сечении проволоки.
Если бы эксперименты Гука с металлической проволокой про-
водились для максимально широкого диапазона прикладываемых
нагрузок, то график зависимости напряжения от деформации имел
бы вид, показанный на рис. 7.1, В (предполагается, что экспери-
мент проводится статически, т. е. в момент измерения деформации
проволока неподвижна). Первая часть кривой вплоть до точки А
линейна. Это означает, что, пока прикладываемые напряжения не
превышают Sa, остается справедливым закон Гука, и, кроме того,
деформация уменьшается практически до нуля, когда до нуля
уменьшается напряжение. В этом диапазоне напряжений материал
является линейно-упругим.
Если напряжение превышает Sa, кривая «напряжение — дефор-
мация» становится нелинейной. При уменьшении напряжения до
нуля наблюдается необратимая остаточная деформация1); иными
словами, деформация не возвращается к нулю (штриховая линия
на рис. 7.1, В). Подобное поведение указывает, что произошло
некоторое течение материала. Напряжение Sa, выше которого
наблюдается течение, называют пределом текучести, а само тече-
ние материала — пластической деформацией. Пластические дефор-
мации необратимы; это означает, что они сопровождаются частич-
ным разрушением структуры молекулярной решетки исследуемого
материала.
При дальнейшем увеличении напряжения пластическая дефор-
мация продолжается до тех пор, пока материал не разрушится
(точка, отмеченная на графике буквой С). После того как достиг-
’) Для ряда материалов остаточных деформаций нет и за пределами линей-
ного участка кривой напряжение — деформация (см. рис. 7.5); материал в этом
рлучае называют нелинейно-упругим. — Прим, ред.
Гл. 7. Механика твердого тела 113
нуто максимальное значение приложенного напряжения Sb, мате-
риал продолжает течь, даже если напряжение несколько умень-
шить. Дело в том, что в этом диапазоне деформаций наблюдается
быстрое уменьшение площади поперечного сечения некоторых уча-
стков проволоки (образование «шеек»; рис. 7.1,Г), так что напря-
жение, измеренное как натяжение, деленное на площадь исход-
ного поперечного сечения, уже не отражает достаточно хорошо
истинное напряжение в этих местах, т. е. натяжение, деленное на
истинную площадь сечения образца. Истинное напряжение в ме-
стах, где площадь минимальна, продолжает расти при движении
точки на графике от В к С.
Приведенное выше описание чрезвычайно упрощено. В дей-
ствительности, если напряжение, лежащее в диапазоне упругости
(т. е. меньшее, чем Sa), поддерживать очень долго (для стальных
проволок несколько дней), то окажется, что деформация очень
медленно нарастает. Это означает, что происходит медленное те-
чение материала. Если затем снять нагрузку, то деформация не
уменьшится строго до нуля, а будет составлять небольшую вели-
чину, определяемую интенсивностью имевшего место течения. По-
добное явление носит название ползучести. Оно отличается от
пластического течения тем, что является обратимым: если дли-
тельное время прикладывать сжимающие напряжения, будет на-
блюдаться обратное медленное течение, и при определенных усло-
виях можно добиться того, что деформация в отсутствие напря-
жения снова станет нулевой. Ползучесть не приводит к необра-
тимым разрушениям молекулярной структуры материала, как это
имеет место при пластическом течении. В случае ползучести от-
носительное движение соседних молекул напоминает движение
молекул вязкой жидкости и направление его можно изменить пу-
тем обращения приложенного напряжения. Как и течение вязкой
жидкости, ползучесть сопровождается диссипацией механической
энергии. С ползучестью тесно связано явление, наблюдающееся
в том случае, когда длительное время поддерживают не напря-
жение, а постоянное удлинение: оказывается, что необходимое
для этого напряжение медленно уменьшается от своего первона-
чального, обусловленного упругостью уровня. Указанное явление
носит название релаксации напряжения. Ползучесть и релаксация
напряжения являются примерами того, что называется вязкоуп-
ругим поведением. Тот факт, что каждый материал в той или иной
степени является вязкоупругим, означает, что идеально упругих
материалов в том смысле, в каком было дано определение этому
понятию, не существует. Однако для очень многих материалов,
таких, как сталь, камень или бетон, деформация ползучести, ко-
торая наблюдается за представляющие интерес промежутки вре-
мени, пренебрежимо мала, и их можно считать чисто упругими
в широком диапазоне прикладываемых напряжений. Как мы уви-
дим далее, стенки кровеносных сосудов являются вязкоупругими,
114 Часть I. Основы механики
и это оказывает существенное влияние на механику течения крови
в сосудах.
Итак, проводя эксперимент, аналогичный описанному выше,
можно ожидать, что в определенном диапазоне напряжений
(и времен) кривая напряжение — деформация будет линейной.
Для этого диапазона мы можем записать следующее соотношение:
Т“ = £-Г- (7Л)
to
Коэффициент пропорциональности Е называется модулем упруго-
сти Юнга исследуемого материала. Модуль Е является количе-
Рис. 7.2, Уменьшение радиуса проволоки (—а')
при ее продольном растяжении (/'). Связь между
этими величинами задается уравнением (7.2).
ственной характеристикой материала, и для простейших материа-
лов его величина не зависит от того, каким образом приклады-
ваются напряжения; Е измеряется в тех же единицах, что и на-
пряжение, т. е. в Н-м-2. Отметим, что уравнение (7.1) может с
таким же успехом применяться к деформациям цилиндрических
твердых тел и под действием продольных сжимающих напряже-
ний (подобных напряжениям в большой берцовой кости вертикаль-
но стоящего человека или напряжениям в гвозде, по которому
ударяет молоток); в этом случае и Т и I' отрицательны.
ХВ ходе описанного эксперимента для большинства материалов
одновременно с продольным растяжением происходит уменьшение
размера образца в поперечном направлении. Например, если ис-
ходный радиус круглой проволоки составлял ао, то после растя-
жения он может оказаться равным а0 + а', где величина а' обыч-
но отрицательна (рис. 7.2; мы будем считать, что уменьшение
длины есть отрицательное растяжение). Для большинства простых
материалов поперечная деформация а'/а0 прямо пропорцио-
нальна продольной:
Гл. 7. Механика твердого тела 115
Здесь о — еще одна константа, характеризующая материал и на-
званная коэффициентом Пуассона (по имени французского есте-
ствоиспытателя XIX в.). Коэффициент Пуассона является безраз-
мерной и обычно положительной величиной. Если объем круглого
цилиндрического элемента, представленного на рис. 7.2, не ме-
няется в ходе малой деформации, то коэффициент о равен */2.
Действительно, новый объем я(а0 + а')2(1о + /') должен равняться
старому ла^0, а для малых а'/ай и 1'/1й это требование сводится
к уравнению
а' 1 Г
а0 2 Iq
представляющему собой уравнение (7.2), в котором <т = -|-. В об-
щем случае, если материал несжимаем, его коэффициент Пуассона
равен ’/2. Ни для одного материала он не может быть больше */2,
Л Б В
l/П/ со---- -л
Рис. 7.3. А. Напряжение сдвига Б. Напряжение изгиба б. Напряжение кручения,
так как это означало бы увеличение объема цилиндра под дей-
ствием сжимающих напряжений, приложенных к его концам.
Почти все биологические материалы и, в частности, стенки кро-
веносных сосудов практически несжимаемы.
Уравнения (7.1) и (7.2) описывают реакции твердого тела на
деформирующее напряжение определенного типа, а именно на
продольное растяжение или сжатие, и только до тех пор, пока
напряжения лежат в диапазоне, в котором реакция является уп-
ругой. Как и в случае уравнений, описывающих связь между на-
пряжением и скоростью деформации в жидкости, уравнения (7.1)
и (7.2) легко обобщить на случай малых трехмерных деформа-
ций.
Помимо продольного растяжения или сжатия и связанных с
ними поперечных деформаций (рис. 7.2) образец может подвер-
гаться напряжениям сдвига, которые стремятся вызвать скольже-
ние слоев материала один относительно другого (рис. 7.3,Л). Об-
разец может также подвергаться изгибу (рис. 7.3,Б), кручению
(рис. 7.3, В) или произвольной комбинации деформирующих воз-
действий. В каждом случае деформации будут прямо пропорцио-
нальны приложенным напряжениям и коэффициент пропорцио-
нальности может быть выражен через модуль Юнга Е и коэффи-
циент Пуассона о материала. Таким образом, для упругих твер-
дых тел описанного выше типа (т. е. подобных металлам) Е и о
полностью определяют реакции образца на приложенные напря-
жения.
116 Часть 1. Основы механики
7.2. Свойства стенок кровеносных сосудов
Посмотрим теперь, подчиняются ли материалы, образующие
стенки артериального сосуда, закону Гука (напряжение пропор-
ционально деформации), и если нет, то как велики отклонения от
этого закона. В классической теории упругости предполагается,
что материал достаточно однороден и изотропен. Это означает, что
измеряемые значения Е и о одинаковы для разных точек мате-
риала (однородность) и не зависят от направления, в котором
приложено напряжение (изотропность). Иными словами, в каком
бы направлении и из какого бы участка ни вырезался маленький
образец материала, экспериментально измеряемые значения £ и о
одинаковы. Однако биологические ткани и, в частности, стенки кро-
веносных сосудов не являются ни однородными, ни изотропными.
Обычно они состоят из нескольких разных материалов, связанных
друг с другом сложным образом. Структура стенок сосудов опи-
сана более детально в гл. 12—15. Ниже мы приведем данные о
свойствах наиболее важных материалов, составляющих эти стенки.
Стенки артерий и вен примерно на 70% состоят из воды, ко-
торая не является упругим материалом и проявляет упругие
свойства только при всестороннем сжатии. Однако остальная
часть стенки представляет собой систему переплетенных волокон,
которые и обладают упругими свойствами. Упругие свойства сте-
нок сосуда как целого определяются волокнами трех типов — эла-
стическими, коллагеновыми и гладкомышечными. Эластин являет-
ся резиноподобным материалом, модуль Юнга которого равен при-
мерно 3-105 Н-м~2. График зависимости натяжение — деформация
для волокон чистого эластина представлен на рис. 7.4, Л. Колла-
ген гораздо более жесткий, чем эластин, его модуль Юнга равен
примерно 108 Н-м~2. График зависимости напряжение — деформа-
ция для коллагена представлен на рис. 7.4, Б. Хотя эластин на-
столько податлив, что может быть растянут до длины, вдвое боль-
шей исходной, напряжение, при котором наступает его разрыв
(это напряжение называется пределом прочности при растяже-
нии), составляет менее 5% от аналогичного показателя для кол-
лагена. Модуль Юнга для гладких мышц примерно такой же, как
у эластина. Значения его, определяемые в эксперименте, зависят
от уровня физиологической активности и могут меняться от значе-
ния 1 -105 Н-м~2, характерного для полностью расслабленных
гладких мышц, до примерно 2-106 Н-м~2 в активном состоянии.
Каково значение модуля Юнга гладких мышц in vivo, сказать
трудно, так как степень активности их при этом обычно неиз-
вестна. В крупных артериях (по крайней мере у собак) на долю
коллагена и эластина приходится около 50% сухого веса. В груд-
ной аорте соотношение между эластином и коллагеном равно
примерно 1,5, в то время как для других артерий этот показатель
близок к 0,5, а для вен уменьшается до 0,3. Аналогичные зиаче-
Гл. 7. Механика твердого тела 117
Рис. 7.4. А. Графики зависимостей натяжения от деформации для пяти свеже-
приготовленных волокон эластина, находящихся в растворе Рингера — Локка при
37 °C. Для каждого волокна приведены экспериментальные точки (X), линии рег-
рессии (—) и общая для всех волокон линия регрессии (....). По оси ординат
отложено натяжение в отдельном волокне, по оси абсцисс — деформация. [Car-
ton, Dainauskas, Clark (1962). Elastic properties of single elastic fibres, p. 549,
J. appl. Physiol. 17.1 Б. График зависимости напряжения от деформации для
волокон коллагена. [Benedict, Walker, Harris, (1968). Stress — strain characteris-
tics and tensile strength of unembalmed human tendon, p. 58, J. Biomech. 1.]
ния характерны и для сосудов человека, хотя переход к преобла-
Данию коллагена над эластином происходит дальше к перифе-
рии — в нижней части брюшной аорты.
Содержание гладких мышц в стенке также зависит от положе-
ния артерии: чем она дальше к периферии, тем больше приходя-
щаяся на них доля сухого веса стенки. В грудной аорте доля глад-
ких мышц составляет 25%, а в мельчайших артериях и артерио-
лах она достигает 60%. Структура вен в принципе аналогична
структуре артерий, но стенки их более тонкие и содержат меньше
эластина.
Поведение кровеносного сосуда как целого определяется не
только упругими свойствами его составляющих, но и геометриче-
скими параметрами. В этом смысле наиболее важными величи-
нами являются внутренний диаметр сосуда d и толщина стенки й,
которые, естественно, различны для разных сосудов. Характерные
118 Часть I. Основы механики
значения этих параметров для некоторых сосудов, измеренные in
vivo у собак (мельчайшие сосуды, измерялись у других млекопи-
тающих), представлены в таблице, помещенной в начале книги.
Здесь же приведено большое количество других данных, имею-
щих отношение к механическим явлениям, которые наблюдаются
в системе кровообращения; эти данные будут детально обсуж-
даться в следующих главах. Как видно из таблицы, величины h
и d уменьшаются к периферии. Интересно отметить, однако, что
отношение толщины стенки к диаметру в артериях большого круга
меняется не очень сильно и составляет величину около 0,07. Этот
параметр возрастает в артериолах, стенки которых содержат
больше мышц, но снова уменьшается в капиллярах и гораздо
ниже в венах и легочных артериях, чем в артериях большого
круга, имеющих близкий диаметр.
Перемещения стенок кровеносного сосуда, обусловленные пе-
ременными силами, с которыми кровь действует на внутреннюю
поверхность сосуда, сильно ограничены механическими воздей-
ствиями со стороны ткани, по которой сосуд проходит и к которой
он прикреплен. Этот «эффект прикрепления» в большой мере за-
висит от механических свойств окружающей ткани. Особенно су-
ществен он для капилляров: их механические свойства почти пол-
ностью обусловлены свойствами окружающей среды, так как сама
их стенка состоит лишь из одного слоя эндотелиальных клеток.
Поскольку стенка кровеносного сосуда неоднородна по составу
и организована таким образом, что вряд ли является изотропной,
к ней нельзя прямо применить классическую теорию упругости
и определить единственный модуль Юнга. Однако описать реак-
цию стенки на различные прикладываемые напряжения все же
необходимо, и для этого была бы очень удобна некая величина,
аналогичная модулю Юнга. Чтобы найти эту величину, измеряют
деформацию стенки как целого в ответ на известное приложенное
напряжение и вычисляют значение модуля Юнга, которым обла-
дал бы материал стенки, если бы он был однородным и изотроп-
ным. При этом мысленно объединяют в единое целое все состав-
ляющие стенки и получают эффективный модуль Юнга.
На изолированных артериях и венах обычно проводят экспе-
рименты двух типов. В ходе одних сосуд данной длины растяги-
вают (раздувают), задавая различные трансмуральные давления,
и измеряют его диаметр (длина сосуда сохраняется постоянной).
Как мы увидим далее (разд. 7.3), давление в кровеносном сосуде
создает в стенке растягивающее напряжение, направленное по ка-
сательной к окружности, которое называют окружным или азиму-
тальным (рис. 7.10,5). При этом возникает и продольное напря-
жение. Из окружного напряжения и соответствующего ему изме-
ненного значения диаметра на основании закона Гука [уравнение
(7.1)] и вычисляется величина Е. В экспериментах другого типа
участок сосуда при поддерживаемом постоянным диаметре растя-
Гл. 7. Механика твердого тела 119
гивают в продольном направлении (как в опытах по растяжению
проволоки). При этом величину Е рассчитывают по продольным
напряжениям и деформации. Результаты подобных эксперимен-
тов, выполненных на сегментах артерии, представлены на рис. 7.5.
Характерной чертой приведенных зависимостей в отличие от
соответствующих зависимостей для металлов (рис. 7.1, Б) являет-
ся то, что они не прямолинейны. Таким образом, даже при объ-
единении всех компонентов стенки в единое целое линейная зави-
симость [уравнение (7.1)] для описания результатов не годится
и однозначно определить модуль Юнга не удается. Эта трудность
снимается, когда нас интересуют только небольшие деформации
около некоторого положения равновесия. Это положение не обя-
зательно должно соответствовать полностью ненапряженному со-
стоянию, так как сосуд может быть предварительно растянут в
окружном и продольном направлениях. Предположим, что суще-
ствует равновесная конфигурация, в которой напряжение (про-
дольное или окружное) равно So (рис. 7.6), а соответствующая
деформация — ео- Тогда для небольших отклонений от этого со-
стояния кривая напряжение — деформация может быть прибли-
женно представлена прямой, касательной к кривой в точке, где
напряжение равно So, и небольшие деформации можно анализи-
ровать, используя модуль Юнга, задаваемый наклоном этой каса-
тельной. В таком случае говорят, что кривая напряжение — де-
формация линеаризована (разд. 8.5). В качестве примера рас-
смотрим эксперимент с продольным растяжением, когда неболь-
шие деформации происходят на фоне исходного напряженного со-
стояния, в котором натяжение, длина и площадь поперечного се-
чения образца равны соответственно Ti, /1 и Для определения
эффективного модуля Юнга мы можем воспользоваться уравне-
нием (7.1), записав его в виде
T — Tt (t-h)
где Т—— малое увеличение натяжения, а / — А— малое уве-
личение длины. Полученную таким образом величину Е иногда
называют касательным (или дифференциальным) модулем Юнга.
Ясно, что его значение зависит от 7’1, /1( /1ь и поэтому важно
всегда указывать те значения этих величин, при которых измерено
данное Е.
Далее, все кровеносные сосуды in vivo прикреплены к окру-
жающей ткани и постоянно подвержены значительному продоль-
ному растяжению: когда сегмент артерии вырезают из ткани, его
длина уменьшается на 30—40% Кроме того, среднее давление
крови во всех артериях значительно больше, чем давление, дей-
ствующее на них снаружи: превышение составляет примерно
1,3-104 Н-м-2 (100 мм рт. ст.) на уровне сердца, увеличиваясь
в соответствии с законом изменения гидростатического давления
120 Часть I. Основы механики
Рис. 7.5. А. График зависимости растягивающего (в окружном направлении) дав-
ления от диаметра для сегмента артерии, длина которого поддерживается по-
стоянной (кривая /); d0 — диаметр артерии в нерастянутом состоянии. Обратите
внимание на нелинейность графика в том диапазоне, в котором изменяется диа-
метр в условиях in vivo. Здесь же построен график для эффективного (касатель-
ного) модуля Юнга Е, связывающий окружное напряжение с окружной дефор-
мацией (кривая II). Когда растягивающее давление превышает нормальную ве-
личину, этот модуль резко возрастает. [McDonald (1974). Blood flow in arte-
ries, p. 260, Edward Arnold, London.] Б. График зависимости продольной рас-
тягивающей силы от длины, полученный для сегментов двух артерий (диаметр
которых поддерживается в ходе измерений постоянным); 1П — длина in vivo.
Как н ранее, отметим нелинейность графика. [McDonald (1974). Blood flow in
arteries, p. 261, Edward Arnold, London.]
Рис. 7.6. Схема, иллюстрирующая процедуру линеаризации кривой напряжение —
деформация. При небольших отклонениях от положения равновесия, в котором
напряжение равно So, а деформация во, кривая может быть приближенно пред-
ставлена касательной к ней в точке равновесия; при этом касательный модуль
Юнга равен ее наклону.
Гл. 7. Механика твердого тела 121
в областях, расположенных ниже этого уровня, и уменьшаясь
выше него. Таким образом, в естественном состоянии артерии рас-
тянуты в окружном и продольном направлениях. Точки, соответ-
ствующие состоянию in vivo, отмечены на рис. 7.5. Видно, что
наклон кривой напряжение — деформация (т. е. касательный мо-
дуль Юнга) in vivo значительно больше, чем в ненапряженном
состоянии, и мы должны иметь это в виду при анализе неболь-
ших деформаций в состоянии, близком к состоянию in vivo (гра-
фик для касательного модуля Юнга также построен на рис. 7.5, А).
В гл. 12 будет показано, что переменное давление и напряжение
сдвига, обусловленные потоком крови, действительно вызывают на
фоне растяжений, характерных для состояния in vivo, довольно
небольшие деформации сосудистой стенки; с помощью соответ-
ствующего касательного модуля Юнга можно связать их с пере-
менными напряжениями. С этого момента везде, говоря о мо-
дуле Е для стенок кровеносного сосуда, мы будем иметь в виду
касательный модуль для состояния in vivo. Обычно это будет ве-
личина, связывающая изменение диаметра с изменением давле-
ния (и, следовательно, окружным напряжением), а не соответ-
ствующая величина для продольного растяжения, поскольку при-
крепление сосудов к тканям препятствует продольным смеще-
ниям стенок в большей степени, чем смещениям радиальным, и
последние оказываются более важными1)- Следует отметить, од-
нако, что модуль Юнга для продольного растяжения артерий
(рассчитанный, например, по кривой рис. 7.5, Б) обычно отличается
от модуля для окружного растяжения. Это обусловлено как раз-
личием начальных напряжений по этим двум направлениям, так
и анизотропией стенок кровеносного сосуда.
*) Величина Е определяется из уравнения
„ &рв аУвО-р2)
Мн ' (d2H-<) ’
(7.3а)
где Арв— изменение давления внутри сосуда, Ad„— соответствующее изменение
наружного диаметра сосуда d„, dB— внутренний диаметр сосуда, о — коэффи-
циент Пуассона, о котором известно, что он равен 0,5.
Если сосуд тонкостенный, толщина его стенки h = 75- (dH — dB) мала по
сравнению с dB, а следовательно, dB и dH примерно равны (и потому могут быть
обозначены одним символом d). Тогда уравнение (7.3а) можно записать так:
2Ad _ d (1 - а2)
dAp Eh
Но ILd/dAp равно AAfAbp, где А — площадь поперечного сечения сосуда, и это
отношение по определению есть растяжимость D сосуда (разд. 12.3). В случае
материала, для которого о2 пренебрежимо мало по сравнению с единицей, урав-
нение (7.3а) практически эквивалентно уравнению
E[h/d) ’
(7.36)
122 Часть 1. Основы механики
В табл. I указаны значения модулей Юнга для ряда крове-
носных сосудов собаки (окружное растяжение), измеренные при
длине и диаметре, соответствующих условиям in vivo. Значение
0,7-105 Н-м-2 для вен получено при растягивающем давлении
650 Н-м-2 (около 5 мм рт. ст.). Такое давление как раз доста-
точно для того, чтобы поперечное сечение вены было круглым.
При трансмуральном давлении 2000 Н-м-2 (около 15 мм рт. ст.)
модуль Юнга Е для вен составляет около 5-10® Н-м-2, т. е. срав-
ним со значением Е для артерий большого круга. При физиологи-
ческих условиях вены функционируют в широком диапазоне дав-
лений и Е может изменяться в 100 раз, поэтому в случае больших
деформаций, например при спадении вены, мы должны прини-
мать во внимание эти изменения Е в зависимости от площади по-
перечного сечения. Отметим, что Е для артерий большого круга
увеличивается к периферии, т. е. периферические артерии более
жесткие. Значения Е, приведенные в табл. I, измерены на выре-
занных сегментах кровеносного сосуда и поэтому не отражают эф-
фект тонуса гладких мышц. Его наличие in vivo стремится уве-
личить (при данном диаметре) Е для артерий и вен, но если
сокращение гладких мышц вызывает уменьшение диаметра, то
значение Е может уменьшиться, так как интересующая нас точка
кривой напряжение — деформация будет смещаться вниз
(рис. 7.5, Л или 7.6), где наклон кривой меньше.
Считают, что резкое увеличение жесткости стенок кровеносного
сосуда при растяжении связано с различием в организации вхо-
дящих в их состав волокон коллагена и эластина. При малых де-
формациях большая часть волокон коллагена расслаблена и не
вытянута по прямой; все напряжение при этом создается волок-
нами эластина. С увеличением деформации волокна коллагена вы-
прямляются и все в большей мере участвуют в создании напря-
жения. Так как они гораздо более жесткие, чем волокна эластина
(рис. 7.4), то и вся стенка становится более жесткой. Аналогом
этой ситуации служит надувание воздушного шара, помещенного
в веревочную сетку. Сначала шар надувается сравнительно легко,
но, когда сетка натягивается, надувать его дальше становится
практически невозможно.
Затруднения в применении теории упругости к стенкам крове-
носных сосудов обусловлены также тем, что эти стенки обладают
четко выраженными вязкоупругими свойствами (разд. 7.1). Когда
к ним прикладывается стационарное напряжение, соответствую-
щие стационарные деформации достигаются не мгновенно; за воз-
никающей в начальный момент деформацией следует более мед-
ленная (ползучесть), нарастающая вплоть до достижения равно-
весного значения. Аналогичным образом напряжение, необхо-
димое для поддержания данной деформации, постепенно падает
от начального уровня до нового равновесного значения (релакса-
ция напряжения). Считают, что за вязкоупругие свойства стенок
Гл. 7. Механика твердого тела 123
Рнс. 7.7. Три последовательно полученные кривые давление — объем для бедрен-
ной артерии. Порядок наполнения артерии н ее опорожнения показан стрел-
ками (кривые для двух последних опорожнений совпадают). Оказывается,
что в ходе последовательных наполнений сосуд становится несколько менее рас-
тяжимым. [Bergel (1961). The static elastic properties of the arterial wall, p. 451,
J. Physiol. 156.]
артерий ответственны в основном гладкие мышцы, хотя вязкоуп-
ругость в экспериментах in vitro проявляет и коллаген (несмотря
на свою большую жесткость). Эластин, с другой стороны, являет-
ся чисто упругим; именно этот упругий компонент стенок крове-
носного сосуда предотвращает неограниченное течение и релакса-
цию напряжения. Вязкоупругость существенна только в тех слу-
чаях, когда напряжение или деформация меняется во времени;
например, если сегмент артерии подвергается периодически ме-
няющимся деформациям данной частоты и амплитуды, то напря-
кение, необходимое для достижения данной деформации, оказы-
вается больше во время растяжения, чем во время фазы возвра-
щения к исходному состоянию (рис. 7.7). Явление подобного рода
называется гистерезисом.
Свойства вязкоупругих материалов в отличие от материалов
линейно-упругих, которые характеризуются модулем Юнга и
коэффициентом Пуассона, обычно нельзя охарактеризовать с по-
мощью небольшого числа констант. На самом деле, описать в точ-
ной математической форме реакцию вязкоупругих материалов на
прикладываемые нестационарные напряжения различных видов,
как правило, не удается, а тогда, когда это оказывается возмож-
124 Часть I. Основы механики
Рис. 7.8. Когда к образцу прикладывается периодически меняющееся напряже-
ние, возникающая в ответ деформация совпадает с ним по фазе, только если
материал чисто упругий (кривая I). Если же он обладает вязкоупругими свойст-
вами, то деформация отстает от напряжения и амплитуда ее изменений меньше,
чем в первом случае (кривая II).
ным, математическое описание становится громоздким и трудно-
обозримым. В ряде случаев, однако, можно получить простое при-
ближенное описание свойств материала, и оно оказывается чрез-
вычайно полезным. Это, в частности, делается для кровеносных
сосудов, когда деформации малы и периодически меняются во
времени под действием периодически меняющегося напряжения
(т. е. давления крови и вязкого напряжения сдвига, которые ме-
няются во времени в ходе каждого сердечного цикла). Изменение
деформации в ответ на изменение напряжения слегка запазды-
вает, поэтому, когда приложенное напряжение периодически ме-
няется, колебания деформации отстают от колебаний напряжения
и имеют меньшую амплитуду, чем для чисто упругого материала
(рис. 7.8) (см. разд. 8.1, где дано точное определение амплитуды).
Охарактеризовать подобное поведение можно с помощью двух
констант: «динамического модуля Юнга» ЕДИН и «эффективной вяз-
кости» т]1). Как мы увидим в гл. 8 (разд. 8.6), изменения во вре-
мени произвольной периодически изменяющейся величины (в дан-
ном случае — напряжения) может быть представлено в виде сум-
мы некоторого числа чисто синусоидальных колебаний (рис. 8.14).
*) Удобно использовать не вязкость т), а произведение Т]со, имеющее ту же
размерность, что и Едия, и называемое «модулем потерь». — Прим, ред
Гл 7. Механика твердого тела 125
Частота, Гц
Рис. 7.9. Экспериментально полученные зависимости динамического модуля Юнга
Един от частоты ю/2л прн периодическом растяжении сегментов артерий. ЕДИп
примерно постоянен в широком диапазоне частот. I — грудная аорта; II — брюш-
ная аорта; III—бедренная артерия; IV—сонная артерия. [McDonald (1974).
Blood flow in arteries, p. 270, Edward Arnold, London.]
Если одно из таких колебаний напряжения имеет угловую час-
тоту со и амплитуду S, то соответствующее ему колебание дефор-
мации будет иметь амплитуду Е/д/еднн + Ц2<о2. а фазовый угол,
на который оно отстает от напряжения, составит примерно г)ш/Един
(в радианах). Используя эти формулы, можно вычислить значе-
ние величин Един и т), прикладывая к сегменту артерии известное
периодически меняющееся напряжение и измеряя возникающую
переменную деформацию.
Некоторые результаты подобных экспериментов представлены
на рис. 7.9. Установлено, что Един обычно больше, чем Е, измеряе-
мое в статических экспериментах. При частотах ниже примерно
2 Гц (т. е. когда со/2л меньше, чем 2 с-1) Едии меняется с часто-
той, достигая статической величины, когда частота приближается
к нулю. При частотах от 2 до 20 Гц величина Едни примерно по-
стоянна и превышает ЕСтат в 1,1—1,8 раза (первое из чисел отно-
сится к грудной аорте собаки, второе — к сонной артерии). Мо-
дуль потерь т](о также зависит от частоты, но при частотах, пре-
вышающих 5 Гц, примерно постоянен и составляет от 0,1 до
0,2 Един. Величина т)(о тем больше, чем артерия мельче и чем
126 Часть I. Основы механики
больше мышечных волокон в ее стенке, поэтому подобные арте-
рии обладают более четко выраженными вязкоупругими свой-
ствами.
7.3. Статика упругой трубки
В гл. 12 мы рассмотрим пульсирующее движение крови в круп-
ных сосудах и сопровождающие его движения стенок сосудов.
Другими словами, мы изучим динамику крови и стенок сосудов.
Однако сначала нам нужно рассмотреть баланс сил, действующих
на стенки совершенно неподвижной трубки, которая содержит по-
коящуюся жидкость, т. е. изучить статику стенки сосуда. Рассмот-
рим, например, артерию, которая 'натянута в продольном направ-
лении силами, действующими со стороны окружающей ее ткани,
и раздута внутренним давлением, превышающим давление сна-
ружи (последнее во многих случаях равно атмосферному) при-
мерно на 1,3-104 Н-м-2 (100 мм рт. ст.), что эквивалентно сред-
нему давлению в артерии на уровне сердца; в ходе сердечного
цикла давление меняется примерно от 1,0-104 до 1,6-104 Н-м-2
(80—120 мм рт. ст.). Силу тяжести в балансе сил мы учитывать
не будем, поскольку обусловленные высоким трансмуральным
давлением напряжения значительно выше напряжений, возникаю-
щих под действием сил гравитационного притяжения. Когда же
трансмуральное давление невелико, сила тяжести может стать су-
щественной (например, она может вызвать спадение на воздухе
тонкостенной гибкой трубки), и выводы, которые мы сейчас по-
лучим, будут верны для этого случая только тогда, когда сосуд
заполнен и окружен жидкостью с плотностью, равной плотности
стенки. При этом вес стенки будет уравновешиваться гидростати-
ческим давлением в жидкости. В системе кровообращения силой
тяжести не всегда можно пренебрегать еще и потому, что давле-
ние внутри кровеносных сосудов имеет составляющую, которая
меняется гидростатически (совершенно независимо от изменения
давления, связанного с потоком крови). Например, у стоящего че-
ловека давление в сосудах ступни будет превышать давление на
уровне сердца примерно на 1,3-104 Н-м-2 (100 мм рт. ст.). Соот-
ветственно в сосудах головы давление крови будет несколько
ниже, чем на уровне сердца. Давление вне сосудов всюду при-
мерно одинаково и близко к атмосферному. Исключение состав-
ляет только грудная клетка: при интенсивном дыхании давление
может отклоняться от атмосферного в ту и другую сторону при-
мерно на 3-103 Н-м-2 (23 мм рт. ст.). Имея в виду, кроме того,
что трансмуральное давление различается даже для кровеносных
сосудов, расположенных на одном уровне, но относящихся к раз-
ным типам (в артериях оно выше, чем в венах), мы приходим к
выводу, что в сердечно-сосудистой системе трансмуральное давле-
ние может изменяться в широком диапазоне. Как мы увидим
Гл. 7. Механика твердого тела 127
ниже, это обстоятельство оказывается весьма существенным при
ответе на вопрос о том, могут или нет конкретные кровеносные
сосуды полностью спадаться.
Проанализируем теперь статический баланс сил, действующих
на малый элемент стенки сосуда, пренебрегая при этом силой тя-
жести (возможно, однако, что она участвует в создании давления
внутри трубки). Обозначим давление внутри и снаружи сосуда со-
ответственно через рв и рн и предположим, что трубка имеет круг-
лое поперечное сечение с внутренним радиусом гв и наружным гн
(рис. 7.10, Д). Толщина стенки h равна гн — гв. Рассмотрим эле-
мент, длина которого вдоль оси трубки составляет Д/ и который
«стягивает» небольшой угол 0 (рис. 7.10,Б). Результирующая
сила, действующая на этот элемент, обусловлена перпендикуляр-
ными (нормальными) напряжениями на шести его поверхностях;
поскольку система в целом симметрична, тангенциальные напря-
жения отсутствуют. Составляющие действующей на элемент ре-
зультирующей силы в трех взаимно перпендикулярных направле-
ниях должны быть равны нулю. Выберем эти направления сле-
дующим образом: 1—параллельно оси трубки, 2 —по радиусу от
оси, 3 — по перпендикулярной к осям 1 и 2 касательной к внутрен-
ней поверхности трубки (рис. 7.10, Б).
Условие баланса сил в продольном направлении (т. е. в на-
правлении оси 1) показывает, что продольное натяжение Т на
SehM
трубки,
которой
₽нД/г„в
Sehbl
h
Рис. 7.10. А. Схема толстостенной
внутренний и наружный радиусы
равны соответственно гв и гн; толщина стен-
ки h = гк — гв; давление внутри и снаружи
равно рв и рк соответственно. Б. Элемент
стенки толстостенной трубки имеет длину
AZ, толщину h и «стягивает» угол 0. Силы,
действующие на этот элемент, показаны бо-
лее жирными стрелками. Силы, действую-
щие на искривленные поверхности, обуслов-
лены внутренним и внешним давлениями, а
на плоские грани, параллельные осн труб-
ки, — окружным напряжением. Растягиваю-
щие силы, приложенные к плоским торцам,
взаимно уравновешены и не показаны на
рисунке.
128 Часть I Основы механики
каждом торце должно быть одним и тем же (оно равно площади
торца, умноженной на среднее нормальное напряжение на этом
торце). Аналогично баланс сил в тангенциальном направлении
(т. е. по оси 3) показывает, что нормальные силы на двух пло-
ских боковых поверхностях элемента, параллельных оси сегмента,
также должны быть одинаковы (они равны площади поверхности
ЛА/, умноженной на среднее нормальное напряжение Se на этой
поверхности). Однако поскольку эти поверхности наклонены друг
к другу, а следовательно, и к радиальной координатной оси 2, то
результирующая сила имеет составляющую, направленную по ра-
диусу внутрь и равную 2SGh Д/sin у6. Еще один вклад в радиаль-
ную составляющую силы вносит давление внутри и снаружи труб-
ки; направленная наружу сила, обусловленная внутренним давле-
нием, равна рв, умноженному на площадь внутренней изогнутой
поверхности (Д/гв6), а направленная внутрь сила, обусловленная
внешним давлением, равна рн, умноженному на площадь наруж-
ной изогнутой поверхности (Д/гн0). Таким образом, баланс сил
в радиальном направлении дает соотношение
рвД/гв0 — рн • Д/гн0 = 2SeftAZ sin -|-0. (7.4)
Далее, когда 0 мало (как в нашем случае), sin у 0 примерно ра-
вен ~0 (это соотношение справедливо с точностью до 1% для
0 вплоть до 0,24 рад, или примерно 14е). Поэтому можно со-
кратить обе части выражения (7.4) на Д/0 и получить соотноше-
ние
/Vb~ /Vh = Sg/i. (7.5)
Величина Sg есть среднее окружное напряжение в стенке трубки;
величина же Sgh— это приложенное в окружном направлении
натяжение в стенке, приходящееся на единицу длины трубки.
Наш опыт обращения с воздушными шарами подсказывает, что
раздувание, вызванное превышением внутреннего давления над
внешним, должно сопровождаться натяжением в стенке шара,
т. е., когда ръ > рн, натяжение So/i положительно. Однако из урав-
нения (7.5) видно, что, если стенка трубки достаточно толстая,
т. е. гн существенно превышает гв, это «натяжение» может быть
отрицательным, несмотря на то, что рв > рн. Другими словами, на-
пряжение в стенке является сжимающим1)- Этот результат со-
вершенно не зависит от упругих свойств материала стенки, но они
’) Это утверждение, кажущееся на првый взгляд парадоксальным, на самом
деле легко объяснимо Следует помнить, что речь идет о средних по толщине
стенки напряжениях, а не об истинных их значениях. Последние же распределе-
ны так, что в стенке при рв > рв имеется область растяжения (оиа примыкает
к внутренней поверхности трубки) и, возможно, область сжатия (у наружной
Г л. 7. Механика твердого тела 129
становятся существенными при оценке влияния, которое подоб-
ное сжимающее напряжение оказывает на форму поперечного се-
чения трубки. Прежде чем обратиться к этому вопросу, посмот-
рим, каким является окружное напряжение в кровеносных сосу-
дах— растягивающим или сжимающим.
Рассмотрим, например, артерию, давление внутри которой пре-
вышает атмосферное на 1,3-104 Н-м~2 (около 100 мм рт. ст.);
давление снаружи равно атмосферному (105 Н-м~2). Если это,
скажем, бедренная артерия собаки, свойства которой приведены
в табл. I, помещенной в начале книги, то внутренний радиус ее
составляет 0,002 м при толщине стенки 0,0004 м. Рассчитав по
уравнению (7.5) натяжение в стенке Soft, мы увидим, что оно
отрицательно (т. е. напряжение сжимающее) и составляет при-
мерно —14,0 Н-м~’. При внутреннем давлении, большем всего на
50 мм рт. ст. (что может наблюдаться в ступне), напряжение
в стенке было бы растягивающим; если бы внутреннее давление
было меньше, чем указано, сжатие было бы еще более сильным.
Подобный анализ напряжения в стенках всевозможных кровенос-
ных сосудов показывает, что для огромного большинства из них
характерно именно сжимающее окружное напряжение. Исключе-
ние составляют лишь самые крупные сосуды — аорта и полая
вена, а также крупные сосуды, расположенные ниже уровня серд-
ца, в которых внутреннее давление относительно высоко. Наибо-
лее высоки сжимающие напряжения в мелких артериях и арте-
риолах, стенки которых очень толстые, так что ги значительно пре-
вышает гв-
Тот факт, что окружное напряжение может быть сжимающим,
не имеет большого значения с точки зрения механики кровенос-
ных сосудов in vivo. Дело в том, что при изучении кровообраще-
ния мы обычно анализируем лишь небольшие отклонения от не-
которого положения равновесия. Как уже говорилось, в рамках
подхода, при котором упругие свойства материала характеризуют-
ся эффективным модулем Юнга, измеренным для небольших от-
клонений от положения равновесия, не имеет значения, есть ли в
этом исходном положении равновесия начальное напряжение или
нет. По этой причине обычно указывают не абсолютные значения
напряжения и давления, а относительные, определенные по отно-
шению к их значениям в данном исходном состоянии равновесия.
В физиологии давление, как правило, отсчитывается от атмосфер-
ного. Соответствующие натяжения отсчитываются от их значений
для случая, когда давления внутри (рв) и снаружи (рн) сосуда
поверхности). С ростом внутреннего давления внутренний диаметр трубки уве-
личивается, тогда как наружный может уменьшаться Кроме того, если материал
стенки несжимаем, то напряжения в ней имеют некоторую составляющую, ана-
логичную гидростатическому давлению в несжимаемой жидкости. Эта составляю-
щая может быть сжимающей, ио, как и в случае жидкости, она не вызывает в
несжимаемом материале никаких деформаций.— Прим. ред.
130 Часть I. Основы механики
Рис. 7.11. Формы, которые может принимать поперечное сечение спавшейся, ис-
ходно круглой трубки.
(трубки) равны атмосферному (ра™). В этом случае окружное
напряжение (обозначим его So) равно
$0 Ратм (^в Г „)/Й Ратм>
как следует из уравнения (7.5). Необходимо отметить, что не
только окружное, но и все нормальные напряжения являются при
этом сжимающими и равны атмосферному давлению. Если при
отклонении от подобного состояния равновесия рн остается рав-
ным атмосферному давлению, рв превышает ратм на величину р',
a Se превышает —ратм на величину S', то из уравнения (7.5) сле-
дует, что
З'Л = р'гв,
или
Р' = ~- (7.6)
' в
Это есть не что иное, как хорошо известный «закон Лапласа».
Из него видно, что давление, необходимое для растяжения труб-
ки против данного натяжения в стенке, обратно пропорционально
радиусу трубки. Воздушный шар, например, труднее надувать
когда его радиус очень мал, чем когда он уже несколько увели-
чен. Применительно к кровообращению этот закон показывает, что
натяжение, уравновешивающее определенно растягивающее давле-
ние (в относительных величинах, отсчитываемых от атмосферного
давления), уменьшается при уменьшении радиуса сосуда. Иногда
с помощью этого факта пытаются объяснить, почему стенки капил-
ляров так тонки, указывая, что они должны обеспечить совсем
небольшое натяжение. Однако этот довод не учитывает гораздо
более существенного воздействия со стороны окружающей ткани
и не является удовлетворительным.
Таким образом, закон Лапласа может использоваться для ана-
лиза лишь небольших отклонений от такого состояния равновесия,
в котором все давления равны атмосферному. Его нельзя приме-
нять для точного определения абсолютных значений напряжения
в стенке сосуда до тех пор, пока растягивающее давление не бу-
Гл. 7. Механика твердого тела 131
дет настолько большим, что напряжение Se [уравнение (7.5) ] ста-
нет велико по сравнению с атмосферным давлением. Этого, од-
нако, обычно не происходит. При анализе ситуаций, в которых
возможно спадение (коллапс) сосудов, важно знать именно абсо-
лютное значение напряжения. Под спадением сосуда понимается
резкое уменьшение площади его поперечного сечения, которое
происходит самопроизвольно в ответ на малое изменение прило-
женных напряжений. Обычно спадение сопровождается изменением
формы поперечного сечения сосуда: из круглого оно может стать
эллипсоидным или еще более сложным (рис. 7.11). Ясно, что
трубка спадется, если давление снаружи существенно превысит
давление изнутри. Спадение обусловлено возникновением боль-
ших сжимающих окружных напряжений, которые, согласно урав-
нению (7.5), должны сопровождать такие изменения давления.
Аналогом этому явлению может служить прогиб прямого прута,
к концам которого приложены достаточно большие сжимающие
силы. При этом теоретически прут еще может остаться прямым,
но такое его состояние является неустойчивым в том смысле, что
очень небольшие отклонения от него приводят к быстрому про-
гибу. Изогнутое состояние является уже устойчивым. Аналогично
карандаш теоретически может стоять на своем острие, но это по-
ложение равновесия неустойчиво, так как любое малое отклоне-
ние от него быстро нарастает и карандаш падает, принимая устой-
чивое положение. Точно так же и круглая форма трубки неустой-
чива, если в ее стенках существуют достаточно большие сжимаю-
щие напряжения. Стенка изгибается и приобретает устойчивую
искаженную форму (рис. 7.11); это и означает, что трубка спа-
дается. х
Величина сжимающих напряжений, приводящих к спадению,
как и форма, которую приобретает трубка, зависят от всех упру-
гих свойств стенки. В частности, процесс спадения во многом оп-
ределяется степенью противодействия сильному изгибу со сто-
роны материала стенки, которая в свою очередь зависит от мо-
дуля Юнга, коэффициента Пуассона и толщины стенки. На спа-
дение влияют также степень продольного удлинения и характер
механического воздействия со стороны окружающей ткани. Совер-
шенно свободная трубка с достаточно тонкими стенками и незна-
чительным продольным натяжением спадается чрезвычайно легко,
т. е. при очень небольшом сжимающем напряжении в ее стенках.
Однако in vivo кровеносный сосуд растянут в продольном направ
лении, прикреплен к окружающей ткани и имеет довольно тол-
стые стенки; все это препятствует спадению. Тем не менее уста-
новлено, что некоторые вены и артерии малого круга и в нормаль-
ных условиях имеют поперечные сечения в виде эллипсов, что
указывает на некоторое их совпадение, связанное с наличием сжи-
мающего напряжения в стенках. Другие примеры спадения кро-
веносных сосудов in vivo рассмотрены в гл, 14,
Глава 8
КОЛЕБАНИЯ И ВОЛНЫ
8.1. Простое гармоническое движение
Давление в аорте и других крупных артериях возрастает в мо-
мент изгнания крови из сердца во время систолы и затем падает
во время диастолы. Рост давления сопровождается радиальным
движением стенок сосудов (растяжением), а когда давление па-
дает, они, будучи упругими, возвращаются в исходное состояние.
Этот процесс происходит во время каждого сердечного цикла, и,
стало быть, элементы стенок сосудов колеблются периодически с
частотой, равной частоте сокращений сердца. В ответ на пульси-
рующее изменение давления кровь движется также толчками, и,
как мы увидим в гл. 12, в действительности при этом происходит
распространение волны давления по артериальному дереву. По-
этому имеет смысл в этой главе рассмотреть предварительно ме-
ханику колебаний в целом и процесс распространения волн в
частности.
Проанализируем вначале колебательное движение отдельной
частицы. Предположим, что частица находится в состоянии равно-
весия в некоторой точке Р и, будучи смещенной, стремится вер-
нуться в эту точку. Примеров подобной ситуации множество: под-
вешенное на нити тело, которое отклоняют в сторону (простой
маятник), тело, прикрепленное к упругой нити и оттягиваемое
вниз от положения равновесия, и т. д. Во всех подобных случаях
возвращающая сила растет с увеличением расстояния, на которое
смещена частица от точки Р. На самом деле для достаточно ма-
лых смещений возвращающая сила оказывается примерно пропор-
циональной расстоянию от точки Р (разд. 8.5). Если частицу сме-
стить, а затем освободить, то она начнет возвращаться к точке Р,
однако в силу инерции «проскочит» положение равновесия и от-
клонится в противоположную сторону. При этом на нее будет
действовать возвращающая сила, замедляющая частицу вплоть до
полной остановки. После остановки частица вновь начнет возвра-
щаться к точке Р, проскочит ее, отклонится в сторону, куда она
была отклонена вначале, еще и еще раз проскочит положение рав-
новесия — т. е. будет колебаться около точки Р.
Математический анализ этого процесса чрезвычайно прост, и
мы приведем его здесь целиком, поскольку считаем, что ои поле-
зен для более полного понимания механики колебательного дви-
жения. Пусть частица массой т колеблется по прямой около сво-
Гл. S. Колебания и волны 133
его положения равновесия Р (рис. 8.1). Если у — расстояние час-
тицы от Р, то величина возвращающей силы, направленной к Р,
пропорциональна у и равна, скажем, Ку, где К — положительная
постоянная, измеряемая в соответствующих единицах. Примем,
что у положительно при смещении вправо от Р и отрицательно
при смещении влево; тогда сила, действующая на частицу в поло-
жительном направлении у, будет равна —Ку. Согласно второму
Сипа К у
Рис. 81 Частица, движущаяся вдоль прямой под действием возвращающей
силы, которая равна произведению К на смещение у от положения равновесия Р.
Ускорение частицы по направлению от точки Р равно A2y/At2. Такое движение
описывается уравнением (8.1).
закону Ньютона, сила равна произведению массы частицы на ее
ускорение в направлении положительных у [это ускорение равно
d2t//d^2; см. уравнение (2.5)]. Итак, мы приходим к уравнению
(8.1)
которое может быть представлено в виде
(8.2)
где со2 = К/т. Уравнение (8.2) применимо к колебаниям любой
частицы около положения равновесия, если только эти колебания
настолько малы, что возвращающая сила пропорциональна у. По-
лученное уравнение выражает условие равновесия между дей-
ствующей на частицу возвращающей силой и инерцией частицы;
для существования любого колебательного движения необходимо,
чтобы система обладала этими двумя свойствами.
По-видимому, наиболее типичным примером подобной системы
служит простой маятник, в котором частица массой m подвешена
почти вертикально на нити длиной I (рис. 8.2). Предположим, что
нить нерастяжима, и пренебрежем сопротивлением воздуха, а так-
же всеми другими силами трения; тогда единственными действую-
щими на частицу силами будут ее вес, равный mg и направлен-
ный вертикально вниз, и сила F, действующая со стороны нити
(натяжение нити) и направленная вдоль нее. Так как a priori ве-
личина F неизвестна, удобнее всего рассмотреть баланс сил в на-
правлении, перпендикулярном нити: в этом направлении F не
имеет составляющей. Обозначим через 0 угол (в радианах), ко-
торый нить образует с вертикалью. Единственной действующей
134 Часть I. Основы механики
в выбранном направлении силой будет составляющая веса тела,
равная mg sin 0 и направленная вдоль дуги к положению равно-
весия (последнее отмечено точкой Р на рис. 8.2). Составляющую
ускорения в данном направлении мы получим, приняв во внима-
ние, что частица движется по дуге вертикальной окружности ра-
диусом I. Составляющая ускорения по касательной к этой окруж-
ности равна d2s/d/2, где s — расстояние по окружности, пройден-
ное от точки Р. Расстояние s равно /0, следовательно, танген-
циальная составляющая ускорения равна d2(Z0)/d/2 и направлена
Рис. 8.2. Простой маятник. Ча-
стица подвешена на иерастяжи-
мой нити длиной I и. колеблется
в вертикальной плоскости. Си-
лы, действующие на частицу: ее
вес mg, направленный верти-
кально вниз, н натяжение нити
F. Удобнее всего рассматри-
вать баланс сил в направле-
нии, перпендикулярном нити;
составляющие силы и ускоре-
ния в этом направлении пока-
заны иа рисунке и обсуждают-
ся в тексте.
в сторону увеличения 0 (см. рис. 8.2). Уравнение «сила равна про-
изведению массы на ускорение» дает в этом случае соотношение
• n d2 (16)
mg sin 0 = — m —At2~.
где знак минус появился из-за того, что справа должна стоять со-
ставляющая ускорения в направлении силы, а не в противопо-
ложном направлении. Данное уравнение может быть переписано
в виде
+ ё sin 0 = 0. (8.2а)
Заметим, что если горизонтальное смещение частицы равно у,
то
sin 6 = у/1.
Предположим теперь, что угол 0 все время остается очень малым;
это означает, что частица фактически колеблется вдоль горизон-
тальной прямой и что sin0 примерно равен 0 (см. разд. 7.3), по-
этому /0 примерно равно у. В этом случае уравнение (8.2а) упро-
щается и приобретает вид уравнения (8.2), в котором роль <о2
играет g/l.
Каким бы образом мы ни освобождали частицу, ее последую-
щее движение должно быть таким, чтобы изменение у во вре-
Рл. 8. Колебания и волны 135
мени удовлетворяло уравнению (8.2) при данной величине со. [На
языке математики это звучит так: функция y(t) должна быть ре-
шением дифференциального уравнения (8.2).] Оказывается, од-
нако, что число движений, удовлетворяющих этому уравнению,
бесконечно; то единственное из них, которое сооответствует дан-
ной ситуации, полностью определяется смещением у и скоростью
dy/d/ в момент освобождения (мы можем, например, при данном
исходном смещении освободить частицу без начальной скорости
или же оттолкнуть ее в сторону от Р с некоторой скоростью).
В наиболее общем виде движение, удовлетворяющее уравнению
(8.2) (его общее решение), дается выражением
у — A cos со/ -р В sin со/, (8.3)
где А и В — произвольные константы. Соответствующую этому
движению скорость частицы dy/dt получают путем дифференци-
рования уравнения (8.3) по времени:
— —со Л sin со/ + соВ cos со/. (8.4)
Значения констант А и В можно определить, зная начальцые зна-
чения у и dy/dt. В качестве первого примера рассмотрим простой
маятник, гирю которого (будем считать ее частицей) отводят в
сторону на расстояние а по горизонтали от положения равнове-
сия, а затем в момент времени / = 0 отпускают с нулевой ско-
ростью. Таким образом, при / = 0 у—а, -^- = 0. Заметим, что
sin0 = 0, a cosO = 1. Подставив второе из этих равенств в (8.4),
мы увидим, что В должно равняться нулю; подстановка первого
равенства в (8.3) даст соотношение А = а. Таким образом, ре-
шение задачи о движении частицы на протяжении всего времени
имеет вид
у— a cos со/, (8.5)
а скорость, как и раньше, получается из этого выражения путем
дифференцирования:
-4т- = —соя sin а/. (8.6)
dr
Графики зависимости двух этих величин от времени приведены
на рис. 8.3. Видно, что у периодически меняется от -j-a до —а, а
dy/dt — от 4-соя до —соя, проходя через нуль при у, равном ±а.
Величина а называется амплитудой колебания положения точки;
соя есть амплитуда скорости. Периодом колебания Т (рис. 8.3) на-
зывают время, за которое завершается полный цикл колебания;
186 Часть I. Основы механики
Рис. 8.3. Графики зависимости от времени величин у и &y!6t, заданных соотно-
шениями (8.5) и (8.6). Амплитуда колебаний положения равна а, а скорости —
соа. Период колебаний равен 7'(=2л/<в). Эти графики представляют простое
гармоническое движение. Штриховые линии, являющиеся их продолжением до
новой точки начала координат, соответствуют соотношениям (8.7) и (8.8) при
V = по.
(х положительно,
у положительно)
6 = 0
6=2п
х положительно,
У отрицательно)
Рис. 8.4. Схема, иллюстрирующая периодический характер функций sin0 и
cos0, определенных тригонометрически В первом квадранте обе функции поло-
жительны; во втором sin0 положителен, a cos0 отрицателен; в третьем обе
функции отрицательны; в четвертом sin 0 отрицателен, a cos 0 положителен.
При увеличении 0 иа 2л (360°) функции sin 0 и cos 0 остаются неизменными.
Дальнейшие пояснения даны в сноске на стр. 137.
Гл. 8. Колебания и волны 137
он равен 2n/(o’). Частота колебаний, т. е. число циклов или пе-
риодов в единицу времени, равна <о/2л; величина <о(= 2л/) на-
зывается угловой частотой. Единицей измерения всех частот яв-
ляется обратное время, т. е. с_|; это связано с тем, что как угло-
вая единица — радиан, так и число циклов безразмерны. Вели-
чины <в и а совершенно независимы друг от друга: частота <о пол-
ностью определяется параметрами рассматриваемой системы
(массой частицы m и константой, характеризующей возвращаю-
щую силу К), в то время как амплитуда а определяется исклю-
чительно смещением и скоростью, которую имеет данное тело в
момент его освобождения. Интересно отметить, что для маятника
частота не' зависит также и от массы т, так как со равно g/l-
Таким образом, период колебаний маятника не зависит от массы
гири; имейте, однако, в виду, что механизм действия маятнико-
вых часов и их регулировка не столь просты, как это может по-
казаться из всего сказанного выше.
Не составляет особого труда найти решение, аналогичное урав-
нениям (8.5) и (8.6), для случая, когда маятник начинает дви-
гаться из положения равновесия с начальной скоростью V. В дан-
ной ситуации в момент / = 0 у==0, — Подставив первое
из этих условий в уравнение (8.3), получим А = 0; подстановка
второго условия в уравнение (8.4) дает соотношение соВ = V,
Следовательно, движение частицы описывается выражением
у=^- sin at, (8.7)
*) Это следует из тригонометрического определения синуса и косинуса. Рас-
смотрим изменение величин sin 0 и cos 0 с увеличением 0 от нуля. Обратимся
для этого к рис. 8 4, где изображена окружность единичного радиуса с центром
в точке С — начале координат Угол 0 представлен точкой А: он равен углу,
образованному прямой СА с осью х. Координаты точки А суть (х, у). Тогда
sin 0 определяется из прямоугольного треугольника АВС как отношение длины
противолежащей стороны А'В к длине гипотенузы СА, т. е. sin 0 = у/1 = у. Ана-
логично cos 0 есть отношение длины прилежащей стороны треугольника к длине
гипотенузы: cos 0 = х/1 = х. При увеличении 0 sin 0 и cos 0 изменяются, оста-
ваясь положительными до тех пор, пока 0 не станет равным л/2 (90°); здесь
точка А переходит во второй квадрант и х становится отрицательным, а у
остается положительным. Таким образом, при значениях 0 от л/2 до л (180°)
cos 0 отрицателен, a sin 0 положителен. В третьем квадранте [от л до Зл/2
(270°)] как х, так и у отрицательны, д поэтому отрицательны и cos0 и sin0.
В четвертом квадранте [от Зл/2 до 2л (360°)] х снова положителен, и поэтому
cos 0 также положителен, a sin 0 отрицателен. Когда 0 становится больше 2л,
точка А снова попадает в первый квадрант, и все повторяется сначала: cos0 и
sin 0 положительны. Видно, что, увеличивая значение 0 на 2л, мы проходим
полный круг и снова попадаем туда, откуда начинали движение; следовательно,
эта операция не изменяет значений cos 0 и sin 0. «Период» sin 0 и cos 0 равен
2л. Отсюда ясно, почему расстояние между последовательными максимумами
на графике функции a cos cot (рис 8 3) равно Т — 2л/ы,
138 Часть 1. Основы механики
а ее скорость равна
= V cos со/. (8.8)
Амплитуда колебаний положения равна V/со, а амплитуда ско-
роста—V. Угловая частота, как и прежде, равна со. Графическое
представление соотношений (8.7), (8.8) можно получить из гра-
фиков, изображенных на рис. 8.3, сместив точку начала отсчета
времени на четверть периода назад (см. штриховую линию на
этом рисунке) и заменив везде а на Г/со.
«0/си
Рис. 8.5. Графики функций (8.9) (сплошная линия) и (8 9а) (штриховая линия).
Сплошная кривая целиком сдвинута вправо относительно штриховой на вели-
чину, соответствующую времени ф/ы; ф — фазовый угол, на который колебание,
заданное соотношением (8.9), опережает колебание, заданное соотношением
(8.9а).
Любое движение, удовлетворяющее уравнению (8.2) [частные
случаи такого рода движения описываются уравнениями (8.5) и
(8.7)], называется простым гармоническим колебанием. Выраже-
ние (8.3) представляет наиболее общий случай простого гармони-
ческого колебания с данной угловой частотой со; это выражение
может быть преобразовано к виду
у = A' cos (со/ — ср), (8.9)
где А' = -\М2+ В2, а ср — угол, такой, что tg ср ~ В/A. Амплитуда
Колебания равна А'; величина ср называется его фазой. Говорят,
что такое колебание отстает по фазе на угол ср от колебания, опи-
сываемого выражением (8.5). Графическое представление зависи-
мости (8.9) дано на рис. 8.5; здесь же приведен график функции
у — A' cos со/. (8.9а)
Видно, что график (8.9а) целиком сдвинут вправо, как если
бы величина со/ везде была уменьшена на ср. При колебаниях, опи-
сываемых соотношением (8.9а), все события происходят раньше,
чем в случае (8.9), на время <р/со — в этом и заключается смысл
Гл. 8. Колебания и волны 139
выражения «отставание по фазе». Если величина <р оказывается
отрицательной, то говорят, что наблюдается опережение по фазе.
Величина —sin ю/ в выражении (8.6) может быть записана в виде
cos(co^4* л/2), т. е. при таком колебании скорость опережает сме-
щение по фазе на л/2. Из рис. 8.3 видно, что скорость достигает
данной точки (например, минимума) на время Т/4 раньше, чем
смещение, т. е. с опережением по величине со/ на л/2.
С помощью рис. 8.3 мы можем описать все основные черты
простого гармонического колебания: когда смещение у макси-
мально, скорость di//d/ равна нулю; возвращающая сила в этот
момент максимальна, а поэтому максимально и направленное на-
зад, к точке равновесия Р (рис. 8.2), ускорение. По мере возвра-
щения частицы к точке Р смещение у уменьшается, а скорость
возрастает; когда частица проходит через точку Р, смещение и
ускорение равны нулю, тогда как скорость максимальна После
прохождения точки Р скорость опять уменьшается, а смещение и
ускорение растут до своих максимальных значений. В отсутствие
трения этот процесс повторяется бесконечно долго.
Описанное выше простое гармоническое колебание является
движением с одной степенью свободы-, оно полностью задано, если
указан характер изменения во времени одной переменной (коор-
динаты частицы у). Система из двух подвешенных одна над дру-
гой на нити частиц, которые могут колебаться в одном и том же
направлении (двойной маятник), или система из одной частицы,
которая может колебаться в плоскости (примером такой системы
служит маятник с упругой нитью), имеют две степени свободы,
так как для описания такого движения требуются две коорди-
наты. Системы частиц более общего вида имеют большее, но ко-
нечное число степеней свободы. С другой стороны, для полного
описания движения любой непрерывной системы, такой, как скри-
пичная струна, вода в сосуде или стенки артерии, необходимо
указать координаты бесконечно большого числа элементов (на-
пример, элементарных объемов жидкости), поэтому подобная си-
стема имеет бесконечное число степеней свободы. Ситуация, од-
нако, не настолько сложна, как можно подумать, поскольку Все
эти элементы связаны друг с другом, — на самом деле описать
волновые движения в непрерывных системах не составляет осо-
бого труда.
8.2. Простые примеры волнового движения
В качестве простейшего примера рассмотрим колебания натя-
нутой упругой струны, закрепленной на обоих концах. Предполо-
жим, что в некоторый начальный момент струна испытала щипок,
удар или была немного смещена вбок каким-то другим способом,
а затем отпущена (рис. 8.6,Д). Как и раньше, будем пренебрегать
всеми видами трения и примем, что никакой диссипации энергии
140 Часть I Основы механики
не происходит. Натяжение струны обусловливает возникновение
возвращающей силы, которая стремится вернуть каждый смещен-
ный элемент в положение равновения (ясно, например, что на
элемент АВ действует результирующая сила, направленная вниз)
Каждый элемент имеет массу, т. е. обладает инерцией, и по-
этому он не останавливается в положении равновесия, а проска-
кивает его. Возникающая при этом возвращающая сила замед-
ляет движение до полной остановки, и далее процесс повторяет-
ся — происходит простое гармоническое колебание Вид струны в
Рис 86 Если закрепленную на концах натянутую струну отклонить в сторону
(Д) и отпустить, то она начнет колебаться, последовательно занимая ряд положе
ний, два из которых показаны на рисунках Б и В Такое движение называется
стоячей волной Изображенное колебание является основным и происходит с уг-
ловой частотой <0о = (л/1) V(F/М). На рисунках Г и Д представлены две более
высокие гармоники с частотами 2<Оо и 4ш0, содержащие соответственно одни и
три узла по длине струны. В общем случае колебание представляет собой нало-
жение многих гармоник.
разные моменты времени показан на рис 8 6, Б и В. Тип движе-
ния, при котором струна закреплена на концах и каждый ее эле-
мент совершает простое гармоническое колебание, называется
стоячей волной. Частоту колебаний можно оценить, прибегнув
к анализу размерностей. Частота может зависеть только от трех
величин: от возвращающей силы, обусловленной в свою очередь
натяжением струны F (единица измерения силы — кг-м-с~2), от
инерции, определяемой массой М единицы длины струны (еди-
ница— кг-м-1), и от геометрии системы, т. е. длины струны I
(единица — м). Получить из трех этих величин величину с раз-
мерностью частоты (единица — с-1) можно единственным спосо-
бом, а именно составив комбинацию вида ^jFIMll, и именно ей
должна быть пропорциональна частота колебаний струны. В дей-
ствительности для закрепленной на обоих концах струны конеч-
ной длины существует бесконечное число частот возможных коле-
баний, значения которых есть в> = ял/FjM/I, 2n^F[M /I,
Зпл/FjM/l и т. д. Отметим, что, хотя число частот бесконечно, они
отстоят друг от друга на конечную величину. Эти частоты назы-
ваются собственными в том смысле, что, каким бы ни было на-
чальное возмущение, последующее движение струны будет пред-
Гл. 8. Колебания и волны 141
ставлять собой суперпозицию (т. е. результат наложения друг на
друга) колебаний именно с этими, свойственными струне часто-
тами (см. разд. 8 6). Колебание с частотой л у/Г/М // называется
основным колебанием. Если струна колеблется только с этой час-
тотой, то она движется вверх и вниз, как это показано на
рис. 8.6,Л, Б, В. Колебания, происходящие с более высокими соб-
ственными частотами, называются высшими гармониками. Первая
гармоника, частота которой равна 2л -у/F/M /1, предполагает та-
кое движение, при котором средняя точка струны остается непод-
вижной (рис. 8.6,Г); такая точка называется узлом. Следующие
Рис 8 7 Распространение бегущей волны в бесконечной натянутой струне Точку
Р(х = 0) заставляют колебаться, она совершает простое гармоническое движе-
ние Начальное смещение (Л) приводит к тому, что элемент ВС действует с не-
которой силой на элементы АВ и CD, стремясь сместить их за собой Элемент ВС
возвращается к положению равновесия но, не останавливаясь, проскакивает его
Непосредственно после этого струна выглядит так, как показано на рисунке Б
Еще через некоторое время струна приобретает вид, представленный на рисунке
В, причем возмущение распространяется в обе стороны со скоростью с
гармоники имеют больше неподвижных точек (узлов) (рис 8.6, Д).
Колебание произвольного вида, являясь суперпозицией основного
колебания и всех его гармоник, обычно узлов не имеет.
Рассмотрим теперь очень длинную струну, один из элементов
которой, скажем ВС (рис. 8 7, А), заставляют непрерывно совер-
шать простое гармоническое колебание с угловой частотой ю и
небольшой амплитудой а. В начальный момент струна выглядит
так, как это изображено на рис. 8.7, Л. Поскольку в струне есть
натяжение, элемент ВС будет действовать на соседние элементы
АВ и CD, создавая направленные вверх результирующие силы,
под действием которых они также начнут двигаться вверх. Эти
элементы из-за инерции будут продолжать движение вверх и
тогда, когда элемент ВС начнет возвращаться к положению рав-
новесия, и вскоре картина станет примерно такой, как изображено
на рис. 8.7, Б. Элементы АВ и CD в свою очередь вызовут смеще-
ние вверх соседних элементов струны, тогда как сами устремятся
к исходному положению и «проскочат» его. Этот процесс непре-
рывно повторяется, и колебательное движение распространяется
142 Часть 1. Основы механики
вдоль струны в виде бегущей волны. Каждый элемент струны со-
вершает простое гармоническое колебание, но, чем дальше распо-
ложен он от точки начального возмущения Р, тем позже начинает
колебательный цикл; это означает, что растет отставание по фазе
колебаний этих элементов от колебания в точке Р. Частота и ам-
плитуда колебаний каждого элемента равны частоте и амплитуде
колебаний, заданных элементу в точке Р. Чтобы оценить скорость,
с которой волна распространяется по струне в обоих направле-
ниях, мы можем, как и прежде, использовать анализ размерно-
стей. Единица измерения скорости распространения волны (обо-
значим ее через с) есть м-с-1, а единственные величины, от кото-
рых эта скорость может зависеть, суть F, М и со. Получить из них
величину с требуемой размерностью можно только одним спосо-
бом, и оказывается, что с должна быть пропорциональна ^FfM
(более строгий расчет показывает, что с в точности равна -yjFIM>)
последнее выражение не зависит от частоты со.
Мы не будем выводить выражения для скорости волны и дру-
гих величин (вывод этот довольно сложен), но поскольку далее
нам часто придется прибегать к математическому представлению
бегущей волны, его мы здесь получим. Рассмотрим точку X
(рис. 8.7), расположенную справа от Р на расстоянии х (х здесь
положительно), и движущуюся слева направо волну, проходящую
через эту точку. В первый момент возмущение в точке равно
нулю; оно достигает X только через время х/с, и после прохожде-
ния «фронта волны» элемент струны в точке X совершает простое
гармоническое движение с частотой со и амплитудой а. Тогда по-
перечное смещение элемента в точке X в момент времени t (обо-
значим смещение через у) задается выражением *)
у~а cos {со (/ — х/с)}* 2). (8.10)
Это выражение описывает картину смещения, распространяюще-
гося вправо со скоростью с. Чтобы убедиться в этом, рассмотрим
графики зависимости у от х для двух последовательных моментов
времени t\ и t% (рис. 8.8). Для произвольного Х| в момент времени
t\ смещение такое же, как в точке x = xi4-c(f2 —t\) в момент
времени /2, поскольку величина (t — х/с) имеет в обоих случаях
’) Подчеркнем, что рис. 8 7 и пояснение к нему относятся к процессу воз-
никновения волны, формула (8 10) — к установившемуся в конце концов волно-
вому процессу.— Прим. ред.
2) Выражение (8.10) предполагает, что в начале координат в момент t = 0
у — 0, a dyldt = —иа. Если это не так, необходимо либо изменить определение
момента времени, который мы принимаем за начало отсчета (/ — 0), либо доба-
вить в фигурные скобки постоянный фазовый угол ф:
у == a cos {и (t — х/с) — ф).
Тогда
у = a cos ф при х = 0 и / = 0.
Гл. 8. Колебания и волны 143
Рис. 8.8. Графики заданной уравнением (8.10) зависимости у от х, которые по-
строены для Двух последовательных моментов времени Г и 1%. Кривая для мо-
мента /2 целиком сдвинута вправо на расстояние с(/2— ti): значение у, получае-
мое из уравнения (8.10) для положения xi и времени tt, в точности равно зна-
чению у для положения xt + с(/2 — й) и времени /2. Таким образом, волна рас-
пространяется со скоростью с. Длина волны равна 1.
одно и то же значение (tt— xi/c). Полученный результат озна-
чает, что за интервал времени t2— Л вся картина целиком сдви-
гается вправо на расстояние c(t2 — ti), а следовательно, волна
распространяется со скоростью с.
Длина волны X в случае, представленном выражением (8.8)
или графиком на рис. 8.8, есть расстояние по оси х, занятое одним
полным колебанием. Оно равно расстоянию, пройденному волной
за время одного периода колебания, который в свою очередь со-
ставляет 2л/со, так что длина волны, равная произведению ско-
рости волны на период, есть
Х = 2лс/со. (8.11)
Как и для простого гармонического колебания одной частицы,
для существования волн необходимо наличие возвращающей силы
и инерции. В приведенном примере возвращающая сила обуслов-
лена натяжением струны, а инерция — ее массой, приходящейся
на единицу длины. В случае волн на поверхности жидкости (за
исключением очень коротких волн, когда существенным становит-
ся поверхностное натяжение) возвращающей силой является сила
тяжести, а инерция связана с наличием у жидкости конечной плот-
ности *)•
) Скорость распространения волн с по поверхности глубокого бесконечного
океана в случае, когда поверхностное натяжение несущественно, может зависеть
только от ускорения силы тяжести g, плотности воды р и длины волны Л.
Из анализа размерностей следует, что с должно быть пропорционально V&X.
Аналогично, если преобладает поверхностное натяжение, а сида тяжести не-
144 Часть I. Основы механики
Распространение звуковых волн в жидкости определяется плот-
ностью среды и ее объемной упругостью, которая вынуждает среду
возвращаться в исходное состояние после любого локального сжа-
тия. Волны давления распространяются и вдоль упругих стенок
артерий, при этом возвращающая сила обусловлена упругостью
стенок, а инерция — содержащейся в сосуде кровью (и в какой-то
степени самой стенкой); в гл. 12 мы покажем, как в терминах рас-
пространения волн может быть описана в основных чертах меха-
ника течения крови в артериях.
8.3. Затухание
Все обсуждавшиеся выше колебательные движения обуслов-
лены равновесием между инерцией и возвращающей силой; при
этом никакие другие силы, действующие на частицу, мы не рас-
сматривали. В таких условиях колебание маятника будет продол-
жаться как угодно долго, а бегущая волна будет распространяться
бесконечно далеко без уменьшения амплитуды. Однако все реаль-
ные колебания затухают, если для их поддержания не приклады-
ваются дополнительные внешние силы; это связано с тем, что в
системе всегда присутствуют силы другого рода, называемые демп-
фирующими. Все демпфирующие силы обусловлены наличием тре-
ния или вязкости в той или иной форме. Колебания маятника
затухают из-за сопротивления воздуха и трения в подвесе; волны
на воде и звуковые волны затухают из-за наличия вязких напря-
жений; колебание струны затухает вследствие сопротивления воз-
духа (вязкие напряжения в окружающем воздухе), а также слабо
выраженных вязкоупругих свойств материала струны. Вечное дви-
жение в этом случае невозможно.
Одним из подходов к рассмотрению явления затухания яв-
ляется энергетический подход. Стоячая волна, как и любое дру-
гое колебание, при котором частица совершает простое гармониче-
ское движение без затухания, несет постоянное количество меха-
нической энергии; это следует из принципа сохранения энергии
[уравнение (2.10)]. Можно доказать, что сумма кинетической и
потенциальной энергий маятника (рис. 8.2), движение которого
существенна, с может зависеть только от р, 1 и о (а — коэффициент поверхност-
ного натяжения на границе воздух — вода, имеющий размерность [МТ-2]).
В этом случае анализ размерностей показывает, что с должно быть пропор-
ционально V°/P^- Когда обе эти оценки скорости волны дают близкие значения,
существенны и сила тяжести, и поверхностное натяжение. Это имеет место в
том случае, если длина волны примерно равна величине Лс, где Лс = V°/P£
Для границы раздела воздух — вода значение о равно примерно 0,07 кг-с~2,
.поэтому Хс составляет 0,002—0,003 м (2—3 мм) Если длина волны намного
больше, чем Хс, поверхностное натяжение несущественно, что, как правило, и
рмрет место для волн ца поверхности моря.
Гл. 8. Колебания и волны 14 g
описывается выражениями (8.5) и (8.6), является величиной по-
стоянной и равна
£ = ±а2-^-. (8.12)
A t
Демпфирующие силы, коль скоро они имеются, всегда направ-
лены так, чтобы противодействовать движению, и совершаемая
ими работа оказывается отрицательной, т. е. вследствие трения
механическая энергия отбирается из системы. Эта энергия пре-
вращается в тепло, и хотя в принципе можно снова превратить это
тепло в механическую работу, мы никогда не получим при этом
столько же механической энергии, сколько перешло в тепло (это,
грубо говоря, есть второй закон термодинамики). Если затухание
происходит чрезвычайно медленно, так что практически система
в каждый момент совершает простое гармоническое колебание, то
общее количество энергии в данный момент по-прежнему будет
определяться выражением (8.12), но амплитуда а этих колеба-
ний будет со временем постепенно уменьшаться, пока не станет
совсем ничтожной.
Если колебания настолько малы, что возвращающая сила про-
порциональна смещению (именно такая ситуация и рассматрива-
лась нами ранее), то обычно столь же хорошим приближением
оказывается представление демпфирующей силы как силы, про-
порциональной быстроте смещения, т. е. скорости. Когда частица
совершает простое гармоническое движение (как, например, в
простом маятнике), к правой части уравнения (8.1) необходимо
добавить член, соответствующий дополнительной силе и имеющий
вид — Cdy/dl, где С — константа. Эта запись означает, что сила
пропорциональна скорости dy/dt и противоположна ей по на-
правлению. Теперь уравнение движения частицы содержит три
члена, а не два, как уравнение (8.1): произведение массы на уско-
рение равно сумме возвращающей и демпфирующей сил. Оно
имеет вид
и может быть переписано в следующей форме:
-§- + 2₽-^- + И2£/=0, (8.13)
где
2р = С/т, ю2 = К/т.
В отсутствие затухания (8.13) совпадает с уравнением (8.2).
- Здесь мы рассмотрим не наиболее общий случай движения,
удовлетворяющего уравнению (8.13), а вернемся еще раз к част-
ному случаю, когда частица начинает двигаться из точки равно-
146 Часть I. Основы механики
весия (у — 0) с данной скоростью V; в математических символах
эти условия имеют вид
г/ = 0, ПРИ ^==0- (8-14)
В отсутствие затухания (р = 0) движение в этом случае описы-
вается выражениями (8.7) и (8.8). При наличии затухания ре-
шение уравнения (8.13), а следовательно, и движение частицы
имеют различные формы в зависимости от того, каков параметр
0— меньше частоты ю, равен ей или больше нее. Рассмотрим три
этих случая раздельно.
а) Р < и.
В этом случае решение имеет вид
у = е~₽* sin Ю1/0 , (8.15)
где Ю] = V®* 2 — ₽2 и называется собственной частотой затухаю-
щего колебания. График зависимости у от t для этого случая (при
некоторых конкретных значениях 0 и ю) приведен на рис. 8.10, Л.
Движение по-прежнему остается колебательным с частотой Ю] (пе-
риодом Ti = 2л/(01), но его амплитуда а задается выражением
а = Хе"₽', (8.16)
т. е. уменьшается в е₽Г| раз за каждый период (множитель ерГ1
часто называют коэффициентом затухания). Рассмотренный пример
иллюстрирует затухающие колебания2).
) Число е равно примерно 2,718. Графики функций у = е*, у = и
у = 1 — е~* приведены на рис. 8.9. Когда х становится бесконечно большим,
е* (экспоненциальная функция) стремится к бесконечности, а — к нулю.
Оказывается, что стремится к нулю быстрее, чем х в любой отрицательной
степени, скажем х-100 или х-1000 °00. Особенность функции е* заключается в том,
что она равна по абсолютной величине своей производной, т. е. d/dx(e*) = ех,
d/dx(e-*) = —е~х. С этим связано столь частое появление экспонент при решении
задач, подобных описанной выше.
2) Подобным образом часто анализируют'работу используемых в физиологи-
ческих исследованиях датчиков давления, которые соединены с заполненными
жидкостью насадочными иглами или катетерами. Диафрагма датчика действует
как пружина и обычно проскакивает положение равновесия, когда смещается под
действием давления; инерция при этом обусловлена по большей части содержа-
щейся в катетере жидкостью. «Переброс» затухает за счет вязкости жидкости в
катетере. Величина и скорость спада «переброса» представляют интерес, посколь-
ку очевидно, что именно они определяют степень достоверности записей давления,
получаемых с помощью подобной системы. При изучении кровообращения обычно
применяются системы, заполненные жидкостью, и датчики с жесткой диафраг-
мой; при этом наблюдаются затухающие колебания, аналогичные изображенному
на рис. 8.10, А. Определение коэффициента затухания н собственной частоты
затухающих колебаний этой измерительной системы с помощью специальной ка-
либровки позволяет предсказывать поведение системы в широком диапазоне ча-
стот,
Гл. 8. Колебания и волны 147
б) Р > (О.
В этом случае движение описывается выражением
!/ = -^-е-3Че"'-е-"'), (8.17)
где п = V₽2 — ®2 и> следовательно меньше, чем р.
График зависимости у от t для конкретных значений р и со
приведен на рис. 8.10,5. Видно, что в этом случае колебания от-
сутствуют: маятник, отклонившись один раз, возвращается назад
и мягко останавливается в исходном положении равновесия. Строго
говоря, движения, описываемые выражениями (8.15) и (8.17), не
затухнут полностью, пока не пройдет достаточно много времени,
так как е~₽' становится равным нулю лишь при /~>оо. Однако на
практике такие движения становятся незаметными через конечное
время; например, амплитуда движения, заданного выражением
(8.16), уменьшается в 100 раз за время, для которого р/= 4,6;
поэтому приемлемой оценкой времени затухания может служить
выражение
/ = 4,6/р. (8.18)
Полностью задемпфированное движение, описываемое выраже-
нием (8.17) (оно называется колебанием со сверхкритическим за-
туханием), представляется суммой двух экспоненциально убываю-
щих членов. Тот из них, который убывает медленнее, имеет вид
так что для него время уменьшения амплитуды в 100 раз
равно 4,6/(р — п). Интересно отметить, что, когда р очень велико
по сравнению с со (т. е. в случае чрезвычайно сильного затуха-
ния), п примерно равно р, а р — п чрезвычайно мало. В самом
деле, р — п при этом примерно равно со2/2р, а время затухания
составляет 8,бр/со2; оно очень велико по сравнению с периодом
незатухающих колебаний 2л/ю (о> называют собственной частотой
незатухающих колебаний). Таким образом, если мы хотим как
можно быстрее задемпфировать нежелательные колебания, то не
следует делать коэффициент р очень большим; на самом деле для
быстрейшего затухания (без «переброса») р должно как можно
меньше превышать со.
в) р = со.
Это критический случай, промежуточный между а) и б). Движе-
ние описывается выражением
y==Vte-^\ (8.19)
График этой функции, представленный на рис. 8.10, В, не слишком
сильно отличается от графика рис. 8.10,5; данный случай интере-
сен только тем, что именно здесь р принимает то минимальное
значение (равное со), при котором еще отсутствуют колебания.
Подобное движение называют колебанием с критическим затуха-
нием.
148 Часть 1 Основы механики
Рис 8 10 Графики зависимости у от t для у, заданного уравнением (8 15) при
р=-^-и(Л), уравнением (817) при Р = 2со(Б) и уравнением (819) при
р = со (В). Во всех случаях V/со принято равным одной единице длины.
Гл 8. Колебания и волны 149
Аналогично простым гармоническим колебаниям, затухающим
во времени, бегущие волны, вызванные колебаниями данной ампли-
туды и частоты, затухают (ослабляются) в пространстве. Если мы
будем двигаться вместе с волной со скоростью с, то увидим, что
волна затухает во времени с неким временем затухания Т3. Ампли-
туда волны уменьшается по мере удаления от начала координат,
и за приемлемую меру «расстояния затухания» можно принять ве-
личину сТ3- Во многих интересующих нас случаях (волны в на-
тянутой струне, волны на воде, звуковые волны) ослабление мало,
и волны наблюдаются на расстоянии многих длин волн от начала
координат, так что рассматривать волны с критическим или сверх-
критическим затуханием нет необходимости (существуют, однако,
исключения — например, волны на поверхности густого сиропа, ко-
лебания давления в микрососудах; разд. 13.4). Степень затуха-
ния на расстоянии, равном длине волны, для волн давления в ар-
териях весьма велика, но, как мы увидим в гл. 12, в действитель-
ности в этом случае длина волны настолько превышает длину
самой артерии, что реально наблюдаемое затухание в артерии не
столь уж значительно.
Затухающее простое гармоническое колебание отличается от
незатухающего наличием экспоненциально убывающего члена.
В этом же состоит отличие и затухающей бегущей волны от неза-
тухающей. Затухающая волна частоты со, соответствующая чисто
синусоидальной волне, описываемой выражением (8 10), представ-
ляется следующим соотношением, связывающим смещение у с хи/:
у = ae~kxlK cos {со (/ - х/с)}. (8.20)
Это выражение того же вида, что и (8 10), — в обоих случаях рас-
пространение колебания описывает член cos{ }, а отличие заклю-
чается в том, что амплитуда колебаний экспоненциально умень-
шается с увеличением х. В выражении (8.20) X — длина волны,
a k — константа, характеризующая затухание: амплитуда cze~ft*/z
уменьшается в 100 раз на расстоянии 4,6/6 длин волн. Другими
словами, на расстоянии, равном длине волны, амплитуда умень-
шается в е* раз (величина е* называется коэффициентом затуха-
ния).
8.4 Отражение волн и резонанс
Рассмотрение затухающих и незатухающих бегущих волн было
основано на предположении, что среда, по которой они распро-
страняются (в нашем примере — натянутая струна), имеет беско-
нечную протяженность, и поэтому препятствия для неограничен-
ного распространения волн отсутствуют. Конечно, в действитель-
ности ни одна реальная система не является бесконечно протя-
женной, и поэтому мы должны выяснить, что же происходит,
150 Часть 1. Основы механики
когда бегущая волна встречается с препятствием. Предположим,
что длинная натянутая струна, которую мы уже рассматривали
раньше в качестве примера, приводится в колебательное движе-
ние на одном конце, а второй ее конец (х = /) надежно закреп-
лен, так что в этом месте невозможны никакие смещения
(рис.8.11). Посмотрим, что произойдет со смещением у, которое
раньше задавалось выражением (8.10), предполагающим, что все
точки струны, включая и точку х = I, колеблются с амплитудой
Конец, который
вьнуждаот
колебаться
Рис. 8.11. Струна конечной длины, один конец которой закрепили, а другой вы-
нуждают колебаться с частотой и Наложение падающей волны на отраженную
создает стоячую волну. В узлах N смещения не происходит; в пучностях А ам-
плитуда колебания максимальна.
±а. Для анализа удобно перенести начало координат в точку
х= I, положив х — 14- х', где х' — новая координата, отрицатель-
ная во всех точках струны. Тогда выражение (8.10) приобретет
вид
у = a cos {<о (/ — x'lc)}. (8.21)
Теперь точка закрепления, в которой смещение у должно быть
равно нулю, имеет координату х' = 0. Оказывается, что наличие
закрепления приводит к наложению на исходную волну (называе-
мую падающей) еще одной волны, имеющей такую же амплитуду
и скорость, но распространяющейся в противоположном направ-
лении. Это есть отраженная волна. Полное уравнение для смеще-
ния принимает теперь вид
у — a (cos {со (/ — х'/с)} — cos {со (I + х'/с)}]. (8.22)
На фиксированном конце (х' = 0) и во всех точках, отстоящих от
него на расстояние, кратное длине полуволны (х' = —лс/ю,
—2лс/(о, —Злс/to и т. д.), смещение в каждый момент времени
равно нулю; посредине между этими узлами находятся точки
(пучности-, см. рис. 8.11), где амплитуда смещений при колеба-
Гл. 8. Колебания и волны 151
ниях равна удвоенной исходной. Фактически подобное движение
является стоячей волной; чтобы показать это, достаточно преоб-
разовать (8.22) к виду
г/= 2а sin ю/sin. (8.23)
При каждом данном значении х' смещение у колеблется, как в
простом гармоническом движении, причем амплитуда колебания
пропорциональна величине sincox'/c, которая, как видно из этого
выражения, равна нулю в узлах и единице в пучностях. Колеба-
ния не распространяются, и теперь ясно, что они представляют
собой стоячую волну.
Падающая волна, амплитуда а
Отраженная волна, амплитуда Па
Рис. 8.12. Отражение при подвижном концевом креплении струны (например,
при наличии шарнира), в котором происходит рассеяние энергии. Амплитуда от-
раженной волны меньше, чем падающей.
Рассмотренный случай является примером полного отражения,
при котором амплитуда (а следовательно, и энергия) отраженной
волны равна амплитуде (и энергии) падающей. Обычно, однако,
отражается не вся энергия падающей волны — некоторая ее часть
либо поглощается, либо передается дальше. Предположим, что
натянутая струна закреплена не неподвижно, а в держателе, ко-'
торый в принципе может перемещаться, но этому перемещению
противодействует сила трения, обусловливающая рассеяние энер-
гии (рис. 8.12). Тогда некоторая часть энергии падающей волны
будет теряться из-за этого рассеяния и отраженная волна будет
обладать меньшей энергией, т. е. меньшей амплитудой. Это при-
мер частичного отражения. Если бы амплитуда отраженной вол-
ны, описываемой выражением (8.22), составляла, скажем, только
80% от амплитуды падающей волны, то смещение имело бы вид
и — a [cos {со (/ — х'/с)} — R cos {со (/ + х'/с)}], (8.24)
где R — константа, равная в данном случае 0,8. Иногда R назы-
вают коэффициентом отражения1). Падающая волна частично от-
*) Здесь R есть отношение амплитуд отраженной и падающей волн. Однако
чаще коэффициент отражения определяют как отношение потока энергии в от-
раженной волне к потоку в падающей, а эта величина равна R2. Важно зиать,
какое из этих определений подразумевается в каждом конкретном случае, когда
привлекается понятие коэффициента отражения.
152 Часть 1. Основы механики
ражается и тогда, когда в среде, где она распространяется, имеет-
ся неоднородность (примером может служить локальное измене-
ние плотности или толщины струны) или когда в каком-либо ме-
сте возникает препятствие для волнового движения (например,
бусинка, закрепленная в середине струны; см. рис. 8.13). В этом
случае волна продолжает распространяться вдоль струны и по
другую сторону неоднородности, но с меньшей амплитудой. Это
прошедшая волна-, ее амплитуда равна произведению а на число
Т, меньшее единицы. Иногда Т называют коэффициентом прохож-
дения, но чаще это понятие относится к величине Т2. Если энер-
гия при прохождении через препятствие не поглощается, то Т2 —
= 1 — R2.
Падающая волна (амплитуда а)
* Частица
Прошедшая волна
(амплитуда Та)
Отраженная волна (амплитуда На)
Рис. 8.13. Отраженная и прошедшая волны при наличии в струне неоднород-
ности (в данном случае к струне прикреплена бусинка массы т, которая пре-
пятствует быстрому смещению в силу сиоей инерции).
Источником отражений может стать любая неоднородность в
системе, а если таких неоднородностей много, то волна в произ-
вольной точке будет представлять собой сложный результат на-
ложения основной волны, отраженных и проходящих волн, соот-
ветствующих каждой неоднородности, волн, возникающих при вто-
ричном отражении и прохождении ранее отраженных и прошед-
ших волн и т. д. Процесс отражения, повторного отражения и т. д.
не будет продолжаться бесконечно в основном из-за затухания
волн, а частично вследствие поглощения энергии на границе. Та-
кое поглощение имеет место при плохой обработке шарнирного
концевого держателя в приведенном выше примере нежесткого
крепления натянутой струны или в том случае, когда волны набе-
гают на песчаный или галечный пляж (такой берег почти пол-
ностью поглощает энергию волн).
В разд. 8.2 говорилось о том, что закрепленная на концах на-
тянутая струна конечной длины может совершать свободные коле-
бания только с определенными собственными частотами. Если же
некоторую точку струны заставить колебаться с частотой ю, не
равной ни одной из собственных частот, то волны будут распро-
страняться от этой точки к обоим концам, отражаться там, вновь
отражаться на противоположных концах и т. д., и установившееся
в результате движение будет представлять собой стоячую волну,
частота которой равна частоте воздействия <о. Если, однако, час-
Гл. 8. Колебания и волны 153
тота со равна одной из собственных частот, то картина резко ме-
няется. В этом случае фазы падающих и отраженных волн таковы,
что волны значительно усиливают друг друга при каждом отра-
жении, а ампли’гуда колебаний продолжает расти до тех пор, пока
диссипативные эффекты, которые также растут, не станут суще-
ственными. Кроме того, в этом случае система обычно перестает
быть линейной (см. ниже). Колебания с собственной частотой не-
сут определенную энергию, и, если дополнительная энергия в си-
стему не поступает, амплитуда этих колебаний остается постоян-
ной. В случае же подвода энергии извне амплитуда катастрофи-
чески увеличивается. Подобное явление называется резонансом.
На практике амплитуда резонансных колебаний ограничена дис-
сипативными потерями (трением) или нелинейностью, но, пока
эти эффекты не слишком велики, она остается гораздо больше,
чем амплитуда исходного смещения под действием вынуждающей
силы. В одних случаях резонансные явления оказываются полез-
ными, в других — весьма нежелательными. Музыкальные инстру-
менты, в частности, устроены таким образом, что из широкого
диапазона вынужденных колебаний (например, колебаний язычка)
резонансными являются только некоторые; поэтому они и усили-
ваются до уровня слышимости. Напротив, в артериальной системе
резонанс нежелателен, так как необычно большие колебания дав-
ления могут сильно изменить характер течения крови и вызвать
повреждение ткани.
8.5. Линейность
В гл. 12 мы рассмотрим применение изложенных выше сооб-
ражений о свойствах волн при анализе волновых явлений в сер-
дечно-сосудистой системе млекопитающих, к которой, как будет
показано, эти и ряд других свойств имеют непосредственное от-
ношение. Важно, однако, уже сейчас отдавать себе отчет в том,
что использованные нами относительно простые подходы справед-
ливы только в том случае, когда амплитуда колебаний достаточно
мала, так что возвращающая сила пропорциональна смещению,
а демпфирующая — скорости смещения. В этом случае колебания
и волны называют линейными. Линейность — сугубо математиче-
ское понятие, но оно настолько важно в теории распространения
волн, что заслуживает более подробного рассмотрения.
Предположим,. что система функционирует так, что какие-то
две переменные величины всегда остаются определенным образом
связанными друг с другом. Это могут быть измеренные в данном
месте скорость и давление крови в артерии, смещение и ускорение
частицы при колебательном движении или масса живого орга-
низма и потребление им кислорода; в принципе это может быть
система самого общего вида. Обозначим две указанные перемен
иые через икр. Связь между икр линейна, если график зависи-
154 Часть I. Основы механики
мости и от р является прямой, в частности, когда и прямо про-
порционально р, т. е.
и = ар, (8.25)
где а — константа. Можно получить также и более общее опре-
деление линейной системы: если щ и р< являются одной парой
значений соответствующих друг другу величин и и р, а и2 и р2—
другой такой парой, то система линейна, если щ и2 и pi + р2
также являются парой значений соответствующих величин. Ясно,
что система, представленная уравнением (8.25), линейна согласно
этому определению1), а система, описываемая, скажем, уравне-
нием и = р2, график которого есть парабола, таковой не является.
Аналогичным образом система, определяемая уравнением (8.2),
которое описывает простое гармоническое движение, и соответ-
ствующими начальными условиями, также является линейной. Мы
видели, например, что решение у — a cos го/ соответствует на-
чальному смещению а [выражение (8.5)], а решение у = — sin го/—
начальной скорости V [выражение (8.7)]. Легко проверить, что
у
сумма этих решений, т. е. у — a cos го/ + — sin го/, соответствует
состоянию, в котором начальное смещение равно а, а начальная
скорость — V. Описанное ранее отражение синусоидальных волн
является процессом линейным, если удвоение амплитуды падаю-
щей волны приводит к удвоению амплитуды отраженной волны
и т. д. Более того, чтобы полное смещение представляло собой
просто сумму смещений, связанных отдельно с падающей н отра-
женной волнами, линейной должна быть и среда, по которой рас-
пространяются волны (натянутая струна).
Уравнения, определяющие поведение почти всех реальных фи-
зических систем, в действительности нелинейны. Например, ис-
тинным уравнением, описывающим колебания простого маятника,
является (8.2а), а не (8.2); оно сводится к виду (8.2), только
если угол 6 достаточно мал, так что sin 6 примерно равен 6. По-
грешность этого приближения пропорциональна О3 2) — величине,
гораздо меньшей, чем 6, если 6 много меньше единицы: при 0 = 0,1
03 равно 0,001. Пока 0 остается достаточно малым, реальная не-
линейная система, описываемая уравнением (8.2а), ведет себя
практически как линейная, и мы получим достаточно точное пред-
ставление о поведении реальной системы, анализируя гораздо бо-
лее простую линейную систему. Сведение низкоамплитудной не-
линейной системы к линейной называется линеаризацией. Эта про-
’) Более точно следующее определение: связь между и и р линейна, если
и ар + Ь, где а, b — постоянные; уравнение (8.25) выражает линейную Одно-
родную связь. — Прим. ред.
*) Поскольку приближением для sin 0, лучшим, чем просто 0, является
0 — 03/6.
Г л. 8. Колебания и волны 165
цедура очень часто используется во многих прикладных задачах,
в частности, при исследовании потока крови в артериях, посколь-
ку линейные системы несравненно проще анализировать. Однако
при всех практических приложениях необходимо убедиться, что
отброшенные нелинейные члены уравнения, описывающего реаль-
ную систему, действительно малы по сравнению с линейными.
Если это не так, нелинейности существенны и необходимо оценить
их влияние.
Примером системы, которая обычно трактуется как линейная,
но при определенных обстоятельствах все же проявляет нелиней-
ные свойства, является натянутая струна конечной длины, которую
вынуждают колебаться с частотой, близкой к одной из собствен-
ных частот. Волны, создаваемые источником колебаний, много-
кратно отражаются таким образом, что усиливают сами себя (ре-
зонанс), вследствие чего их амплитуда становится очень большой.
В этих условиях линейное приближение, в котором волны анали-
зировали первоначально, перестает быть справедливым. Деталь-
ный анализ резонансных систем с учетом нелинейностей обычно
очень сложен.
Уравнения механики жидкостей нелинейны из-за члена, пред-
ставляющего конвективные ускорения: примером может служить
уравнение Бернулли (4.6), которое связывает давление р со ско-
ростью и жидкости при стационарном движении в случае, когда
вязкостью можно пренебречь. Предположим, что жидкость течет
в горизонтальной трубке (и поэтому сила тяжести несущественна)
с переменной площадью поперечного сечения (и поэтому и ме-
няется вдоль трубки; разд. 4.6); если трубка еще и упругая, то,
как мы увидим в гл. 12, рассматриваемая система может служить
моделью потока крови в крупных артериях. Предположим также,
что течение колебательное, а уравнение Бернулли можно модифи-
цировать, введя член, пропорциональный локальному ускорению
dw/d/, так что оно будет учитывать нестационарность потока.
Тогда мы получим
y+4«2 + A:-^-=const, (8.26)
где k — константа (с размерностью длины), а р — считающаяся
постоянной плотность жидкости. В точном уравнении, описываю-
щем нестационарный поток, член, представляющий локальное ус-
корение, имеет более сложный вид, но он, как и в рассмотренном
здесь случае, линеен, поэтому уравнение (8.26) является адекват-
ной моделью системы в данном контексте. Если бы член, представ-
ляющий конвективные ускорения (у^2), был пренебрежимо мал,
то уравнение (8.26) свелось бы к виду
-J-+ ft= const, (8.27)
156 Часть I. Основы механики
Из приведенного определения линейности следует, что уравнение
(8.27) линейно, тогда как (8.26) — нелинейно. Предположим, что
в каждой точке скорость жидкости колеблется с угловой частотой
го и пропорциональна, скажем, cos roZ. Если поведение системы
описывается уравнением (8.27), это движение будет сопровож-
даться колебанием давления той же частоты, пропорциональным
sinroZ. Но если определяющим является уравнение (8.26), то из
решения, в котором и пропорционально cos roZ (и, следовательно,
dw/dZ пропорционально sinroZ), будет следовать, что и2 пропор-
ционально cos2roZ, т. е. у+ у cos2coZ. Таким образом, волне ско-
рости, характеризующейся единственной частотой о, соответствует
существование среднего давления и его колебательных компонент
не только с частотой го, но также и с частотой 2го. Как мы увидим
далее, это значительно затрудняет анализ несинусоидальных ко-
лебаний.
8.6. Фурье-анализ
В общем случае периодическое волновое движение обычно не
является чисто синусоидальным. График зависимости смещения
струны от положения х в произвольный момент времени или гра-
фик зависимости скорости крови в данном месте артерии от вре-
мени нередко имеют чрезвычайно сложную форму (см. послед-
ний график на рис. 8.14). Однако такой график может быть пред-
ставлен в виде суммы некоторого набора синусоид с различными
фазами, амплитудами и частотами, кратными основной частоте
го(== 2л/Т), где Т—полный период колебаний. Процесс разло-
жения волны сложной формы на синусоидальные составляющие
называется фурье-анализом. Различные по частоте составляющие,
которые в совокупности создают полную волну, как и прежде, на-
зываются гармониками основного колебания (разд. 8.2). Число
гармоник, необходимых для достаточно точного описания полного
колебания, может быть различным. Например, волна, представ-
ленная на рис. 8.14, которая аналогична по форме волне скорости
крови в артерии, практически может быть представлена в виде
суммы средней величины, основного колебания и трех гармоник.
Соответствующая формула имеет вид
и = Ло 4- AI cos (roZ + 0i) 4- А-> cos (2roZ 4* 62) 4*
4" А3 cos (3roZ 4" Оз) 4~ A4 cos (4roZ 4" 0<)> (8 • 28)
где Ло/Д1 = О,44, Д2/Д1 = 0,97, Д3/Д1 = 0,47, Д4М1 = 0,14, =
=—58° = —1,01 рад, 02=-—151° = —2,64 рад, 03 = 4-124° =
= 2,16 рад, 04 = 4-86°= 1,5 рад. На рис. 8.14 показано, каким
образом, складывая эти составляющие, мы получаем полную
форму волны.
Гл. 8. Колебания и волны 157
Фурье-анализ можно применять для волн любой формы. Эта
процедура значительно упрощает изучение колебаний и волн в
линейных системах, но для1 анализа нелинейных систем она мало-
пригодна. Предположим, что мы хотим с помощью уравнения
(8.27) рассчитать давление крови р в артерии, измерив скорость
и, которая носит колебательный характер. Для этого мы можем
представить скорость в виде суммы синусоидальных составляю-
щих, как это сделано в уравнении (8.28). После этого можно при-
Рис. 8.14. Графическое представление периодического волнового движения, ана-
логичное графику зависимости скорости крови в артерии от времени. (McDonald.
Blood flow in arteries, p. 129. Edward Arnold, London) Показано, как при сло-
жении средней величины, основного колебания и трех гармоник можно пол-
ностью воссоздать форму волны. Аналитически каждая из этих составляющих
дается уравнением (8.28) )•
влечь уравнение (8.27) и рассчитать колебание давления, соответ-
ствующее колебанию одной из составляющих скорости. Так, для
составляющей и = Ап cos (пи/ + 0„) это уравнение, в котором
const = 0, даст р = pknmAn sin (пи/ -f- 0„) [константа в правой
части уравнения (8.27) равна средней величине левой части, т. е.
ро/р]. Сложив полученные таким образом колебания давления
всех видов, мы найдем результирующее давление; это сложение
правомочно, поскольку система линейна. Однако в нелинейной
системе, представленной уравнением (8.26), все частотные состав-
ляющие взаимодействуют друг с другом (как в рассмотренном
выше примере). В таком случае независимое сложение решений,
полученных при подстановке в уравнение (8.26) на место и только
одного синусоидального члена, необоснованно.
’) На схеме, как и во многих работах, принята система обозначений, при
которой основное колебание для единообразия называется первой гармоникой, —
Прим, перев.
158 Часть I. Основы механики
Как будет показано в гл. 12, волны давления, распространяю-
щиеся в крупных артериях, являются приблизительно линейными
и их основные параметры могут быть получены из линейной тео-
рии. Однако некоторые характерные свойства этих волн можно
объяснить только с помощью нелинейного подхода. Среди таких
свойств — увеличение в отдельных случаях крутизны фронта волн
давления при распространении по сосудистому руслу, а также на-
личие высокоамплитудных колебаний, благодаря которым про-
слушиваются характерные звуки при сдавливании артерии ман-
жеткой тонометра (так называемые тоны Короткова; разд. 14.4),
Нелинейность наблюдается также в венах (гл. 14).
Г лава 9
ВВЕДЕНИЕ В ТЕОРИЮ МАССОПЕРЕНОСА
Термин «массоперенос», или «массопередача», охватывает ши-
рокий круг процессов, при которых происходит перемещение ве-
щества внутри некой системы. Дать простое определение совокуп-
ности таких явлений довольно трудно; можно только сказать, что
речь идет о движении молекул разных видов внутри систем и о
факторах, влияющих на это движение. Начнем с двух простых
примеров, далеко не исчерпывающих, однако, всего содержания
предмета.
Лужа воды на дороге медленно испаряется; при этом вода, на-
ходящаяся в жидком состоянии, постепенно переходит в окружаю-
щий воздух в виде пара. Скорость испарения зависит от таких
факторов, как влажность, температура воздуха относительно зем-
ли и скорость ветра над поверхностью лужи.
Если кристалл сульфата меди поместить в пробирку с водой,
он начнет медленно растворяться и около его поверхности создает-
ся концентрированный раствор. Со временем растворенное веще-
ство будет диффундировать все дальше и дальше в воду. Ско-
рость растворения кристалла и скорость переноса растворенного
сульфата меди в водную фазу зависят от ряда факторов. Напри-
мер, в горячей воде эти процессы будут протекать быстрее, чем
в холодной, ускорятся они и при встряхивании пробирки.
Данные примеры иллюстрируют два фундаментальных меха-
низма переноса массы внутри одной среды (или фазы) — диффу-
зию и конвекцию (или объемное движение). Они демонстрируют
также важный класс явлений — перенос веществ через границу
между фазами, в данных примерах — через границы газ — жид-
кость и жидкость — твердое тело. В биологических системах часто
наблюдается перенос массы между двумя жидкими фазами, раз-
деленными полупроницаемой перегородкой, например клеточной
мембраной. Такой перенос может протекать как благодаря объем-
ному движению, так и вследствие диффузии, а детали его опре-
деляются структурными и химическими свойствами мембраны и
такими параметрами, как концентрация и давление в каждой
фазе. В этой главе мы покажем, как можно количественно изучать
простые процессы переноса массы, и получим некоторые оценки
для более сложных систем.
160 Часть I. Основы механики
Введем несколько определений. Во-первых, рассмотрим такую
величину, как концентрация. Она может быть определена по-раз-
ному— как массовая концентрация, массовая доля, молярная кон-
центрация, мольная доля (определение моля дано в гл. 3) и т. д.
Все эти определения можно применять с равным успехом, но по-
скольку мы в основном будем рассматривать перенос между жид-
кими и твердыми фазами и внутри них, то будем использовать
молярную концентрацию. Молярная концентрация Сь вещества А
в растворе определяется как число молей этого вещества в еди-
нице объема и обычно выражается в г-моль-л-1. Измерив коли-
чество вещества А, переносимого в данном направлении, мы мо-
жем определить его молярный поток J — число молей, проходя-
щих за единицу времени через единичную площадку, перпендику-
лярную направлению движения вещества. Молярные потоки обыч-
но выражаются в моль • с-1 • см-2. В определение потока входит и
указание направления, в котором происходит данное движение;
поток является величиной векторной, и следует всегда иметь это
в виду.
9.1. Диффузия
В 1827 г. шотландский ботаник Роберт Броун поставил очень
простой, но вместе с тем очень важный эксперимент, который
привел к более глубокому пониманию поведения молекул в жид-
кости. В ходе этого эксперимента Броун обнаружил, что частицы
пыльцы на поверхности воды почти безостановочно и хаотично
движутся во всех направлениях (рис. 9.1). Такое совершенно слу-
чайное, или хаотическое, движение впоследствии наблюдали для
самых разных мелких взвешенных частиц, и сейчас оно называется
броуновским движением. Движение подобных частиц обусловлено
Движением молекул жидкости, в которой они взвешены. Эти моле-
кулы находятся в состоянии постоянного хаотического движения,
в результате чего сталкиваются друг с другом и со взвешенными
частицами. Случайное движение частиц отражает именно это хао-
тическое молекулярное движение. В то время как отдельные мо-
лекулы и их смещения слишком малы, чтобы их можно было
увидеть непосредственно, обусловленное ими движение частиц уже
удается наблюдать. Смещение взвешенной в жидкости частицы яв-
ляется результатом происходящих в каждый момент времени уда-
ров со стороны многочисленных молекул жидкости.
Движение молекул жидкости обусловлено тем, что они обла-
дают кинетической энергией, которая увеличивается с температу-
рой. Естественно, что и смещение частиц возрастает с температу-
рой. Движение частиц зависит также от вязкости жидкости, при-
чем с увеличением вязкости среды смещение уменьшается.
В 1855 г. немецкий физиолог Адольф Фик (его именем назван
метод разведения, используемый для определения минутного объ*
Гл. 9. Введение в теорию массопереноса 161
ема сердца) сформулировал свой известный закон массопереноса,
который гласит, что, как только в среде возникают различия в
концентрации сА некоторого вещества А, образуется поток J этого
вещества в направлении уменьшения концентрации (скажем, в
Рис. 9.1. Схематическое изображение
броуновского движения малой части-
цы на поверхности жидкости. Отдель-
ные смещения изображены прямоли-
нейными отрезками. Частица, нахо-
дившаяся вначале в точке 1, через
некоторое время оказывается в точ-
ке 2. Дх и Д{/ — проекции смещения
на оси х в у.
направлении х), который пропорционален градиенту концентрации
deA/dx. Таким образом,
~ dcA
—D ——
A dx
(9.1)
Константа £)А называется коэффициентом диффузии данного ве«
щества. Экспериментально показано, что это уравнение приме-
нимо для описания диффузии как молекул, так и микрочастиц при
условии, что концентрация и тех и других мала (см. стр. 164).
Таблица 9.1
Сравнение коэффициентов молекулярной диффузии для разных
веществ при комнатной температуре
Вещество
Коэффициент диффузии, см2«с
О2 в воздухе
Н2 в воздухе
О2 в воде
Н2 в воде
Ацетат в воде
Липопротеид низкой плотности в воде
0,21
0,41
1,8 • 10"°
5.1 •
0,8 • 10
10~7—10-8
Значения коэффициентов диффузии варьируют в широких пре-
делах; для газов они, как правило, во много раз выше, чем для
жидкостей. Коэффициенты диффузии зависят от относительного
размера диффундирующих молекул или частиц и молекул среды
(табл. 9.1), а также (как и интенсивность броуновского движения)
от температуры. Можно показать1), что если абсолютная темпе-
') В предположении, что форма и размер молекул не меняются с темпера-
турой, такие факторы, как изменение степени гидратации, здесь не учитываются.
162 Часть I. Основы механики
ратура среды равна Т, то
D = kJ 1ц,
где р — вязкость жидкости (которая в свою очередь также зави-
сит от температуры), a k*— константа, зависящая от молекуляр-
ных характеристик среды.
На первый взгляд кажется, что связать хаотическое движение
частиц пыльцы в опытах Броуна с формальным законом Фика
довольно трудно. Однако можно установить связь между вероят-
ным смещением (Ах) частицы или молекулы за данное время с
коэффициентом диффузии D этого вещества. Представим себе по-
„А Р D
Концентрация с. Концентрация с7 . А х _
Вос
Рис 9 2 Прямоугольный участок поверх-
ности воды; пояснения см. в тексте.
верхность воды с плавающими на ней частицами пыльцы, кото-
рые совершают броуновское движение. Рассмотрим прямоуголь-
ник ABCD на поверхности и прямую PQ (рис. 9.2), рассекающую
его пополам так, что линии АВ н CD находятся от PQ на одинако-
вом расстоянии Ах. Поставим перед собой цель — подсчитать
число частиц пыльцы, которые пересекут прямую PQ с каждой
стороны за небольшой интервал времени. Концентрация частиц
в прямоугольнике ABQP равна Ci (в этом случае концентрация
определяется как число частиц на единичной площади), а в пря-
моугольнике CDPQ — с2. Если среднее смещение частиц за ин-
тервал времени t есть Ах, то слева прямую PQ пересекут только
те частицы, которые исходно находились на прямой АВ или ближе
к PQ. Аналогичным образом частицы, пересекающие PQ справа,
первоначально были не дальше, чем на прямой CD. Кроме того,
только половина из числа частиц, которые могут пересечь эту пря-
мую с каждой стороны, пересекут ее на самом деле, поскольку
в среднем половина частиц в каждом отсеке будет двигаться от
PQ. Таким образом, число частиц, пересекающих PQ слева на-
право, дается выражением
Число частиц = X Площадь ABQP X Локальная концентрация=
= Ci X Площадь ABQP.
Гл. 9. Введение в теорию массопереноса 163
Аналогично общее число частиц, пересекающих PQ справа на-
лево, есть
Число частиц = у с2Х Площадь CDPQ.
Следовательно, суммарное число частиц, пересекших PQ слева
направо за время t, равно
Число частиц = 4- <?1Х Площадь ABPQ —5- с2 X Площадь CDPQ=
£ £
= y(ci — с^'ХаЬх,
где а — длина отрезков АВ и CD. Суммарный поток п (число час-
тиц, проходящих за единицу времени через единичную длину пря-
мой PQ) равен
п == ~ с^'
Умножив числитель и знаменатель правой части этого уравнения
на Ах, получим
___ (Ах)2 ct — с2
п~~ 21 ' Ах ‘
Но если мы рассматриваем изменения концентрации на неболь-
шом расстоянии, то
с2 “ С| de
Ах dx
Таким образом,
Сравнение (9.2) с математическим выражением закона Фика
[уравнение (9.1)] показывает, что
D==-^~ • М
Таким образом, коэффициент диффузии и вероятное расстояние,
на которое частица (или молекула) смещается за данное малое
время, непосредственно связаны друг с другом.
Хотя Фик понимал, что движущая сила процесса диффузии
обусловлена кинетической энергией молекул, физический смысл
коэффициента диффузии был ему неясен. Позднее Нернстом было
показано, что если считать молекулы сферическими частицами, то
коэффициент диффузии связан с действующими на них силами
трения следующим образом:
D = RT/fN,
(9.4)
164 Часть I. Основы механики
где R — универсальная газовая постоянная, Т — абсолютная тем-
пература, N — число Авогадро (число молекул в 1 моле), f—сила
трения, противодействующая движению молекул. Если диффун-
дирующие молекулы велики по сравнению с окружающими их мо-
лекулами среды, то для представления силы трения можно при-
влечь закон Стокса [уравнение (6.1)]
/ = 6лр.а, (9.5)
где р.—вязкость жидкости, а — «эффективный» радиус молекулы,
обычно называемый радиусом Стокса — Эйнштейна. Это не истин-
ный радиус молекулы, а радиус эквивалентной сферической час-
тицы, проявляющей такую же диффузионную способность. Объ-
единив уравнения (9.4) и (9.5), мы получим выражение для а:
RT /л
о = -7—тттг • (9.6)
Таким образом, радиус а можно определить, измерив коэффициент
свободной диффузии данного вещества в растворе.
До сих пор мы молчаливо предполагали, что концентрация
диффундирующих веществ низка (подобные растворы называются
разбавленными). Считалось, что диффундирующие молекулы стал-
киваются только с молекулами жидкости, в которой они раство-
рены. Однако, если концентрация раствора растет, увеличиваются
и взаимодействия между молекулами диффундирующего вещества,
в результате чего уменьшается активность молекул, а, следова-
тельно, в какой-то степени и их диффузионная способность. Об-
суждение деталей механизма этого явления и величины самого
эффекта выходит за рамки данной книги; читатель должен просто
знать, что этот эффект существует и может быть важен в тех
случаях, когда перенос вещества происходит в концентрирован-
ных растворах.
9.2. Коллоидное состояние
Дать точное определение этому состоянию довольно трудно, но
в данном случае это и не столь уж необходимо. Коллоидные рас-
творы, или золи, как их обычно называют, представляют собой
растворы либо крупных молекул, либо агрегатов из более мелких
молекул. Между золем и суспензией частиц (или эмульсией, кото-
рая является суспензией жидких капель) нет принципиального
различия. Если молекулы или частицы растворенного вещества
можно увидеть под световым микроскопом, то считается, что это
суспензия, если нельзя — золь.
Часто оказывается, что отдельные крупные молекулы золя на-
ходятся не в «простом» растворенном состоянии, а адсорбируют
на своей поверхности молекулы растворителя, образуя с ними
комплексы, гораздо более крупные, чем сами исходные молекулы.
Гл. 9. Введение в теорию массопереноса 165
Склонность крупных молекул присоединять («впитывать») воду
зависит от распределения электрического заряда на их поверхно-
сти и от pH окружающей среды. Если постепенно удалять сво-
бодный растворитель из золя, то в конце концов золь затвер-
деет и превратится в гель. При этом абсорбированный раствори-
тель остается в твердой матрице, как в ловушке, причем его уда-
ление приводит к разрушению структуры геля.
Гели часто встречаются в биологических системах, например,
они заполняют межклеточное пространство (разд. 13.2). Здесь со-
держатся различные макромолекулы, обладающие очень большим
сродством к воде, вследствие чего их можно считать находящи-
мися в коллоидном растворе.
9.3. Коэффициенты массопередачи
При исследовании многих реальных систем, в частности био-
логических, бывает неудобно, а нередко и просто невозможно,
анализировать скорости переноса в терминах молекулярной диф-
фузии. Это, в частности, относится к диффузии веществ через
клеточные мембраны, когда часто неизвестны ни эффективная
Рис. 9.3. Схематическое изображение
трехслойной мембраны. Показан про-
филь концентрации в стационарных
условиях.
площадь, через которую осуществляется диффузия, ни толщина
мембран. Чтобы преодолеть подобные трудности, принимают, что
при данной разности концентраций ЛсА в отсеках по разные сто-
роны мембраны поток / вещества А через нее определяется соот-
ношением
/ = КДвА, (9.7)
где К. называется коэффициентом массопередачи. Таким образом,
при этом подходе все диффузионные пути рассматриваются в со-
вокупности и характеризуются одним общим коэффициентом. Диф-
фузионное сопротивление мембраны определяется как величина
1/Л.
Во многих биологических системах диффузия идет через ряд
последовательно расположенных клеточных мембран и слоев жид-
кости. Рассмотрим систему с тремя слоями, схематически изо-
браженную на рис. 9.3. Суммарная разность концентраций в ней
равна ci — Cf, но распределение концентрации внутри системы
166 Часть I. Основы механики
(трехслойной мембраны) не является простой линейной функ-
цией координаты, а сильно зависит от относительных значений
коэффициентов массопередачи для трех слоев. Ясно, что в ста-
ционарном случае поток J через все три слоя должен быть оди-
наковым. Последовательно используя уравнение (9.7) для каж-
дого из трех слоев, получаем
1 — с2
Л1 “ / *
1 ___ С2 — Сз
k2 J '
1 _ с3 — Ct
k% J
где ki, kz, kz — коэффициенты массопереноса для первого, второго
и третьего слоев соответственно. Сложив три этих уравнения, по-
лучим
1 г 1 г 1 1 1 А
+ k2 + k3~ J (C1 Ci’~
Суммарный коэффициент массопереноса К для многослойной
мембраны можно определить как
1 Ct — Ct
К J ‘
Тогда мы получим, что
1 = 1 , 1 1
К kl ^2 ^3
„ 1 .1
Поскольку у есть сопротивление переносу слоя 1, у — сопро-
тивление слоя 2 и т. д., то общее сопротивление переносу равно
сумме сопротивлений отдельных слоев мембраны. Ясно, что если
сопротивление одного из слоев очень велико по сравнению с дру-
гими, то полное сопротивление и полная скорость массопередачи
определяются переносом именно через этот слой мембраны. Раз-
ность концентраций с двух сторон подобной части мембраны близ-
ка к суммарной разности концентраций; про такую часть мемб-
раны говорят, что она определяет скорость массопереноса через
всю мембрану.
9.4. Диффузия через поры и мембраны
В последующих главах мы рассмотрим диффузионный перенос
молекул через стенки кровеносных сосудов. Это очень сложное
явление, и здесь дано только элементарное введение в диффузию
и фильтрацию через поры и мембраны. Обсудим сначала некото-
рые наиболее простые аспекты этого процесса, а затем ограничен'
ную, или несвободную, диффузии?.
Гл. 9. Введение в теорию массопереноса 167
Пусть имеется сосуд, содержащий раствор некоего вещества
концентрации который отделен пористым фильтром площадью
Лм от второго сосуда с аналогичным раствором концентрацией сг.
Градиент концентрации, обусловленный разностью концентраций
на двух поверхностях фильтра, обеспечивает диффузионный поток
растворенного вещества из одного сосуда (с более высокой кон-
центрацией) в другой. Одновременно происходит встречная диф-
фузия растворителя. Если диаметр пор фильтра гораздо больше
размера молекул растворенного вещества, легко рассчитать поток,
воспользовавшись уравнением (9.1); эффективная площадь диф-
фузионной поверхности равна в этом случае суммарной площади
пор фильтра. Пористость фильтра А„ равна площади пор на еди-
ничной поверхности фильтра или мембраны, и уравнение (9.1)
может быть записано в виде
Дс,
/==-Л"0-дГ’ <9-8)
где J, как и раньше, — поток через единичную поверхность фильт-
ра, а Аса — разность концентраций с двух сторон фильтра толщи-
ной Ах.
Если данный фильтр последовательно заменять на другие с
той же пористостью, но все меньшим диаметром пор, то поток
растворенного вещества сначала меняться не будет, однако ниже
определенного критического размера поры станут слишком малы,
чтобы пропускать молекулы растворенного вещества, и поток по-
следних уменьшится до нуля. Более мелкие молекулы по-преж-
нему смогут проходить через поры беспрепятственно; при этом го-
ворят, что они фильтруются.
Именно этот механизм лежит в основе диализа, при котором
раствор, содержащий крупные и мелкие молекулы, отделяют мемб-
раной от чистого растворителя. Крупные молекулы через мемб-
рану не проходят, более мелкие же могут проходить через нее
практически свободно, в результате чего исходный раствор обо-
гащается крупными молекулами. Адсорбция молекул растворен-
ного вещества на поверхности биологических мембран, их после-
дующий трансмембранный транспорт и высвобождение по другую
сторону зависят от ряда сложных изменений в ориентации вхо-
дящих в состав мембраны молекул. Для описания различных
свойств мембранного транспорта построен целый ряд молекуляр-
ных «моделей» биологических мембран; поскольку ни одну из них
нельзя считать полностью удовлетворительной, мы не будем ка-
саться этой темы в дальнейшем.
Ограниченная диффузия. Все сказанное выше применимо к
диффузии через пористые фильтры и мембраны, в которых диа-
метр пор во много раз превосходит диаметр молекул растворен-
ного вещества или растворителя. Было бы неверно считать, что
молекулы проходят через поры свободно, если только их диаметр
168 Часть I. Основы механики
меньше диаметра пор. Когда размер пор становится сравним с
хаотическими тепловыми смещениями молекул, диффузия стано-
вится ограниченной — как из-за столкновений молекул с краями
пор, так и из-за наличия близкодействующих электрических сил
взаимодействия между молекулами и стенкой. Вследствие этого
эффективная площадь пор Аэфф, через которую может идти пере-
нос, по мере увеличения размера молекул уменьшается до нуля.
Многие молекулы, транспортируемые через биологические
мембраны, сравнимы по размеру с заполненными водой порами,
через которые они проходят. Так, поры радиусом 4 нм ограничи-
вают диффузию глюкозы (радиус молекулы 0,37 нм) по сравне-
нию с ее свободной диффузией примерно на 35%, они ограничи-
вают даже диффузионный перенос воды (радиус 0,15 нм) при-
мерно на 15%. Чтобы количественно описать данный процесс, пре-
образуем сначала уравнение (9.8) к виду
An J
t\x D Ьск •
(9.9)
Левая часть этого уравнения — площадь пор, приходящаяся на
единичную длину диффузионного пути, — задается через легко из-
меряемые величины только при условии, что поры имеют гораздо
больший диаметр, чем диффундирующие молекулы. Когда же раз-
мер пор уменьшается, проникновение в них молекул становится
стерически затруднено и, кроме того, увеличивается сопротивле-
ние диффузии в порах. Поэтому коэффициент диффузии должен
быть заменен на £>огр— коэффициент ограниченной диффузии, а
истинная площадь пор АП — на эффективную площадь ЛЭфф. Тогда
уравнение (9.9) будет иметь вид
^эфф __ /________
“ ДогрДсА '
(9.10)
Из этого уравнения можно получить выражение для £)ОгР через
эффективную площадь пор, а также рассчитать поток молекул че-
рез мембрану с маленькими порами, когда известны Аэфф и Ап-
Подробно этот вопрос рассмотрен в гл. 13.
Многие биологические и искусственные мембраны обладают
Свойством полупроницаемости. Это означает, что они избирательно
пропускают одни молекулы и ионы и задерживают другие.
Первоначально явление полупроницаемости объясняли тем, что
мембрана действует просто как сито или пористый фильтр, задер-
живая крупные молекулы и пропуская более мелкие. Хотя этот
механизм в самом деле лежит в основе разделения молекул при
фильтрации и пассивной диффузии через поры мембран, в настоя-
щее время установлено, что он не является единственной причи-
ной полупроницаемости. Известны, например, случаи, когда моле-
кулы растворителя проходят через мембрану, а молекулы раство-
Гл. 9. Введение в теорию массопереноса 169
ренного вещества сделать этого не могут, хотя диаметр их иногда
меньше, чем у молекул растворителя.
Эта очевидная аномалия обусловлена рядом причин. Напри-
мер, эффективный размер молекул растворенного вещества мо-
жет существенно увеличиться вследствие гидратации (т. е. связы-
вания молекул растворителя), молекулы в растворе могут агре-
гировать. Прохождению молекул через поры иногда препятствует
электростатическое взаимодействие со стенками пор.
Внутриклеточная
среда
Внеклеточная
жидкость
Мембрана
рация вещества А
=> А
Высокая концентрация
вещества А
Рис. 9.4. Схема, иллюстрирующая механизм активного транспорта молекул веще-
ства А из среды внутрь клетки. Молекулы переносчика В все время находятся
внутри мембраны.
Идеальные полупроницаемые мембраны совершенно не про-
пускают молекулы растворенного вещества. Многие мембраны не
являются идеальными, растворенные молекулы в какой-то степени
проходят через них — наблюдается так называемая «утечка». По-
добная утечка объясняется либо внутренними свойствами мемб-
раны, либо ее неспособностью противостоять нагрузкам — она раз-
рывается или частично разрушается под действием большого пе-
репада давления, возникающего вследствие осмоса (разд. 9.7).
Активный транспорт. Многие биологические мембраны пере-
носят вещества быстрее, чем можно было бы ожидать, или же
переносят их против градиента концентрации в ходе процессов,
для осуществления которых необходима метаболическая энергия.
Такое явление называется активным транспортом и играет важ-
ную роль в поддержании определенного состава внутриклеточной
среды. Многие детали этого явления еще недостаточно выяснены,
но его механизм в своей основе, по-видимому, одинаков для всех
транспортируемых веществ и определяется наличием внутри мемб-
раны так называемых переносчиков (рис. 9.4). Молекулы веще-
ства А из внеклеточной среды адсорбируются на мембране и здесь
связываются с переносчиком В, образуя комплекс АВ. Последний
траспортируется через мембрану и на внутренней ее поверхности
диссоциирует, высвобождая вещество А во внутриклеточное про-
странство. После этого переносчик диффундирует обратно к на-
ружной поверхности мембраны, с тем чтобы еще раз присоеди-
нить молекулу вещества А. Метаболическая энергия необходима
для образования и разрыва химических связей — процессов, лок4’
лизованных в мембране.
170 Часть I. Основы механики
9.5. Проницаемость
Ввиду сложности структуры и свойств биологических мембран
для описания транспорта молекул через них широко используют
понятие мембранной проницаемости. Так, если поток вещества А
через мембрану при разности концентраций этого вещества Дс ра-
вен J, то
/ = РА-ДсА> (9.11)
где Рд — проницаемость мембраны для растворенного вещества А.
Ра в этом уравнении есть коэффициент массопередачи, согласно
определению, данному в уравнении (9.7). Проницаемость мемб-
раны для произвольного вещества зависит от многих факторов,
в частности от его концентрации и от температуры, и значения
этих величин необходимо указывать всякий раз, когда приводят-
ся значения проницаемости. Если концентрация не указана, го
предполагается, что раствор разбавленный (разд 9.1). Темпера-
тура существенно влияет на проницаемость, поскольку она опре-
деляет количество энергии, необходимой для осуществления пере-
носа через мембрану. Это связано с затратами энергии на адсорб-
цию растворенного вещества поверхностью мембраны и конфор-
мационные изменения в молекулярной структуре растворенного
вещества или самой мембраны.
9.6. Фильтрация через мембраны
До сих пор мы рассматривали только диффузионный перенос
веществ через мембраны. Однако если на мембране существует
перепад давления, то перенос может происходить также вслед-
ствие фильтрации через поры, и на самом деле этот процесс со-
провождает любой диффузионный перенос. Характерной его чер-
той является способность подвижных молекул в том случае, когда
градиент давления достаточно большой, перемещаться незави-
симо от существующих градиентов концентраций. Если поры мемб-
раны достаточно малы, чтобы не пропускать некоторые компо-
ненты жидкости, то сам процесс фильтрации может привести к об-
разованию перепада концентраций на мембране. Значение этого
эффекта в физиологии станет ясно в гл. 13, где будет рассмотрен
транскапиллярный обмен (разд. 13.8). Если через поры не про-
никают макромолекулы (скажем, белки), то процесс называют
ультрафильтрацией, если же не проникают мелкие молекулы (та-
кие, как неорганические соли), то он называется обратным осмо-
сом, однако механизм обоих процессов одинаков, различие лишь
в масштабе.
Если перепад давления на мембране есть Др, а обусловленный
им поток — J, то для простых мембран
l = (9.1?)
Гл 9 Введение в теорию массопереноса 171
где константа F называется коэффициентом фильтрации по ана-
логии с коэффициентом массопередачи в уравнении (9 7). Коэф-
фициент фильтрации зависит от таких факторов, как число пор,
приходящихся на единицу площади, их размер, форма, длина и
извилистость При данной геометрии F уменьшается с ростом вяз-
кости среды. Уравнение (9.12) может быть получено теоретически,
если предположить, что поток внутри пор подчиняется закону
Пуазейля, однако численное значение F при этом не может быть
предсказано достаточно точно.
9.7. Осмос
Осмос (от греч. «проталкивать») играет чрезвычайно важную
роль во всех процессах массопереноса, протекающих в биологи-
ческих системах.
Представим себе две камеры, разделенные жесткой полупро-
ницаемой мембраной так, как показано на рис. 9 5. Если камеру В
заполнить водным раствором белка, а камеру А — водой, то на пе-
регородке создастся градиент концентрации белка Если при этом
Рис 95 Схематическое изображение двух
камер А и В, разделенных жесткой полу-
проницаемой мембраной В камере В со-
держится водный раствор белка, в камере
А белка нет
размер пор фильтра достаточно мал, то молекулы белка не смогут
диффундировать через перегородку в камеру. А. Вода же из ка-
меры А будет диффундировать в камеру В, уменьшая разность
концентраций. В результате объем жидкости в камере В будет
увеличиваться.
Если обе камеры имеют вертикальные трубки, как показано
иа рис. 9 5, то при диффузии воды уровень жидкости в трубке ре-
зервуара В будет расти, а в трубке А — падать. Через некоторое
время уровни перестанут меняться, указывая на то, что суммар-
ный поток воды равен нулю. В этот момент достигается равно-
весие между «силой», обеспечивающей диффузию, и «силой»,
обусловленной превышением давления в отсеке В. Это превыше-
ние давления, устанавливающееся в состоянии равновесия, назы-
вают осмотическим давлением П раствора белка при данной рав-
новесной концентрации. Если мембрана пропускает мелкие рас-
творенные молекулы, но задерживает крупные коллоидные моле-
кулы, то устанавливающееся осмотическое давление называют
коллоидным осмотическим, или онкотическим.
172 Часть I. Основы механики
Итак, осмос наблюдается всегда, когда два раствора разной
концентрации разделены перегородкой, которая пропускает моле-
кулы растворителя, но не растворенного вещества; он может
быть результатом разделения веществ.
Можно показать, что осмотическое давление разбавленного
раствора, суммарная молярная концентрация которого (т. е. сум-
ма молярных концентраций всех растворенных веществ) равна с,
при данной абсолютной температуре Т подчиняется закону Вант-
Гоффа:
П = сЯГ, (9.13)
где R = 8,31 Н • м • К-1 • моль-1 — универсальная газовая постоян-
ная. Таким образом, разность осмотических давлений ДП с двух
сторон мембраны, где концентрация растворенного вещества равна
соответственно ct и сг, задается выражением
ДП^ТОй-Сг). (9.14)
Если бы разность концентраций составляла 1 моль-см-3, а темпе-
ратура раствора была 25°С (298 К), то перепад осмотического дав-
ления на мембране был бы равен примерно 1360 атмосферам! Из
этого следует, что в биологических системах даже при довольно
низких концентрациях растворенного вещества осмотическое дав-
ление может быть достаточно велико — для плазмы, например,
оно составляет примерно 3,3-103 Н-м~2 (25 мм рт. ст.).
Рассмотрим еще раз систему из двух камер, изображенную на
рис. 9.5. Предположим теперь, что в трубку камеры В введен
поршень,’ приложив к которому силу, можно увеличить гидроста-
тическое давление в этой камере. Пусть в какой-то определенный
момент времени концентрация белка в камере В равна сй тогда
осмотическое давление раствора П] будет задано уравнением
(9.13). Если с помощью поршня создать в камере В дополнитель-
ное давление pi, равное по величине Пь и воспрепятствовать тем
самым дальнейшему проникновению воды в камеру, то концент-
рация белка в ней перестанет уменьшаться. Осмотический пере-
нос воды остановится. Это не означает, что молекулы воды больше
не пересекают мембрану — они продолжают диффундировать че-
рез нее, но в обоих направлениях, так что суммарный поток ра-
вен нулю.
В действительности закон Вант-Гоффа применим только к иде-
альной полупроницаемой мембране, разделяющей два раствора;
если по какой-либо причине у мембраны есть утечка, то разность
осмотических давлений оказывается меньше, чем предсказывает
уравнение (9.13). Например, если два заполненные водой резер-
вуара разделены мембраной с порами диаметром 6 нм, то добав-
ление в один из них идеально растворимого вещества с диамет-
ром молекул, равным диаметру пор, вызовет в соответствии с за-
Гл. 9. Введение в теорию массопереноса 173
коном Вант-Гоффа осмотический поток воды. Если, однако, такая
же молярная концентрация будет создана не крупными, а мел-
кими молекулами (скажем, мочевиной, диаметр молекул которой
равен 0,5 нм), то вызванный осмотический поток составит менее
5% от ожидаемой величины, потому что большая часть мелких
молекул в этом случае может диффундировать через мембрану.
Это обстоятельство приводит к эмпирическому преобразованию
закона к следующему виду:
11= а с RT, (9.15)
где <т—осмотический коэффициент отражения. Его величина ме-
няется от единицы (для идеальной полупроницаемой мембраны)
до отрицательного значения (когда подвижность молекул раство-
ренного вещества выше, чем молекул растворителя). Сопряжение
между осмотическим и гидростатическим давлениями раствора иг-
рает важную роль в обменных процессах, протекающих в биологи-
ческих системах, так как перенос молекул почти всегда сопровож-
дается изменениями обоих видов давления. Действительно, изу-
чение как капиллярной фильтрации, так и транскапиллярного об-
мена (гл. 13) основано на рассмотрении взаимодействия между
осмосом и фильтрацией.
9.8. Простая модель массопереноса
В системе кровообращения непрерывно происходит перенос
массы между циркулирующей кровью и тканями. Отчасти массо-
перенос осуществляется благодаря фильтрации через стенки со-
суда, но в основном он обусловлен диффузией, идущей как в об-
ход клеток стенки сосуда, так и непосредственно через них.
Прежде чем рассматривать далее явление переноса массы в си-
стеме кровообращения, обсудим теории и модели, созданные для
описания некоторых простых динамических процессов массопере-
носа.
Рассмотрим более подробно, что происходит, когда кристаллы
сульфата меди (вещество А, рис. 9.6) помещают в воду. Вначале
в водной фазе совсем нет молекул А, но уже в следующий момент
времени в результате растворения и диффузии какая-то часть
молекул переходит в воду, прилегающую к поверхности кри-
сталла. В этом тонком слое воды раствор быстро становится
насыщенным в том смысле, что при данной температуре он не
может содержать больше молекул, чем в нем растворено. Кон-
центрация здесь называется равновесной поверхностной концент-
рацией сд и все время остается постоянной. Несколько дальше
от поверхности кристалла концентрация сд ниже сд, но она рас-
тет со временем. Растворение и перенос вещества путем диффу-
зии будут продолжаться до тех пор, пока всюду в водной фазе
концентрация не достигнет значения, равного равновесной поверх-
174 Часть I. Основы механики
Рис. 9.6. Диффузия молекул растворимого вещества А в неподвижной жидкости
(например, сульфата меди в воде). Показано последовательное изменение во вре-
мени профиля концентрации в сосуде.
ностной концентрации сд. До этого момента, как показано на
рис. 9.6, в воде происходит непрерывное изменение градиента кон-
центрации, а следовательно, и диффузионного потока. Концентра-
ция уменьшается с расстоянием от поверхности кристалла при-
мерно по экспоненте, и с помощью уравнения (9.3) можно опре-
делить, как быстро меняется профиль концентрации во времени.
Например, расстояние Ах от поверхности, на котором концентра-
ция составляет заданную величину (например, 0,5с1), прямо про-
порционально 'yjDt. Таким образом, вещество движется тем бы-
стрее, чем больше D, а пройденное расстояние растет как корень
квадратный из времени движения.
9.9. Взаимодействие объемного потока и диффузии
Нередко массоперенос протекает в условиях, когда одновре-
менно есть и объемный поток вещества, и тогда скорость пере-
носа зависит от характеристик потока. Такая ситуация, например,
наблюдается в кровеносных сосудах, где, в частности, представ-
ляет интерес перенос веществ к стенкам и от них при течении
крови. Следует иметь в виду, что сопротивление переносу может
определяться в основном стенкой, а не кровью, и, таким образом,
сопротивление последней слабо сказывается на общей скорости
переноса. Чтобы понять природу совместного действия потока и
диффузионного обмена, рассмотрим чрезвычайно упрощенную мо-
дель. Представим себе длинную прямую трубку (рис. 9.7); поток
на входе в нее имеет плоский профиль скорости, который посте-
пенно развивается при продвижении вдоль трубки так, как это
Гл. 9. Введение в теорию массопереноса 17В
описано в разд. 5.2. Стенка трубки содержит вещество А (или
сделана из него), очень медленно растворяющееся в данной жид-
кости, а жидкость, втекающая в трубку, не содержит этого веще-
ства совсем. По мере прохождения потока над растворимой частью
трубки стенки последней постепенно растворяются. Справа от на-
чала растворимой части стенки (от точки х — 0) концентрация
вещества А в жидкости у стенки равна Сд (равновесной поверх-
ностной концентрации), но в ядре потока вещество А сначала от-
сутствует. По мере продвижения вниз по течению происходит диф-
фузия вещества А из пристеночной области, и оно все больше
Стенка из растворимого вещества А
Рис. 9.7. Длинная трубка с
медленно растворяющейся стен-
кой, по которой течет стацио-
нарный ламинарный поток.
Изображено увеличение толщи-
ны диффузионного погранич-
ного слоя с расстоянием от
входа.
проникает в ядро потока. Однако, так как жидкость в ядре дви-
жется вдоль трубки, диффундирующие молекулы также перено-
сятся в этом направлении. Если для последовательно располо-
женных по длине трубки точек мы отложим радиальное расстоя-
ние, на которое диффундирует А (т. е. расстояние от стенки до то-
чек, где концентрация сА становится практически нулевой, как
показано на рис. 9.7), то полученная кривая (в трехмерном слу-
чае— поверхность) укаже’г нам границу той области, которая на-
зывается диффузионным пограничным слоем (ср. с вязким погра-
ничным слоем, описанным в гл. 5) 1).
При стационарных условиях мы можем рассчитать примерную
скорость роста толщины диффузионного пограничного слоя с рас-
стоянием таким же образом, как в гл. 5 мы рассчитывали уве-
личение толщины вязкого пограничного слоя. Рассмотрим неболь-
шой кубический элемент жидкости внутри пограничного слоя
(рис. 9.8) и примем, что каждая грань кубика имеет единичную
площадь. Тогда диффузионный поток в кубик снизу, через грань 1,
будет определяться уравнением (9.1), в которое надо подставить
значение градиента концентрации Ас^/Ау на этой грани. Диффу-
зионный поток из кубика через верхнюю грань 2 также опреде-
ляется уравнением (9.1), но уже с градиентом концентрации на
верхней грани. Таким образом, суммарный диффузионный приток
) На практике локальная толщина бд диффузионного пограничного слоя
определяется как расстояние от стенки, на котором местная концентрация рас-
сматриваемого вещества отличается от концентрации в ядре потока жидкости не
более чем иа 1% от общей разности концентраций.
176 Часть I. Основы механики
внутрь элемента имеет вид:
Суммарный поток = Коэффициент диффузии X
X Скорость изменения градиента концентрации с ростом у
или
J=D
d
Ay \ Ay )
(9.16)
Это соотношение полностью аналогично полученному в гл. 5 выра-
жению для суммарной действующей на элемент силы, обусловлен-
ной вязкими напряжениями.
Рис. 9.8. Кубический элемент жидкости внутри диффузионного пограничного слоя.
Такой диффузионный поток внутрь элемента уравновешивается
конвективным переносом вещества вниз по течению. Конвектив-
ный поток внутрь элемента через грань 3 равен произведению
концентрации на этой поверхности на скорость втекающей в эле-
мент жидкости. Таким же образом можно определить конвек-
тивный поток наружу через грань 4.
Следовательно, суммарный конвективный поток из элемента
задается выражением
Поток = Скорость изменения величины (скорость X
X концентрация) с ростом х,
или
_ d (исА)
йх
Если скорость массопереноса мала, а диффузия молекул внутрь
элемента не приводит к большим изменениям его объема, ско-
рость и не будет зависеть от х и
, йсА
J==U -j--
» Ах
(9,17)
Гл. 9 Введение в теорию массопереноса 177
Так как система стационарна, потоки, определяемые уравнениями
(9.16) и (9.17), должны быть равны. Таким образом,
dx dy \ dy )
ИЛИ
= (9-18)
dx dy2 ' '
Как и при расчете скорости, с которой увеличивается толщина
вязкого пограничного слоя, скорость роста диффузионного погра-
ничного слоя проще всего оценить, приписав каждому параметру
в этом уравнении разумные масштабы (порядки величины). Пусть
толщина диффузионного пограничного слоя на расстоянии х от
начала растворимой части трубки равна бд. Перпендикулярно
этому пограничному слою концентрация вещества А в жидкости
падает от сд (у стенки) до сА = 0 (на расстоянии бд от стенки),
и, следовательно, мы можем принять за масштаб dcA величину
Аса, равную (сА — 0). Порядок величины скорости внутри диффу-
зионного пограничного слоя зависит от того, какова толщина вяз-
кого пограничного слоя б по сравнению с 6Д. Если вязкий по-
граничный слой гораздо тоньше, чем диффузионный, то достаточно
хорошим масштабом скорости может служить величина Uo, т. е.
скорость в ядре потока жидкости. Если же диффузионный погра-
ничный слой гораздо тоньше вязкого, то можно считать градиент
скорости в диффузионном слое постоянным, и тогда разумным
масштабом скорости будет величина П0(бд/б).
Подставив полученные масштабы величин для случая, когда
вязкий пограничный слой гораздо тоньше диффузионного, в урав-
нение (9.18), мы придем к соотношению
r, kt „ Дс
^0—
д
откуда ___
(9Л9>
Здесь k — константа, значение которой необходимо рассчитывать
отдельно для каждого случая. Таким образом, толщина слоя рас-
тет с расстоянием от входа в трубку как х1/2. Сравнение (9.19)
с (5.3) показывает, что они весьма сходны. В частности, можно
видеть, что
(6/6J2 = v/D. (9.20)
Проделав то же самое для случая, когда толщина вязкого по-
граничного слоя больше диффузионного, приходим к соотношению
I] &л „ Г)
д
178 Часть I. Основы механики
Отсюда
Около входа в трубку, где величина 6 задается уравнением (5.3),
(6/6J3 = v/D. (9.21)
С другой стороны, для части трубки, достаточно удаленной от на-
чала, где профиль скорости развитый и 6 постоянно,
6дсхх,/3.
Толщина диффузионного пограничного слоя в этом случае растет
с расстоянием вдоль растворимой части трубки как х1/3.
Число Шмидта. Отношение v/D, как и число Рейнольдса, яв-
ляется безразмерной величиной; оно называется числом Шмидта
(Sc). Для газов числа Шмидта близки к единице, но в жидко-
стях гораздо больше. Так, при комнатной температуре для кисло-
рода в воде число Шмидта равно примерно 600, для мочевины
в воде — 950, а для лактозы в воде — 2400. В биологических си-
стемах для мелких молекул и ионов (например, для ацетат-ионов)
число Шмидта составляет 103, при увеличении размеров молекул
оно также растет и для крупных белковых молекул в воде или
плазме достигает 10®.
Столь огромные значения чисел Шмидта оказываются весьма
существенными при рассмотрении процессов массопереноса в био-
логических системах. Если мы рассчитаем ожидаемую толщину
диффузионного пограничного слоя для крупных белковых моле-
кул в небольшом сосуде диаметром, скажем, 100 мкм (толщина
вязкого пограничного слоя в нем не может быть больше радиуса
сосуда, т. е. 50 мкм), то увидим из уравнения (9.21), что она со-
ставляет примерно 5-10-2 мкм, или 50 нм. Это означает, что по
толщине подобного слоя укладываются всего две молекулы! Ясно,
что в таких ситуациях среду нельзя считать сплошной, но приве-
денный расчет не претендует на точность и был сделан только для
того, чтобы показать, насколько тонким может быть диффузион-
ный пограничный слой в некоторых биологических системах.
Часть II
МЕХАНИКА КРОВООБРАЩЕНИЯ
Глава 10
КРОВЬ
В этой главе рассмотрены механические свойства крови и ее
компонентов. Анализом движения крови в кровеносных сосудах,
а также вопросами, связанными с взаимодействием крови и стенки
сосудов, мы займемся в следующих главах—12 и 13. Основой
для изложения всех этих сведений послужат обсуждавшиеся в
гл. 4 и 5 элементы гидромеханики.
Кровь представляет собой суспензию форменных элементов
(различных клеток крови) и особых жидких частиц (хиломикро-
нов) в плазме. Сама плазма — это водный раствор, содержащий
в низких концентрациях множество низкомолекулярных органиче-
ских и неорганических веществ и примерно 7% (по массе) белков
Таблица 10.1
Примерные данные о составе плазмы
Вещество Концентра ЦИЯ, г на 100 мл Молекуляр- ный вес‘IO-3 Размер молекул, им
Вода Белки 90—92 15X4
Сывороточный альбумин 3,3—4,0 69
а,-Глобулины (включая липо- протеиды) 0,31—0,32 44—200
аа-Глобулииы (включая глико протеиды) - 0,48—0,52 150—300 20-50
P-Глобулины (включая липопро- теиды) 0,78—0,81 90—1300
у-Глобулииы 0,66—0,74 160-320 23X4
Фибриноген Неорганические компоненты Катионы Натрий Калий Кальций Магний Анионы Хлорид Бикарбонат Фосфат 0,34—0,43 0,31-0,34 0,016—0,021 0,009—0,011 0,002—0,003 0,36—0,39 0,20—0,24 0,003—0,004 400 (50-60) X (3-8)
180 Часть II. Механика кровообращения
(табл. 10.1). Очень важным механическим свойством крови яв-
ляется ее вязкость. Чтобы понять, чем определяется вязкость цель-
ной крови, мы должны сначала рассмотреть вопрос о том, от чего
зависит вязкость простых жидкостей и суспензий, а также озна-
комиться с механическими свойствами плазмы и взвешенных в
ней частиц и, наконец, с механическими свойствами цельной крови.
10.1. Вязкость жидкостей и суспензий
Как указывалось в гл. 1, физические свойства жидкостей, га-
зов и твердых тел прямо связаны с их молекулярной структу-
рой, и как собственно жидкости, так и газы относятся к классу
жидких сред, поскольку под действием сил, приводящих к воз-
никновению в них напряжения сдвига, они текут. Свойство веще-
ства, заключающееся в наличии связи между скоростью сдвига и
напряжением сдвига, называют вязкостью (разд. 4.3). Рассмот-
рим факторы, от которых зависит вязкость жидких сред.
Понять физику жидкостей помогает сопоставление свойств
жидкостей и газов. Газы имеют значительно меньшую плотность,
чем жидкости, и их молекулы находятся на большем расстоянии
друг от друга, чем молекулы жидкости. В связи с этим при хаоти-
ческом движении молекулы газа имеют большую длину свобод-
ного пробега, поскольку они реже сталкиваются друг с другом.
Этим же обусловлен и тот факт, что характерные коэффициенты
диффузии в газах примерно в 10 000 раз больше, чем в жидкостях.
Ввиду такого большого различия подвижностей молекул в га-
зах и в жидкостях механизмы возникновения вязкости этих ве-
ществ также различаются. Представим себе два параллельных
потока газа, движущихся с разными средними скоростями. На-
правления движения отдельных молекул в каждом потоке слу-
чайны, а диапазон их скоростей велик. Некоторые молекулы неиз-
бежно будут пересекать плоскость, разделяющую оба потока. Мо-
лекулы, переходящие из более медленного потока в более быст-
рый, сталкиваясь с молекулами последнего, будут ускоряться, и
их скорость будет приближаться к средней скорости движения
молекул более быстрого потока. Молекулы же, переходящие из бо-
лее быстрого потока в более медленный, будут замедляться. В ре-
зультате разность скоростей потоков уменьшается. Вязкость газа
определяется характером движения молекул. Например, при повы-
шении температуры газа тепловое движение составляющих его мо-
лекул становится более интенсивным и вязкость газа возрастает.
Молекулярную структуру жидкостей можно представить себе
как нечто среднее между структурой кристаллических твердых тел,
характеризующихся упорядоченным расположением молекул, и
структурой газов, молекулы которых расположены хаотически.
Когда твердое вещество плавится и переходит в жидкое состоя-
ние, его решетка разрушается, но небольшие скопления (класте-
Гл. 10. Кровь 181
ры) молекул, имеющих некую упорядоченность, еще сохраняются.
По мере повышения температуры жидкости энергия колебатель-
ного движения молекул в кластерах увеличивается, что вызывает
постепенное их разрушение на более мелкие, пока в точке кипе-
ния жидкости кластеры фактически не исчезнут.
Вязкость жидкостей во много раз превышает вязкость газов
как из-за более тесной упаковки молекул, так и из-за наличия
сравнительно больших упорядоченных групп молекул, которые
увеличивают сопротивление жидкости деформациям. В отличие от
вязкости газов вязкость жидкостей с повышением температуры
уменьшается. Полагают, что это происходит несмотря на увеличе-
ние подвижности отдельных молекул жидкости и обусловлено по-
степенным разрушением упорядоченных групп.
Когда суспензия с хаотически распределенными в ней части-
цами (будь то абсолютно твердые, деформируемые или жидкие
частицы) течет в трубке, размеры которой велики по сравнению с
размерами частиц и с расстоянием между ними, суспензию, как
было показано в гл. I, можно считать однородной жидкостью.
Изучив механические свойства такой суспензии, мы сможем по-
нять, от чего зависит ее вязкость и ведет ли себя жидкость как
ньютоновская (определение ньютоновской жидкости дано в
разд. 4.3). Кроме того, мы узнаем, какие факторы определяют
свойства крови как вязкой жидкости.
Сферические частицы. Пусть взвешенные в жидкости частицы
сферические и не оседают, т. е. имеют ту же плотность, что и
жидкость, которая сама по себе является ньютоновской. В этом
случае напряжение сдвига при любом движении будет пропор-
ционально скорости деформации (скорости сдвига), и суспензия
будет вести себя как ньютоновская жидкость, если только кон-
центрация сферических частиц не слишком высока. Это положе-
ние подтверждается, как описано в разд. 13.6, опытами, проводи-
мыми в условиях стационарного течения с суспензиями твердых
сферических частиц. Эффективная вязкость такой суспензии
т. е. вязкость, измеренная данным вискозиметром в данных усло-
виях (см. также разд. 10.7), при объемной концентрации частиц с
вплоть до 30% не зависит от скорости сдвига. Однако при кон-
центрациях, больших, чем примерно 10%, эффективная вязкость
зависит от способа измерения, т. е. от характера движения жид-
кости в конкретном вискозиметре *)• Следовательно, когда гово-
рят, что суспензия твердых сферических частиц является ньюто-
новской жидкостью, то обычно имеют в виду, что ее вязкость не
зависит от скорости сдвига при условии измерения некоторым
’) Эффективная, нлн кажущаяся, вязкость не является, таким образом, ха-
рактеристикой только собственных свойств суспензии и не совпадает, вообще
говоря, с истинным коэффициентом вязкости р, входящим в формулы типа
(4.4) — Прим. ред.
182 Часть И. Механика кровообращения
конкретным способом. Если взвешенные частицы не являются сфе-
рическими, если они деформируемы или притягиваются друг к
другу и образуют агрегаты, то при концентрациях, сравнимых с
30%, напряжение сдвига становится непропорциональным скоро-
сти сдвига. Так, например, вязкость суспензии эритроцитов в
плазме начинает зависеть от скорости сдвига уже при концентра-
циях, лишь немного превышающих 12% (рис. 10.16). Эффектив-
ная вязкость суспензии всегда выше эффективной вязкости ее
жидкой фазы. Чтобы понять, почему это так, рассмотрим ньюто-
новскую жидкость, движение которой вызвано перемещением с
постоянной скоростью ограничивающих ее поверхностей (рис. 10.1).
Жидкость между движущимися поверхностями сдвигается, в ре-
зультате чего в ней происходит диссипация энергии, причем тем
интенсивнее, чем больше вязкость жидкости.
Предположим теперь, что в жидкость введены твердые сфе-
рические частицы (рис. 10.2). Они могут вращаться, но в отличие
от той жидкости, место которой они заняли, не могут деформиро-
ваться. Следовательно, при таком же, как и ранее, перемещении
ограничивающих поверхностей жидкость будет деформирована в
большей степени и средняя скорость сдвига увеличится. Кроме
того, поскольку жидкость не может проскальзывать по поверхно-
сти частиц (разд. 4.3), в той ее части, которая прилегает к части-
цам, возникает дополнительный сдвиг. Оба этих эффекта приво-
дят к увеличению диссипации энергии в жидкости, и, таким обра-
зом, эффективная вязкость ее возрастает. При увеличении отно-
сительного объема взвешенных частиц должно происходить даль-
нейшее повышение вязкости, что и подтверждается эксперимен-
тально. Но если концентрация частиц не слишком высока, соот-
ношение между скоростью сдвига и напряжением сдвига при лю-
бой заданной концентрации постоянно, т. е. суспензия ведет себя
как ньютоновская жидкость.
Вязкость суспензии капель или деформируемых частиц также
растет с увеличением их относительного объема, но в меньшей
степени, чем при таком же увеличении концентрации твердых час-
тиц. Это обусловлено тем, что капли все же могут деформировать-
ся, и при движении суспензии добавочная скорость деформации
дисперсионной среды оказывается меньше. Однако при увеличе-
нии скорости сдвига в таких суспензиях капли не только деформи-
руются, но и постепенно ориентируются в направлении течения
(рис. 10.3). Это означает, что напряжение сдвига растет с увеличе-
нием скорости сдвига уже нелинейно. В результате вязкость ока-
зывается зависящей от скорости сдвига, а суспензия — соответ-
ственно неньютоновской жидкостью. Подробнее этим вопросом мы
займемся чуть дальше, когда будем рассматривать поведение
(тоже неньютоновское) суспензии твердых несферических частиц.
Поведение суспензий твердых и деформируемых частиц может
усложняться и становиться неньютоновским и в результате взаи-
Гл. 10. Кровь 183
Рис. ЮЛ. Стационарный сдвиг ньютоновской жидкости, заключенной между дву-
мя параллельными пластинами. Проскальзывание жидкости на границах отсут-
ствует (разд. 4.3). Нижияя пластина относительно наблюдателя неподвижна; длина
стрелок соответствует скорости движения жидкости относительно этой пласти-
ны. Очевидно, что наблюдателю, движущемуся вместе с верхней пластиной, жид-
кость и нижняя пластина будут казаться передвигающимися в противоположном
направлении (штриховые линии). Скорость движения жидкости в каждой точке
линейно связана с расстоянием от этой точки до границы жидкости, поэтому
скорость сдвига постоянна (гл. 4).
Рис. 10.2. Деформация сдвига в суспензии твердых сферических частиц. Хотя
частицы вращаются, их присутствие в жидкости приводит при заданном дви-
жении ограничивающих поверхностей к увеличению скорости сдвига в жидкости
(рис. Л). Кроме того, видно (рис. Б), что, поскольку жидкость не может про-
скальзывать по поверхности частиц, в той ее части, которая прилегает к части-
цам, возникает дополнительный сдвиг.
модействия между частицами. Это взаимодействие обусловлено
силами притяжения и отталкивания, а также тем, что жидкость,
изменившая свое движение под действием одной частицы, изме-
няет движение других частиц. Единственная точная теория, кото-
рой мы располагаем, относится к разбавленным суспензиям не-
взаимодействующих частиц. Эффективную вязкость рс разбавлен-
ной суспензии твердых невзаимодействующих сферических частиц
одинакового размера, обладающих нейтральной плавучестью (т. е.
не оседающих и не всплывающих), в жидкости с вязкостью Ро
рпервые вычислил в 1906 г. Альберт Эйнштейн. Он предсказал.
184 Часть II. Механика кровообращения
что если объемная концентрация частиц с (в долях единицы!)
мала по сравнению с 1, то относительная вязкость суспензии цОТн
(равная отношению эффективной вязкости к вязкости жидкой
фазы суспензии) определяется соотношением
ЦотН=4г = 1+4С- (ЮЛ)
но z
Этот результат подтвержден экспериментально для значений с, не
превышающих примерно 0,1. Для больших значений с нужно учи-
тывать сложное взаимодействие частиц, а это связано с введе-
нием членов, пропорциональных с2. В 1932 г. Тэйлор обобщил вы-
вод Эйнштейна на суспензии капель, которые сохраняют сфериче-
скую форму, например, благодаря поверхностному натяжению. Со-
ответствующее соотношение имеет вид
/ < 5 - \
„ ( Но + o' Ц 1
Н07И=-^=1+Д—(Ю.2)
где ц— вязкость жидкости, образующей капли. Когда Д стано-
вится бесконечно большим, т. е. когда капли оказываются в сущ-
ности твердыми частицами, это соотношение сводится к (10.1).
Рис. 10.3. Деформация сдвига в суспензии жид-
ких или деформируемых частиц. Частицы вра-
щаются, как показано на рис. 10.2, но, кроме
того, деформируются и ориентируются в направ-
лении течения.
Асимметричные частицы. При сдвиговом течении суспензии
твердых сферических частиц наблюдается вращение последних с
постоянной угловой скоростью. Если частицы асимметричны, то
они также вращаются, но угловая скорость вращения оказывается
непостоянной во времени: она максимальна, когда продольная ось
частиц перпендикулярна направлению течения, и минимальна, ког-
да параллельна. Поэтому большую часть времени частицы нахо-
дятся в таком положении, при котором их продольная ось почти
параллельна плоскости действия напряжений сдвига. В результате
подобного «выстраивания» частиц, обусловленного действием на-
пряжения сдвига, вязкость суспензии уменьшается и поведение ее
оказывается неньютоновским.
При одинаковой объемной концентрации с относительная вяз-
кость суспензии случайно ориентированных асимметричных твер-
дых частиц превышает вязкость суспензии сферических твердых
частиц. Это связано с тем, что при «кувыркании» под действием
напряжения сдвига каждая частица приводит в движение Объем
Гл. 10. Кровь 1Й5
жидкости, больший объема самой частицы и являющийся проме-
жуточным между объемами сфер, диаметры которых равны наи-
большему и наименьшему размерам частиц (рис. 10.4). Относи-
тельная вязкость суспензии твердых асимметричных частиц воз-
растает при увеличении как концентрации частиц, так и степени
их асимметричности. Эта зависимость определяется соотношением
Нотн=-^=1+Кг, (10.3)
но
где К (геометрический фактор) больше 5/2 [см. уравнение (10.1)].
С увеличением асимметричности частиц величина этого коэффи-
циента возрастает.
Рис. 10.4. «Кувыркание» асимметричных частиц
в суспензии при сдвиге. Для каждой частицы схе-
матически показан объем жидкости, приводимой
ею во вращательное движение.
Если асимметричные частицы гибкие, то ситуация еще больше
усложняется. По причинам, касаться которых мы здесь не будем,
во время части цикла своего вращения в потоке со сдвигом такие
частицы стремятся изогнуться (рис. 13.24). В результате этого
объем жидкости, который они приводят в движение, по мере воз-
растания скорости сдвига уменьшается, а следовательно, умень-
шается и эффективная вязкость суспензии.
При соответствующих условиях — т. е. при определенных раз-
мерах частиц, температуре и вязкости жидкой фазы — броунов-
ское движение (разд. 9.1) приводит к тому, что ориентация асим-
метричных частиц становится хаотической. Как мы видели, уве-
личение скорости сдвига в таких суспензиях уменьшает их эф-
фективную вязкость, если скорость сдвига достаточно высока,
чтобы преодолеть разориентирующее действие броуновского дви-
жения. Подобное свойство суспензий представляет интерес по-
тому, что в плазме крови содержится значительное количество та-
ких белков, как фибриноген и глобулины, молекулы которых су-
щественно асимметричны. Эти молекулы настолько велики по срав-
нению с молекулами воды, что их вполне можно считать взвешен-
ными частицами, способными придавать плазме неньютоновские
свойства. В действительности, как мы сейчас узнаем, этого не
происходит.
186 Часть II. Механика кровообращения
Рис. 10.5. Сопоставление температурных зависимостей вязкости для плазмы (Д)
и воды (□). [Cokelet (1972). The rheology of human blood. In: Biomechanics,
its foundations and objectives (eds Fung, Perrone, Anliker), Chapter 4, Prentice-
Hall, New Jersey.]
Вязкость плазмы. Плазма представляет собой бледно-желтую
прозрачную жидкость, которую можно получить, удалив из цель-
ной крови форменные элементы; предварительно предотвращают
свертывание добавлением особых веществ — антикоагулянтов. Ти-
пичные значения концентраций и размеров некоторых компонен-
тов плазмы здорового человека приведены в табл. 10.1; в нее
включены данные о веществах, которые предположительно ока-
зывают влияние на механику плазмы или цельной крови. Сыво-
ротка по внешнему виду сходна с плазмой, но для ее получения
позволяют крови свернуться, после чего удаляют сгусток, поэтому
в сыворотке отсутствуют те компоненты плазмы, которые участ-
вуют в процессе свертывания крови, в частности фибриноген.
Если производить вискозиметрию таким образом, чтобы плаз-
ма не соприкасалась с воздухом (это позволяет предотвратить де-
натурацию белков и устраняет механические эффекты, обусловлен-
ные наличием границы плазма — воздух), то при скоростях сдвига,
сравнимых с существующими in vivo (разд. 12.9), плазма здоро-
вого человека будет вести себя как ньютоновская жидкость. Это
подтверждает предсказание теории, согласно которому в физиоло-
гическом диапазоне значений скорости сдвига влияние последней
на вязкость плазмы должно быть неизмеримо мало. По всей ви-
димости, при таких скоростях сдвига броуновское движение под-
держивает хаотическую ориентацию асимметричных молекул. Ти-
Гл. 10. Кровь 187
пичные значения вязкости плазмы и сыворотки здорового чело-
века при 37°С составляют 1,2 и 1,1 мН-с-м-2 соответственно. При
некоторых заболеваниях, например при множественной миеломе
(злокачественном перерождении клеток костного мозга, синтези-
рующих глобулины), концентрация глобулинов в плазме может
повышаться в несколько раз и тогда вязкость плазмы увеличи-
вается. Ведет ли себя такая плазма как ньютоновская жидкость,
в настоящее время неясно *).
При изменении температуры вязкость плазмы здорового чело-
века меняется точно так же, как вязкость ее растворителя — воды:
повышение температуры (в пределах нормальных физиологиче-
ских значений) на 5°С уменьшает вязкость плазмы примерно на
10% (рис. 10.5).
10.2. Осмотическое давление плазмы
В гл. 9 мы установили, в чем заключается физическая сущ-
ность осмотического давления, и теперь представим некоторые ко-
личественные данные об осмотических свойствах плазмы. Кроме
того, мы приступим к рассмотрению влияния осмотического дав-
ления плазмы на механику кровообращения.
Осмотическое давление раствора определяется количеством
присутствующих в нем частиц и температурой. Например, одно-
молярный раствор вещества (1 моль, растворенный в 1000 г воды)
содержит 6,02-1023 молекул, и при 0°С его осмотическое давление
равно 2,27 МН-м~2 (22,4 атм.). Однако в жидкостях тела содер-
жатся как электролиты, так и неэлектролиты, а электролиты в
воде диссоциируют на ионы. Поэтому в одномолярном водном рас-
творе хлорида натрия при полной диссоциации содержится 12,04 X
ХЮ23 ионов, и при 0°С его осмотическое давление составляет
4,54 МН-м~2 (44,8 атм.). Литр одномолярного раствора неэлектро-
лита содержит один осмоль, а литр такого же раствора электро-
лита (например, хлорида натрия) при полной диссоциации — два
осмоля.
Существует несколько механизмов влияния осмотического дав-
ления плазмы на механику течения крови. Изменение перепада
осмотического давления на мембране эритроцита приводит к из-
менению содержания в нем воды и к изменению его объема. На-
пример, снижение концентрации хлорида натрия снаружи эритро-
цита ведет к его набуханию. При особо сильном набухании об-
разуются сферические эритроциты; поскольку такие эритроциты не
могут деформироваться без растяжения их мембраны (стр. 193),
жесткость их значительно превышает жесткость обычных эрит-
*) Добавим, что большинство реологических опытов с плазмой поставлено
в условиях стационарного течения, когда некоторые типы отклонений от ньюто-
новского поведения могут просто не проявляться.— Прим. ред.
188 Часть П. Механика кровообращения
роцитов, имеющих форму дисков. Как мы увидим ниже (разд. 10.7),
изменение как формы эритроцитов, так и их гибкости влияет на
механические свойства цельной крови. Кроме того, изменение ос-
мотического давления плазмы влияет на гематокрит (объемную
концентрацию эритроцитов в цельной крови) вследствие перерас-
пределения воды между внутрисосудистым и внесосудистым про-
странством. Это в свою очередь сказывается на движении цель-
ной крови.
Растворы, имеющие такое же осмотическое давление, как и
плазма здорового организма (примерно 0,7 МН-м-2 при 0°С), при-
нято называть изотоническими, а растворы с более высоким или
более низким осмотическим давлением — соответственно гипер- и
гипотоническими. Нередко используется и еще один термин — кол-
лоидное осмотическое давление; он характеризует осмотическое
давление, обусловленное присутствием в растворе макромолекул,
в отличие от осмотического давления, определяемого мелкими мо-
лекулами или ионами. Коллоидное осмотическое давление жид-
кости, находящейся в сосудах, и жидкости, заполняющей межкле-
точное пространство (т. е. жидкостей, находящихся по разные
стороны стенок капилляров), несколько различается. Это разли-
чие обусловлено меньшим содержанием белков в межклеточной
жидкости. Именно транскапиллярная разность коллоидного осмо-
тического давления вместе с транскапиллярной разностью гидро-
статического давления в значительной степени определяют рав-
новесное распределение жидкости между внутрисосудистым и вне-
сосудистым пространством (стр. 408). Коллоидное осмотическое
давление плазмы определяется главным образом содержанием
в ней альбумина. Вклад фибриногена в эту величину очень мал,
поскольку его молярная концентрация, а следовательно, и осмоти-
ческое давление составляют лишь около */50 молярной концент-
рации и осмотического давления альбумина.
10.3. Взвешенные элементы
В плазме взвешены клетки крови и капельки жидкости, назы-
ваемые хиломикронами. Присутствие в крови этих капелек диа-
метром 0,2—0,5 мкм связано с механизмом переноса жира. Од-
нако концентрация хиломикронов в крови настолько мала, что
они не оказывают сколько-нибудь заметного влияния на вязкость
или осмотическое давление плазмы. Поэтому мы не будем далее
рассматривать указанные элементы и лишь отметим, что они по-
являются в крови главным образом после приема жирной пищи,
попадают в кровь из лимфатических сосудов кишечника и уда-
ляются посредством ряда механизмов, включая перенос жира на
растворимых в воде молекулах-переносчиках — липопротеидах
(табл. 10.1).
Гл. 10. Кровь 189
10.4. Клетки крови
В циркулирующей крови присутствуют клетки нескольких ти-
пов, но мы остановимся в основном на эритроцитах, поскольку
только они оказывают на механические свойства крови существен-
ное влияние. Это обусловлено тем, что эритроциты содержатся в
крови в очень высокой концентрации (примерно 5-Ю6 мм-3), со-
ставляя около 45% ее объема (табл. 10.2).
Таблица 10.2
Клетки крови
Клетки
Число клеток
в 1 мм®
Форма и размеры* мкм
Объемная
концент-
рация
в крови,
%
Эритроциты
Лейкоциты
Общее содержание
Гранулоциты
Нейтрофилы
Эозинофилы
Базофилы
Лимфоциты
Моноциты
Тромбоциты
(4-6) X 106
(4-11) ХЮ3
(1,5—7,5) X 103 ]
(0-4) ХЮ2
(0—2) ХЮ2 I
(1—4,5) ХЮ3 j
(0-8) ХЮ2)
(250-500) X Ю3
Двояковогнутый диск,
8-Х (1-3)
Приблизительно
сферические, 7—22
Сфероидальные, 2—4
Для сравнения укажем, что в нормальных условиях на
1000 эритроцитов приходится только 1—2 лейкоцита. Тромбоциты
(кровяные пластинки) более многочисленны, чем лейкоциты (50—
100 на 1000 эритроцитов), но, поскольку они очень малы, их влия-
ние на механические свойства крови по сравнению с влиянием
эритроцитов ничтожно. Как видно из табл. 10.2, объем тромбоцита
составляет лишь около ’/ю объема эритроцита.
Эритроциты. Прежде чем приступить к анализу механических
свойств эритроцитов, следует упомянуть о некоторых физиологи-
ческих свойствах этих клеток. Внутри эритроцитов находится
почти весь гемоглобин крови (около 15 г на 100 мл крови). Имен-
но гемоглобин (мол. вес 67 000), состоящий из белка глобина, свя-
занного с гемом — органической группой, содержащей атом же-
леза, придает эритроцитам их красный цвет, а способность гемо-
глобина обратимо соединяться с кислородом обеспечивает боль-
шую кислородную емкость крови. На уровне моря кислородная
емкость 100 мл цельной крови составляет околб 20 мл, тогда как
кислородная емкость 100 мл плазмы — всего лишь около 0,3 мл.
Эритроциты образуются в костном мозгу, и их незрелые формы
имеют ядра. Однако у большинства млекопитающих при созре-
190 Часть П. Механика кровообращения
вании эритроцитов ядра исчезают, и в нормальных условиях эрит-
роциты с ядрами в циркулирующую кровь не поступают. Несмотря
на отсутствие ядра и митохондрий1), эритроцитам присуща мета-
болическая активность. Исследования с применением меченных изо-
топами эритроцитов показывают, что средняя продолжительность
их жизни составляет примерно 120 сут. Следовательно, для под-
держания в крови постоянной концентрации этих клеток необхо-
димо, чтобы каждые сутки разрушалось и образовывалось вновь
примерно 0,8% всего их количества. По мере старения эритроциты
становятся более хрупкими. In vivo они, вероятно, разрушаются
механически и фрагменты их поглощаются лейкоцитами и дру-
гими клетками — «чистильщиками»; этот процесс называется фа-
гоцитозом (см. ниже). Полагают, что механическое разрушение
Рис. 10.6. Схематическое изображение недеформироваиного эритроцита.
клеток происходит в основном в микрососудах: именно здесь
кровь испытывает наибольшее сопряжение сдвига (табл. 12.3).
Поскольку эритроциты оказывают существенное влияние на ме-
ханические свойства цельной крови, мы уделим им особое внима-
ние. Изучение механических свойств эритроцитов оказывается, од-
нако, сложной задачей, и по некоторым вопросам здесь еще не
выработана единая точка зрения.
Когда эритроциты находятся во взвешенном и недеформирован-
ном состоянии в плазме или в другой изотонической жидкости,
то они представляют собой очень гибкие двояковогнутые диски
(рис. 10.6). Их плотность равна примерно 1,08-103 кг-м-3. Пола-
гают, что они состоят из очень тонкой мембраны и жидкого со-
держимого, которым является почти насыщенный раствор гемо-
глобина (концентрация его равна примерно 32% по массе). В том,
что это действительно так, мы убедимся ниже. Некоторые веще-
ства, входящие в состав эритроцитов, перечислены в табл. 10.3.
Вязкость находящейся внутри эритроцитов жидкости считается
равной примерно 6 мН-с-м-2 — это приблизительно в 5 раз выше
вязкости плазмы крови. Предполагают, что при некоторых забо-
') Это органеллы клетки, ответственные за метаболические процессы, в част-
ности связанные с высвобождением энергии в результате окисления,
Гл. 10. Кровь 191
Вещество
Состав эритроцитов
Таблица 10.3
Содержание
Вода
Компоненты мембраны
(белок, фосфолипиды, холестерин)
Гемоглобин
Неорганические элементы
Калий
Натрий
Магний
Кальций
65% (по массе)
3% (по массе)
'<32% (по массе)
0,420 г на 100 мл
0,025 г на 100 мл
0,006 г на 100 мл
Небольшое количество
леваниях, сопровождающихся кристаллизацией гемоглобина (на-
пример, при серповидноклеточной анемии), вязкость этой жидко-
сти изменяется.
Изучение мембраны эритроцитов, на долю которой приходится
не более 3% массы клетки, показывает, что она содержит двой-
ной слой фосфолипидов толщиной около 7,5 нм. Проницаемость
мембраны для анионов (хлорида и бикарбоната) и для воды весь-
ма высока, тогда как для катионов (калия и натрия), а также
для макромолекул, включая белки плазмы, крайне низка. Обмен-
ный Na+—К+-насос клетки перекачивает Na+ и К+ против гра-
диента их электрохимических потенциалов, Na+ — наружу, К+—
внутрь клетки (табл. 10.1 и 10.3). Энергию для этого насоса обес-
печивает расщепление глюкозы, а поскольку митохондрии в эрит-
роцитах отсутствуют, процесс этот протекает главным образом
анаэробно.
Представление о том, что содержимое эритроцитов является
жидкостью, подтверждается результатами ряда наблюдений. На-
пример, проходя по капилляру, диаметр которого меньше их соб-
ственного диаметра, эритроциты очень сильно изменяют свою
форму. Форма, которую они в этом случае принимают, — притуп-
ленная спереди и заостренная сзади (рис. 13.33,Л), — указывает,
что их содержимое перетекает в переднюю часть клетки. К та-
кому же выводу приводит изучение поведения эритроцитов, при-
липших к предметному стеклу, покрытому слоем сывороточного
альбумина. Под действием сил, вызывающих в эритроците на-
пряжение сдвига, он деформируется (рис. 10.7). При увеличении
напряжения сдвига клетка удлиняется и выглядит так, будто ее
содержимое выдавлено из растянутого «хвоста» в тело клетки.
Эти и другие наблюдения (включая электронно-микроскопи-
ческие) показывают, что неповрежденные эритроциты не имеют
определенной внутренней структуры, т. е. их содержимое является
жидким. Кроме того, эти наблюдения наводят на мысль, что фор-
192 Часть II. Механика кровообращения
ма эритроцитов определяется свойствами их мембраны. Такая
точка зрения подтверждается исследованиями теней эритроцитов.
Если постепенно снижать осмотическое давление жидкости, в ко-
торой взвешены эритроциты, то они не только становятся сфери-
ческими, но и теряют свое содержимое — сначала низкомолеку-
лярные вещества, а затем, после разрыва мембраны, и гемогло-
бин. Этот процесс называется гемолизом. Если поврежденные та-
ким способом клетки поместить в подходящий изотонический рас-
твор, то мембрана их смыкается, они поглощают раствор и вновь
Рис. 10.7. Действие напряжения сдвига на эритроцит, удерживаемый на предмет-
ном стекле. Поток направлен справа налево. Крайнее слева изображение полу-
чено при наибольшей величине напряжения сдвига, крайнее справа — iwn наи-
меньшей. [Hochmuth, Mohandas, Blackshear (1973). Biophys. J., 13, 754.]
напоминают по форме и размерам нормальные эритроциты. «Те-
нями» такие эритроциты названы потому, что они лишены гемо-
глобина.
Есть, однако, один факт, указывающий на то, что эритроциты
имеют какую-то внутреннюю структуру: их можно разделить на
небольшие части без потери содержимого. Для объяснения этого
явления предполагают, что внутри эритроцитов имеется сеть нз
гибкого материала, но ни электронная микроскопия, ни изучение
теней эритроцитов не дают каких-либо подтверждений этому. Воз-
можно, мембрана эритроцитов способна смыкаться вокруг их
фрагментов уже в момент образования последних.
Поскольку толщина мембраны эритроцитов составляет лишь
около 7,5 нм, а наименьший радиус кривизны эритроцита — около
1 мкм (рис. 10.6), для анализа его свойств можно использовать
теорию тонких оболочек. Согласно этой теории, сопротивление
мембраны изгибу должно быть пренебрежимо мало по сравне-
нию с ее сопротивлением растяжению (см. также разд. 13.7). По-
этому если содержимое эритроцитов является жидким и, следо-
вательно, способно оказывать деформации вязкое, но не упругое
сопротивление, то можно ожидать, что эритроциты должны быть
очень гибкими.
Гл. 10. Кровь 193
Способность эритроцитов сильно деформироваться (они могут,
например, проходить не разрушаясь через трубочку диаметром
3 мкм и длиной 12 мкм) обусловлена, однако, не только свой-
ствами их мембраны и тем, что внутри этих клеток находится
жидкость, но и их формой. Подобно другим несферическим телам,
двояковогнутые эритроциты могут деформируясь принимать самую
разную форму без изменения объема или площади поверхности,
т. е. без растяжения мембраны. Значение этого факта легче всего
оценить, сравнив механические свойства обычных и сферических
эритроцитов. Можно показать, что тело сферической формы имеет
при заданной площади поверхности максимальный объем. Значит,
площадь мембраны сферического эритроцита должна при его де-
формации увеличиваться, так что гибкость клетки будет опреде-
ляться жесткостью мембраны в тангенциальном (т. е. вдоль по-
верхности) направлении. И действительно, экспериментально об-
наружено, что сферические эритроциты не могут деформироваться
столь же сильно, как обычные, — например, они не проходят без
разрушения через узкие каналы. Однако при очень сильной де-
формации (возникающей, например при движении в потоке с высо-
ким напряжением сдвира) мембрана натягивается, стремясь огра-
ничить деформацию эритроцита, и может, как показано ниже, ра-
зорваться.
Модуль Юнга для мембраны эритроцитов, определенный экс-
периментально, имеет порядок 106 Н-м~2. Эти измерения произ-
ведены на мембране, растянутой почти до разрыва, а для боль-
шинства биологических тканей характерно увеличение модуля
Юнга при увеличении их растяжения (гл. 7). Исследования, в ко-
торых сравнивали движение эритроцитов при возвращении их
после деформации к ненапряженному состоянию с движением мо-
делей клеток с известными свойствами, при пересчете дали для
модуля Юнга мембраны величину порядка 10~5 Н-м-2, т. е.
в 10 раз меньшую.
Мембрана эритроцитов обладает свойством сохранять постоян-
ными площадь поверхности и толщину при любой деформации.
Например, когда при изменении осмотического давления эритро-
циты становятся сферическими, никакого изменения площади их
поверхности не выявляется. С другой стороны, под влиянием боль-
ших напряжений сдвига — в опытах с прикреплением эритроци-
тов (рис. 10.7) или в вискозиметре (рис. 10.9) — эритроциты за-
метно удлиняются.
До сих пор мы не пытались объяснить, почему эритроциты,
содержимое которых является жидким, а форма, которую они
имеют, будучи взвешенными в ненапряженном состоянии в изо-
тоническом растворе, является дискообразной, могут разбухать и
становиться сферическими. Теоретически клетка, мембрана кото-
рой имеет однородную растяжимость, не может вести себя подоб-
ным образом. Как показано на рис. 10.8, переход от диска к сфере
194 Часть 11. Механика кровообращения
Рис. 10.8. Погруженная в воду тонкостенная резиновая модель эритроцита, за-
полненная водой. А — исходная форма. При увеличении трансмурального давле-
ния (Б) полюса выпячиваются, а вблизи экватора образуются прогибы, показан-
ные стрелками. При высоком внутреннем давлении (В) увеличившаяся в разме-
рах «клетка» опять гладкая и по форме близка к сфере. [Fung (1966). Theore-
tical considerations of the elasticity of red cells and small blood vessels, Fedn
Proc. Fedn Am. Socs exp. Biol., 25, 1761—72.]
требует уменьшения длины окружности клетки по экватору. Зна»
чит, при раздвижении полюсов эта область должна сморщиваться,
что и наблюдается на моделях эритроцитов, имеющих мембрану
с однородными свойствами, но не на реальных эритроцитах. По-
ведение эритроцитов при переходе их от дискообразной формы
к сферической можно объяснить, если предположить, что модуль
Юнга мембраны, или ее толщина, или и то и другое в области
Гл. 10. Кровь 195
экватора больше, чем у полюсов клетки. Однако никаких прямых
данных по этому вопросу пока нет.
Обратимся теперь к деформации эритроцитов, вызываемой на-
пряжением сдвига, и рассмотрим данные, полученные при иссле-
довании разбавленной суспензии эритроцитов в условиях сдвиго-
вого течения. Ясно, что эритроцит может войти в очень узкую
трубку и затем передвигаться по ней, лишь если он при этом де-
формируется. Кроме того, из уже проведенного в этой главе ана-
лиза поведения нежестких взвешенных частиц мы знаем, что в
потоке со сдвигом клетка крови тоже должна деформироваться,
и это подтверждают прямые наблюдения при помощи движуще-
гося, следящего за клеткой, микроскопа (гл. 13). Когда в суспен-
зии возникает напряжение сдвига, эритроциты действительно де-
формируются и вращаются, что обусловлено наличием в потоке
скорости сдвига, и при этом как скорость деформации, так и ско-
рость вращения зависят от скорости сдвига и от концентрации
клеток. При скорости сдвига около 1 с~‘ клетки, вращаясь, изги-
баются, а с увеличением скорости сдвига они перестают вращать-
ся как единое целое и движутся вдоль трубки так, что их про-
дольная ось оказывается направленной под некоторым углом к
потоку. Тщательные исследования показывают, что клетки при
этом непрерывно меняют очертания, что обусловлено вращением
мембраны вокруг клеточного содержимого и весьма похоже на
движение гусеницы танка. В этом случае поведение клеток в ка-
кой-то степени аналогично поведению капель жидкости (разд. 10.1).
Исследование суспензии эритроцитов в условиях ламинарного
течения показывает, что для их гемолиза необходимо напряже-
ние сдвига около 200 Н-м~2, но эта величина не является на-
дежно установленной, а некоторые исследователи утверждают,
что причина гемолиза — в первую очередь контакт эритроцитов
с чужеродной поверхностью, а не наличие достаточно большого
напряжения сдвига1)- На рис. 10.9,Л и Б представлены электрон-
ные микрофотографии сильно разбавленных (0,2%) суспензий
эритроцитов человека, фиксированных глутаральдегидом во время
воздействия на них напряжений сдвига разной величины в рота-
ционном вискозиметре с коаксиальными цилиндрами. При 10 Н-м-2
клетки деформируются довольно слабо, а при 300 Н-м-2 они до-
вольно сильно вытягиваются (до 20—30 мкм). Если клетки фик-
сировали после снятия напряжения сдвига, большинство их имело
обычный внешний вид, т. е. возникающие в них изменения были
в данном случае обратимы. Напряжение сдвига 200 Н-м-2, оче-
видно, слишком высокая для системы кровообращения величина:
*) Величина 200 Н-м-2 считается абсолютным порогом, ниже которого ие
происходит гемолиза в объеме (т. е. не на стенках прибора), выше этого порога
степень гемолиза зависит еще и от длительности действия напряжений. — Прим
ред.
196 Часть II. Механика кровообращения
Ри?. 10.9. Микрофотографии эритроцитов, фиксированных глутаральдегидом во
время воздействия на них В вискозиметре напряжения сдвига, равного 10 Н-м-2
(И) и 300 Н-м“2 (£). [Sutera, Mehrjardi, Mohandas (1974), личное сообщение.]
Рис. 10.10. Образование эритроцитами монетных столбиков. [Bessis (1973). Li-
ving blood cells and their ultrastructure (transl. Weed), p. 141, Springer Verlag,
Heidelberg.]
Гл. 10. Кровь 197
уже при 40 Н-м-2 происходит отрыв или разрушение эндоте-
лиальных клеток, выстилающих аорту. Полагают также, что ге-
молиз возникает в потоке с высокой турбулентностью (которая
может развиваться, например, за стенозированным аортальным
клапаном), с чем и связывают наблюдаемую иногда в этих усло-
виях анемию. Однако величина и длительность воздействия на-
пряжения сдвига на эритроциты в этом случае неизвестны.
Еще одним характерным свойством эритроцитов является тен-
денция к образованию агрегатов. Если нанести мазок крови на
предметное стекло микроскопа, то можно видеть, как эритроциты
«склеиваются» друг с другом, образуя скопления, которые полу-
чили название монетных столбиков (рис. 10.10). Этот процесс на-
чинается через 10—20 с, и увеличение длины столбиков продол-
жается в течение 1—2 мин. Механизм образования столбиков
до конца не выяснен. По-видимому, эритроциты притягиваются
друг к другу, и это притяжение определяется локализованными
на поверхности клеток заряженными группами1)- Процесс агре-
гации зависит также от наличия в окружающей эритроциты среде
некоторых крупных асимметричных макромолекул, в частности
фибриногена и глобулинов. В отсутствие таких молекул (напри-
мер, если эритроциты взвешены в солевом растворе или в рас-
творе сывороточного альбумина нормальной концентрации) про-
цесс не развивается. Более того, с повышением концентрации фиб-
риногена и глобулинов скорость его увеличивается. Поэтому в
стационарном потоке с ненулевой скоростью сдвига в любой мо-
мент времени относительное количество эритроцитов, объединен-
ных в монетные столбики, зависит от концентрации названных ве-
ществ.
Наибольший диаметр эритроцитов в составе монетных столби-
ков превышает диаметр недеформированных эритроцитов, а тол-
щина их соответственно меньше. Это согласуется с представле-
нием о взаимном притяжении клеток. Сферические эритроциты,
как и эритроциты, жесткость которых увеличена, например, глута-
ральдегидом, не образуют монетных столбиков, возможно, из-за
неспособности их соответствующим образом деформироваться.
Монетные столбики могут образовываться при низких скоро-
стях сдвига не только in vitro, но и in vivo, и в обоих случаях
зависимость их образования от концентрации фибриногена и гло-
булинов одинакова. Эти вещества важны с точки зрения меха-
ники, потому что они влияют на вязкость крови при низких ско-
ростях сдвига и на скорость оседания эритроцитов (разд. 10.7).
*) Если даже такое притяжение существует (а большинство исследователей,
наоборот, считают, что существует электростатическое отталкивание), оио, как
показывают оценки, может иметь значение только после сближения эритроцитов
на расстояние ~ 0,01 мкм. Само же сближение обычно обусловлено чисто меха-
ническими факторами — неодинаковостью скоростей движения эритроцитов, ув-
лекаемых потоком, оседанием и т. п. — Прим. ред.
198 Часть II. Механика кровообращения
Лейкоциты. Эти клетки крови мало влияют на ее механические
свойства, и потому мы расскажем о них очень кратко. Концент-
рация лейкоцитов в крови весьма низка: на их долю приходится
лишь около 1% ее объема. Лейкоциты играют решающую роль
в защите организма от инфекций — они уничтожают микроорга-
низмы и образуют антитела. Более подробно об этих клетках
можно прочитать в руководствах по физиологии и гематоло-
гии.
В циркулирующей крови имеются лейкоциты трех видов —
гранулоциты, лимфоциты и моноциты (табл. 10.2). Разновидно-
сти лейкоцитов можно отличить по внешнему виду под микроско-
пом и по их. сродству к различным органическим красителям.
В нормальных условиях лейкоциты имеют форму, близкую к
сферической; диаметр клеток разных видов составляет примерно
от 7 до 22 мкм (табл. 10.2). Электронно-микроскопическое иссле-
дование этих клеток обнаруживает развитую внутреннюю струк-
туру, включая присутствие митохондрий. Если принимать во вни-
мание объем эритроцитов и лейкоцитов, то последние биохимиче-
ски значительно более активны, причем в лейкоцитах протекают
как аэробные, так и анаэробные процессы. Лейкоциты способны
к активному амебоидному движению; они используют для этого
особые выросты — псевдоподии. Кроме того, они способны к фаго-
цитозу, т. е. к поглощению чужеродных веществ, в том числе бак-
терий, а также погибших или разрушенных клеток. Лейкоциты
могут также захватывать и выделять и более мелкие частицы, на-
чиная с макромолекул, но это обеспечивает другой механизм, на-
зываемый пиноцитозом (разд. 13.8).
О механических свойствах лейкоцитов известно немного. По-
скольку при столкновении в движущейся крови эритроцита с лей-
коцитом деформируется в основном эритроцит, полагают, что лей-
коциты более жесткие, чем эритроциты. Однако не нужно забы-
вать о вязкоупругих свойствах клеток. Обнаружено, например,
что если лейкоцит переходит из широкой трубки в капилляр, диа-
метр которого меньше диаметра недеформированного лейкоцита,
то для того, чтобы он настолько изменил свою форму, что смог бы
продвигаться по капилляру, требуется 10—20 с. Поместив лейко-
циты в соответствующий гипотонический раствор, можно заста-
вить их разбухать и разрываться.
Тромбоциты. Кровяные пластинки, или тромбоциты, образуют-
ся в костном мозгу из очень крупных клеток, называемых мега-
кариоцитами. При наблюдении in vivo, например в капилляре,
тромбоциты выглядят как маленькие сфероидальные тельца диа-
метром 2—4 мкм и объемом 5-^10 мкм3. Ядер в них нет, но при
электронно-микроскопических исследованиях выявляются мемб-
рана толщиной около 8-10"3 мкм и органеллы, типичные н для
других клеток, в том числе митохондрии, микрофибриллы и мик-
ротрубочки.
Гл, 10. Кровь 199
X
Эндогенный путь
IXa 1
+VIII-—[д
+Пл+Са.2^'
Экзогенный путь
Ткань
+VIl+Ca?+
Ха-«—
+V-e-
+Пл+Сар+
•X
II—-----▼---------
(протромбин)
1-----
(фибриноген)
эд II а (тромбин)
XIII-
—1-1-------1 а-f- фибринопепти дь)
[ (фибрин-мономер)
[ | (спонтанный процесс}
' Фибрин-попимер
। (растворимый)
-Д’ - >ХШа—(сшивание)
Сгусток фибрина
(нерастворимый)
Фагоцитоз —-----------Фибринолиз
Продукты расщепления
фибрина
Рис. 10.11. Поздние стадии процесса свертывания крови (см. текст). Римские
цифры соответствуют международной номенклатуре факторов свертывания. На-
звания факторов, указанных на рисунке только цифрами: V — проакцелерин;
VII — проконвертин, или акцелератор преобразования сывороточного протром-
бина; VIII—антигемофилический фактор; IX — фактор Кристмаса; X — фактор
Стюарта — Прауэра; XIII — фибринстабилизирующий фактор. Активированные
производные обозначены индексом а. Пл — плазма. [Hardisty, Weatherall (1974).
The haemostatic mechanism. In: Blood and its disorders, Blackwell Scientific Pub-
lications, Oxford.]
При соприкосновении с АДФ (аденозиндифосфат, органическая
молекула, играющая важную роль во многих механизмах высво-
бождения энергии у живых организмов) тромбоциты образуют
агрегаты. В настоящее время считают, что в нормальных усло-
виях АДФ способствует агрегации тромбоцитов in vivo. Он высво-
бождается из разрушенных эритроцитов, а также из соедини-
тельной ткани (ее коллагена) в местах повреждения. Но основ-
ным источником АДФ при агрегации тромбоцитов являются, по-
видимому, они сами: вполне возможно, что, прилипая к повреж-
денной ткани, тромбоциты выделяют АДФ, способствующий их
последующей агрегации.
В присутствии тромбина (см. следующий раздел, Свертыва-
ние крови, и рис. 10.11), коллагена, АДФ или адреналина тром-
боциты быстро теряют свою округлую форму, выпуская множе-
ство небольших отростков (псевдоподий). Такое обратимое изме-
нение формы предшествует агрегации. Когда происходит эгрета-
200 Часть II. Механика кровообращения
Рис. 10.12. Тромбоциты с псевдоподиями (микрофотографии получены при помо-
щи сканирующего электронного микроскопа). [Bessis (1973). Living blood cells
and their ultrastructure (transl. Weed), p. 381, Springer Verlag, Heidelberg.]
ция, клетки приходят в более тесный контакт и выпускают зна-
чительно более крупные псевдоподии (рис. 10.12). Эти изменения
необратимы.
Агрегация тромбоцитов — лишь один из процессов, служащих
для прекращения кровотечения из поврежденного сосуда; той же
цели служат образование сгустков крови и сужение сосудов. Эти
процессы входят в арсенал защитных реакций организма. Есть
основания считать, что при нормальном состоянии системы крово-
обращения процесс агрегации тромбоцитов также приводится в
действие для закупорки спонтанно возникающих небольших ка-
налов утечки крови.
10.5. Свертывание крови
Кровь свертывается, когда оказывается в необычных условиях,
например в поврежденном кровеносном сосуде, в ране или в стек-
лянной пробирке. Процесс свертывания крови чрезвычайно сло-
жен и пока еще выяснен не полностью. Поэтому мы изложим
здесь лишь основные его этапы.
Сгусток крови состоит из сети фибрилл — тонких волокон, об-
разованных белком фибрином, и из попавших в эту сеть клеток —
Гл. 10. Кровь 201
эритроцитов, лейкоцитов и тромбоцитов (относительные концент-
рации их в сгустке крови примерно такие же, как и в цельной
крови). Обычно в крови содержится очень мало фибрина; он об-
разуется в ней при свертывании. Основные стадии процесса об-
разования фибрина схематически представлены на рис. 10.11.
Схема составлена на основе одного из последних обзоров, к ко-
торому читатель может обратиться для получения более подроб-
ных сведений. Заметим, что предпоследняя важная стадия (пре-
вращение протромбина в тромбин) может быть активирована
двумя путями: один (экзогенный) связан с выделением факторов
свертывания из поврежденной ткани, а другой (эндогенный) за-
висит от соприкосновения крови с чужеродными поверхностями.
Препятствующее свертыванию крови вещество гепарин действует
на оба процесса н, кроме того, тормозит реакцию между тром-
бином и фибриногеном.
Фибрин, образующийся в кровеносных сосудах in vivo, может
быть удален из крови как путем фагоцитоза, так и путем разру-
шения, т. е. процесса, называемого фибринолизом. Фибринолиз,
как и образование фибрина, складывается из сложной последо-
вательности ферментативных реакций.
После образования сгусток крови сокращается (ретракция
сгустка), т. е. нити фибрина укорачиваются, сгусток становится
более плотным и из него выдавливается сыворотка. По-видимому,
в процессе ретракции принимают участие прикрепляющиеся к
фибриновым нитям тромбоциты. Показано, что последние содер-
жат сократительный белок, который сходен с актомиозином мышц.
10.6. Тромбоз
Тромбозом ’) называется прижизненное свертывание крови в
кровеносных сосудах. В различных областях сосудистого русла
этот процесс протекает по-разному — возможно, из-за существен-
ного влияния на него местных гидромеханических условий. Тром-
бы, как и сгустки крови, свернувшейся вне сосудов, состоят из
различных элементов крови, попавших в сеть фибрина. Механизм
образования фибрина при тромбозе, вероятно, тот же, что и при
формировании сгустков — та же цепь процессов, которая пока-
зана на рис. 10.11. Однако механизм, запускающий процесс тром-
боза, обычно несколько иной. По этой причине, а также из-за
различий гидромеханических влияний на процесс образования
тромбов они отличаются по структуре от сгустков и в разных час-
') Более точное определение тромбоза таково: это процесс прижизненного
образования в просвете сосудов или в полостях сердца тромбов — плотных масс,
формирующихся из составных частей крови и способных полностью или частично
перекрывать просвет сосуда. Далее авторы уточняют, что процесс образования
тромбов не тождествен прижизненному свертыванию крови в сосудах, поскольку
формирование тромба начинается с агрегации тромбоцитов. — Прим. ред.
202 Часть Л. Механика кровообращения
тях сосудистого русла бывают различными. Наиболее сходен с
процессом образования сгустков венозный тромбоз, особенно при
весьма медленном течении крови. Тромбоз такого типа особенно
часто встречается у больных преклонного возраста, перенесших
хирургическое вмешательство или находящихся в течение дли-
тельного времени на постельном режиме. Тромбы, образующиеся
в артериях, имеют другую структуру. Прежде всего в этом случае
формируется «головка» тромба, состоящая главным образом из
агрегатов тромбоцитов, на которых затем осаждаются слои фиб-
рина и клеток крови.
Причины тромбоза разнообразны; этот процесс может проис-
ходить в любой области сосудистого русла, но, по крайней мере
в артериях, решающим начальным событием является формиро-
вание тромбоцитарного агрегата. В нормальных условиях тром-
боциты не прилипают друг к другу или к стенкам сосудов; как
сказано выше, способность слипаться появляется у них в присут-
ствии АДФ. Кроме того, есть основания полагать, что агрегаты
тромбоцитов и поврежденная ткань способны выделять веще-
ство, которое запускает процесс свертывания крови; такой путь
является «экзогенным» (рис. 10.11). Местные гидромеханические
факторы могут влиять на этот процесс, воздействуя как на диф-
фузию содержащихся в крови веществ, которые ответственны за
агрегацию тромбоцитов и свертывание крови, так и на доставку
тромбоцитов и других клеток крови к месту образования тромба.
Свертывание крови в сосудистом русле можно вызвать экспери-
ментально, введя в кровь экстракт из ткани или тромбин. Воз-
можно поэтому, что изменение содержания в крови веществ, по-
добных АДФ, способно, вызывая агрегацию тромбоцитов, оказы-
вать влияние на вероятность образования тромбов in vivo.
К образованию артериальных тромбов наиболее часто пред-
располагают атероматозные бляшки (гл. 12). Однако необходи-
мым условием является, по-видимому, соприкосновение крови с
коллагеном или с другими веществами, находящимися глубоко
в сосудистой стенке, поскольку тромбы не возникают, если нет
обширного повреждения эндотелия, приводящего к обнажению
этих веществ.
Тромбоз возможен и на уровне микрососудов — при таких, на-
пример, заболеваниях, как полицитемия (чрезмерно высокое со-
держание в крови эритроцитов) и серповидноклеточная анемия.
Последняя представляет собой врожденное заболевание, при кото-
ром эритроциты содержат аномальный гемоглобин. Из-за этого
при снижении парциального давления кислорода в крови, что на-
блюдается, например, при подъеме на большие высоты, эритро-
циты становятся серповидными и менее гибкими.
Некоторые вены мышц голени имеют тенденцию с возрастом
расширяться, что сопровождается наклонностью их к тромбозу.
Это заболевание поражает преимущественно больных преклонного
Гл. 10. Кровь 203
возраста, которые, как отмечено выше, перенесли хирургическое
вмешательство или находились в течение длительного времени на
постельном режиме. Замедление течения крови в расширенных
венах считают фактором, предрасполагающим к тромбозу, осно-
вываясь, в частности, на том, что применение процедур, увеличи-
вающих местный кровоток, например сокращения мышц голеней
или ритмическое сжатие их (разд. 14.5), предохраняет от тром-
боза. Однако экспериментальные исследования показывают, что
повышение свертываемости крови также играет важную роль,
хотя оно и не является причиной преимущественного развития
тромбов в определенных венах. Тромбы могут возникать также
в левом предсердии, особенно когда оно расширено и имеет место
его фибрилляция (слабые неэффективные нерегулярные сокраще-
ния его мышцы). Полагают, что механизм тромбоза в этом случае
сходен с механизмом возникновения тромбов в венах в том отно-
шении, что движение крови в предсердии при этом также замед-
лено.
Тромбоз, по-видимому, редко возникает у больных, находив-
шихся в неподвижном состоянии менее двух-трех дней, но после
хирургического вмешательства тромбы иногда возникают уже че-
рез час. При экспериментальном повреждении стенки сосуда
агрегацию тромбоцитов в области повреждения можно наблю-
дать уже через секунды. Скорость роста агрегата в этом случае
пропорциональна местной скорости сдвига в потоке у стенки, что,
вероятно, обусловлено увеличением числа тромбоцитов, достав-
ляемых в область повреждения. В противоположность этому у
больных, у которых тромбы формируются в венах или в предсер-
дии, скорости сдвига вблизи стенки в области возникновения
тромба, очевидно, малы. Важно иметь в виду, что in vivo
существуют также механизмы удаления тромбов, так что воз-
можно, например, восстановление кровотока в закупоренном со-
суде.
В нормальных условиях свертывание крови не является един-
ственным процессом, участвующим в остановке кровотечения при
повреждении сосуда, и, пожалуй, не относится к наиболее важ-
ным из таких процессов. Действительно, люди, у которых врож-
денно отсутствует фибриноген (афибриногенемия), могут вести
почти нормальный образ жизни. Полагают, что основными факто-
рами, обеспечивающими безотлагательную остановку кровотече-
ния, являются на самом деле образование тромбоцитарного агре-
гата и сокращение разорванного сосуда. Затем на месте повреж-
дения образуется сгусток крови, который, приклеиваясь к тканям,
действует подобно пломбе. У больных с пониженным содержанием
тромбоцитов или с аномальными тромбоцитами возникают спон-
танные кровотечения из мелких сосудов и длительные кровотече-
ния из ран.
204 Часть II. Механика кровообращения
10.7. Механические свойства цельной крови
Итак, согласно сказанному выше, с точки зрения механики
цельную кровь можно рассматривать как ньютоновскую жидкость
(плазму), в которой взвешены гибкие эритроциты. Рассмотрим
теперь оседание эритроцитов в крови, принципы измерения вяз-
кости крови и ее вязкие свойства.
Будем считать, что пробы, на которых исследуются свойства
крови, достаточно велики, чтобы можно было йренебречь влия-
нием стенок ячейки прибора, в которую заключена проба. Такое
предположение упростит анализ. На практике, однако, при виско-
зиметрии крови используются и малые пробы, так что это пред-
положение не всегда соответствует действительности.
Оседание эритроцитов. Этот вопрос весьма интересен, посколь-
ку оседание эритроцитов может оказывать влияние на механику
течения крови и затруднять измерение вязкости крови. Кроме
того, определение скорости оседания эритроцитов в крови имеет
важное диагностическое значение.
В гл. 5 мы рассматривали обтекание вязкой жидкостью твер-
дого сферического тела в стационарных условиях в случае, когда
инерцией жидкости можно пренебречь. Такое течение имеет ме-
сто также, когда сферическая частица медленно опускается в вяз-
кой жидкости, а стенки сосуда, в который налита жидкость, нахо-
дятся на достаточно большом расстоянии от частицы, чтобы не
оказывать влияния на протекающие процессы. В этом случае на-
правленная вниз гравитационная сила, действующая на частицу,
уравновешена силой вязкого трения. Исходя из этого равновесия
сил, можно показать, что скорость оседания частицы Uo опреде-
ляется законом Стокса [уравнение (6.1)]:
iJ X ✓
где а — радиус частицы, рч и рж(рч>рж)— плотности частицы и
жидкости соответственно, ц — вязкость жидкости, g— ускорение
силы тяжести. Существенно, что скорость оседания частицы зави-
сит от квадрата ее радиуса.
Если частицу удерживать в жидкости в состоянии покоя и за-
тем освободить, то скорость ее оседания U будет увеличиваться
до тех пор, пока не достигнет указанной выше постоянной вели-
чины Uo, которую называют конечной скоростью. Скорость час-
тицы является функцией времени t и приближенно описывается
соотношением
t/ = t/0(l -е"ет)’
так что при больших значениях t(t^T) U становится практи-
чески равной Uo. Величина Т — это постоянная времени процесса,
определяемая соотношением
гос-^е-4.
Гл. 10. Кровь 205
Таким образом, она увеличивается с ростом размера частицы. За-
метим, что, когда t = ЗТ, U составляет приблизительно 95% Uo
(гл. 8).
Для твердой сферической частицы, близкой по размерам к
эритроциту (8 мкм в диаметре), Т равно примерно 5-10—5 с. Сле-
довательно, частица достигнет скорости, равной 95% Uo, чрезвы-
чайно быстро — через 10-4 с.
Эту теорию нельзя безоговорочно применять для описания
движения эритроцитов, поскольку они не являются сферическими
и легко деформируются. Тем не менее теория движения твердых
сфер дает некоторые указания относительно поведения одиноч-
ных оседающих эритроцитов. В настоящее время нельзя точно
описать оседание в суспензии множества эритроцитов, но теория
движения твердых сфер позволяет высказать некоторые сообра-
жения и по этому вопросу. Если цельную кровь, свертывание ко-
торой предотвращено, поместить в капиллярную трубочку, уста-
новленную вертикально, эритроциты будут объединяться в монет-
ные столбики и затем медленно оседать. В клинических лабора-
ториях измерение расстояния, на которое за первый час смещает-
ся верхняя граница области, занимаемой эритроцитами, исполь-
зуют в качестве диагностического теста. Результат такого измере-
ния называют скоростью оседания эритроцитов (СОЭ). В норме
СОЭ составляет примерно 3 мкм-с-1; условно она указывается
как 10 мм-ч-1. В действительности скорость оседания не является
постоянной, потому что сначала должны образоваться агрегаты,
а когда эритроциты начинают упаковываться в нижней части тру-
бочки, скорость оседания снижается.
При различных заболеваниях СОЭ может увеличиваться в не-
сколько раз (вплоть до 10); при этом одновременно повышается
концентрация в плазме фибриногена и/или глобулинов. Эти белки
способствуют образованию монетных столбиков; предполагают,
что именно быстрое оседание таких агрегатов является причиной
увеличения СОЭ. Возрастание СОЭ не обусловлено изменением
вязкости плазмы, которая в действительности увеличивается и,
следовательно, сама по себе должна уменьшать СОЭ.
Наблюдая при определенных условиях за течением крови по
горизонтальной трубке, можно видеть, что эритроциты отходят от
верхней стенки трубки и, оседая, упаковываются более плотно
вблизи ее нижней стенки, изменяя таким образом условия дви-
жения крови в этих областях. Например, в верхней части трубки
вязкость будет уменьшаться, в нижней — увеличиваться. Не ис-
ключено, что такие же явления происходят и in vivo — например,
если скорость течения крови становится очень низкой или если
белковый состав плазмы изменяется так, что образование монет-
ных столбиков усиливается.
Оседание эритроцитов может привести к ошибкам при изме-
рении вязкости крови, особенно если используется вискозиметр
206 Часть It. Механика кровообращения
типа конус — плоскость (см. ниже), который имеет фактически
горизонтальные рабочие поверхности. При очень малых скоростях
сдвига эритроциты будут образовывать агрегаты и перемещаться
от верхней поверхности к нижней. Детально этот вопрос, по-види-
мому, не изучен ’).
Принципы измерения вязкости крови. Прежде чем переходить
к анализу механики сдвигового течения цельуой крови, следует
рассмотреть вопросы, связанные с методами измерения вязкости
крови. Это необходимо сделать потому, что кровь является сус-
пензией частиц и при физиологически нормальных скоростях
сдвига может вести себя как неньютоновская жидкость.
Если есть основания полагать, что некая среда является не-
ньютоновской, то вязкость ее необходимо измерять при несколь-
ких скоростях сдвига. Отсюда следует, что нужно уметь либо оп-
ределять скорость сдвига, либо задавать ее. Но для неньютонов-
ских жидкостей решить эту задачу особенно сложно, поскольку
вискозиметр, который обеспечивает практически постоянную ско-
рость сдвига во всем объеме пробы ньютоновской жидкости, мо-
жет не сделать этого в неньютоновской жидкости. Как мы видели,
именно из-за присутствия эритроцитов кровь здорового орга-
низма является неньютоновской жидкостью. Возникающие из-за
этого технические сложности вискозиметрии связаны преимуще-
ственно с тем, что кровь не является однородной жидкостью. Так,
например, вязкость крови зависит от размеров пробы: особенно
большие трудности возникают, когда размеры пробы, исследуе-
мые в вискозиметре, сравнимы с размерами эритроцитов или —
если в пробе образуются монетные столбики — с размерами стол-
биков. Подробно этот вопрос рассматривается в гл. 13, посвящен-
ной движению крови в микрососудах. Ни один вискозиметр не
является идеальным, но в одних вискозиметрах эти затруднения
преодолеть легче, в других — труднее.
Обычно используются вискозиметры двух типов: капиллярные
и ротационные. В капиллярном приборе жидкость протекает по
трубке с точно известными размерами под действием заданной
разности давлений между концами трубки. Прибор сконструиро-
ван так, чтобы для представляющих интерес жидкостей течение
было ламинарным и полностью развитым (разд. 5.1). В этом
случае кажущуюся вязкость жидкости можно определить вполне
корректно исходя из формулы Пуазейля [формула (5.1)].
Капиллярный вискозиметр вполне подходит для исследования
ньютоновских жидкостей, каковой является, например, плазма,
но, по уже упомянутым причинам, для неньютоновских жидкостей,
в том числе и для крови, он малопригоден. В вискозиметре такого
') Оседание эритроцитов может вносить погрешности и в другие измерения —
например, расхода крови при помощи электромагнитного расходомера. — Прим,
ред.
Гл. 10. Кровь 207
типа жидкость подвергается действию напряжения сдвига, меняю-
щегося в широких пределах, причем это напряжение всегда мак-
симально вблизи стенки, а у осевой линии равно нулю (разд. 5.1).
Кроме того, поскольку для получения полностью развитого ла-
минарного течения при приемлемых значениях разности давлений
приходится применять трубки с малым внутренним диаметром
(например, 100 мкм), последний оказывается сравнимым с раз-
мером эритроцитов (или монетных столбиков)—он превышает его
лишь в несколько раз. В этих условиях кажущаяся вязкость
крови, как показано дальше (разд. 13.6), зависит от диаметра
трубки.
Широкое распространение получили ротационные вискози-
метры двух типов: с коаксиальными цилиндрами (рис. 10.13) и
типа конус — плоскость (рис. 10.14). Как те, так и другие состоят
из двух основных частей, отделенных друг от друга исследуемой
жидкостью. Одна из этих частей приводится во вращение отно-
сительно другой с постоянной скоростью, и вращающий момент
передается неподвижной части благодаря вязким свойствам жид-
кости. Этот момент можно измерить, определив угловое переме-
щение неподвижной части, если последняя подвешена на нити.
Действительно, в таком случае поворот и закручивание нити бу-
дут продолжаться до тех пор, пока возвращающие силы, связан-
ные с напряжениями в закрученной нити, не сравняются с переда-
ваемым неподвижной части вращающим усилием.
При исследовании свойств крови ротационные вискозиметры
предпочтительнее капиллярных; это связано главным образом с
тем, что при правильной конструкции таких приборов и доста-
точно высоком напряжении сдвига большая часть пробы подвер-
гается сдвигу, скорость которого близка к постоянной. Кроме
того, ротационные вискозиметры позволяют исследовать пробы в
течение длительного периода времени и изучать изменение их
свойств во времени.
Если в зазоре вискозиметра с коаксиальными цилиндрами уста-
новилось стационарное ламинарное течение ньютоновской жидко-
сти, то скорость сдвига в жидкости пропорциональна угловой ско-
рости вращения подвижного цилиндра и падает по ширине зазора
обратно пропорционально квадрату расстояния от оси цилиндров
(рис. 10.15). Следовательно, скорость сдвига можно считать по-
стоянной лишь в том случае, если это расстояние меняется мало,
т. е. ширина зазора много меньше радиусов цилиндров. Момент,
передаваемый наружному цилиндру при вращении внутреннего
(обычно вращается именно внутренний цилиндр), прямо пропор-
ционален напряжению сдвига, а значит, кажущейся вязкости
жидкости и угловой скорости вращения внутреннего цилиндра.
Если исследуемая жидкость неньютоновская, то ни напряжение
сдвига, ни измеряемый вращающий момент, действующий на на-
ружный цилиндр, не будут прямо пропорциональны угловой око-
208 Часть II. Механика кровообращения
Рис. 10.13. Схема вискозиметра с коаксиальными цилиндрами. [Van Waser, Lyons,
Kim, Colwell (1963). Viscosity and flow measurement, p. 50, Interscience Publi-
shers, New York.]
Исследуемая проба
Вращающийся конус
Неподвижная
плоскость
Рис. 10.14. Схема вискозиметра типа конус — плоскость, а —угол между конусом
и плоскостью. [Van Wazer, Lyons, Kim, Colwell (1963). Viscosity and flow measu-
rement, p. 52, Interscience Publishers, New York.]
роста вращения внутреннего цилиндра. Уравнения, описывающие
поведение в вискозиметре различных неньютоновских жидкостей,
основываются на сильно идеализированных моделях жидкостей,
поэтому целесообразнее определять кажущуюся вязкость реаль-
ных жидкостей путем калибровки вискозиметра по ньютоновской
жидкости с известной вязкостью. При этом, исследуя поведение
реальной жидкости при разных скоростях сдвига, обнаруживают,
что ее кажущаяся вязкость зависит от скорости сдвига. Еще
одна трудность возникает, если изучаемая жидкость обладает
предельным напряжением сдвига; в этом случае при достаточно
низких напряжениях сдвига определенные части жидкости сов-
сем не приходят в движение. В нормальных условиях кровь ве-
дет себя так лишь при исключительно низких напряжениях
сдвига.
В вискозиметре типа конус — плоскость (рис. 10.14) исследуе-
мая жидкость заключена между конусом с большим углом при
вершине и пластиной, плоская поверхность которой перпендику-
лярна оси конуса. Обычно конус приводят во вращение и изме-
Гл. 10. Кровь 209
ряют вращающее усилие, передаваемое исследуемой жидкостью
неподвижной пластине. Такое устройство имеет перед цилиндри-
ческой конструкцией некоторые преимущества. В частности,
можно использовать пробы меньшего объема, а при малых углах
между конусом и пластиной (а < 3°) скорость сдвига ньютонов-
ской жидкости можно считать одинаковой на любом расстоянии
от оси. В некотором приближении это справедливо даже для не-
ньютоновской жидкости, если ее вязкость зависит от скорости
сдвига не слишком сильно. Как и в цилиндрическом вискозиметре,
л б в г
Рис. 10.15. Распределение скорости движения жидкости по линиям тока в зазоре
вискозиметра с коаксиальными цилиндрами. Внутренний цилиндр вращается, на-
ружный неподвижен. А. Прибор не приведен в движение. Б. Сдвиг в ньютонов-
ской жидкости; обратите внимание, что скорость движения жидкости по ширине
зазора падает (см. текст). В. Если ширина зазора очень мала по сравнению с
радиусом цилиндра, то скорость сдвига почти постоянна, как между двумя па-
раллельными плоскостями (рис. 10.1). Г. Жидкость, обладающая предельным
напряжением сдвига. [Van Wazer, Lyons, Kim, Colwell (1963). Viscosity and
flow measurement, p. 75, Interscience Publishers, New York.]
неньютоновские жидкости исследуют в указанном приборе, пред-
варительно откалибровав его по ньютоновской жидкости с извест-
ной вязкостью. Однако, поскольку зазор, в котором происходит
сдвиговое течение, в вискозиметре типа конус — плоскость очень
узок, допущение о том, что жидкости, подобные крови, однородны,
может оказаться неправомочным. Вблизи вершины конуса вели-
чина зазора может быть сравнимой с размерами клеток крови или
их агрегатов.
Вязкие свойства крови. Приведем теперь результаты экспери-
ментального изучения вязкости крови и прежде всего обратимся
к зависимости вязкости крови от скорости сдвига. Данные, пред-
ставленные на рис. 10.16, основаны на нескольких исследованиях
крови человека; они показывают, что с изменением скорости
сдвига кажущаяся вязкость крови меняется. Когда скорость
сдвига близка к 1000 с-1, т. е. к величине, типичной для многих
кровеносных сосудов in vivo, отклонение поведения крови от
ньютоновского становится очень малым и кажущаяся вязкость
приближается к своему асимптотическому значению, лежащему
в пределах 3—4 мН-с-м~2. Однако при уменьшении скорости
сдвига кажущаяся вязкость постепенно возрастает, пока при ско-
210 Часть II. Механика кровообращения
Рис. 10.16. Зависимость кажущейся вязкости крови человека от скорости сдвига.-
[Whitmore (1968). Rheology of the circulation, p. 67, Pergamon Press, Oxford.]
ростях сдвига меньше 1 с-1 не начинает увеличиваться очень
резко.
Чтобы объяснить эти данные, рассмотрим сначала поведение
крови при очень малых скоростях сдвига. Известно, что при этом
в крови образуются монетные столбики, а когда сдвиговое течение
отсутствует, можно наблюдать неупорядоченную сеть агрегирован-
ных эритроцитов. Если такая кровь подвергается воздействию на-
пряжения сдвига, не превышающего некоторой критической вели-
чины, то структура, образованная агрегированными клетками, как
предполагают, деформируется, но течение крови не возникает, т. е.
такая кровь обладает предельным напряжением сдвига. В пользу
этого представления имеются некоторые экспериментальные дан-
ные; так, тщательные измерения показывают, что предельное на-
пряжение сдвига крови здорового человека составляет около 1,5—
5,0 мН-м"2. Как видно из рис. 10.17, оно снижается с уменьше-
нием гематокрита, а если гематокрит не превышает определенной
критической величины, то и вообще отсутствует. Возможно, в этом
случае единица объема крови содержит слишком мало эритроци-
тов, чтобы при агрегации они связали всю пробу в единую струк-
туру. С увеличением концентрации фибриногена или у-глобулина
в крови предельное напряжение сдвига увеличивается. Этого и
следовало ожидать, имея в виду, что такие асимметричные бел-
корые молекулы способствуют образованию монетные столбиков.
Рис. 10.17. Зависимость предельного напряжения сдвига (то)
кровн от гематокрита (для пяти проб крови). При гематокрите
ниже примерно 5% предельное напряжение сдвига не выяв-
ляется. Значения гематокрита сопоставлены со значением ку-
бического корня из предельного напряжения сдвига, поскольку
такое соотношение оказалось близким к линейному. [Merrill,
Gilliland, Cokelet. Shin, Britten, Wells (1963). Rheology of
human blood near and at zero flow, Biophys. J., 3, 210.]
Рис. 10.18. Зависимость вязкости суспензии эритроцитов чело-
века, находящихся в плазме той же крови, из которой их пер-
воначально удалили (к плазме добавлено противосвертываю-
щее вещество), от скорости сдвига при различных объемных
концентрациях эритроцитов (значения концентраций в % ука-
заны около кривых). Температура суспензии 25°C. [Brooks,
Goodwin, Seaman (1970). Interactions among erythrocytes un-
der shear, J. appl. Physiol., 28, 174.]
212 Часть II. Механика кровообращения
Рис. 10.19. Влияние гематокрита на
относительную вязкость суспензий
обычных эритроцитов и эритроцитов
с повышенной жесткостью в физиоло-
гическом растворе (скорость сдвига
230 с-1). За единицу принята вяз-
кость чистого раствора. [Whitmore
(1968). Rheology of the circulation,
p. 81, Pergamon Press, Oxford.]
Влияние гематокрита на механику течения крови изучено для
широкого диапазона скоростей сдвига; поведение вязкости крови
при скоростях сдвига вплоть до 700 с-1 иллюстрируется рис. 10.18.
При значениях гематокрита, достигающих примерно 12%, свойства
суспензий эритроцитов в плазме оказываются ньютоновскими при
всех скоростях сдвига, хотя вязкость этих суспензий выше вязко-
сти плазмы. Однако при дальнейшем повышении гематокрита не
только увеличивается вязкость суспензии, но и выявляются ее не-
ньютоновские свойства, причем прежде всего неньютоновские свой-
ства обнаруживаются при очень низких скоростях сдвига. При-
чины увеличения вязкости при повышении гематокрита мы уже
рассматривали в этой главе, когда знакомились с поведением сус-
пензий. Неньютоновское поведение крови при очень низких ско-
ростях сдвига частично обусловлено свойствами образующихся в
ней монетных столбиков. Наблюдения при помощи микроскопа за
поведением крови в прозрачном вискозиметре типа конус — пло-
скость показали, что по мере повышения скорости сдвига сред-
няя длина монетных столбиков уменьшается вплоть до полного их
распада, и можно думать, что такое изменение сопровождается
снижением вязкости (разд. 10.1). В связи с этим следует напом-
нить, что при вращении в сдвиговом потоке монетные столбики то
изгибаются, то распрямляются, и это, вероятно, также уменьшает
диссипацию энергии и соответственно снижает вязкость. Однако
такое объяснение нельзя признать полностью удовлетворительным,
Гл. 10. Кровь 213
поскольку суспензия эритроцитов в физиологическом растворе так-
же проявляет неньютоновские свойства, хотя в такой среде эритро-
циты и не образуют монетных столбиков.
Итак, следует сделать вывод, что частично за неньютоновское
поведение крови ответственны одиночные эритроциты. Как уже
отмечалось, в суспензии, в которой существует напряжение сдвига,
капли жидкости деформируются. Данные, полученные при изуче-
нии разбавленных суспензий эритроцитов, указывают на то, что
находящиеся в сдвиговом потоке эритроциты тоже деформируются
(см. выше). Поэтому представляется вероятным, что одной из
причин неньютоновского поведения крови являются именно де-
формация и вращение эритроцитов. Веские доводы в пользу та-
кого представления дают исследования, проведенные с эритроци-
тами, жесткость которых была искусственно увеличена. Даже при
гематокрите 40% суспензия таких клеток в солевом растворе ве-
дет себя (подобно суспензии жестких сфер, разд. 10.1) почти
как ньютоновская жидкость.
Эти наблюдения позволяют предположить, что вязкость крови
в значительной мере определяется способностью эритроцитов де-
формироваться и что при заданных значениях гематокрита и ско-
рости сдвига вязкость суспензии эритроцитов с повышенной же-
сткостью будет превышать вязкость суспензий обычных эритроци-
тов. Вообще говоря, как показано на рис. 10.19, это предположе-
ние подтверждается экспериментально: при гематокрите выше
30% и при скорости сдвига 230 с-1 вязкость суспензии эритроци-
тов, жесткость которых искусственно увеличена, значительно пре-
восходит вязкость суспензии обычных эритроцитов и приближает-
ся к вязкости суспензии твердых сферических частиц. Аналогич-
ные данные получены и для суспензий сферулированных и, следо-
вательно, менее гибких эритроцитов (разд. 10.4), а также для сус-
пензий более жестких эритроцитов, присутствующих в крови боль-
ных, страдающих серповидноклеточной анемией. Когда объемная
концентрация (эквивалент гематокрита) жестких сфер в суспензии
приближается к 60%, вязкость ее стремится к бесконечности, т. е.
в такой суспензии не может возникнуть деформации сдвига, и, сле-
довательно, она не может течь. Поразительно, что суспензия
обычных эритроцитов способна течь даже при гематокрите 98%!
Глава 11
СЕРДЦЕ
Сердце млекопитающего состоит из Двух насосов, соединенных
друг с другом последовательно, т. е. так, что выход одного оказы-
вается в конечном счете входом второго, а выход второго — вхо-
дом первого. Поскольку эмбриологически оба этих насоса разви-
ваются путем дифференцировки единой структуры, неудивительно,
что они тесно связаны анатомически и имеют ряд общих свойств.
У них единый механизм возбуждения, обеспечивающий почти син-
хронное действие обоих насосов. Далее — совершенно особый тип
мускулатуры, сердечная мышца, которая по гистологическому
строению близка к скелетным мышцам, но функционально суще-
ственно от них отличается. Сходным является также расположе-
ние камер и наличие клапанов одностороннего действия. Поэтому
нередко полагают, что и работать оба насоса должны одинаково.
В итоге стало общепринятым исследовать свойства одного насоса
(обычно левого) и считать, что результаты приложимы также и к
другому. Однако такой подход нередко бывает неоправдан, осо-
бенно при изучении механики сердца, и потому наши знания о ме-
ханике правых камер сердца и легочного кровообращения остают-
ся весьма неполными. Кроме того, следует помнить, что, поскольку
возможности экспериментов на сердце человека крайне ограни-
чены, приходится опираться главным образом на эксперименталь-
ные данные, полученные на животных, так что дальнейшее описа-
ние будет относиться главным образом к сердцу собаки.
Многие факторы, оказывающие влияние на работу сердца, не
будут предметом рассмотрения в этой главе. К наиболее важным
из них относятся разнообразные рефлексы. Например, нервные
окончания, расположенные в стенке аорты и в каротидных сину-
сах, чувствительны к растяжению и, соответственно, к изменениям
артериального давления. Снижение артериального давления, чем
бы оно ни вызывалось, уменьшает частоту нервных импульсов,
возникающих в этих окончаниях. Такое изменение передается в
мозг и, изменяя импульсацию в нервах, подходящих к сердцу, вы-
зывает увеличение частоты и силы его сокращений. Это в свою
очередь увеличивает производительность сердца и способствует
восстановлению давления в аорте. Ряд таких рефлексов, а также
гормоны, поступающие к сердцу с током крови, непрерывно изме«
няют его деятельность. Но, поскольку действие подобных факто-
Гл. 11. Сердце 215
ров обусловлено физиологическими процессами, которые исходят
не от самого сердца, мы не будем учитывать их в этой главе. Та-
ким образом, не ставя перед собой задачу дать всестороннее опи-
сание работы сердца в целом организме, мы рассмотрим лишь осо-
бенности сердечной мышцы и механические свойства сердца как
насоса.
Два насоса, составляющие сердце, должны действовать в ши-
роком диапазоне параметров и быть при этом очень хорошо согла-
сованными. Действительно, минутный объем сердца у человека,
составляя в покое около 5 л-мин-1, при напряженной мышечной
работе может достигать 25 л-мин-1. Однако способность сосудов
малого круга кровообращения изменять свой объем (составляю-
щий в нормальном состоянии 0,5—1 л) весьма ограничена, и по-
тому производительность обоих насосов должна быть одинакова
всегда, за исключением лишь очень небольших отрезков времени.
При мышечной работе увеличивается как частота сокращений
сердца, так и его производительность за каждый удар (ударный
объем). Поэтому представляется вероятным, что мышечные во-
локна стенки сердца способны изменять и временной ход, и ампли-
туду своего сокращения. Отсюда следует, что познание собствен-
ных сократительных свойств сердечной мышцы имеет основопола-
гающее значение для любых исследований механики сердца. В те-
чение последних 20 лет интенсивное изучение этих свойств вдох-
новлялось предшествовавшими успехами количественного описа-
ния механики сокращения скелетных мышц, и теперь стали оче-
видными глубокие различия функциональных свойств сердечной и
скелетных мышц. Эти различия усложняют изучение особенностей
сердечной мышцы, а различие экспериментальных методов и тер-
минологии еще более усугубляет ситуацию. Из-за сложности строе-
ния и геометрии желудочков сердца попытки связать присущие
выделенным из сердца мышечным полоскам способность развивать
напряжение и укорачиваться со способностью целого желудочка
развивать давление и, уменьшая свой объем, изгонять кровь по-
влекли за собой много интересных вопросов. Вопросы эти допол-
нительно усложняются из-за отсутствия точных сведений о внут-
ренних потерях энергии в мышце и о последовательности и син-
хронности активации сердечной мышцы.
Таким образом, хотя теперь все полнее раскрываются детали
ультраструктуры сократительного аппарата сердца, наше понима-
ние механики миокарда все еще остается неполным. Несмотря на
постоянно возрастающую точность измерений различных гидроди-
намических параметров как у животных, так и у человека и изу-
чение изменений этих параметров в различные фазы сердечного
цикла, поведение сердечной мышцы все еще рассматривается на
основе весьма приблизительных моделей, а отсутствие точных гео-
метрических представлений позволяет описать поведение сокра-
щающихся камер сердца только качественно,
216 Часть II. Механика кровообращения
11.1. Анатомия сердца
Каждый из сердечных насосов имеет камеру низкого давления
(предсердие), которая наполняется кровью из венозной системы
и через клапан одностороннего Действия перекачивает ее в ка-
меру высокого давления (желудочек). В свою очередь желудочек
через второй клапан одностороннего действия направляет кровь в
артериальную систему (рис. 11.1). По венам большого круга пра-
вые камеры сердца получают кровь, оттекающую от тканей тела,
и перекачивают ее в артерии малого круга (к легким). Левые ка-
меры сердца получают эту кровь из вен малого круга и через аор-
ту и ее ветви направляют в артериальное русло большого круга.
Строение обоих предсердий сходно: их стенки тонки и относи-
тельно податливы. Друг от друга их отделяет общая стенка —
межпредсердная перегородка. Вены, по которым в предсердия по-
ступает кровь, соединяются с ними без клапанов. Строение клапа-
нов, отделяющих предсердия от желудочков (атрио-вентрикуляр-
ные клапаны), несколько различно: клапан правых камер (трех-
створчатый) имеет три створки, а клапан левых (двухстворчатый,
или митральный)—две. Створки представляют собой лепестки, ко-
торые одним краем прикреплены к фиброзному кольцу в стенке
сердца, а двумя свободными краями обращены в полости желу-
дочков. Створки клапанов исключительно тонки (около 0,1 мм)
и состоят главным образом из сети коллагеновых и эластических
волокон, покрытой слоем таких же клеток, которые выстилают
внутреннюю поверхность камер сердца и кровеносных сосудов
(эндотелий). Свободные края створок митрального и трехстворча-
того клапанов соединены со стенками соответствующих желудоч-
ков тонкими фиброзными тяжами (сухожильными нитями), кото-
рые соединяются с мышечными пучками (сосочковые, или папил-
лярные мышцы), идущими от стенок желудочков. Каждая нить
соединяется со свободными кромками двух створок в тех местах,
где они соприкасаются друг с другом при закрывании клапана. Та-
ким образом, натягиваясь при сокращении сосочковых мышц в на-
чале систолы, нити не позволяют клапану вывернуться в предсердие
под действием нарастающего внутрижелудочкового давления и
тем самым препятствуют обратному движению крови. Вероятно,
существует самоподдерживающийся гидродинамический меха-
низм закрытия этих клапанов (разд. 11.5) в конце систолы пред-
сердия.
Выходные клапаны желудочков (легочный и аортальный) очень
похожи друг на друга. Каждый состоит из трех створок, свобод-
ные края которых начинаются непосредственно от стенок клапан-
ного кольца. Такое расположение их позволяет открывать всю
площадь поперечного сечения клапанного кольца без изменения
формы створок. Эти створки лишены натягивающих сухожильных
нитей, но тем не менее могут противостоять значительной разно-
Гл. И. Сердце 217
Рис. 11.1. Схема анатомического строения сердца и направлений движения крови.
сти давлений [в случае аортального клапана приблизительно
1,3-104 Н-м~2 (100 мм рт. ст)]. Кроме того, выходные клапаны
чрезвычайно эффективны, поскольку закрываясь (что происходит
свыше 30 миллионов раз в год) они пропускают лишь очень не-
большой обратный поток крови. Неудивительно, что механизм
их работы издавна вызывал интерес ученых. В XV столетии Лео-
нардо да Винчи исследовал строение этих клапанов и построил
предположительные схемы движения крови через них, оказав-
шиеся впоследствии поразительно близкими к действительности.
В последнее время механизм действия клапанов подробно изучен
(разд. 11.5).
Четыре клапанных отверстия в сердце расположены прибли-
зительно в одной плоскости. Основания створок каждого клапана
прикрепляются к жесткому кольцу из фиброзной ткани (рис. 11.2).
Все четыре кольца в свою очередь соединены между собой фиб-
розной тканью, так что клапанные устройства располагаются в
жестком каркасе, к которому прикрепляются мышечные волокна
каждой камеры: предсердия с одной стороны, желудочки — с дру-
гой; легочная артерия и аорта также прикреплены своими началь-
ными участками к этой «пластине» фиброзной ткани. Сердце в
целом заключено в перикард — сумку из тонкой фиброзной тка-
ни, — который в свою очередь прикрепляется к другим структурам
внутри грудной клетки, а через них — к позвоночному столбу. Со-
отношение между напряжением и деформацией для перикарда,
как следует из рис. 11.3, очень далеко от линейного. При нормаль-
ном диастолическом объеме сердца перикард растянут лишь не-
значительно и, вероятно, не оказывает существенного влияния на
наполнение сердца. Однако при некоторых заболеваниях, когда в
полости перикарда накапливается жидкость, он может в силу
своей жесткости ограничить диастолическое наполнение желудоч-
218 Часть И. Механика кровообращения
Рис. 11.2. Взаиморасположение различных структур сердца. К «скелету» сердца,
состоящему в основном из фиброзных клапанных колец, прикрепляются также
предсердия и желудочки и две главные артерии. (Рашмер Р.Дицамика сердеч-
но-сосудистой системы. Пер. с англ. — М.: Медицина, 1981.)
ков, в результате чего производительность сердца резко умень-
шается (см. стр. 559).
Левый желудочек является единственной камерой сердца,
строение стенки которой изучено систематически. Произведенные
в самое последнее время микроскопические исследования серий-
ных срезов миокарда левого желудочка показали, что старые
представления, согласно которым стенки желудочков образованы
четко определяемыми мышечными слоями, неверны. Оказалось,
что в действительности ориентация волокон имеет строго упоря-
доченное непрерывное распределение (рис. 11.4). Самые внутрен-
ние (субэндокардиальные) волокна идут преимущественно про-
дольно от фиброзной области вокруг клапанов (основание сердца)
к другому концу приблизительно эллипсоидной камеры (верхуш-
ка); чуть более удаленные волокна лежат под небольшим углом
к оси камеры и к верхушке они идут как бы по спирали. Для во-
локон более глубоких слоев угол постепенно увеличивается, так
что приблизительно на середине стенки они проходят параллельно
наименьшей оси камеры, т. е. по окружности. Затем угол продол-
жает увеличиваться, и волокна наружной поверхности стенки же-
лудочков (эпикардиальные) снова идут продольно. Такое распо-
ложение позволяет желудочку развивать значительные усилия, не-
Гл. 11. Сердце 219
Рис. 11.3. График давление — объем для перикарда собаки. [Holt (1971). Circu-
lation Res., 8, 1171. С любезного разрешения American Heart Association, Inc.]
Рис. 11.4. Ориентация мышечных волокон в стенке левого желудочка. А. Ориен-
тация продольной оси волокон на разной глубине от наружной поверхности. Б.
Изменение угла между волокнами и плоскостью, перпендикулярной продольной
оси левого желудочка, по толщине стенки. [Streeter et al. (1969). Circulation
Res., 24, 339.]
220 Часть II. Механика кровообращения
смотя на то что по отдельности мышечные волокна могут выдер-
живать только аксиальную нагрузку. Это обусловлено тем, что на-
пряжение в стенке, действующее в любом направлении, всегда
встречает сопротивление по крайней мере части мышечных воло-
кон и не существует каких-либо направлений или плоскостей, ко-
торые отличались бы меньшей способностью нести нагрузку. Заме-
тим, что волокна не должны обязательно оканчиваться у верхуш-
ки; они могут поворачивать и по спирали направляться обратно
к основанию, подобно шнуру, спиралью навитому на палку. Как
видно из рис. 11.4,£>, наибольшее количество волокон проходит
по окружности желудочка.
11.2. Сердечный цикл
Электрические явления. Цикл сокращения сердца начинается
в определенной области стенки правого предсердия, которую на-
зывают водителем ритма, или сино-атриальным узлом. Мышечные
клетки этой области особые: они способны периодически деполя-
ризоваться и реполяризоваться (второй процесс зависит от энер-
гии, извлекаемой в конечном итоге из метаболических процессов
в самих клетках). Начавшаяся в водителе ритма деполяризация
распространяется со скоростью порядка 1 м-с-1 по окружающим
эту область мышцам стенок правого и левого предсердий (вызы-
вая их сокращение). Затем она передается клеткам атрио-вентри-
кулярного узла, расположенного в межпредсердной перегородке.
Далее деполяризации подвергается особый пучок мышечных воло-
кон (атрио-вентрикулярный пучок, или пучок Гиса), который про-
ходит через фиброзную ткань, окружающую трех створчатый кла-
пан, в межжелудочковую перегородку; здесь этот пучок делится
и проникает в мышечную массу каждого желудочка, оканчиваясь
среди обычных мышечных клеток сетью особых тонких мышечных
волокон (волокна Пуркине). Волна деполяризации распростра-
няется по этому проводящему пути быстро (5 м-с-1), и потому
деполяризация и сокращение всех мышечных волокон как левого,
так и правого желудочков происходят относительно синхронно:
потенциал деполяризации желудочков на электрокардиограмме —
комплекс QRS— длится меньше 0,1 с. Определенные нервные и
гормональные влияния, берущие начало вне сердца, могут изме-
нять частоту работы водителя ритма, а также скорость распро-
странения по сердцу волны деполяризации. Но организованная по-
следовательность сокращения предсердий и желудочков, возникаю-
щая после деполяризации водителя ритма, обеспечивается глав-
ным образом расположением и свойствами проводящих путей.
Циклы деполяризации и реполяризации, происходящие в сер-
дечной мышце, генерируют слабые электрические потенциалы, и
при соответствующем расположении электродов эти потенциалы
могут быть восприняты на поверхности тела, усилены и зареги-
Гл. 11. Сердце 221
Время, с
.Рис. 11.5. Полусхематическое изображение процессов, происходящих за время
сердечного цикла в левых камерах сердца. Все значения давлений даны по отно-
шению к атмосферному давлению. Происхождение а- и v-волн на записи давле-
ния в предсердии обсуждается в тексте.
стрированы в виде электрокардиограммы. Типичная запись пока-
зана в верхней части рис. 11.5. Деполяризация предсердий вызы-
вает небольшое отклонение, называемое зубцом Р\ с задержкой
около 0,2 с за этой волной следует более резкое колебание потен-
циала, часто многофазное, именуемое комплексом QPS. Оно от-
ражает деполяризацию обоих желудочков. Вслед за ним возни-
кает последний компонент, зубец Т, который генерируется во вре-
мя реполяризации желудочков. Временные соотношения между
этими суммарными электрическими потенциалами и механиче-
скими событиями в левых камерах сердца можно проследить по
рис, 11.5, который показывает изменения давления в левом пред-
222 Часть П. Механика кровообращения
150т
Рнс. 11.6. Полусхематическое изображение изменений за время сердечного цикла
давления в левом и правом желудочке, а также давления и скорости кровотока
в аорте и в главной легочной артерии.
сердии, левом желудочке и в аорте, а также расхода крови в ней
за время полного сердечного цикла.
Механические явления. Как было только что сказано, о на-
чале сокращения желудочков (точнее, о деполяризации их мышцы)
сигнализирует комплекс Q/?S электрокардиограммы. Оба желу-
дочка сокращаются почти синхронно (рис. 11.6), но для простоты
мы рассмотрим лишь явления, происходящие в левых камерах
сердца. Последовательность событий показана на рис. 11.5. Через
очень короткий интервал времени после деполяризации мышечные
волокна стенки желудочка начинают развивать активное напря-
жение (см. стр. 228), и давление в желудочке возрастает. На этой
стадии аортальный клапан еще остается закрытым, так как дав-
ление в аорте превышает давление в левом желудочке, а створки
митрального клапана сближаются по мере уменьшения потока
крови из предсердия в желудочек. Такое состояние весьма кратко-
временно, так как давление в желудочке почти сразу становится
выше давления в предсердии, и в течение очень короткого периода
времени кровь течет обратно — из желудочка в предсердие. Этот
период заканчивается закрытием митрального клапана, что сопро-
вождается характерным звуком, который прослушивается через
Гл. II. Сердце 223
стенку грудной клетки; этот звук — первый тон сердца — отмечает
начало систолы, т. е. периода сокращения желудочков. Второй тон
сердца соответствует началу диастолы, т. е. периода расслабления
желудочков. Отметим, что эти периоды определены в соответ-
ствии с тонами сердца, а не с механическим поведением его мышц
или с электрокардиограммой *).
Напряжение стенки желудочка теперь начинает расти исклю-
чительно быстро, и этот рост продолжается до тех пор, пока дав-
ление в полости желудочка не превысит давление в аорте. В этот
период никакого изменения объема желудочка не происходит, по-
скольку кровь фактически несжимаема, и потому этот период на-
зывают изоволюмическим. (В старой литературе его часто назы-
вали изометрическим периодом; однако в настоящее время ясно,
что в действительности желудочек в эту фазу изменяет свою фор-
му, хотя объем его сохраняется постоянным, и потому термин изо-
волюмический является предпочтительным.) Как только давление
внутри желудочка превысит давление в аорте, возникает система
сил, действующих так, чтобы открыть аортальный клапан, и на-
чинается следующая фаза систолы — фаза изгнания крови. По-
скольку объем левого желудочка уменьшается, кровь, находя-
щаяся в желудочке и в проксимальной части аорты, быстро при-
обретает направленное вперед ускорение. Затем напряжение стен-
ки желудочка падает, разность давлений в желудочке и в аорте
меняет знак и движение крови в аорте замедляется. Наконец, пе-
ред закрытием аортального клапана, сопровождающимся вторым
тоном сердца, существует небольшой период времени обратного
движения крови через этот клапан — из аорты в желудочек. За
этим следует второй изоволюмический период, во время которого
мышца желудочка расслабляется и давление в нем падает. В то
же самое время давление .в левом предсердии повышается по мере
того, как оно наполняется кровью из легочных вен (v-волна). Как
только это давление превысит давление в левом желудочке, мит-
ральный клапан вновь открывается и под влиянием разности дав-
лений, создаваемой сначала пассивно, а затем за счет активного
укорочения мышечных волокон стенки предсердия, происходит на-
полнение желудочка. Это активное сокращение предсердия (си-
стола предсердия) проявляется на электрокардиограмме зубцом Р,
а механически — кратковременным увеличением давления в пред-
сердии (a-волна). Вскоре после этого происходит активация
мышцы желудочка и весь цикл начинается снова.
У здоровых людей частота сокращений сердца может изменять-
ся от 45 (у спортсменов в состоянии покоя) до 200 в 1 мин (при
максимальной мышечной работе). При низких частотах систола
!) Поэтому систола не соответствует строго периоду изгнания крови из же-
лудочков, хотя использование этого термина в таком смысле широко распро-
страняю.
224 Часть II. Механика кровообращения
значительно короче диастолы и занимает около одной трети дли-
тельности всего цикла (рис. 11.5). По мере увеличения частоты
сокращений сердца диастола укорачивается значительно быстрее
систолы, и при самых высоких частотах длительность обеих фаз
может быть почти одинаковой. В нормальных условиях объем
крови, изгоняемой из желудочка при каждом ударе (ударный
объем), составляет в покое 70—100 см3; несколько меньше крови
остается в желудочке —т. е. желудочек изгоняет лишь 60—70%
своего содержимого. Изменение ударного объема при мышечной
работе выражено в значительно меньшей степени, чем изменение
частоты сокращений сердца. Поэтому при тяжелой мышечной ра-
боте увеличение производительности (минутного объема) сердца
(в 5 раз и выше) обеспечивается в первую очередь повышением
частоты его сокращений, а не ударного объема1). Давление крови
в легочной артерии и в аорте при мышечной работе повышается,
но это повышение весьма умеренно по сравнению с увеличением
общего потока крови, поскольку раскрытие ранее закрытых мель-
чайших артериальных сосудов и расширение открытых сосудов
снижает периферическое сопротивление течению крови.
11.3. Свойства сердечной мышцы
Строение. Под световым микроскопом миокард предстает в
виде ряда тяжей, состоящих из тонких вытянутых мышечных кле-
ток с центрально расположенным ядром и многочисленными мито-
хондриями (рис. 11.7). Как и в скелетных мышцах, волокна мио-
карда выглядят поперечноисчерченными, что обусловлено струк-
турой сократительных элементов, или миофибрилл, лежащих внут-
ри этих мышечных клеток. Однако в сердечной мышце отсутствуют
имеющиеся в скелетной мышце двигательные нервные волокна,
нервно-мышечные соединения и мышечные веретена, контролирую-
щие ее длину. Другое отличие заключается в том, что мышечные
клетки миокарда многократно ветвятся, а между ними имеется
множество коллагеновых волокон.
') Необходимо специально подчеркнуть, что само по себе повышение ча-
стоты сокращений сердца способно привести лишь к незначительному повышению
его производительности — всего на несколько процентов прн удвоении частоты,
а при ее дальнейшем росте (более 120—140 в 1 мин) производительность сердца
даже снижается вследствие уменьшения длительности диастолы, а соответствен-
но и степени наполнения желудочков Точно так же само по себе увеличение
силы сокращения желудочков и рост ударного объема не в состоянии обеспечить
устойчивое увеличение минутного объема кровообращения. Рост и частот®, и
силы сокращений сердца — это лишь условия возможности повышения его про-
изводительности, для реализации которого необходимо увеличение притока крови
к сердцу. При мышечной работе последнее достигается сочетанием снижения со-
противления сосудов работающих мышц, ограничения роста объема вен (за счет
увеличения их жесткости), работы мышечного насоса (гл. 14), повышения цен-
трального венозного давления и изменения характеристик работы сердца. —
Прим. ред.
Гл. 11. Сердце ЯМ
Рив. 11.7. Электронная микрофотография части трех волокон миокарда, приле-
жащих к капилляру (/); продольный срез. Две верхние клетки соединяются
друг с другом конец в конец через типичный вставочный диск (2). Ряды мито-
хондрий (3) как будто бы делят сократительные структуры на единицы, подоб-
ные миофнбриллам, но в отличие ог истинных миофнбрилл скелетной мышцы эти
единицы разветвляются и вновь соединяются; кроме того, они различаются по
ширине. Между концами митохондрий в разных местах препарата видны капель-
ки жира (4); форма некоторых из них искажена; X15000. [Fawcett, McNutt
(1969). The ultrastructure of the cat myocardium, J. Cell Biol., 42, 1—46.]
Предел разрешения светового микроскопа составляет около
0,25 мкм, и на этом уровне различить дальнейшие детали невоз-
можно. Поскольку в этих условиях не были обнаружены клеточ-
ные мембраны, идущие поперек волокон, раньше полагали, что по-
следние образуют синцитий, т. е. допускали непрерывность их ци-
топлазмы. Однако электронная микроскопия показала, что кле-
226 Часть tl. Механика кровообращения
точные мембраны имеют два слоя, причем внутренний слой пере-
секает волокна и делит их через каждые 50—100 мкм на струк-
турно изолированные ячейки диаметром 10—20 мкм. С интерва-
лами порядка 2 мкм по всей длине каждой такой ячейки имеются
исключительно тонкие впячивания мембраны, названные Т-ка-
нальцами. Показано, что при деполяризации мембраны эти ка-
нальцы обеспечивают почти одновременную активацию всех мио-
фибрилл в клетке.
Описано множество других тонких деталей структуры клеток, и
многие современные достижения связаны с выяснением биохими-
ческих реакций, при которых высвобождается энергия для сокра-
щения и реполяризации. Однако здесь мы остановимся подробно
лишь на сократительном аппарате, поскольку нас интересует ме-
ханика мышечных волокон.
Как было отмечено выше, сократительные элементы каждой
клетки — это миофибриллы. Они ориентированы вдоль длинной
оси клетки и заметны вследствие повторяющегося узора — попе-
речных полос. Миофибриллы состоят из пучков микронитей (мио-
филаментов), а поперечная исчерченность возникает из-за того,
что миофибриллы построены из повторяющихся цепочек саркоме-
ров. Саркомер является элементарным сократительным аппара-
том; его структура была впервые описана для скелетной мышцы
и оказалась такой же для миокарда лишь с некоторыми отли-
чиями. Каждый саркомер ограничен двумя соседними узкими по-
лосами (или линиями),соответствующимиZ-пластинкам (рис. 11.8).
Участок саркомера между этими линиями состоит из широкой
центральной полосы (A-диска), отделенной от каждой Z-пластин-
ки узкой, более светлой полосой (1-диском). Когда мышечное во-
локно растягивается, расстояние между Z-пластинками увеличи-
вается, но размер A-дисков сохраняется неизменным. Все это раз-
личимо под световым микроскопом. При увеличении общей длины
мышцы длина саркомеров увеличивается и для сердечной мышцы
при ее диастолических размерах составляет около 2,2 мкм. Ши-
рина A-диска составляет при этом 1,5 мкм, а каждого из 1-дис-
ков — по 0,35 мкм.
Изящные электронно-микроскопические исследования показали,
что изображения дисков возникают благодаря частичному пере-
крыванию параллельных пучков нитей, расположенных как пока-
зано на рис. 11.8. Более толстые нити, образующие A-диски, со-
стоят преимущественно из белка миозина-, входящие между ними
более тонкие нити состоят из белка актина. Когда мышца растяги-
вается, Z-пластинки «раздвигаются»; актиновые нити при этом
скользят вдоль миозиновых и частично выходят из области пере-
крывания. В результате 1-диски расширяются. Когда мышца со-
кращается, происходит обратное, пока при очень малой длине
мышцы 1-диски не исчезают, а в центрах A-дисков, где концы ак-
тиновых нитей частично перекрываются друг с другом, не возни-
Гл. 11. Сердце 227
' ' - Саркомер"(2,2:мкм)
М-линия
4 V
г- линия; н-зона jz-
диск' ' (Ъ5 мкм) ; ДР
: : . *
Б'
актиновая J ~~....’.... ' ‘ миозинов:
нить нить
Рис 11 8. Тонкая структура волокон миокарда А Электронная микрофотогра-
фия саркомера Саркомер ограничен двумя Z-пластинками. Между ними виден
темный центральный A-диск (в середине его выделяется еще более темная М-ли-
ния), а также два более светлых I диска Б Схема расположения нитей сократи-
тельных белков актина и миозина, придающих саркомеру исчерченность Обра-
тите внимание, что при такой длине саркомера актиновые нити в середине не
перекрываются и «полосы сокращения» не видны. Подробнее см. в тексте.
[Sonnenbick, Spiro, Spotmtz (1964). Am Heart J., 68, 336.]
кают новые темные полосы («полосы сокращения»). Это происхо-
дит при длине саркомеров меньше 1,5 мкм.
В скелетных мышцах между актиновыми и миозиновыми ни-
тями имеются тонкие поперечные мостики, идущие от каждой тол-
стой (миозиновой) нити. Вероятно, на каждую молекулу миозина
приходится один мостик; основания мостиков отделены друг от
друга расстоянием около 6 нм и расположены вокруг миозиновой
нити с шагом 60° вдоль спирали. Каждая миозиновая нить окру-
жена шестью актиновыми нитями и, следовательно, на расстоя-
нии около 40 нм с каждой из них образует один поперечный мо-
стик.
Поперечные мостики, по-видимому, играют важную роль в про-
цессе сокращения поперечнополосатых мышц, так как прямое
взаимодействие между нитями маловероятно: они слишком уда-
лены друг от друга. Одно из предположений заключается в том,
что во время сокращения поперечные мостики движутся взад и
вперед, зацепляясь за определенные участки актиновых нитей, пе-
редвигая их вплоть до разрыва связи и возвращаясь к новому
участку зацепления. Таким образом, при активации мостики дей-
288 Часть II. Механика кровообращения
Длина
Рис. 11.9. Типичные кривые длина — напряжение для скелетной (А) и сердечной
(Б) мышцы. В каждом случае напряжение в покое (/) и активное напряжение
(П) отложены по оси ординат и выражены в килограммах на квадратный сан-
тиметр площади поперечного сечения мышцы. Обратите внимание на различие
масштаба на этих двух графиках. Длину мышцы устанавливали предварительно
и поддерживали постоянной во время сокращения, которое вызывали электриче-
ским раздражением. По оси абсцисс отложена длина саркомера, отнесенная к той
его длине Lmax, при которой развивается максимальное активное напряжение;
в этих опытах Lmax равнялась 2,2 — 2,3 мкм как для скелетной, так и для сердеч-
ной мышцы. [Spiro, Sonnenblick (1964). Comparison of the ultrastructural basis
of the contractile process in heart and skeletal muscle, Circulation Res., 15, sup-
plement 2, pp. 14—37. По разрешению American Heart Association, Inc.]
ствуют подобно храповому механизму; по прекращении активации
нити освобождаются и Пассивно раздвигаются обратно. В настоя-
щее время такое объяснение остается умозрительным, особенно
в отношении сердечной мышцы.
Статические механические свойства сердечной мышцы. Приве-
денное выше описание свойств саркомеров соответствует гипотезе
«скользящих нитей». Эта гипотеза имеет две сильные стороны: она
не только согласуется с наблюдаемой ультраструктурой мышцы,
но и предлагает объяснение одного из ее основных механических
свойств — специфического соотношения между длиной и напря-
жением (усилием). Если при раздражении электрическим током
длину мышцы удерживать постоянной, то она развивает напряже-
ние (активное, или развиваемое, напряжение), превышающее лю-
бое напряжение пЬкоя, имевшееся до раздражения. Если такой
эксперимент повторять, последовательно увеличивая длину мышцы
небольшими шагами, то оказывается, что активное напряжение
постепенно растет до максимума, а затем падает (рис. 11.9). Это
справедливо для самых разных мышц, и для скелетной мышцы
(а, вероятно, также и для сердечной) график длина — напряже-
Гл. 11. Сердце 229
ние достигает пика, когда растяжение доводит длину саркомера
до величины порядка 2,2 мкм. Как видно из рис. 11.9, в абсолют-
ных величинах (т. е. в расчете на единицу площади поперечного
сечения) сердечная мышца значительно «слабее», чем скелетная.
Быть может, это скорее кажущееся различие, чем действительное,
так как сердечная мышца содержит больше элементов, н₽ относя-
щихся к сократительным (коллаген и митохондрии), и не все ее
мышечные волокна параллельны. В мышце, сократившейся до
длины саркомеров меньше 2,2 мкм, актиновые нити проходят за
середину миозиновых и перекрываются с актиновыми нитями про-
тивоположной стороны. Полагают, что это препятствует образо-
ванию поперечных связей, в результате чего развиваемое напряже-
ние оказывается меньше максимального. Такое перекрывание по
мере растяжения мышцы уменьшается до тех пор, пока при длине
саркомеров 2,2 мкм не исчезнет двойное перекрывание актиновых
и миозиновых нитей. Именно при этой длине может быть образо-
вано наибольшее количество поперечных связей и развито макси-
мальное активное напряжение. При дальнейшем растягивании сар-
комеров актиновые нити постепенно выдвигаются из пространства
между миозиновыми нитями и могут образовывать с ними все
меньше поперечных связей. В этом диапазоне длин активное на-
пряжение, развиваемое при сокращении, линейно уменьшается с
увеличением длины до тех пор, пока при длине саркомеров около
3,5 мкм актиновые нити не выдвинутся полностью и мышца не
перестанет развивать какое бы то ни было активное напряжение.
Для скелетных мышц такая связь между длиной саркомеров и
напряжением твердо установлена и представление о ее функцио-
нальном значении общепринято. Поведение саркомеров сердечной
мышцы изучено значительно менее основательно. В физиологиче-
ских условиях они, по-видимому, работают в диапазоне длин при-
мерно от 1,5 до 2,2 мкм. Длина саркомеров приближается к 2,2 мкм,
когда диастолическое давление в желудочке составляет около
12 мм рт. ст., что является верхним пределом нормы, и потому
гипотеза скользящих нитей дает очень заманчивое объяснение со*
отношения между длиной и напряжением для миокарда. Однако
именно для миокарда исследований зависимости между длиной
саркомеров и напряжением проведено сравнительно мало, и ре-
зультаты их до некоторой степени противоречивы. Возможно, втй
обусловлено методическими трудностями, особенно связанными е
усадкой ткани при гистологической обработке. Недавно раарабо*
таны методы прижизненного измерения длины саркомеров, и они
могут помочь решению этого вопроса. В настоящее время цред-
ставляется установленным, что при длине саркомеров мвЙЬШе
1,5 мкм активное напряжение не развивается и пик кривой дли-
на — напряжение лежит где-то между 2,0 и 2,2 мкм, как это имеет
место в случае скелетной мышцы. Однако для миокарда все еще
не ясно, является ли увеличение напряжения следствием .посте-
230 Часть II. Механика кровообращения
пенного изменения длины саркомеров (как в случае скелетных
мышц) или же оно обусловлено включением все большего числа
саркомеров тех мышечных волокон, которые при малой длине
мышцы были изогнуты, а по мере удлинения выпрямляются и за-
тем растягиваются. Более того, недавно доказано, что для сердеч-
ной мышцы график зависимости напряжения от длины саркоме-
ров (рис. 11.9) может смещаться вдоль оси абсцисс при измене-
нии химического состава окружающей мышцу среды. Чтобы разо-
браться с этой неопределенностью, необходимы более детальные
сведения о поведении актино-миозиновых поперечных связей.
В скелетной мышце линейное соотношение между длиной мыш-
цы и длиной саркомеров поддерживается до тех пор, пода послед-
няя не достигнет по крайней мере 3,5 мкм, и прогрессирующее
уменьшение числа возможных мест поперечных связей при посте-
пенном раздвигании актиновых и миозиновых нитей дает адекват-
ное объяснение линейного снижения активного напряжения на
нисходящей ветви кривой длина — напряжение. Для сердечной
мышцы при большой степени растяжения возникает другая ситуа-
ция. Саркомеры допускают удлинение только до 2,6 мкм, и даже
в этой области отмеченное выше линейное соотношение не имеет
места. Еще большее увеличение полной длины мышцы требует
создания очень высокого напряжения покоя и, вероятно, связано
с перемещением целых волокон одно относительно другого. В здо-
ровом сердце подобная ситуация не возникает и, по-видимому,
очень маловероятна даже в случае ослабленного или патологиче-
ски измененного сердца. Однако в остром опыте, в котором рас-
слабленный левый желудочек растягивали давлением до 100 мм
рт. ст. (это значительно выше уровня, наблюдаемого даже при тя-
желых заболеваниях сердца), средняя длина саркомеров в стенке
желудочка не превышала 2,3 мкм.
Динамические механические свойства сердечной мышцы.
Кривая длина — напряжение отображает важное свойство мышцы
в статических условиях, т. е. при сохранении постоянной длины
мышцы как до, так и во время ее сокращения, но ничего не гово-
рит о динамике мышечного сокращения, которая принципиально
важна для понимания механизма действия сердечной мышцы.
При возбуждении мышца переходит в состояние механической
активации («активное состояние»), которое отражает высвобож-
дение энергии в ходе химических реакций. Удается измерить как
длительность, так и интенсивность этого состояния. Большие ус-
пехи достигнуты в выявлении и количественном описании этапов
биохимических процессов, служащих источником энергии, а также
в исследовании механического поведения мышцы, отражающего
это высвобождение энергии. Литература, касающаяся связанных
с исследованием миокарда методических трудностей и присущей
ему сложности, очень обширна, и эти вопросы мы можем рассмот-
реть лишь вкратце.
Гл. 11. Сердце 231
Для изучения механических свойств сердечной мышцы обычно
используют сосочковую мышцу, удаленную из правого желудочка
молодых наркотизированных животных. Таким образом удается
получать исключительно тонкие полоски мышцы длиной несколько
миллиметров, состоящие из множества достаточно параллельных
друг другу мышечных волокон. Такие сосочковые мышцы, поме-
щенные в снабжаемую кислородом питательную среду с соответ-
ствующими ионным составом и осмотическими свойствами, в те-
чение длительного времени сохраняют способность сокращаться в
ответ на электрическое раздражение. Сократительные свойства
этих мышц интерпретируют исходя в основном из очень простых
механических моделей. Чтобы разъяснить, почему были выбраны
именно такие модели, необходимо вкратце описать опыты, выпол-
ненные ранее на скелетной мышце.
Целые скелетные мышцы можно легко извлечь у мелких жи-
вотных (скажем, у лягушки), и многие исследователи в начале
двадцатого столетия изучали эти мышцы, раздражая их электри-
ческим током и определяя механические свойства и теплопродук-
цию во время сокращения (последнее явление открыл Гельмгольц
более чем на 50 лет раньше).
Как уже отмечалось, электрическое раздражение мышцы при-
водит к развитию в ней напряжения. В состоянии покоя на мемб-
ране мышечной клетки поддерживается разность потенциалов
около 90 мВ — потенциал покоя. Приложенный снаружи толчок
электрического тока может вызвать временное изменение поляр-
ности этого потенциала с последующим медленным восстановле-
нием. Этот разряд, названный потенциалом действия, инициирует
процесс высвобождения ионов кальция в мышечной клетке, и они
каким-то образом активируют поперечные связи между актино-
выми и миозиновыми нитями сократительного аппарата саркоме-
ров, Весь этот процесс осуществляется за миллисекунды, и затем
в течение короткого промежутка времени — пока метаболические
процессы внутри клетки не удалят ионы кальция и мышца не вер-
нется в состояние покоя — мышечная клетка способна сокращаться
(т. е. укорачиваться и/или развивать напряжение).
Таким образом, одиночный электрический стимул, приложен-
ный к мышечному волокну, вызывает кратковременное сокраще-
ние, названное соответственно одиночным сокращением. Серия
стимулов вызывает повторяющиеся сокращения, и если частота
стимулов достаточно высока, эти одиночные сокращения скелет-
ной мышцы сливаются и дают поддерживающееся сокращение.
Такое сокращение названо тетанусом, а соответствующая серия
стимулов — тетаническим раздражением.
Развиваемое мышцей при каждом одиночном сокращении или
при тетанусе напряжение с усилением стимулов увеличивается
232 ¥acn IL Механика кровообращения
--- -'f**- - — - - - - - - - _ ______
до тех пор, пока не будет достигнут максимум напряжения1),
который хорошо воспроизводится при повторных раздражениях
мышцы в течение больших периодов времени. Если длину мышцы
поддерживают постоянной, то одиночное сокращение (или тета-
нус) называют изометрическим. Если же мышца имеет возмож-
ность укорачиваться и силу, противодействующую укорочению
(если она имеется) и называемую нагрузкой, поддерживают по-
Отоянной (последнее подразумевает, что все ускорения, возникаю-
щие при укорочении мышцы, очень малы по сравнению с ускоре-
нием Силы тяжести), то сокращение называют изотоническим. По-
скольку было обнаружено, что максимальные изометрические со-
кращения очень хорошо воспроизводятся, их использовали в опы-
тах как исходное состояние. В этом случае во время цикла стиму-
ляции мышца выделяет тепло, но так как не происходит никакого
укорочения, то не совершается и никакой внешней работы (ра-
бота, или затраченная энергия, равна произведению силы на рас-
стояние) Если же мышце позволяют укорачиваться на длину х
против нагрузки или силы Р, то не только совершается работа Рх,
нй и выделяется добавочное количество тепла [Такое влияние
укорочения на теплопродукцию названо по имени автора этого от-
крытия «эффектом Фенна»; американский физиолог У. Фенн (сде-
лавший эту работу в 1923 г.) показал также, что справедливо и
обратное- если мышцу во время раздражения растягивать, то она
выделяет меньше тепла, чем при поддержании ее длины постоян-
ной. Описывая этот опыт, Фенн ввел понятие «отрицательной ра-
боты», против которого всегда возражали физики. Это очень пе-
чально, поскольку смысл опыта, состоящий в том, что механиче-
ские условия, при которых совершается сокращение, контролируют
количество высвобождающейся энергии, весьма примечателен и не
оценен по достоинству.]
Объяснение выделению дополнительного тепла при укорочении
мышцы было дано через несколько лет, когда появились при-
боры, способные определять теплопродукцию непрерывно, в каж-
дый момент цикла сокращения и расслабления. Именно тогда
Арчибальд Хилл в знаменитой серии экспериментов установил, что
дополнительное количество тепла, связанное с укорочением, про-
порционально величине укорочения х. Поэтому оно эквивалентно
работе ах, где постоянная а имеет размерность силы. Итак, со-
вершенная механическая работа равна Рх; следовательно, полная
дополнительная энергия, высвобождающаяся при укорочении,
есть (Р-\-а)х, а скорость высвобождения энергии — (Р + о)о, где
v и йх/At — скорость укорочения. Экспериментально было пока-
зано, что скорость высвобождения энергии увеличивается при
’) Рост напряжения при увеличении амплитуды электрического раздраже-
ния мышц определяется включением в сокращение все большего количества мы-
шечных волокон — Прим ред.
Гл. 11. typtyf ЯЗЗ
уменьшении нагрузки, достигая максимума, когда нагрузка стано-
вится равной нулю, и обращаясь в нуль, когда мышца развивает
максимальную силу в условиях изометрического сокращения. Эти
соотношения, как следовало из опытов, лучше всего представ-
ляются в виде
(P + a)v = b(P0-P), (11.1)
где b — константа с размерностью скорости. Данное уравнение
можно переписать так:
(Р + a) (v + Ь) = Ь (Ро + а), (41.2)
где правая часть является константой Из (11.2) следует, что сила
Р и скорость сокращения v связаны между собой гиперболической
Рис. 11 10 Соотношение между нагрузкой Р (в граммах веса) н скоростью уко-
рочения v (в см-с-1) скелетной мышцы лягушки при изотоническом сокращении.
Точки — экспериментально измеренные величины; кривая проведена в соответ-
ствии с уравнением (11 2) [Hill (1938). Proc. R. Soc. В, 126, 136.]
зависимостью. Таким образом, свойства, предсказанные на осно-
вании тепловых измерений, оказалось возможным исследовать в
помощью чисто механических экспериментов, что и было сделано
в опытах по определению скорости, с которой мышца может изо-
тонически укорачиваться при различных нагрузках (рис. 11.10).
Тепловые эксперименты имели, однако, большое значение по
другой причине: они подсказали первую простую механическую
модель мышечного волокна. Наблюдения за темпом выделения
тепла на самых ранних стадиях стимуляции выявили, что qh оди-
наков при изометрическом и изотоническом сокращениях. Это на-
водило на мысль об аналогичности механических событий, проис-
ходящих на ранних стадиях сокращения обоих типов. А так как
длина мышечного волокна не может изменяться при изометриче-
ском сокращении, родилась идея о сократительном элементе мыш-
цы, укорачивающемся при стимуляции, но связанном последова-
тельно с упругим элементом, который может растягиваться, если
234 Часть II. Механика кровообращения
длина мышцы поддерживается постоянной (рис. 11.11,Л). При
равномерном укорочении в изотонических условиях длина упругого
элемента должна быть постоянной, и, следовательно, он не дол-
жен оказывать влияние на соотношение сила — скорость. Поэтому
считалось, что это соотношение характеризует свойства сократи-
тельного элемента.
Следует подчеркнуть, что А. Хилл исследовал свойства скелет-
ной мышцы в совершенно особых условиях. Во-первых, мышцу
раздражали сериями стимулов высокой частоты (тетанически),
так что развивался длительный и притом максимальный ответ.
Кроме того, каждое наблюдение производилось при постоянной
Сократительный
элемент
Последователь-
ный упругий
элемент
Рис. 11.11 Механические модели мышцы.
нагрузке и постоянной скорости укорочения. Реальные физические
свойства последовательного упругого элемента не рассматрива-
лись, так как его длина всегда оставалась постоянной. Не изу-
чался и временной ход развития (или спада) силы. Более того,
при использовавшихся длинах мышцы напряжение покоя было
очень малым (приблизительно 2% от активного напряжения), и
поэтому модель из двух элементов была адекватной. Однако при
больших длинах мышцы необходимо учитывать напряжение покоя
(рис. 11.9), а это требует введения в модель соответствующего
компонента — параллельного упругого элемента, как показано на
рис. 11.11,15 и В. Влияние этого компонента сколько-нибудь де-
тально не анализировали. Его представляли простым упругим
элементом, не играющим никакой роли в активном укорочении, и
его природа, а также способ соединения с другими двумя элемен-
тами модели преднамеренно не рассматривались. Наконец, точная
природа и локализация сократительного элемента и последова-
тельного упругого элемента также оставались неопределенными;
подлинно упругим элементом мышцы могли бы быть такие струк-
туры, как сухожилия, но упругостью могли быть наделены и соб-
ственно сократительные структуры Тем не менее «двухэлемент-
ная» модель активной скелетной мышцы получила широкое рас-
Гл. 11 Сердце 23S
пространение, поскольку она объясняла результаты целого ряда
механических и тепловых экспериментов. Ввиду аналогичности
структуры саркомеров сердечной и скелетной мышц было есте-
ственным рассмотреть применимость такой модели также и к сер-
дечной мышце.
Проблемы возникли сразу же, как только попытались это сде-
лать. Первая из них связана с тем, что в препаратах сердечной
мышцы развивается заметное пассивное напряжение во всем диапа-
зоне длин, при которых они могут сокращаться. Поэтому парал-
лельный упругий элемент становится существенной частью модели,
и соотношение сила — скорость может быть исследовано лишь час-
тично, так как добиться, чтобы сила была равна (или хотя бы
близка) нулю, в этих условиях невозможно. Вторая, значительно
более важная проблема заключается в том, что в обычных усло-
виях сердечная мышца в отличие от скелетной тетаническое со-
кращение не развивает, и потому получить длительно поддержи-
вающееся и хорошо воспроизводимое изометрическое сокращение
сердечной мышцы невозможно. Сердечная мышца реполяризуется
относительно медленно, и повторные стимулы не вызывают тета-
нического сокращения. Напротив, возникают одиночные сокраще-
ния, которые даже при высокой частоте раздражения сливаются
лишь частично, так что изменение во времени напряжения или
укорочения имеет «зубчатый» вид. Таким образом, в ответ на каж-
дый стимул осуществляется неполный цикл сокращения и расслаб-
ления. При более низких частотах стимулы вызывают одиночные
сокращения, которые четко отделены друг от друга и могут очень
хорошо воспроизводиться, но ни один из этих типов ответов не
представляет собой установившееся состояние активности, так как
способность мышцы развивать напряжение и укорачиваться (ее
активное состояние) может во время сокращения непрерывно изме-
няться. Серия одиночных сокращений препарата сосочковой мыш-
цы показана на рис. 11.12.
Все это значительно усложняет постановку опытов и интерпре-
тацию результатов. Интенсивность активного состояния (каким бы
образом ее ни оценивали) является, очевидно, важным свойством
мышцы. Но это свойство становится крайне неопределенным, если
оно меняется на всем протяжении сокращения. Возникающие здесь
проблемы лучше всего пояснить на примерах. Хилл первоначально
определил активное состояние как усилие, которое сократительный
элемент может развивать, не изменяя свою длину, и которое по-
этому могло быть измерено, только когда скорость укорочения
сократительного элемента равна нулю. Даже для скелетной мыш-
цы это возможно лишь при условии, что развиваемое усилие до-
стигло поддерживающейся максимальной величины в состоянии
изометрического тетануса. При тетаническом изометрическом со-
кращении активированный сократительный элемент начинает уко-
рачиваться и растягивает последовательный упругий элемент. При
Я8в Чаоть II. Механика кровообращения
«том в упругом элементе возникает усилие, пропорциональное его
удлинению, что эквивалентно увеличению нагрузки, против кото-
рой укорачивается сократительный элемент. В результате скорость
укорочения последнего постепенно уменьшается до нуля
(рис. 11.10). Интенсивность активного состояния в этом случае
характеризуется величиной Ро [уравнение (11.1)]. С другой сто-
роны, очевидно, что когда развивается одиночное изометрическое
Рис. 11.12. Серия наложенных
друг на друга записей, показываю-
щая изменение длины сосочковой
мышцы кошки и развиваемого ею
усилия во время сокращения этой
мышцы против различных по ве-
личине нагрузок. Начальная длина
мышцы поддерживалась одинако-
вой; приведенное внизу семейство
кривых показывает, что в каждом
случае развиваемое мышцей уси-
лие сначала растет до значения,
равного нагрузке, а затем остается
постоянным, тогда как длина мыш-
цы в это время уменьшается. Сте-
пень укорочения при каждой на-
грузке показана на верхнем семей-
стве кривых. Последняя (верхняя)
кривая развиваемого мышцей уси-
лия показывает ответ мышцы в
условиях, когда она не может под-
нять нагрузку, т. е. при одиночном
изометрическом сокращении. [Son-
nenblick (1962). Am. J. Physiol.,
202, 931.]
сокращение, при котором интенсивность активного состояния на-
растает, а затем падает, то крайне сомнительно, что нагрузка пре-
кратит укорочение сократительного элемента как раз в то время,
когда интенсивность активного состояния максимальна. Но если
этого не происходит, максимальное регистрируемое напряжение
не соответствует величине Ро и оценка интенсивности активного
состояния оказывается заниженной. Соответствующие измерения
можно выполнить, создав какое-то специальное устройство, спо-
собное поддерживать длину сократительного элемента постоянной,
например путем контролируемого растяжения мышцы во время
сокращения, но возникающие здесь экспериментальные сложности
очень велики.
Другой подход (который в какой-то мере даже увеличил пута-
ницу, поскольку он ввел другое определение активного состояния)
основан на использовании в качестве меры активного состояния
скорости укорочения в отсутствие нагрузки. Эту величину опре-
делить проще: мышцу освобождают от нагрузки в различные мо-
менты повторяющихся циклов одиночного сокращения и каждый
Гл. 11. Сердца 237
раз, тотчас после скачкообразного перехода мышцы к своей длине
в ненагруженном состоянии, измеряют скорость укорочения. При-
мер полученных в таком опыте результатов приведен на рис. 11.13:
активное состояние достигает пика примерно через 150 мс после
раздражения (несколько раньше, чем достигается пиковое напря-
жение при изометрическом одиночном сокращении), а затем рав-
номерно спадает.
Рис. 11.13. Изменение во времени актив-
ного состояния сердечной мышцы, изме-
ренного методом освобождения ее от
нагрузки во время изотонических сокра-
щений. Стрелкой отмечен момент време-
ни, в который усилие, развиваемое мыш-
цей при одиночном изометрическом со-
кращении, достигает максимума. [Ed-
man, Nilsson (1968). Acta Physiol.
Scand., 72, 205.]
Недавно, однако, было показано, что методы, основанные на
определении скорости укорочения сократительного элемента, об-
ладают одним недостатком: обнаружено, что длительность актив-
ного состояния зависит от изменений длины сократительного эле-
мента. Если в какой-либо момент цикла сокращения он укорачи-
вается, то активное состояние спадает раньше. Поэтому в послед-
нее время применяется более изощренный метод: сначала для
последовательного упругого элемента мышцы определяют характе-
ристику напряжение — длина, а затем, используя управляемую
при помощи вычислительной машины механическую обратную
связь, растягивают мышцу во время сокращения так, чтобы она
непрерывно удлинялась именно на величину, необходимую для со-
хранения длины сократительного элемента постоянной1). В таком
случае в каждый момент цикла сокращения интенсивность актив-
ного состояния мышцы определяется развиваемым напряжением.
Временной ход и интенсивность активного состояния, определен-
ные этим методом, лишь немного отличаются от аналогичных ха-
рактеристик одиночного изометрического сокращения. Основным
отличием являются несколько более ранние повышение и сниже-
’) Разумеется, длина сократительного влемента не измеряется, а вычисляется
о принятой исследователем модели. — Прим. ред.
238 Часть И. Механика кровообращения
ние активного состояния. Однако даже эти результаты едва ли яв-
ляются окончательными, так как совсем недавно было показано,
что свойства последовательного упругого элемента зависят от вре-
мени, и потому степень растяжения, необходимая для фиксации
длины сократительного элемента в различные моменты цикла,
зависит не только от мгновенного значения напряжения.
Эти исследования, которые привели в конце концов к пара-
доксу, заключающемуся в том, что сократительную способность
препарата сосочковой мышцы лучше всего определять, заставляя
ее удлиняться, описаны здесь довольно подробно, поскольку они
хорошо освещают трудности, возникающие при эксперименталь-
ном изучении механики сердечной мышцы на этом уровне. Теперь
должно быть очевидно, что важными являются по крайней мере
четыре переменные — время, длина, напряжение и скорость, а це-
лый ряд внешних факторов, оказывающих влияние на сократи-
мость, таких, как температура, насыщение кислородом и ионный
состав окружающей среды, должен строго контролироваться. Весь-
ма ценный обзор по этому вопросу написан Блинксом и Джуел-
лом1). Авторы указывают, что, поскольку в одном опыте обычно
определяют соотношение только между двумя переменными, для
установления взаимосвязи между временем, длиной, напряжением
и скоростью необходимо провести эксперименты шести типов.
Мы не будем рассматривать эти вопросы детально, но некото-
рые общие выводы настолько важны, что их необходимо изложить.
Обратная зависимость между развиваемым усилием и скоростью
укорочения (рис. 11.10), характерная, как установлено, для ске-
летной мышцы, справедлива также и для сердечной мышцы (хотя
здесь эта зависимость обычно не является строго гиперболиче-
ской). Следует сразу отметить, что на зависимость сила — ско-
рость оказывают влияние длина мышцы и интенсивность ее ак-
тивного состояния. Влияние длины можно качественно предска-
зать исходя из зависимости длина — напряжение, изображенной
на рис. 11.9: с увеличением начальной длины кривая сила — ско-
рость смещается вверх и вправо. Поэтому, чтобы представить свой-
ства мышцы графически, необходимо использовать систему коор-
динат, по осям которой отложены сила, развиваемая сократитель-
ным элементом, скорость его укорочения (как на рис. 11.10) и
длина мышцы. При любом уровне активного состояния мышцы
совокупность различных возможных сочетаний этих переменных
образует в такой системе координат трехмерную поверхность.
Строить подобные графики так, чтобы они достаточно полно
отражали поведение реальных систем, довольно трудно, поскольку
на них необходимо указывать и положительные, и отрицательные
’) Blinks J. R., Jewell В. R. (1972) The meaning and measurement of myo-
cardial contractility, Chapter 7 in Cardiovascular fluid dynamics, ed. Bergel D. H,
Academic Press.
Гл. 11. Сердце 239
скорости (т. е. и укорочение, и удлинение), а если мы хотим рас-
смотреть полный цикл сокращения, то должны быть представлены
также и возрастающая, и спадающая ветви кривой длина — на-
пряжение. Чтобы понять, как связаны между собой различные
свойства мышцы, проще ограничиться анализом двух типов сокра-
щения— изометрического и изотонического.
Более простым является одиночное изометрическое сокраще-
ние, так как оно исключает изменение длины мышцы. Сокращение
начинается в некоторой точке кривой длина — напряжение, снятой
в состоянии покоя (рис. 11.9). Сначала сократительный элемент
укорачивается очень быстро, так как он нагружен сравнительно
слабо. Но, поскольку укорочение сократительного элемента сопро-
вождается удлинением последовательного упругого элемента, воз-
никает противодействующая укорочению сила, и скорость укоро-
чения сократительного элемента постепенно уменьшается, пока не
станет равной нулю. В этот момент развивается максимальное уси-
лие (равное сумме активного усилия и усилия покоя; см. рис. 11.9).
Таким образом, точка, соответствующая состоянию мышцы, сдви-
гается вдоль кривой сила — скорость, показанной на рис. 11.10, до
пересечения этой кривой с осью нагрузки (силы). Затем происхо-
дит расслабление, и мышца возвращается в исходное состояние,
которому соответствует начальная точка на кривой длина — сила
для состояния покоя.
Если мышца укорачивается против действующей во время со-
кращения нагрузки (изотоническое сокращение), то ситуация не-
сколько усложняется. Сначала сократительный элемент укорачи-
вается и развивает усилие, растягивая последовательный упругий
элемент, как в случае изометрического сокращения. Затем насту-
пает состояние, когда развитое усилие становится равным нагруз-
ке; после этого усилие остается постоянным, и вся мышца начи-
нает укорачиваться при постоянной длине последовательного уп-
ругого элемента. При уменьшении длины мышцы усилие может
оставаться постоянным только за счет уменьшения скорости уко-
рочения сократительного элемента (рис. 11.14). Поэтому мышца
укорачивается все более медленно, пока не начнет расслабляться,
и затем вновь возвращается в исходное состояние, соответствую-
щее начальной точке на кривой длина — напряжение в состоянии
покоя.
Во время сокращения целого сердца реализуются как изомет-
рическая, так и изотоническая фазы, и, как будет показано ниже,
нагрузка (т. е. давление в желудочках) во время изотонической
фазы непрерывно изменяется. Совершенно очевидно, что детально
описать такое сокращение чрезвычайно сложно. Более того, мы
до сих пор не принимали во внимание основной фактор, услож-
няющий все описанные выше соотношения, — время. Интенсив-
ность активного состояния в течение всего цикла сокращения не
остается постоянной, но растет и уменьшается. Поэтому кривая
240 Часть II Механика кровообращения
Рис 11.14 Кривые, иллюстрирующие зависимость соотношения сила — скорость
для сердечной мышцы от ее длины. Каждая кривая построена по данным, полу-
ченным при сокращении мышцы против различных по величине нагрузок
(см. рис. 11.12), Последовательное увеличение начальной длины мышцы сопро-
вождается все большим смещением кривых вправо. [Sonnenblick (1962). Am. J.
Physiol., 202, 931.)
сила — скорость, аналогичная приведенной на рис. 11.10, отражает
свойства сердечной мышцы только в какой-то конкретный момент
времени. Эта кривая после стимуляции мышцы смещается вверх
от исходной и возвращается к ней, когда активное состояние спа-
дает. Все графики, связывающие силу, скорость и длину, во время
сокращения и расслабления мышцы сдвигаются относительно ко-
ординатных осей.
Такой способ описания представляет определенную ценность
как для иллюстрации существующих сложностей, так и для выяв-
ления того факта, что, как бы мы ни рассматривали сокращение
(принимая за его главный показатель скорость или силу), суще-
ствуют два главных определяющих его фактора. Первый — это
длина мышечного элемента, второй — интенсивность активного со-
стояния. На последнем из них стоит остановиться особо: интуи-
тивно ясно, что большая интенсивность активного состояния при
прочих равных условиях будет давать более сильное сокращение
независимо от того, как его измеряют—исходя из силы или из
скорости. Значительно труднее найти способ отображения или
измерения уровня активного состояния: нередко оказывается, что
некое свойство мышцы — обычно называемое «сократимостью» —
Гл. II. Сердце 241
не поддается измерению и не имеет четкого определения1). Ряд
спорных вопросов остается даже тогда, когда мы имеем дело с
изолированным мышечным препаратом. Эти вопросы связаны глав-
ным образом с формой «краев» трехмерной поверхности, которая
графически отображает рассмотренные выше взаимосвязи между
силой, скоростью и длиной. Указанную форму не всегда удается
получить прямо из данных эксперимента, и нередко ее устанавли-
вают экстраполяцией. Такая экстраполяция должна основываться
на модели мышцы (изображенной, например, на рис. 11.11, Б или
В или несколько более сложной), и результаты могут очень сильно
зависеть от того, какая именно модель выбрана.
Для иллюстрации рассмотрим один пример, имеющий важное
практическое значение. Он касается максимальной скорости уко-
рочения, которую может достичь ненагруженный сократительный
элемент. Для сердечной мышцы эту скорость (обозначаемую через
Vmax) измерить невозможно, поскольку, как видно из рис. 11.9, при
любых длинах в покоящейся сердечной мышце существует конеч-
ное напряжение, и потому, чтобы задать начальную длину, необхо-
димо наличие некой нагрузки (называемой предварительной на-
арузкой). Таким образом, Утах можно определить только экстрапо-
ляцией. Несколько лет назад американский физиолог Зонненблик
произвел такую экстраполяцию кривых сила — скорость, получен-
ных при разных длинах мышцы. Он пришел к выводу, что все
кривые (показанные на рис. 11.14) можно экстраполировать к об-
щей точке их пересечения с осью скоростей.
Поэтому казалось, ЧТО Утах не зависит от длины мышцы. Это
Предположение вызвало большой интерес, так как оно означало, что
Уmax можно использовать для различения влияний, оказываемых на
сокращение мышцы изменением ее длины и интенсивности актив-
ного состояния (т. е. сократимости). Совершенно очевидно, на-
сколько полезным критерием была бы такая величина для опытов
на изолированной мышце, и тем более — при исследовании дея-
тельности целой сердечной камеры, когда гораздо труднее поддер-
живать длину ее мышечных волокон и их сократимость на по-
стоянном уровне.
Однако для проведения экстраполяции, дающей в результате
Утах, нужно знать форму экстраполируемых кривых сила — ско-
рость, а именно при определении формы становится существенным
то, какая модель мышцы используется. В данном случае модель,
предложенная А. Хиллом (рис. 11.11.А), неприменима, так как
в сердце существует напряжение покоя. Поэтому до экстраполя-
ции необходимо скорректировать результаты измерений так, чтобы
*) Разумеется, «сократимость» можно определить (и определяют) как спо-
собность мышцы укорачиваться и/или развивать силу при стимуляции Однако
суть в том, что такое определена не указывает меры для этой способности. —
Прим- ред.
242 Часть II. Механика кровообращения
учесть влияние параллельного упругого элемента. Более подробно
этот вопрос изложен в работе Поллака ); здесь же достаточно за-
метить, что необходимая коррекция зависит от вида выбранной
модели (на рис. 11.11, Б и В представлены наиболее простые из
них) и влияет на получаемое значение Утах. Это означает, что ис-
ходная гипотеза о независимости Утах от начальной длины, кото-
рая основывалась на экстраполяции кривых рис. 11.14 к общей на-
чальной точке на оси скоростей, может быть верна только как ча-
стный случай. В результате соотношение длина — скорость при ну-
левой нагрузке снова стало предметом дискуссий.
В течение примерно десяти лет, когда при исследовании сердца
использовали эти простые механические модели, весьма детально
была изучена ультраструктура скелетных мышц, а также созданы
изящные методы измерений на уровне саркомеров. Соответственно
механические свойства скелетной мышцы интерпретируют теперь
на основе известной структуры сократительного аппарата. Таким
образом, моделью стал сам саркомер. Применять такой подход
при исследовании сердечной мышцы мешает отсутствие подходя-
щих препаратов, и механические модели все еще являются един-
ственной полезной аналогией реальной сердечной мышцы.
Заключение. Прежде чем приступить к анализу поведения целой
сердечной камеры, полезно подвести итоги некоторых исследова-
ний изолированных мышечных препаратов. Такие мышцы содер-
жат большое число мышечных клеток, которые, вероятно, все ле-
жат достаточно параллельно продольной оси мышцы. В покое и
во всем диапазоне длин, который можно считать нормальным для
их работы, эти мышцы сопротивляются растяжению с силой, резко
возрастающей при увеличении их длины. Ответственный за это
свойство структурный элемент пока не идентифицирован. В про-
стых механических моделях его называют параллельным упругим
элементом и представляют как пружину (хотя пружина плохо от-
ражает действительное положение вещей, так как мышца не на-
чинает колебаться, когда ее освобождают, и заметно «ползет»,
когда ее растягивают, что указывает на наличие как упругих, так
и вязких свойств). Мышечные клетки сердца можно деполяризо-
вать толчком электрического тока, и эта деполяризация приводит
через несколько миллисекунд к развитию механической актив-
ности, во время которой мышца укорачивается. Если, нагрузив
мышцу, создать сопротивление ее укорочению, она разовьет неко-
торое усилие, и, когда это усилие сравняется с нагрузкой, мышца
начнет укорачиваться, причем скорость укорочения будет обратно
пропорциональна нагрузке. Своей способностью развивать усилие
и укорачиваться мышца несомненно обязана структурам сарко-
мера, длина которого в широком диапазоне строго пропорцио-
') Pollack G Н (1970). Maximum velocity as an index of contractility in
cardiac muscle, Circulation Research, 26, 111—127.
Гл 11. Сердце 243
нальна длине мышцы. Однако, поскольку детали механизма уко-
рочения не выяснены, здесь опять прибегают к простой механиче-
ской модели, содержащей сократительный элемент, который при
стимуляции укорачивается, а с нагрузкой (когда она существует)
соединен через последовательный упругий элемент. При увеличе-
нии длины мышцы в пределах физиологически нормального диапа-
зона развиваемые мышцей усилие и скорость укорочения также
увеличиваются. Поэтому для описания сокращений, начинающихся
при различных длинах мышцы, необходимо располагать семейст-
вом кривых сила — скорость. Растяжение мышцы сверх опреде-
ленной длины, вероятно соответствующей некоторому оптималь-
ному расположению в саркомере белковых нитей, приводит к сни-
жению развиваемого при сокращении усилия. Но эта длина, по-
видимому, лежит вне нормального диапазона длин для целого
сердца. Максимальное укорочение отдельного саркомера, оче-
видно, не превышает примерно 30%: в состоянии покоя его длина
составляет около 2,2 мкм, а при максимальном сокращении —
1,5 мкм. Возможно, однако, что эти данные не совсем точны из-за
усадки ткани при гистологической обработке. Скорость и степень
изменения сократимости мышцы ограничены. На практике пре-
делы их изменения, по-видимому, достаточно хорошо выявляют-
ся при одиночных изометрических сокращениях. Эти величины
чувствительны к ряду внешних воздействий. Факторы, оказываю-
щие влияние на временные параметры сокращений сердца (т. е
на ча'стоту его сокращений), называют хронотропными, а факторы,
изменяющие интенсивность сокращений, — инотропными. К ино-
тропным факторам относятся определенные химические агенты —
ряд гормонов, лекарственных препаратов и ионов, которые могут
увеличивать или уменьшать сократимость и, по-видимому, имеют
большое значение для регуляции и коррекции работы целого
сердца.
11.4. Механическое поведение целого сердца
До сих пор мы рассматривали свойства сердечной мышцы как
таковой. Все это, однако, не представляет большой практической
ценности для анализа механических свойств сердечных камер или
их действия как насоса, если необходимые физические параметры
не могут быть переопределены в соответствующей форме. Напри-
мер, нагрузкой, против которой сокращается мышца, теперь яв-
ляется сила, действующая на стенку желудочка и возникающая
благодаря наличию в нем крови. Однако эта сила распределена
по некоторой площади, изменяющейся вместе с объемом желудоч-
ка, и, чтобы учесть это изменение при любом анализе баланса снл,
следует отнести силу к площади, т. е. перейти к давлению. Натя-
жение стенки желудочка, которое противодействует этому давле-
нию, тоже следует пересчитывать в силу, действующую на еди
244 Часть II. Механика кровообращения
ницу площади, т. е. в напряжение. Подобным же образом из-за
сложной геометрии и изменений формы, которые претерпевает
желудочек за время сердечного цикла, никакая отдельно взятая
длина не будет характеризовать состояние всех мышечных воло-
кон. Наиболее простой характеристикой размера, пригодной для
данного случая, является, по-видимому, объем желудочка, но для
определения такого показателя или для более сложного описания
распределения длин мышечных волокон важно учитывать форму
желудочка. Скорость укорочения волокон отражается в скорости
Рис. 11.15. Схематическое представление ста-
тического равновесия сил, действующих на ма-
лый элемент стенки желудочка. Р — давление
в желудочке, Si и S2 — нормальные напряже-
ния в стенке.
изменения объема желудочка, которую обычно проще всего изме-
рять по скорости поступления крови в аорту или в легочную арте-
рию.
Чтобы представить себе связь между напряжением в стенке
желудочка и давлением в его полости в статических условиях,
рассмотрим силы, действующие на мысленно выделенный малый
участок стенки желудочка (рис. 11.15). С внутренней стороны
стенка нагружена давлением Р- оно уравновешивается противо-
действующими силами в стенке, наиболее существенными компо-
нентами которых являются напряжения Si и 5г, действующие па-
раллельно поверхности стенки1). Поскольку эти напряжения дей-
ствуют перпендикулярно (нормально) к боковым поверхностям
элемента, их называют нормальными. Могут существовать также
силы, параллельные указанным поверхностям, т. е. касательные
(тангенциальные) напряжения.
Связь между давлением в полости и напряжением в стенке
оказывается довольно сложной. Отчасти это обусловлено слож-
ной геометрией системы, но дело не только в ней. Мы в этом убе-
димся, рассмотрев простейшую из возможных форм желудочка —
сферу.
Если мы хотим использовать элементарную теорию упругости,
то сначала должны определить напряжения в стенке так, чтобы
учесть возможные изменения (градиенты) их от внутренней по-
верхности стенки к наружной. Такие изменения напряжения за-
ведомо существуют, но, поскольку значения их неизвестны, мы
*) См. также гл. 7, где аналогичный баланс сил описан для кровеносных
сосудов.
Гл. 11. Сердце 241
-Ц. "П ' • --
должны, как и в Гл. 7, определить напряжения как величины,
средние по толщине стенки. Таким образом, нормальные напря-
жения (Si и S2 на рис. 11.15) являются средними по всей толщине
стенки. Далее мы должны предположить, что стенка однородна
и изотропна. Тогда для стенки сферической формы Si = S2.
Теперь можно, как в гл. 7, применить закон Лапласа [см. урав-
нение (7.6)], но при этом принять во внимание, что мы имеем
дело с двумя взаимно перпендикулярными нормальными напря-
Рис. 11 16 Напряжения в стен
ке левого желудочка, представ-
ленной в виде эллипсоида (см.
текст).
жениями, а не с единственным окружным напряжением, характе-
ризующим натяжение трубки по окружности. Поэтому вместо
уравнения (7.6) нужно положить
, 2S'h
Р = ~'
где р' — разность давления по разные стороны стенки, гв — внут-
ренний радиус, h — толщина стенки, S' равно Si и S2. Если ма-
териал стенки не является однородным и изотропным, то, даже
если она будет оставаться сферической, Si уже не будет равно
Тогда
/ (Si + S2) h
п -----------.
гв
Простейшим после сферы телом вращения является эллипсоид
вращения. Такой случай потенциально значительно сложнее, так
как теперь нужно учитывать и тангенциальные силы. Но если мы
выберем элемент стенки, ориентированный по главным осям эл-
липсоида (рис. 11.16), то тангенциальных сил на гранях такого
элемента не будет, и мы опять сможем написать уравнение, свя-
зывающее трансмуральное давление с напряжением в стенке:
246 Часть II. Механика кровообращения
St и Хг, как и ранее, средние нормальные напряжения, но га и
гъ уже не просто радиусы: первая из этих величин является ра-
диусом кривизны поверхности в точке Р в продольной (меридиа-
нальной) плоскости. При движении точки Р по меридиану эллип-
соида этот радиус будет, очевидно, непрерывно изменяться, стано-
вясь очень малым на вершинах эллипсоида и наибольшим на его
экваторе. Вторая величина, гъ, — это длина перпендикуляра, опу-
щенного на ось эллипсоида из точки Р. При движении последней
по меридиану он также будет непрерывно изменяться и станет
истинным радиусом (равным меньшей полуоси), только когда точ-
ка Р окажется на экваторе эллипсоида. Когда точка расположена
на вершине, длина перпендикуляра равна нулю.
Форма левого желудочка и напряжение в его стенке. Из всего
сказанного ясно, что если форма тела не является сферической,
то в рамках даже самой упрощенной теории упругости связь ме-
жду трансмуральным давлением и напряжениями в стенке будет
довольно сложной. Поэтому едва ли стоит удивляться, что во мно-
гих исследованиях форму левого желудочка считали сферической,
причем в самых первых работах были приняты тонкостенные сфе-
рические модели. Однако они совершенно неадекватны, в частно-
сти потому, что даже в диастолу толщина стенки левого желу-
дочка составляет 20—30% его наружного радиуса, а во время
систолы она на 10—12% больше, чем во время диастолы. Сле-
дующая модель, которую следует рассмотреть, — толстостенная
сфера; эта модель вполне приемлема для оценки изменений длины
мышечных волокон. Чтобы объем крови, заключенной в такой
сфере, за время систолы уменьшился вдвое (что в случае желу-
дочка близко к истине), волокна самого внутреннего слоя стенки
должны укоротиться примерно на 20% • Эта величина согласуется
с тем, что мы знаем о нормальном диапазоне длин саркомера.
Естественно, при сокращении желудочка толщина стенки будет
увеличиваться, так как мышца несжимаема, и уменьшение ра-
диуса наружной стенки не достигнет 20%. Поэтому укорочение от-
дельных волокон будет непрерывно меняться по толщине стенки,
достигая максимума на внутренней поверхности. Это конечно,
свойственно всем толстостенным структурам, как и изменение на-
пряжения по толщине стенки, о котором упоминалось ранее.
Однако, несмотря на простоту сферической модели и разум-
ность получаемых исходя из нее изменений длины волокон, она
слишком далека от действительности — форма желудочка сердца
гораздо сложнее. Сложность формы левого желудочка человека
и животных выявлена многими методами; наиболее наглядно это
показывают кинофильмы, снятые с экрана рентгеновского аппа-
рата. Для получения изображения сердца в него через катетер,
конец которого находится в полости желудочка, вводят водный
раствор рентгеноконтрастного вещества. (Этот метод, названный
киноангиографией, широко используют как в клиниках, так и в
Гл. 11. Сердце 247
Рис. 11.17. Очертания полости левого
желудочка человека (воспроизведен-
ные по киноангнограммам) через
равные промежутки времени в пе-
риод от конца диастолы до конца си-
столы. Выход нз желудочка в аорту
находится вверху слева, а верхушка
желудочка — внизу справа. [Gibson,
Brown (1976), личное сообщение.]
Рис. 11.18. Схематическое изображе-
ние изменений наружных размеров ле-
вого желудочка собаки н перемеще-
ний его стенок за время от конца
диастолы до конца систолы. Стрел-
ками отмечены направление движе-
ния стенок и расстояние, на которое
они перемещаются. [Hinds et al.
(1969). Instantaneous changes in the
left ventricular lengths in dogs during
the cardiac cycle. Fedn Proc. Fedn
Am. Socs Biol., 28, 1351.]
исследовательских лабораториях для изучения структуры и функ-
ционирования сердца и кровеносных сосудов.) Получаемое таким
путем изображение левого желудочка (рис. 11.17) похоже по
форме на притупленный наконечник стрелы, причем верхушка же-
лудочка образует острие наконечника, а два клапана — его осно-
вание. Внутренняя стенка желудочка из-за проходящих здесь про-
дольных мышечных тяжей (трабекул) различной толщины имеет
вид неупорядоченно гофрированной поверхности, а сосочковые
мышцы выступают в полость и делают ее форму еще более слож-
ной. В поперечном сечении желудочек обычно выглядит приблизи-
тельно круглым, и наибольший поперечный диаметр его при нор-
мальных диастолических объемах составляет примерно половину
длины продольной (большой) оси.
При сокращении желудочка его продольная ось сначала не-
сколько укорачивается, а поперечный диаметр соответственно уве-
личивается. Это происходит до начала фазы изгнания крови и,
вероятно, вызвано сокращением продольно-ориентированных мы-
шечных волокон наиболее глубоких слоев стенки сердца. Дей-
ствительно, электрофизиологические исследования показали, что
деполяризация мышцы желудочка начинается у внутренне!)
Я48 Часть II. Механика кровообращения
стенки. В начале фазы изгнания крови основным изменением яв-
ляется укорочение поперечных осей, которые уменьшаются при-
близительно на одну треть. Но это укорочение несимметрично —
оно максимально вдоль передне-задней оси. Продольная ось уко-
рачивается очень мало. К концу систолы отношение продольной
(большой) оси к поперечной (малой) составляет примерно 2,5:1.
Вся последовательность изменений, совершающихся во время со-
кращения, представлена на рис. 11.18. Приведенная на нем диа-
грамма построена по данным исследований, в которых при по-
мощи рентгеновских лучей прослеживали движение непроницае-
мых для этих лучей маркеров, прикрепленных к стенке желудочка
в различных ее точках.
Из этих описаний ясно, что действительная форма желудочка
настолько далека от сферической, что считать его сферой нель-
зя— и не только потому, что желудочек имеет продольную и по-
перечные оси, но и вследствие того, что первая почти не меняется
за время сердечного цикла. В последнее время значительно более
успешно используют эллипсоидальные модели, причем предпола-
гают, что длина продольной оси эллипсоида постоянна, а измене-
ние объема происходит из-за укорочения поперечных осей. Это
вполне удовлетворительно согласуется с рассмотренными только
что результатами наблюдений за реальным желудочком. Требуе-
мая такой моделью степень укорочения мышечных волокон сов-
местима также с известными свойствами саркомера: для того что-
бы содержащийся в эллипсоиде вращения объем крови умень-
шился вдвое, а длина его продольной оси при этом не изменилась,
поперечные оси, а следовательно, и идущие по окружности во-
локна должны укоротиться (по внутренней стенке) самое большее
на 29%. Наконец, предсказываемые этой моделью значения на-
пряжений в стенке желудочка ближе к величинам, полученным
при прямом измерении тензодатчиками, чем значения, следующие
из сферической модели.
Обратим, однако, внимание еще на одно обстоятельство: в рас-
смотренных модельных исследованиях неизбежно предполагают,
что свойства стенки по всей ее толщине одинаковы, а следова-
тельно, пренебрегают действительным строением стенок желудоч-
ка (см. рис. 11.4), равно как и тем, что активное усилие, разви-
ваемое по-разному растянутыми мышечными волокнами, может
различаться. Правда, эти факторы были учтены в детальном ис-
следовании Стритера и др. *).
В этой работе, как и в предыдущих, желудочек считали эллип-
соидом, а кроме того, предполагали, что мышечные волокна мо-
гут сопротивляться только продольному растяжению. Поскольку
’) Streeter D. D. Jr.. Vaishnav R. N, Patel D. J., Spotnitz H. M., Ross J. Jr.,
SonnenbHck E. H. (1070). Stress distribution on the canine left ventricle during
diastole and systole, Biophysical Journal, 10, 84’5—603, 4
Гл. 11. Сердце М9
мышечные волокна, проходя в изогнутой стенке левого желу-
дочка, непрерывно меняют направление (рис. 11.4), изгиб их чрез-
вычайно сложен. Поэтому ориентацию волокон и главные радиусы
кривизны стенок желудочка определяли на трупах собак и исходя
из этих данных вычисляли кривизну волокон. Полученные резуль-
таты использовали затем для расчета распределения по толщине
стенки нормальных и радиальных напряжений1). Стенку рассмат-
ривали при этом как набор вложенных друг в друга связанных
эллипсоидальных оболочек, в каждой из которых ориентация во-
локон постоянна и соответствует ориентации, определенной для
данного слоя стенки экспериментально. В средней части стенки
кривизна волокон оказалась наибольшей в конце систолы, а
ближе к эндокарду — во время диастолы. Если считать натяже-
ние волокон всюду одинаковым, то распределение по толщине
стенки радиального напряжения (или, что то же самое, давле-
ния, оказываемого одним слоем на другой) получается не совсем
линейным: наиболее быстро оно спадает во внутренних слоях стен-
ки. Если же натяжение волокон во время систолы не считать оди-
наковым (а мы знаем, например, из рис. 11.9, что активное натя-
жение волокон зависит от их длины и, кроме того, что длина сар-
комеров во время систолы меняется по толщине стенки желу-
дочка), то предсказываемое распределение радиального напряже-
ния поперек стенки будет приблизительно линейным. Рассчитан-
ные для систолы и для диастолы распределения нормальных на-
пряжений, направленных по двум главным осям, изображены на
рис. 11.19. Здесь, как и раньше, предполагается, что натяжение
волокон меняется по толщине стенки во время систолы, но сохра-
няется постоянным во время диастолы. Громадное значение учета
ориентации волокон в таких исследованиях очевидно: в моделях,
предполагающих однородность свойств стенки, как продольное
(Si), так и направленное по окружности (5г) напряжения оказы-
ваются самыми высокими на поверхности эндокарда, что, как
видно из рис. 11.19, неверно.
Проведенное в этом же исследовании сопоставление предска-
занных значений направленных по окружности сил с величинами,
измеренными непосредственно при помощи датчиков силы in vivo,
показало хорошее соответствие теории и эксперимента. Таким об-
разом, даже в предположении эллипсоидальной формы такие мо-
дели левого желудочка несомненно близки к действительности й
*) Радиальные напряжения также принадлежат к категории нормальных
(а не тангенциальных) и действуют перпендикулярно площадкам, мысленно вы-
деленным в толще миокарда «параллельно» ею поверхностям. Термин «нормаль-
ные» авторы выделяют здесь для напряжений, ориентированных так, как это по-
казано на рис. 11.15. Важно, однако, что в отличие от предыдущего речь идет
ие о средних по толщине стенки напряжениях, а об истинных распределениях
напряжений. Поэтому Si, S2 на рис. 11.19 и в формулах, приведенных выше,
Имеют разный физический смысл.— Прим. ред.
2б0 Часть II. Механика кровбобращения
Рис. 11.19. Отношения напряжений, развивающихся в стенке левого желудочка
в конце диастолы и в конце систолы. Оси абсцисс, положение элемента стенки,
для которого указывается отношение напряжений, между ее эндокардиальной (0)
и эпикардиальной (100) поверхностями. Оси ординат: напряжения, отнесенные
к среднему по всей толщине стенки напряжению. Si — продольное напряжение,
Sz—окружное напряжение (см. рис. 11.16). [Streeter, Vaishnav, Patel, Spotnitz,
Ross, Sonnenblick (1970). Stress distribution in the canine left ventricle during
diastole and systole, Biophys. J., 10, 357.]
весьма полезны, хотя их можно усовершенствовать с учетом более
подробных данных о геометрии волокон левого желудочка ') и ре-
зультатов более тщательных измерений соотношений длина —
'сила для саркомера.
Вероятно, следует подчеркнуть, что для построения адекват-
ной модели распределений напряжения в стенке левого желудоч-
ка имеется очень важный практический стимул: кровь, питающую
сердечную мышцу, доставляют к ней коронарные сосуды, которые
проходят внутри стенки желудочка и поэтому подвергаются воз-
действию развивающихся в ней местных сил. По методическим
причинам трансмуральное давление в сосудах, находящихся внутри
стенки, и кровоток в них измерять трудно, и наши знания о рас-
пределении коронарного кровотока остаются весьма неполными.
Большая часть крови попадает из аорты в русло коронарных арте-
рий, несомненно, во время диастолы (рис. 11.20). Это естественно,
потому что в это время давление в желудочке, а следовательно, и
в его стенке (интрамуральное давление) значительно ниже давле-
') Streeter D. D., Jr., Powers W. E., Ross A. M., Torrent-Guasp F. (1976)
Thre-edimensional fibre-orientation in the mammalian left ventricular wall. In
Cardiovascular System Dynamics, eds. J. Baan, A. Nordergraaf, J. Raines, M. 1.1
Press, Cambridge, Mass.
Гл. 11. Сердце 261
Систола Диастола
Давление в аорте
105 мм рт. ст.
(среднее давление)
I 225 мп.Мин-1
Кровоток в левой \ Л \ А 90 мЛ-мии -Ina 100 г
коронарной артерииХ \ \ \ 2.8 мл на один удар
у VJ у Vсе₽вца
Кровоток в аорте
3000 мл - мин-1
37 мл на один удар
сердца
Частота сердцебиений 80 мин-1
Рис. 11.20. Записи давления и кровотока в аорте и кровотока в левой коронарной
артерии. Измерения произведены иа бодрствующей собаке через 14 сут после
(живления датчиков кровотока. Обратите внимание, что во время диастолы через
коронарную артерию протекает значительно больший объем крови, чем во время
систолы. [Gregg, Fisher (1962). Handbook of physiology, Section 2, p. 1534.]
ния в аорте. В результате распределение кровотока по сердечной
мышце во время диастолы оказывается равномерным. В систолу
картина более сложна, так как давление во внутренних слоях
стенки желудочка должно быть близким к давлению в его по-
лости. И поскольку в коронарных сосудах должно происходить
некоторое падение давления, обусловленное вязкостью крови,
трудно представить себе, каким образом может поддерживаться
кровоснабжение этих слоев. Действительно, измерение давления
в этой внутренней (субэндокардиальной) области стенки, произво-
дившееся многими исследователями, показало, что во время си-
столы интрамуральное давление иногда превышает давление в
желудочке. Это возможно, если ориентация волокон неодно-
родна. Такая неоднородность может возникнуть естественным об-
разом или как артефакт в эксперименте, если, например, изгиб
какой-то группы волокон окажется противоположным изгибу ос-
новной части стенки желудочка. Если учесть это условие, то к та-
ким выводам можно прийти исходя из математической модели.
Форма правого желудочка. Характер сокращения правого же-
лудочка значительно менее ясен, так как этому вопросу было по-
священо очень мало исследований. Но его насосное действие от-
личается от насосного действия левого желудочка. Одна стенка
правого желудочка — перегородка — функционально является
частью стенки левого желудочка. Другая, свободная стенка, зна-
252 Часть fl. Механика кровообращения
чительно тоньше и имеет большую площадь, так что правый желу-
дочек как бы пришит к одной стороне левого, подобно карману,
который сверху открывается в легочный ствол и в правое пред-
сердие. Поэтому площадь стенок, приходящаяся на единицу объ-
ема камеры, велика, и относительно слабое перемещение их по
направлению друг к другу вытесняет большой объем.
Такое действие, подобное действию кузнечных мехов, несом-
ненно важно для опорожнения правого желудочка. Однако оно мо-
жет быть усилено двумя другими механизмами. Существование
первого убедительно показано рентгенологически. Во время си-
Начало диастолы
Конец диастолы
Рис. 11.21. Принцип действия клапан-
ного насоса, работающего без изме-
нения объема камеры Насосное дей-
ствие осуществляется за счет измене-
ния положения камеры и согласован-
ной с этим изменением работы
клапана. Опорожнение выходного
тракта правого желудочка собаки мо-
жет частично обеспечиваться этвм
механизмом [Carlsson (1969). Expe-
rimental studies of ventricular mecha-
nics in dogs using the tantalum-labe-
led heart. Fedn Proc. Fedn Am. Socs
Biol, 28, 1324.]
столы свободная стенка движется вниз, т. е. как бы тангенциаль-
но по отношению к содержащейся в камере крови. Во время диа-
столы она снова движется вверх. Амплитуда этого движения
увеличивается по мере удаления от верхушки желудочка. Таким
образом, свободная стенка ведет себя подобно упругой мембране,
прикрепленной к верхушке сердца и продольно растягивающейся
во время диастолы. Когда она сокращается, то легочный клапан
смещается вниз, надвигаясь на заключенную в камере кровь, так
что кровь переходит в выходной тракт по другую сторону легочного
клапана. Принцип действия такого насоса, который может рабо-
тать без изменения объема камеры, показан на рис 11.21.
Другой механизм, который может способствовать изгнанию
крови из правого желудочка, — само сокращение левого желу-
дочка, которое натягивает свободную стенку правого желудочка
по мере того, как уменьшается радиус кривизны стенки левого же-
лудочка. О значении этого механизма можно лишь догадываться,
так как он еще не выявлен рентгенологически. Это неудивительно,
потому что относительное перемещение, которое такой механизм
может вызвать в какой бы то ни было плоскости, невелико и
должно осуществляться очень быстро и притом в то время, когда
все сердце находится в движении. Тем не менее как в опытах иа
животных, так и при клинических наблюдениях четко доказано,
Гл 11 Сердце 253
что при резко выраженном повреждении мышечных волокон стен-
ки правого желудочка он может продолжать действовать удиви-
тельно хорошо
Механика желудочка в целом. Если форма желудочка извест-
на, можно приступить к вычислению для него тех же величин,
которыми характеризуют поведение изолированной сердечной
мышцы. При таком способе изучения деятельности сердца желу-
дочек рассматривают как специализированную мышцу, геометрия
и механические свойства которой являются определяющими фак-
торами сокращения. Поскольку логически этот подход развивает
сказанное в предыдущих разделах, сначала мы займемся именно
им. Но есть и второй подход, к которому мы обратимся позднее
(хотя исторически он предшествовал первому). Суть его заклю-
чается в том, что желудочек рассматривают как насос, игнорируя
детали механики сокращения и исследуя в опытах соотношение
между наполнением и опорожнением (входом и выходом), а так-
же факторы, изменяющие это соотношение.
Отправной точкой второго подхода явился характер экспери-
ментально установленного соотношения «вход — выход». Оно из-
вестно как закон Старлинга для сердца и до сих пор остается
чрезвычайно важным для понимания многих аспектов деятельно-
сти сердца. Этот подход в значительной степени дополняет под-
ход, основанный на рассмотрении механики мышцы, и в последние
годы наблюдается тенденция объединить оба подхода и описать
насосное действие желудочка исходя из мгновенных соотношений
между нагрузкой и свойствами мышцы. Создание такого «синте-
тического» способа описания деятельности желудочка сопряжено
с большими трудностями и далеко от завершения.
Характеристика работы желудочка как мышцы. Как мы уже
видели, есть веские основания полагать, что форма и движение
левого желудочка геометрически значительно проще, чем пра-
вого, и большая часть изложенного ниже материала относится к
левому желудочку (хотя закон Старлинга несомненно справедлив
также и для правого желудочка). Если мы примем, что форма ле-
вого желудочка может быть с достаточной степенью точности
представлена эллипсоидом, малые оси которого во время систрлы
укорачиваются, а длина большой оси остается постоянной, то смо-
жем приступить к расчетам напряжения и скорости укорочения
мышечных элементов стенки. Среднее окружное напряжение в пло-
скости малых осей может быть получено из давления в желудоч-
ке, если известна кривизна стенки и ее толщина. Скорость укоро-
чения кольцевых элементов стенки в этой плоскости можно рас-
считать по скорости изгнания крови в аорту, если известна форма
эллипсоида. Наконец, мгновенные значения периметра эллипсоида
в плоскости его малых осей можно рассчитать, если известны ско-
рость изгнания крови и начальный объем желудочка (конечно-
диастолический объем).
254 Часть II. Механика кровообращения
Определить все эти величины экспериментально чрезвычайно
трудно, и некоторые из них удается получить только косвенно.
Например, измерить точно конечно-диастолический объем желу-
дочка в настоящее время невозможно. Его приходится рассчиты-
вать по результатам измерений линейных размеров, произведен-
ных тем или другим методом прижизненно — рентгенологическими
методами, при помощи ультразвука или закрепленных вокруг же-
лудочка датчиков изменения длины либо (с дополнительными до-
пущениями) посмертно. Положение осложняется еще и тем, что
сокращения сердца не являются ни изометрическими, ни изотони-
ческими: чтобы рассматривать изоволюмическую фазу как изомет-
рическую, мы должны предположить, что изменения формы сердца
несущественны, а чтобы фазу изгнания можно было рассматри-
вать как изотоническую, рабочую нагрузку на всем протяжении
этой фазы надо считать постоянной. Первое предположение, ве-
роятно, не слишком далеко от действительного положения вещей,
но второе несомненно неправдоподобно, так как абсолютно оче-
видно, что во время систолы давление в аорте не является по-
стоянным— см. рис. 11.5. Напомним и о более общих предполо-
жениях, состоящих в том, что во время эксперимента ионный и
химический состав внутренней среды сердца остается постоянным,
что активное состояние каждого мышечного волокна возникает
мгновенно и в течение систолы не меняется и что все мышечные
волокна активируются синхронно.
Если принять во внимание все отмеченные сложности, едва ли
покажется странным, что попытки изучать эту проблему на рабо-
тающем целом сердце почти не предпринимались. Чаще для ис-
следования тех или иных сторон сократительного поведения же-
лудочка или проверки конкретных предположений создавали осо-
бые условия (например, поддерживали постоянным конечно-диа-
столический объем или давление в аорте или, предотвращая изгна-
ние крови из желудочка, превращали сокращение в изометриче-
ское). Поэтому наши знания о целом желудочке, так же как и об
изолированной сердечной мышце, получены на основе результатов
многих разнородных экспериментов и являются в основном каче-
ственными. Литература по этим вопросам, естественно, столь же
обширна и сложна, как и литература, рассматривающая свой-
ства изолированных мышечных волокон.
По ряду причин трудно установить, действительно ли мышеч-
ное усилие связано со скоростью сокращения гиперболической
зависимостью. Во-первых, как мы видели выше, расчеты напря-
жения в мышце становятся чрезвычайно сложными, если учиты-
вать сложную геометрию стенки сердца и ориентацию в ней мы-
шечных волокон. Чтобы характеризовать развиваемое мышцей
усилие, в экспериментальных исследованиях обычно рассчитывают
некоторое среднее напряжение. Во-вторых, нужно помнить, что
соотношение сила — скорость для мышцы, строго говоря, отно-
Гл. 11. Сердце 255
сится только к сократительному элементу. Если бы во время со-
кращения напряжение поддерживалось постоянным, так что длину
последовательного упругого элемента можно было бы считать по-
стоянной, то это соотношение было бы справедливо и для целой
мышцы. В экспериментах на изолированной мышце такое условие
часто выполняется, и это привело к необоснованному представле-
нию о том, что гиперболическая зависимость характерна для по-
ведения мышцы всего желудочка. Во время сокращения целого
сердца напряжение быстро изменяется, и должны быть такие мо-
менты, когда последовательный упругий элемент удлиняется, а со-
кратительный укорачивается. Рассчитывая скорость укорочения
сократительного элемента, на это нужно делать поправку, а вве-
дение ее влечет за собой целый ряд новых допущений, включая
использование для описания свойств последовательного упругого
элемента значений модулей упругости, полученных в опытах на
изолированной мышце. При расчете скорости укорочения возни-
кает еще одна трудность. Поскольку мышца представляет собой
сплошное тело, скорость укорочения ее нужно рассматривать как
скорость, с которой уменьшается малый элемент данной окруж-
ности. Выбранная окружность должна быть именно той, для ко-
торой предстоит рассчитать напряжение. Но поскольку стенка же-
лудочка толстая, укорочение, отнесенное к единице длины во-
локна, будет разным в зависимости от радиального положения по-
следнего. Связанные с этим затруднения можно преодолеть, если
представить стенку как ряд бесконечно тонких оболочек, меняю-
щих объем с одинаковой скоростью. В таком случае можно опре-
делить среднюю по пространству скорость укорочения. Последнее
ограничение рассматриваемого подхода заключается в том, что ра-
боту целого сердца удается исследовать только в относительно не-
большом диапазоне изменения силы и скорости.
Несмотря на все эти затруднения, было четко установлено, что
факторы, существенные для деятельности изолированной сердеч-
ной мышцы, имеют самое прямое отношение и к сердцу в целом.
Это иллюстрирует рис. 11.22, на котором изображена зависимость
скорости укорочения сократительного элемента от мгновенного на-
пряжения. В этом исследовании объемы и толщину стенок вычис-
ляли по результатам измерений длины поверхностных сегментов
в известных плоскостях, считая форму желудочка эллипсоидаль-
ной. Скорость укорочения определяли по результатам измерения
скорости изгнания крови в аорту, а напряжение — по данным из-
мерения давления в полости левого желудочка. Каждая прямая
отражает соотношение величин, полученных при одном мгновен-
ном объеме желудочка. Разброс данных в таких экспериментах
весьма велик (это неудивительно, если принять во внимание слож-
ность опытов, не говоря уже о допущениях, необходимых для рас-
четов). Тем не менее из них следуют два четких вывода. Во-пер-
вых, при каждом значении объема желудочка имеет место обрат
2бв Часть It. Механика кровообращения
S
О 100 200 300 400 500 600
Напряжение, г-см-2
Рис. 11.22. Соотношения напряжение — скорость для целого желудочка, вычислен-
ные rto данным, полученным в экспериментах при различных значениях объема
желудочка. Скорость укорочения определена для сократительного элемента сер-
дечной мышцы и рассчитана на 1 см длины окружности желудочка. [Fry (1962).
Fedn Proc. Fedn Am Socs Biol., 21, 991.]
__L_
700
иая зависимость между скоростью укорочения и напряжением; во-
вторых, с увеличением объема желудочка прямые сдвигаются
вверх и вправо. Если мы примем, что увеличение объема желу-
дочка аналогично увеличению длины волокон, то сходство поведе-
ния целого сердца (рис. 11.22) и препарата изолированной мышцы
(рис. 11.14) окажется поразительным.
Подобные результаты были получены в ряде разного рода экс-
периментов, и можно с уверенностью сказать, что напряжение,
начальная длина волокон и скорость укорочения сократительного
элемента — важйые факторы, определяющие деятельность целого
сердца. И все-таки проблема остается — во многих эксперимен-
тальных исследованиях, не говоря уже о клинических условиях,
эти переменные не удается ни измерить, ни рассчитать точно. По-
этому приходится использовать другие, более доступные для из-
мерения параметры, отражающие изменения названных основных
свойств. Некоторые из таких параметров уже упоминались. Од-
нако в экспериментальных исследованиях им придают столь боль-
шое значение, что имеет смысл рассмотреть их несколько подроб-
нее.
Мы уже видели, что, изучая деятельность сердца в целом, для
представления длины мышечных волокон обычно используют
объем желудочка. Но из-за сложной геометрии желудочка связь
между этими двумя величинами оказывается по своей природе ста-
тистической— в том смысле, что объем отражает некоторую сред-
нюю-длину волокон. Кроме того, мы отмечали, что измерить объем
желудочка прижизненно весьма трудно. К счастью, это не относит*
Гл. ti. Сердце 28?
ся к давлению, и диастолическое давление в желудочке повсе-
местно используют в качестве меры его объема, а следовательно,
и меры длины мышцы в состоянии покоя. Именно поэтому ко-
нечно-диастолическое давление в литературе часто называют
«предварительной нагрузкой» по аналогии с натяжением изолиро-
ванной мышцы в состоянии покоя. В клинике нередко прибегают
к дальнейшему упрощению, предполагая, что в конце диастолы
атрио-вентрикулярный клапан все еще открыт, но поток крови
через него мал или отсутствует совсем, так что давление в пред-
сердии равно давлению в желудочке. В частности, это использует-
ся для правой стороны сердца: в этом случае давление в венах
можно грубо оценить визуально, уложив испытуемого на спину и
используя вены на шее как манометр («давление в яремных ве-
нах»), или измерить точно, введя через вену предплечья или шеи
в крупные вены внутри грудной клетки или в правое предсердие
тонкий зонд для измерения «центрального венозного давления»
(«давления наполнения»).
Совершенно очевидно, что использование конечно-диастоличе-
ского давления для определения начальной длины мышцы стенки
желудочка является чисто качественным методом. Прежде всего
такой метод предполагает, что соотношение между давлением в
желудочке и его объемом не подвержено влиянию времени, т. е.
что оно не может изменяться при заболеваниях и что материалу
желудочка не присуща ползучесть. Справедливость последнего
предположения, по-видимому, не подвергалась критической про-
верке. Для непродолжительных опытов и, в частности, для опытов,
в которых изменяют наполнение сердца от удара к удару,
оно может быть несущественным. Однако в покоящейся изо-
лированной сердечной мышце ползучесть наблюдается совер-
шенно отчетливо и может иметь значение в условиях, когда в фазе
диастолы сердце растянуто в течение длительного времени. Еще
одно обстоятельство, о котором иногда забывают, заключается в
том, что степень растяжения стенки желудочка определяется
именно разностью давлений по обе стороны этой стенки (трансму-
ральным давлением), а когда грудная клетка закрыта, давление
снаружи желудочка меняется при дыхании. Например, во время
вдоха давление в грудной клетке становится субатмосферным, и
если трансмуральное давление низкое, то и давление в желудочке
может быть тоже субатмосферным. Это не слишком утрирован-
ный пример: в норме пределы изменения конечно-диастолического
давления в левом желудочке имеют тот же порядок, что и проис-
ходящие даже при спокойном дыхании колебания давления в груд-
ной полости.
В настоящее время измерение у человека конечно-диастоличе-
ского давления при помощи зонда не является сложной задачей.
Большой объем желудочков — характерная черта многих заболе-
ваний сердца у человека, особенно при сердечной недостаточности.
2Б8 Часть 11. Механика кровообращения
и потому все приведенные выше соображения имеют большое
Практическое значение. Немаловажное недавнее достижение — со-
здание особых ультразвуковых приборов, которые работают по
принципу корабельного эхолота. Они позволяют определять раз-
меры левого желудочка (эхокардиография) через стенку грудной
клетки и следить за движением стенки желудочка (а следова-
тельно, и за скоростью укорочения мышцы) на протяжении всего
сердечного цикла. Тем самым ультразвуковые приборы откры-
вают путь к бескровному способу оценки размеров сердечной ка-
меры, а вместе с измерением давления при помощи зондов — и к
возможности впрямую исследовать соотношения давление — объем
в разнообразных экспериментальных и клинических условиях.
Итак, для описания условий, при которых в любом экспери-
менте начинается сокращение желудочка, конечно-диастолическое
давление может быть чрезвычайно полезным параметром, хотя
нельзя утверждать, что его абсолютное значение однозначно свя-
зано с длиной мышечных волокон в состоянии покоя. Этот метод
неоценим для изучения целого сердца, в частности как простой
способ наблюдения за длиной покоящейся мышцы сердечной
стенки.
Таким образом, измерение конечно-диастолического давления
позволяет, по крайней мере частично (для фазы диастолы), избе-
жать необходимости измерения объема как показателя длины мы-
шечных волокон. Для фазы систолы, во время которой объем же-
лудочка изменяется по мере изгнания крови, проблема возникает
снова. Правда, в этом случае с ней справиться легче: в опытах на
животных можно измерить скорость изгнания крови из желудочка,
используя расходомер, датчик которого расположен на аорте или
на легочном стволе (этим методом получены записи скорости кро-
вотока на рис. 11.6). Измеренная скорость кровотока несомненно
связана со скоростью укорочения мышечных волокон, однако и
здесь есть свои сложности: по геометрическим причинам это соот-
ношение нелинейно. Например, если обратиться к эллипсоидаль-
ной модели желудочка с фиксированной большой осью, то ока-
жется, что малая ось, а следовательно, и сегменты окружности
укорачиваются пропорционально квадратному корню из измене-
ния объема, а абсолютное укорочение зависит от начальной длины
этой оси. Таким образом, при установлении количественного соот-
ношения между поведением целого желудочка и мышечных воло-
кон опять возникают геометрические проблемы!).
’) Если, рассматривая возможность использования моделей мышечного во-
локна для описания работы целого желудочка, мы хотим использовать как можно
более строгий подход, то иам нужно знать скорость укорочения сократительного
элемента мышцы, а ие всего волокна Для этого следует либо предположить,
что напряжение во время систолы остается постоянным (так что последователь-
ный упругий элемент сохраняет постоянную длину), либо (чтобы можно было
использовать для пересчета данные о скорости укорочения всего волокна) обра-
Гл. 11. Сердце 259
За исключением очень специальных условий, измерить мгно-
венную скорость кровотока в аорте человека невозможно, и един-
ственным выходом является измерение минутного объема сердца
обычными методами, например методом разведения индикатора.
Этот метод дает величину объемной скорости кровотока (расхода
крови), усредненную по времени. Ее можно разделить на частоту
сокращений сердца и получить величину ударного объема, но
нельзя использовать для описания мгновенных событий во время
систолы.
Еще одним представляющим интерес параметром является на-
грузка, против которой кровь изгоняется из желудочка. Оценить
ее достаточно точно наиболее трудно. Когда в начале систолы дав-
ление в желудочке повышается до уровня, слегка превышающего
давление в аорте, аортальный клапан открывается и начинается
фаза изгнания (укорочения мышцы). На первый взгляд давление
в аорте (которое при нормальной систоле лишь ненамного отли-
чается от давления в желудочке, см. рис. 11.5) во время этой
фазы представляет собой аналог той «рабочей нагрузки», против
которой укорачивается изолированная мышца (см. рис. 11.12),
так же как конечно-диастолическое давление аналогично «предва-
рительной нагрузке», которая определяет начальную длину мыш-
цы и ее предварительное натяжение. Однако, как мы видели
раньше, чтобы выразить нагрузку на мышцу желудочка в вели-
чинах, аналогичных нагрузке, применяемой в опытах на изолиро-
ванной мышце, необходимо от давления в полости желудочка пе-
рейти к напряжению в его стенке. Эти величины связаны между
собой через геометрические параметры — объем и толщину стенки
желудочка. Последние же, как и давление, в течение фазы изгна-
ния меняются. Объем желудочка во время систолы снижается
довольно равномерно, но давление в нем сначала возрастает, а
потом падает (рис. 11.5). Поэтому если мы предположим, что же-
лудочек обладает упругостью, и рассмотрим окружное напряже-
ние, то обнаружим, что в начале систолы оно может быть при-
мерно постоянным, но затем, по мере уменьшения радиуса кри-
визны и увеличения толщины стенки, уменьшается. Таким обра-
зом, напряжение падает быстрее, чем давление [см. уравнение
(7.6)]. Другими словами, соотношение объем — давление для со-
кращающегося желудочка отличается по форме от соотношений
длина — натяжение для изолированных мышечных волокон, и ис-
пользование систолического давления для описания нагрузки на
уровне мышечных волокон в лучшем случае будет очень грубым
приемом. Мы опять сталкиваемся с проблемой, обусловленной
титься к сведениям о последовательном упругом элементе, полученным в опытах
на изолированной мышце Но даже этот подход — он применен при построении
рис. 11.22 — не является полностью удовлетворительным, так как предполагает,
что полученные в предельно искусственных экспериментальных условиях коли-
чественные результаты применимы и в общем случае, т. е. для целого сердца.
260 Часть // Механика кровообращения
сложной геометрией желудочка: величины, которые можно изме'
рить, не соответствуют величинам, которые действительно опре-
деляют поведение мышечных волокон.
Подводя итог, повторим, что свойства, определяющие поведе-
ние изолированных препаратов мышцы желудо.чка, определяют
также и поведение сердечной мышцы в целом. Но при переходе
к изучению сердца свойства мышцы оказываются отделенными от
свойств целой камеры барьером неопределенностей — в основном
геометрических. Поэтому описать работу желудочка исходя из
рассмотренных в этой главе основных свойств мышцы обычно не
удается.
Работа желудочка как насоса. Обратимся к другому под-
ходу:— эмпирическому рассмотрению деятельности сердца в пер-
вую очередь как насоса, а не как мышцы. Этот подход более ста-
рый: его основы заложил на исходе XIX в. немецкий физиолог
О. Франк, а спустя несколько лет тщательно разработал Э. Стар-
линг в Англии. Таким образом, происхождение данного подхода
чисто экспериментальное, и основные его идеи предшествовали
познанию механики мышц.
В этих ранних опытах исследовали в основном взаимосвязь ме-
жду притоком и оттоком крови. Франк работал на сердце лягушки
и использовал довольно грубые методы, а Старлинг достиг исклю-
чительных успехов, развив метод сердечно-легочного препарата,
который позволяет отъединить работающие сердце и легкие нар-
котизированного животного от сосудов всех остальных органов.
Сущность метода заключается в том, что кровь, изгоняемую из
левого желудочка в аорту, собирают в резервуар и возвращают в
сердце через правое предсердие. Такой препарат позволяет неза-
висимо контролировать давление наполнения предсердия и желу-
дочка и давление в аорте. Старлинг обнаружил, что по мере уве-
личения притока крови в предсердие давление в нем постепенно
растет и ударный объем увеличивается (рис. 11.23). Указанное
соотношение оказалось нелинейным: при равномерном ступенча-
том увеличении венозного давления каждое последующее его при-
ращение сопровождалось все меньшим приращением ударного
объема, пока в некоторых случаях последний не начинал умень-
шаться. Далее Старлинг установил, что этот эффект относится в
равной мере как к правым, так и к левым камерам сердца, так
что, когда правые камеры в ответ на увеличение наполнения по-
вышают свою производительность, соответствующий дополнитель-
ный объем крови попадает в левые камеры, где вызывает тот же
самый эффект. В следующей серии опытов, в которых измерялись
также изменения объема желудочка, Старлинг показал, что при
повышении аортального давления, когда сердце вынуждено выпол-
нять большую работу, ударный объем сначала падает и желудо-
чек расширяется; затем ударный объем восстанавливается до
своей исходной величины.
Гл 11 Сердце 261
Рис. 11.23. Воспроизведение одной из записей, полученных Старлингом на сер-
дечио-легочном препарате. Запись на закопченной ленте кимографа показывает
влияние увеличения венозного давления наполнения (1, без калибровки) на диа-
столический и систолический объемы сердца II — артериальное давление в мм
рт. ст.; калибровочные отметки (100 и 75 мм рт. ст) нанесены ниже; III — не-
прерывная запись объема обоих желудочков, полученная при помощи кардио-
метра (жесткой латунной камеры, в которую помещены желудочки, во время диа-
столы воздух из этой камеры вытесняется в измерительный цилиндр), шкала
прокалибрована в мл Отметка времени 1 с А. Исходное состояние Венозное
давление 95 мм вод. ст. Производительность желудочков 520 мл-мин-1. Б. Бы-
строе повышение венозного давления до 145 мм вод. ст Обратите внимание на
соответствующее увеличение как диастолического объема желудочков, так и их
ударного объема. Производительность желдочков 840 мл мин-1. В Быстрое
снижение венозного давления до 55 мм вод ст Как диастолический, так и удар-
ный объем желудочков падают ниже исходного уровня. Производительность же-
лудочков 200 мл-мин-1. [Patterson, Piper, Starling (1914). The regulation oi the
heart-beat. J. Physiol, 48, 465.]
262 Часть II. Механика кровообращения
В свете современных представлений о механике сердечной
мышцы (которые начали складываться с конца 50-х годов) мы
можем сказать, что Старлинг продемонстрировал поведение этой
мышцы при укорочении от различных исходных длин. А зная те-
перь ультраструктуру саркомеров, мы имеем по крайней мере ка-
кие-то ключи к пониманию того, каким образом это может проис-
ходить. Представление о том, насколько глубоко понимал Стар-
линг поведение сердца, дают его слова, сказанные в 1914 г.: «За-
кон сердца заключается, следовательно, в том... что механическая
энергия, рысвобождающаяся при переходе из состояния покоя в
состояние сокращения, зависит от площади «химически активных
поверхностей», т. е. от длины мышечных волокон».
Таким образом, закон Старлинга для сердца (или, как его
по-иному называют, механизм Франка — Старлинга) отражает
важную связь между начальным наполнением сердца (т. е. дли-
ной мышцы) и объемом изгоняемой крови. Правда, это соотноше-
ние было выражено в весьма расплывчатых механических тер-
минах— такие слова, как «энергия» и «работа», оставались недо-
статочно определенными, — но оно четко устанавливало тот спо-
соб, при помощи которого согласуется производительность двух
«последовательных» насосов, составляющих сердце млекопитаю-
щего, и обеспечивается их ответ на изменения давления наполне-
ния (предварительная нагрузка) и артериального давления (ра-
бочая нагрузка). Говоря в двух словах, Старлинг эксперимен-
тально установил, что и правые, и левые камеры сердца в силу
присущих им свойств способны при самых разных условиях изго-
нять весь тот объем крови, который возвращается к ним.
Сам Старлинг хорошо сознавал ограниченность такого описа-
ния деятельности сердца. Очевидным недостатком этого описания
является то, что оно не количественное; даже для одного и того
же сердца не существует постоянного соотношения между давле-
нием наполнения и производительностью. Например, во втором из
упомянутых выше опытов за увеличением объема желудочка, со-
провождающим ступенчатое повышение давления в аорте, может
после нескольких ударов наблюдаться постепенное уменьшение ко-
нечно-диастолического объема желудочка (т. е. давления напол-
нения) до его исходного уровня без какого-либо падения произ-
водительности сердца.
Старлинг полагал, что это обусловлено увеличением коронар-
ного кровотока и соответственно «улучшением питания мышцы».
В действительности указанное явление наблюдается, даже когда
коронарный кровоток поддерживается постоянным, и дело здесь,
вероятно, скорее в улучшении распределения крови в миокарде,
обеспечивающем лучшее питание его внутренних слоев. Мы упо-
мянули об этом лишь для того, чтобы наглядно показать, что на-
грузка может оказывать влияние иа соотношение между давле-
нием наполнения и ударным объемом желудочка. Чтобы предста-
Гл. 11. Сердце 268
<4000
3000
2000
1000
120
180
240
, 3000
CJ'
, 180
в*о 240
'е ,
о
ч3"
ч.
300
4000
3600
3000
2600
2000
1500
1000
600
20
18 X’
14Х
8 ;♦$£
X’
ё
“ё
Рис. 11.24. Производительность сердца собаки (вес тела 10 кг) при разных зна-
чениях венозного давления наполнения и давления в аорте (сердечно-легочный
препарат). Обратите внимание, что во всем физиологическом диапазоне давлений
производительность сердца очень чувствительна к изменению давления наполне-
ния (предварительной нагрузки), но не к изменению артериального давления
(рабочей нагрузки). [Herndon, Sagawa (1969). Am. J. Physiol., 217, 65.]
вить это графически, нужно построить семейство кривых, отра-
жающих такое соотношение при различных нагрузках, т. е. при
различном среднем артериальном давлении, либо использовать
для графика три оси (см. рис. 11.24).
Уже после опытов Старлинга было установлено, что снижение
производительности сердца из-за дополнительного увеличения и
без того высокого давления наполнения (в течение ряда лет кли-
ницисты считали это снижение основным механизмом развития
сердечной недостаточности, которую приписывали растяжению
мышечных волокон сверх каких-то функциональных пределов) в
действительности обусловлено недостаточностью атрио-вентрику-
лярного клапана; при значительном расширении желудочка кла-
панное отверстие оказывается сильно увеличенным, и в результате
часть изгоняемой крови возвращается через него в предсердие.
Опыты Старлинга были повторены в самых разных условиях,
причем мерой наполнения служило конечно-диастолическое дав-
ление в левом желудочке, а показателем «достаточности» силы
сокращения — работа за удар (рис. 11.25). Последняя опреде-
264 Часть II. Механика кровообращения
ляется как произведение ударного объема на давление в аорте
и потому имеет соответствующую для описания внешней работы
размерность: [сила]X[расстояние]. Введение для оценки деятель-
ности желудочка вместо принятого Старлингом ударного объема
именно этой величины вызвано двумя обстоятельствами. Во-пер-
вых, многие из своих выводов Старлинг представил в энергетиче-
ских терминах, так что работа (которая имеет ту же самую раз-
мерность) казалась вполне подходящей мерой. Во-вторых, вели-
чина работы учитывает (по крайней мере грубо) нагрузку, против
Рис. 11.25. Статическая характеристи-
ка желудочка — зависимость работы,
совершаемой левым желудочком за
удар, от величины среднего давления
в левом предсердии. [Sarnoff (1955).
Physiol. Rev, 35, 107.]
которой осуществляется изгнание, потому что она включает в себя
величину среднего давления в аорте. Для сеодечно-легочного пре-
парата, в котором аортальное давление можно контролировать
и поддерживать постоянным, это обоснованно. Но при изучении
кровообращения в целом организме работу за удар следует ис-
пользовать весьма осторожно, так как давление в аорте не свя-
зано непосредственно с конечно-диастолическим давлением. На-
пример, работа за удар не остается постоянной при изменении
давления в аорте: она равна нулю, когда желудочек изгоняет
кровь без противодействующего давления, достигает максимума
при повышении этого давления, а когда давление увеличивается
настолько, что изгнание становится невозможным, снова снижает-
ся до нуля. Поскольку в системе кровообращения всего организма
аортальное давление отражает не только эффективность изгнания
крови из желудочка, но и свойства периферического сосудистого
русла, работа за удар может быть в высшей степени ненадежным
показателем деятельности сердца. Работа за удар представляет
собой весьма несовершенную оценку деятельности миокарда и по
другим причинам. В частности, она не учитывает ни работы, за-
трачиваемой на приведение крови в движение, ни внутренних по-
терь энергии в мышце (обусловленных, например, трением).
Гл. 11. Сердце 265
Среднее давление в левом предсердии см вод ст
Рис. 11.26. Влияние раздражения симпатических нервов, обладающих сильным
положительным инотропным действием, на деятельность желудочка в условиях
сердечно-легочного препарата. Каждая кривая отражает соотношение между
средним давлением в левом предсердии и работой левого желудочка за удар
(статические характеристики желудочка) при последовательном повышении ча-
стоты раздражения симпатических нервов сердца (частота раздражения — число
импульсов в секунду — указана около каждой кривой). [Sarnoff, Mitchell (1961).
Am. J. Med., 30, 747.]
Если условия соответствующим образом контролируются, то
такие исследования показывают (как подразумевал, но никогда
прямо не формулировал Старлинг), что кривая, связывающая ко-
нечно-диастолическое давление и работу за удар (обычно назы-
ваемая «кривой функционирования желудочка», или статической
характеристикой его), не единственная. Для любого конкретного
сердца имеется семейство таких кривых, так что помимо длины
волокон в конце диастолы деятельность желудочка должен опре-
делять и какой-то другой фактор.
Этот второй определяющий фактор обычно называют сократи-
тельным состоянием, или сократимостью желудочка. Воздействия,
изменяющие сократимость желудочка, как упоминалось ранее, на-
зывают инотропными. Они могут оказывать положительный эф-
фект,— сдвигая статическую характеристику желудочка так, что
при заданном конечно-диастолическом давлении в левом желу-
дочке ударный объем или работа за удар увеличиваются
(рис. 11.26), — или отрицательный. Такие сдвиги кривой обычно
266 Часть II. Механика кровообращения
описывают как изменение сократимости, и они могут быть конеч-
ным результатом влияния самых разнообразных факторов — тем-
пературы, кровоснабжения, насыщения кислородом, ионного и хи-
мического состава внутренней среды. Наиболее важными из груп-
пы химических положительно инотропных агентов (которые вполне
могут иметь общей точкой приложения внутриклеточные про-
цессы высвобождения энергии) являются катехоламины — природ-
ные или синтетические вещества, аналогичные гормону адрена-
лину. Одно из них, изадрин (изопреналин), особенно эффективно,
и потому его широко применяют как в исследованиях, так и в
клинике. Аналогичное влияние оказывает на деятельность желу-
дочков раздражение симпатических нервов, идущих к сердцу (по-
видимому, это действие осуществляется через посредство норад-
реналина, высвобождающегося из нервных окончаний). Увеличи-
вают сократимость сердца и некоторые лекарственные препараты,
а также повышение концентрации кальция и других ионов в цир-
кулирующей крови. Возбуждение симпатической нервной системы
и поступление в кровь адреналина из надпочечников — это нор-
мальные составляющие некоторых реакций у животных (обычно
относимых к разряду оборонительных реакций — «нападения или
бегства»), а также мышечной работы и эмоционального напряже-
ния у людей. Значение их как дополнительных к механизму Фран-
ка— Старлинга факторов, определяющих поведение здорового
сердца, выявлено лишь относительно недавно. Представления Стар-
линга о связи наполнения сердца с его производительностью были
приняты сразу, но расширение этих представлений с учетом дей-
ствия внешних инотропных факторов произошло лишь почти через
сорок лет. Детальное изучение влияния на деятельность желу-
дочка адреналина и возбуждения симпатической системы и появ-
ление концепции «сократимости» приблизительно совпали с раз-
витием представлений о свойствах изолированной сердечной
мышцы.
Концепция сократимости наталкивается на определенные труд-
ности. Первая из них относится к области определений. Легко
видеть, каким образом, скажем, раздражение симпатических нер-
вов может «устанавливать» статическую характеристику желудоч-
ка на различные уровни (рис. 11.26). Но дать определение этому
эффекту, сказав, что увеличение ударного объема при заданном
конечно-диастолическом давлении обусловлено увеличением со-
кратимости, невозможно, потому что к тому же самому резуль-
тату могло бы привести снижение аортального давления (см.
рис. 11.24), а в этом случае причина эффекта может вовсе не
быть связанной с сердцем. Следовательно, для определения сокра-
тимости таким образом нужно сначала описать статическую ха-
рактеристику желудочка, а это, очевидно, очень громоздко. Кроме
того, изучать сократимость желудочка путем исследования такой
характеристики чрезвычайно трудно: для того чтобы действие на
Гл. 11. Сердце
ударный объем или на работу за удар можно было приписать из-
менениям сократимости, необходимо строго контролировать актив-
ность симпатических нервов, конечно-диастолическое давление и
давление в аорте. Наконец, сравнивать количественно деятель-
ность сердца в разных экспериментах — дело весьма сложное.
Эти причины, а также постепенное осознание того, что деятель-
ность сердечной мышцы можно было бы охарактеризовать более
основательно с тех же позиций, что и поведение скелетной мышцы,
лишили попытки описывать сердце как насос побуждающего сти-
мула и привели к поискам способов количественного определения
Рис. 11.27. Влияние положительного ино-
тропного воздействия (добавления в ван-
ночку, в которой находится препарат,
норадреналина) на соотношение сила —
скорость для изолированной сердечной
мышцы Начальная длина мышцы под-
держивается постоянной. [Sonnenblick
(1962). Am. J. Physiol, 202, 931.]
сократимости мышцы сердца на основе уже рассмотренного в этой
главе соотношения ее свойств: сила — скорость — длина.
Конечная цель заключалась, конечно, в том, чтобы найти не-
кий показатель, который можно было бы как в эксперименте, так
и в клинике применять для целого сердца. Мы уже видели, какие
трудности возникают на пути применения механики мышцы к це-
лому желудочку, и поэтому нет ничего удивительного в том, что
для целого сердца «сократимость» все еще остается нечетко опре-
деленной величиной. С другой стороны, для препаратов сердечной
мышцы ее можно определить: влияние изменений длины удается
в этом случае устранить, поддерживая длину мышцы постоянной,
и тогда воздействия на инотропное состояние мышцы отражают-
ся, как показано на рис. 11.27, в изменении положения кривой
сила — скорость. Положительно инотропные агенты сдвигают эту
кривую вверх. Таким образом, инотропное влияние было опреде-
лено как такое влияние, которое, действуя на мышцу в условиях
постоянства ее длины, смещает кривую сила — скорость. Это
сразу же наводило на мысль, что точки пересечения кривой
сила — скорость с осями координат можно использовать как пока-
затели сократимости мышцы, получая путем экстраполяции кри-
вой либо силу при нулевой скорости (изометрическое усилие Ро),
268 Часть II. Механика кровообращения
либо скорость укорочения сократительного элемента при нулевой
нагрузке Утах-
Однако попытка использовать в качестве показателя сократи-
мости изометрическое усилие приводит к непреодолимым трудно-
стям. Мы уже видели, с какими сложностями и допущениями свя-
зан расчет действующих в стенке желудочка напряжений по ве-
личине давления в его полости. А в данном случае к тому же
важно быть уверенным, что длина мышцы (т. е. конечно-диастоли-
ческий объем желудочка) за время между измерениями не изме-
няется. (Очень сильное влияние начальной длины мышцы на изо-
метрическое усилие видно из рис. 11.14, где точки пересечения
кривых с осью нагрузок дают параметры изометрического сокра-
щения.) Грубый подход — приравнять максимальное напряжение
в стенке к максимальному давлению, развиваемому в желудоч-
ке,—довольно прост, но в данном случае неприменим, так как
максимальное давление достигается уже после открытия клапана
(см. рис. 11.4), когда длина мышцы изменяется.
Гораздо большие надежды возлагали всегда на осуществи-
мость и обоснованность использования в качестве показателя со-
кратимости величины Утах. Обусловлено это было главным обра-
зом следующим: полагая, что все кривые на рис. 11.14 можно
экстраполировать к одной и той же начальной точке на оси ско-
ростей, Утах считали независящей от длины мышцы, но в то же
время существенно зависящей от инотропного состояния, так как
кривые на рис. 11.27 экстраполируются к разным точкам на оси
скоростей. Предположение о независимости Утах от длины об-
суждалось раньше (разд. 11.3) и по приведенным там причинам
было подвергнуто резкой критике. Но из-за практической ценно-
сти, которую мог бы иметь показатель сократимости, а также
потому, что другие способы его определения оказались бесплод-
ными, эта идея все еще стимулирует множество работ и служит
предметом живой полемики.
Поначалу оправданием служило то, что вычисленное по абсо-
лютной величине и скорости повышения давления в желудочке и
при определенных предположениях о свойствах последовательного
упругого элемента, о синхронности сокращения и о геометрии же-
лудочка значение Утах могло бы быть показателем сократимости,
независящим от изменений длины волокон желудочка (т. е. от ме-
ханизма Франка — Старлинга). Однако в последние годы сомне-
ния относительно постоянства Утах при изменениях начальной
длины волокон неуклонно росли. Все это вместе с неопределенно-
стями и сложностями требуемых вычислений заставило обратить-
ся к более простым, непосредственно измеряемым параметрам,
которые могли бы изменяться в ту же сторону, что и сократи-
мость. Наибольшее внимание привлекла скорость повышения дав-
ления в желудочке — одна из величин, которые необходимо было
измерять для расчета Утах- Ход развития давления в желудочке и
Гл. 11. Сердце 269
скорость его изменения (dp/d/) показаны на рис. 11.28. Скорость
увеличения давления достигает максимума (dp/df)max в конце
изоволюмического периода, и поскольку этот максимум может
иметь какое-то отношение к максимальной скорости укорочения
сократительных элементов мышцы и несомненно значительно из-
меняется при воздействии инотропных агентов, внимание было
сфокусировано именно на нем. Строго говоря, dp/d/ непосред-
ственно не измеряется; эту величину получают, дифференцируя
Рис 11.28. Давление в левом желудочке (верхняя запись) и скорость его измене-
ния (нижняя запись). Давление зарегистрировано при помощи высокоточного
манометра, помещенного на конце введенного в желудочек зонда; манометр вос-
производит достаточно широкий спектр частот; сигнал скорости изменения дав-
ления получен путем дифференцирования сигнала давления при помощи соответ-
ствующего электронного устройства. [Gabe (1972). Cardiovascuar fluid dynamics,
p. 14, Academic Press, New York.]
при помощи электронного устройства сигнал давления в желу-
дочке, которое действительно измеряют. (Более традиционный ме-
тод— проведение касательной к возрастающей части записи дав-
ления— весьма ненадежен, потому что временной интервал, в те-
чение которого наклон кривой записи давления максимален, очень
мал — см. рис. 11.28). Сигнал давления должен быть неискажен-
ным, чего трудно достичь, используя длинные заполненные жид-
костью зонды. Но с помощью современных манометров, располо-
женных на конце зонда, такие измерения могут быть выполнены
не только у животных, но и у человека.
Вопрос об интерпретации величины (dp/dOraax и целесообраз-
ности ее измерения является в настоящее время предметом дис-
куссий. Эта величина должна, например, зависеть от начальной
длины мышцы, так как она отражает скорость увеличения напря-
жений, развиваемых в стенке, а скорость эта, как мы знаем, за-
висит от длины. Далее, поскольку значение (dp/d/)max достигает-
ся в конце изоволюмического периода, оно может также умень-
шиться, если произойдет раннее открытие аортального клапана и
мышца желудочка начнет укорачиваться, например в случае низ-
270 Часть II. Механика кровообращения
кого аортального давления. Другими словами, эта величина мо-
жет быть чувствительна как к рабочей, так и к предварительной
нагрузке. Большинство исследований свидетельствует о том, что
параметр (dp/df)max подвержен влиянию обоих этих факторов и
его несомненно никак нельзя считать универсальным показателем
сократимости желудочка. Но при некоторых определенных усло-
виях, например когда давление в аорте и конечно-диастолическое
давление в левом желудочке стабильны или контролируются, он
может оказаться весьма ценным. Между тем поиски применимого
во всех случаях количественного показателя сократимости про-
должаются, хотя поборники «чистой науки», детально изучившие
проблемы механики сердца, убеждены, что никакого универсаль-
ного показателя существовать не может. Прагматики же, озабо-
ченные необходимостью оценивать состояние сердца у больных,
готовы (даже с оптимизмом) испытывать эмпирически почти лю-
бой подающий надежды параметр, доступный измерению, и их
можно понять. Что в конечном счете необходимо, так это, по всей
видимости, измерение параметров, характеризующих биохимиче-
ские процессы внутри клетки. Процессы высвобождения энергии
в мышце (хотя мы здесь на них не останавливались) настойчиво
и весьма плодотворно изучаются. Однако над этой проблемой
как над задачей механики надо еще упорно работать.
Заключение. Деятельность сердца изучали с двух основных
точек зрения. В рамках первой из них, исторически более старой,
к сердцу подходят как к насосу и изучают соотношение между
входом (обычно выражаемым через давление в венах или в пред-
сердии) и выходом (обычно измеряемым как ударный объем), не
пытаясь понять лежащие в основе э*гого соотношения механизмы.
С этой точки зрения поступающая в каждую камеру сердца кровь
растягивает ее, и изгоняемый затем во время сокращения объем
крови оказывается пропорциональным этому растяжению. Такая
зависимость известна как механизм Франка — Старлинга и ответ-
ственна не только за управление производительностью сердца в
соответствии с величиной венозного притока, но и за согласова:
ние производительности левых и правых камер сердца. Ясно, од-
нако, что такое описание не дает полного представления о дея-
тельности сердца, так как не учитывает нагрузки («рабочей на-
грузки»), против которой сердце должно работать, изгоняя удар-
ный объем. Чтобы учесть ее, надо принять во внимание зависи-
мость между конечно-диастолическим давлением в желудочке и
работой за удар. Последняя определяется как произведение удар-
ного объема на среднее давление в аорте и в действительности
является несовершенной мерой производимой мышцей желудочка
внешней работы. Указанную зависимость называют «кривой функ-
ционирования желудочка», или его статической характеристикой.
Она не является единственной для какой бы то ни было конк-
ретной сердечной камеры, так как на деятельность сердечной
Гл. It. Сердце 271
мышцы оказывает влияние еще и третий существенный фактор —
состояние сократимости мышцы. Поэтому для полного описания
поведения сердца необходимо располагать семейством статиче-
ских характеристик желудочка, отражающим влияния предвари-
тельной нагрузки, рабочей нагрузки и сократимости.
Вообще говоря, такой подход чрезвычайно полезен. Действи-
тельно, легко представить себе кривую, которая отражает зави-
симость между входом и выходом и в ответ на требование орга-
низма изменить общий объем циркулирующей крови сдвигается
вверх или вниз. Однако применимость указанного подхода в экс-
периментальных исследованиях ограничена, и, кроме того, по-
скольку все параметры в этом случае рассматриваются как усред-
ненные по времени, его совсем нельзя использовать для описания
хода событий в пределах одного сердечного цикла. Таким обра-
зом, этот подход дает ясное, но далеко не полное описание ра-
боты сердца.
Второй подход заключается в том, чтобы исследовать деятель-
ность сердца на основе механики мышцы и попытаться установить
соответствие между свойствами, обнаруженными в опытах на изо-
лированных мышечных волокнах, и поведением целой камеры.
Из-за сложной геометрии желудочков и наличия определенной
ориентации волокон в их стенках для этого необходимо произ-
вести ряд чрезвычайно сложных вычислений. Серьезные попытки
сделать это предпринимались до сих пор только для левого же-
лудочка. Значительное количество данных, полученных к настоя-
щему времени, подтверждают ту точку зрения, согласно которой
свойства, обнаруживаемые у изолированных мышц, действительно
определяют поведение камеры в целом, но реального способа
оценки работы сердца в клинике и в эксперименте пока не суще-
ствует, так как необходимые для этого измерения слишком
сложны, а многие предположения, используемые при вычислениях,
весьма ненадежны. К сожалению, об этих ограничениях нередко
забывают перед лицом растущих потребностей медицины в пока-
зателях качества работы сердца, пригодных как для диагностики
заболеваний, так и для оценки правильности выбора и эффектив-
ности различных вццов лечения — хирургического и медикамен-
тозного.
Следует, наконец, подчеркнуть еще один важный момент: все
сказанное в этой главе относится к серду, изолированному от со-
судистой системы. Мы исследуем давление и расход крови на входе
и на выходе исключительно с точки зрения работы сердца. Однако
в целом организме внешние влияния, действуя через механизмы,
которые мы обсуждали, непрерывно изменяют поведение сердца.
И если иногда нам удается предсказать, как отразятся на работе
сердца те или другие реакции сосудистой системы, то сделать об-
ратное мы не можем. Система кровообращения постоянно нахо-
дится в состоянии рефлекторного приспособления, обусловленного
272 Часть II. Механика кровообращения
как потребностями тканей в крови, так и механизмами «обратной
связи», которые быстро отвечают на ряд стимулирующих воздей-
ствий типа изменения давления и химического состава крови. Дру-
гими словами, множество процессов в сосудистом русле тела осу-
ществляется совершенно независимо от работы сердца.
11.5. Гидродинамические аспекты деятельности сердца
Правые камеры сердца. Факторы, управляющие притоком
крови в правое предсердие, весьма многообразны, но главной дви-
жущей силой является артериальное давление. При прохождении
крови через микрососуды большого круга пульсации, видимые на
записях давления в аорте, в основном сглаживаются, а средний
уровень давления в значительной степени снижается, но на веноз-
ном конце капиллярного русла он еще достаточно высок, чтобы
обеспечить движение крови по направлению к сердцу. Сглажива-
ние пульсаций означает, что эта движущая сила является в основ-
ном стационарной. Однако существуют и другие факторы, благо-
даря которым на поток накладываются колебания. Некоторые из
этих факторов, например работа «мышечного» насоса (гл. 14),
являются по происхождению периферическими и вызывают коле-
бания давления, распространяющиеся по сосудам преимущественно
к сердцу. Другие колебания являются побочным следствием коле-
баний давления в грудной полости, обусловленных дыханием. На-
конец, изменения давления происходят в самом предсердии. Они
обусловлены сокращениями сердца и из-за отсутствия клапанов
при входе вен в сердце (см. гл. 14) распространяются по венам в
обратном направлении.
Таким образом, приток крови в правое предсердие во времени
непостоянен, и давление в правом предсердии, а также в соседних
венах, расположенных в грудной полости, на всем протяжении сер-
дечного цикла колеблется, даже если человек находится в состоя-
нии покоя и дышит спокойно (рис. 11.5, 11.29 и 14.20). Волна давле-
ния имеет две основные составляющие: а- и v-волны. Первая об-
условлена систолой предсердия; вторая растет медленнее, совпадая
с систолой желудочка, и заканчивается быстрым спадом (у-спад),
возникающим при открытии трехстворчатого клапана, когда в на-
чале диастолы кровь начинает поступать в желудочек. При гори-
зонтальном (или почти горизонтальном) положении испытуемого
эти колебания заметны в яремной вене на шее. При заболеваниях
их форма может дать ценные сведения о функциональном состоя-
нии правых камер сердца. Более подробно механизмы притока
крови в вены грудной полости мы разберем в гл. 14.
Движение крови в правых камерах сердца детально не изучено.
Структурное сходство позволяет предположить, что поведение их
клапанов такое же, как и клапанов левых камер сердца. Работа
Последних будет рассмотрена ниже. Вместе с тем механика сокра-
Гл. 11. Сердце 273
125 ->
Давление в аорте,
мм рт.ст. 1ПП _
75 J
Кровоток в
в легочном
стволе,
см3-мин-*
Давление в
в передней
полой вене,
мм вор и. ст.
Рис. 11.29. Одновременная запись давления в аорте, кровотока в легочном стволе
и кровотока и давления в передней полой вене собаки. Обратите внимание, что
в передней полой вене максимум кровотока, направленного к сердцу (положи-
тельные сигналы), приходится на фазу систолы желудочка, а максимум давления
(обусловленный сокращением правого предсердия) совпадает с коротким перио-
дом обратного движения крови [Brecher (1956). Venous return, Grune and Strat-
ton Inc., New York.]
щения правого и левого желудочков несомненно различается как
по способу изменения объема, так и по временным параметрам:
правый желудочек сокращается медленнее (см. рис. 11.6). В на-
чале систолы давление в правом желудочке повышается медлен-
нее, чем в левом. Но среднее давление в легочном стволе настолько
ниже среднего давления в аорте (10 мм рт. ст. против 100 мм рт. ст.),
что легочный клапан открывается раньше аортального и изгнание
крови из правого желудочка начинается за несколько миллисекунд
до начала ее изгнания из левого желудочка. Ускорение и замедле-
ние движения крови в легочном стволе, а также пиковая скорость
кровотока в нем значительно меньше, чем в аорте. Поэтому вся
волна кровотока более плавная и сглаженная (рис. 11.6), фаза
изгнания длительнее и клапан закрывается позднее, чем аорталь-
ный. По этой причине второй тон сердца имеет две составляющие
(рис. 11.5).
Рассчитанное в соответствии с уравнением (5.5) по значениям
пиковой скорости движения крови во время систолы, диаметра ле-
гочного ствола и кинематической вязкости крови пиковое число
Рейнольдса для легочного ствола несколько меньше, чем для аор-
ты. Приводить точные значения не имеет смысла, так как они, ве-
роятно, даже для здоровых животных в состоянии покоя лежат в
очень широком диапазоне, а различным экспериментальным уело*
274 Часть II. Механика кровообращения
виям, несомненно, соответствует еще более широкий диапазон. Тем
не менее значения пиковых чисел Рейнольдса — от 2000 до 4000
для легочного ствола и от 2500 до 7000 для аорты — близки к дей-
ствительности. К вопросу о возникновении турбулентности в сосу-
дистом русле мы обратимся позднее. Существование турбулентно-
сти в легочном стволе при нормальных условиях экспериментально
еще не исследовано.
Левые камеры сердца. Изменения давления и потока в легоч-
ных венах и в левом предсердии несколько отличаются от таковых
в венах большого круга и в правом предсердии, но этим вопросом
мы займемся в гл. 14. Здесь же, описывая гидромеханические яв-
ления в левом предсердии и желудочке, легочные вены и левое
предсердие можно рассматривать как резервуар, который напол-
няется во время систолы желудочка и начинает опорожняться в
желудочек в начале диастолы, когда перепад давления на мит-
ральном клапане в результате повышения давления в предсердии
и падения давления в желудочке становится отрицательным. Во
время диастолы скорость течения крови через митральный клапан
быстро растет и может превышать 1 м-с-1. Числа Рейнольдса в
это время соответственно весьма велики (около 8000). Затем ско-
рость спадает до того момента, пока в конце диастолы не возник-
нет кратковременное ускорение, обусловленное сокращением левого
предсердия. После этого, прежде чем клапан закроется и начнется
сокращение желудочка, скорость быстро уменьшается.
Механика закрытия митрального клапана. Экспериментально
достоверно установлено, что, когда в начале систолы желудочек
сокращается, обратный поток крови через митральный клапан
очень мал. Кроме того, при помощи различных способов визуали-
зации движения крови в сердце, а также в исследованиях на моде-
лях показано, что во время обусловленного систолой предсердия
движения крови через митральный клапан в желудочек створки
этого клапана уже движутся по направлению друг к другу, т. е.
клапан начинает закрываться. Ко времени, когда перепад давле-
ния на клапанном отверстии меняет знак, они достаточно сближены
друг с другом, чтобы закрыть отверстие п®чти мгновенно. Этот яв-
ный парадокс первоначально приписывали раннему сокращению
сосочковых мышц, которые притягивают сухожильные нити, а вме-
сте с ними и створки клапана (см. рис. 11.2). Но по ряду причин
это кажется маловероятным, и недавние исследования при помощи
киноангиографии, а также опыты на моделях свидетельствуют
о том, что сосочковые мышцы, видимо, лишь предотвращают вы-
ворачивание клапана и утечку крови обратно в предсердие во
время систолы желудочка, а начало закрытия клапана обеспечи-
вается гидродинамическими механизмами.
В основе механизма, который, как полагают, вызывает раннее
Закрытие митрального клапана, лежит возникновение в желудочке
Гл. 11. Сердце 276
Рис. 11 30 Схема модели левого желу-
дочка, показывающая характер течения
крови во время диастолы. [Bellhouse
(1972) The fluid mechanics of heart val-
ves. Cardiovascular fluid dynamics,
p. 275, Academic Press, New York.]
во время диастолы кольцевого вихря'). Он образуется за створка-
ми клапана, т. е. там, где кровь, прошедшая через отверстие кла-
пана, после отклонения от верхушки желудочка к его стенкам по-
падает как бы в ловушку и вращается (рис. 11.30). В опытах на
моделях установлено, что движение створок клапана по направ-
лению друг к другу зависит от интенсивности этого вихря. Обрат-
ное движение крови через клапан значительно возрастает, если
интенсивность вращения вихря снижена за счет приводящей к его
затуханию задержки сокращения желудочка после прекращения
прямого движения крови через клапан или в результате увеличения
размеров модели желудочка: в этом случае образуется лишь
очень слабый вихрь.
Сущность процессов, приводящих к закрытию клапана именно
в то время, когда через него еще проходит прямой поток крови,
понять нетрудно. Обратимся к рис. 11.31, где схематически пред-
ставлен желудочек, в котором область, занимаемая вихрями, зна-
чительно больше области, заключенной между створками клапана.
Во время почти равномерного движения жидкости через клапан,
которое устанавливается перед замедлением, давление по обе сто-
роны створки, например в точках Р и Q, одинаково, и потому
створки клапана неподвижны. Когда перепад давления на клапан-
ном отверстии уменьшается, скорость движения жидкости через
него снижается, но соответствующие изменения скорости в обла-
сти вихрей оказываются меньше. Это обусловлено как относитель-
но большими размерами этой области, так и инерцией вращаю-
*) Наглядным примером кольцевого (тороидального) вихря служит колечко
дыма Вращение в таком вихре происходит вокруг круговой оси тора —Прим. ред.
276 Часть II. Механика кровообращения
щейся жидкости. В результате давление с наружной стороны створ-
ки клапана, скажем в точке Р, изменяется не так быстро, как
давление с ее внутренней стороны, например в точке Q. Грубо го-
воря, мы можем считать, что давление в области между точками Р
и R — с наружной стороны створок — не изменяется, в то время
как на внутренней стороне, от точки R к точке Q, оно должно па-
дать, чтобы обеспечить наблюдаемое замедление потока. Поэтому
давление в точке Р оказывается больше, чем в точке Q, и клапан
стремится закрыться. Следует подчеркнуть, что это рассуждение
очень грубое и не учитывает многих деталей движения, которые
Рнс 11.31. Схематическое изображение
отверстия митрального клапана и левого
желудочка Во время диастолы кровь из
левого предсердия через клапан движет-
ся в направлении, показанном стрелкой,
со скоростью U. За створками клапана
образуются вихри.
могли бы оказывать влияние на величину разности давлений по
обе стороны створок. Однако основной механизм закрытия кла-
пана оно, вероятно, отображает верно.
Механика закрытия аортального клапана. О строении аорталь-
ного клапана вкратце уже говорилось. Он имеет три створки, при-
крепленные к кольцу фиброзной ткани, которое образует в основа-
нии левого желудочка «выходной тракт». Свободные края каждой
створки начинаются от самых стенок желудочка, и поэтому клапан
может без изменений формы открыть всю площадь поперечного се-
чения выходного тракта. Непосредственно позади каждой створки
клапана в стенке аорты имеется закругленный карман, называе-
мый синусом Вальсальвы. В двух из этих синусов находятся устья
левой и правой коронарных артерий, которые снабжают кровью
мышцу самой сердечной стенки. Створки клапана исключительно
тонки (0,1 мм) и состоят в основном из эластической ткани и кол-
лагена, покрытых слоем эндотелия, подобного тому, который вы-
стилает стенки артерий.
Анатомия легочного клапана почти такая же. Единственное су-
щественное отличие заключается в том, что в синусах Вальсальвы
легочного ствола нет отверстий, открывающихся в коронарные ар-
терии. Как мы видели, абсолютные величины и изменение во вре-
мени давления в легочном стволе и в аорте несколько различаются.
Амплитуды волн кровотока в этих сосудах и изменение их во вре-
мени также различаются. Важны ли эти различия для механики
Гл. 11. Сердце 277
закрытия клапанов — неясно. Исследования на моделях, как и не-
многочисленные опыты in vivo, осуществленные методом визуали-
зации, относятся к аортальному клапану. Поэтому и приведенные
далее сведения касаются в первую очередь этого клапана.
Прямые данные о поведении даже аортального клапана крайне
ограничены. Киноангиография, конечно, подтверждает, что у здо-
ровых животных и у человека аортальный клапан не препятствует
прямому движению крови во время систолы, но она не выявляет
каких-либо деталей поведения створок этого клапана. В одном
или двух исследованиях, в которых использовали высокоскорост-
Рис. 11.32. Линии тока в корне аорты в момент пика систолы. [Bellhouse (1972).
The fluid mechanics of heart valves Cardiovascular fluid dynamics, p 264, Acade-
mic Press, New York ]
иую киносъемку рентгеноконтрастных частиц, сообщалось о круго-
вом движении крови в синусах Вальсальвы. Но существование вих-
ревого механизма, управляющего движением аортального клапана
(хотя это и очень старая идея), пришлось выявлять в основном
в опытах на моделях. Конечно, уже давно было ясно, что движе-
ниями клапана управляет какой-то механизм: регистрация мгно-
венной скорости кровотока в проксимальном отделе аорты (напри-
мер, рис. 11.5, нижняя запись) достоверно показала, что в обычных
условиях во время закрытия клапана обратно в желудочек попа-
дает меньше 5% ударного объема. Кроме того, какое-то количество
йрови поступает во время систолы в коронарные артерии
(рис. 11.20), а это означает, что отверстия коронарных артерий в
это время не закрыты створками аортального клапана.
Исследования на достаточно близких к изучаемым системам
моделях области аортального клапана, осуществленные как мето-
дом непосредственной визуализации, так и путем изучения рас-
пределений давления и скорости в неустановившемся потоке, по-
казали, что во время систолы вихревое движение возникает позади
каждой створки клапана. Когда створки клапана распахнуты,
кромка синусного кармана, расположенная ниже по течению, иг-
рает роль критической точки, т. е. своего рода водораздела: часть
жидкости отделяется от основного потока, заворачивая в синус,
где образуется вихрь (рис. 11.32). Во время увеличения скорости
278 Часть II. Механика Кровообращения
потока через клапан, а также когда величина потока максимальна,
вихревое движение жидкости в синусах удерживает створки кла-
пана в таком положении, что они не закрывают коронарные арте-
рии и в то же время не препятствуют выходу крови из желудочка.
При снижении скорости потока створки клапана плавно сбли-
жаются и окончательно закрываются в конце систолы, допуская
при этом, как и в митральном клапане, очень небольшую утечку
крови обратно в желудочек. Когда такие опыты повторяли на мо-
делях без синусов, т. е. в условиях, когда после клапана жидкость
Рис. 11.33. Линии тока во время закрытия клапана. [Bellhouse, Talbot (1969).
The fluid mechanics of the aortic valve. J. Fluid Meeh, 35, 721. Имеется перевод:
Механика (периодический сборник переводов иностранных статей). — М.: Мир,
1970, № 1, 96—100.]
попадала в трубку в виде простого цилиндра, эффективность ра-
боты клапана значительно снижалась: створки его закрывались
нерегулярно (т. е. не в какой-либо определенный момент времени)
и с опозданием, а обратная утечка жидкости была велика.
Приближенный теоретический анализ действия аортального кла-
пана провел Беллхауз ’). В основе лежат два предположения. Пер-
вое заключается в том, что влиянием вязкости можно пренебречь,
и потому между локальными давлением, скоростью и ускорением
имеется простая связь [обобщающая теорему Бернулли* 2), уравне-
ние (4.6)]. Это предположение вполне оправданно, потому что пол-
ный сердечный цикл длится не более одной секунды, а время, не-
обходимое для того, чтобы вязкость вызвала существенное зату-
хание вихревого движения крови в синусе, составляет около 20 с.
Второе предположение заключается в том, что скорость в вихре
(т. е. интенсивность вихревого движения) прямо пропорциональна
скорости движения крови в аорте сразу за синусами Вальсальвы.
Обосновать это предположение не так-то просто, особенно для не-
стационарного состояния, возникающего при закрытии клапана,
когда, как показывают наблюдения, поток жидкости быстро уст-
ремляется за закрывающиеся створки (рис. 11.33). Часть общего'
потока идет на заполнение расширяющейся полости позади кла-
пана и поддержание там вихревого движения, а остальная часть
образует очень плавное течение вниз по аорте. Однако, приняв
*) Bellhouse В. J. (1972). The fluid mechanics of heart valves, Chapter 8 in
Cardiovascular Fluid Dynamics, ed. D H Bergel, Academic Press
2) Имеется в виду обобщение иа случай нестационарного течения —Прим,
ред.
Гл. 11. Сердце 279
упомянутое предположение, мы сможем показать, что механизм
закрытия этого клапана очень похож на уже описанный механизм
закрытия митрального клапана. Теоретический анализ закрытия
клапана с использованием для предсказания местного давления в
аорте и в синусах реальных значений меняющейся во времени ско-
рости течения крови через клапан и скорости закрытия клапана
приводит, конечно, к результатам, очень хорошо соответствующим
результатам измерений на моделях. Еще одно предположение по-
зволяет довольно просто объяснить и высокую стабильность поло-
жения створок клапана при ускорении крови во время систолы. Это
предположение заключается в том, что интенсивность вихря ме-
няется с изменением размеров области в синусе позади створки.
Если створка вдавливается в синус глубже, чем в нормальных ус-
ловиях (рис. 11.32), то интенсивность вихря снижается. Как сле-
дует из теоремы Бернулли, это означает, что давление за створкой
растет и стремится, следовательно, сдвинуть створку назад к по-
ложению равновесия. То же самое рассуждение справедливо и в
обратном случае, когда створка смещается так, что область, зани-
маемая вихрем, растет. Хотя теоретическое рассмотрение движе-
ния створок клапана основано на весьма сильных дбпущениях, ко-
торые невозможно оправдать априорно, значение вихревого движе-
ния крови для контроля положения створок представляется четко
установленным.
Изгнание крови из левого желудочка. До сих пор ничего не
было сказано о давлении и потоке, создаваемых в аорте левым же-
лудочком при его опорожнении. Подробно об этом будет идти речь
в следующей главе, и тогда станет ясно, что давление и кровоток
в аорте существенно зависят как от условий в периферической
части сосудистого русла, так и от состояния крупных артерий.
Здесь мы очень коротко очертим связь между процессами, проте-
кающими в желудочке и в аорте во время систолического изгнания
крови, рассмотрев для этого баланс сил, действующих на находя-
щуюся в желудочке кровь.
Аортальный клапан открывается, как только давление в желу-
дочке оказывается выше давления в корне аорты. До этого момен-
та желудочек сокращается изоволюмически, но затем объем ле-
вого желудочка (и, возможно, его форма), так же как и давление
крови, контактирующей с его стенками, изменяются во времени.
Сила, которая заставляет кровь вытекать в аорту с ускорением,
обусловлена разностью между давлением, развиваемым сокращаю-
щейся мышцей желудочка, и давлением в аорте. Величина этой
силы приближенно описывается уравнением
Сила изгнания = (рж — ра) А, (11-3)
где рж — давление, развиваемое желудочком при его сокращении,
ре — давление в корне аорты, А— площадь входного отверст 1Щ
аортц.
280 Часть II. Механика кровообращения
Разность давлений в левом желудочке и в корне аорты и изме-
нение ее во времени можно найти из рис. 12.13, на котором пока-
заны записи этих давлений, произведенные одновременно при по-
мощи точных манометров. В начале систолы давление в левом
желудочке быстро повышается, пока не превысит давление в аорте.
Именно в этот момент времени кровь начинает поступать в аорту.
Кровь продолжает приобретать ускорение примерно до середины
систолы, когда записи давлений вновь пересекаются. После этого
давление в аорте превышает давление в желудочке, и сила, дей-
ствующая на кровь во входном отверстии аорты [уравнение (11.3)],
меняет знак и начинает замедлять ее движение. В действительно-
сти движение крови не прекращается и не меняет своего направле-
ния до значительно более позднего момента времени, так как для
того, чтобы действие обратного градиента давления преодолело
инерцию движения крови, приобретенную ею при ускорении в на-
чале систолы, требуется некоторое конечное время. Таким обра-
зом, ускорение совпадает по фазе с градиентом давления, тогда
как скорость кровотока отстает от градиента давления. Когда кро-
воток действительно меняет свое направление, аортальный клапан
закрывается, что отмечается на записи аортального давления
(рис. 12.13) зубцом (дикротический зубец). После этого, естест-
венно, сообщение между желудочком и аортой прекращается и вы-
ражение (11.3) для силы, действующей на кровь, теряет смысл.
Однако если мы рассматриваем период, когда клапан открыт,
и считаем, что влиянием вязкости можно пренебречь (это допу-
стимо, так как длительность систолы недостаточна для образова-
ния вязких пограничных слоев, которые могли бы оказать значи-
тельное сопротивление движению), то сила, обеспечивающая из-
гнание крови из желудочка [уравнение (11.3)], в любой момент
времени должна быть равна сумме двух составляющих:
1) скорости, с которой количество движения выносится из же-
лудочка;
2) скорости изменения количества движения крови, находя-
щейся в желудочке.
[Это утверждение представляет собой просто второй закон Ньюто-
на, выраженный через количество движения; такая формулировка
закона рассмотрена в гл. 2, уравнение (2.9).]
Величину первой из этих двух составляющих можно оценить,
рассмотрев движение крови, изгоняемой из желудочка за малый
промежуток времени б/ (рис. 11.34). Масса этой крови равна
рАи^Ы, где р — плотность крови, иа — мгновенная скорость крови
в корне аорты. Количество движения (масса X скорость) изгоняе-
мой крови составляет, следовательно, (рЛиа-б/)иа, и это есть ко-
личество движения, выносимое из желудочка за время б/. А ско-
рость, с которой количество движения выносится из желудочка
(т. е. само это количество, деленное на б/), есть рАи*. Во время
Систолы эта реличина непрерывно меняется.
Гл. 11. Сердце 281
Вторая составляющая обусловлена тем, что количество дви-
жения крови, остающейся в желудочке, также непрерывно ме-
няется. Это количество движения равно сумме количеств движения
всех малых элементов крови в желудочке, движущихся по на-
правлению к аорте. Чтобы точно его рассчитать, необходимо знать
форму желудочка и скорость изменения его объема во время фазы
изгнания, однако мы можем предсказать, что оно будет равно
массе крови в желудочке рУ, где V — мгновенный объем желудоч-
ка, умноженной на скорость изгнания wa и на коэффициент, учиты-
вающий изменение формы, который мы обозначим через /. Таким
Левый
желудочек
Рис. 11.34. Схематическое изображение левого желудочка и аорты во время си-
столы. За короткий отрезок времени б/ кровь в аорте (движущаяся со скоростью
иа) проходит расстояние «а-б/.
образом, скорость изменения количества движения крови, нахо-
дящейся в желудочке, будет d(IpVua)/dt. Если бы форма желу-
дочка во время сокращения не изменялась, то коэффициент / был
бы постоянным. Поскольку в действительности форма желудочка
меняется, этот коэффициент, вероятно, тоже меняется, но в каж-
дом случае его можно в принципе вычислить.
Итак, мы можем записать следующее соотношение, справедли-
вое, пока аортальный клапан открыт, в любой момент времени:
Рж Ра ___ 2 I__L (^иа) /1 | л\
р A At ’
Имеется еще одно, более очевидное соотношение между объе-
мом желудочка V и скоростью крови в аорте: поскольку кровь не-
сжимаема, скорость, с которой она вытекает в аорту, должна быть
равна скорости изменения объема желудочка, т. е.
(11.5)
^Ыа ~ а/ •
Именно эти уравнения связывают давление в желудочке и его
объем с давлением и скоростью движения крови в аорте. В прин-
282 Часть II. Механика кровообращения
ципе они дают способ исследования деятельности желудочка по
данным изменения давления и потока в аорте. Если, например, мы
знаем величины I и А, то по результатам измерения скорости кро-
вотока и давления в аорте (с появлением зондовых датчиков такие
измерения удается проводить даже в клинике) можем получить
представление об изменениях систолического давления в желудочке
и его объема. Однако на практике здесь возникают большие труд-
ности. Например, величина I не определена ни для какой формы
желудочка, и изменчивость ее in vivo не изучена. Кроме того, пол-
ный объем желудочка V из уравнения (11.5) найти невозможно,
если неизвестен конечно-диастолический объем, который заведомо
трудно измерить с достаточной точностью. Пока подход, основан-
ный на соотношениях (11.4) и (11.5), не разработан ни теорети-
чески, ни экспериментально.
Наконец, следует указать, что все это описание касается только
сил, относящихся к движению крови из желудочка, и не учитывает
очень высокого давления (диастолическое давление в аорте), ко-
торое должен преодолеть желудочек, чтобы открыть аортальный
клапан и положить начало движению крови. При рассмотрении
всей механической деятельности желудочка диастолическое давле-
ние в аорте выступает как один из доминирующих факторов. Это
можно видеть из рис. 12.13: максимальное давление, которое раз-
вивает желудочек, превышает давление в аорте на величину, очень
малую по сравнению с абсолютными уровнями этих давлений.
Тоны и шумы сердца. Тоны. Общепринятое представление о том,
что нормальные тоны сердца возникают при закрытии клапанов,
существует уже много лет. Однако в объяснении лежащих в ос-
нове этого механизмов так много недоразумений и путаницы, что
имеет смысл рассмотреть эти механизмы детально.
В качестве примера выберем митральный клапан и, следова-
тельно, первый тон сердца. Мы видели, что к концу диастолы
створки клапана уже сближены друг с другом. Последняя стадия
закрытия клапана вызывается обратным движением крови из же-
лудочка в предсердие. Когда клапан закрывается, кровь, находя-
щаяся вблизи него, в силу инерции продолжает свое движение по
направлению к предсердию и растягивает клапан. Поскольку створ-
ки клапана упруги, они после растяжения стремятся вернуться
назад, приводя кровь в движение в противоположном направлении.
Клапан вновь проскакивает положение равновесия, и процесс по-
вторяется. Эти колебания продолжаются до тех пор, пока не про-
изойдет их затухания из-за обусловленных вязкостью потерь энер-
гии в крови и в самих створках клапана.
Поскольку разность давлений по обе стороны клапана увели-
чивается по мере повышения давления в желудочке, положение
равновесия клапана в ходе его колебаний, конечно, смещается.
В результате процесс колебаний становится несколько более слож-
ным, поскольку упругие свойства створок клапана нелинейны (как
Гл 11 Сердце 283
и в случае артерий, и, вероятно, по той же причине — см. гл. 12 —
их жесткость увеличивается при растяжении) Тем не менее этот
механизм является главным в возникновении высокочастотных ко-
лебаний давления в предсердии и в желудочке, и именно эти ко-
лебания лежат в основе генерации первого тона сердца.
Бытовавшие в прошлом ошибочные суждения относительно
этого механизма имели, по-видимому, два источника. Во-первых,
неправильно толковали физические процессы (например, полагали,
что изменение знака разности давлений между предсердием и же-
лудочком мгновенно изменяет направление движения крови; на
самом же деле очень важным свойством рассматриваемого потока
крови является инерция, и такой поток не может изменить направ-
ление своего движения мгновенно). Во-вторых, несовершенство из-
мерительных приборов приводило к искажению наблюдаемых явле-
ний и неправильному их толкованию1). Самое бесспорное доказа-
тельство того, что источником тонов сердца являются колебания его
клапанов, дает одновременная регистрация звуковых колебаний в
левом предсердии и в левом желудочке при помощи двух микро-
фонов. Такая регистрация показывает, что колебания, обусловли-
вающие первый тон, в этих двух камерах противоположны по
фазе. Объяснить этот факт можно, по-видимому, только колеба-
тельным движением клапана.
Все это не означает, что другие процессы не принимают ника-
кого участия в генерации тонов сердца. Например, флуктуации
давления крови приводят в движение стенки сердца, в результате
чего, как показано в гл. 12, по ним распространяются упругие
волны (от аортального и легочного клапанов эти волны распро-
страняются также и по артериям). Все эти колебания в совокуп-
ности перекрывают широкий спектр частот. Если относительно
высокочастотные составляющие колебаний (превышающие по ча-
стоте 50 Гц) достигают поверхности грудной клетки, их удается
услышать через стетоскоп. Кроме того, их можно зарегистрировать
точными приборами, такими, как расположенные на кончике зонда
датчики давления или микрофоны. Низкочастотные составляющие
(вероятно, до 20—30 Гц) регистрируют при помощи обычных зон-
довых датчиков давления. В случае, например, закрытия аорталь-
’) Одним из особенно уместных здесь примеров является форма искажений,
называемая «звоном» Такие искажения могут встретиться при измерении давле-
ния обычной системой зонд-манометр Они представляют собой высокочастотные
колебания в измерительном устройстве (которые можно ошибочно принять за
колебания, происходящие в желудочке) Механизм возникновения «звона» в точ-
ности совпадает с механизмом возникновения сердечных тонов Здесь также
имеется упругая мембрана (диафрагма манометра и стенки зонда), связанная
с массой жидкости, находящейся в зонде, и когда эта масса резким изменением
давления на конце зонда приводится в движение, диафрагма смещается, при об-
ратном движении она проскакивает положение равновесия, и в системе возникают
кратковременные колебания Демпфирование и в этом случае обусловлено вяз-
кими свойствами жидкости в зонде.
284 Часть II. Механика кровообращения
него клапана низкочастотные колебания регистрируются на запи-
сях давления в аорте, и их изображение называют дикротическим
зубцом (рис. 11.5), а высокочастотные прослушиваются через стен-
ку грудной клетки с помощью стетоскопа и дают аортальный ком-
понент второго тона сердца. Легочный компонент образуется точно
так же, но несколько позднее, так как фаза изгнания крови из пра-
вого желудочка длится дольше, чем из левого (рис. 11.5 и 11.6).
Хотя диагностическое значение тонов сердца очень велико,
о механизме их распространения известно мало. Сразу же поясним,
что в истинно акустическом смысле тоны сердца вовсе не являются
звуками. По определению акустические звуковые волны — это вол-
ны сжатия, которые распространяются внутри самого материала
и сопровождаются изменениями его плотности. Скорость распро-
странения их в жидкостях очень велика (в крови — около
1500 м-с-1). Колебания, передаваемые стенкой грудной клетки
диафрагме фонендоскопа или раструбу стетоскопа, вызывают вол-
ны такого типа в воздухе, который находится в трубке стетоскопа,
и поэтому доходят до уха как истинный звук *). Но они не рас-
пространяются в такой форме от места их происхождения до стеи-
ки грудной клетки. Показано, что тоны сердца распространяются
по сосудам (таким, как аорта) и через ткани к стенке грудной
клетки со скоростью, составляющей менее одной сотой скорости
распространения в крови акустических звуковых волн. Например,
по аорте второй тон сердца распространяется со скоростью пуль-
совой волны (около 5—10 м-с-1) и быстро затухает (т. е. «остана-
вливается»), если стенка сосуда в данной области закреплена и
не может двигаться в радиальном направлении. Таким образом,
второй тон сердца распространяется по упругому сосуду как волна
давления точно так же, как и пульсовая волна, и является фак-
тически частью ее, хотя составляющие тон частоты лежат вне
диапазона частот, обычно регистрируемых датчиками давления.
При прохождении по сосудам второй тон сердца очень быстро за-
тухает: на расстоянии от клапана 20 см и больше в сущности уже
ничего не обнаруживается.
Показано, что первый тон сердца (т. е. колебания давления,
возникающие при закрытии митрального клапана) распростра-
няется по стенке желудочка точно таким же образом — в виде
волны, движущейся к верхушке сердца, а затем, после отражения,
*) В упругих материалах и конструкциях могут распространяться волны
деформации самых разных типов, в том числе н такие, при которых не происхо-
дит объемной деформации и, стало быть, колебаний плотности. Например, рас-
пространение пульсовых волн по сосудам ие сопровождается изменениями плот-
ности крови и стенки сосудов, которые практически несжимаемы. Однако, когда
деформационные колебания, частота которых лежит в диапазоне истинных зву-
ковых волн, достигают поверхности тела, они приводят к возникновению в воз-
духе истинных звуков, которые и улавливает ухо врача. См. также конец разд.
12.7.— Прим. ред.
Гл. 11. Сердце 285
обратно приблизительно с той же скоростью. Это движение про-
исходит так же, как описано в гл. 12 для волн давления в арте-
риях. Наконец, показано, что оба тона сердца могут распростра-
няться о г места их возникновения как вверх по течению крови, так
и вниз. Например, первый тон проходит от митрального клапана
и через левое предсердие, и через левый желудочек, а второй тон
(от аортального клапана) — как по стенке левого желудочка, так
и по аорте. Скорость затухания этих волн зависит от их частоты
и от жесткости стенки, по которой они распространяются: если
волны распространяются по относительно «свободно висящей»
стенке или если их частота высока, то они затухают на более ко-
ротком расстоянии.
Влияния частоты колебаний и жесткости стенки на затухание
связаны друг с другом через скорость волны. Зависимость от ча-
стоты объяснить довольно просто, потому что затухание волн та-
кого типа характеризуется постоянной величиной, отнесенной к
длине волны (см. гл. 12). Таким образом, волны с более высокой
частотой (с меньшей длиной волны) исчезают, проходя меньшее
расстояние (и это занимает также меньшее время). Например,
если волна распространяется по аорте или назад и вперед по
стенке желудочка, она может иметь скорость 5 м-с-1 и затухать
на 50% на пути в одну длину волны. Тогда при прохождении рас-
стояния, равного 5 длинам волны, ее амплитуда уменьшится при-
мерно на 97%. Для волны с частотой 100 Гц это расстояние со-
ставит 25 см при времени прохождения 50 мс. Для волны с часто-
той 50 Гц как расстояние, так и время такого затухания будут в
2 раза больше. Итак, если колебания создаются быстропротекаю-
щими процессами, подобными закрытию клапана, то высокоча-
стотные составляющие их быстро затухают, в чем можно убе-
диться, наблюдая локально затухание при многократном отраже-
нии или прослеживая волну на некотором определенном пути, ска-
жем в аорте.
Влияние жесткости стенки на затухание обусловлено тем, что
скорость волны зависит от жесткости (см. гл. 12) и уменьшается,
когда стенка становится более «свободной». Поэтому в провисаю-
щей трубке длина волны, имеющей частоту 100 Гц, будет меньше
и такие волны будут затухать на меньшем расстоянии (но не за
более короткое время), чем в рассмотренном выше случае.
Если мы отвлечемся от сосудистых структур как таковых и пе-
рейдем к рассмотрению способа передачи этих волн к стенке груд-
ной клетки, то окажемся перед гораздо менее понятной ситуацией.
Предполагается, что сердце и сосуды вызывают колебания стенки
грудной клетки либо благодаря прямому контакту, либо через
другие структуры, расположенные между ними. Тщательная «кар-
тография» стенки грудной клетки при помощи большого числа
микрофонов показала, что каждый тон сердца обнаруживается сна-
чала в определенной точке (которая в каждом случае хорошо соот-
286 Часть И. Механика кровообращения
ветствует принятой в клинике классической точке прослушивания
стетоскопом) и затем, быстро затухая, распространяется по стенке
во все стороны. Скорость распространения при этом также значи-
тельно меньше скорости распространения истинной звуковой вол-
ны, и высокочастотные составляющие затухают гораздо быстрее,
чем низкочастотные. Однако, кроме указанных фактов, о физиче-
ской природе этих волн ничего не известно.
Шумы. Большая часть приведенных выше рассуждений отно-
сится также и к колебаниям, которые соответствуют прослушивае-
мым шумам, в том смысле, что и они распространяются по струк-
турам грудной полости относительно медленно, появляются сначала
в определенных точках стенки грудной клетки, а их высокочастот-
ные составляющие при распространении быстро затухают. Однако
если происхождение тонов сердца как следствие колебаний клапа-
нов четко доказано, то физический источник шумов точно неизве-
стен. Согласно классическому представлению, их источником яв-
ляются колебания давления, сопровождающие появление турбу-
лентности, а последняя может возникать при одном из трех основ-
ных условий: если патологически изменен клапан, если клапан не
изменен, но расширена камера (образование струи) и, наконец,
если скорость движения крови в области клапана чрезвычайно вы-
сока (при нормальном клапане и размере камеры). Осуществить
удовлетворительно визуализацию движения крови в сердце или
прямые измерения, которые могли бы обнаружить турбулент-
ность,— дело чрезвычайно сложное, и потому рассуждения о шу-
мах носили по большей части умозрительный характер. Несом-
ненно, существуют и другие типы возмущений течения крови, ко-
торые могли бы действовать как источник звука. Вопрос о турбу-
лентности в сосудистой системе будет рассмотрен более подробно
при обсуждении движения крови в крупных артериях (гл. 12).
Здесь же достаточно сказать, что прямое сопоставление частотного
спектра шумов и флуктуаций скорости движения крови дает до-
статочные основания полагать, что источником шумов действи-
тельно является турбулентность. Остается, однако, много неясно-
стей относительно источников других звуков, возникающих в
сердце при заболеваниях. В настоящее время нельзя с уверенно-
стью сказать, благодаря каким физическим явлениям возникают
третий и четвертый тоны сердца — щелчки при изгнании крови
и треск при открывании клапанов.
СПИСОК ЛИТЕРАТУРЫ
Bergel D. Н. (ed.). (1972). Cardiovascular fluid dynamics, Chapter 7: The meaning
and measurement of myocardial contractility, by J. R. Blinks, B. R. Jewell;
Chapter 8: The fluid mechanics of heart valves, by B. J. Bellhouse, Academic
Press, London and New York. (2 vols).
Challice С. E., Viragh S. (eds). (1973). Ultrastructure in biological systems,
Vol. 6: Ultrastructure of the mammalian heart, Academic Press, London and
New York,
Гл. 11. Сердце 287
CIBA Foundation Symposium (1974). New Series No. 24, The physiological basis
of Starling’s law of the heart, Associated Scientific Publishers, Amsterdam.
Fung Y. C., Perrone N., Anliker M. (eds). (1972). Biomechanics: its foundations and
objectives, Chapter 12; Mechanics or contraction in the intact heart, by
J. W. Covell; Chapter 13; Determinants of cardiac performance, by C. Urschel,
E. H. Sonnenblick, Prentice-Hall, New Jersey.
Hamilton W. F. (ed.). (1962). Handbook of physiology, Section 2: Circulation,
Vol. 1: several chapters on different aspects of cardiac physiology, American
Physiological Society, Washington, D. C.
Hill A. V. (1970). First and last experiments in muscle mechanics, Cambridge
University Press. (Хилл А. Механика мышечного сокращения. — M.: Мир.
Taylor М. G. (1973). Haemodynamics, Ann. Rev. Physiol., 35, 87—116.
Глава 12
АРТЕРИИ БОЛЬШОГО КРУГА
В этой главе мы рассмотрим механизмы движения крови в круп-
ных артериях большого круга. Артерии малого круга обладают
особыми свойствами, и им посвящена заключительная глава книги.
Таким образом, предметом обсуждения будут здесь аорта и ее
ветви — сосуды, доставляющие богатую кислородом кровь к орга-
нам и тканям тела. Как и ранее, основным примером послужит со-
судистая система собаки, так как она изучена наиболее полно;
однако мы -будем обращаться и к сосудистой системе человека,
если возникнет необходимость подчеркнуть особенности ее функций
или строения. Не касаясь, как и прежде, активных физиологиче-
ских процессов — механизмов рефлекторного сужения сосудов, спо-
собных приводить к изменениям кровотока или распределения
крови, мы сосредоточим внимание на тех физических свойствах
изучаемой системы, которые изменяются в результате действия
этих активных процессов.
Материал, посвященный артериальной системе большого круга,
разбит нами на две части: артерии рассмотрены в данной главе,
а система микрососудов — в гл. 13. Поэтому мы должны прежде
всего объяснить причины и принцип такого разделения; затем мы
дадим краткое описание анатомической организации и гистологи-
ческого строения артерий большого круга, а также особенности
картин давления и потока крови в них. Далее мы привлечем для
рассмотрения этих особенностей определяющие их основные прин-
ципы механики и, вводя шаг за шагом все более сложные детали,
подойдем к полному описанию картин давления и течения крови
в артериях. В ходе изложения мы будем многократно обращаться
к основам механики, о которых шла речь в первой части книги.
По традиции физиологи подразделяют сосуды большого круга
на крупные и мелкие, условно выбирая границу между ними на
уровне самых мелких артерий — артериол. Как будет показано в
гл. 13, к такому подразделению прибегают в силу различных сооб-
ражений, опираясь иа доводы как физиологии, так и механики.
Здесь нам представляется наиболее удобным и разумным объяс-
нить это деление, опираясь на основное различие гидромеханиче-
ских свойств течения крови в крупных и мелких сосудах. Им яв-
ляется, как это уже говорилось в гл. 5, различие между тече-
ниями, в которых преобладает действие инерционных сил, и вяз-
Гл 12. Артерии большого круга 289
ким течением, при котором роль инерции движения жидкости пре-
небрежимо мала. Относительное значение инерционных и вязких
сил характеризуют числом Рейнольдса Re, равным
и, таким образом, режим течения зависит от скорости течения U,
кинематической вязкости жидкости v и диаметра сосуда d. На-
помним (гл. 5), что свойства течений с числами Рейнольдса, много
большими единицы, определяются в основном инерционными си-
лами, тогда как особенности течения с числами Рейнольдса, много
меньшими единицы, — главным образом вязкими силами. В этой
главе мы будем иметь дело-с течениями, характеризующимися
большими числами Рейнольдса, т. е. с такими течениями, которые
почти полностью определяются инерционными силами. По суще-
ству, таково течение крови лишь в крупных сосудах (до мелких
артерий, диаметр которых равен примерно 100 мкм и в которых
число Рейнольдса становится меньше единицы), поскольку после-
довательные ветвления артерий приводят к сильному увеличению
площади поперечного сечения сосудистого русла, и поэтому по мере
перемещения крови в мелкие артерии скорость ее движения умень-
шается1). Гл. 13 охватывает механику движения крови в сосудах
малого диаметра, в которых преобладают силы вязкого трения
Важно осознавать, что, поскольку числа Рейнольдса зависят от па-
раметров, которые могут меняться в широких пределах, граница
раздела между течениями с высокими и низкими числами Рей-
нольдса в сосудистой системе подвижна. Представим себе, напри-
мер, что поток крови через какой-либо орган, скажем работающую
мышцу, повысился вследствие расширения ее мелких сосудов и
роста скорости кровотока в них; тогда на каждом из уровней со-
судов мышцы возрастет число Рейнольдса и граница раздела сме-
стится в сторону капилляров. Сужение сосудов органа приведет
к обратному явлению.
12.1. Анатомическая организация и гистологическое строение
Анатомия крупных кровеносных сосудов. Анатомическая схе-
ма аорты и ее главных ветвей у собаки представлена на рис. 12,1,
а ряд характерных размеров указан в табл. I. Аорта выходит из
левого желудочка и почти сразу над аортальным клапаном раз-
ворачивается примерно на 180° от своего первоначального направ-
’) Заметим, что вязкость крови в крупных сосудах вполне можно считать
постоянной величиной, так как, во-первых, диаметр этих сосудов много больше
диаметра индивидуальных клеток крови и, следовательно, кровь здесь можно
рассматривать как сплошную жидкую среду, а, во-вторых, скорость сдвига в
этих сосудах достаточно велика, чтобы вязкость была независящей от нее
(см. гл. 10).
290 Часть II Механика кровообращения
Позвоночная
ft
Восходящая аорта
Крестцовая
Бедренная
Большеберцовая
большая
лодкожная
Подвздошная
Уровеиь
_"пупартовой
связки
Брюшная аорта
----Почечная
=Поясничная
Правая подключичная
Плечеголовная
Подкрыльцовая
-Левая подключичная
—Дуга аорты
Грудная аорта
Межреберная
Грудная аорта
Чревная артерия
Передняя брыжеечная
Задняя брыжеечная
Диафрагма
Рис 12 1 Схема главных ветвей артериального дерева собаки. [McDonald D. А.
(1974). Blood flow in arteries Edward Arnold, London]
Гл 12 Артерии большого круга 2gj
Рис 122. Слепок аорты собаки, полученный при физиологическом значении дав.
ления: А. Вид спереди Б Внд слева На слепке видны три синуса Вальсальвы и
отходящие от аорты коронарные артерии Рисунок показывает также сложный
изгиб аорты в двух плоскостях Определить углы, под которыми от аорты отхо.
дят плечеголовная и левая подключичная артерии, трудно из-за сильного изгиба,
ния этих сосудов сразу по выходе из аорты.
ления, образуя дугу. От дуги аорты отходят ветви к сердцу, голове
и передним конечностям (рис. 12.2). Продолжаясь в виде прямого
ствола, аорта проходит сквозь диафрагму в брюшную полость и
посылает ветви к расположенным здесь органам. Заканчиваясь в
брюшной полости, аорта делится на две подвздошные артерии,
питающие задние конечности, и среднюю крестцовую, снабжающую
хвост (напомним, что речь идет о собаке).
По мере удаления от сердца аорта конусообразно суживается
Степень ее «конусности», по-видимому, различается как для жи.
вотных разных видов, так и для разных особей одного вида. Для
собак изменение площади поперечного сечения аорты по длине
достаточно хорошо описывается экспоненциальной функцией вида
А = Аое~Вх1в°,
где А — площадь поперечного сечения аорты в данном месте, Ло
и До — площадь и радиус аорты в некотором другом ее месте вверх
292 Часть П. Механика кровообращения
по течению, х—-расстояние между двумя указанными сечениями,
В — «коэффициент конусности», который, согласно результатам
измерений, равен 0,02—0,05. Таким образом, «конусность» аорты
собаки весьма значительна, что видно из рис. 12.3, Л, который по-
строен по результатам измерений, выполненных на посмертных
слепках аорты собак. Значение В, соответствующее степени конус-
ности аорты на этом рисунке, составляет около 0,033. Согласно
имеющимся данным, конусообразное сужение аорты человека не
столь плавно, как это следует из приведенной выше формулы.
Коэффициенты и углы ветвления. Хотя аорта и все дру-
гие сосуды конусообразно сужаются, общее поперечное сечение со-
судов артериального русла по мере удаления от сердца возрастает.
По ряду причин количественные данные, на которые обычно ссы-
лаются как на подтверждающие это положение, скорее всего не-
точны: в частности, измерения не всегда производились при фи-
зиологических значениях давления, и на их результатах могла
сказываться усадка тканей, вызванная применением химических
фиксирующих средств. Прижизненные измерения у животных, есте-
ственно, сопряжены с большими трудностями, и поэтому наши зна-
ния о размерах артерий даже для представителей одного вида до
сих пор весьма неполны. Тем не менее имеются достаточно точные
сведения о размерах ветвей аорты собаки, полученные как путем
прямых прижизненных измерений, так и посредством измерений
посмертных слепков, зафиксированных при физиологических зна-
чениях давления в артериях. Аортальное дерево некой «средней»
собаки изображено на рис. 12.3, Б на основе средних данных для
десяти довольно крупных собак.
Диаметр ветвей аорты в общем хорошо соответствует расходу
крови в них (исключение составляют только почечные артерии:
если учесть, что через них протекает 20—25% минутного объема
крови, они представляются непропорционально узкими). Однако,
рассчитав коэффициенты ветвления (их вычисляют как отношение
суммы площадей поперечного сечения ветвей, отходящих от об-
щего для них сосуда, к площади поперечного сечения этого по-
следнего), мы обнаружим, что они значительно варьируют — от
0,79 до 1,29. Если коэффициент ветвления больше 1, это означает,
что суммарный просвет артерий стал больше, и напротив, значение
коэффициента, меньшее 1, свидетельствует о том, что он стал
меньше.
Для ветвей грудной аорты коэффициент ветвления близок к
единице, тогда как для ветвей начального отдела брюшной
аорты он несколько больше единицы, и это означает, что здесь
просвет артерий становится шире. Однако это расширение почти
полностью компенсируется сужением суммарного просвета тех ар-
терий, которыми заканчивается аорта (коэффициент ветвления
0,85—0,90). Такое же сужение просвета конечных ветвлений аорты
обнаружено и у человека. В целом изменение суммарной площади
Гл. 12. Артерии большого круга 293
Рис. 12.3. А. Конусообразное сужение аорты. Средние значения наружного радиуса
аорты в разных ее участках рассчитаны по результатам измерении иа десяти
собаках при физиологических значениях давления. [Fry, Griggs jr., Greenfield jr.
(1963). In vivo studies of pulsatile blood flow. В кн. Pulsatile blood flow (ed.
Atfinger), p. ПО. С любезного разрешения McGraw-Hill Book Company, New
York.] Б. Площадь поперечного сечення (в см2) различных участков аорты и ее
главных ветвей (измерения произведены прн тех же условиях, что и ддя
рис. 12.3, A). [Patel, de Freitas, Greenfield jr., Fry (1963). Relationship of radius
to pressure along the aorta in living dogs. J. appl. physiol., 18, 1111—1117.]
294 Часть П Механика кровообращения
поперечного сечения крупных артерий незначительно: площадь се-
чения ветвей аорты, показанных на рис. 12.3, Б, лишь на 7% пре-
вышает площадь сечения восходящей аорты. Согласно другим экс-
периментальным данным, увеличение суммарной площади сечения
ветвей аорты значительно больше — до 60%; такой разброс дан-
ных обусловлен тем, что трудно точно поддерживать физиологиче-
ский уровень давления при изготовлении слепков путем заполнения
сосудов затвердевающей массой.
Несомненно, однако, что по мере ветвления артерий, отходя-
щих от аорты, суммарная площадь артериального дерева начинает
очень резко возрастать. Наиболее подробное прижизненное изме-
рение размеров сосудов произведено на крыле летучей мыши: здесь
их удается непосредственно наблюдать под микроскопом в прохо-
дящем свете. Результаты этого исследования приведены в
табл. 12.1. Конечно, размеры всех сосудов, кроме капилляров и
ближайших к ним микрососудов, у такого маленького животного
уменьшены соответственно размеру тела — диаметр даже аорты
у летучей мыши составляет всего лишь около 1 мм, но тем не ме-
нее увеличение суммарной площади поперечного сечения сосудов
по мере их последовательных ветвлений поразительно.
На животных других видов столь детальных прижизненных из-
мерений не производили, и сведения о размерах сосудов собаки
приходится собирать из многих источников, причем в основном это
данные для посмертного состояния, подобные уже упомянутым дан-
ным по слепкам крупных артерий. Поэтому общая картина, пред-
ставленная на рис. 12.4, менее точна. Все же она сходна с резуль-
татами, полученными для крыла летучей мыши. Таким же обра-
зом увеличивается суммарная площадь поперечного сечения сосу-
дов артериального русла у человека. Прижизненное исследование
ветвления артерий у человека возможно во время хирургических
вмешательств или при диагностической рентгенографии сосудов
после введения в кровь контрастного вещества (артериография)
Именно таким образом получен снимок, приведенный на рис. 12.5,
где виден ход внутренней сонной артерии и ее ветвей в черепе
у человека.
Углы, под которыми крупные артерии отходят от аорты, разли-
чаются в неменьшей степени, чем коэффициенты ветвления, и ясно,
что центральное артериальное дерево несимметрично; симметрия
отсутствует как для углов и коэффициентов ветвления, так и для
длин сосудов. Это следует помнить при количественном описании
поведения артерий. Симметричное ветвление крупных артерий —
явление исключительное; у человека симметрична лишь бифур-
кация аорты — область ее разветвления на две подвздошные арте-
рии, а у собаки, как видно из рис. 12.1, вообще нет симметричных
ветвлений. Особенности течения крови в бифуркациях артерий
рассмотрены в разд. 12.8.
Таблица 12.1
Размеры кровеносных сосудов крыла летучей мыши (прижизненное измерение) ’)
Сосуд Средняя длина, мм Средний диаметр, мкм Среднее число ветвей Число сосудов Суммарная площадь поперечного сечения, мкм2 Объем X ю+3, мм’ Процент от общего объема
Артерия 17,0 52,6 12,3 1 2 263 г) 38,4 10,1
Мелкая артерия 3,5 19,0 9,7 12,3 4 144 14,4 3,8
Артериола 0,95 7,0 4,6 119,3 5101 4,7 1,2
Капилляр 0,23 3,7 3,1 3) 548,7 6548 1,5 0,39
Посткапилляриая вену- ла 0,21 7,3 1727,0 78233 16,4 4,3
Венула 1,0 21,0 5,0 345.4 127 995 127,9 33,7
Мелкая вена 3,4 37,0 14,1 24,5 27 885 94,7 25,0
Вена 16,6 76,2 24,5 1 4 882 81,0 21,4
') По Wledeman (1963) Clrculat Res , 12, 376
’) Получено усреднением площадей поперечного сечения отдельных сосудов.
•) Результат вычислений
296 Часть П Механика кровообращения
Рис. 12.4. Схема увеличения площади поперечного сечення сосудистого русла по
мере удаления от сердца. Разброс результатов не является следствием ошибок
измерений, а отражает тот факт, что сосуды всех уровней имеют различные диа-
метры и коэффициенты конусности. [Данные собраны из многих источников.
Iberall (1967). Anatomy and steady flow characteristics of the arterial system.
Math. Biosci, 1, 375—395.]
Более мелкие артерии достаточно часто образуются путем ди-
хотомического деления общего ствола, причем углы ветвления до-
статочно малы. Конечно, это не общее правило: организация со-
судистой сети некоторых органов высоко специализирована и уни-
кальна. В большинстве органов существуют анастомозы, т. е. со-
суды, соединяющие различные артериальные ветви. Такие ана-
стомозы иногда развиваются, образуя-пути для крови в обход за-
купоренного или поврежденного сосуда. Одну из наиболее важ-
ных систем анастомозов представляет виллизиев круг — особое
артериальное кольцо у основания головного мозга, которое объ-
единяет системы нескольких артерий и посылает ветви к различ-
ным отделам головного мозга. (Назван в честь английского врача
Т. Willis, который в 1664 г. опубликовал иллюстрированный Кри-
стофером Ренном и пользовавшийся признанием трактат по ана-
томии нервной системы.)
Строение артериальной стенки. Все артерии (а также вены,
см. разд. 14.1) имеют общий план строения и образованы из одних
и тех же материалов, хотя соотношения между компонентами сте-
нок различных сосудов неодинаковы. Рассмотрим сначала гистоло-
гическое строение артериальной стенки, а затем обсудим функ-
Гл. 12. Артерии большого круга "297
Рис. 12.5. Рентгенограмма черепа человека (вид сбоку), сделанная сразу после
введения в сонную артерию рентгеноконтрастного вещества Стрелкой указана
внутренняя сонная артерия Внутричерепной ее участок короток и извилист; да-
лее она участвует в образовании виллизиева круга, от которого отходят много-
численные ветви, снабжающие кровью головной мозг.
циональное значение различных ее слоев и компонентов, а также
некоторые возрастные изменения.
Артериальная стенка состоит из трех оболочек: внутренней
(интима), средней (медия) и наружной (адвентиция). Для каждой
из этих оболочек характерны преобладающие структурные эле-
менты и особые виды клеток. Важнейшие особенности строения
артериальной стенки показаны на рис. 12.6, на котором представ-
лена микрофотография поперечного среза брюшной аорты морркой
свинки.
Внутренняя оболочка состоит из двух слоев. Самый глубокий,
соприкасающийся с кровью слой — эндотелий. Он представляет
собой один ряд клеток, которые в виде непрерывной выстилки по-
крывают всю поверхность сердечно-сосудистой системы, соприка-
сающуюся с кровью, — внутреннюю поверхность артерий, капилля-
ров, вен, клапанов и полостей сердца. Эндотелиальный слой меха-
нически очень непрочен и легко повреждается, например воздей-
ствием высокого напряжения сдвига. Однако, обладая значитель-
ной способностью к регенерации и росту, Эндотелий постоянно во-
зобновляется. Разрастаясь, он способен покрывать оживляемые
298 Часть Н Механика кровообращения
Рис 126 А Фазово-контрастиая микрофотография продольного среза брюшной
аорты морской свинки Изображена вся стенка небольшого участка аорты. 1 —
просвет, 2 — эндотелий, 3 — внутренняя эластическая мембрана, 4 — средняя обо-
лочка, 5 — наружная оболочка Это достаточно типичный пример артерии мы-
шечного типа Четко видны пластины средней оболочки, состоящие нз слоев
гладкомышечных клеток, разделенных более темными н тонкими пластами кол-
лагеновых н эластических волокон Б Электронная микрофотография того же
препарата, что и на рнс 12,6, Л, при большом увеличении Обратите внимание
на слой эндотелиальных клеток и светлую соединительную ткань, окружающую
гладкие мышцы внутреннего слоя В наружном слое на нижней части среза за-
метны интенсивно окрашивающиеся фибробласты н коллагеновые волокна.
[Любезно предоставлена д-ром G Gabel 1а ]
хирургами протезы сосудов или искусственные клапаны сердца
независимо от того, изготовлены ли такие протезы из живых тка-
ней или из синтетических материалов. Строение эндотелиальных
клеток подробно рассмотрено в гл. 13. К эндотелию прилежит тон-
кий субэндотелиальный слой, состоящий из небольшого числа фиб-
робластов (клеток, синтезирующих коллаген) и коллагеновых во-
локон. Далее следует слой ветвящихся эластических волокон —
внутренняя эластическая мембрана (рис. 12.6, Л), которая осо-
бенно хорошо выражена в мелких артериях и образует внутреннюю
границу следующей оболочки — средней. Эта оболочка обычно
представляет собой самый толстый слой стенки сосудов, строение
и свойства которого наиболее существенно различаются в разных
областях системы кровообращения. Соответственно этим разли-
чиям артерии принято подразделять на артерии эластического и
мышечного типов. Поскольку, однако, резкой границы между ними
не существует и к тому же артерии тех или иных органов или
Гл 12. Артерии большого круга 299
300 Часть П. Механика кровообращения
у животных разных видов специализированы, такое деление пред-
ставляется чересчур упрощенным. Однако оно привлекает внима-
ние к тем изменениям строения сосудов, которые происходят по
мере их удаления от сердца. Средняя оболочка аорты и ее круп-
ных ветвей, отходящих в грудной полости, состоит из множества
концентрических слоев эластической ткани, разделенных тонкими
слоями соединительной ткани, коллагеновых волокон и небольшого
числа гладкомышечных клеток, которые в косом направлении со-
единяют последовательные эластические слои. Гладкомышечные
клетки длинные (25—50 мкм) и тонкие (5 мкм), часто с щупаль-
цеобразными отростками. Они содержат сократительные миофиб-
риллы, строение которых в основном такое же, как и в миокарде
(разд. 11.3). Под микроскопом миофибриллы гладких мышц видны
не столь отчетливо, как в миокарде, так как их меньше и они
расположены не столь упорядоченно. Однако функция тех и дру-
гих одинакова — в ответ на деполяризацию мембраны клетки они~
сокращаются и развивают усилие. Гладкомышечные клетки круп-
ных артерий ориентированы наклонно или продольно; в мелких
более периферических артериях их расположение ближе к кольце-
вому — здесь они образуют спирали.
По-видимому, у всех видов млекопитающих средняя оболочка
крупных артерий имеет слоистое, или «пластинчатое», строение.
В стенке аорты, которая в этом отношении изучена наиболее под-
робно, толщина пластин примерно постоянна и составляет около
15 мкм, так что общее число пластин почти пропорционально ра-
диусу аорты как у животных разных видов, так и у различных осо-
бей одного вида. В аорте мыши, например, пять пластин, а в аорте
свиньи — более семидесяти.
Строение средней оболочки более мелких артерий, расположен-
ных дальше от сердца, иное: они относятся к сосудам мышечного
типа. В средней оболочке таких артерий преобладают спирально
расположенные гладкие мышцы. Они размещаются в виде слоев,
разделенных небольшим количеством соединительной ткани, кол-
лагена и эластической ткани. С уменьшением радиуса сосуда
уменьшается и число слоев гладких мышц; в мельчайших артериях
и артериолах может быть всего один или два слоя. Однако во всех
мелких артериях без исключения средняя оболочка остается ос-
новной структурой, образующей стенку сосуда. Внешняя граница
средней оболочки отмечена очень тонким слоем эластической
ткани, которая называется наружной эластической мембраной.
Наружная оболочка стенки некоторых артерий может быть
столь же толстой, как средняя, или даже толще. Тем не менее под
микроскопом она менее заметна, так как состоит из рыхлой соеди-
нительной ткани с редкими эластическими и коллагеновыми во-
локнами, расположенными в основном продольно, и ее граница,
постепенно сливающаяся с окружающими тканями, неопреде-
ленна.
Гл 12. Артерии большого круга 301
Стенки всех артерий, диаметр которых превышает 1 мм, имеют
собственные питающие их кровеносные сосуды — «сосуды сосу-
дов»— vasa vasorum; последние берут начало от той же артерии,
которую питают, или от соседней с ней. Разветвляясь, они обра-
зуют сеть капилляров в наружной оболочке, а в крупных артериях
эта сеть достигает внутренних слоев средней оболочки. Питание
внутренней оболочки и самых глубоких слоев средней осущест-
вляется в основном за счет переноса веществ непосредственно из
просвета артерии. Мелкие лимфатические сосуды и тончайшие
нервные волокна, идущие к гладкомышечным клеткам артерии,
разветвляются, проходя через наружную оболочку.
Давно известно, что упругие свойства артерий нелинейны. При
растяжении артерии становятся гораздо более жесткими (как по-
казано на рис. 7.5). Объяснение этого факта дано в разд. 7.2.
Кроме того, модуль Юнга стенки артерий по мере их удаления от
сердца постепенно возрастает. Это может быть обусловлено рядом
факторов, в том числе изменениями соотношения между коллаге-
ном, эластином и гладкими мышцами и различной организацией
их в сосудах.
Как эластин, так и коллаген построены из длинных цепей бел-
ковых молекул и входят в состав волокнистых решеток и пластов,
находящихся в различных слоях стенки. Физические свойства обоих
этих материалов описаны ранее (разд. 7.2), и здесь нет необхо-
димости возвращаться к ним. Тем не менее следует напомнить,
что для крупных артерий существует совершенно четкая связь ме-
жду свойствами эластина и коллагена и упругим поведением их
стенки как целой структуры. С другой стороны, вопрос о том, ка-
ким образом влияют гладкие мышцы на свойства крупных арте-
рий, издавна является предметом споров. Причина этого кроется
главным образом в том, что большинство исследований проводилось
на удаленных из организма сосудах, так что результаты относятся
к состоянию, при котором степень сокращения гладких мышц
(«тонус сосудов») скорее всего не такова, как в организме жи-
вотного. Но даже в случаях, когда удается исследовать неповреж-
денные артерии in situ, сохраняются значительные трудности. Они
определяются тем, что в этих условиях сокращение гладких мышц
мелких артерий настолько уменьшает общую площадь попереч-
ного сечения сосудистого русла, что давление в крупных артериях
повышается. Поэтому маскируется любое уменьшение диаметра
крупных артерий, создаваемое увеличением активного напряжения
в их стенке. В недавно проведенном исследовании указанные эф-
фекты удалось разделить, и это позволило установить, что сокра-
щение гладких мышц даже самых крупных артерий, где таких
мышц довольно мало, способно значительно изменять напряжение
в стенке и фактически приводит к сужению этих сосудов.
В случае аорты подобные изменения диаметра невелики. На-
пример, в ответ на электрическое раздражение .нервов, идущих
302 Часть II. Механика кровообращения
к гладким мышцам стенки аорты, ее диаметр уменьшается прибли-
зительно лишь на 5% (среднее артериальное давление при этом
поддерживали постоянным). В естественных условиях большинство
раздражителей менее интенсивны. Поэтому сомнительно, чтобы
физиологические изменения активности гладких мышц могли при-
водить к более выраженному изменению диаметра аорты. Дей-
ствие гладких мышц периферических (мышечных) артерий выра-
жено сильнее. В подобных же опытах диаметр, например, бедрен-
ной артерии уменьшался на 20%, а перерезка волокон, иннерви-
рующих-гладкие мышцы этой артерии, приводила приблизительно
к такому же увеличению ее диаметра относительно исходной вели-
чины.
В зависимости от условий опыта и способа регистрации влия-
ние сокращения гладких мышц сказывается на свойствах артерий
различным образом. Если, например, во время сокращения этих
мышц диаметр сосуда поддерживают неизменным, то его стенка
становится более жесткой, а касательный модуль упругости воз-
растает. Если же допустить изменение диаметра, то сокращение
гладких мышц приведет к сужению сосуда, а модуль упругости
может уменьшиться (это явление объяснено в разд. 7.2). В есте-
ственных условиях у бодрствующего животного сокращение глад-
ких мышц стенки крупных артерий повышает их жесткость (если
только сосуд не растянут столь сильно, что коллаген в его стенке
почти полностью принимает нагрузку на себя; в этом случае со-
кращение гладких мышц никак не проявится, поскольку модуль
упругости даже максимально сократившихся гладких мышц мень-
ше модуля упругости коллагена).
Выше было отмечено, что гладкомышечные клетки крупных ар-
терий обычно располагаются наклонно или продольно и, по-види-
мому, соединены с эластической тканью, в частности с внутренней
и наружной эластическими мембранами средней оболочки. По-
этому гладкие мышцы меняют напряжение в стенке двумя путями:
во-первых, они натягивают эластическую ткань и, во-вторых, вос-
принимают нагрузку параллельно с этой тканью и коллагеном.
Слабое влияние гладких мышц на диаметр крупных артерий даже
при максимальном сокращении этих мышц обусловлено, по-види-
мому, их наклонным расположением и ограниченной способностью
к укорочению (в предельном случае не более чем на 25% их длины
в расслабленном состоянии). Способность некоторых сосудов, и в
том числе весьма крупных артерий мышечного типа, спазмиро-
ваться в ответ на грубые воздействия или при повреждениях также,
вероятно, есть проявление каких-то свойств гладких мышц. Од-
нако эта способность еще не получила удовлетворительного объяс-
нения: при спазме просвет артерии может практически исчезнуть,
а не только уменьшиться на 20—25%, как это происходит в ответ
на импульсы симпатических волокон и что согласуется с извест-
ными сократительными свойствами гладких мышц артерий.
Гл. 12. Артерии большого круга 303
Итак, следовательно, в обычных обстоятельствах действие глад-
ких мышц на диаметр и жесткость крупных артерий оказывается,
по всей видимости, сравнительно слабым, в особенности при срав-
нении с действием в системе мелких артерий, где гладкие мышцы
являются основным компонентом стенки, а их почти кольцевое рас-
положение означает, что они способны сильно изменять просвет
сосуда1). Интенсивность сокращения гладких мышц этих сосудов
определяет как местное распределение кровотока, так и общее
периферическое сопротивление, что еще предстоит обсудить в гл. 13.
Однако, как мы выяснили в гл. 7, гладкие мышцы существенно
влияют на динамическое поведение крупных артерий. Вязкие свой-
ства артерий (релаксация напряжения, ползучесть, гистерезис),
которые приписывают главным образом наличию гладких мышц,
сильно усложняют механическое поведение стенки (раз. 7.2). Еще
одним динамическим свойством гладких мышц, физиологическая
значимость которого окончательно не установлена, является «мио-
генная реакция». Это свойство гладких мышц заключается в том,
что они деполяризуются (и, следовательно, сокращаются) в ответ
на растяжение. Миогенная реакция развивается во времени мед-
ленно (слишком медленно, чтобы изменить свойства стенки арте-
рий в течение, например, одного цикла сокращения сердца), но нет
сомнений в том, что она может существенно изменять зависимость
деформации стенки от напряжения в ней. Физиологическая значи-
мость миогенной реакции остается предметом дискуссий.
Толщина артериальной стеики. Толщина стенки разных арте-
рий существенно различна. Характерные значения приведены в
табл. I. Значительный разброс результатов измерений неудивите-
лен, так как в процессе изготовления препаратов мелких артерий
трудно сохранить должные значения давления в этих сосудах и
предотвратить изменения их формы. Кроме того, — и это касается
не только мелких, но и крупных артерий, — наружная оболочка
может постепенно сливаться с окружающими тканями, и потому
трудно определить, где именно заканчивается стенка артерии. Мик-
рососуды собак систематически не исследовались, так что для
табл. I пришлось собирать данные о размерах мельчайших сосу-
дов у млекопитающих других видов: кошек, крыс и мышей. Это,
по-видимому, не очень важно, так как сравнение показывает, что
размеры мелких сосудов у млекопитающих, значительно различаю-
щихся по массе тела, поразительно одинаковы.
Такое утверждение совершенно неприменимо к крупным сосу-
дам: толщина их стенок у мелких млекопитающих резко меняется
*) Недавно было высказано предположение о том, что сокращение гладких
мышц, благодаря их косому расположению, приводит и к уменьшению дливы
сосуда. Тогда, как показывает простой геометрический расчет, уменьшение про-
света может быть намного большим, чем при чисто кольцевом расположении
волокон. См. Alexander R. S. (1977), Critical closure reexamined, Circulat. Res.,
40, 531. — Прим. ped.
И1.ЙГ
Вес, кг
Рис. 12.7. Толщина средней оболочки грудной аорты у де-
сяти видов млекопитающих. Толщина этой оболочки в ряду
мелких млекопитающих быстро растет с увеличением веса
взрослой особи, тогда как у крупных млекопитающих ее
рост выражен значительно слабее или отсутствует вовсе.
[Wolinsky, Glagov (1967). Circulation Res. 20, 99.]
Рис. 12.8. Диаметр грудной аорты у десяти видов млеко-
питающих. Увеличение диаметра с ростом веса взрослой
особи для мелких млекопитающих более значительно, чем
для крупных. [Wolinsky, Glagow (1967). Circulation Res.,
20, 99.]
Гл. 12. Артерии большого круга 305
с изменением массы тела. Этот факт очевиден из рис. 12.7, где
приведены результаты соответствующего исследования, в котором
при взятии материала тщательно следили за тем, чтобы сохранить
размеры сосудов такими же, какими они были in vivo, а при изго-
товлении гистологических препаратов старались свести до мини-
мума изменения формы сосудов и вводили поправки на такие из-
менения. Разумеется, с изменением массы животных сильно ме-
няется также и диаметр крупных артерий. Это следует из рис. 12.8,
заимствованного из того же источника. Сравнивая две кривые,
можно прийти к выводу, что отношение толщины стенки крупных
артерий к их диаметру у разных особей одного вида и у животных
разных видов примерно одинаково. Таким образом, отношения h/d,
приведенные в табл. I, не зависят от массы животного.
Итак, внутренний диаметр d и толщина стенки h крупных цен-
тральных артерий зависят от массы животного, тогда как их отно-
шение (hid) остается примерно одинаковым для всех артерий этой
группы у животных разных видов. По мере продвижения к пери-
ферии, к малым артериям, их стенка становится все тоньше, но
отношение h/d возрастает, так что наружный диаметр мельчайших
артерий может вдвое превосходить диаметр просвета, даже когда
гладкие мышцы расслаблены. И наконец, для самых мелких сосу-
дов, и особенно для капилляров, совершенно исчезает зависимость
d и h от массы животных — диаметр и толщина стенки оказывают-
ся почти одинаковыми у млекопитающих всех видов.
Возрастные изменения стенки артерий. Размеры и свойства
крупных артерий зависят не только от их места в системе крово-
обращения и от размеров тела, но и от возраста животного. Си-
стематические исследования изменений в процессе роста и старе-
ния проведены главным образом на тканях человека, причем зна-
чительная часть измерений осуществлена посмертно — на спав-
шихся тканях трупа. Поэтому мы лучше осведомлены о возраст-
ных изменениях не столько точных размеров сосудов, сколько ги-
стологического строения их стенки. Тем не менее очевидно, что на
протяжении жизни толщина стенки артерий и ее строение медленно
меняются и что такие изменения отражаются на их упругих свой-
ствах. Понятно также, что сходные изменения артерий возникают
и у млекопитающих других видов, хотя данные о них менее полны.
У растущего животного и у ребенка диаметр и толщина стенки
крупных артерий постепенно увеличиваются. Данные о размерах
артерий при физиологических значениях растяжения отсутствуют,
и в табл. 12.2 приведены размеры аорты человека при нулевом
трансмуральном давлении. Величина h/d при этом условии прак-
тически постоянна для любого возраста: такой результат был не-
однократно подтвержден.
Увеличение толщины стенки крупных эластических артерий оп-
ределяется в основном утолщением и разрастанием эластических
Пластин средней оболочки. Этот процесс заканчивается с насту-
306 Часть II. Механика кровообращения
Таблица 12.2
Размеры нерастянутых артерий человека в разном возрасте1)
-ч Внутренний диаметр d. Возраст, годы мм Толщина стенкн Л, мм hld
1 11 3-6 13 15-18 17 20 19 0,83 0,08 1,14-1,23 0,09 1,41-1,58 0,09 1,58 0,08
*) Вычислено по данным Dittmer, Grebe (1959), Handbook of circulation, p 20, W B. Saun-
ders Company, Philadelphia.
плением зрелости, и далее он незаметно сливается с гистологиче-
скими изменениями другого типа, которые обычно относят к деге-
неративным. Именно, эластические элементы стенки начинают из-
нашиваться, фрагментироваться и могут подвергаться обызвест-
влению; увеличивается количество коллагеновых волокон, которые
замещают гладкомышечные клетки в одних слоях стенки и разрас-
таются в других. В целом это приводит к увеличению диаметра
сосудов и к утолщению их стенки, так что она становится значи-
тельно менее растяжимой. Такое повышение жесткости затраги-
вает как крупные артерии, так и артерии среднего калибра *). По-
скольку этот процесс характерен для всех индивидуумов, но вы-
ражен далеко не одинаково, трудно сказать, является ли он физио-
логическим или его следует отнести к разряду патологических про-
цессов.
Существует и безусловно патологическое поражение артерий —
атеросклероз. Это заболевание встречается все чаще и в развитых
странах является причиной почти половины смертей. В связи с
этим и учитывая также, что возможному значению механических
явлений в развитии атеросклероза посвящены специальные иссле-
дования, мы подробнее обсудим их результаты в разд. 12.8.
Возрастные изменения размеров и упругих свойств артерий
человека явились предметом ряда исследований. Рис. 12.9, заим-
ствованный из одной работы такого типа, обобщает происходящие
изменения. Исследованию подвергали непораженные артерии, взя-
тые посмертно у трупов «молодых» (до 20 лет) и «пожилых»
(после 35 лет) людей. Перед измерением артерии растягивали до
прижизненной длины и создавали в них физиологический уровень
давления. Видно, что с возрастом диаметр аорты увеличивается,
а ее стенка утолщается, но относительно несколько быстрее, чем
возрастает диаметр. Поэтому отношение h/d увеличивается. Диа-
*) Этот процесс нередко называют артериосклерозом, и поэтому возникает
путаница: этим же словом обозначают иногда и поздние стадии атеросклероти-
ческого поражения стенки артерий.
Гл 12 Артерии большого круга Stiff
метр меньших, «мышечных», ветвей аорты практически не меняет-
ся, хотя их стенка значительно утолщается. Таким образом, с воз-
растом сосуды становятся шире и толще, однако эти явления вы-
ражены в различной степени для разных сосудов. Для крупных
артерий характерна тенденция как к увеличению просвета, так и
к утолщению стенки, тогда как для артерий меньшего диаметра —
только к утолщению стенки.
Эти изменения размеров артерий сопровождаются значитель-
ными изменениями их упругих свойств. Касательный модуль Юнга
Е (разд. 7.2) грудной аорты, измеренный при физиологическом
значении давления (1,33-104 Н-м-2 « 100 мм рт. ст.), с возрастом
неуклонно увеличивается (рис. 12.10, Л), тогда как модуль Юнга
более мелких артерий либо не меняется, либо уменьшается
(рис. 12.10, Б). Объяснение заключается, по-видимому, в том, что
с возрастом диаметр аорты увеличивается. Поэтому состояние со-
суда, изображаемое точкой на кривой длина — напряжение, как бы
продвигается вправо вдоль ее нелинейного участка (рис. 7.5, Л)
и соответственно увеличивается Е.
Многое из обсуждавшегося в этом разделе можно обобщить,
сказав, что размеры и упругие свойства артерий зависят от воз-
раста, массы тела и от места сосуда в системе кровообращения.
Большинство таких зависимостей очевидно: представив себе, на-
пример, мышь и слона, все мы предскажем некое соответствие ме-
жду размерами аорты и массой тела. Тем не менее, поскольку
многие свойства связаны друг с другом не столь очевидным обра-
зом, ряд подробностей оказывается существенным: мы перейдем
в следующем разделе к изучению механических явлений (таких,
как форма пульсовых волн давления и скорости кровотока), а их
можно до конца понять, только опираясь на обстоятельные знания
анатомической организации сосудистого русла и поведения стенки
артерий. В конечном счете содержание этого раздела служит сво-
его рода предостережением: оно напоминает о том, что биологиче-
ским системам присуща изменчивость. Физики, изучающие крово-
обращение, хотят получить о нем точные количественные сведения
и нередко разочаровываются, выяснив, что физиологи ими не рас-
полагают. Поэтому и представлялось поучительным показать, что
измерение даже такой незамысловатой величины, как диаметр
аорты, — дело не только исключительно трудное, но что резуль-
таты будут зависеть от того, на каком участке аорты измеряют
диаметр, от вида животного, от возраста, массы тела, кровяного
давления и степени сокращения гладких мышц. Чтобы определить
число Рейнольдса для аорты, мы должны знать не один только ее
диаметр, но и две другие величины — скорость течения крови и ее
вязкость. Чтобы найти скорость пульсовой волны, используя урав-
нение Мсенса — Кортевега (разд. 12.3), требуются данные о диа-
метре и толщине стенки артерии, о ее модуле Юнга и о плотности
крови. И все эти величины, подобно диаметру аорты, подвергаются
308 Часть П Механика кровообращения
Рис. 12.9. Возрастные изменения радиуса (А) и толщины (Б) артерий человека
(данные получены прн физиологических значениях давления). Средние значения
этих величин для разных артерий «молодых» (/) и «пожилых» (//) людей пред-
навлены в соответствии с местом артерий в системе кровообращения. [Learoyd,
Taylor (1966). Circulation Res., 18, 278.]
A
Рис. 12.10. Возрастные изменения упругих свойств артерий человека. А. Зависи-
мость от возраста касательного модуля Юига аорты при давлении 100 мм рт. ст.
Б. Подобная зависимость для подвздошной артерии. [Learoyd, Taylor (1966).
Circulation Res., 18, 279,]
Гл. 12. Артерии большого круга 309
влияниям многих биологических факторов и поэтому изменчивы.
Из сказанного следует, что вычисляемые параметры, подобные
числам Рейнольдса, могут изменяться в широких пределах, даже
если использовать для их подсчета результаты точных и совмести-
мых измерений. Когда же такие параметры вычисляют исходя из
плохо подобранных данных, разброс становится фантастическим.
12.2. Давление и кровоток в артериях большого круга
Вследствие периодичности изгнания крови из левого желудочка
в большой круг давление во всех его артериях колеблется. Типич-
ная запись давления в аорте представлена на рис. 12.15 (и на мно-
гих последующих рисунках). Говоря о давлении крови в артериях,
мы будем следовать двум традициям, и чтобы понять, что именно
подразумевают под давлением в артериях и какое отношение оно
имеет к механическим явлениям, определяющим движение крови,
на этих традициях следует остановиться более подробно.
Первая из них заключается в* том, что кровяное давление при-
нято измерять в миллиметрах ртутного столба (мм рт. ст.). Такой
единицы нет ни в одной из международных систем единиц — ни в
старых, ни в новых; появилась же она потому, что в подавляющем
большинстве случаев кровяное давление на самом деле измеряют
в мм рт. ст. С этой целью используют ртутный манометр, соеди-
ненный с манжеткой, надувая которую, пережимают артерию ко-
нечности. Такой прибор называется сфигмоманометром. В защиту
мм рт. ст. как единицы измерения давления следует, вероятно,
сказать, что ее бессменно используют с 1828 г., когда ее ввел Пуа-
зейль, и, следовательно, она пережила многие другие единицы из-
мерения давления. Имея в виду, что мм рт. ст. — единица общеиз-
вестная, в этой книге мы всюду используем ее наряду с соответ-
ствующей единицей системы СИ ’).
Вторая традиция состоит в том, что, говоря о кровяном давле-
нии, всегда имеют в виду давление, отсчитываемое относительно
атмосферного. Обычно принимают, что давление в тканях тела,
непосредственно у наружной стенки артерии, равно атмосферному,
так что давление крови рассматривают как трансмуральное дав-
ление (трансмуральное давление есть рв — Рн, где рв — давление
внутри артерии, а рв — наружное давление, равное атмосферному).
Трансмуральное давление. Причины, в силу которых необхо-
димо знать трансмуральное давление, весьма разнообразны. Эта
величина нужна, например, в том случае, когда при анализе рав-
новесия стенки сосуда с учетом сил, действующих на нее со сто-
роны крови, удается измерить параметры этой стенки лишь по-
смертно, а нам необходимо определить, каковы они при жизни.
Значения трансмурального давления говорят нам нечто о нагрузке,
*) 1 мм рт. ст. = 133,32 H-m-s.
310 Часть II. Механика кровообращения
против которой должно работать сердце, о состоянии русла пери-
ферических сосудов, объеме крови и рефлекторных реакциях сер-
дечно-сосудистой системы, а также о многих других физиологиче-
ских процессах. И однако, располагая данными о значениях транс-
мурального давления, мы решительно ничего не знаем о силе, ко-
торую необходимо приложить, чтобы обеспечить движение крови
по сосудам. Для этого следует определить разность давлений ме-
жду двумя точками системы кровообращения, точно так же как
необходимо установить разность давлений по обе стороны стенки
артерии, чтобы исследовать действующие в стенке силы. Суть
сказанного здесь состоит в том, что давления, измеряемые физио-
логом, — это нередко вовсе не те давления, которые необходимы
инженеру для рассмотрения физических процессов в системе кро-
вообращения. Например, разность давлений по обе стороны стенки
крупной артерии руки составляет обычно 1.33-104 Н-м-2
(100 мм рт. ст.). (Именно эту разность по традиции называют
«артериальным давлением».) Однако сила, которая на самом деле
обеспечивает движение крови из восходящей аорты в крупную ар-
терию руки, есть разность давлений между этими сосудами, а она
обычно равна всего лишь 0,03-104 Н-м-2 (2—3 мм рт. ст.). Эту
разность редко удается измерить из-за больших методических труд-
ностей.
Важным фактором, влияющим на величину трансмурального
давления, является сила тяжести. Почему это так — легко понять,
представив состояние сосудистой системы у мертвого животного.
Гладкие мышцы всех сосудов расслаблены, произошло перераспре-
деление крови. Увеличение объема периферических артериальных
сосудов и мелких вен столь значительно, что большинство крупных
сосудов фактически запустевает, и они могут оказаться в состоянии
коллапса, т. е. спадения ’). Если бы сила тяжести отсутствовала,
давление во всех сосудах оказалось бы одинаковым и практически
равным атмосферному.
Чтобы понять, как влияет сила тяжести, рассмотрим, что про-
изойдет, если придать трупу вертикальное положение. Кровь снова
перераспределится, и все сосуды, расположенные в теле выше того
уровня, на котором давление в сосудах равно атмосферному
(«точка равенства давлений»), спадутся. Давление в сосудах, на-
ходящихся выше этого уровня, поскольку они запустевают, срав-
няется с давлением в окружающих тканях и, следовательно, ока-
жется приблизительно равным атмосферному давлению р&. Ниже
этого уровня (а при неполном запустевании и выше его) давле-
ние в сосудах рв определяется соотношением
Рв = Ра + Р£Л, (12.1)
’) Понятие коллапса сосудов вкратце рассмотрено в разд. 7.3. Спавшиеся
сосуды вовсе ие обязательно абсолютно пусты.
Г л. 12. Артерии большого круга 311
Рис. 12.11. Среднее избыточное (по от-
ношению к атмосферному) артериальное
и венозное давление в разных отделах
системы кровообращения человека. [Раш-
мер Р. Динамика сердечно-сосудистой
системы. Пер. с англ. — М.: Медицина,
1981.]
Среднее артериальное
давление.
где h — расстояние от «точки равенства давлений» вниз по вер-
тикали (для сосудов, лежащих выше этой точки, h отрицательно),
р — плотность крови, g — ускорение силы тяжести. Это соотноше-
ние, конечно, тождественно уравнению (4.2), так что то давление
(pg/i), которое мы обсуждаем, есть гидростатическое давление.
Гидростатическое давление существенно влияет на распределе-
ние крови не только у трупа, но и в сосудистой системе живого
организма. У вертикально стоящего человека нулевой уровень
(h = 0) находится примерно на уровне сердца (рис. 12.11). По-
этому, когда человек стоит в такой позе, вены на шее обыкновенно
спадаются (правда, не полностью, кровь все же течет по ним),
а давление, под которым наполняется сердце, — «давление напол-
нения» (гл. 11) —близко к атмосферному. Однако оно может ме-
няться при изменении либо объема крови, либо объема сосуди-
стого русла нижних конечностей.
Наиболее хорошо известное проявление действия силы .тяжестй
на сердечно-сосудистую систему — обморок. Один из важных ме-
ханизмов обморока заключается в том, что у стоящего человека
резко возрастает объем крови в венах ног, а давление наполнения
сердца падает. Это приводит к снижению производительности
сердца и, следовательно, к уменьшению кровоснабжения мозга.
Когда в результате обморока человек падает или когда ему при-
дают горизонтальное положение, устраняется сила, удерживающая
кровь в сосудах ног, и тем самым обморок становится менее глу-
боким. К механизмам, обычно защищающим каждого из нас от
обморока, когда мы стоим, относится рефлекторно обусловленное
сужение венозных сосудов ног, которое сильно уменьшает способ-
312 Часть II Механика кровообращения
ность этих сосудов растягиваться и накапливать кровь. Еще один
механизм защиты от обморока — сужение артериол, которое при-
водит к увеличению сопротивления кровотоку и не позволяет ар-
териальному давлению заметно понизиться *). Если сосудосужи-
вающий эффект блокирован (например, некоторыми лекарствен-
ными препаратами) или нарушен в результате заболевания, резкое
вставание часто сопровождается обмороком или головокружением.
То же самое наблюдается, когда сосуды сильно расширяются в
жару или после принятия горячей ванны. У пилотов высокоско-
ростных самолетов, когда они выполняют крутые виражи и испы-
тывают большие перегрузки, может возникнуть потеря зрения (так
называемая «черная слепота»), если напряжение в стенке сосудов,
даже подвергающихся активным суживающим влияниям, недоста-
точно велико, чтобы уравновесить увеличившееся из-за перегрузок
гидростатическое давление. В таком случае стенки сосудов будут
растягиваться (становясь при этом все более жесткими, см.
рис. 14.2) до тех пор, пока натяжение стенки не уравновесит транс-
муральное давление.
Количественно связь между трансмуральным давлением
(рв — рн) и натяжением стенки трубки определяется законом Ла-
пласа [уравнение (7.6)]:
Т = (ръ — pn)-R,
где Т — натяжение на единицу длины стенки трубки, 7? — ее ра-
диус. Когда одновременно увеличиваются и трансмуральное дав-
ление, и радиус сосуда, равновесие сил может быть достигнуто
только при условии, что натяжение стенки растет быстрее, чем
любая из этих двух величин. Поэтому устойчивость стенки сосудов
зависит от нелинейности их упругих свойств. Отметим, что для
растяжения сосуда совершенно безразлично, вызывается ли оно
снижением рн или повышением ръ. Если подвергнуть нижнюю по-
ловину тела здорового человека действию давления, которое мень-
ше атмосферного, у него может возникнуть обморок. В физиологи-
ческих условиях сосуды большого круга, заключенные в грудной
полости, подвергаются действию давления рн, которое при каждом
цикле дыхания становится то выше, то ниже атмосферного, и та-
кие колебания приводят к важным последствиям, в частности ка-
сающимся устойчивости формы вен (см. гл. 14).
Из сказанного выше ясно, что гидростатические силы являются
важной составляющей сил, действующих на стенку сосудов, осо-
бенно вен. В обычных условиях, как это показано на рис. 12.11,
гидростатическое давление действует и на артерии, и на вены,
*) В этом состоит одно из важных назначений «миогенной реакции» глад-
ких мышп артериол, о которой упоминалось выше. Такая реакция осуществляется
вследствие растяжения стенки артериол гидростатическим давлением. — Прим,
ред.
Гл. 12. Артерии большого' круги 313
однако среднее артериальное давление (1,2-104 Н-м'2, или
90 мм рт. ст.) прибавляется к гидростатическому только в арте-
риях, но не в венах. Трансмуральное давление в артериях голени,
может быть в четыре раза больше, чем в артериях поднятой руки;
в то же время, если поднять руку над грудной клеткой, вены этой
руки запустеют и спадутся.
Таким образом, в самых обычных обстоятельствах значения1
трансмурального давления могут меняться в очень широких пре-
делах; об этом нередко забывают при анализе баланса сил, дей-
ствующих на стенку артерий. А с другой стороны, граница «без-
опасных» значений артериального давления лежит в не очень шиА
роком диапазоне: в гл. 7 (разд. 7.3) мы отмечали, что артерии спо-
собны спадаться, даже когда трансмуральное давление все еще
остается слегка положительным (т. е. растягивающим)'. Здесь за<--
ключен еще один источник биологических различий: чтобы, пред-
отвратить спадение артерий, снабжающих кровью мозг, среднее?
артериальное давление у стоящего жирафа должно быть много
больше, чем у человека.
О растягивающем действии гидростатического давления на ар-
терии и вены, а тем самым — ио его влиянии на упругие свойства
этих сосудов нередко забывают. Обусловлено это тем, что врачи
измеряют кровяное давление, накладывая манжетку на область
плеча, а поскольку во время этой процедуры плечо располагается
на уровне верхней части грудной клетки, гидростатическая состав-
ляющая трансмурального давления в плечевой артерии у стоящего
или лежащего пациента равна нулю. При исследованиях кровооб-
ращения, в которых применяют заполненный жидкостью и соеди-
ненный с манометром зонд, мембрану манометра принято распо-
лагать на уровне «середины груди», что также устраняет гидро-
статическую составляющую давления. Для таких измерений сла-
гаемое pgh в уравнении (12.L) равно нулю, и результаты говорят
только о величинах «избыточного давления», создаваемого дея-
тельностью сердца. Однако, обращаясь к упругим свойствам ар-
терий, строит вспомнить, что 200 мм рт. ст. может быть обычным
значением трансмурального давления в сосудах опущенных голе-
ней и стоп.
Пульсирующее давление в крупных артериях. Весьма сложные,,
изменяющиеся картины распределения трансмурального давления,,
которые могут возникать в сердечно-сосудистой системе, очень
важны с точки зрения механики стенки сосудов, но эти картины
не должны отвлекать нас от выяснения действительной природы
сил, ответственных за движение крови. Указанные силы не имеют
ничего общего с трансмуральным давлением: кровь может течь
даже в почти полностью спавшихся сосудах, как это происходит,
например, когда она возвращается в сердце по венам шеи стоя-
щего человека. Несущественно и гидростатическое давление: чтобы
заставить жидкость, течь по трубке, расположена ли она гориЗон-
314 Часть !!• Механика кровообращения
Рис. 12.12. Форма волны избыточного давления и его среднее значение в раз-
личных отделах артериальной системы.
тально или вертикально, требуется одно и то же количество энер-
гии (если только размеры трубки не изменяются *). Поэтому,
когда мы обращаемся к силам, определяющим движение крови,
важным становится только давление, создаваемое сердцем, — дав-
ление, которое мы будем называть избыточным (рИзб)- Эта вели-
чина есть разность между фактически существующим давлением
и его гидростатической составляющей, и как раз эту величину
имеют в виду, когда говорят о кровяном давлении. Градиент имен-
но избыточного давления определяет движение жидкости. Распре-
деление избыточного давления в системе кровообращения показано
на рис. 12.12. Можно видеть, что, заставляя кровь протекать через
систему микрососудов, оно в значительной мере «растрачивается».
Это явление мы обсудим в гл. 13, пока же нас интересуют значи-
тельно меньшие градиенты давления, которые заставляют кровь
двигаться по крупным артериям. На этих градиентах давления и
обусловленном ими движении крови мы и сосредоточим свое вни-
мание до конца главы. Как это принято, обычно мы будем назы-
вать избыточное давление кровяным давлением или же просто
давлением, поскольку из контекста ясно, что имеется в виду.
Сначала обратимся к давлениям. На рис. 12.13 приведены за-
писи давлений в левом желудочке и в восходящей аорте сразу за
аортальным клапаном. Кривые зарегистрированы при помощи
двух чрезвычайно тщательно подобранных один к другому датчи-
б Подразумевается замкнутая трубка. — Прим. ред.
Гл. 12. Артерии большого круга 315
ков давления, поэтому они точно отражают события, совершаю-
щиеся в каждое мгновение цикла сокращения сердца1).
Как говорилось в гл. 11, в самом начале систолы левого желу-
дочка давление в нем быстро возрастает и вскоре становится выше
давления в аорте, так что аортальный клапан открывается, кровь
изгоняется в аорту и давление в ней повышается. В течение неко-
торого времени после начала периода изгнания крови давление
в желудочке превышает давление в аорте. Примерно в середине
этого периода кривые давления пересекаются, и теперь в области
Рис. 12.13. Зарегистрированные одновременно кривые давления в левом желудоч-
ке и в восходящей аорте собаки. [Noble (1968). The contribution of blood mo-
mentum to left ventricular ejection in the dog. Circulation Res., 23, 663—670.]
аортального клапана создается обратный градиент давления, ко-
торый сохраняется до тех пор, пока оба давления не начинают
уменьшаться. В этот момент на кривой давления для аорты появ-
ляется зубец (дикротический зубец), который отмечает закрытие
аортального клапана, после чего сердечная мышца расслабляется
и давление в желудочке очень быстро падает. Снижение давления
в аорте происходит значительно медленнее. Причину этого легко
понять, если вспомнить, что крупные центральные артерии, в осо-
бенности аорта, — артерии эластического типа, и поэтому в тече-
ние систолы они действуют как емкость, накапливающая часть
изгнанного объема крови, которая затем во время диастолы про-
талкивается в периферические сосуды.
Переместимся теперь немного вниз по течению крови и проана-
лизируем кривые давления, зарегистрированные в двух точках по
оси аорты (например, в точках 0 и 4 см на рис. 12.14). Мы можем
убедиться, что форма этих кривых почти одинакова, но та из них,
что записана несколько ниже по потоку, чуть запаздывает во вре-
') Дальше мы убедимся, что именно пульсации давления и скорости потока
крови в крупных артериях представляют собой определяющую особенность гидро-
механических явлений в них, а чтобы точно регистрировать такие процессы, при-
боры, применяемые в экспериментальных исследованиях, должны обладать до-
статочным быстродействием Поэтому к обычным требованиям стабильности и
чувствительности добавляется еще одно — приборы должны обладать хорошей
частотной характеристикой. Нередко это требование оказывается очень трудно
осуществимым, и в прошлом оио далеко не всегда выполнялось (см. стр. 324).
316 Часть 11. Механика кровообращения
мени. Другими словами, пульсовое колебание давления, порожден-
ное сокращением желудочка, распространяется вдоль аорты в виде
волны. Зная расстояние между точками измерения давления, по
запаздыванию волны можно грубо оценить скорость ее распро-
странения, а также составить представление о разности давлений,
обусловливающей движение крови по этому участку, вычитая зна-
чение давления в точке, находящейся ниже по течению, из значе-
ния в вышележащей точке.
Если одновременно измерить давление в нескольких точках
по длине аорты (или, что методически гораздо проще, произвести
Время, мс
Рис. 12.14. Кривые артериального давления, зарегистрированные одновременно в
различных участках аорты собаки: точка «О см» находится в самом начале ии-
исходящей аорты. [Olson (1968) Aortic blood pressure and velocity as a function
of time and position, J. appl. physiol 24, 563—569.]
с помощью зонда ряд последовательных измерений в разных точ-
ках аорты и затем совместить кривые во времени, пользуясь, как
опорными, сигналами от другого зонда, положение которого фикси-
ровано), то мы обнаружим, что форма волны давления по мере ее
перемещения по аорте изменяется (рис. 12.14). Разумеется, волна
все больше и больше запаздывает, но, кроме того, она становится
круче, ее амплитуда возрастает, тогда как резкий дикротический
зубец исчезает. Таким образом, наблюдается удивительное явле-
ние: фактический рост систолического давления по мере удаления
от сердца. (И это характерно не только для рассматриваемого нами
вида животных — собак: такое же явление постоянно наблюдается
как у животных других видов, так и у человека.) Конечно, в дей-
ствительности при удалении волны давления от сердца среднее
давление уменьшается. Но установить это непосредственно по за-
писям, представленным на рис. 12.14, трудно, поскольку падение
среднего давления по всей аорте составляет всего 0,05-104 Н-м~2
(4 мм рт. ст.), тогда как амплитуда колебаний давления между
максимумом в систолу и минимумом в диастолу возрастает при-
мерно вдвое.
Гл. 12. Артерии большого круга 317
Этот процесс увеличения амплитуды пульсовых колебаний дав-
ления продолжается и в ветвях аорты, вплоть до сосудов (у со-
баки) приблизительно третьего поколения ветвления, например до
большой подкожной артерии (a. saphena magna), внутренний диа-
метр которой на голени равен 1—2 мм (см. рис. 12.19). Далее как
амплитуда пульсовых колебаний, так и среднее давление круто
падают до уровня, характерного для микрососудов.
Радиальное расширение и спадение стенки артерий при прохож-
дении пульсовой волны давления измеряли рядом способов. На
рис. 12.15 приведены произведенные одновременно записи изме-
нений диаметра артерии и давления в ней. Видно, что движения
Рис. 12.15. Зарегистрированные одновре-
менно изменения диаметра аорты собаки
и давления в течение одного сердечного
цикла. [Barnett, Mallos, Shapiro (1961).
Relationship of aortic pressure and dia-
meter in the dog, J. appl. Physiol., 16,
545—548.]
стенки артерии происходят практически в фазе с изменениями дав-
ления, иными словами, стенка ведет себя как чисто упругий ма-
териал, почти не проявляя гистерезиса, обусловленного ее вязко-
упругими свойствами (разд. 7.2). Заметим также, что изменение
радиуса артерии мало; при пересчете в относительное удлинение
(разд. 7.1) оно составляет всего 7%. Эти наблюдения оправды-
вают применение для описания поведения стенки артерий линей-
ной теории упругости, поскольку в ней свойства стенки характе-
ризуются постоянным модулем упругости Е (разд. 7.2). Как мы
увидим в следующем разделе этой главы, линейная теория упру-
гости в качестве первого приближения с успехом используется при
описании пульсовых волн давления и позволяет предсказать ско-
рость их распространения.
Движение крови. Чтобы завершить описание процессов в круп-
ных артериях и перейти к детальному анализу механики этих про-
цессов, необходимо остановиться на движении крови в аорте и ее
ветвях. Поскольку все давления и их градиенты меняются во вре-
мени, естественно предположить, что это относится и к движению
крови. Во всей системе кровообращения мы имеем дело с неуста-
новившимся течением, и, как выяснится в дальнейшем, это сильно
усложняет физическую теорию, которую нам предстоит использо-
вать. Более того, это означает, что для понимания происходящих
процессов необходимо уметь точно измерять мгновенные значения
расхода или скорости крови, а весьма часто одновременно произ-
водить и запись давления. Осуществление таких измерений даже в
опытах на животных, когда возможен хирургический доступ к круп-
318 Часть П. Механика кровообращения
ным сосудам, является непростой методической задачей. У чело-
века же доступ к сосудам большого круга крайне ограничен, по-
этому данные такого рода весьма скудны. Запас таких сведений
пополняется в настоящее время (хотя и медленно) лишь за счет
проведения в кардиологических клиниках сложных диагностических
исследований с использованием метода зондирования.
На рис. 12.16 представлены кривые давления и расхода крови
в восходящей аорте вблизи клапана. При частоте сокращений
сердца, характерной для состояния покоя, фаза «прямого» (т. е.
Рис. 12.16. Зарегистрированные одновременно кривые давления и расхода крови
в восходящей аорте бодрствующей собаки, находящейся в состоянии покоя.
[Noble (1968). The contribution of blood momentum to left ventricular ejection
in the dog. Circulation Res., 23, 663—670.]
от сердца) движения крови по аорте составля.ет примерно от одной
четверти до одной трети продолжительности цикла сокращения
сердца. Когда частота сокращений сердца возрастает (как это
происходит, например, при мышечной работе), продолжительность
как систолы, так и диастолы уменьшается, но в разной степени,
причем укорочение диастолы гораздо значительнее. Поэтому при
частых сокращениях сердца относительная продолжительность пря-
мого движения крови увеличивается и может достигать 50% вре-
мени полного цикла.
Прямое движение крови в восходящей аорте начинается, есте-
ственно, в момент открытия аортального клапана вместе с изгна-
нием крови из желудочка. Кровоток резко возрастает до пикового
значения и тут же начинает быстро снижаться, хотя и несколько
медленнее, чем повышался. При закрытии аортального клапана
возникает короткая фаза обратного движения крови, когда она
течет по направлению к аортальному клапану, а затем в течение
остальной части цикла сокращения сердца движение крови прак-
тически прекращается. Пиковое значение скорости крови зависит
от многих факторов (в острых опытах на животных не в послед-
нюю очередь от выбора средства для наркоза и от глубины по-
следнего); у здоровой бодрствующей собаки оно, по-видимому,
больше 1 м-с-1. (Значения, представленные в табл. I, собраны из
многих источников, и разброс их отражает влияние различных
факторов.) Ускорение крови в начале фазы изгнания приобре-
Гл. 12. Артерии большого круга 319
Рис. 12.17. Записи скорости и ускорения
крови в восходящей аорте человека. Кри-
вая ускорения получена дифференцирова-
нием сигнала скорости с помощью элек-
тронного устройства. [Bennett, Else, Miller,
Sutton, Miller, Noble (1974). Maximum
acceleration of blood from the left ventricle
of patients with ischaemic heart desease
Chn. Sci. mol. med., 46, 49—59.]
тается очень быстро, но оно кратковременно: картина такая, как
если бы по столбу крови нанесли удар молотком. В правомерности
такого сравнения можно убедиться, взглянув на рис. 12.17. Здесь
наряду с записью волны скорости крови приведена полученная од-
новременно запись ускорения и замедления ). Пиковое значение
ускорения крови у здоровой собаки составляет около 5—10 g и
может увеличиваться на 50% при усилении сокращений сердца
соответствующими веществами или во время мышечной работы.
Значения скорости крови и ее ускорения у человека точно не уста-
новлены, но, вероятно, они несколько ниже (см. рис. 12.17).
О соотношении во времени изменений давления и расхода крови
на входе в аорту позволяет судить рис. 12.18, на котором соответ-
*) Ускорение есть скорость изменения скорости, и поэтому его определяют
по наклону кривой скорости к оси времени. На языке математики ускорение
есть производная скорости и по времени: dn/dt. В опытах ускорение обычно оп-
ределяют, дифференцируя сигналы скорости кровотока при помощи электронного
устройства, напряжение на выходе которого пропорционально скорости измене-
ния входного сигнала. Такое же устройство можно применить и для того, чтобы
из сигнала давления извлечь сигнал скорости изменения давления.
320 Часть II Механика кровообращения
ствующие записи приведены в сопоставимом масштабе и наложены
друг на друга. В начале систолы давление и расход растут одно-
временно — факт, который указывает еще раз, что именно гра-
диент давления, а не само давление как таковое определяет дви-
жение крови. Далее в течение систолы синфазность изменений дав-
ления и расхода крови нарушается, что обусловлено главным обра-
зом появлением отраженных компонентов волны давления (этот
вопрос мы подробно разберем позднее).
Рис. 12.18. Записи давления (/) и расхода крови (II) в восходящей аорте собаки.
Масштаб выбран таким образом, чтобы амплитуды колебаний обеих величии
были примерно одинаковыми. [Kouchoukos, Sheppard, McDonald (1970). Esti-
mation of Stroke volume in the dog by a pulse contour method. Circulation Res.,
26, 611—623.1
Анализ формы волны скорости крови во все более удаленных от
сердца участках артериальной системы (нижние записи на
рис. 12.19) показывает, что амплитуда этой волны постепенно
уменьшается в противоположность увеличению амплитуды и кру-
тизны волны давления, на что мы уже обращали внимание (верх-
ние записи на рис. 12.19). Далее (разд. 12.3) мы постараемся
объяснить природу как этих, так и других уже упоминавшихся яв-
лений.
Мы убедимся, что в принципе по кривым пульсовых волн давле-
ния или скорости движения крови в корне аорты можно предска-
зать форму пульсовых волн давления и скорости кровотока в ка-
ком угодно участке сосудистого русла, если только свойства этого
русла известны достаточно подробно. В частности, давление в
корне аорты определяет скорость кровотока в нем, и наоборот. Бо-
лее того, хотя мы не знаем полностью геометрию сосудистого русла
и механические свойства кровеносных сосудов, имеющихся данных
все же достаточно, чтобы с определенной степенью точности пред-
сказать соотношение между давлением и расходом крови на про-
тяжении всей аорты.
Терминология. Поскольку движение крови во времени не-
стационарно и скорость по сечению артерии неодинакова (суще-
ствует профиль скорости), необходимо точно определить, что имен-
но мы будем понимать под скоростью. Если в некоторый момент
времени расход (объемная скорость) крови в артерии с площадью
Гл. 12. Артерии большого круга 321
Рис. 12.19. Сопоставление кривых давления и скорости крови в разных участках
артериального русла собаки. [McDonald (1974). Blood flow in arteries, Edward
Annold, London.]
поперечного сечения А равен Q, то в этот момент средняя по сече-
нию скорость равна Q/Л. Эта величина меняется во времени точно
так же, как и расход крови, и отличается от него только множи-
телем 1/Л. Именно эта мгновенная средняя скорость приведена на
большинстве тех рисунков данной главы, где изображена форма
волн скорости кровотока.
Однако в любой момент времени скорость движения крови по
любому данному сечению сосуда неодинакова, например из-за на-
личия пограничного слоя. В случае пуазейлевского течения
(разд. 5.1) скорость меняется от нуля у стенки сосуда до величины,
вдвое большей средней по сечению сосуда скорости, — на оси по-
тока. По причинам, которые мы обсудим ниже, в крупных арте-
риях профиль скорости почти плоский (см., например, рис. 12.39),
с узким пограничным слоем вблизи стенки. Вне этого пограничного
слоя жидкость перемещается почти с одинаковой скоростью, обра-
зуя «ядро потока», скорость в котором лишь ненамного больше
средней по сечению скорости. Поэтому для крупных артерий ско-
рость в ядре потока (например, скорость на оси потока) с доста-
точно хорошим приближением (с ошибкой, вероятно, не больше
10%) можно считать равной средней скорости по всему сечению
сосуда.
Показав, что путем усреднения по пространству можно полу-
чить среднюю по сечению сосуда скорость движения крови, ука-
жем теперь на возможность усреднения и по времени — в случае
нестационарного, т. е. непостоянного во времени течения. При этом
322 Часть II. Механика кровообращения
среднюю по времени скорость можно вычислить как для некоторой
точки в потоке, так и для всего сечения сосуда. Например, на
рис. 12.38 для нескольких сечений сосуда нанесены средние по вре-
мени скорости течения в ряде точек, отстоящих на разные расстоя-
ния от оси сосуда, и таким образом построен профиль средней по
времени скорости.
Наконец, в некоторых случаях, когда на записи пульсовых волн
скорости течения накладываются случайные флуктуации скорости,
обусловленные, например, турбулентностью, для того чтобы от-
сеять эти последние, применяют усреднение особого рода, называе-
мое усреднением по ансамблю. Сущность такого усреднения изло-
жена в разд. 12.8.
Анализ Фурье. Ценность этого метода для исследования свойств
волн давления и скорости кровотока в крупных артериях на-
столько велика, что стоит не только вкратце повторить то, что уже
было сказано об анализе Фурье в главе, посвященной колебаниям
и волнам (разд. 8.6), но и дополнить этот материал. Как отмеча-
лось, анализ волновых процессов существенно упрощается, если
волна имеет форму синусоиды. Однако, судя по многим приведен-
ным в этой главе кривым, ни пульсовые волны давления, ни волны
скорости кровотока в крупных артериях не являются, к сожале-
нию, синусоидальными, а имеют сложную форму.
Анализ Фурье основан на том, что любое сложное колебание,
подобное пульсовым волнам артериального давления, может быть
представлено в виде некоторого набора (суммы) синусоидальных
составляющих, равно как и восстановлено по ним. Каждое из этого
набора синусоидальных колебаний (называемых гармониками)
имеет свою частоту и амплитуду, и все они сдвинуты одно относи-
тельно другого по фазе. Этот прием разложения ценен тем, что ма-
тематические операции над представляющими отдельные гармони-
ки синусоидальными функциями сравнительно просты. Например,
основываясь на несложной теории, описанной ниже, для каждой из
ряда синусоидальных волн давления в артерии можно рассчитать
соответствующую синусоидальную волну расхода. Далее, сложив
все синусоидальные волны расхода, каждая из которых характе-
ризуется своей частотой, амплитудой и фазой, можно построить
суммарную волну расхода сложной формы и сравнить ее с наблю-
даемой в опыте. Равным образом, зная форму волны давления в
некотором месте артерии, в принципе можно рассчитать форму
волны давления или скорости кровотока в другом месте артериаль-
ной системы. Это, правда, возможно только в том случае, если из-
вестны геометрия и упругие свойства артерий на соответствующем
участке системы, и могут быть рассчитаны особенности распро-
странения каждой синусоидальной составляющей. (Разрыв между
тем, что возможно «в принципе», и тем, что удается осуществить
на практике, в значительной степени определяется неполнотой на-
ших знаний о геометрии и упругих свойствах артерий.) Основное
Гл. 12. Артерии большого круга 323
ограничение названных приложений анализа Фурье обусловлено
тем, что результат оказывается правильным только в том случае,
когда изучаемая система линейна. Если же система нелинейна, от-
дельные гармоники не являются независимыми и сложение сину-
соидальных волн не позволяет предсказать свойства сложной
волны1). Это очень важное ограничение, и оно подробно обсуж-
дается в разд. 12.3. Как будет показано, поведение артерий в фи-
зиологических условиях с хорошей точностью можно считать ли-
нейным.
Проводить разложение в ряд Фурье «вручную» — дело очень
трудоемкое, поэтому сейчас эту операцию осуществляют с помощью
ЭВМ. Частоты гармоник, на которые должна быть разложена ана-
лизируемая волна, предписываются фактической частотой повторе-
ний самой этой волны (основной частотой). Частота первой гармо-
ники равна основной частоте, а частота второй и последующих гар-
моник — произведению основной частоты на номер гармоники. Так,
если основная частота равна 2 Гц, то частота второй гармоники
будет 4 Гц, третьей — 6 Гц и т. д. С увеличением номера гармоник
их амплитуды, вообще говоря, постепенно уменьшаются и в конце
концов становятся столь малыми, что ими можно пренебречь. Что-
бы проверить, достаточно ли полон в этом отношении полученный
набор гармоник ряда Фурье, следует сложить все гармоники и
сопоставить результат с исходной формой волны. Такую процедуру
повторяют несколько раз, добавляя все более высокие гармоники
до тех пор, пока не будет получено удовлетворительное совпадение.
Если амплитуда исходной волны ( например, волны давления кро-
ви) колеблется вокруг ненулевого среднего (по времени) значения,
то этот средний уровень, естественно, необходимо включить в ряд
Фурье в виде постоянной составляющей.
Волны давления и расхода крови в аорте могут быть с очень
хорошей точностью представлены в виде суммы постоянной состав-
ляющей и примерно 10 гармоник2). Амплитуды этих гармоник
вплоть до десятой приведены на рис. 12.20. Как и следовало ожи-
дать, в случае волны давления наибольшую амплитуду имеет по-
стоянная составляющая; низкоамплитудные высшие гармоники не-
*) Разумеется, любое колебание можно разложить на гармоники. Но для
нелинейной системы не имеет смысла (или, по меньшей мере, затруднительно)
оперировать с отдельными гармониками В самом деле, если величины а и Ь
связаны линейно, то точно такая же связь есть и между их гармониками и, ста-
ло быть, b можно легко реконструировать по гармоникам а. Если же линейная
связь отсутствует, то каждая гармоника b своим собственным образом (обычно
очень сложным для высоких гармоник) зависит от многих гармоник а. Ясно,
что при этом принципиальная возможность реконструкции Ь сохраняется, но те-
ряется главное преимущество анализа Фурье — практическая простота реали-
зации. — Прим. ред.
2) Это соответствует частотам гармоник вплоть до 20 Гц для собаки и до
10 Гц для человека. Следовательно, характеристики измерительной и регистри-
рующей аппаратуры должны быть известны вплоть до этих частот.
324 Часть II. Механика кровообращения
Рис. 12.20. Результат разложения сигналов давления и расхода крови в корне
аорты на гармонические составляющие (средние данные по восьми собакам).
Указано стандартное отклонение. Штриховой горизонтальной линией обозначен
уровень шума регистрирующих приборов [Patel, de Freitas, Fry (1963). Hydraulic
input impedance to aorta and pulmonary artery in dogs. J. appl. Physiol, 18,
134—140.]
обходимы главным образм для описания резких колебаний
при развитии дикротического зубца. В случае волны расхода
крови постоянная составляющая относительно невелика, а ам-
плитуда основной гармоники вносит значительно больший вклад.
Здесь высшие гармоники позволяют описать очень крутой рост
кровотока в начале систолы и его резкие колебания во время фазы
«обратного» кровотока в конце систолы. Убедительным свидетель-
ством точности, с которой сумма постоянной составляющей и де-
сяти гармоник отображает форму исходных волн, является резуль-
тат синтеза этих волн, представленный рис. 12.21. По мере удале-
ния от сердца волны давления и расхода крови сглаживаются и
могут быть достаточно точно описаны меньшим числом гармоник.
В разд. 8.6, например, приближенная форма волны расхода была
восстановлена по четырем гармоникам.
В заключение необходимо сделать одно замечание: с помощью
анализа Фурье нельзя получить больше, чем было заложено.
Иными словами, если форма исходной волны по тем или иным при-
чинам искажена (например, из-за несовершенства приборов), то
разложение в ряд Фурье передаст эти искажения. Предположим,
что мы регистрируем артериальное давление с помощью датчика,
частотная характеристика которого такова, что он может «отсле-
живать» частоты не выше пятой гармоники. В таком случае при
Гл. 12. Артерии большого круга 325
разложении зарегистрированного сигнала в ряд Фурье может со-
здаться впечатление, что амплитуды всех гармоник выше пятой
несущественны. Но когда на основании испытаний датчика мы
точно знаем, что последний способен надежно охватывать область
частот выше пятой гармоники, ситуация будет иной. Если и в этом
случае анализ Фурье обнаружит только пять первых гармоник
с достаточно большой амплитудой, мы безусловно можем быть
уверены в том, что не введены в заблуждение и зарегистрировали
истинную форму волны. Фактически такой прием позволяет в на-
стоящее время проверять надежность записи формы волн, да и
ранее он был одним из самых полезных применений анализа
Фурье. Другое, примыкающее к этому применение данного метода
заключается в коррекции формы зарегистрированных волн. Рас-
полагая датчиком давления, который с известным нам искажением
воспроизводит определенные частоты (это достаточно распростра-
ненная ситуация), мы можем установить истинную форму волн.
Для этого зарегистрированные кривые необходимо подвергнуть
анализу Фурье и, скорректировав амплитуду и фазу гармоник,
приходящихся на область искажений, восстановить путем синтеза
истинную форму волн. Таким образом, анализ Фурье служит
исключительно полезным инструментом исследования, а также ос-
новой для выяснения свойств волн давления и расхода крови в
артериях.
Рис. 12.21. Форма волн давления и рас-
хода крови в восходящей аорте. Штрихо-
вой линией изображены результаты прямого
измерения этих величии, а сплошной —
кривые, полученные суммированием постоян-
ной составляющей и первых десяти гар-
моник ряда Фурье. Видно, что кривые, по-
лученные синтезом фурье-составляющих, с
высокой точностью совпадают с результа-
тами непосредственных измерений. [McDo-
nald (1974). Blood flow in arteries, Edward
Arnold, London.]
326 Часть П. Механика кровообращения
12.3. Распространение волн в артериях
При изучении механики движения крови в крупных артериях
удобно и физически оправданно считать «входом» в систему пуль-
совое колебание давления в корне аорты (рис. 12.18). Избыточное
давление (рИзб. а) колеблется здесь относительно среднего значе-
ния, равного примерно 1,3-104 Н-м-2 (100 мм рт. ст.). Эта вели-
чина много больше избыточного давления (рИзб, в) в полой вене,
которое близко к нулю. Именно артерио-венозная разность давле-
ний ризб. а — Ризб, в и вызывает движение крови. Если бы сосуди-
стая система представляла собой единственную длинную прямую
жесткую трубку, течение в которой является пуазейлевским, то в
каждый момент времени во всех участках такой трубки расход Q
был бы одинаков и прямо пропорционален разности давлений на
концах трубки. Сказанное означает, что
Ризб, а Ризб, в == " Q> (12.2)
где R — постоянное сопротивление, a Q, рИЗб, а и рИЗб, в меняются
во времени [см. уравнение (5.1)]. Этот вывод оказался бы примерно
таким же и для более сложной сети трубок при условии, что все
они жесткие, а числа Рейнольдса достаточно малы, чтобы влияние
явлений на начальном участке (разд. 5.3) и вторичных течений
было пренебрежимо малым (разд. 5.8). Будь система кровообра-
щения совокупностью жестких трубок, форма пульсовых волн дав-
ления в каждой ее точке была'бы подобна форме волны давления
в корне аорты и эти волны возникали бы везде одновременно
с последней, но их амплитуда уменьшалась бы соответственно по-
ложению данной точки относительно входа. На самом же деле та-
кая картина не может служить удовлетворительным приближением
к действительности уже потому, что длина начального участка при
характерных для артерий значениях чисел Рейнольдса велика [см.
уравнение (5.4)]. Но и при любых числах Рейнольдса, будь си-
стема жесткой, существенные изменения давления возникали бы
в ней повсюду почти одновременно, а объемная скорость притока
жидкости в любую часть такой системы была бы всегда равна
объемной скорости оттока из нее.
Модель упругого резервуара. На самом деле нигде в реальной
сосудистой системе ни форма волн давления, ни их ход во вре-
мени не совпадают (рис. 12.14). Временной ход волн расхода
крови (или средней по сечению скорости крови) в полой вене со-
всем не таков, как в аорте, и нигде пульсовые изменения скорости
не пропорциональны пульсовым колебаниям разности давлений.
Причина этих явлений кроется в растяжимости кровеносных сосу-
дов. Когда сердце изгоняет кровь и давление в крупных артериях
растет, они растягиваются. При последующем падении давления
диаметр артерий уменьшается, благодаря чему расход через мел-
кие периферические сосуды не снижается тотчас до нуля. Впервые
Гл. 12. Артерии большого круга 327
мысль о таком действии артерий, запасающих кровь во время си-
столы и выталкивающих ее в мелкие сосуды во время диастолы,
высказал в 1733 г. Хейлс (сельский священник, который, кстати,
первым измерил кровяное давление). Но только в 1899 г. немецкий
физиолог Франк теоретически развил эту идею.
В модели Франка (рис. 12.22) артерии представлены в виде
растяжимой камеры (ее называют еще упругим резервуаром),
объем которой V пропорционален избыточному давлению в ней.
Рис. 12.22. Модель упругого резервуара, состоящая из растяжимой камеры,
объем которой V пропорционален избыточному давлению рИзб в ней. Расход Q
жидкости, вытекающей из камеры, пропорционален разности между артериаль-
ным и венозным давлениями (рИзв, а — рИЗб, в), причем коэффициент пропорцио-
нальности есть сопротивление системы микрососудов. Расход жидкости, посту-
пающей из сердца в систему, Qc, может отличаться от Q.
Система микрососудов представлена в этой модели как единствен-
ное постоянное сопротивление, расход крови через которое Q прямо
пропорционален разности давлений на его концах. Уравнение
(12.2)., таким образом, описывает связь между расходом крови че-
рез систему микрососудов и артерио-венозной разностью давлений.
Если учесть, что избыточное давление в венах много меньше, чем
в артериях, то, не слишком изменяя выводы, следующие из модели,
давление в венах можно считать равным нулю, и тогда:
Ризб, a = ^Q- (12.3)
Объем артериальной камеры связан с давленйем в ней соотноше-
нием
Ризб,а=У/С, (12.4)
где С — податливость камеры, считающаяся постоянной (в гЛ. 7
убедительно показано, что это слишком сильное упрощение даже
для единственной артерии). Наконец, скорость увеличения объема
камеры dV/dt должна равняться разности скоростей притока в Нее
крови из сердца Qc и оттока крови в систему микрососудов Q:
-^ = Qc-Q. (12.6)
Три уравнения—(12.3), (12.4) и (12.5)—могут быть объединены
в одно, содержащее одну неизвестную — рИЗб, а, V или Q. Решение
328 Часть II. Механика кровообращения
должно дать взаимосвязь между давлением в артериях и притоком
крови в них из сердца для любого момента времени.
Из модели следует, что в течение диастолы, когда приток фак-
тически равен нулю, давление в артериях снижается во времени
экспоненциально, т. е.
рИзб,аОсе-ВДс, (12.6)
и такой характер спада давления близок к наблюдаемому в дей-
ствительности. Однако в начале систолы, когда приток крови Qc
велик, согласно теории, пропорционально ему должна возрастать
скорость повышения давления брИз($, a/dt; опыты же показывают,
что пропорционально притоку растет само давление (точнее, ве-
личина, на которую оно превышает давление в конце диастолы
Ризб, д), т. е.
Ризб. а Ризб, д °* Qc-
Это расхождение показывает, что модель упругого резервуара не
в состоянии точно отобразить динамические процессы даже в круп-
ных артериях. Конечно, простота этой модели привлекает, но ей
присущ один принципиальный недостаток: она не учитывает того,
что пульсовые колебания давления перемещаются по артериальной
системе в виде волны.
Распространение волны давления. Этот недостаток модели
упругого резервуара вытекает из предположения, что все артерии
растягиваются одновременно. Как мы уже выяснили (рис. 12.19),
в периферических артериях максимум давления достигается позже,
чем в аорте, и пульсовое колебание давления распространяется
по сосудам в виде волны. Когда сердце изгоняет кровь в началь-
ный участок аорты, давление здесь повышается, стенка этого
участка растягивается и напряжение в ней возрастает. По мере
снижения скорости изгнания крови давление в аорте начинает па-
дать и растянутая стенка возвращается к равновесному положе-
нию. Кровь по инерции продолжает двигаться вперед и после того,
как обусловивший ее движение градиент давления уменьшился.
Последнее приводит к тому, что стенка наиболее близкого к сердцу
участка аорты «проскакивает» равновесное положение и начи-
нается ее колебательное движение. В это же самое время стенка
следующего участка аорты оказывается растянутой. А так как и
этот участок вследствие упругих свойств стремится вернуться в ис-
ходное состояние, вытесняемая из него кровь растягивает соседний
участок стенки, который затем тоже возвращается назад благо-
даря упругости и так далее. Таким образом, по артериальной си-
стеме распространяется возмущение в виде волны давления. Этот
процесс подобен распространению волны в натянутой струне, опи-
санному в гл. 8 (разд. 8.2). Между возвращающей силой, обеспечи-
ваемой упругостью стенки артерий, и гидродинамическими силами,
обусловленными инерцией крови, существует равновесие; незначи*
Гл. li. Артерии большого круга 329
тельный вклад в процесс вносит и инерция самой стенки. Как и
волны, о которых говорилось в гл. 8, волна давления в артериях
может распространяться в любом из двух направлений. Однако в
артериальной системе большого круга волна давления зарож-
дается в сердце и перемещается от него в дистальном направле-
нии; правда, как мы узнаем дальше, на нее накладываются компо-
ненты, отраженные от периферических сосудов.
Обусловленные волной давления радиальные движения стенки
сосуда могут также вызывать некоторые ее продольные переме-
щения, поскольку она подвергается продольному растяжению или
сжатию, но такие перемещения имеют второстепенное значение и
их влияние на распространение волны пренебрежимо мало. Кроме
того, можно представить себе волны совсем другого рода, когда
колебательные перемещения стенки исходно являются продоль-
ными и благодаря вязкости вызывают продольные движения жид-
кости в пограничном слое, преодолевая ее инерцию. Однако физио-
логические доказательства существования таких волн отсутствуют,
и, по-видимому, они не оказывают существенного влияния на ре-
гистрируемые величины давления и скорости крови '). Подробнее
волны такого рода будут рассмотрены в разд. 12.7.
Скорость волны. В гл. 8 мы установили, что скорость рас-
пространения волны с в натянутой струне определяется возвра-
щающей силой (натяжением струны F) и инерцией (массой на
единицу длины, М) и равна
(12.7)
Следовательно, при одном и том же натяжении чем больше масса
струны, тем меньше скорость распространяющейся по ней волны,
и наоборот. Если струну натянуть сильнее, то возвращающая сила
увеличится и скорость возрастет. Подобным же образом возвра-
щающая сила и инерция определяют скорость распространения
волн давления в кровеносных сосудах. Инерция обусловлена мас-
сой крови (и стенки сосуда) и может быть охарактеризована плот-
ностью крови р. Возвращающая сила обязана своим происхожде-
нием упругости стенки сосуда; если растяжимость ее велика, ско-
рость волны низкая, и наоборот, если стенка жесткая, скорость
высокая. В гл. 7 мы говорили о том, что самой прямой мерой
жесткости стенки сосуда является величина, которая была опре-
делена как растяжимость D. Эта величина представляет собой
меру относительного увеличения площади поперечного сечения
*) В принципе возможно существование еще одного типа волн в артериях,
а именно крутильных упругих волн, бегущих по стенке и возникающих благо-
даря тому, что иа нее вблизи сердца могут действовать периодические крутящие
моменты (из за некоторых движений сердца и вторичных течений в дуге аор-
ты) — Прим ред г
330 Часть II. Механика кровообращения
(ДЛ/Л) участка артерии неизменной длины, вызываемого неболь-
шим повышением давления ДрИзб, т. е.
0=4- • (12.8)
Л Лризб
Далее можно вновь обратиться к тем опирающимся на анализ
размерностей рассуждениям, которые в гл. 8 привели к уравнению
вида (12.7), и показать, что скорость распространения волн давле-
ния в кровеносных сосудах пропорциональна (р£))~1/2. Подробный
математический анализ показывает, что для упрощенной модели
артерии это действительно так и что коэффициент пропорциональ-
ности, как и в уравнении (12.7), равен единице, а потому
с = (рП)-1/2_ (12.9)
Когда D уменьшается, с увеличивается, и наоборот, т. е., как и сле-
довало ожидать, по более жесткой артерии волны распростра-
няются быстрее.
Рассуждения, которые приводят к формуле (12.9), опираются
на два основных принципа — закон сохранения массы и второй
закон Ньютона (сила = масса X ускорение). Эти принципы при-
менены для описания движения жидкости в бесконечно длинной
растяжимой трубке, диаметр которой не изменяется, пока к ней
не прикладываются возмущения, подобные пульсовому колебанию
давления. Такая трубка уже представляет собой очень сильно уп-
рощенную модель артерии. Но чтобы вывести уравнение (12.9),
необходимо сделать еще два упрощающих предположения: 1) вяз-
кость крови не влияет на ее движение и 2) амплитуда возмущения,
т. е. пульсового колебания давления, достаточно мала, и потому
соотношения, описывающие упругие свойства стенки трубки и дви-
жение жидкости, линейны. Проверим, обоснованны ли эти предпо-
ложения.
Пренебрежение вязкостью. По определению вязкость играет су-
щественную роль только в тех областях потока, где жидкость под-
вергается сдвигу. Скорость сдвига принимает наибольшие значе-
ния в пограничном слое вблизи стенки сосуда, так как скорость
движения жидкости непосредственно у стенки равна нулю. При
пульсирующем течении в жесткой трубке толщина пограничного
слоя равна расстоянию от стенки, на которое распространяется су-
щественное влияние вязкости в течение одного периода колебания.
Следовательно, толщина пограничного слоя пропорциональна Vv/®,
где v — кинематическая вязкость жидкости, <о — угловая частота
колебаний. Поэтому параметр, определяющий, какая часть сечения
трубки занята пограничным слоем, равен отношению радиуса
трубки yd к -y/v/a). Это и есть параметр Уомерсли ос:
Гл. 12. Артерии большого круга 331
определение которому было дано в гл. 5. Смысл параметра а
можно представить и по другому: заметим, что величина а2 равна
отношению времени, в течение которого действие вязких сил рас-
пространяется на всю ширину трубки (d2/4v), к периоду колебаний
(1/<о). Параметр а пригоден для характеристики процессов и в
упругих трубках.
При больших а пограничный слой тонкий, и объем жидкости,
движение которой замедлено из-за ее прилипания на стенке
трубки, много меньше объема, приводимого в движение волной
давления. Расчет а для крупных артерий показывает, что у всех
(кроме самых мелких) млекопитающих при колебаниях потока
крови с частотой, равной частоте сокращений сердца (основная
гармоника), значения этого параметра много больше единицы.
Типичное значение а для аорты человека равно 20, собаки — 14,
кошки — 8, крысы — 3. Значения, характерные для других артерий
собаки, приведены в табл. I, а соответствующие значения для ар-
терий человека обычно примерно на 50% больше. Таким образом,
пренебрежение вязкостью крови при вычислении скорости распро-
странения волн давления в крупных артериях приводит, по-види-
мому, лишь к незначительным ошибкам. Насколько велики эти
ошибки в действительности и какие другие явления могут быть
связаны с вязкостью крови, мы обсудим в разд. 12.6.
Предположение о линейности. Как говорилось в гл. 8
(разд. 8.5), линейность означает, что синусоидальное колебание
давления какой-то одной частоты вызывает синусоидальное коле-
бание скорости потока такой же частоты. Есть две возможные
причины нелинейности, которые могут быть существенными для
процесса распространения пульсовых волн. Обе эти причины про-
истекают из того обстоятельства, что колебания давления сопро-
вождаются изменением площади поперечного сечения сосуда. Из
менения площади приводят к тому, что скорость течения жидкости
в разных участках трубки оказывается различной, и, следователь-
но, возникают не только местные, но и конвективные ускорения
(разд. 4.5) и соответствующие силы инерции, которые должны
уравновесить градиент давления. Именно эти конвективные уско-
рения нелинейны: они выражаются через квадрат скорости, а не
скорость как таковую, что уже обсуждалось в разд. 8.5 в связи
с уравнением Бернулли (4.6). Это первая причина нелинейности,
но нелинейность такого типа становится несущественной, если кон-
вективные ускорения малы по сравнению с местными. Для выпол-
нения этого условия необходимо, чтобы характерное время значи-
тельных местных изменений скорости течения жидкости (т. е. пе-
риод колебаний 2 л/со) было меньше времени, за которое элемент
жидкости конвективно перемещается в другую точку, где скорость
течения существенно иная Характерное время такого перемещения
равно длине волны 2лс/о>, отнесенной к характерной скорости жид-
332 Часть 11. Механика кровообращения
кости й. Таким образом, при 1/со, много меньших с1и>й, т. е. при
Ъ (12.10)
местные ускорения всегда преобладают. Как будет показано ниже,
для крупных артерий это условие примерно выполняется.
Вторая причина нелинейности заключается в том, что артерии
при растяжении становятся более жесткими (см. рис. 7.5, Л). Рас-
суждения, приводящие к уравнению (12.9), исходят из предполо-
жения о линейности, и, следовательно, это уравнение применимо
только в том случае, если растяжимость D остается примерно по-
стоянной на протяжении всего цикла движения стенки артерии.
Однако с ростом растягивающего давления D уменьшается, по-
этому для выполнения условия линейности необходимо, чтобы ам-
плитуда пульсового колебания давления была мала по сравнению
со средним давлением.
В действительности при обычных условиях отношение этих дав-
лений в артериях большого круга близко к 0,2, и потому возни-
кает вопрос, достаточно ли оно меньше единицы, чтобы систему
можно было приближенно считать линейной. Примерно таково же
(~ 0,25) для обычных условий максимальное значение отношения
й/с, и здесь возникает такой же вопрос. Далее (разд. 12.5) мы
увидим, что нелинейности приводят к ощутимым эффектам; эти
нелинейности несущественны для оценки скорости волны, но сказы-
ваются на форме пульсовых волн давления и расхода крови.
Сравнение теории с экспериментом. Измерить растяжимость
D участка артерии сравнительно несложно, особенно в статических
условиях: для этого необходимо шаг за шагом повышать растяги-
вающее давление и регистрировать вызванные им изменения пло-
щади. Затем, поскольку плотность крови известна, можно, поль-
зуясь формулой (12.9), вычислить скорость волны.
На первый взгляд измерение скорости распространения волны
давления также не составляет особого труда. Для этого требуется
зарегистрировать пульсовые колебания давления в двух участках
сосуда, установить, на каком расстоянии друг от друга находятся
эти участки, и определить время за которое определенная точка
волны давления (например, ее максимум или «подножие», т. е. на-
чало фронта) пробегает это расстояние. Если бы в процессе рас-
пространения по сосуду волна сохраняла свою форму, то результат
такого измерения был бы однозначен. Однако форма пульсовой
волны меняется. Как будет показано ниже, это связано с отраже-
ниями волны от мест разветвления артерий и от самих ветвей,
а также с непрерывным сужением («конусностью») каждой арте-
рии. Форма волны изменяется еще и потому, что различные ее со-
ставляющие имеют разные частоты *) и, следовательно, могут про-
*) Выявляемые путем разложения аолны в ряд Фурье — см. разд. 86 и 12.2.
Гл. 12. Артерии большого круга 333
бегать один и тот же участок сосуда с неодинаковой скоростью и
затухать в разной степени Наличие отражения означает, в част-
ности, что кажущаяся скорость волны, определенная для участка
артерии, может не совпадать с собственной скоростью, с которой
волна распространялась бы в бесконечно длинной трубке, обла-
дающей такими же упругими свойствами. Но в таком случае зна-
чения скорости, оцененные из уравнения (12.9), нельзя сравнивать
с результатами, получаемыми в опытах. На самом деле, однако,
оказывается, что если определять скорость волны для начальной
части крутого фронта волны, то отраженная волна не успевает вер-
нуться и потому нигде не искажает форму этой части волны, кроме
как в артериях, расположенных непосредственно вблизи области
отражения (разд. 12.4). Следовательно, значения скорости волны,
вычисленные для начального участка фронта волны («от подно-
жия до подножия»), можно использовать в качестве меры соб-
ственной скорости распространения волны давления в артериях.
Следует, однако, помнить, что упругие свойства артерии на участке
между местами, выбранными для измерения скорости волны, могут
существенно различаться (если, например, на этом участке отходят
ветви), и потому скорость волны на данном участке будет непо-
стоянной и возможно измерение только средней скорости волны.
Вычисляя скорость волны, многие исследователи используют
видоизмененное уравнение (12.9), вводя в него касательный мо-
дуль Юнга Е стенки артерии (разд. 7.2). Тем самым они приме-
няют простую модель этой стенки, в которой ее упругие свойства
(растяжимость) уподобляются упругим свойствам тонкой однород-
ной оболочки. В таком случае D определяется уравнением (7.36)
E(h/d) ’
где h/d — отношение толщины стенки к диаметру сосуда. Подстав-
ляя это выражение в уравнение (12.9), получаем формулу для оп-
ределения скорости волны:
c0 = (Eft/pd)1/2, (12.11)
где символ Со выбран для обозначения вычисляемой именно таким
способом скорости. Это хорошо известное соотношение называется
формулой Моенса — Кортевега, по имени двух голландских уче-
ных, опубликовавших его в 1878 г. (фактически впервые его полу-
чил Томас Юнг в 1809 г.). Так как для вывода формулы (12.11)
использована простая модель упругой стенки артерий, на практике
она скорее всего приведет к менее точным результатам, чем урав-
нение (12.9). Ошибки могут возникать при определении тем или
иным способом значения модуля Юнга Е, а главным образом из-за
неточностей измерения отношения h/d. Однако при условии, что эти
величины определяют по результатам измерений растяжимости,
использование формулы (12.11) оправданно.
334 Часть II. Механика кровообращения
Значения скорости волны, полученные с помощью соотношения
(12.11), (со), и измеренные непосредственно (с) приведены в табл. I.
Наиболее удивительным при сравнении этих величин представ-
ляется то, что, несмотря на принятые при выводе уравнения (12.11)
допущения о линейности, упругости системы, роли вязкости и т. д.,
скорость с0 для артерий отличается от скорости с не более чем на
15%, что вполне укладывается в пределы обычных ошибок измере-
ний и изменчивости физиологических величин. Это означает, что
принятые упрощения не приводят к большим неточностям при вы-
числении скорости волны.
Точность линейного приближения. Используя приведенные в
табл. 1 значения измеренной скорости волны с и пиковой скорости
кровотока йщах, можно проверить, насколько хорошо выполняется
в действительности допущение о том, что й/с всегда меньше еди-
ницы. Максимальные значения й/с для аорты и легочного ствола
равны 0,2—0,25, что, конечно, меньше единицы, но не пренебре-
жимо мало по сравнению с ней. Однако в случае более перифери-
ческих артерий отношение й/с меньше: скорость кровотока здесь
падает, а скорость волны возрастает, поскольку к периферии по-
вышается жесткость стенок артерий (см. табл. 1). Таким образом,
для обычных условий предположение о линейности, принятое при
анализе распространения пульсовых волн, справедливо для всех
артерий собаки (и человека), кроме самых крупных, что под-
тверждает и простая оценка отношения амплитуды пульсовых ко-
лебаний давления ДрИзб к среднему избыточному давлению рИзб
в данной артерии. Это отношение для крупных артерий изменяется
примерно от 0,2 (восходящая аорта) до 0,4 (бедренная артерия,
в которой амплитуда колебаний пульсового давления наибольшая),
но для мелких артерий оно много меньше.
При некоторых патологических состояниях отношение й/с для
крупных артерий может принимать значения, весьма близкие
к единице. К таким состояниям относится, например, редкое забо-
левание системы соединительной ткани, одним из признаков кото-
рого является вялость артериальной стенки, когда скорость волны
становится низкой; встречаются также состояния, при которых
увеличен ударный объем (а именно атрио-вентрикулярная блокада,
недостаточность аортального клапана и шунты крупных артерий,
например незаращение боталлова протока). При всех этих со-
стояниях передний фронт волны давления оказывается очень кру-
тым, а скорость кровотока значительно выше нормальной вели-
чины.
Другие ограничения простой упругой модели. Согласно изло-
женной выше теории, волна давления должна была бы распростра-
няться по артерии, не изменяя своей формы. Однако записи, при-
веденные на рис. 12.14 и 12.19, свидетельствуют об изменении
формы волны давления в ходе ее перемещения по артериальной
системе Кроме того, из этой простой теории следует, чго форма
Гл. 12. Артерии большого круга 335
пульсовой волны скорости кровотока (зависимость и от t) должна
быть такой же, как форма волны давления, а амплитуда в рс раз
меньше1). На самом же деле форма волны скорости совершенно
отлична от формы волны давления (рис. 12.18), и амплитуда ее
при удалении от сердца уменьшается, тогда как амплитуда пуль-
совой волны давления в крупных артериях растет (рис. 12.19). Это
означает, что, хотя изложенная выше теория позволяет достаточно
точно найти скорость волны, и предположение о линейности можно
считать в этом смысле обоснованным, некоторые другие допуще-
ния, положенные в основу этой простой теории, неправомерны. Мы
должны принять во внимание четыре реально существующих фак-
тора, которые изменяют выводы, следующие из простой теории.
1. Трубка предполагалась бесконечно длинной, тогда как кро-
веносные сосуды относительно короткие, поэтому происходит отра-
жение на концах. В самом деле, в случае аорты человека длина
волны основной гармоники составляет примерно 4 м (скорость
волны равна приблизительно 5 м-с-1, частота — около 1,2 с-1), в то
время как длина аорты — примерно 0,5 м. Следовательно, в каж-
дое мгновение на такой длине может уложиться лишь небольшая
часть всей волны. Именно это обстоятельство позволило Франку —
исследователю, хорошо знакомому с фактами, касающимися тео-
рии распространения волн, отстаивать свою модель упругого ре-
зервуара. Роль отражения волн мы обсудим в следующем разделе.
2. Свойства всех артерий неоднородны по длине. В частности,
аорта на дистальном конце более узкая и более жесткая, чем
вблизи сердца (см. табл. I). Именно «конусность» аорты, как до-
казано, отчасти ответственна за увеличение амплитуды пульсо-
*) Это связано с тем, что, согласно уравнению движения, произведение плот-
ности жидкости на местное ускорение pdu/df должно равняться градиенту избы-
точного давления — dpKS(,/dx. Как й, так и "рИЗб могут быть представлены в виде
суммы постоянной составляющей и синусоидальных функций вида
В cos {<о (f — х/с) — 0}, (12.12)
где В — амплитуда, равная амплитуде й0 или ро соответственно. Чтобы из ско-
рости и давления получить ускорение и градиент давления, продифференцируем
каждую из этих составляющих:
р = — ®йор sin {<о (t — х/с) — 0),
= ^£0. sin {<в {z _ х/с) _ в}.
Так как левые части этих выражений должны быть равны, то йо должно рав-
няться ра/рс. Если речь идет о волне, распространяющейся в противоположном
направлении, то знак при х/с в выражении (12.12) должен быть изменен, тогда
Йо будет равно — р0/рс — Прим. авт.
Добавим, что в (12 12) сначала нужно было бы угол 0 принять равным 0!
для й и 02 — для ризе После приравнивания правых частей двух последующих
уравнений мы получим, кроме выражения для й0, еще и равенство 0| = 02,
означающее совпадение по фазе ускорения и градиента давления. — Прим, ред.
836 Часть II. Механика кровообращения
вых колебаний давления (см., например, рис. 12.19). Следствие
конусности артерий и повышения их жесткости по мере удаления
от сердца также будет рассмотрено в следующем разделе.
3. О нелинейных свойствах артерий уже упоминалось. Обстоя-
тельства, при которых такие свойства существенны, обсуждаются
в разд. 12.5.
4. Наконец, следует напомнить о том, что всякое движение вяз-
кой жидкости и вязкоупругого твердого тела связано с диссипа-
цией энергии. Как указывалось в разд. 8.3, это приводит к затуха-
нию волн. Роль этого явления в распространении пульсовых коле-
баний мы обсудим в разд. 12.6.
12.4. Отражение и прохождение волн в местах ветвления артерий
Любой участок артерии, на котором изменяются ее свойства,
является местом частичного отражения волны давления, подобно
тому как всякая неоднородность натянутой струны служит источ-
ником отраженных волн (разд. 8.4). В каждой артерии на всем
ее протяжении имеются многочисленные неоднородности и неболь-
шие изгибы, приводящие в той или иной степени к отражению
волн, однако наиболее четко выраженные неоднородности, где от-
ражение особенно существенно — это места ветвления артерий
(рис. 12.1). Именно отражение ответственно за наиболее значи-
тельное изменение формы распространяющейся к периферии пуль-
совой волны. Вот почему мы должны подробно проанализировать
это явление, начав с отражения от отдельно взятого ветвления,
а затем рассмотрев отражение волн от последовательности неод-
нородных структур.
Отражение на одном ветвлении. Рассмотрим сначала случай,
когда артерия делится на две ветви (рис. 12.23). Диаметры этих
ветвей могут различаться (в сосудистой системе это обычное яв-
ление), а сами ветви могут отходить от порождающего их ствола
Рис 12.23. Схема бифуркации артерии. Падающая волна частично отражается
у осцовной ствол О, а частично проходит в ветви 1 и 3.
Гл. 12. Артерии большого круга 337
под разными углами. Волна, распространяющаяся по стволу, ча-
стично отражается от места ветвления (причем отраженная волна
бежит назад, в общий ствол), а частично проходит в его ветви.
Предсказать свойства отраженной и прошедшей волн весьма не-
сложно.
В месте ветвления при прохождении и отражении волн должны
выполняться два условия. Во-первых, давление во всех трех сосу-
дах должно быть строго одинаковым: даже очень малая разность
давлений, действуя на малый объем крови, мгновенно сообщила
бы ему весьма большое ускорение, что свело бы разность давлений
к нулю. Во всех трех сосудах должны быть одинаковы не только
значения среднего давления, но и переменные составляющие волны
давления (в дальнейшем эти переменные составляющие мы будем
обозначать символом р). Если переменная (пульсирующая) состав-
ляющая падающей волны давления равна рпад, отраженной — ротр
и прошедших волн — рпР11 и рпр, г, то для любого момента времени
должно выполняться условие
Рпад “Ь Ротр == Рпр» 1 == Рпр» 2* (12.13)
Во-вторых, должен выполняться закон сохранения массы: ко-
личество крови, втекающее в единицу времени в область ветвле-
ния, всегда должно равняться количеству крови, вытекающему из
нее, т. е. в каждый момент времени объемная скорость притока
крови из общего сосуда в область ветвления должна равняться
сумме объемных скоростей ее оттока в отходящие от этой области
сосуды. (Объем крови, содержащийся собственно в месте ветвле-
ния, считается столь малым, что любыми его изменениями можно
пренебречь.) Обозначим расход крови в направлении распростра-
нения волны через Q с соответствующим индексом. Согласно за-
кону сохранения массы, в месте ветвления
Спад QoTp == Qnp» 1 + Qnp> 2- (12.14)
Qotp взято со знаком минус, так как отраженная волна распро-
страняется в данном случае справа налево (рис. 12.23). Напомним,
что средняя по сечению скорость кровотока й' в направлении рас-
пространения волны изменяется- во времени так же, как и избы-
точное давление, и что амплитуда ее изменений равна амплитуде
пульсовых колебаний давления, деленной на рс. Следовательно,
расход Q, равный Ай, где А — площадь поперечного сечения арте-
рии в состоянии равновесия, дается соотношением
0=-^Э- (12.15)
Величины А, р и с характеризуют свойства крови и той арте-
рии, по которой распространяется волна, а не свойства водны кай
338 Часть II. Механика кровообращения
(12.18)
(12.19)
таковой. То же самое относится, следовательно, и к величине рс/Л,
которую обычно обозначают символом Z:
Z = pc/A. (12.16)
Величину Z называют характеристическим импедансом трубки и
определяют как отношение пульсирующего давления к расходу
жидкости в трубке, когда в ней распространяется синусоидальная
волна некоторой частоты; эта величина одинакова для всех точек
трубки1)- Размерность Z есть [M-L-4?-1], т. е. импеданс изме-
ряется в кг-м~4-с_|. Уравнение сохранения массы (12.14) можно
переписать, введя пульсирующие давления и импедансы трех тру-
бок:
Рпад Ротр Рпр-1 Рпр» 2
----Тй----+ —• (12.17)
Уравнения (12.13) и (12.17) должны выполняться для места
ветвления в любой момент времени. Они образуют систему трех
уравнений [(12.13)—это фактически два уравнения], решив кото-
рую, можно найти три неизвестные величины, ротр, рпр, i, рпР, 2, по
известному для места ветвления значению давления в падающей
волне рпад. Решение системы имеет вид
Ротр 1 — (zt 1 + Z2 ______
Рпад “ Z0-,+(Zr, + Z2-1)~
Рпр___________2Z0 1_______у,
Рпад 1 -)- (Z। 1 -J- Z2
(Индекс при рПр опущен, так как в месте ветвления давление в
обеих отходящих от ствола ветвях одинаково: рпр, i = рпр, 2-) Вели-
чину, обратную импедансу, Z-1, обычно называют адмиттансом
(комплексной, или полной, проводимостью).
Уравнения (12.18) и (12.19) весьма содержательны. Обратимся
сначала к частному случаю, когда падающая волна имеет синусои-
дальную форму:
Рпад = Ро cos {® (/ — х/с0)}. (12.20)
Поместим начало координат для каждой трубки в точку ветвления
и примем, что х отрицательно для ствола и положительно для от-
ходящих от него ветвей (см. рис. 12.23). На этом этапе анализа
') Термин «импеданс» заимствован из теории электрических цепей и в на-
стоящее время широко используется в физиологии сердечно-сосудистой системы.
Импеданс подобен сопротивлению в том отношении, что он представляет собой
меру противодействия, которое система сосудов оказывает движению крови через
нее в ответ на определенное колебание разности давления Существует несколь-
ко различных характеристик типа импеданса; этот вопрос обсуждается более
подробно, с привлечением математического аппарата, в конце главы.
Гл. 12. Артерии большого круга 339
конкретная форма волны еще неважна; выражение (12.20) для ее
представления выбрано из соображений простоты и удобства.
В точке ветвления (х — 0) падающая волна давления имеет,
согласно уравнению (12.20), вид
Рпад = Ро COS
Из уравнений (12.18) и (12.19) следует, что ротр и рпр в точке
ветвления также пропорциональны cos и/. Кроме того, нам из-
вестно, что отраженная и прошедшая волны распространяются так
же, как и падающая, поэтому зависимость от х и t для них имеет
вид
Ротр = RpO C0S {“ + х/Со)},
(12.21)
Рпр 1, 2 = Тро COS {и (t — х/ci, 2)}.
Знаки при х/со в первом из этих уравнений и в уравнении (12.20)
различаются, так как отраженная волна распространяется в отри-
цательном направлении по оси х. Второе уравнение в равной сте-
пени применимо к обеим отходящим ветвям; при этом величины,
относящиеся к первой ветви, отмечены индексом 1, а ко второй —
индексом 2.
Эти соотношения дают нам полную информацию об отраженной
и прошедшей волнах: 1) их форма такая же, как у падающей
волны (в рассмотренном случае — синусоидальная); 2) амплитуда
отраженной волны отличается в R раз, а прошедших в каждую
ветвь — в Т раз от амплитуды падающей волны; 3) скорости рас-
пространения отраженной и прошедших волн такие же, как при
распространении обычных синусоидальных волн в этих трубках
(со, Ci и с2 соответственно); 4) переменная составляющая давления
в стволе равна сумме аналогичных составляющих падающей и
отраженной волн:
Р Рпад “Ь Ротр-
Теорию отражения волн от места ветвления общего (т. е. про-
ксимального по отношению к сердцу) сосуда на два отводящих
(дистальных) сосуда можно легко обобщить на случаи отхождения
более чем двух дистальных ветвей. Подобная ситуация имеет
место, например, при отхождении от аорты двух почечных артерий
или при ветвлении аорты собаки на пару подвздошных и крестцо-
вую артерии (область трифуркации аорты). Уравнения, описываю-
щие отраженную и прошедшую волны, аналогичны уравнениям
(12.18) и (12.19) с тем лишь отличием, что вместо суммы адмиттан-
сов двух отходящих ветвей в них входит сумма адмиттансов трех
или более дистальных ветвей, т. е. вместо Zf1 + Z^1 подставляется
Z11 Z21 + Z3 1 ... .
Отношение амплитуд прошедшей и падающей волн [Г в уравне-
нии (12.19)] всегда положительно. Это означает, что в точке вет-
340 Часть II. Механика кровообращения
вления прошедшие волны давления всегда находятся в фазе с па-
дающей волной, каковы бы ни оказались амплитуды и скорости
(ci или с2) прошедших волн. Отношение амплитуд отраженной и
падающей волн R может быть, однако, как положительным, так и
отрицательным. Если это отношение отрицательно, то отраженная
волна достигает максимума при минимуме падающей, и наоборот,
т. е. эти волны сдвинуты по фазе на 180°. Поэтому знак R имеет
важное значение и этот вопрос заслуживает дополнительного рас-
смотрения.
Согласование импедаисов. Когда сумма адмиттансов (обратных
импедансов) ветвей равна адмиттансу общего ствола, коэффициент
R [см. (12.18)] равен нулю и отраженная волна отсутствует. В та-
ком случае говорят о ветвлении с согласованными импедансами.
Используя определение импеданса и связь скорости волны с рас-
тяжимостью [уравнение (12.9)], мы можем исследовать условия,
при которых импедансы согласованы. Импеданс равен
<|222>
Выражая растяжимость через модуль Юнга и отношение толщины
стенки к диаметру [уравнение (7.36)], получим
Z=^(^-)/2- (1223)
Таким образом, если у отходящих ветвей и общего ствола модули
Юнга и отношения толщины стенки к диаметру одинаковы (т. е.
сосуды обладают одинаковой растяжимостью), то импедансы будут
согласованы тогда, когда сумма площадей поперечного сечения
ветвей равна площади сечения ствола. Однако, как видно из табл. I,
при удалении от сердца Е возрастает, а отношение h/d остается
более или менее постоянным. Значит, ветви обычно менее растя-
жимы, чем ствол, и для полного согласования импедансов необхо-
димо, чтобы суммарная площадь поперечного сечения отходящих
сосудов несколько превышала таковую для общего ствола. Далее
мы ознакомимся с некоторыми данными, указывающими на нали-
чие достаточно хорошего согласования импедансов многих ветвле-
ний в сосудистой системе.
Едва ли удастся найти хоть одно ветвление артерий, которое
было бы идеально согласованным, однако можно показать, что при
неполном согласовании, когда величина R очень мала, хотя все
же измерима, энергия, переносимая отраженной волной, пренебре-
жимо мала, так как она пропорциональна квадрату амплитуды.
Энергию, переносимую синусоидальной волной в единицу времени
(IT), легко определить, так как она почти полностью соответствует
работе сил давления по перемещению крови. Эта энергия для лю-
бого момента времени оказывается равной произведению пульси-
Гл. 12. Артерии большого круга 341
Рис. 12.24. Сравнение функций cos at и cos2 at Видно, что среднее значение
cos2 at равно ’/а, тогда как cos at — нулю.
рующего давления р на пульсирующий расход крови Q, а так как
Q — p/Z, то
№ = pQ = p7Z. (12.24)
Хотя р — периодически меняющееся давление и его среднее зна-
чение равно нулю [см. уравнение (12.20)], среднее значение р2 нулю
не равно. Например, среднее за период значение функции
cos2{®(/ — х/£о)} равно '/г (см. рис. 12.24). Таким образом, сред-
няя энергия W, переносимая волной давления через любое сечение
сосуда, равна
W = ±p2Q!Z,
где ро — амплитуда колебаний давления [см. уравнение (12.20)].
Отсюда находим, что отношение средней энергии, переносимой
в единицу времени отраженной волной, к соответствующей энер-
гии для падающей волны равно
Величина R2 называется коэффициентом отражения (разд. 8.4).
Отношение энергии, переносимой обеими прошедшими волнами,
к соответствующей энергии для падающей волны равно
г~1 '
Z0
Эта величина называется коэффициентом пропускания. Из опреде-
лений R и Т [формулы (12.18) и (12.19)] следует, что сумма ко-
342 Часть II. Механика кровообращения
эффициентов отражения и пропускания равна единице, т. е. энер-
гия на ветвлениях такого типа не рассеивается.
Предположим, что ветвление несколько рассогласовано, так что
отличается от Zo', скажем, на 5%. В этом случае, как
показывают вычисления, отраженная энергия составляет всего
0,06% падающей, т. е. подавляющая часть энергии проходит. Это
означает, что рассогласование может быть весьма значительным и
тем не менее еще не оказывать существенного влияния на пере-
дачу энергии.
Сделанные выводы справедливы для «ветвлений» двух типов,
встречающимся в сосудистой системе: 1) ветвлений, при которых
достаточно мелкие артерии отходят от крупной (причем диаметр
и растяжимость последней при этом не меняются, например от-
хождение межреберных артерий от аорты); 2) участков артерий,
в которых их растяжимость или площадь сечения немного ме-
няются; постепенное конусообразное сужение большинства круп-
ных артерий можно рассматривать как последовательность таких
участков (см. далее в этом разделе).
Положительное и отрицательное отражение. При неполном
согласовании R может быть как положительным, так и отрица-
тельным. В предельном случае, когда сумма адмиттансов отходя-
щих ветвей много меньше адмиттанса порождающей ветви (что
должно быть, например, если общая площадь поперечного сечения
отходящих сосудов много меньше сечения общего ствола, а растя-
жимости всех этих сосудов почти одинаковы), R близко к единице.
Отражение при этом должно быть подобно отражению от закры-
того конца, поток через который невозможен. В последнем случае
отраженная волна потока сдвинута по фазе относительно падаю-
щей волны на 180° (волны находятся в противофазе), а поскольку
фаза отраженной волны давления не меняется, амплитуда колеба-
ний давления удваивается, т. е.
Оотр Опад | п <
„ _ J и, следовательно, R = 1.
Ротр г Рпад )
Если бы наблюдатель измерял давление в данной точке беско-
нечно длинной упругой трубки, в которой распространяется сину-
соидальная волна, описываемая уравнением (12.20), то обнаружил
бы синусоидальное изменение давления с амплитудой ро- Если бы
трубку перекрыли заглушкой, то по-прежнему наблюдались бы
синусоидальные колебания давления, однако их амплитуда зави-
села бы от расстояния между точкой измерений и закрытым кон-
цом. У самого конца трубки амплитуда оказалась бы равной 2рс
и уменьшалась бы по мере удаления точки наблюдения от закры-
того конца, пока не обратилась бы в нуль на расстоянии четверти
длины волны от конца трубки (см. рис. 8.11; эта точка называется
узлом). Далее амплитуда опять начала бы увеличиваться, до-
Гл. 12. Артерии большого круга 343
Рис. 12 25 Изменение амплитуды волны с расстоянием от конца закрытого типа,
на котором происходит частичное отражение (амплитудный коэффициент отра-
жения 0,8) Штриховая линия — амплитуда волны в отсутствие отражения.
стигнув максимума, равного 2р0, на расстоянии половины волны от
конца (в пучности), и т. д. Если бы вместо закрытого конца по-
явилось ветвление или другая неоднородность, коэффициент R на
которой положителен, но не равен единице (составляет, скажем,
0,8), то амплитуда колебаний давления ни в одной точке не умень-
шилась бы до нуля, а равнялась бы 0,2 р0 в узлах и лишь 1,8 ро
в пучностях (см. рис. 12.25). Это был бы случай частичного отра-
жения, но того же типа, как отражение от закрытого конца.
Важно отметить, что, когда R близко к единице, Т, как следует
из уравнения (12.19), близко к двум. Следовательно, амплитуда
волны давления, прошедшей через ветвление, эквивалентное почти
полностью закрытому концу, вдвое больше, чем амплитуда падаю-
щей волны. Этот удивительный на первый взгляд вывод объяс-
няется тем, что, как мы знаем [уравнение (12.15)], амплитуда волны
расхода равна амплитуде волны давления, умноженной на Z~l. По-
этому, даже когда амплитуда давления удваивается, амплитуда
прошедшей волны расхода все равно остается очень малой. Удвое-
ние амплитуды колебаний давления в ответвляющейся трубке есть
просто следствие удвоения амплитуды давления вблизи почти за-
крытого конца основной трубки.
Другой предельный случай отражения волн имеет место тогда,
когда сумма адмиттансов отходящих ветвей много больше адмит-
танса общего ствола, например при очень большой суммарной
площади поперечного сечения отходящих ветвей. В этом случае R
близко к —1, а Т очень мало. Эта ситуация аналогична картине,
наблюдаемой при отражении волн от свободного, полностью от»
344 Часть II. Механика кровообращения
крытого конца трубки. Поскольку давление на этом ее конце
должно быть таким же, как в камере, в которую открывается
трубка, колебания давления на открытом конце должны быть
равны нулю. При этом амплитуда колебаний расхода на открытом
в камеру конце трубки удваивается. На этом конце и через каждые
полдлины волны от него находятся узлы давления; пучности рас-
полагаются на расстоянии в четверть длины волны от конца и да-
лее через каждые полдлины волны. При отражении от открытого
конца фаза волны расхода остается неизменной, тогда как фаза
волны давления меняется на противоположную. Во всех случаях
отрицательного отражения (т. е. когда R < 0) изменение фазы
волн давления и расхода такое же, как при отражении от конца
открытого типа.
Итак, при положительном отражении фаза волны давления не
меняется, фаза волны расхода изменяется на 180°, а амплитуда
прошедшей волны давления становится больше амплитуды падаю-
щей волны. В случае отрицательного отражения фаза волны рас-
хода остается прежней, фаза волны давления изменяется на 180°,
а амплитуда прошедшей волны давления становится меньше, чем
у падающей волны.
Физиологические данные, свидетельствующие об отражении
волн. Изучению пульсовых волн давления и кровотока в различ-
ных участках системы крупных артерий человека посвящен ряд
работ. На рис. 12.26 приведены записи этих волн в восходящей
аорте, в грудной и брюшной аорте, а также в общей подвздошной,
подключичной, плечеголовной и почечной артериях человека. Опи-
раясь на все сказанное выше, попытаемся предсказать некоторые
возникающие в результате отражений особенности пульсовых
волн ).
Прежде всего определим, через какое время от начала систолы
а) фронт пульсовой волны достигнет некоторого участка системы
крупных артерий и б) отраженные волны вернутся в тот же уча-
сток. Место наблюдений в восходящей аорте можно рассматривать
как расположенное на выходе левого желудочка. Зная скорость
волны — около 5 м-с-1 (из табл. I видно, что различие скорости
волны у собаки и человека незначительно), можно оценить время,
за которое пульсовая волна достигнет места отхождения ветвей
дуги аорты, отстоящего от сердца примерно на 8 см. Это время
составляет около 0,016 с, и, следовательно, уже через 0,032 с от-
раженная от этих ветвей волна вернется к корню аорты. Но на-
чальная часть систолы, когда кровь ускоряется, а давление и
расход быстро растут, длится обычно 0,1 с. Таким образом,
*) Картина отраженных волн, наблюдаемая в данном месте системы круп-
ных артерий, у животных разных видов по-видимому, неодинакова. Обсуждае-
мые здесь явления относятся к человеку. Соответствующие данные, касающиеся
собаки, приведены в гл 2 книги McDonald D А, Blood flow in arteries, Edward
Arnold, 1§74.
Гл. 12. Артерии большого круга 345
Правая
подключичная
артерия
Плечеголовная
артерия
Давление,
мм рт. ст
Давление
мм рт. ст
Рис. 12.26. Зарегистрированные одновременно кривые пульсовых волн давления
и скорости крови в различных участках артериального дерева человека. (Все
кривые зарегистрированы у одного пациента, за исключением кривых дли правой
почечной и правой общей подвздошной артерий.) [Mills et al. (1970). Cardiovacs.
Res., 4, 405.]
отраженная волна едва ли окажется заметной: ее фронт достигает
корня аорты в период быстрого повышения давления, а ее ампли-
туда будет, вероятно, невелика, поскольку общая площадь по-
перечного сечения всех артерий после отхождения ветвей от дуги
аорты немногим больше площади сечения аорты до их отхожде-
ния.
Другое, более важное место отражения распространяющейся
по аорте пульсовой волны, характеризующееся наибольшим коэф-
фициентом отражения, — это бифуркация аорты. Измерения пока-
зали, что общая площадь поперечного сечения обеих подвздошных
артерий обычно на 20% меньше площади сечения брюшной аорты,
поэтому на бифуркации аорты следует ожидать значительного по-
ложительного отражения. (Это предположение может не выпол-
няться для собак, у которых последнее разветвление аорты, ее три-
346 Часть tl. Механика кровообращения
фуркация, может быть хорошо согласовано.) Далее, время, в те-
чение которого волна распространяется от сердца до бифуркации
аорты, равно отношению расстояния (у человека — около 60 см)
к средней скорости волны в аорте (равной примерно 6 м-с-1)- Это
время составляет приблизительно 0,1 с, и, следовательно, отра-
женная волна вернется к корню аорты (равно как и к местам из-
мерения в подключичной и плечеголовной артериях) спустя 0,2 с.
Отметим, что на нисходящем фронте волн скорости крови
имеется ступенька (рис. 12.26; измерения производили в ядре по-
тока, и поскольку профиль скорости в артериях почти плоский, за-
писи отображают также и расход крови, см. разд. 12.8). Эта сту-
пенька представляет собой результат кратковременного повышения
и последующего замедления спада скорости кровотока. Ступенька
появляется спустя некоторое время от начала систолы, причем это
время тем больше, чем дальше от бифуркации аорты вверх по те-
чению крови расположено место измерения, и для плечеголовной
артерии это время достигает примерно 0,2 с. Поэтому наличие сту-
пеньки на записи волны скорости крови можно расценить как при-
знак прихода отраженной волны. Но, поскольку предполагается,
что область разветвления аорты на пару подвздошных артерий
является местом положительного отражения, т. е. отражения по
типу закрытого конца, отраженная и падающая волны скорости
крови не совпадают по фазе и максимуму падающей волны соот-
ветствует минимум (впадина) отраженной волны. Легко видеть,
что такая впадина должна вызвать ускоренный спад волны скоро-
сти кровотока в аорте, что и наблюдается в действительности.
Флуктуации на записях давления (рис. 12.26) столь многочис-
ленны, что расценить какую-либо из них как следствие отражения
волн от бифуркации аорты весьма трудно. Зубец, наблюдаемый
в конце систолы, отмечает момент закрытия аортального клапана
и перемещается вместе с основной волной давления, хотя с удале-
нием от сердца его амплитуда быстро падает (разд. 12.6). Отме-
тим, что амплитуда волны давления по мере удаления от сердца
возрастает (на протяжении аорты она увеличивается примерно на
60%; рис. 12.19), тогда как амплитуда волны скорости кровотока
уменьшается. Такое явление находится в соответствии с представ-
лением об отражении волн на бифуркации аорты как на конце за-
крытого типа, хотя частично оно может определяться плавным
конусообразным сужением аорты и увеличением ее жесткости,
о чем пойдет речь в следующем разделе.
Другое явление, наблюдающееся в системах, где распростра-
няются волны, — резонанс (разд. 8.4). На расстоянии четверти
длины волны от места отражения типа закрытого конца располо-
жена точка (узел давления), в которой амплитуда колебаний дав-
ления минимальна (рис. 12.25), а амплитуда колебаний скорости
кровотока максимальна. Узлы давления располагаются далее че-
рез каждую половину длины волны. Между ними находятся пуч-
Гл. 12. Артерии большого круга 347
ности, в которых амплитуда достигает наибольшего значения. Если
положение какой-либо пучности или узла, обязанных своим проис-
хождением отражению от конца закрытого типа, совпадает с дру-
гим местом отражения, например с противоположным концом труб-
ки конечной длины, то в месте первого отражения возникает пуч-
ность, обусловленная наличием этого второго отражающего участ-
ка. И без того большая амплитуда давления (связанная с первым
положительным отражением) еще более увеличивается и так про-
исходит при каждом следующем отражении. Такое явление, назы-
ваемое резонансом, должно наблюдаться, если длина системы рав-
на целому числу полуволн. Длина пульсовой волны давления в
аорте много больше длины сосуда, и поэтому не следует ожидать
Падающая
волна --------
Отраженная •*—
волна --------
Прошедшая
волна
Прошедшая
волна
Рис. 12.27. Ветвление, при котором от общего ствола отходит небольшая ветвь,
а свойства ствола при этом не меняются (пример — отхождение межреберной
артерии от аорты).
резонанса для всей волны. Однако у человека четвертая гармо-
ника пульсовой волны давления имеет длину волны, примерно
вдвое большую длины аорты, а у собак такую длину волны имеет
шестая гармоника. Следовательно, используя анализ Фурье, нуж-
но разложить пульсовую волну на ее синусоидальные составляю-
щие и посмотреть, не окажется ли амплитуда четвертой гармоники
(для собак — шестой) непропорционально большой по сравнению
с амплитудами других гармоник. Однако даже для этих гармоник
никаких экспериментальных данных о наличии резонанса не полу-
чено (рис. 12.20). По-видимому, указанные гармоники быстро за-
тухают и после нескольких отражений становятся несуществен-
ными.
До сих пор мы рассматривали распространение волн в основ-
ных артериальных стволах; обратимся теперь к явлениям, которые
наблюдаются в местах, где от крупных артерий отходят более
мелкие боковые ветви (рис. 12.27), например в местах ответвления
межреберных или коронарных артерий от аорты. Здесь адмиттанс
одной из отходящих ветвей остается примерно таким же, как у об-
щего ствола, тогда как адмиттанс другой (или других) ветвей
очень мал. Поэтому сумма адмиттансов ветвей лишь немного
больше адмиттанса ствола и ветвление достаточно хорошо согласо-
вано. Это означает, что энергия, переносимая отраженной волной,
318 Часть II. Механика кровообращения
пренебрежимо мала (см. ниже), так что форма основной волны
давления в аорте практически не должна меняться, тогда как ам-
плитуда пульсового давления в небольшой ветви оказывается та-
кой же, как в аорте. Это и наблюдается в действительности.
Множественные отражения. Для установления соотношения
между волнами давления и расхода до и после отдельного ветвле-
ния артерий нет необходимости привлекать к рассмотрению всю
артериальную систему. Однако, чтобы детально проанализировать
особенности формы пульсовых волн давления и кровотока, зареги-
стрированных в некотором месте этой системы, например в вос-
Рис. 1228 Схема двух последовательных ветвлений. Волна, падающая из трубки
О, в точке А частично отражается, а частично проходит в трубки 1 и 2. Прошед-
шая в трубку 1 волна в точке В частично отражается, а частично проходит даль-
ше. Отраженная в точке В волна вновь частично отражается в точке А и т. д.
ходящей аорте, нельзя не учитывать наличие в артериальной си-
стеме многочисленных мест отражения, расположенных на самых
разных расстояниях от места измерения. Следовательно, необхо-
димо изучить, как влияют на пульсовую волну повторные отра-
жения.
Рассмотрим сначала два последовательных ветвления, изобра-
женных на рис. 12.28. Волна, однажды отразившаяся на бифурка-
ции В, повторно отражается на бифуркации А, и т. д. Амплитуды
отраженной и прошедшей волн всякий раз определяются характе-
ристиками ветвления. Если ветвление В достаточно хорошо согла-
совано, то отраженная на нем волна будет иметь небольшую ам-
плитуду. Когда эта волна вновь отразится на ветвлении А, ампли-
туда дважды отраженной волны станет еще меньше, хотя и нена-
много, так как, если ветвление А хорошо согласовано для волны,
которая приходит из общего сосуда, оно не будет столь же хорошо
согласовано для волны, возвращающейся к нему из ветви. Однако
в любом случае амплитуда такой многократно отраженной от кон-
цов участка АВ («запертой» в нем) волны будет быстро умень-
Г л. 12. Артерии большого круга 349
шаться п станет весьма малой уже после небольшого числа отра-
жений даже в отсутствие затухания ’).
Единственным исключением из этого правила является случай,
когда длина сосуда АВ равна целому числу полуволн — в такой
системе должен возникать резонанс. При резонансе прохождение
волны через сосуд АВ не требует притока энергии и волны, прошед-
шие ветвление В, будут иметь такие характеристики, как если бы
сосуда АВ не было, т. е., пройдя через сосуд АВ, волны в случае
резонанса не претерпевают никаких изменений.
С другой стороны, если длина сосуда АВ в нечетное число раз
больше четверти длины волны, отраженная волна будет всякий раз
находиться в противофазе с падающей, когда отражение на вет-
влении В положительно, и в фазе, когда отражение отрицательно.
Амплитуда прошедшей волны будет уменьшаться в случае поло-
жительного отражения и увеличиваться в случае отрицательного.
Вообще говоря, на практике эти предельные случаи не встре-
чаются, однако особенности отраженной и прошедшей волн все же
зависят от отношения длины I промежуточного сосуда к длине рас-
пространяющейся волны. Именно это отношение определяет вре-
менную задержку, а следовательно, и разность фаз между волной,
прошедшей ветвление А, и волной, вернувшейся к нему после от-
ражения на ветвлении В. Значит, поведение волны при прохожде-
нии двух или более последовательных ветвлений зависит от ее ча-
стоты, а потому поведение волны, имеющей сложную форму, ко-
торую можно представить в виде суммы синусоидальных состав-
ляющих с разными частотами (рис. 8.14), будет, по-видимому,
весьма сложным и форма волны, прошедшей через пару последо-
вательных ветвлений, будет отличаться от формы падающей волны.
Не претерпят изменения только те ее составляющие, длина кото-
рых много больше I, а также составляющие, для которых I при-
близительно равно целому числу полуволн. По отношению к та-
ким составляющим волн два ветвления и сосуд между ними можно
рассматривать как единый отражающий участок. Что касается
системы кровообращения, то длина самой крупной артерии —
аорты (60 см у человека и 40 см у собаки) много меньше, чем
длина пульсовой волны (Б м у человека и 3 м у собаки), так что
последовательные отражения способны влиять только на высокие
гармоники. Поэтому значительные изменения будут претерпевать
именно эти, высокочастотные составляющие пульсовой волны.
Выше в этом разделе мы рассмотрели, как определить свойства
прошедшей и отраженной на одиночном ветвлении волн. Исполь-
’) До сих пор термин «затухание» использовался практически как синоним
уменьшения амплитуды. В действительности уменьшение амплитуды (т. е. ослаб-
ление) волны может быть вызвано как диссипацией энергии в системе, например
вследствие вязкости, так и различными недиссипативными механизмами Затуха-
нием называют обычно именно те виды ослабления, которые обусловлены дис-
сипацией энергии. — Прим. ped.
350 Часть II. Механика кровообращения
зуя те же условия (12.13), (12.14)—отсутствие скачкообразных
изменений давления и полного расхода крови при переходе каж-
дого ветвления, можно найти параметры синусоидальных волн дан-
ной частоты и в случае множественных отражений на последова-
тельных ветвлениях. Зная, например, какие волны распростра-
няются в сосудах 3 и 4 (рис. 12.28), можно определить, что проис-
ходит в сосуде 1. Установив таким же способом характеристики
волн в сосуде 2, объединив результаты для сосудов 1 и 2 и оценив
форму колебаний давления и расхода крови на ветвлении А, можно
найти характеристики волн в сосуде 0. Более того, если сосуд 0
есть результат предшествовавшего ветвления, следует повторить
эту процедуру и получить характеристики волны выше этого вет-
вления и т. д. Такой способ позволяет вычислить параметры пуль-
совых волн давления и расхода в любом участке сложной сети
ветвящихся сосудов, если известны соотношения между давлением
и расходом в самых мелких сосудах системы, а также длины и ха-
рактеристические импедансы всех образующих ее сосудов. Чтобы
проделать подобный расчет для артериальной системы, можно
было бы, например, предположить, что мгновенный расход крови
в микрососудах пропорционален мгновенному перепаду давления в
них (т. е. что периферическое сопротивление постоянно). Однако
в настоящее время провести такие расчеты невозможно, ибо необ-
ходимые для этого данные об анатомической организации и свой-
ствах сосудов, соединяющих крупные артериальные стволы с мик-
рососудами, отсутствуют. Соответствующие расчеты были выпол-
нены лишь для модели артериальной системы, которая состояла
из нескольких поколений ветвящихся трубок. При этом считалось,
что диаметр и вязкоупругие свойства сосудов одного поколения
одинаковы, а длины их распределены случайным образом. Пред-
полагалось также, что скорость волны и общая площадь попереч-
ного сечения ветвей по направлению к периферии возрастают.
К результатам, полученным для этой модели, мы обратимся ниже.
Указанные расчеты позволяют предсказать характер связи ме-
жду давлением и расходом крови в любой точке системы сосудов,
даже если неизвестно, какие именно волны распространяются в ее
конечных разветвлениях. Так, зная форму волны давления в неко-
торой точке, можно определить форму волны расхода, и наоборот.
Кроме того, регистрация в каком-либо месте сосудистой системы
пульсовых волн давления и расхода крови позволяет сделать за-
ключения о некоторых свойствах этой системы. Связь между дав-
лением и расходом в случае синусоидальной волны, имеющей не-
которую определенную частоту, характеризуют двумя постоян-
ными: одна из них — отношение амплитуд колебаний давления и
расхода, обозначаемое обычно символом М, а другая — отставание
волн расхода от волн давления, т. е. сдвиг по фазе 0. Если на
входе в систему синусоидальная составляющая давления с угловой
частотой © имеет вид р cos ©/, то соответствующая составляющая
Гл. 12. Артерии большого круга 351
расхода будет (p/Af)cos((o/— 0). Чтобы полностью охарактеризо-
вать систему, необходимо построить графики зависимости М и 0
от ю; такие графики позволяют описать поведение в системе волн
любой формы, которые представлены в виде суммы синусоидаль-
ных составляющих. Зарегистрировав пульсовые волны давления и
расхода крови, например, в восходящей аорте, те и другие следует
подвергнуть анализу Фурье и сравнить амплитуды и фазы каждой
из гармоник волны расхода крови с соответствующими параме-
трами гармоник волны давления. Это позволит определить значе-
ния М и 0 для каждой из частот, представленных в волнах, т. е.
присутствующих в разложении в ряд Фурье. Результаты многих
экспериментальных исследований отображены именно в таком
виде.
Величина М, получаемая делением амплитуд гармоник давления
и расхода крови, имеет размерность импеданса. Однако, поскольку
М зависит от расположения места измерения, равно как и от ча-
стоты, — это не то же самое, что характеристический импеданс
сосуда, в котором проводят измерения (например, аорты). Вели-
чины М и 0 в совокупности представляют собой эффективный (или
входной) импеданс всей системы в точке измерения на частоте со.
Обычно М называют модулем эффективного (или входного) импе-
данса артериальной системы, а 0 — его фазой. Эти термины обя-
заны своим происхождением тому, что волновые процессы принято
описывать с помощью комплексных чисел, чего мы до сих пор пы-
тались избегать. В дополнении к данной главе пояснен смысл этих
названий и некоторых других величин, которые иногда также на-
зывают «импедансом». Быстрый рост числа таких величин спосо-
бен привести к изрядной путанице, и мы надеемся, что упомяну-
тое дополнение внесет ясность в этот вопрос и послужит предосте-
режением от того, чтобы использовать термин «импеданс» без та-
ких определяющих его прилагательных, как, например, «характе-
ристический» или «входной».
Анализ формы пульсовых волн давления, наблюдающихся в
крупных артериях. Представление о входном импедансе сер-
дечно-сосудистой системы для волн давления различной частоты
позволяет объяснить особенности формы пульсовых волн в аорте
и других крупных артериях гораздо более полно, чем это удается
сделать при простом просматривании записей волн. В последнем
случае удается выявить лишь такие грубые эффекты, как влияние
на форму волны единственного места, достаточно сильно отражаю-
щего волны. Для получения более детальной информации необхо-
димо разложить зарегистрированные в каком-либо месте волны
давления и расхода на синусоидальные составляющие различной
частоты. Далее следует сопоставить амплитуды и фазы гармоник
волн расхода крови с амплитудами и фазами соответствующих гар-
моник волн давления, выразить результаты через модуль М и фазу
362 Часть И Механика кровообращения
6 входного импеданса и построить графики зависимости этих ве-
личин от частоты.
Анализ таких графиков позволяет сделать некоторые выводы
о свойствах системы. Например, если рассматриваемая система
представляет собой просто однородную трубку с отражением на
конце, а точка измерения отстоит от конца трубки на четверть
длины волны, то, как следует из уравнения (12.32) (см. дополне-
ние), участок трубки, заключенный между ее концом и местом из-
мерения, не оказывает влияния на величину входного импеданса.
В этом случае модуль М входного импеданса минимален. Кроме
того, при повышении частоты волны до величины, при которой /
(расстояние от точки измерения до конца трубки) становится
больше четверти длины волны, фаза 6 входного импеданса меняет
знак: из отрицательной она становится положительной. Такие со-
ображения привели экспериментаторов, изучающих артериальную
систему, к идее построения графиков зависимости М и 0 от <в, из
которых, если М имеет минимум (а 6 обращается в нуль) при
<о==б, следует, что существует место отражения, расположенное
на расстоянии лс/2б от точки измерения, где с — определенная не-
зависимым методом скорость распространения волны давления по
артериям. Для сложных ветвящихся систем подобное истолкова-
ние результатов неизбежно оказывается сугубо предположитель-
ным. Определенное таким способом место отражения (ветвления)
почти наверняка не совпадет в точности с действительным его по-
ложением, поскольку на результаты определения влияют отраже-
ния от всех других ветвлений. В сложных системах отраженная
волна представляет собой сложную результирующую отражений
от многих участков. Однако если в силу анатомических особенно-
стей преобладающее влияние оказывает одно из ветвлений (напри-
мер, бифуркация аорты), а анализ указывает, что место отраже-
ния находится вблизи этого ветвления, то именно его и разумно
считать главным местом отражения волн.
На рис. 12 29 приведены типичные графики зависимости модуля
и фазы входного импеданса от частоты, построенные по резуль-
татам измерений давления и кровотока в восходящей аорте чело-
века. Эти результаты получены из той же серии измерений, в кото-
рой производились записи формы пульсовых волн, представленные
на рис. 12.26. Видно, что минимум М достигается при частоте 3 Гц,
а из расчетов следует, что тогда место отражения должно распо-
лагаться приблизительно на уровне бифуркации аорты. Измерения
в других участках аорты приводят к тому же выводу.
Еще одно общее свойство входного импеданса, выявляющееся
из рис. 12.29, состоит в резком увеличении значений М при очень
низких частотах (при этом 6 близко к нулю). Постоянная, т. е.
имеющая нулевую частоту, составляющая волны расхода представ-
ляет собой средний по времени расход крови, и, следовательно,
эта огромная величина входного импеданса при частоте, равной
Рл. t2. Артерии большого круга 353
£
Рис. 12.29. Зависимости фазы (Л) и модуля (Б) входного импедвиса восходящей
аорты человека от частоты f Наличие единственного минимума модуля импедан-
са позволяет предположить, что ниже по течению имеется лишь одии участок,
где происходит значительное отражение. [Mills et al. (1970). Cardiovasc. Res.,
4, 405—417]
нулю, есть сопротивление, оказываемое среднему потоку крови.
Таким образом, мы устанавливаем, что «противодействие», оказы-
ваемое системой артерий нескольким первым гармоникам пульси-
рующей части потока, много меньше, чем «противодействие» сред-
нему потоку. Но именно эти несколько первых гармоник переносят
большую часть энергии волны. Отсюда следует, что энергия, тре-
бующаяся для поддержания пульсирующего течения крови а аорте,
лишь ненамного (примерно на 10%) превосходит энергию, необ-
ходимую для поддержания стационарного (непульсирующего) по-
тока с тем же средним расходом.
354 Часть tl Механика кровообращения
Рис. 1230. Зависимости модуля (Д) и фазы (В) входного импеданса восходя-
щей аорты собаки от частоты f Наличие двух минимумов модуля импеданса
позволяет предположить, что имеются два участка, где происходит значительное
отражение, находящиеся на разном удалении от сердца [O’Rourke, Taylor
(1967). Input impedance of the systemic circulation Circulation Res., 20, 365—
380.]
Гл. 12. Артерии большого круга 355
Результаты подобного анализа входного импеданса для восхо-
дящей аорты собаки представлены на рис. 12.30. Они отличаются
от предыдущих тем, что модуль импеданса имеет два минимума.
Этот факт был объяснен наличием двух отражающих участков, на-
ходящихся на разном удалении от сердца. А поскольку при ана-
лизе волн, зарегистрированных в нисходящей аорте собаки, второй
минимум обнаружен не был, заключили, что одно из основных мест
отражения находится в передней половине тела, т. е. в той части
Рис. 12.31. Модель большого круга кровообращения
в виде асимметричного Т-образного тройника, пред-
ложенная О’Рурком для объяснения результатов,
приведенных на рис. 12 30. Ближайший участок от-
ражения связывают с артериальными сосудами голо-
вы и передних конечностей, а более удаленный — с
сосудами органов полости таза и задних конечностей.
[O’Rourke (1967). Pressure and flow-waves in sys-
temic arteries and the anatomical design of the ar-
terial system. J. appl. Physiol, 23, 148.J
Сердце
Г олова
и передние
коначности
Задние конечности
и органы полости
газа
кровеносной системы, которая снабжает кровью голову и передние
конечности. Это привело к построению модели большого круга
кровообращения в виде несимметричного Т-образного «тройника»
с двумя основными местами отражения волн (рис. 12.31). Резуль-
таты измерений пульсирующего давления и расхода в восходящей
аорте собаки могут, конечно, истолковываться в рамках этой мо-
дели, однако пользоваться ею следует с осторожностью, так как
она не отражает всей сложности системы кровообращения. Отсут-
ствие второго минимума на графике зависимости модуля входного
импеданса от частоты для человека может быть следствием ряда
факторов: а) незначительности волны, отраженной от крупных ар-
терий верхней части тела, вследствие того, что в них направляется
относительно меньший поток крови; б) лучшего согласования вет-
влений артерий в этой части тела; в) различия анатомической ор-
ганизации ветвей аорты человека и собаки (у человека от дуги
аорты отходят три ветви, у собаки — две); г) того обстоятельства,
что входной импеданс определяли у больных, страдающих забо-
леваниями сердца. Не исключено, что у здоровых людей мог бы
быть выявлен второй минимум модуля импеданса.
Результаты анализа входного импеданса бедренной артерии
собаки позволили заключить, что место существенного отражения
волн находится чуть ниже коленного сустава. Кроме того, эти ре-
зультаты показали, что для бедренной артерии отношение модулей
импеданса нескольких первых гармоник к сопротивлению сред-
356 Часть П. Механика кровообращения
нему потоку крови больше, чем для аорты. Такое различие есть
следствие меныпей растяжимости более периферических артерий.
Единственный теоретический расчет входного импеданса, ре-
зультат которого можно сопоставить с экспериментальными дан-
ными (рис. 12.29 и 12.30), основан на использовании упомянутой
выше модели, в которой артериальная система представлена в
виде сети случайным образом ветвящихся трубок. Результаты рас-
четов приведены на рис. 12.32. Видно, что в области низких частот
модель воспроизводит основные свойства входного импеданса, вы-
явленные экспериментально, но детального соответствия между
кривыми нет.
Влияние конусообразного сужения артерий. Чтобы понять,
как изменяются параметры волны, распространяющейся по трубке,
площадь сечения и растяжимость которой плавно меняются по
длине, можно воспользоваться уже рассмотренной выше относи-
тельно простой теорией отражения на отдельном отражающем
участке. Полученный результат следует далее применить к вол-
нам, распространяющимся по артериям, которые, подобно аорте,
по мере удаления от начала конусообразно суживаются и стано-
вятся более жесткими. Влияние конусообразного сужения аорты
наряду с эффектом отражения волны от области ее разделения на
подвздошные артерии позволяет объяснить, почему возрастает ам-
плитуда пульсовой волны давления (рис. 12.19), хотя реальное ее
увеличение оказывается значительно меньше предсказываемого
этой простой теорией.
Чтобы использовать эту теорию, представим постепенно сужаю-
щуюся артерию в виде ряда последовательных отражающих участ-
ков, распределенных по ее длине (рис. 12.33). Все составляющие
волны будут многократно отражаться на каждом из участков. Сов-
местное влияние таких отражений равносильно постепенному не-
прерывному изменению исходной волны, так что по мере ее пере-
мещения вдоль трубки амплитуда, форма и скорость распростра-
нения будут меняться. В случае достаточно плавного изменения
свойств стенки артерии анализ изменений формы пульсовой волны
оказывается весьма несложным, так как каждое из мест отраже-
ния, дающих вклад в постепенное изменение волны, соответствует
очень малому изменению характеристического импеданса дисталь-
ной части сосуда. Следовательно, амплитуда отраженной на каж-
дом участке волны также мала, и тем более мала энергия, пере-
носимая отраженной волной (см. выше). Это означает, что при рас-
пространении волны по сосуду, свойства которого меняются очень
плавно, «отражения» энергии практически не происходит и изме-
нение формы волны можно анализировать так, как если бы вся
энергия переносилась прошедшей волной.
Согласно уравнению (12.24), энергия, переносимая в единицу
времени перемещающейся волной давления, должна быть равна
квадрату амплитуды пульсаций давления р%, деленному на харак-
Гл 12. Артерии большого круга 357
Рис. 12.32. Зависимости модуля и фазы входного импеданса модели арте-
риального дерева большого круга от частоты (ср. с результатами непосредствен-
ных измерений, представленными на рис 12 29 и 12 30) Масштаб для моду-
ля импеданса выбран так, чтобы модуль характеристического импеданса аорты
был равен единице. [Taylor (1966). The input impedance of an assembly of ran-
domly branching elastic tubes. Biophys. J, 6, 29—51.]
Рис. 1233. Постепенное конусообразное сужение трубки, представленное в виде
последовательности малых ступенчатых изменений просвета. На каждой из сту-
пенек происходит пренебрежимо малое отражение.
358 Чисть П. Механика кровообращения
теристический импеданс Z трубки. Если вся энергия переносится
прошедшей волной, это отношение для всех участков сосуда оди-
наково, откуда
р0 = const • Z1/2. (12.25)
Таким образом, в случае постепенного конусообразного сужения
артерий амплитуда волны давления пропорциональна корню квад-
ратному из характеристического импеданса.
Наконец, из (12.22) следует, что импеданс сосуда увеличивается
с уменьшением как площади поперечного сечения А, так и растя-
жимости D. Поскольку и А, и D для аорты в направлении к пери-
ферии уменьшаются, то, как можно предсказать, амплитуда пуль-
совой волны давления должна возрастать. Амплитуда пульсовой
волны расхода крови в аорте, равная po/Z [уравнение (12.15)], про-
порциональна Z-1/2 и потому будет уменьшаться.
Из данных, приведенных в табл. I, следует, что площадь попе-
речного сечения брюшной аорты составляет около 40% площади
грудной. Кроме того, модуль Юнга стенки брюшной аорты при-
мерно вдвое больше, чем грудной. Следовательно, импеданс брюш-
ной аорты почти в 3,5 раза превосходит импеданс грудной аорты
[уравнение (12.23)], и можно ожидать, что амплитуда пульсовой
волны давления возрастет примерно на 85%. По мере приближе-
ния этой волны к области отражения на бифуркации аорты проис-
ходит дальнейший рост ее амплитуды, и в итоге, как следует из
теории, в конце аорты амплитуда пульсовой волны давления долж-
на по меньшей мере вдвое превосходить ее амплитуду в начале
аорты. Действительный рост не столь велик, и это можно припи-
сать двум обстоятельствам: 1) приближенному характеру теорети-
ческого анализа и 2) затуханию волны вследствие диссипации энер-
гии, обусловленной вязкими свойствами крови и вязкоупругими
свойствами стенки крупных артерий. Последнее мы рассмотрим
в разд. 12.6, а здесь остановимся на первом факторе.
Представленная теория точно предсказывает величину, на ко-
торую возрастает амплитуда волны давления вдоль конусообразно
сужающегося сосуда, пока выполняется условие пренебрежимо ма-
лого переноса энергии отраженной волны. Однако, если конусность
трубки значительна, перенос энергии отраженной волной стано-
вится существенным и предсказанное теорией увеличение ампли-
туды волны давления будет меньше наблюдаемого. Критерием
правомерности использования теории служит выполнение требова-
ния, состоящего в том, что длина конусообразно суживающейся
трубки должна быть в несколько раз больше длины волны. Это
условие, естественно, не выполняется для нескольких первых гар-
моник— составляющих пульсовых волн давления: длина волны
этих гармоник превышает длину аорты. Поэтому неудивительно,
что реально наблюдаемое увеличение амплитуды пульсовых волн
давления меньше предсказываемого теорией.
Гл. 12 Артерии большого круга 359
12.5. Влияние нелинейности
До сих пор мы принимали, что артериальная система линейна,
т. е. что распространение и отражение каждой из синусоидальных
составляющих пульсовой волны давления можно рассматривать
независимо от других составляющих. Это положение опирается на
два допущения: 1) средняя скорость движения крови й при любых
обстоятельствах много меньше скорости волны с и 2) амплитуда
волны давления в артериях мала по сравнению со средним
избыточным давлением в них, так что при прохождении этой волны
растяжимость артерий не меняется. Однако в разд. 12.3 было пока-
зано, что для наиболее крупных артерий — аорты и легочного
ствола — эти допущения не вполне справедливы и в действитель-
ности нелинейные свойства могут, вероятно, в заметной степени
изменять процесс прохождения пульсовых волн по этим сосудам.
Нелинейность проявляется в виде двух эффектов. Один из
них — упругий нелинейный эффект — есть следствие нелинейной
зависимости напряжения в стенке артерии от ее диаметра. Соот-
ветствующая кривая (рис. 7.5) показывает, что с ростом давления,
растягивающего артерию в радиальном направлении, она стано-
вится более жесткой, т. е. растяжимость ее уменьшается. Значит,
скорость волны с, как следует из уравнения (12.9), должна воз-
растать. Результаты опытов как in vitro на вырезанных сегментах
артерий, так и in vivo подтверждают это предположение. В послед-
нем случае у собак пережимали аорту и во время диастолического
спада давления при различных значениях этого давления в ней
создавали короткие пакеты высокочастотных волн давления. По-
скольку длина волны таких колебаний была мала, удавалось заре-
гистрировать несколько их циклов до того, как в место измерения
давления (ниже по течению от места генерации колебаний) воз-
вращалась волна, отраженная от трифуркации аорты, и искажала
записи. Результаты одного из таких опытов представлены на
рис. 12.34. Из них следует, что скорость волны при обычных зна-
чениях максимального давления, достигаемых во время систолы,
существенно отличается от скорости волны при обычном диастоли-
ческом давлении. Таким образом, систолическая часть пульсовой
волны давления перемещается быстрее, чем диастолическая.
Второй нелинейный эффект имеет гидродинамическое присхож-
дение. Он становится существенным в случаях, когда отношение
й/с не мало. Можно доказать, что пульсовая волна давления рас-
пространяется с локальной скоростью с относительно движущейся
крови, а не относительно стенки сосуда. Поэтому если локальная
скорость движения крови равна й, то скорость прямой волны, пере-
мещающейся по течению, есть с + й, тогда как скорость обратной
волны, которая, возвращаясь от какого-либо дистально располо-
женного места отражения, перемещается против течения, равна
с — й. Этот вывод также был подтвержден в ходе наблюдений за
360 Часть П. Механика кровообращения
Рис. 12 34 Зависимость скорости волны в аорте собаки от давления в аорте
Высокочастотные колебания накладывали на пульсовую волну во время диа-
столы. Необходимое диастолическое давление устанавливали, пережимая аорту
в разных участках Темные точки соответствуют волнам, распространяющимся
вниз по течению, светлые — волнам, распространяющимся вверх [Histand, Anliker
(1973) Circulation Res, 32, 524.]
наложенными на пульсовую волну давления небольшими по ам-
плитуде высокочастотными колебаниями. Последние возбуждались
в различные моменты пульсовых циклов, а скорость их распростра-
нения измерялась как по течению крови, так и против него. Влия-
ние нелинейного гидродинамического эффекта на пульсовую волну
заключается в том, что во время систолы, когда скорость движения
крови наибольшая, скорость перемещения пульсовой волны давле-
ния в направлении потока крови больше, чем во время диастолы
Таким образом, это явление также приводит к ускорению пере-
мещения начальной части пульсовой волны относительно ее после-
дующих частей.
Учитывая нелинейные эффекты обоих видов, можно, следова-
тельно, заключить, что в начале систолы направленная вниз по
течению скорость волны больше, чем во время диастолы. Поэтому
по мере перемещения пульсовой волны к периферии вершина волн
и давления, и скорости кровотока стремится догнать подножие этих
волн, в результате чего фронт волны становится более крутым. Та-
кой эффект можно наблюдать, производя одновременно записи
давления в различных участках аорты и в других крупных арте-
риях (рис. 12.14 и 12.19). Полагают, что у здоровых людей это
явление не играет сколько-нибудь заметной роли, однако при та-
ких заболеваниях, при которых растяжимость артерий существенно
повышается (так что с становится мала) или когда амплитуда
Гл. 12. Артерии большого круга 36|
пульсовых волн сильно возрастает (и потому возрастают й и
Др/Ризб), это явление может оказаться весьма существенным. У со-
ответствующих больных увеличение крутизны фронта пульсовой
волны происходит быстрее, ее вершина может нагнать подножие, и
фронт волны станет весьма крутым.
Подобные явления, не охватываемые линейными теориями, хо-
рошо известны для волн различной физической природы. Напри-
мер, когда фронт волны на воде становится вертикальным, волна
разрушается, обыкновенно пенясь и бурля, и это сопровождается
значительной диссипацией энергии. Если звуковая волна достигает
большой интенсивности, то ее вершина может нагнать подножие,
возникает ударная волна, при образовании которой большая часть
энергии рассеивается в виде тепла. И в том и в другом случае до-
стигается равновесие между нелинейным эффектом увеличения
крутизны фронта волны и рассеянием энергии, так что устанавли-
вается стационарное состояние. (Когда амплитуда волн на воде
или звуковых волн весьма мала, крутизна фронта не слишком воз-
растает, и небольшого рассеяния энергии, связанного с самим вол-
новым движением, достаточно, чтобы противодействовать этому
эффекту.) Соответственно можно предвидеть, что и в артериальной
системе при достаточно большой амплитуде пульсовых волн давле-
ния способна развиваться своего рода «ударная волна». Из выпол-
ненных на ЭВМ расчетов нелинейных эффектов при распростра-
нении волн по артериям следует, что у больных с недостаточностью
аортального клапана, у которых ударный объем левого желудочка
очень велик, а потому велика и скорость й, должны наблюдаться
явления типа ударной волны. Тем не менее еще ни один исследо-
ватель не изучал в подробностях динамику таких «ударных волн»
ни теоретически, ни экспериментально и никто не выяснил меха-
низма сильной диссипации энергии, аналогичной диссипации, на-
блюдаемой при разрушении волн на воде. Следует отметить, од-
нако, что в артериях больных с недостаточностью аортального кла-
пана иногда улавливают звуки типа «пистолетного выстрела», ко-
торые, вполне возможно, порождаются эпизодически возникаю-
щими «ударными волнами».
В отличие от артерий большого круга его вены, а также арте-
рии малого круга значительно более растяжимы, а колебания дав-
ления в этих сосудах составляют большую долю характерного для
них среднего давления. Поэтому роль нелинейности для указанных
сосудов может быть весьма существенной. Однако соответствую-
щие детальные исследования столь малочисленны, что доказатель-
ства этого утверждения почти отсутствуют. Данным вопросом мы
займемся в гл. 14 и 15, посвященных венам большого круга и ма-
лому кругу ').
’) В разд 14 4 приводятся также и важные дополнительные сведения о не-
линейных эффектах в артериях большого круга — в связи с проблемой возник-
новения тонов Короткова. — Прим, ред.
362 Часть II. Механика кровообращения
12.6. Влияние вязкости
Итак, мы уже составили довольно полное представление как
о механизмах распространения пульсовых волн давления, так и
о причинах изменений их формы по мере перемещения по арте-
риям. Все эти изменения, кроме увеличения крутизны фронта пуль-
совой волны, объяснимы в рамках линейной теории. Однако ни
один из рассмотренных выше факторов не позволяет объяснить
форму реальной пульсовой волны расхода (рис. 12.19), которая,
согласно линейной теории, должна иметь такую же форму, как
пульсовая волна давления, и быть в фазе с ней (см. сноску на
стр. 335). Объяснить наблюдаемую форму пульсовой волны рас-
хода можно лишь в том случае, если учесть вязкие свойства крови.
Вязкость играет существенную роль также и в ослаблении (за-
тухании) пульсовых волн давления. Как отмечалось в разд. 8.3,
затухание волн должно происходить в любой реальной системе:
причиной превращения механической энергии волн в тепло служит
в таких системах наличие трения или вязкости. Реально рассеяние
энергии пульсовой волны, обусловленное вязкоупругими свойст-
вами стенки сосудов, более значительно, чем вызываемое вязко-
стью крови.
Влияние вязкости крови на форму пульсовой волны расхода.
На рис. 12.35 изображена форма волн давления и расхода крови,
зарегистрированных одновременно в бедренной артерии собаки.
Для фазы ускоренного изгнания крови обе волны можно практи-
чески совместить (выбрав подходящий масштаб по оси ординат),
однако затем их форма становится существенно различной. Более
того, вершина волны расхода заметно опережает вершину волны
давления, тогда как, согласно упрощенной теории, не учитывающей
вязкие свойства, обе волны должны быть синфазны. Следователь-
но, влияние вязкости здесь существенно.
Чтобы понять, как вязкость приводит к таким особенностям
волны расхода, необходимо снова обратиться к анализу Фурье и
рассмотреть связь между соответствующими синусоидальными со-
ставляющими волн давления и расхода крови. Как отмечалось
в гл. 4, движение крови определяется не давлением как таковым,
а местным градиентом давления. Однако если синусоидальная вол-
на давления имеет вид
р = ро cos [<о (t — х/с)]
[см. уравнение (12.20)], то действующий в направлении х гра-
диент давления (—dp/dx) есть
— -^p-sin [©(/ — х/с)].
Следовательно, градиент давления также изменяется синусои-
дально, с той же угловой частотой и опережает давление пр
Гл. 12. Артерии большого круга 363
Рис. 12.35. Зарегистрированные одновременно волны давления и расхода крови
в бедренной артерии собаки. Изображены также волна градиента давления (вни-
зу) и построенная по ней с помощью теории нестационарного течения в жесткой
трубке волна расхода крови. [McDonald (1974). Blood flow in arteries. Edward
Arnold, London.]
фазе на л/2, т. е. на 90°. (В разд. 8.1 объяснялось, что значит опе-
режает или отстает по фазе; сказанное здесь — «градиент давле-
ния на л/2 опережает давление» — означает, что при данном х
градиент давления достигает максимума на л/2© раньше, чем
давление.) В гл. 5 мы уже обсуждали характер течения жидкости
в прямой жесткой трубке под действием синусоидального градиен-
та давления. Тот же самый анализ применим, как оказывается, и
к артериям, которые, не будучи жесткими и растягиваясь, все же
сохраняют при этом примерно цилиндрическую форму (разд. 12.8).
В ответ на синусоидальные изменения градиента давления с ча-
стотой © возникают синусоидальные же по форме волны расхода
той же частоты (поскольку волны взаимосвязаны линейно), од-
нако амплитуда и фаза волн расхода зависят от параметра
Уомерсли а. Когда значение а велико, толщина пограничного слоя
много меньше радиуса трубки и расход вслед за изменением гра*
диента давления меняется почти так же, как при течении невязкой
жидкости. Однако при малых значениях а вязкость жидкости су-
щественна во всей толще потока и расход в любой момент вре-
мени связан с градиентом давления так, как это присуще пуазей-
левскому течению. Подробности связи между градиентом давления
364 Часть II. Механика кровообращения
и расходом жидкости рассчитаны для всех значений а. Как отме-
чалось выше, для самых больших артерий крупных млекопитаю-
щих значение а велико (больше 10), а для микрососудов оно много
меньше единицы.
Чтобы проверить, в какой мере применима к артериям теория,
разработанная для течения вязкой жидкости в жесткйх трубках,
необходимо: 1) разложить зарегистрированные волны градиента
давления на гармонические (т. е. синусоидальные) составляющие;
2) найти составляющие волны расхода, соответствующие каждой
составляющей волны градиента давления, и 3) просуммировав гар-
монические составляющие волны расхода, получить ее предпола-
гаемую форму. Затем расчетную форму волны следует сравнить
с полученной в опыте. На рис. 12.35 приведены результаты подоб-
ного сопоставления; здесь представлены кривые пульсовых волн
расхода крови: зарегистрированная в бедренной артерии собаки и
построенная по кривой для градиента давления. (Значение а для
основной гармоники, соответствующей частоте сокращений сердца,
равно 1,4, т. е. лишь немногим больше единицы.) Для построения
расчетной кривой были использованы только первые четыре гар-
моники, и тем не менее соответствие с экспериментальной кривой
очень хорошее, исключая лишь несколько меньшие, чем в действи-
тельности, колебания скорости кровотока во время диастолы. Та-
ким образом, ясно, что теория, разработанная для жестких трубок,
вполне применима для расчета колебаний расхода крови в арте-
риях, соответствующих определенным колебаниям градиента дав-
ления, хотя упругость сосудов (но не вязкость крови) сильно
влияет на форму таких колебаний градиента давления.
Влияние вязкости крови на распространение волн. Наличие у
крови вязкости приводит как к уменьшению скорости распростра-
нения волн давления, так и к их затуханию. Оба этих эффекта
возникают потому, что при течении вязкой жидкости проскальзы-
вание крови относительно стенки сосуда в продольном направле-
нии, которое допускается теорией невязкой жидкости, невозможно.
Это означает, во-первых, что при одном и том же градиенте давле-
ния амплитуда колебаний расхода вязкой жидкости оказывается
несколько меньше, чем невязкой; следовательно, за счет вязкости
как бы увеличивается эффективная инерционность жидкости, что
и приводит к уменьшению скорости волны. Во-вторых, в погранич-
ном слое развиваются высокие скорости сдвига, обусловливающие
диссипацию механической энергии, а следовательно, и затухание
волн. Так как толщина пограничного слоя и скорость сдвига в нем
зависят от величины а, следует ожидать, что скорость волны с и
степень затухания волны (на расстоянии, равном длине волны)
также будут зависеть от а.
Метод количественного описания затухания волн рассмотрен
в разд. 8.3. Синусоидально меняющиеся компоненты волны давле-
ния р зависят от времени t и расстояния х по длине сосуда еле-
Гл. 12. Артерии большого круга 365
дующим образом:
р = poe~kx/K cos — у) — <р
(12.26)
где ро — амплитуда колебания давления, <р — фазовый угол, с —
скорость волны, со — угловая частота, Л = 2пс/ш — длина волиы.
Далее, поскольку при распространении волны на расстояние Л ее
амплитуда уменьшается в ek раз, степень затухания волны опре-
деляют константой k. Детальный теоретический анализ, учитываю-
щий влияние вязкости, показывает, что с и e~k меняются с ростом а
так, как это изображено на рис. 12.36.
Рис. 12.36. А Зависимость отношения рассчитанной теоретически скорости волны
с к скорости Со> полученной по формуле Моенса — Кортевега, от а. I — попереч-
ные волны давления, II— продольные сдвиговые волны (см. разд 12.7). Б. Зави-
симость рассчитанного теоретически фактора затухания (е-*) от а. I — волны
давления, II — сдвиговые волны. [Atabek (1968). Wave propagation through a
viscous fluid contained in a tethered, initially stressed, orthotropic elastic tube.
Biophys. J., 8, 626—49 ]
Из приведенных кривых видно, что при всех а > 1,5 скорость
волны примерно постоянна и равна величине Со, определяемой по
формуле Моенса — Кортевега [формула (12.11)]. Поэтому у круп-
ных млекопитающих, у которых значение а для основной гармо-
ники, соответствующей частоте сердцебиений, больше 10, все си-
нусоидальные составляющие пульсовой волны давления распро-
страняются по аорте с одинаковой скоростью. Однако в случае
значительно более мелких артерий величины а для основной гар-
моники меньше 1,5 и, как это следует из рис. 12.36, А, в этих ар-
териях высокочастотные составляющие пульсовой -волны распро-
страняются с большей скоростью, чем низкочастотные. Это явле-
ние, называемое дисперсией, — еще одна возможная причина изме-
нения формы пульсовой волны при ее распространении по арте-
риям. Но, как мы видели, для крупных артерий оно, по-видимому,
несущественно.
Расчеты показывают, что затухание волн даже в наиболее круп-
ных артериях может быть весьма заметным. Из рис. 12.36,5 видно,
366 Часть И. Механика кровообращения
что оцениваемое по ek затухание волны при прохождении ею
расстояния, равного длине волны Л {затухание на длине волны),
уменьшается с увеличением а (т. е. с повышением частоты1)). Од-
нако ослабление волны при прохождении ею единицы длины сосуда
(затухание на единицу длины) увеличивается с ростом частоты,
так как К уменьшается. Величина kfk [см. уравнение (12.26)] про-
порциональна (й1/2, и потому высокочастотные составляющие волны
при прохождении ими некоторого определенного расстояния вдоль,
сосуда затухают быстрее, чем низкочастотные.
Степень затухания волны на участке артерии, имеющем дан-
ную длину, легко оценить из рис. 12.36, Б [или по формуле (12.27)].
В случае аорты собаки при частоте сокращений сердца около 2 Гц
(со = 4л) для основной гармоники а равно примерно 11. Соответ-
ствующее значение k составляет примерно 0,4, и, следовательно,
степень затухания волны на расстоянии, равном ее длине, согласно
расчетам, достигает 33%. Поскольку длина волны равна примерно
3 м, степень затухания ее при прохождении аорты будет всего лишь
5°/о- Для волн с частотой 40 Гц получаем k ж 0,1, а для волн с ча-
стотой 100 Гц—еще меньше — 0,06. На расстоянии, равном длине
аорты собаки, волны такой частоты должны были бы, таким обра-
зом, затухать на 24 и 33% соответственно. Возвращаясь к уже упо-
минавшимся опытам, в которых на пульсовые волны давления в
аорте собаки накладывали искусственно созданные высокочастот-
ные— от 40 до 150 Гц—колебания, отметим, что в этих опытах
были произведены и очень тщательные измерения степени затуха-
ния. Она оказалась гораздо более высокой, чем это следовало из
теории. Результаты этих опытов показали, что k практически не
зависит от частоты, и его значения почти всегда лежат в интервале
0,7—1,0.
Эти данные находятся в соответствии с результатами измере-
ний степеней затухания на значительно более низких частотах
(рис. 12.37). Следовательно, теория, учитывающая только вязкие
свойства крови, не способна правильно предсказать коэффициент
затухания волн, особенно высокочастотных. К дополнительному
затуханию должны приводить процессы в стенке сосудов, связан-
ные с диссипацией энергии из-за вязкоупругих свойств как мате-
риалов стенки, так и ткани, к которой эта стенка прикреплена
(разд. 7.2). Прежде чем обратиться к этому вопросу, следует от-
метить, что, согласно измерениям in vivo, затухание высокочастот-
’) Значение k (рис. 12,36, Б) для больших а (превышающих примерно 6)
приближенно определяется соотношением
k ta (12.27)
Соответствующая скорость волны с равна
с = с0(1- 1/ад/2*).
где Со — скорость волны, определяемая по формуле Моенса — Кортевега.
Гл. 12. Артерии большого круга 367
ных волн, распространяющихся против течения, значительно боль-
ше (k — 1,3—1,5), чем волн, перемещающихся по течению (k =
= 0,7—1,0). Вероятной причиной этого различия является измене-
ние амплитуды волн, обусловленное конусообразным сужением
аорты и увеличением ее жесткости по направлению к периферии.
Такое изменение амплитуды, согласно теории, должно приводить
Рис. 12.37. Зависимость фактора затухания е-* от а для пульсовых воли дав-
ления в аорте собаки. I — теоретическая кривая, полученная в предположении,
что затухание обусловлено лишь вязкостью крови, // и III — величины, измерен-
ные в двух сериях опытов. [McDonald (1974). Blood flow in arteries, Edward
Arnold, London.]
к уменьшению затухания распространяющихся в периферическом
направлении волн и к увеличению затухания волн, перемещаю-
щихся в противоположном направлении.
Влияние вязкоупругих свойств стенки. В гл. 7 (разд. 7.2) го-
ворилось о том, что при учете вязкоупругих, а не одних только
чисто упругих свойств стенки сосуда изменяется соотношение, свя-
зывающее избыточное давление с площадью поперечного сечения
сосуда. Если напряжение в его стенке колеблется с амплитудой S
и частотой со, то возникающие деформации, т. е. в данном случае
изменения периметра сосуда — имеют амплитуду S/VЕдин + т)2©2
и отстают по фазе от напряжения на величину, равную примерно
т)ш/Едии, где Един — динамический модуль Юнга (который больше
статического модуля Юнга £), а цю — так называемый модуль вяз-
ких потерь1).
Соответственно роль растяжимости играет величина
DE/д/Ё^ии + т)2©2, где D определено формулой (7.36), а изменение
площади поперечного сечения отстает по фазе от изменений давле-
ния на величину, также равную г]и/ЕдИН. В результате оказывается,
*) Используя комплексные числа, модуль Юнга Е в уравнении (7.1) можно
заменить выражением Един + й]ю. Аналогично растяжимость D в уравнении
(7.36) можно с некоторым приближением записать в виде DE (Едии—щ<в/(Е^ +
+ П2®2).
368 Часть II. Механика кровообращения
что из-за уменьшения фактической растяжимости стенки сосуда ско-
рость волны несколько возрастает (в отличие от уменьшения ее,
обусловленного вязкостью крови), но вместе с тем усиливается за-
тухание волны в процессе ее перемещения по сосуду. В этом слу-
чае степень затухания на длине волны также может быть охарак-
теризована константой k [уравнение (12.26)], которая приблизи-
тельно равна произведению 2л на фазовый сдвиг т](о/Ддии, выра-
женный в радианах, при условии, что последний мал. Но, как уже
говорилось в гл. 7, отношение tjoj/Z?ДИи примерно одинаково для всех
частот выше 5 Гц и равно 0,1—0,2. Следовательно, значения k
должны лежать в интервале 0,6—1,2, что и было обнаружено в ука-
занных выше опытах (рис. 12.37). Поэтому представляется, что
простая теория вязкоупругих свойств стенки достаточно хорошо
описывает затухание высокочастотных волн. Для более низких ча-
стот (менее 5 Гц), т. е. для основной и первой следующей за ней
гармоник пульсовой волны, фазовый сдвиг и, следовательно, зна-
чение k меньше. Вместе с тем на затухании низкочастотных состав-
ляющих волны в большей мере сказывается вязкость крови, и по-
тому роль последней и вязкоупругих свойств стенки артерий в за-
тухании основной гармоники пульсовой волны давления сопоста-
вима. Поскольку, однако, длина волны основной гармоники очень
велика, измерить в опытах вычисленную для этой гармоники сте-
пень затухания (k < 1) весьма непросто. Возникающие здесь труд-
ности связаны, в частности, с изменением формы волны, обуслов-
ленным ее отражением.
12.7. Волны других типов
Помимо волн давления по артериям могут распространяться
волны других типов. Предположим, что артерия — это длинная
тонкостенная упругая трубка, которая не прикреплена к окружаю-
щим тканям и содержит вязкую жидкость. Допустим также, что
проводится математический анализ такой модели, основанный на
следующих весьма правдоподобных предположениях: а) жидкость
движется лишь вдоль оси трубки и по ее радиусу и б) длины всех
волн много больше радиуса трубки. В таком случае в результате
вычислений мы получим два возможных значения скорости волны с
и два соответствующих каждой из них показателя затухания e~ft
(расчет последних основан на значении со, полученном без учета
вязкости жидкости). Одна пара величин, ci и e_ftl, соответствует
волнам давления; их зависимость от а приведена на рис. 12.36.
Вторая пара величин, с2 и е-*2, зависимость которых от а
также изображена на рис. 12.36, соответствует волнам другого
типа. Обычно их называют быстрыми волнами, так как с2 при лю-
бых а больше, чем сь а при а, стремящемся к бесконечности, ста-
новится бесконечно большой. Волны давления сопряжены в основ-
ном с радиальными движениями стенки трубки, тогда как волны
Гл. 12. Артерии большого круга 369
второго типа — главным образом с ее продольными движениями.
Элементы стенки трубки колеблются в продольном направлении
под действием возвращающей силы, зависящей от продольного
растяжения стенки. Инерция, противостоящая возвращающей силе,
определяется массой вязкой жидкости, которая благодаря напря-
жениям сдвига вовлекается в колебательное движение вместе со
стенкой трубки. При больших значениях а вместе со стенкой ко-
леблется только тонкий пограничный слой жидкости, а основная
ее масса не участвует в этом движении. С увеличением а погра-
ничный слой становится еще тоньше, и, следовательно, влияние
инерции жидкости уменьшается. Поэтому скорость волны возрас-
тает, что объясняет приведенные на рис. 12.36, Л результаты для
больших а. Естественно, что в реальных условиях скорость волны
с2 никогда не становится бесконечно большой, так как ее рост
ограничен инерцией стенки сосуда.
На первый взгляд кажется удивительным, что при очень ма-
лых а скорость распространения быстрых волн не становится рав-
ной нулю, а затухание оказывается пренебрежимо малым. Все это,
однако, можно понять, если учесть, что в данном предельном случае
продольные колебания участка стенки трубки заставляют всю на-
ходящуюся в этом участке жидкость колебаться как твердое тело.
Движение жидкости в любой точке синфазно с движением стенки,
и, следовательно, напряжение сдвига равно нулю. Поэтому при
таком движении энергия не рассеивается в жидкости и волна не
затухает.
В принципе возможны еще и волны третьего типа: когда в
стенке существуют крутильные колебания, вязкая жидкость под
действием напряжения .сдвига движется по окружности. Такое
движение создает инерционные эффекты, уравновешивающие уп-
ругую возвращающую силу. Мы не будем далее рассматривать
крутильные колебания, так как в сосудах их не наблюдали.
Быстрые сдвиговые волны второго типа in vivo также факти-
чески не наблюдали, и скорее всего потому, что стенка артерий
прикреплена к окружающим тканям. Такое прикрепление практи-
чески полностью подавляет продольные (и крутильные) движения
стенки, но допускает ее значительные радиальные перемещения.
Это явление удалось промоделировать, приняв, что артериальная
стенка прикрепляется к жесткому основанию пружинками, обла-
дающими заданными упругими и вязкими свойствами. Даже если
эти свойства выбирали так, чтобы скрепление было слабее, чем в
действительности, расчеты показывали, что быстрые сдвиговые
волны почти полностью подавлены.
Введение представления о закреплении артерии в окружающей
ткани — это лишь одно из многочисленных усовершенствований
простой модели артерии, которые были сделаны для того, чтобы
оценить значение тех или иных факторов. Например, была иссле-
дована роль инерции собственно стенки артерии, которая ограничи-
370 Часть II. Механика кровообращения
вает максимальное значение скорости распространения сдвиговых
волн, увеличивая наряду с этим роль инерции в распространении
пульсовой волны давления и несколько уменьшая тем самым ее
скорость. Было построено множество моделей упругой стенки арте-
рии, отличных от той, в которой стенку считают тонкой, а ее мо-
дуль Юнга — однородным. При этом рассмотрены возможные след-
ствия, которые привносит толстая стенка сосуда, обладающая ани-
зотропными упругими свойствами и подвергающаяся различным
начальным напряжениям; учтено также влияние вязкоупругих
свойств стенки. Однако все эти факторы сказываются только на
связи растяжимости D сосуда как конструкции с упругими свой-
ствами его стенки. На связи же скорости волны и растяжимости
[уравнение (12.9)] они не отражаются и, следовательно, не влияют
на основные механические закономерности распространения волн
(за исключением того, что процессы, сопровождающиеся диссипа-
цией энергии, приводят к изменениям фазового сдвига и затуха-
нию волн)1).
В заключение следует упомянуть об еще одном типе волн, кото-
рые могут существовать в артериях, — об истинных звуковых вол-
нах. Возвращающая сила в этом случае обусловлена не растяжи-
мостью стенки артерий, а сжимаемостью жидкости (или твердого
тела), в которой они распространяются. Звуковые волны в крове-
носных сосудах и других тканях, несомненно, существуют, но, по-
скольку скорость их очень велика (около 1500 м-с-1), они пере-
даются по сердечно-сосудистой системе практически мгновенно,
а их динамические воздействия ничтожно малы. Пренебрегать зву-
ковыми волнами при анализе пульсовых явлений — то же самое,
что считать кровь несжимаемой (как это и предполагается во всей
книге). Физическая природа тонов сердца рассмотрена в гл. 11.
12.8. Картины течения крови в артериях
Обратимся теперь к изучению картин течения крови в артериях,
имея в виду две цели: во-первых, объяснить особенности кривых,
получаемых при измерениях скорости движения крови в арте-
риях и, во-вторых, понять, каким образом локальные особенности
течения крови в артериях влияют на физиологические процессы,
протекающие в их стенке. Рассмотрев экспериментальные данные
о картинах течения крови в крупных артериях, мы перейдем затем
к обсуждению физических механизмов, лежащих в их основе.
Профили скорости в крупных артериях. В последние годы был
разработан ряд методов измерения скорости течения крови в ар-
’) Подробный обзор различных видоизменений простой упругой модели ар-
терий приведен в работе Сох R. Н. (1969). Comparison of linearized wave pro-
pagation models for arterial blood flow analysis. Journal of Biomechanics, 2,
251—265. Имеется перевод: Кокс P. Г. Сравнение моделей артериального дви-
жения кровн, основанных на линеаризованных теориях распространения волн.
В кн.: Гидродинамика кровообращения. М.: Мир, 1971, с. 43—60.
Гл. 12. Артерии большого круга 371
териях животных in vivo. Некоторые из этих методов позволяют
определить расход крови через весь просвет сосуда (электромаг-
нитный метод с использованием «манжеточных» датчиков) или че-
рез большую часть просвета (тот же метод с зондовыми датчи-
ками). При помощи таких методов измеряют среднюю по сечению
сосуда скорость кровотока. Другие методы, особенно метод пле-
ночной термоанемометрии, а также ультразвуковой импульсный
доплеровский метод дают возможность найти скорость движения
крови в отдельных точках просвета сосуда и построить распреде-
ление скорости по сечению. Весьма многообещающим представ-
ляется ультразвуковой доплеровский метод, так как с его помощью,
не вводя датчик в сосуд, можно получать в виде графика практи-
чески мгновенное распределение скоростей движения крови по се-
чению сосуда (в принципе даже у бодрствующих животных). Од-
нако этот метод до конца не разработан и объем собранных с его
помощью данных о картинах кровотока в артериях пока невелик.
Поэтому мы остановимся на исследованиях, в которых применялся
метод термоанемометрии.
Миниатюрные, чувствительные к скорости движения крови дат-
чики пленочного термоанемометра помещают на кончике иглы для
подкожных инъекций или зонда, которые вводят в артерию. Эти
датчики «чувствуют» скорость движения крови только в пределах
одного миллиметра (или менее) своей поверхности, поэтому они и
позволяют измерять локальные скорости течения крови в разных
точках сечения сосуда. Эти датчики можно использовать и для
изучения возмущенного и турбулентного потока крови в сосудах,
поскольку они способны воспринимать колебания скорости с часто-
тами до 500 Гц.
Метод пленочной термоанемометрии выявил многие детали рас-
пределения скорости течения крови в аорте собак. С его помощью
удалось провести ряд наблюдений и на человеке. Но ни в том ни
в другом случае даже самые миниатюрные датчики не позволяют
изучать течение крови в пограничном слое. Этот слой так тонок,
что здесь, у самой стенки, любые датчики становятся ненадеж-
ными: отчасти из-за влияния на сигнал близости стенки, отчасти
из-за радиальных смещений последней. Дело в том, что в течение
цикла сокращения сердца положение фиксированного в просвете
артерии датчика относительно ее стенки изменяется, а кроме того,
происходит перемещение крови в радиальном направлении с за-
метной скоростью, в результате чего искажаются записи скорости
продольного течения. Таким образом, поскольку измерения скоро-
сти у самой стенки ненадежны, метод пленочной термоанемоме-
трии можно использовать для изучения профиля скорости только
в крупных артериях, у собак — в аорте. Но и здесь измерения осу-
ществимы лишь в определенных участках. Это обусловлено труд-
ностями хирургического доступа и необходимостью сохранения
формы сосуда. Уже не один раз было показано, что изменение
372 Часть Л. Механика кровообращения
Рис. 12 38 Профили скорости в разных участках аорты собаки. Для всех участков
скорости в разных точках просвета сосуда отнесены к скорости на оси аорты
[Schultz (1972). Pressure and flow in large arteries Cardiovascular fluid dyna-
mics (ed. Bergel), vol. I, Academic Press, New York.]
формы сосуда коренным образом меняет профиль скорости. И на-
конец, последняя сложность, с которой приходится сталкиваться
при изложении и объяснении данных, полученных этим методом,
заключается в том, что исследователи не всегда точно определяют
место измерения и те сечения сосуда, по которым перемещали дат-
чик. Как будет показано ниже, местные геометрические особенно-
сти сосуда существенно влияют на течение крови, и расхождение
результатов, полученных в различных исследованиях или на жи-
вотных разных видов, можно, вероятно, связать именно с этим фак-
тором. Дальше мы остановимся лишь на твердо установленных
основных особенностях картин течения крови.
Анализ профиля средних скоростей (т. е. профиля, построен-
ного по результатам измерений средних по времени скоростей)
в различных участках аорты собаки (рис. 12.38) показывает, что на
всем протяжении аорты профиль остается практически плоским, и
только измерения, произведенные непосредственно у стенки, выяв-
ляют, что здесь течение медленнее, чем на оси сосуда. Вблизи
Гл. 12. Артерии большого круга 373
Рис. 12.39. Мгновенные профили скорости в восходящей аорте собаки в шесть
различных моментов пульсового цикла. Эти моменты отмечены точками на схеме
пульсовой волны скорости крови (внизу).
сердца такое уменьшение скорости фактически не обнаруживается.
Кровь движется здесь в виде как бы единого столбика, а присте-
ночный пограничный слой столь тонок, что скорость в нем не
удается измерить. На практике это означает, что толщина слоя
менее 2 мм.
Сказанное справедливо для картин течения крови в грудной
аорте на протяжении всего пульсового цикла. На рис. 12.39 изоб-
ражены мгновенные профили скорости в восходящей аорте для
шести разных моментов этого цикла. Видно, что в фазу ускоряю-
щегося движения крови имеет место значительный перекос про-
филя скорости. Однако какие-либо признаки того, что хотя бы на
одной стадии пульсового цикла преимущественное снижение ско-
рости приходится на слои вблизи стенки, отсутствуют. Пытаясь
приблизиться к изучению сложного вопроса о распределении ско-
рости в этих тонких пограничных слоях, некоторые исследователи
ставили опыты на животных, аорта которых больше по диаметру,
чем у собаки (свинья, лошадь). Чтобы устранить радиальные дви-
жения стенки аорты, в опытах на свиньях соответствующий уча-
сток сосуда фиксировали в жесткой манжетке. Но даже при этих
условиях не удалось сколько-нибудь подробно исследовать тече-
ние крови в пограничном слое, поскольку его толщина оказалась
374 Часть II. Механика кровообращения
меньше 1,5 мм. Как мы увидим несколько ниже, теоретически по-
граничный слой и должен быть тонким.
Тем не менее эти исследования выявили важные детали свойств
течения крови в ядре потока. В области входа в аорту, на 1—2 см
ниже аортального клапана, профиль скорости во время систолы
обычно симметричный, по крайней мере в плоскости главной кри-
визны дуги аорты (на данном уровне аорты это единственно до-
ступная для измерения плоскость, и здесь она проходит фактиче-
ски в передне-заднем направлении — рис. 12.40, внизу). Именно
такого профиля скорости и следовало ожидать для течения в об-
ласти входа (разд. 5.2), но здесь же обнаружены и некоторые дру-
гие, не столь хорошо известные особенности течения.
Прежде всего на записях волн скорости крови в области входа
в аорту часто выявляются наложенные на систолический фронт
волны низкочастотные возмущения. Они могут существенно ме-
няться от одного сокращения сердца к другому или, же, напротив,
повторяться с таким постоянством, что приводят к перекосу про-
филя скорости при каждом сокращении сердца, обычно в конце
систолы. Эти возмущения могут представлять собой «следы» за-
вихрений потока, которые, как известно, развиваются в желудочке
(разд. Н.5), или же возникать при прохождении крови через аор-
тальный клапан. Измерения скорости движения крови в желудочке
с помощью датчиков, закрепленных на конце зонда, показали, что
возмущения там существуют в течение как систолы, так и диа-
столы. Это позволяет предположить, что возмущения, возникаю-
щие в желудочке во время диастолы, продолжают существовать
и во время изгнания крови и поэтому могут быть источником воз-
мущений скорости ее течения в аорте. Однако подобного рода дан-
ные не являются бесспорными, так как зонды, находящиеся в желу-
дочке, могут испытывать в течение систолы толчки, а они приводят
к искажению сигнала скорости. Поэтому природу низкочастотных
возмущений скорости течения крови в восходящей аорте нельзя
считать твердо установленной.
Другая любопытная особенность течения крови состоит в том,
что симметричность профиля скорости, наблюдающаяся во время
систолы вблизи сердца, утрачивается по мере перемещения пото-
ка по восходящей аорте в ее дугу. Скорость течения крови у внут-
ренней (задней) стенки дуги возрастает, что приводит к четко
выраженному перекосу профиля (рис. 12.40, вверху).
Асимметричность профиля скоростей на входе в дугу аорты на-
блюдается постоянно. Это явление объясняют по-разному. Напри-
мер, утверждают, что на профиль скорости оказывает влияние
близость крупных ветвей, отходящих от дуги аорты, и это приво-
дит к существенным местным возмущениям течения (см., в част-
ности, рис. 12.47). Согласно другому предположению, перекос про-
филя скорости связан с сильно выраженной кривизной дуги аорты.
В потоке, движущемся по подобному изгибу, возникает поперечный
Гл. 12. Артерии большого круга 375
Рис. 12.40. Профили скорости в двух участках восходящей аорты собаки при
пиковых значениях скорости крови в течение систолы. Все значения скорости
(и) отнесены к скорости на оси сосуда (ио), а расстояния от оси — к радиусу
сосуда. В обоих участках аорты профили построены по данным исследований
на нескольких животных, причем во всех случаях скорости измеряли в плоскости
главной кривизны дуги аорты.
градиент давления, и, таким образом, вдоль диаметра аорты в пло-
скости оси дуги давление возрастает от внутренней стенки к на-
ружной. Это приводит к относительному замедлению скорости те-
чения невязкой жидкости (каковой можно считать кровь в ядре
потока в аорте) вблизи наружной (передней) стенки. Подробно это
явление рассмотрено в гл. 5 (разд. 5.8). Такое предположение на-
ходится в соответствии и с данными о том, что профиль становится
376 Часть fl. Механика кровообращения
симметричным, когда скорость течения крови уменьшается, а в на-
чале диастолы, в фазе обратного движения крови, вновь возникает
асимметрия. Кроме того, изложенное представление объясняет,
почему степень асимметричности профиля скорости неодинакова
у различных животных: она определяется индивидуальными осо-
бенностями геометрии дуги аорты (ее диаметром и радиусом кри-
визны). Последнее, конечно, остается в силе и в том случае, если
перекос профиля скоростей определяется отхождением артериаль-
ных ветвей, и весьма вероятно, что в нарушении симметрии про-
филя скорости играют роль оба механизма.
Профили скорости были исследованы и в направлениях, пер-
пендикулярных плоскости дуги аорты. Вблизи аортального
клапана этого сделать не удается, так как здесь аорту загоражи-
вает легочный ствол, но на входе в дугу аорты (примерно на уров-
не, указанном верхней стрелкой на рис. 12.40) такие измерения
возможны. Перекос профиля скорости обнаруживается и в этом
случае, причем на протяжении систолы скорость больше по на-
правлению к левой стенке дуги аорты. Поскольку в плоскости, пер-
пендикулярной плоскости дуги, ход сосуда относительно пря-
мой, едва ли это явление связано с кривизной; четкого объяснения
ему пока не найдено.
В восходящей аорте средняя по сечению скорость течения кро-
ви, определенная, например, с помощью манжеточного датчика
электромагнитного расходомера, в течение диастолы близка к
нулю. Однако измерения, произведенные пленочным термоанемо-
метром, показывают, что на самом деле движение крови во время
диастолы не прекращается. В части просвета аорты может наблю-
даться местное течение, направленное обратно к сердцу и согла-
сующееся с оттоком крови в коронарные артерии. Кроме того, на
входе в дугу аорты возможно одновременное движение крови впе-
ред в одной части сечения сосуда и назад — в другой. Это позво-
ляет высказать предположение о наличии движения крови по за-
мкнутым траекториям, возможно способствующего ее оттоку в
ветви дуги аорты, где в течение диастолы может поддерживаться
направленный к периферии поток.
Для проксимального отдела нисходящей аорты измерения воз-
можны только в плоскости, перпендикулярной плоскости дуги.
И здесь во время диастолы кровь может двигаться по замкнутым
траекториям; что же касается систолы, то данные о каком-либо
влиянии крупных артерий, отходящих сразу же перед этим отде-
лом аорты, на картину течения крови в ней практически отсут-
ствуют. Систолический профиль скорости остается плоским, с не-
большим перекосом в сторону правой стенки; профили средних по
времени скоростей обычно симметричны (рис. 12.38). В более ди-
стальных отделах аорты появляется начальное «скругление»
профиля средних по времени скоростей, видимо, вследствие влия-
ния вязких сил и увеличения толщины пограничного слоя.
Гл. 12. Артерии большого Круга 377
Пленочные термоанемометрические датчики обычно слишком
йелики, чтобы детально исследовать картины течения крови й
брюшном отделе аорты собак и в крупных ветвях аорты. Однако
недавно удалось провести соответствующие исследования на лоша-
дях, у которых возможен доступ к гораздо более периферическим
артериям, чем у собак (у крупных лошадей диаметр восходящей
аорты может составлять 5 см, а плече-головной и подвздошной ар-
терий— 1—3 см). На первый взгляд опыты на крупных животных
представляются также очевидным путем для решения сложной
задачи исследования течений в пограничных слоях. Но в действи-
тельности толщина пограничного слоя для пульсирующего течения
йе зависит от диаметра сосуда и пропорциональна (разд. 12.3).
Кинематическая вязкость крови при высоких скоростях сдвига
практически одинакова для большинства млекопитающих, так что
толщина пограничного слоя зависит от 1/V®» и следовательно, из-
меняется обратно пропорционально квадратному корню из часто-
ты сокращений сердца. Таким образом, если сравнивать толщину
пограничного слоя у животных разных видов, то она больше
у того вида, у особей которого сердце сокращается с меньшей
частотой.
Вообще говоря, чем крупнее животное, тем с меньшей частотой
сокращается у него сердце. Вот почему есть все основания иссле-
довать пограничный слой на сосудах крупных млекопитающих.
Например, частота сокращений сердца у лошади составляет 30—
50 мин-1, в то время как у собаки — 80—120 мин-1; следовательно,
можно ожидать, что в сосудах лошади пограничный слой примерно
на 60% толще, чем в сосудах собаки, и, как доказано, его действи-
тельно можно исследовать, хотя пока и не очень детально. На
рис. 12.41 представлены профили скорости в нисходящей грудной
аорте лошади для трех разных моментов систолы. Эти данные под-
тверждают, что во время направленного вперед, к периферии,
движения крови толщина пограничного слоя здесь весьма мала
(3—4 мм). Но, исследуя свойства этого слоя, нужно помнить, что
результаты измерений скорости непосредственно вблизи стенки не
могут быть очень точными из-за перемещений стенки и наличия
радиального течения. В случае грудной аорты часто наблюдаются
высокочастотные колебания скорости, указывающие на наличие
сильного возмущения течения или турбулентности. В более ди-
стальных отделах аорты это явление, однако, отсутствует.
На распределение скорости течения в брюшной аорте оказы-
вают влияние несколько факторов. Сразу же за местами отхожде-
ния от аорты чревной, краниальной брыжеечной и почечной арте-
рий (у лошади анатомическая организация брюшной аорты такая
же, как у собаки: см. рис. 12.1) скорость течения резко умень-
шается, что связано со значительным оттоком крови в соответ-
ствующие артерии. Появляются признаки действия вязких сил-
378 Часть 11. Механика кровообращения
Рис. 12.41. Профили скорости от осн сосуда до правой стенки в нисходящем
участке грудной аорты лошади. Измерения произведены в 10 см ниже дуги аор-
ты. На графике пульсовой волны скорости крови точками отменены моменты
пульсового цикла, для которых исследовали профили скорости. По оси абсцисс
отложено расстояние от осн сосуда в сантиметрах. Диаметр сосуда d = 2,69 см;
а = 19,4; пиковое значение числа Re — 2536. [Nerem, Rumberger jr., Gross, Ham-
lin, Geiger (1974). Hot-film anemometer velocity measurements of arterial blood
flow in horses. Circulation Res., 34, 193—203.]
профиль скорости становится более выпуклым, появляется пере-
кос, связанный, вероятно, с оттоком крови в вышележащую ветвь.
Пиковые значения числа Рейнольдса для этой области лежат в
интервале 200—800, возмущения течения крови отсутствуют. Еще
ниже, у самого конца аорты, наиболее примечательная особен-
ность течения заключается в том, что максимальные скорости на-
блюдаются не на оси сосуда, так что профиль скорости во время
систолы приобретает М-образную форму. Как связано (и связано
ли вообще) установление такого профиля в фазу движения крови
вперед, к периферии, с расположением поблизости области ветвле-
ния аорты — неясно. Такое же явление наблюдается в прямой, не-
разветвляющейся трубке (рис. 5.9), так что оно видимо есть осо-
бенность пульсирующего течения.
Немногочисленные измерения скорости течения крови в отхо-
дящих от аорты подвздошных артериях показали, что во время
Гл. 12. Артерии большого круга 379
систолы профили скорости здесь относительно симметричные и
заметно выпуклые. Картины течения крови в более дистальных
артериях методом пленочной термоанемометрии ни у одного вида
животных не изучали.
Физические механизмы, определяющие профили скорости.
В гл. 5 и в разд. 12.3 данной главы мы рассматривали пульсирую-
щее течение жидкости в длинной прямой жесткой трубке под дей-
ствием меняющейся во времени разности давлений на ее концах.
Результаты этих экспериментов, находящиеся в хорошем согласии
с теоретическими выводами, представлены на рис. 12.42. Эти ре-
зультаты не вполне применимы к течению крови в аорте потому,
что, как станет ясно чуть позже, среднее по времени течение в аор-
те не является полностью развитым. С другой стороны, выявлен-
ные в этих опытах пульсирующие компоненты течения, видимо,
соответствуют таковым в аорте.
В данном разделе мы рассмотрим стационарное и нестацио-
нарное пульсирующее течение крови в области входа в сосуд,
влияние кривизны сосуда на течение, а также картины течения
вблизи ветвлений. Мы будем пренебрегать растяжимостью стенок
артерий: играя главную роль в формировании местного градиента
давления при распространении пульсовой волны, растяжимость
артерий не влияет ни на расход, ни на профили скорости в движе-
нии, вызываемом этим градиентом. Объясняется это тем, что длина
пульсовой волны много больше расстояния, на которое переме-
щается выделенный элемент жидкости за время одного пульсового
цикла, поскольку скорость волны много больше скорости движе-
ния крови (разд. 12.3). Например, для аорты собаки длина пуль-
совой волны составляет 3—4 м, тогда как расстояние, которое
проходит выделенный объем крови (оно равно произведению сред-
ней скорости ее движения на длительность пульсового цикла), при-
мерно 10—20 см. Это значит, что участок артерии, по которому
в течение одного цикла сокращения сердца успевает переместиться
выделенный элемент жидкости, всюду имеет примерно одинаковую
площадь сечения. Тот факт, что на протяжении одной длины вол-
ны эта площадь меняется, здесь несуществен, в частности, потому,
что такие изменения составляют лишь около 10%. Растяжимость
артерии была бы важна только в том случае, если бы прохожде-
ние пульсовой волны приводило к значительному изменению пло-
щади поперечного сечения на длине порядка 10 см. Такое может
произойти, если фронт волны станет очень крутым (разд. 12.5),
но в обычных условиях этого не бывает.
Начальный участок. Для стационарного течения в прямой труб-
ке длину начального участка можно оценить, вычислив, где имен-
но в трубке пограничный слой начинает заполнять весь ее просвет
[уравнение (5.4)]. Развитие среднего по времени профиля скоро-
сти для пульсирующего потока такое же, как и для стационарного,
С тем лишь отличием, что в первом случае на средний профиль
380 Часть Л Механика кровообращения
Рис. 12 42. Мгновенные профили скорости потока в длинной прямой жесткой
трубке в восемь разных моментов времени одного цикла колебаний. Эта момен-
ты времени отмечены на схеме волны скорости {внизу слева). Волну создавали
путем наложения синусоидального колебания на стационарный поток, так, чтобы
период возвратного течения был очень коротким. Измерения производили на
таком расстоянии от начала трубки, чтобы среднее течение было полностью раз-
витым и устанавливался пуазейлевский профиль скорости (он также показан
на рисунке). Характеристики течения соответствовали характеристикам потока
крови в аорте, среднее значение числа Re — 1785, параметр Уомерсли а — 20,5;
пиковая скорость в 2,74 больше средней скорости Течение визуализировали,
создавая линии из мелких пузырьков водорода путем пропускания импульсов
электрического тока по натянутой перпендикулярно течению тонкой проволочке.
Эти пузырьки сносились потоком, н их фотографировали на одни негатив через
заданные интервалы времени. По такой фотографии {внизу справа) можно вы-
числить скорость в данный момент времени в каждой точке просвета трубки.
[С любезного разрешения Mr. Р, Minton и Dr, М. Clamen.]
Гл. 12. Артерии большого круга 381
всюду накладывается профиль, меняющийся во времени, форма
которого определяется значением параметра а1)- При исследова-
нии профиля скорости для любого момента сердечного цикла вы-
является, что этот профиль представляет собой сочетание стацио-
нарной (средней) и пульсирующей составляющих (как на
рис. 12.42). Длину начального участка для пульсирующей состав-
ляющей потока можно оценить, вычислив расстояния до того се-
чения трубки, где толщина пограничного слоя, возрастающая от
значения 61 непосредственно на входе в трубку, становится равной
толщине этого слоя 62, которую он имел бы при полностью разви-
том пульсирующем течении. Толщина пограничного слоя для ста-
ционарного течения была определена (разд. 5.3) как расстояние
от стенки трубки до линии, на которой скорость потока составляет
99% ее значения вне слоя. Это расстояние равно 3,5 (vx/«)1/2, где
v — кинематическая вязкость жидкости, и — скорость в ядре пото-
ка, х—расстояние от входа (рис. 5.5). Эта же формула справед-
лива для величины di, определяемой в любой момент времени,
если и — мгновенная скорость в ядре потока. Толщина погранич-
ного слоя 62 для пульсирующей составляющей потока, определен-
ная таким же образом, равна 6,5 (v/gj)1/2, где ы— угловая частота
колебаний потока. Это приводит к следующему соотношению для
длины /1 начального участка для пульсирующей составляющей по-
тока:
Zi^3,4«/®. (12.28)
Так, для аорты собаки при частоте сокращений сердца около
2 Гц (так что и = 4л) и амплитуде колебаний пульсирующей со-
ставляющей скорости около 40 см-с-1 максимально возможная
мгновенная длина начального участка Zi равна примерно 10 см.
Но, поскольку на протяжении большей части цикла мгновенная
скорость и мала, длина начального участка для пульсирующей со-
ставляющей потока будет значительно меньше указанной выше
величины (и обратится в нуль в фазу обратного движения крови).
Мала будет длина Zi также и для высокочастотных составляющих
потока.
Средний по времени поток продолжает развиваться и за пре-
делами участка длиной Zi. Однако пульсирующие составляющие
не оказывают влияния на это развитие, так как вдали от входа их
пограничный слой много тоньше пограничного слоя среднего тече-
ния и, следовательно, влияет лишь на узкую область вблизи стен-
ки, где средняя скорость всегда очень мала. Поэтому для оценки
длины начального участка среднего течения можно использовать
простое соотношение Z = 0,03-Z?e-d [уравнение (5.4)], где d —
*) Сказанное здесь предполагает, что нелинейные эффекты несущественны.—
Прим. ред.
382 Часть II. Механика кровообращения
диаметр трубки, Re — среднее по времени значение числа Рей-
нольдса. Для аорты собаки, например, при диаметре 1,5 см перед-
ней скорости 20 см-с-1 среднее значение числа Рейнольдса равно
приблизительно 800 и соответствующая этим величинам длина на-
чального участка составляет около 36 см, что почти равно длине
аорты. Для аорты человека (d — 2,5 см, и ~ 20 см-с-1) длина
начального участка равна примерно 94 см, т. е. превышает длину
аорты. Эти оценки сугубо приближенные, так как при вычислениях
мы пренебрегали всеми эффектами, которые связаны с изгибами
аорты и отхождением от нее ветвей. Однако основной вывод оста-
ется в силе: для среднего по времени потока крови начальным
участком течения является почти вся аорта. С другой стороны, в
более мелких артериях как d, так и среднее значение Re меньше,
и длина начального участка составляет существенно меньшую
часть длины сосуда. Например, для бедренной артерии собаки
(г/~0,4 см, и = 10 см-с-1; см. табл. I) значение Zi равно пример-
но 1,2 см, что много меньше длины артерии.
Итак, мы выяснили, что в аорте профиль скорости в ядре по-
тока более или менее плоский, а пограничный слой тонкий. Фак-
торами, которые могут изменить такую картину течения, являются
кривизна артерий (в особенности дуги аорты) и наличие ветвей.
Рассмотрим, к каким именно последствиям приводят эти факторы.
Изгиб аорты. Стационарное течение жидкости в изогнутой труб-
ке было описано в гл. 5. Вдали от входа жидкость, движущаяся
в центре трубки и имеющая большую скорость, отбрасывается к
наружной стенке изгиба, так как инерция мешает ей быстро реа-
гировать на поперечный градиент давления, заставляющий жид-
кость двигаться вдоль изгиба. Эта оттесненная к наружной стенке
жидкость замещается имеющей меньшую скорость жидкостью, ко-
торая прилегает к стенкам и движется по направлению к внутрен-
ней стенке изгиба. Таким образом, устанавливается вторичное те-
чение, и исходно осесимметричный профиль скорости приобретает
вид, изображенный на рис. 5.16, с наибольшей скоростью течения
у наружной стенки.
Напротив, вблизи входа в изогнутую трубку первоначально
плоский профиль скорости изменяется так, что наибольшие значе-
ния скорости достигаются у внутренней стенки (рис. 5.17). Имен-
но такая ситуация наблюдается в аорте (рис. 12.40). Здесь, как
обычно на начальном участке, пристеночный пограничный слой
достаточно тонок, но из-за того, что в ядре потока скорость повы-
шается в направлении к внутренней стенке, пограничный слой на
ней тоньше и, следовательно, градиент скорости (а значит, и ско-
рость сдвига) выше, чем на наружной стенке. Однако высокое дав-
ление вблизи наружной стенки трубки (рис. 12.43) действует не
только на жидкость, движущуюся с большой скоростью в ядре
потока вдоль изгиба, но и на медленно движущуюся жидкость в
пограничном слое. Это давление заставляет ее перетекать по стен-
Гл. 12. Артерии большого круга 383
ке от наружной части изгиба к внутренней (как в полностью раз-
витом течении) так, что толщина пограничного слоя на внутренней
стенке становится больше, чем на наружной и скорость сдвига
уменьшается. Следовательно, начальное распределение скоростей
сдвига меняется на обратное. Теория предсказывает, что пере-
стройка течения происходит на расстояниях порядка одного диа-
метра трубки. Такие картины течения крови невозможно выявить
в опытах, в которых изучают профиль скоростей в аорте описан-
ным выше методом (рис. 12.40), так как пограничный слой не под-
дается исследованию. Однако влияние изменения скорости сдвига
Рис. 12.43. А. Профиль скорости в начальном участке изогнутой трубки (погра-
ничный слой во внимание не принимается). Знаком «+» обозначена область
высокого давления, знаком «—»— область низкого давления. Б. Различие дав-
лений между областями «+» и «—» приводит к возникновению в пограничном
слое вторичных течений. Их направление указано стрелками.
на стенке должно проявляться в опытах, в которых изучают пере-
нос веществ через стенку в тех случаях, когда скорость переноса
зависит от скорости сдвига или напряжения сдвига (разд. 12.9).
Сказанное выше, строго говоря, справедливо только для ста-
ционарного потока. Применительно к пульсирующему потоку оно
справедливо, независимо от величины а, лишь для очень неболь-
шой области вблизи входа. Если а велико, то вне этой области
указанные выше особенности характерны только для среднего по-
тока, но можно ожидать, что при малых а (меньших единицы)
рассмотрение будет справедливо для всего начального участка.
Это значит, что наличие пульсаций практически не повлияет на
предсказанный профиль скоростей в проксимальной части аорты.
Однако ниже по течению картина течения крови усложнится из-за
наложения на средний поток, характеризующийся высокой ско-
ростью вблизи наружной стенки, пульсирующих составляющих с
высокими скоростями вблизи внутренней стенки.
Ветвления сосудов. Рассматривая отражение волн от ветвле-
ний артерий, мы не обращали внимания на картину течения крови
в самом месте ветвления. В этом случае деталями геометрии ветв-
384 Часть II. Механика кровообращения
ления можно было пренебречь и характеризовать его только отно-
шением площадей поперечного сечения общего ствола и отходящих
от него ветвей. Но чтобы нарисовать детальную картину течения
крови, необходимо располагать гораздо более полными сведения-
ми о геометрии ветвления. Нужно знать, под каким начальным
углом ветвь отходит от порождающего ее ствола и чему равен
угол, который она в конечном счете с ним образует (эти углы
обычно неодинаковы, см. рис. 12.2). Требуются данные о кривизне
всех стенок, образующих ветвление, включая степень заостренно-
сти делителя потока. Необходимо также знать, как меняются фор-
ма и площадь сечения общего ствола при приближении к точке
ветвления. Однако подобные сведения о геометрии ветвления арте-
рий в настоящее время практически отсутствуют. Получить их
можно лишь путем кропотливых обмеров слепков артерий, изго-
товленных тщательно, при сохранении физиологических значений
трансмурального давления и исходной длины сосудов.
Но даже если бы мы всё знали о геометрии ветвлений артерий,
предсказать картины течения крови в них было бы весьма слож-
но, так как ни экспериментальные, ни теоретические исследования
стационарных или пульсирующих течений в ветвящихся трубках
почти не предпринимались. Поэтому картины течения в ветвле-
ниях артерий можно представить себе лишь в самых общих чертах.
Тип ветвления, о котором известно больше всего, — это симмет-
ричная бифуркация с одинаковым оттоком из обеих отводящих
Ветвей (рис. 12.44), поскольку для такого ветвления на моделях
из стекла или плексигласа осуществлен ряд опытов со стационар-
ным течением воды или воздуха. Однако даже при столь геомет-
рически правильном ветвлении картины течения зависят от не-
скольких параметров. В их число входят: коэффициент ветвления
(отношение суммы площадей сечения отводящих ветвей к площади
основного ствола), угол ветвления, радиусы кривизны наружной
стенки ветвления и делителя потока, число Рейнольдса потока,
попадающего в область ветвления из основного ствола, и, наконец,
профиль скорости в нем. Опыты были поставлены на модели с
коэффициентом ветвления 1,2, углом ветвления 70° и при числах
Рейнольдса от 100 до 1000. Такие параметры характерны для ветв-
лений воздухоносных путей легких. Симметричных ветвлений
крупных артерий крайне мало; для одного же из хорошо извест-
ных — бифуркации аорты человека — коэффициент ветвления ра-
вен обычно 0,8, угол ветвления — около 70°, среднее число Рей-
нольдса— около 400. Тем не менее результаты опытов на модели
позволяют получить представление об основных особенностях те-
чения в области ветвления.
Наиболее очевидная особенность — разделение потока. Следо-
вательно, если предположить, что профиль скорости в основном
стволе близок к симметричному, то, как только течение минует
делитель потока, зона наибольших скоростей окажется в непосред-
Г г /2 Артерии большого круга 385
Рис. 12.44. Схема симметричной бифуркации, поясняющая используемые в тек-
сте обозначения. Стрелки указывают направление течения
ственной близости к внутренней стенке отводящих ветвей, сразу
же за делителем потока. А поскольку скорость жидкости, сопри-
касающейся с этой стенкой, должна равняться нулю, то, начиная
от делителя потока, где скорость сдвига очень велика, формиру-
ется новый пограничный слой. Можно ожидать, что профиль про-
дольной скорости в плоскости ветвления будет сходен с изображен-
ным на рис. 12.45, А.
Другая очевидная особенность состоит в том, что каждый из
двух потоков должен огибать изогнутую стенку. Значит, в них,
так же, как и в изогнутой трубке постоянного сечения, устанавли-
ваются вторичные течения. Такие изображенные на рис. 12.46 те-
чения поддерживают наиболее высокую скорость вблизи наружной
стенки изгиба, т. е. вблизи внутренней стенки ветвления (рис. 12.44).
Кроме того, вторичные течения достаточно сильны, чтобы сместить
часть быстро движущейся жидкости к верхней и нижней стенкам.
В результате профиль скорости в плоскости, перпендикулярной
плоскости отхождения ветвей, приобретает М-образную форму,
как показано на рис. 12.45, Б. Чтобы вторичные течения стали ви-
димыми, в поток воздуха добавляли табачный дым (рис. 12.46).
По мере удаления от места ветвления картины течения меня-
ются, пока на расстоянии, сопоставимом с обычной длиной на-
чального участка [уравнение (5.4)], в каждой из отводящих
ветвей вновь не устанавливается пуазейлевское течение. Однако
начальное развитие течения сразу за ветвлением существенно от-
личается от простого осесимметричного входного потока, потому
что благодаря вторичным течениям положение максимума скорости,
смещенное к внутренней стенке трубки, поддерживается на рас-
стоянии, равном нескольким ее диаметрам
386 Часть fl Механика кровообращения
Радиальное положение
Рис. 12 45. Измеренные профили скорости в отходящих ветвях на расстоянии,
равном двум их диаметрам, за симметричной бифуркацией Число Рейнольдса
в общем стволе равно 700 (пуазейлевское течение) А Профиль скорости в пло-
скости ветвления. Б. Профиль в перпендикулярной плоскости. [Schroter, Sudlow
(1969). Resp Physiol., 7, 341—355.]
Судя по профилям скорости, изображенным на рис. 12.45, ско-
рость сдвига на внутренних стенках области ветвления остается
высокой, и не только на делителе потока, но и на отводящих вет-
вях: на некотором расстоянии вниз по потоку и на определенной
Рис. 12 46 Вид с торца трубки, отходящей от симметричной бифуркации; видны
вторичные течения (поток в общем стволе пуазейлевский) В трубках движется
воздух, картины течения получены путем введения дыма через наружную стенку
отходящей трубки. [Schroter, Sudlow (1969). Resp Physiol, 7, 341 ]
Гл. 12. Артерии большого круга 387
Рис. 12.47. Продольный разрез асимметричного ветвления Штриховая кривая —
поверхность раздела между течениями, одно из которых направляется в боковую
ветвь, а другое остается в основном стволе, сплошные кривые — линии тока.
Обратите внимание иа существование замкнутых вихрей в области отрыва по-
тока.
части их окружности. Однако на наружной стенке ветвления ско-
рость сдвига очень низкая. И действительно, во многих опытах от-
мечали наличие областей обратного течения вблизи наружной
стенки сразу за ветвлением, что указывает на отрыв потока
(разд. 5.7). Возникнет или нет такой отрыв потока после прохо-
ждения симметричного ветвления, зависит от того, существует ли
вблизи стенки обратный градиент давления, стремящийся замед-
лить движение жидкости. В рассматриваемых опытах на моделях
такой градиент существовал вследствие как увеличения площади
поперечного сечения русла после ветвления (при ветвлении аорты
этого обычно не бывает), так и значительного искривления наруж-
ной стенки ветвления. Обратный градиент давления не возникает,
если наружная стенка в модели ветвления изгибается более плавно.
Кривизна стенки в области бифуркации аорты выражена обыч-
но не слишком сильно, и потому полагают, что отрыва потока не
происходит. Однако он мог бы возникнуть, если бы в результате
заболевания произошло изменение формы бифуркации. Это при-
вело бы к появлению местного обратного течения и уменьшению
скорости сдвига на стенке. Такие явления могли бы повысить ве-
роятность развития здесь турбулентности и повлиять на перенос
веществ через стенку артерий. Обе эти возможности обсуждаются
в следующих разделах главы.
Единственный тип асимметричного ветвления, картина течения
в котором была исследована, — это ствол постоянного сечения, от
которого под прямым углом отходит ветвь (рис. 12.47). Эта модель
не самая удачная, даже если иметь в виду такие ветвления арте-
рий, для которых угол отхождения ветви от ствола хотя и оказы-
вается прямым, но лишь в конечном счете (пример тому — ответв-
ление межреберных или почечных артерий от аорты). Поэтому
388 Часть 11. Механика кровообращения
картина течения в модели ветвления такого типа чересчур упро-
щена по сравнению с действительной, но, поскольку геометрические
характеристики модели известны точно и ветвление несимметрично,
такая модель все же представляет некоторый интерес.
Опыты с добавлением красителя в равномерно движущийся
поток воды показали, что существует четко выраженная поверх-
ность, разделяющая слои жидкости, которые устремляются в бо-
ковую ветвь и продолжают течь по главному стволу (рис. 12.47).
Самая дальняя вниз по течению точка этой поверхности раздела
лежит где-то на участке стенки напротив приближающейся к ней
«струи». Этот участок соответствует делителю потока в предыду-
щем случае. Точное местоположение указанной точки зависит от
соотношения расходов жидкости в стволе и ветви. Вблизи этой кри-
тической точки скорость жидкости равна нулю даже на наружной
границе пограничного слоя. Однако пограничный слой здесь очень
тонок, и на небольшом расстоянии вниз по течению от этой точки
в обеих трубках существует область с очень высоким напряжением
сдвига на стенке. В большинстве случаев наблюдается отрыв по-
тока от наружной стенки ответвления, и сразу за ветвлением воз-
никает область обратного течения, а напряжение сдвига на наруж-
ной стенке ответвления оказывается низким. В боковой ветви уста-
навливаются сильные вторичные течения. Иногда в стволе также
возникает отрыв потока — в точке, примерно противоположной
входу в ветвь. Определяется это тем, что «отсасывание» жидкости
в ветвь создает в стволе область низкого давления, ниже которой
возникает обратный градиент давления. На рис. 12.47 нанесены
также характерные линии тока, по которым в плоскости симмет-
рии ветвления перемещается краситель.
Недавно проведены количественные исследования, в ходе кото-
рых на полых слепках аорты собаки было измерено напряжение
сдвига на различных участках стенки. Эти измерения подтвердили
приведенную выше качественную картину течения. Правда, по-
скольку ветви отходят от аорты под малыми углами, а затем рез-
ко изгибаются, области отрыва потока в ветвях могут смещаться
вниз по течению или отсутствовать вовсе. Кроме того, эти опыты
ясно показали, что даже совсем небольшие изменения геометриче-
ских параметров сосуда в области ветвления способны сильно
влиять на местную картину течения крови. В одной из артерий
вершина делителя потока (рис. 12.47) чуть-чуть выпятилась внутрь
просвета (менее чем на 1 мм), но этого оказалось достаточно, что-
бы на стенке главного ствола сразу за делителем потока сформи-
ровалась еще одна небольшая область отрыва.
Итак, ясно, что картины течения крови в ветвящихся артериях
очень сложны, так как они связаны с развитием вторичных тече-
ний и возникновением смежных областей с высоким (там, где по-
граничный слой тонок) и низким (там, где существуют области
отрыва потока) напряжением сдвига на стенке. Сложный характер
Гл. 12. Артерии большого круга 389
распределения сдвиговых напряжений в стенке артерий может
быть важным фактором, определяющим места появления ранних
признаков атероматоза, если только это заболевание хотя бы ка-
ким-то образом связано с напряжением сдвига на стенке сосудов
(разд. 12.9).
Устойчивость и турбулентность. Для течений очень вязких
жидкостей или течений с низкими скоростями (т. е. для течений
с малыми числами Рейнольдса) характерны упорядоченность дви-
жения и наличие четко выраженных линий тока. Перенос массы,
импульса и энергии поперек линий тока при этом осуществляется
только за счет молекулярного движения (т. е. диффузии). Такое
течение называется ламинарным-, один из его частных случаев —
пуазейлевское течение. Следует, однако, помнить, что ламинарное
течение может быть как стационарным, так и нестационарным и
иметь весьма сложную структуру из-за наличия вихрей или вто-
ричных течений. Для ламинарного потока характерно затухание
возмущений в нем под действием вязких сил. Признак турбулент-
ного течения — наличие случайных по амплитуде, частоте и на-
правлению незатухающих возмущений (гл. 5).
По мере увеличения числа Рейнольдса затухание малых воз-
мущений в ламинарном потоке уменьшается. В конце концов до-
стигается критическое число Рейнольдса, при превышении кото-
рого течение становится неустойчивым, и возмущения (по крайней
мере некоторые) усиливаются. Когда число Re оказывается боль-
ше критического, а времени для усиления возмущений достаточно,
возникает турбулентность. Обычно считают, что критическое число
Рейнольдса (его находят по средней скорости жидкости и диамет-
ру трубки) для полностью развитого течения в трубке равно при-
мерно 2300, хотя оно может быть и существенно выше, если поза-
ботиться о том, чтобы не внести в поток возмущений. Когда же,
однако, скорость течения, уже ставшего турбулентным, умень-
шают, уменьшая тем самым Re, поток при всех обстоятельствах
остается турбулентным, пока Re не окажется меньше 2300. Воз-
мущения, приводящие к турбулентности, могут возникать в силу
многих причин, и интенсивность их существенно влияет на значе-
ние критического числа Рейнольдса. Обычные причины возмуще-
ний— существование в потоке, выше по течению (например, в ре-
зервуаре), вихрей или наличие шероховатостей на стенке трубки.
Поскольку те образующиеся в потоке вихри, которые и пред-
ставляют собой турбулентность1), по своим размерам и по скоро-
сти случайны, они выявляются (с помощью, например, датчика
’) Понятие «вихря» в гидродинамике используется ие только в строгом его
смысле (который не обсуждается в этой книге), но и как обозначение для раз-
личных течений с замкнутыми траекториями — например, в зонах отрыва. Ото-
ждествление турбулентности с рождением и затуханием в потоке множества
беспорядочно вращающихся элементов жидкости есть не более чем одно из мо-
дельных представлений.— Прим. ред.
390 Часть II. Механика кровообращения
скорости, введенного в какую-либо точку потока) в виде хаотич-
ных флуктуаций скорости, наложенных на кривую средней скоро-
сти. Соответствующий пример для потока в трубке приведен на
рис. 12.48. Строго говоря, называть поток турбулентным можно,
только если показано, что случайные флуктуации скорости трех-
мерны. Впрочем, процесс этот на практике столь хорошо изучен
для многих случаев течения жидкостей, что флуктуации скорости
лишь в одном направлении, когда они обладают соответствующи-
ми характеристиками, уже достаточны для распознания турбу-
лентности. Кривая на рис. 12.48 не только наглядно показывает
Рис. 12.48. Турбулентное течение. Запись сделана с помощью датчика пленочного
термоанемометра, помещенного в трубку, скорость течения жидкости в которой
постепенно увеличивали вплоть до достижения турбулентного режима течения,
а затем повышение скорости прекращали. Максимальное значение числа Рей-
нольдса равно 9500. Штриховой прямой указано значение скорости, соответствую-
щее Re = 2300, которое обычно считают критическим. [Nerem, Seed (1972).
An in vivo study of aortic flow disturbances. Cardiovasc. Res., 6, 1—14.]
высокочастотные флуктуации скорости, но и позволяет проследить
за некоторыми особенностями появления и затухания турбулент-
ности. Во-первых, турбулентность не развивается, пока число Рей-
нольдса не достигает значения много большего, чем обычное кри-
тическое значение 2300. В какой-то степени это может опреде-
ляться тем, что движение жидкости с ускорением устойчивее
равномерного движения, но, кроме того, и отсутствием возмуще-
ний выше по течению благодаря соответствующей конструкции
установки. Во-вторых, турбулентность сохраняется при уменьше-
нии скорости жидкости, даже когда число Рейнольдса становится
ниже 2300. Отчасти это обусловлено тем, что замедляющийся по-
ток в принципе менее устойчив, чем равномерный, а отчасти —
конечным временем, необходимым для затухания вихрей.
Так как вихри в турбулентном потоке весьма разнообразны по
величине и переносятся течением, сигналы чувствительного к ско-
рости датчийй, фиксированного в потоке, выявляют наличие со-
ставляющих скорости, колеблющихся в широком диапазоне частот,
Т. е. некое распределение скоростей по частоте. Его можно харак-
Гл. 12. Артерии большого круга 391
теризовать различными способами (например, с помощью анализа
Фурье, разд. 12.2) и изобразить в виде кривой зависимости скоро-
сти (или энергии) от частоты. Обычно такая спектральная кривая
располагается по обе стороны от некоторой частоты f0, причем при
частотах, много больших или много меньших этой частоты, ско-
рость мала. Величина fo, называемая «центральной частотой», оп-
ределяется размерами наибольших вихрей, а эти размеры зависят
от геометрии потока. Например, для течения в трубке диаметр
наибольших вихрей сопоставим с диаметром трубки d. Поэтому,
если скорость есть и, то величина f0 равна приблизительно u/d.
В какой именно точке спектральной кривой достигается макси-
мальная амплитуда скорости или энергии, определяется рядом
взаимосвязанных и нередко трудно поддающихся анализу особен-
ностей течения, включая геометрию русла как в данном месте, так
и выше по течению, а также вязкостью жидкости.
Возвращаясь к течению крови в артериях, заметим, что в боль-
шинстве инженерных задач переход к турбулентности и саму тур-
булентность изучали, к сожалению, только в условиях стационар-
ного (исходно, до перехода) потока, и лишь немногие исследования
посвящены устойчивости пульсирующего потока в трубках. Однако
и эти исследования обычно касались таких течений, в которых
пульсирующие составляющие скорости были много меньше сред-
ней скорости, и поэтому характер течения определялся свойствами
именно среднего течения. Конечно, это мало похоже на течение
крови в крупных артериях, где пиковая скорость может в четыре-
пять раз превышать среднюю.
Поэтому до недавнего времени рассуждения о возникновении
турбулентности в артериях носили в основном гипотетический ха-
рактер и опирались на сопоставления с данными о непульсирую-
щих течениях. Такие рассуждения приводили к представлению о
том, что во время систолы в крупных артериях может появляться
турбулентность. Немногочисленные попытки изучить прижизненные
картины течения крови в артериях животных — методом киносъем-
ки движения красителей или рентгеноконтрастных веществ, вводи-
мых в кровь, — как будто подтверждали это представление. Одна-
ко результаты, получаемые подобным методом визуализации пуль-
сирующего потока, с трудом поддаются анализу: наблюдения
кратковременны, они производятся лишь в каком-то одном месте
артерии, и к тому же все возмущения (например, ламинарные
вихри или вторичные течения) могут смещать частицы красителя
и восприниматься как турбулентность. Другой классический метод
обнаружения турбулентности, а именно доказательство нарушения
пуазейлевского соотношения давление — расход, к пульсирующему
течению, разумеется, неприменим.
Как уже говорилось, условия, определяющие переход непуль-
сирующего течения к турбулентному режиму, твердо установлены.
В этом случае определяющим параметром является число Рей-
3&2 Часть tl. Механика кровообращения.
нольдса, критическое значение которого порядка 2000 (оно отча-
сти зависит от степени возмущения потока выше по течению), а
сама турбулентность характеризуется частотным спектром, свой-
ства которого четко определены. Когда же поток пульсирует (как
в артериях), ситуация по ряду причин усложняется. Во-первых,
для столь сильно пульсирующего потока среднее значение Re едва
ли является адекватной характеристикой, и потому неясно, каким
образом следует выбирать подходящее его значение. Вообще го-
воря, более уместным параметром считается пиковое значение чис-
ла Рейнольдса, ибо имеющиеся данные позволяют предполагать,
что возмущения потока в аорте чаще возникают при пиковой или
близкой к ней скорости течения крови во время систолы.
Вряд ли, однако, возникновение турбулентности определяется
единственным параметром — пиковым значением Re. Для того
чтобы возмущения потока при сверхкритических значениях Re
превратились в турбулентность, необходимо, как уже отмечалось,
некоторое конечное время. Значит, по-видимому, важно и то, как
именно изменяется во времени скорость крови в аорте. При высо-
кой частоте сокращений сердца длительность одного пульсового
цикла может оказаться недостаточной для перехода к турбулент-
ному режиму даже при числах Рейнольдса выше критического.
Чтобы обобщить только что сказанное о факторе времени, следует
ввести соответствующий безразмерный параметр; подходящей для
этого величиной является параметр Уомерсли а (разд. 12.3).
Можно ожидать, что при низких значениях а поток будет вести
себя так, как будто он непульсирующий, причем критическое зна-
чение числа Рейнольдса будет порядка 2000. По мере увеличения
а тот промежуток времени — в пределах каждого пульсового цик-
ла,— в течение которого возможно усиление возмущений потока,
становится меньше требующегося для этого времени. Поэтому есть
основания полагать, что с повышением частоты сердцебиений кри-
тическое значение числа Рейнольдса будет возрастать1).
В последнее время появилась возможность исследовать вопрос
о существовании турбулентности в артериях с помощью прямого
метода — пленочной термоанемометрии. Соответствующие измере-
ния убеждают в том, что течение крови в грудном отделе аорты
может быть турбулентным. Из записи скорости, приведенной на
рис. 12.49, видно, что в конце систолы, когда скорость крови умень-
) В добавление к сказанному отметим, что пульсирующее течение в длин-
ной трубе с гладкими стенками характеризуется следующими размерными вели-
чинами: средней по сечеиию и во времени скоростью V, плотностью жидкости
р и ее вязкостью р., диаметром трубы R, а также частотой пульсаций <о и сред-
ней по сечению амплитудой пульсаций скорости 6V. Из этих величин можно
составить три независимые безразмерные комбинации- обычное или пиковое чис-
ло Рейнольдса, параметр Уомерсли а и относительную амплитуду б У/У. Кри-
тическое число Рейнольдса зависит, таким образом, не только от а, но и от
8V/V. Влияние последнего параметра, по-вндимому, велико.—Прим. ред.
Гл. 12. Артерии большого круга 393
шается, на заднем фронте пульсовой волны кровотока появляются
высокочастотные колебания. Анализ показывает, что частоты этих
колебаний лежат в широком диапазоне — от 500 Гц до менее чем
25 Гц. Таким образом, они сливаются с высокочастотными состав-
ляющими пульсовой волны скорости. Но в отличие от последних
амплитуды наблюдаемых высокочастотных составляющих (по
всему их спектру) случайным образом меняются от удара к удару
сердца, и, следовательно, имеет место истинная турбулентность.
Опыты, в которых при помощи соответствующих препаратов или
Рис. 12.49. Волна скорости
крови в нисходящей аорте со-
баки. Видно, что в фазу за-
медленного изгнания крови су-
ществует турбулентность. [Seed,
Wood (1971). Velocity patterns
in the aorta. Cardiovasc. Res.,
5, 319—333.]
100
20
<——------—i----------1
0 0 0.5 l.o 1.5
Время c
путем раздражения нервов меняли скорость и частоту сокраще-
ний сердца — и тем самым характеристики пульсовых волн крово-
тока, — показали, что переход от ламинарного течения к турбу-
лентному зависит и от числа Рейнольдса, и от значения парамет-
ра а (т. е. фактически от частоты сокращений сердца), как и
утверждалось выше. Результаты этих опытов представлены на
рис. 12.50.
Однако, как следует из рис. 12.50, в большинстве опытов поток
крови в аорте не был турбулентным, и прямые данные о том, явля-
ется ли у здоровых бодрствующих собак поток крови в аорте тур-
булентным, отсутствуют.
Тем не менее есть достаточно веские доводы в пользу того, что
турбулентность имеет место. Все опыты, результаты которых пред-
ставлены на рис. 12.50, произведены на наркотизированных живот-
ных, а наркоз сильно изменяет как частоту сокращений сердца,
так и пиковые значения скорости крови, и притом таким образом,
что точки на рис. 12.50 должны смещаться вниз и вправо, т. е. в
область устойчивых течений. Анализ приводимых в литературе
характеристик потока крови в аорте здоровых бодрствующих жи-
вотных (рис. 12.51, А) дает серьезные основания полагать, что тур-
булентность существует. К такому же выводу приводит и анализ
данных для человека; кроме того, в этом случае есть и прямые
данные, полученные с помощью метода пленочной термоаиемо-
394 Часть 11. Механика кровообращения
а
Рис. 12.50. Влияние параметра а и пикового значения числа Рейнольдса иа устой-
чивость течения в нисходящей аорте наркотизированных собак. Прямыми соеди-
нены точки, полученные на одном животном при различных условиях. Светлые
кружки — ламинарное течение; наполовину зачерненные кружки — переходный
режим (кратковременные возмущения существуют лишь при максимальных значе-
ниях скорости); темные кружки — турбулентное течение. Оттенена область устой-
чивого ламинарного течения. [Nerem, Seed (1972). An in vivo study of aortic flow
disturbances, Cardiovasc. Res., 6, 1—14.]
метрик (рис. 12.51,Б). Таким образом, можно считать, что при
физиологических условиях в состоянии покоя поток крови в про-
ксимальных отделах аорты человека, а возможно, и собаки яв-
ляется турбулентным. Что именно порождает турбулентность —
наличие вихрей выше по течению или местные возмущения, свя-
занные с геометрией стенки сосуда, — пока не ясно. Данные же
о том, как далеко вниз по потоку от грудного отдела аорты может
простираться область турбулентности, практически отсутствуют.
С точки зрения гидромеханики нет оснований считать турбу-
лентность нежелательным явлением. Хотя обусловленное ею уве-
личение диссипации энергии должно приводить к увеличению па-
дения давления на крупных артериях, последнее, как мы уже знаем,
очень мало по сравнению с общим падением давления в сосудах
большого круга. Вместе с тем турбулентность причастна к ряду
важных физиологических явлений. Одно из них — генерация зву-
ковых колебаний. Причиной шумов в сердце нередко считают
именно турбулентность потока крови и потому полагают, что у здо-
ровых людей турбулентность обычно отсутствует, тем более что
представление о бесшумности ламинарного потока почти общепри-
нято. Однако ни то ни другое положение не является бесспорным.
Ламинарное течение сопровождается явлениями (например, схо-
дом вихрей за обтекаемым телом, разд. 5.9), которые могут поро-
Гл. 12. Артерии большого круга 395
Рис. 12.51. А. Значения параметра а и пикового значения числа Рейнольдса,
обычно реализующиеся при течении крови в восходящей аорте здоровых бодр-
ствующих собак. Оттенены области, отвечающие такой же области на рис. 12.50.
Обратите внимание на то, что у ненаркотизированных животных числа Рейнольд-
са выше, чем у наркотизированных, и все точки попадают в ту область, для ко-
торой можно ожидать турбулентности. [Noble, Trenchard, Guz (1966). Circula-
tion Res., 19, 139—147.] Б. Собранные из многих источников сопоставимые дан-
ные для здоровых, бодрствующих людей. Здесь также можно предсказать наличие
турбулентного течения почти во всех случаях. Кружками (см. подпись к
рис. 12.50) отмечены результаты, полученные с помощью пленочной термоанемо-
метрии. Они указывают на существование турбулентности.
а
Б
ждать звуковые волны. Наряду с этим нет никаких оснований
полагать a priori, что флуктуации давления при турбулентном по-
токе всегда будут выявляться при прослушивании со стороны
грудной клетки. Вполне вероятно, что для таких клебаний суще-
ствует «порог» слышимости.
Турбулентность может существенно влиять на силы, с которы-
ми кровь действует на стенку артерии, а эти силы способны изме-
нять как механические свойства стенки, так и условия обмена
веществами между стенкой и кровью (разд. 12.9). Влияние турбу-
лентности на такие силы осуществляется как через высокочастот-
ные флуктуации давления и напряжения сдвига, которые сопро-
вождают хаотические изменения скорости течения крови, так и
через изменение среднего профиля скорости, а следовательно, и
среднего напряжения сдвига на стенке. Обратимся сначала к сред-
ним характеристикам турбулентного течения. Если средний рас-
ход жидкости, текущей в прямой трубке, не пульсирует, то про-
филь скорости полностью развитого турбулентного течения отли-
чается от профиля скорости пуазейлевского течения тем, что в
ядре потока он более плоский, а скорость сдвига на стенке больше
(гл. 5). Однако толщина пограничного слоя, формирующегося на
начальном участке трубки, при турбулентном потоке больше, чем
396 Часть II. Механика кровообращения
при ламинарном, что связано с наличием в первом случае хаоти
ческого вихревого движения. Заставляя элементы жидкости пере
мещаться в поперечном направлении, оно приводит к значительно
более эффективному перемешиванию жидкости в этом направле-
нии, чем при ламинарном течении, с участием одних только вяз
ких сил. Толщину пограничного слоя б при турбулентном течении
можно приближенно представить в виде
»=*(4Г-
Здесь в сравнении с уравнением (5.3) кинематическая вязкость v
заменена эффективной «вихревой вязкостью» v*, которая учиты
вает турбулентное перемешивание. Одним из следствий такой за
мены является уменьшение при турбулентном режиме длины на-
чального участка (разд. 5.5).
Если сам по себе исследуемый средний1) поток пульсирует с
угловой частотой го, то толщина колеблющегося пограничного слоя
оказывается пропорциональной д/у7и> так что и в этом случае
его толщина много больше, чем при ламинарном потоке. Таким
образом, при переходе от ламинарного потока, колеблющийся по-
граничный слой в котором сравнительно тонок (ос ^.4), к турбу-
лентному наблюдается увеличение толщины этого слоя. Этот вы
вод относится ко всей аорте.
Турбулентность связана с еще одним важным в физиологиче-
ском отношении явлением — действием вибрации на стенку арте
рий. Клиницистам давно известно, что участок артерии сразу же
за препятствием, ограничивающим поток крови, например за су
жением (стенозом) аортального клапана или за атероматозной
бляшкой, нередко оказывается расширенным. Такое постстеноти
ческое расширение участка артерии развивается вне зависимости
от природы препятствия — важно только, чтобы последнее вызы-
вало турбулентность.
Исследований, в которых действие вибрации на стенку артерий
изучали бы на протяжении достаточно длительного времени, не
предпринималось. Но относительно непродолжительные — до не-
скольких месяцев — опыты показали, что расширение артерии сра-
*) Следует четко определить, что означает слово «средний» в отношении тур-
булентного потока Для того чтобы дать определение средней скорости, предполо-
жим, что мы многократно измеряем скорость, которую приобретает жидкость
в одном и том же месте данного потока через одинаковые промежутки времени
от начала течения Всякий раз из-за случайных турбулентных флуктуаций зна-
чения скорости окажутся различными. Усреднив результаты таких измерений,
мы получим вполне определенную величину, называемую «средней по ансамблю».
Именно эту величину называют средней скоростью жидкости в данной точке
в данный момент времени Такая процедура усреднения устраняет все случайные
флуктуации и позволяет получить среднее значение, которое в каждой точке
может, конечно, меняться во времени, как это происходит, например, при про-
хождении волны.
Гл. 12. Артерии большого круга 397
зу за ее местным сужением (где артерию частично перетягивали)
воспроизводится неизменно, если в участке за сужением возникают
прослушиваемые шумы и воспринимаемые пальцем исследователя
вибрации. Участки артерии за сужением оказались более растя-
жимыми, чем нормальные, хотя микроскопически никаких струк-
турных нарушений стенки артерии не выявилось. Кроме того, было
установлено, что если удаленные из организма участки артерий на
протяжении нескольких часов подвергать действию низкоампли-
тудных вибраций с частотой 30—400 Гц, то они становятся более
растяжимыми. Этот диапазон частот почти полностью соответ-
ствует частотному диапазону спектра флуктуаций скорости при
турбулентном течении крови. Таким образом, есть веские основа-
ния считать, что турбулентность причастна к возникновению пост-
стенотического расширения артерий. В этой связи интересно поду-
мать над тем, почему с возрастом увеличивается диаметр аорты:
не может ли это быть результатом длительного воздействия низко-
амплитудных вибраций на аорту, а не присущих ей дегенератив-
ных изменений? Заметим, наконец, что искусственно созданная в
эксперименте турбулентность способна изменять проницаемость
стенки артерий для крупных молекул и приводить к повреждению
эндотелия (разд. 12.9).
12.9. Смешивание и перенос веществ в артериях
Основной функцией сердечно-сосудистой системы является пе-
ренос различных веществ и тепла к тканям и от них. Обмен ве-
ществами между кровью и тканями происходит главным образом
на уровне капилляров и соответственно обсуждается в следующей
главе. Однако стенки артерий (и вен) также проницаемы, и через
них тоже осуществляется перенос веществ. Этот процесс обеспе-
чивает питание стенки артерий, но может играть роль и в возник-
новении некоторых поражений артерий, таких, как атеросклероз.
Недавно было показано также, что силы, действующие на стенку
артерии со стороны текущей крови, могут быть важным самостоя-
тельным фактором, определяющим проницаемость стенки для
крупных молекул и, следовательно, существенно влияющим на по-
ражения артерий. Некоторые свидетельствующие об этом данные
рассмотрены дальше в этом разделе.
Чтобы вещество, переносимое кровью к тканям по артериям и от
тканей по венам, было равномерно распределено по капиллярам
любого органа, оно должно быть хорошо перемешано в крупных
сосудах, обеспечивающих приток крови к этому органу и ее отток.
Если бы кровь, оттекающая от какого-либо органа, продолжала
свое движение по сердечно-сосудистой системе в виде отдельной
струи, не смешиваясь с остальной кровью, эта система не могла бы
удовлетворительно функционировать. Заметим далее, что широко
используемый метод измерения минутного объема сердца и крово-
S98 Часть II Механика кровообращения
снабжения отдельных областей тела — метод разведения индика-
тора— основан на предположении о том, что «меченный» каким-
либо индикатором объем крови хорошо смешивается с остальной
кровью. Следовательно, вопрос о степени перемешивания крови
в сердце и в крупных сосудах очень важен.
Смешивание в сердце и в крупных кровеносных сосудах.
Принцип метода разведения индикатора для измерения минутного
объема сердца предложил в прошлом веке физиолог А. Фик (он
же сформулировал законы диффузии, рассмотренные в гл. 9). Фик
указал, что если известны две величины, а именно разность кон-
центрации кислорода в крови, оттекающей из легких и притекаю-
щей к ним, и общее потребление организмом кислорода в единицу
времени, то количество крови, протекающей в единицу времени
(1 мин) через легкие (т. е. минутный объем сердца) есть про-
сто отношение второй величины к первой. Значительно позд-
нее, когда были разработаны приемы взятия проб крови и спо-
собы их анализа, предложенный Фиком принцип был положен в
основу нормативного метода измерения минутного объема сердца
как у человека, так и у животных. Были созданы основанные на
этом принципе методы измерения расхода крови при помощи вве-
дения в нее — непрерывно либо в виде малых объемов (порций) —
красителей (метод разведения красителя), радиоактивных изото-
пов или же термической метки, т. е. охлажденной порции крови
пли физиологического раствора. Одни из этих методов применяют
для измерения минутного объема сердца, другие — для измерения
расхода крови в отдельных органах или областях тела. Для этого
берут пробы крови, притекающей к органу и оттекающей от него,
и измеряют в них концентрацию индикатора. Очевидно, что такой
метод может дать надежные результаты только в том случае, ког-
да концентрации индикатора, обнаруженные в пробах крови, пред-
ставительны фактически для всего объема крови, который проте-
кает через орган. Если, например, в пробе артериальной крови,
используемой для расчета минутного объема по принципу Фика,
содержится меньше кислорода, чем в остальном объеме артериаль-
ной крови, то вычисленная величина окажется завышенной.
Чтобы выяснить, в каких областях системы кровообращения
происходит смешивание и какова его полнота, использовали ряд
методов. Непосредственно наблюдали за течением в просвечивае-
мых сосудах или в прозрачных моделях сосудов, визуализируя те-
чение введением красителей; изучали поведение введенного в сер-
дечно-сосудистую систему рентгеноконтрастного вещества; наконец,
вводили в эту систему индикаторные вещества и измеряли их кон-
центрацию в пробах крови, взятых из разных сосудов Вывод из
всех этих исследований один: в момент, когда индикатор, прошед-
ший через сердце, выходит из него, он уже полностью смешан с
кровью. Смешивание же только в одной из камер сердца или где-
либо в сосудистой системе может быть неполным.
Гл 12. Артерии большого круга 399
Смешивание в сердце происходит, по-видимому, главным обра-
зом во время прохождения крови через желудочек. Если быстро
ввести индикатор в какую-нибудь вену, а затем взять пробы крови
из многих участков правого предсердия, в котором сливается ве-
нозная кровь от различных частей тела, можно обнаружить, что
концентрации индикатора в пробах различны. Однако в пробах
крови из правого желудочка или из легочного ствола концентра-
ция индикатора практически всегда одинакова. Это заключение
справедливо также для левых камер сердца.
Смешивание в сердце имеет конвективную природу, т. е. явля-
ется следствием такого движения крови в желудочке, которое пере-
мешивает ее и тем самым способствует выравниванию состава кро-
ви. Поэтому этот процесс протекает значительно быстрее, чем если
бы он определялся одной только диффузией. Правда, диффузия
все-таки имеет существенное значение, поскольку она окончатель-
но выравнивает концентрации растворенного вещества в неболь-
ших объемах крови, в одних из которых его много, а в других
мало, после сближения этих объемов в результате перемеши-
вания. Последнее обеспечивает смешивание растворяемого веще-
ства и растворителя гораздо более эффективно, чем диффузия:
пример тому — ежедневные «опыты» с чашкой чая.
Как показано в гл. 9, время, за которое происходит диффузия
на расстояние Дх, пропорционально to, где
. (Дх)2
tD ~~ 2D ’
a D — коэффициент диффузии вещества, диффундирующего в дан-
ном растворителе [уравнение (9.3)]. Коэффициент диффузии, ска-
жем, кислорода в крови равен примерно 2-Ю-9 м2-с-1, и, следова-
тельно, время, необходимое для диффузии молекул кислорода на
расстояние 1 см (а такое расстояние намного меньше того, на ко-
тором должна была бы осуществляться диффузия кислорода в же-
лудочке в отсутствие конвекции), составляет 2-104 с. Это время
слишком велико для того, чтобы за один удар сердца успело про-
изойти полное смешивание. Коэффициент диффузии тепла в крови
больше (порядка 10-7 м2-с~1), однако и в этом случае время диф-
фузии (выравнивания температуры) очень велико (500 с). Время,
необходимое для диффузионного смешивания кислорода, могло бы
сократиться до 1 с только при условии, что за счет конвекции рас-
стояние, на которое должен диффундировать кислород, умень-
шено до 0,06 см.
В желудочках происходят перемешивающие движения двух ти-
пов. Первое и, вероятно, наиболее важное — это вихревое движе-
ние, возникающее в то время, когда кровь проходит через пред-
сердно-желудочковые клапаны. Такие вихри наблюдали как у айй-
вотных, так и на моделях левого желудочка (гл. 11) Они приво-
дят к цнтенсивному перемешиванию крови, хотя за время одного
400' Часть И. Механика кровообращения
сокращения сердца не обеспечивают полного смешивания индика-
тора, введенного непосредственно в полость желудочка. Основной
вихрь — кольцевой, формирующийся вокруг створок клапана. На
него накладываются более слабые возмущения потока крови, обус-
ловленные наличием сухожильных струн клапана и нерегулярно-
стями внутренней поверхности стенки желудочка. Подобные вихри
существуют вне зависимости от того, является ли течение крови
ламинарным или турбулентным. Второй возможный фактор сме-
шивания— турбулентность сама по себе. Прямых эксперименталь-
ных данных о наличии турбулентности течения крови в сердце нет.
Тем не менее, поскольку число Рейнольдса при прохождении крови
через митральный клапан может достигать 8000, а кровь втекает
в желудочек в виде струи с высокой скоростью сдвига на грани-
цах, в сердце почти несомненно возникает турбулентное течение,
по крайней мере на протяжении какой-то части периода сокраще-
ния (разд. 5.8).
Течение крови в восходящей аорте, судя по рентгенографиче-
ским данным и по результатам опытов с введением охлажденных
индикаторных жидкостей в этот отдел аорты, тоже обеспечивает
весьма эффективное смешивание крови. И в этом случае причиной
перемешивания могут быть вихри (вовсе не обязательно турбу-
лентные), которые образуются при открытии аортального клапана
или конвективно переносятся в аорту из желудочка. Кроме того,
к перемешиванию приводит турбулентность как таковая, которая,
по всей видимости, существует в аорте (см. выше).
Что касается грудной и брюшной аорты, а также более мелких
артерий и вен, то в них, согласно некоторым данным, происходит
лишь частичное смешивание. Так, в пробах крови, взятых из раз-
ных мест одного из таких сосудов, могут оказаться неодинаковые
концентрации индикатора, который был введен в этот сосуд немно-
го выше по течению. Далее, струйки непрерывно вводимого в со-
суд рентгеноконтрастного вещества так и остаются разделенными,
не смешиваясь с кровью. Характер течения в рассматриваемых
сосудах несколько иной, чем в восходящей аорте, так как числа
Рейнольдса в них ниже и турбулентность не возникает. Поэтому
смешивание в этих сосудах может осуществляться только благо-
даря вторичным течениям, возникающим из-за изменений геомет-
рии системы (изгиб или ветвление), а возможно, и вследствие ко-
лебаний крови (как в поперечном, так и в продольном направле-
ниях), сопряженных с распространением пульсовых волн.
Насколько известно, смешивание в мелких сосудах собаки не
изучали. В мелких артериях кролика (d ~ 0,1 см) наблюдали не-
смешивающееся течение струек красителя, однако проследить за
течением таких струек в аорте кролика (d ~ 0,3 см) не удавалось,
так как они быстро разрушались. Данные о смешивании в арте-
риях человека противоречивы. Существует одно очень давнее на-
блюдение — краситель, введенный в дугу аорты, появляется в каж-
Гл. 12. Артерии большого круга 401
дой из двух подвздошных артерий в неодинаковых концентрациях.
С другой стороны, краситель, введенный непосредственно в плече-
вую артерию, почти полностью смешивается, если судить об этом
по анализу концентрации красителя в пробах крови, взятых ниже
по течению одновременно из лучевой и локтевой артерий. В мел-
ких венах кролика, где течение крови практически не пульсирует,
удается наблюдать картины движения отдельных струек красите-
ля, указывающие на наличие ламинарного режима. Правда, чтобы
наблюдать такие картины, необходимо выполнение ряда правил,
в частности раствор красителя следует вводить вдали от места
наблюдения, чтобы не вызвать возмущений течения (присутствие
в просвете мелкого сосуда иглы или зонда может оказаться мощ-
ным источником местных завихрений и перемешивания). Хотя те-
чение крови в более крупных венах (в каудальной полой вене
кролика, d ~ 0,34 см) нестационарно, здесь все же можно разли-
чить движение отдельных струек красителя. Эти струйки хорошо
видны на рентгенограммах вен голени у человека (диаметр этих
вен ~0,5 см). Однако анализ таких данных требует известной
осторожности, так как заметное различие плотностей крови и рас-
твора рентгеноконтрастного вещества может затруднять смеши-
вание.
Существует еще один тип влияния процессов течения на диф-
фузию, который сказывается на результатах, получаемых методом
разведения индикатора, а именно продольное «вытягивание», или
дисперсия, порции индикатора по мере ее движения в сердечно-
сосудистой системе. В какой-то степени дисперсия определяется
тем, что разные части исходной порции «меченой» крови проходят
в сосудистой системе изучаемого органа разные расстояния. Одна-
ко значительная дисперсия индикатора наблюдается даже в про-
цессе прохождения последнего по одному сосуду (например, ка-
кой-нибудь артерии). Она гораздо больше дисперсии, которая
должна была бы наблюдаться, если бы имелась одна лишь про-
дольная диффузия индикатора, характеризуемая коэффициентом
его молекулярной диффузии D, который определяется уравнением
(9.3).
Механизм такой дисперсии индикатора проще всего понять, рас-
сматривая полностью развитое пуазейлевское течение в длинной
прямой трубке (рис. 12.52). Предположим, что в некоторый мо-
мент времени существует четкая плоская поверхность раздела
между «немеченой» кровью А и «меченой» кровью В. С течением
времени эта поверхность вытягивается и приобретает форму пара-
болоида, поскольку скорость движения жидкости на вершине его
(т. е. на оси трубки) вдвое превышает среднюю скорость, тогда
как жидкость, соприкасающаяся со стенкой трубки, из-за прили-
пания остается неподвижной. Если бы эти жидкости были неспо-
собны диффундировать друг в друга, то со временем поверхность
ид раздела должна была бы неуклонно вытягиваться.
402 Часть II. Механика кровообращения
Начальный
момент времени
Последующие
моменты времени
Рис. 12.52. Дисперсия жидкости В
при пуазейлевском течении в
длинной трубке, которая первона-
чально была заполнена жидкостью
А.
Можно рассчитать концентрацию жидкости В (среднюю по се-
чению) вдоль трубки от основания (в точке х0) ДО вершины пара-
болоида «меченой» крови (в точке х = L); результаты этого рас-
чета изображены штриховой кривой на рис. 12.53. При наличии
диффузии получится иная кривая, так как жидкость В будет диф-
фундировать в жидкость А как в направлении оси трубки, так и
в радиальном направлении. Диффузия вдоль оси трубки приводит
лишь к незначительному размыванию поверхности раздела; ра-
диальная же диффузия весьма существенна, поскольку она пере-
носит жидкость В из быстро движущегося ядра потока в область
более низких скоростей, где ее перенос вниз по течению замед-
ляется. Тем самым уменьшается продольное вытягивание поверх-
ности раздела (дисперсия), и концентрация жидкости. В изменя-
ется вдоль оси х по кривой, близкой к приведенной на рис. 12.53
сплошной кривой. На рисунке изображен также профиль концен-
трации для плоской поверхности раздела. Такие же процессы про-
исходят и на задней границе раздела, если объем «меченой» жид-
кости имеет форму таблетки, и в таком случае этот объем распре-
деляется в трубке симметрично относительно центра таблетки
(симметричность слегка нарушается продольной диффузией).
На рис. 12.53 показано распределение концентрации «меченой»
жидкости вдоль трубки лишь в какой-то определенный момент
времени. Между тем метод разведения индикатора предусматри-
вает взятие проб крови из определенного места и измерение кон-
центраций, изменяющихся во времени. Поскольку предполагается,
что время, за которое индикатор достигает точек измерения, до-
статочно для полного смешивания его с кровью по всему попереч-
ному сечению сосуда, важно, чтобы эти точки измерения находи-
лись достаточно далеко вниз по течению от места введения ин-
Гл. 12. Артерии большого круга 403
дикатора. Когда передний фронт порции крови с индикатором
достигает места измерения, концентрация индикатора здесь возра-
стает, а затем, когда это место проходит задний фронт порции,
уменьшается. Однако независимо от того, проходят ли части пор-
ции индикатора одно и то же или разное расстояние, кривая кон-
центрация — время оказывается несимметричной относительно мо-
мента времени, в который центральная часть порции индикатора
проходит место измерения. Несимметричность кривой определяется
тем, что процесс смешивания индикатора с кровью для «хвоста»
Рис. 12 53 Изменение средней по сечению концентрации жидкости В с расстоя-
нием х
меченой порции крови продолжается значительно дольше, чем для
ее переднего фронта, и потому «хвост» оказывается более вытя-
нутым, чем передний фронт. Возвращаясь к физиологическим при-
менениям метода разведения индикатора, следует напомнить, что
«хвост» порции индикатора может также сильно изменяться в ре-
зультате рециркуляции крови.
Для пуазейлевского течения распределение индикатора по дли-
не трубки может быть описано так, как будто жидкость покоится
и происходит только продольная диффузия, но с эффективным
коэффициентом диффузии К, много большим, чем D *). Если течение
не является пуазейлевским или если расстояние, проходимое пор-
цией индикатора вниз по течению от места введения этой порции,
недостаточно велико, то расчеты, основанные на приведенных
здесь соображениях, оказываются количественно неточными. Тем
*) Математически это впервые показал Тэйлор в 1953 г. Согласно получен-
ному им соотношению, К зависит от D, средней по сечению скорости й и диа-
метра трубки d следующим образом:
Это соотношение настолько четко подтверждено экспериментально, что теперь
на нем основан распространенный метод измерения D.
404 Часть II. Механика кровообращения
не менее в принципе механизм дисперсии индикатора, обусловлен-
ной существованием профиля продольной скорости течения крови
и ограничиваемой радиальной диффузией, продолжает действо-
вать.
Турбулентность или вторичные течения, вызываемые изгибом
либо ветвлением сосуда, перемешивая жидкость, приводят к более
интенсивному смешиванию индикатора в плоскости сечения сосу-
да, чем в результате одной только диффузии. Значит, продольная
дисперсия индикатора, ограничиваемая в таких случаях попереч-
ным смешиванием, оказывается существенно меньше дисперсии в
пуазейлевском течении с тем же расходом и тем не менее остается
весьма значительной. Опять-таки Тейлор показал, что в полностью
развитом турбулентном потоке эффективный коэффициент про-
дольной диффузии Кт любого растворенного вещества не зависит
от его коэффициента молекулярной диффузии D. Когда число Рей-
нольдса равно 2300, Кт составляет примерно 3,5-10~3 м2-с-1. Чтобы
сравнить эту величину с соответствующей ей величиной К для пуа-
зейлевского течения, следует рассмотреть какой-либо частный слу-
чай, например дисперсию кислорода в крови; в этом случае
D = 2-10-9 м2-с-1. При Re = 2300 значение К составляет около
250, а Кт меньше К примерно в 105 раз и все же в два миллиона
раз превышает коэффициент молекулярной диффузии D. При зна-
чении числа Рейнольдса 2300 порция кислорода, растворенная в
крови, «расплываясь» по артерии, займет за время t участок дли-
ной 'j'i.Kt [в соответствии с уравнением (9.3), в котором D замене-
но на К]. Если течение крови турбулентное и t = 1 с, длина этого
участка равна 8 см, а при пуазейлевском течении — 20 м. В слу-
чае ламинарного течения, возмущенного вторичными течениями,
рассчитать длину соответствующего участка нельзя. Ясно, однако,
что ее значение лежит между двумя приведенными выше — как мы
ожидаем, ближе к величине, характерной для турбулентного тече-
ния, так как механизмы поперечного смешивания сходны. Отме-
тим, что в принципе эффективный коэффициент диффузии может
быть даже выше значения, характерного для пуазейлевского тече-
ния, в том случае, когда на это течение накладываются продоль-
ные пульсации скорости. Однако экспериментальное подтвержде-
ние этому пока не получено.
Перенос веществ через стенку артерий. Так как в стенке арте-
рий протекают процессы метаболизма, необходимо, чтобы веще-
ства могли проникать внутрь стенки и удаляться из нее. Один
путь такого движения веществ — их перенос по vasa vasorum, ко
торые отходят от мелких артериальных веточек, снабжающих кро-
вью наружную оболочку крупных артерий, или же выходят непо-
средственно из их просвета. Эти сосуды, видимо, обеспечивают
питание наружной и средней оболочек крупных артерий (разд. 12.1).
Однако в лишенную капилляров внутреннюю оболочку стенки та-
ких артерий вещества поступают по другому пути — через эндоте-
Гл. 12. Артерии большого круга 405
лий. Эти выводы основаны на изучении in vivo поступления мече-
ных веществ из крови в стенку артерий. Из-за методических труд-
ностей перенос веществ по vasa vasorum подробно не изучали, и
в дальнейшем мы не будем касаться этого вопроса. С другой сто-
роны, второй путь переноса веществ — сквозь слой эндотелия —•
в последнее время оказался предметом многочисленных исследо-
ваний, в частности в связи с предположением о роли этого пути
в атероматозном поражении артерий, т. е. ранней стадии атеро-
склероза.
В наши дни атероматоз все чаще поражает людей среднего,
а не пожилого возраста. Однако при посмертном исследовании
изменения в крупных артериях нередко встречаются даже у моло-
дых людей, и многие ученые склонны считать эти изменения на-
чальными стадиями атероматоза. Судя по микроскопическим дан-
ным, процесс начинается с накопления липидов во внутренней
оболочке артериальной стенки — образуются так называемые жи-
ровые полоски, которые покрыты слоем неизмененного эндотелия
и не изменяют форму стенки сосуда. На следующей стадии забо-
левания становятся заметны приподнимающиеся над внутренней
поверхностью артерий бляшки. Покрывающий их слой эндотелия
все еще может оставаться неизмененным. Микроскопическое строе-
ние бляшек весьма различно, так как оно зависит от относитель-
ного содержания в них внеклеточных отложений жировых веществ,
нагруженных такими веществами клеток («пенистых» клеток) и
фиброзной (рубцовой) ткани. В дальнейшем эндотелий может раз-
рушиться, в результате чего эти элементы приходят в непосред-
ственное соприкосновение с кровью. А затем может произойти либо
восстановление эндотелиального слоя, либо увеличение размеров
бляшки из-за дальнейшего отложения жировых веществ, разраста-
ния рубцовой ткани и постепенного накопления слоев тромба (об-
разующегося в результате агрегации тромбоцитов, механизмы ко-
торой описаны в гл. 10). Если тромб остается в месте своего обра-
зования, проникающие в него клетки могут постепенно разрушить
его, а вновь образующийся слой эндотелия покроет тромб. В слу-
чаях, когда тромб даже полностью перекрывает просвет сосуда,
такой процесс нередко приводит к восстановлению проходимости
сосуда. Другая возможность — отрыв кусочков тромба в просвет
сосуда и их застревание в находящихся ниже по течению более
мелких ветвях. Оторвавшиеся кусочки тромба, называемые эм-
болами, приводят к различным повреждениям, степень которых оп-
ределяется анатомической организацией нижележащей части со-
судистого русла, в частности наличием анастомозов, т. е. мелких
сосудов-перемычек, позволяющих крови обходить места закупор-
ки. Прекращение кровоснабжения жизненно важных органов в
результате атеросклеротических поражений сосудов и осложнения,
вызываемые закупоркой сосудов этих органов, являются в наше
время причиной около 50% всех смертей в наиболее развитых
406 Часть II. Механика кровообращения
странах. Основные области поражений — мозг (инсульт) и сердце
(инфаркт миокарда).
Атероматозное поражение сосудов проявляется в виде пятен,
развивающихся преимущественно в определенных участках систе-
мы кровообращения. Например, как начальные проявления атеро-
матозных бляшек, так и жировые полоски обычно обнаруживают
не в периферических сосудах, а в крупных центральных арте-
риях1). Здесь первые признаки атероматозного поражения чаще
наблюдаются вблизи отхождения ветвей. Брюшная аорта пора-
жается гораздо тяжелей, чем грудная. На задней стенке нисходя-
щего отдела грудной аорты почти всегда обнаруживаются жировые
полоски, тогда как в передней стенке их обычно не бывает. Как
правило, отмечается тяжелое поражение каротидных синусов. Та-
кая предпочтительная локализация атероматозных изменений мо-
жет быть следствием одной или сразу двух причин: нормальное
строение стенки разных артерий неодинаково — и это определяет
предрасположенность некоторых их участков к развитию атеро-
матозного процесса, или же стенки этих участков артерий подвер-
гаются неодинаковым воздействиям (например, неодинаковым на-
пряжениям сдвига, вызываемым течением крови) — и это изменяет
некоторые физиологические и биохимические процессы. Первую
возможность в настоящее время исследуют, пока же предпочтение
обычно отдают второй.
Обнаруживаемые в артериях жировые полоски образуются в
результате накопления во внутренней оболочке липидов, в основ-
ном сложных эфиров холестерина. В начинающих развиваться
атероматозных бляшках также наблюдается отложение липидов
наряду с другими изменениями, включая накопление белков и не-
большое утолщение фиброзного и мышечного слоев. Полагают,
что некоторые из упомянутых липидов доставляются сюда из кро-
ви и что их накопление — процесс, который определяется неболь-
шим нарушением равновесия между количеством жировых ве-
ществ, поступающих в стенку и распадающихся или удаляющихся
из нее за то же время. Это естественно, так как перенос ве-
ществ— двунаправленный процесс, и для достижения равновесия
необходимо, чтобы число поступающих молекул вещества было
равно числу удаляемых его молекул. Однако другие липиды содер-
жатся внутри клеток и, как считают, синтезируются в них. Для
накопления таких липидов необходимо, чтобы процесс удаления
жировых веществ из стенки артерии в кровь протекал медленнее,
чем процесс переноса из крови в стенку тех веществ-предшествен-
ников (независимо от того, что это за вещества), из которых синте-
*) Мы рассматриваем здесь только ранние проявления атероматоза, так как
уже развившиеся поражения сами по себе способны существенно влиять на те-
чение крови в дистальных областях, а следовательно, и на развитие атероматоз-
ных поражений соседних участков сосуда.
Гл. 12. Артерии большого круга 407
зируются липиды. Наряду с мелкими молекулами в число таких
предшественников входят, вероятно, содержащиеся в плазме крови
липопротеиды, поскольку холестерин нерастворим в плазме и по-
этому должен переноситься в комплексе с белком (т. е. в виде ли-
попротеида). Наблюдения, результаты которых кратко обобщены
выше, привели к гипотезе о том, что атероматоз развивается в
стенке тех участков артерий, где темп удаления липидов или ли-
попротеидов по какой-то причине несколько замедлен по сравне-
нию с таковым для стенок других участков. В пользу этой гипо-
тезы свидетельствуют опыты, в ходе которых животным скармли-
вали большее количество холестерина. Содержание холестерина в
крови этих животных оказалось много выше нормального, и жиро-
вые полоски развивались преимущественно в стенке тех участков
артерий, которые, судя по данным посмертного изучения сосудов
людей, повреждаться не должны. Такой результат позволяет пред-
положить, что под действием разности концентраций холестерина
в крови и в стенке артерий, знак которой, как полагают, изменен
на обратный относительно существующего в обычных условиях,
увеличивается приток холестерина именно в те участки стенки, где
при естественно возникающем атероматозном процессе жировые
полоски не обнаруживаются. (На более поздней стадии опытов с
кормлением животных пищей, содержащей много холестерина,
скопления липидов обнаруживают в тех же самых участках арте-
рий, что и у людей.) Высказанную выше гипотезу проверяли как
in vivo, так и in vitro в ряде опытов, в основе которых лежало ис-
следование проницаемости эндотелия стенки артерий для белков,
липопротеидов и других крупных молекул и изучение влияния гид-
родинамических факторов на проницаемость эндотелия для этих
веществ.
Многие из таких опытов производили in vivo, применяя краси-
тель— синьку Эванса, которая в плазме связывается с альбуми-
ном. Те участки стенки артерии, в которые альбумин проникает
относительно быстро, окрашиваются в синий цвет и выявляются
при посмертном вскрытии в виде синих пятен. Поток меченого
альбумина через стенку пропорционален градиенту его концентра-
ции и не зависит от присутствия любых других альбуминов. По-
следние остаются в динамическом равновесии, как это было и до
введения в кровь меченого альбумина. Скорости проникновения
немеченых альмубинов в те участки, которые окрашиваются в си-
ний цвет, и скорости их удаления из этих участков также вероят-
но, повышены, но это не сказывается на их равновесной концент-
рации. Никаких заключений об общем количестве альбуминов в
стенке артерий из опытов с синькой Эванса сделать нельзя. Су-
дить же о состоянии проницаемости эндотелия можно: опыты до-
статочно непродолжительны, а при этом условии проницаемость
пропорциональна скорости увеличения концентрации меченого аль-
бумина в стенке,
408 Часть II. Механика кровообращения
Естественна мысль о том, что участок стенки артерии с повы-
шенной проницаемостью для альбумина обладает также повышен-
ной проницаемостью для других белков и липопротеидов. Эту
мысль подтвердили результаты опытов, в которых применяли ме-
ченные радиоактивными изотопами фибриноген и холестерин (по-
следний в крови, видимо, связывается с белком). Все эти иссле-
дования показали, что проницаемость эндотелия выше в тех уча-
стках стенки артерий, которые обычно не затрагиваются при на-
чальных стадиях атероматозного процесса, возникающего естест-
венным образом, т. е. у человека. Однако именно в этих участках
появляются первые признаки поражений у животных, получающих
обогащенную холестерином пищу. Таким образом, гипотеза о том,
что в естественных условиях ранние признаки атероматоза свя-
заны с низкой проницаемостью эндотелия, которая ограничивает
отток холестерина из стенки, подтверждена. Теперь возникает воп-
рос: чем же именно определяется проницаемость эндотелия?
Одна из наиболее известных гипотез заключается в том, что за
неодинаковую проницаемость эндотелия ответственно различие
сил, действующих на стенку сосуда со стороны текущей по нему
крови. Этими силами являются давление и обусловленное вяз-
костью крови напряжение сдвига на стенке сосуда. В любой точке
стенки данной артерии обе эти силы имеют вполне определенные
средние значения, относительно которых они колеблются при про-
хождении пульсовой волны, а, кроме того, при наличии турбулент-
ности на них наложены хаотические высокочастотные колебания.
Поскольку установлено, что напряжение сдвига, превышающее
40 Н-м~2, может повреждать слой эндотелиальных клеток, перво-
начально предполагали, что возникновение атероматоза связано
с повреждением эндотелия, вызываемым именно высоким напря-
жением сдвига на стенке сосудов. Однако, как будет показано
ниже, при обычных условиях напряжение сдвига в кровеносных
сосудах значительно меньше указанной величины, а для достиже-
ния столь высокого его значения необходимо, чтобы местная гео-
метрия стенки сосуда уже была изменена. Во всяком случае, позд-
нее в опытах in vitro было показано, что основным результатом
повреждения эндотелия является повышение его проницаемости
для крупных молекул, а это, как следует из изложенных выше со-
ображений, не должно было бы приводить к атероматозу.
Согласно недавно предложенной гипотезе, атероматозные по-
ражения с наибольшей вероятностью развиваются в тех областях
артериальной системы, где среднее напряжение сдвига на стенке
относительно низкое. Эта гипотеза основана, с одной стороны, на
данных о распределении жировых полосок в артериях человека,
а с другой — на вытекающих из гидромеханики выводах о положе-
нии в артериальной системе областей с низким средним напряже-
нием сдвига. Из наблюдений за распределением жировых поло-
сок, в частности тех, о которых говорилось выше, следует, что, за
Гл. 12. Артерии большого круга 409
исключением периферических отделов аорты, жировые полоски
реже всего возникают в отдаленных от сердца участках артериаль-
ной системы. Известны также данные о локальном распределении
жировых полосок вблизи ветвлений артерий [поражения наблю-
даются вблизи наружной стенки ветвления (рис. 12.44), тогда как
делитель потока обычно не затрагивается], в изгибающихся ар-
териях (здесь жировых полосок больше на внутренней стенке из-
гиба) и в каротидных синусах. Области, не склонные к образова-
нию жировых полосок, — это области характеризующиеся обычно
более высокой проницаемостью эндотелия для крупных молекул.
Выводы, следующие из гидромеханики, основаны на данных о
распределении напряжения сдвига в трубках, по которым движет-
ся непульсирующий поток жидкости, и на допущении о том, что
средний поток при пульсирующем течении аналогичен непульси-
рующему потоку с той же средней скоростью. При пуазейлевском
течении жидкости со средней скоростью й напряжение сдвига на
стенке есть
То = 8p.ii/d, (12.29)
где |i — вязкость жидкости, d — диаметр трубки (разд. 5.1). Если
бы во всех сосудах течение крови было пуазейлевским, напряже-
ние сдвига на стенке сосудов разного калибра было бы таким, как
это представлено в табл. 12.3 (при составлении этой таблицы ис-
пользовали данные табл. I, т. е. величины, характерные для сосу-
дов собаки). Явное (примерно десятикратное) увеличение напря-
жения сдвига в направлении от аорты к артериолам (табл. 12.3)
свидетельствует в пользу точки зрения, согласно которой отсут-
ствие жировых полосок в артериях небольшого калибра связано
с высоким напряжением сдвига на их стенках. Конечно, из-за на-
личия начального участка средний поток в артериях не является
пуазейлевским (и потому среднее напряжение сдвига на стенке
восходящей аорты много больше указанного в табл. 12.3, а это
находится в соответствии с тем фактом, что проницаемость эндо-
телия здесь более высокая). Непуазейлевский характер течения в
Таблица 12.3
Напряжение сдвига на стенке различных кровеносных сосудов
при пуазейлевском течении крови (Н • м~2)
Восходящая аорта Брюшная аорта Ведренная артерия Артериола
0,43 0,53 0,80 4,8
Капилляр Венула Нижняя полая вена Легочный ствол
3.7 3,0 0,64 0,28
410 Часть II. Механика кровообращения
артериях обусловлен, кроме того, вторичными течениями, вызван-
ными изгибами и ветвлениями. Большее количество жировых по-
лосок, обнаруживаемых на внутренней стенке изгиба артерии, со-
ответствует ожидаемому для этой части стенки низкому напряже-
нию сдвига. Далее, особенности непульсирующего течения в ветв-
лениях таковы, что напряжение сдвига на делителе потока дей-
ствительно должно быть велико, а на наружной стенке — мало, и
это опять-таки соответствует данным о распределении жировых
полосок *).
Таким образом, гипотеза о возникновении атероматоза в местах
с низким средним напряжением сдвига представляется весьма при-
влекательной. Однако она связана с предположениями о том, что
пульсирующие составляющие напряжения сдвига не приводят к
каким-либо качественным изменениям распределения проницае-
мости эндотелия и что среднее напряжение сдвига при пульсирую-
щем потоке распределено вдоль артериальной системы таким же
образом, как напряжение сдвига при стационарном потоке. Второе
предположение может быть в какой-то степени оправданно, так
как всюду, за исключением небольшой области вблизи входа в со-
суд, пульсирующая составляющая потока является полностью раз-
витой, и можно теоретически показать, что она не влияет на сред-
ний поток. Обосновать же правомочность первого предположения
значительно сложнее, поскольку, по-видимому, пульсирующие со-
ставляющие напряжения сдвига в артериях много больше среднего
напряжения сдвига. Чтобы пояснить сказанное, рассмотрим пол-
ностью развитое течение жидкости в прямой трубке со средней
скоростью й, амплитудой синусоидальных колебаний скорости щ
и при большом значении частотного параметра а. Среднее напря-
жение сдвига на стенке равно то [уравнение (12.29)], а ампли-
туда колебаний напряжения сдвига составляет 2apui/d. Приме-
ним эти соотношения к течению крови в аорте человека, для кото-
рой d~2,5 см, й = 0,2 м-с-1, 1 м-с-1. Полагая а — 20, полу-
чим пиковое значение напряжения сдвига, равное 6,4 Н-м-2, что
примерно в 25 раз превышает среднее значение. Более того, когда
волна скорости потока содержит высокочастотные составляющие
(это имеет место in vivo), пиковое значение напряжения сдвига
становится еще больше — оно может превышать среднее значение
в 50 раз. Представляется маловероятным, что проницаемость
эндотелиальных клеток зависит лишь от низкого среднего напря-
) Это далеко не столь очевидно, как кажется: опыты, в которых изучали
течение жидкости в полом слепке аорты, показывают, что распределение напря-
жений сдвига вблизи ветвлений (например, в месте отхождения чревной или бры-
жеечной артерий) очень сильно зависит от вполне определенных деталей геомет-
рии ветвления. Если наиболее удаленная точка раздела течений (делителя потока
иа рис. 12 47) выступает на 0,5 мм в просвет аорты, то непосредственно за ней
напряжение сдвига ие возрастает, а уменьшается, видимо, из-за отрыва потока.
Гл. 12. Артерии большого круга 411
жения сдвига при том, что реальные мгновенные его значения мо-
гут быть во много раз больше.
Таким образом, гипотезу о возникновении атероматоза в ме-
стах с низким средним напряжением сдвига нельзя считать под-
твержденной до тех пор, пока не будет понят механизм, посред-
ством которого напряжение сдвига регулирует проницаемость эн-
дотелия, а до этого еще далеко. Для проверки гипотезы был по-
ставлен ряд опытов на удаленных из организма сегментах арте-
рий, через которые пропускали кровь и измеряли приток радио-
активно меченных веществ в стенку артерий, изменяя характери-
стики течения. Результаты этих опытов позволили исключить не-
которые из возможных в принципе механизмов влияния напряже-
ния сдвига на проницаемость эндотелия, но каков на самом деле
механизм изменения проницаемости эндотелия, пока не известно.
Одно из предположений состоит в том, что проницаемость эн-
дотелия определяется сопротивлением, которое встречают крупные
молекулы, движущиеся по каналам между эндотелиальными клет-
ками, при диффузии или фильтрации. Однако прямые данные о
том, что крупные молекулы (мол. вес которых более 70000) про-
никают в стенку именно по этому пути, отсутствуют.
Согласно другому предположению, скорость проникновения ве-
ществ в стенку артерий регулируется тонким диффузионным по-
граничным слоем (разд. 9.9), в котором концентрация растворен-
ных веществ меняется от значения, характерного для ядра по-
тока, до значений у стенки. Такое предположение о механизме ре-
гуляции проницаемости эндотелия обладает двумя достоинствами:
оно предусматривает зависимость от напряжения сдвига и позво-
ляет с несомненностью доказать, что существенно именно среднее
напряжение сдвига, а не его колебания. Однако количественные
исследования in vitro показывают, что сопротивление проникнове-
нию веществ в стенку артерий примерно в 1000 раз больше со-
противления, которое мог бы оказывать пограничный слой крови.
Далее, диффузионный пограничный слой в артерии становится
толще по направлению течения крови [уравнения (9.19) или
(9.22)], и следует ожидать, что в том же направлении будет ме-
няться интенсивность перехода веществ из просвета артерии в ее
стенку. Однако в удаленных из организма артериях этого не на-
блюдается.
Таким образом, за сопротивление переносу крупных молекул
через эндотелиальный слой должны быть ответственны сами его
клетки. Возможные механизмы переноса связаны с явлением пи-
ноцитоза. Оно состоит в том, что крошечные пузырьки, образую-
щиеся у мембраны эндотелиальной клетки, захватывают мельчай-
шие порции плазмы, переносят их через клетку к противополож-
ной мембране, где и высвобождают во внутреннюю оболочку стен-
ки артерии. Такой процесс передвижения пузырьков может быть
412 Часть 11. Механика кровообращения
пассивным (диффузионным) или активным. Не исключено, что в
переносе веществ через эндотелиальные клетки участвуют и дру-
гие типы активного транспорта. Но никто еще не предложил та-
кого активного механизма переноса веществ, который позволил
бы объяснить, каким образом напряжение сдвига на стенке арте-
рии влияет на проницаемость эндотелиальных клеток, хотя в опы-
тах in vitro увеличение практически непульсирующего напряжения
сдвига на самом деле приводит к увеличению проницаемости эндо-
телия для макромолекул. Есть также некоторые указания на то,
что турбулентность не только влияет на упругие свойства стенки
артерий, но и повышает проницаемость эндотелия. Это отчасти
объясняет более существенные изменения проницаемости, обнару-
живаемые вблизи ветвлений, — там, где может происходить от-
рыв потока и, следовательно, наблюдаться турбулентность.
12.10. Дополнение: импеданс
Математические соотношения, встречающиеся в этой главе, ши-
роко используются в физиологии, так как они помогают истолко-
вывать результаты измерений. Именно поэтому мы и рассматри-
ваем их здесь, хотя это требует использования комплексных чи-
сел, чего мы до сих пор пытались избежать. В комплексной форме
волна давления, распространяющаяся в положительном направле-
нии по оси х, записывается в виде [сравните с (12.20)]
р = 5?{р+е/<а(<-*/с,>). (12.30)
Здесь Z=V—1»е— основание натурального логарифма, е1г —
экспоненциальная функция, которая обладает тем свойством, что
eIz = cos z -J- i sin z,
а символ 5? означает, что берется действительная часть комплекс-
loro выражения, записанного в фигурных скобках. Таким обра-
юм, если р+ — комплексная величина вида
Р+ = Ра+'1Рь>
го уравнение (12.30) можно переписать так:
р = Ра cos {<0 (t — х/С1)} — pb sin {<0 (/ — x/п)}.
Аналогичные выражения можно написать для волны расхода, а
также для волн, распространяющихся в противоположном направ-
лении. Обычно символ 5? опускают и всегда, когда некоторую фи-
зическую величину записывают в комплексном виде, подразуме-
вают, что речь идет лишь о действительной части ее комплексного
выражения.
Используя комплексные числа, можно записать отношение дав-
ления к расходу в любой точке трубки (сосуда), например в точ-
ке А сосуда АВ (рис. 12.28), и это отношение (которое также яв-
Гл. 12. Артерии большого круга 413
ляется комплексной величиной) называют эффективным импедан-
сом Zi Эфф в точке А:
Qa = 21Эфф • Да- (12.31)
Соответствующие вычисления для двух ветвлений, изображен-
ных на рис. 12.28, показывают, что Zi эфф можно выразить через
характеристический импеданс сосуда АВ Zi (равный pci/Ai, где
Ai и ct — площадь сечения этого сосуда и скорость волны в нем) и
эффективные импедансы сосудов 3 и 4 в точке В:
! _ , (£3-зфф + ^Гэфф) + tg (®'М)
*ЭФФ ~ + ^з-э’фф + гГэ'фф) tg «о//С1) •
(12.32)
Многократно повторяя такое рассуждение и пользуясь всякий
раз этим уравнением, можно (и выше мы говорили об этом) по
известным значениям избыточного давления и расхода в терми-
нальных ветвях сложной ветвящейся системы рассчитать отноше-
ние давления к расходу в приносящем сосуде этой сети. Следует,
однако, помнить, что уравнение (12.31) применимо только к сину-
соидальным волнам определенной частоты ю. Для волн сложной
формы, имеющих составляющие с разными частотами, определе-
ние связи давления и расхода требует проведения таких вычисле-
ний для каждой частоты в отдельности. Эффективный импеданс
Zi эфф в точке А обычно называют входным импедансом части си-
стемы, расположенной дистальнее точки А.
Отметим, что вид уравнения (12.32) зависит от того, чему равно
отношение длины I трубки АВ к величине cj/io (равной в свою
очередь произведению ’Дя на длину волны в трубке АВ). Так, на-
пример, когда (£>l/ci очень мало, величина tg(io//ci) также пре-
небрежимо мала, и уравнение будет иметь вид
Z1 эфф Z3 эфф “1“ Z4 эфф'
Иными словами, если I много меньше длины волны, то свойства со-
суда АВ несущественны и сам этот сосуд можно рассматривать
как часть ветвления в точке В. Как говорилось выше, такой же
результат получается, когда а>1/сх кратно л, т. е. при I, равном
целому числу полуволн. Напротив, при ы1/с\, равном произведе-
нию нечетного числа на л/2 (т. е. если I равно четверти длины
волны, умноженной на нечетное число), tg(io//ci) становится бес-
конечно большой величиной и уравнение (12.32) принимает вид
V—2
7-1 _ Z1____________
Z"1 эфф 1 1
^3 эфф эфф
Отсюда видно, что при Zi-1, меньшем Z-Гэфф + 2Гэ’фф, ЯГэфф будет
также меньше этой величины, и наоборот. Это соответствует сде-
ланному ранее выводу (разд. 12.4), что амплитуда прошедшей
414 Часть II. Механика кровообращения
волны будет большой, если отражение в точке В положительно, и
малой, если оно отрицательно.
Эффективный, или входной, импеданс системы есть комплекс-
ная величина и, следовательно, имеет действительную (Zr) и мни-
мую (Zi) части:
^эфф —' Zr | iZj.
При анализе свойств сердечно-сосудистой системы это выраже-
ние часто переписывают в иной форме:
гэфф = Ме1в, (12.33)
где М — амплитуда, или модуль импеданса, равный
а 0 — его фаза, определяемая соотношением
tg0 = Z,/Zr.
Модуль М характеризует сопротивление потоку со стороны части
системы, расположенной дистальнее точки измерения 2эфф, а 0 —
сдвиг фаз между давлением и потоком.
Понятие импеданса весьма полезно как для вычисления соот-
ношения между волнами давления и расхода в системе, так,и для
толкования результатов соответствующих измерений. Но одновре-
менно это и понятие, способное ввести физиологов в заблуждение,
поскольку наблюдается тенденция применять слово «импеданс»,
не отдавая себе отчета в точном его значении, и называть так
большое число близких, но различных величин. Мы перечисляем
здесь многие из этих величин не потому, что советуем пользовать-
ся ими, но в надежде, что ознакомление с этими понятиями помо-
жет избежать путаницы. Мы уже встречались с характеристиче-
ским импедансом Z сосуда — отношением давления к расходу в
этом сосуде, когда в нем распространяется с постоянной скоростью
и в одном направлении синусоидальная волна. В качестве модели
артерии мы до сих пор использовали чисто упругую трубку, в ко-
торой движется невязкая жидкость; для такой трубки характери-
стический импеданс есть действительная величина, и это означает,
что волны давления и расхода в ней синфазны. Близко к этому
понятие погонного импеданса Zl, который представляет собой от-
ношение градиента давления к расходу, когда в трубке распро-
страняется такая же волна, как и в предыдущем случае. Отметим,
что размерность погонного импеданса отличается от обычной для
импедансов размерности: она равна [ML-5T-*] вместо [ML-4T-1].
Для тех трубок, которые мы до сих пор рассматривали, погонный
импеданс есть чисто мнимая величина (ZL — —iioZ/c), так как
градиент давления и расход сдвинуты по фазе на 90°. Если час-
тота колебаний расхода жидкости в прямой жесткой трубке столь
Гл. 12. Артерии большого круга 415
мала, что в каждый момент времени имеет место пуазейлевское
течение, то градиент давления и расход синфазны и ZL — действи-
тельная величина, a Z — мнимая. Еще одна величина, с которой
мы уже встречались, — эффективный импеданс 2Эфф системы, рав-
ный отношению давления и расхода на входе в систему. Эту вели-
чину называют также входным импедансом, особенно когда речь
идет о всем сосудистом русле, а иногда даже терминальным импе-
дансом— в тех случаях, когда эту величину относят к той части
русла, которая расположена еще ниже по течению, чем та часть,
которую изучают. Наконец, существует понятие импеданса источ-
ника, которое относится к источнику потока или насосу, например
к сердцу. Если насос замкнут «сам на себя», так что поток с выхода
подается прямо на вход, то система, к которой подсоединен насос,
имеет пренебрежимо малый импеданс. Тем не менее производи-
тельность Q любого реального насоса при этом оказывается вполне
определенной величиной, и между его входом и выходом сущест-
вует отличная от нуля разность давлений Др. Отношение &p/Q,
таким образом, не равно нулю, и цепь в целом имеет ненулевой
импеданс, который принято целиком относить к свойствам самого
насоса. «Импеданс источника» существует вне зависимости от того,
к какой системе насос подсоединен.
«Импеданс источника» для левого желудочка при некотором
объеме крови в его полости равен разности между максимальным
давлением, которое способна развить мышца желудочка при дан-
ном объеме (во время фазы изоволюмического сокращения), и те-
кущим давлением в желудочке, деленной на расход изгоняемой
из него крови. Б начале периода изгнания этот импеданс примерно
в шесть раз больше входного импеданса аорты, и это означает,
что сердечная мышца здорового организма сокращается с пре-
дельной скоростью, определяемой ее собственным механизмом ог-
раничения скорости сокращения (она отражена зависимостью
сила — скорость, см. гл. 11), а не величиной нагрузки, против ко-
торой опорожняется желудочек. Однако позднее, в ходе изгнания
крови, импеданс источника уменьшается, и влияние внешней на-
грузки на желудочек — давления в аорте — возрастает.
СПИСОК ЛИТЕРАТУРЫ
Bergel D. И. (ed.). (1972). Cardiovascular fluid dynamics, Chapter 2: Pressure
measurement in experimental physiology, by I. T. Gabe; Chapter 3: Measure-
ment of pulsatile flow and flow velocity, by C. J. Mills; Chapter 9- Pressure
and flow in large arteries, by D. L. Schultz; Chapter 10: Vascular input imped-
ance, by U. Gessner; Chapter 11; The rheology of large blood vessels,
by D. J. Patel and R. N. Vaishnav; Chapter 12: The influence of vascular
muscle on the viscoelastic properties of blood vessels, by B. S. Gow; Chap-
ter 13- Post-stenotic dilatation in arteries, by M. R. Roach; Chapter 19:
Synthesis of a complete circulation, by R. Skalak, Academic Press, London
and New York (2 vols).
Berpel D. H, Schultz D L. (1971). Arterial elasticity and fluid dynamics, Prog.
Biophys. Molec. Biol., 22 (ed. J. A. V. Butler and D. Noble),
416 Часть tl. Механика кровообращения
Charm S. Е., Kurland О. S. (1974). Blood flow and microcirculation, John Wiley,
New York.
CIBA Foundation Symposium (1969). Circulatory and Respiratory Mass Transport.
Articles on: The optimum elastic properties of arteries, and Velocity distribu-
tion and transition in the arterial system, J. & A. Churchill Ltd., London.
CIBA Foundation Symposium (1973). New Series, No. 12, Atherogenesis: Initiating
Factors, Associated Scientific Publishers, Amsterdam.
Fung Y. C., Perrone N., Anliker M. (ed.). (1972). Biomechanics: Its foundations
and objectives, Chapter 5: The properties of blood vessels, by D. H. Bergel;
Chapter 15: Toward a non-traumatic study of the circulatory system, by M. An-
liker; Chapter 16: Flow and pressure in the arteries, by T. Kenner, Prentice-
Hall, New Jersey.
Guyton A. C., Jones С. E. (eds). (1974). Cardiovascular physiology, Chapter 1:
The mechanics of the circulation, by C. G. Caro, T. J. Pedley and W. A. Seed,
M. T. P. International Review of Science, Butterworth and Co., Ltd., London
and University Park Press, Baltimore.
Lighthill M. J. (1975). Mathematical biofluid dynamics, S. I. A. M., Philadelphia.
McDonald D. A. (1968). Haemodynamics, A. Rev. Physiol., 30, 525—556.
McDonald D. A. (1974). Blood flow in arteries (2nd ed.), Edward Arnold, London.
Patel D. J., Vaishnav R. N., Gow B. S., Kot P. A. (1974). Haemodynamics, A. Rev.
Physiol., 36, 125—154.
Taylor M. G. (1973). Haemodynamics, A. Rev. Physiol., 35, 87—116.
Whitmore R. L. (1968). Rheology of the circulation, Pergamon Press, Oxford.
Глава 13
МИКРОСОСУДЫ БОЛЬШОГО КРУГА
В предыдущей главе мы выяснили, что в крупных артериях
кровь можно рассматривать как однородную жидкость, не прини-
мая во внимание присутствие форменных элементов. Кроме того,
характер движения жидкости в крупных сосудах определяется
инерцией, поскольку числа Рейнольдса в них достаточно велики.
В той же главе было дано гидромеханическое обоснование пред-
ставления сосудистой системы в виде совокупности двух отдельных
частей, «граница раздела» между которыми проходит по сосудам
диаметром 100 мкм. Условия движения крови в крупных артериях
и в системе микрососудов, к которой относятся мельчайшие арте-
рии и вены, а также капилляры, весьма различны, и потому свой-
ства этого движения в соответствующих частях сосудистой си-
стемы целесообразно рассматривать раздельно.
Во-первых, в микрососудах кровь нельзя считать однородной
жидкостью — необходимо рассматривать ее как суспензию эрит-
роцитов и других форменных элементов в плазме. Как будет по-
казано в данной главе, это обусловлено тем, что диаметр даже
самых крупных микрососудов лишь в 15 раз превышает размер
эритроцита. Во-вторых, во всех микрососудах характер движения
крови определяется в основном вязкими эффектами, а не инер-
ционными, и числа Рейнольдса здесь очень малы: типичные зна-
чения их для артерии диаметром 100 мкм близки к 0,5, а для
капилляров диаметром 10 мкм они ниже 0,005 (см. табл. I).
Для крупных артерий параметр Уомерсли а (разд. 5.6) всегда
существенно больше единицы, тогда как для микрососудов этот
параметр очень мал: у собаки (если считать, что сердце сокра-
щается с частотой 2 Гц) для сосудов диаметром 100 мкм он равен
примерно 0,08, а для капилляров еще меньше — около 0,005. Это
означает, что в любом участке этих мелких сосудов кровоток со-
впадает по фазе с местным градиентом давления и условия дви-
жения крови квазистационарны. Как будет показано ниже
(разд. 13.4), давление и кровоток здесь еще могут оставаться пуль-
сирующими, но в любой точке и в любой момент времени характер
течения будет определяться только равновесием между силами
давления и вязкими силами. Силы инерции, т. е. силы, связанные
с местными и конвективными ускорениями, в этом случае прене-
брежимо малы [см. уравнение (4.5)]. Другим важным следствием
418 Часть II. Механика кровообращения
малости диаметра микрососудов является то, что, хотя скорость
кровотока в них и числа Рейнольдса низки, скорость сдвига здесь
оказывается значительно большей (порядка 1000 с-1), чем в круп-
ных сосудах. Дальше мы узнаем, что это существенно влияет на
движение крови.
Для подразделения сосудистой системы на две части имеются
не только гидродинамические, но и физиологические основания.
Назначение крупных сосудов — обеспечивать доставку крови в
микрососуды и ее отвод из них, тогда как функцией микрососудов
является перенос веществ между кровью и тканями.
Механические явления в системе микрососудов редко изучают
на собаках, и потому наши представления основаны главным об-
разом на результатах исследований на более мелких лаборатор-
ных животных — крысах, кроликах и лягушках. В таких исследо-
ваниях почти всегда приходится прибегать к методам микроско-
пии, и это ограничило их преимущественно плоскими, почти дву-
мерными сосудистыми системами. К ним относятся сосудистое
русло брыжейки и сальника (тонких прозрачных тканей, поддер-
живающих кишечник), m. cremaster (мышцы, поднимающей яич-
ко,— тонкого поверхностного мышечного слоя в стенке мошонки),
гп. tenuissimus («тончайшей мышцы» — узкого тонкого мышечного
тяжа, расположенного на бедре) и слизистой оболочки защечного
мешка хомячка. Исследовали также сосудистое русло крыла ле-
тучей мыши, которое представляет особый интерес потому, что
здесь микрососуды в отличие от других органов можно изучать на
ненаркотизированном животном. Трехмерные сосудистые русла
(например, сосудистое русло массивной скелетной мышцы) изу-
чены далеко не полно, хотя для разработки необходимых экспе-
риментальных методов были приложены значительные уси-
лия.
Прямое измерение характеристик движения крови в микросо-
судах чрезвычайно сложно не только из-за того, что размеры изу-
чаемых областей малы. Эти сосуды весьма чувствительны к лю-
бым механическим раздражениям, и потому хирургические про-
цедуры, введение в сосуды игл для измерения давления и фикса-
ция тканей на столике микроскопа являются возможными причи-
нами артефактов. Такие затруднения заставили многих исследова-
телей вместо того, чтобы иметь дело с физиологическими объек-
тами, обращаться к моделированию системы микрососудов.
Хотя наши представления о микрососудах составлены на ос-
нове сугубо фрагментарных сведений, все же ясно, что у разных
видов млекопитающих механические свойства и размеры сосудов
конкретных областей не обнаруживают заметных различий. По-
этому в настоящее время стало возможным, опираясь даже на та-
кие сведения, представить схему типичной системы микрососу-
дов.
Гл. 13. Микрососуды большого круга 419
13.1. Организация русла микрососудов
На рис. 13.1 приведена схема микроциркуляторной единицы,
построенная на основе исследования сосудов брыжейки. Посколь-
ку разные исследователи использовали для подразделения микро-
сосудов неодинаковые критерии, сравнение численных характери-
стик микрососудов разных областей крайне затруднительно. Тем
не менее в табл. 13.1 и в табл. 12.2 представлены полученные в
последнее время данные о микрососудах областей, обычно исполь-
зуемых для исследований.
Рис. 13.1. Схема микроциркуляторной единицы. В данном случае метартериола
не оканчивается разделением на капилляры, а продолжается до соединения с ве-
нулой, как основной канал. [Friedman (1971). Microcirculation. In: Physiology
(ed. Selkurt), Chapter 12, p. 260, Churchill Livingston, Edinburg and London.]
Система артериол. Как можно видеть из рис. 13.1, артериолы,
прежде чем достичь собственно капиллярной сети, делятся на ме-
тартериолы. Некоторые авторы не признают существования метар-
териол и просто считают их мельчайшими терминальными арте-
риолами. Легко понять, почему так трудно обнаружить отличи-
тельные признаки: когда размеры двух сосудов, образующих раз-
ветвление, почти одинаковы, усмотреть различие между ними не
просто. К тому же распределение сосудов по размерам непре-
рывно, и потому выделение разных категорий достаточно произ-
вольно. Показанные на схеме метартериолы являются боковыми
ветвями приводящей артериолы. Это означает, что одна артериола
отдает по своему ходу ряд метартериол и, следовательно, обра-
зует несколько капиллярных систем, функционирующих парал-
лельно. Угол отхождения метартериол от приводящей артериолы
Таблица 13.1
Сводка данных о размерах микрососудов и о скоростях движения в них крови у животных разных видов.
Данные о сосудах крыла летучей мыши относятся ко всему сосудистому руслу;
в данные о сосудах брыжейки в некоторых случаях включены данные о сосудах сальника1)
Свойства Артериолы Капилляры Венулы
крупные 5 мелкие артериаль конец ный венозный конец мелкие крупные
Диаметр, мкм
Крыло летучей мыши 19 7 3,7 7,3 21 37
Брыжейка летучей мыши 70 20 10 20 70
М. cremaster крысы 80 14 5,5 6,1 24 74
М. tenuissimus кошки 22 10 4,7 5,9 10 40
Длина, мкм
Крыло летучей мыши 3500 950 450 1000
Брыжейка летучей мыши 380 200 130 130 350
М. cremaster крысы 615 300
М. tenuissimus кошки 300 100 1000 100
Число
Крыло летучей мыши 12 120 1700 350 25
Брыжейка летучей мыши 2 9 20 10 2
М. cremaster крысы
М. tenuissimus кошки
СКОРОСТЬ, ММ'с-1
Брыжейка летучей мыши 1,0-31,7 0-1,7 0,5-11,1
М. cremaster крысы 0,8-12,9 0,2-1,2 0,4-6,6
М. tenuissimus кошки 0-1,5
’) Данные собраны в основном из следующих источников.
Schlechta, Fulton (1963). Biood flow-rates In small vessels of the hamster cheek pouch. Proc Soc. exp Biol. Med , 112, 1076.
Smaje, Zweifach, Intaglietta (1970). Micropressures and capillary filtration coefficients in single vessels of the cremaster muscle of the rat, Mie-
rovasc. Res., 2, 96-110.
Gaehtgens, Meiselman, Wayland (1970). Erythrocyte flow velocities in mesenteric microvessels of the cat, Microvasc. Res., 2, 151 — 162.
Intaglietta, ^Tompkins, Richardson (1970). Velocity measurements in the microvasculature of the cat omentum by on—line methods, Microvasc.
Erikson, Myrhage (1970). Microvascular dimensions and blood flow in skeletal muscle, Acta Physiol. Scand , 86, 211-222.
Wiedeman (1963). Dimensions of blood vessels from distributing artery to collecting vein. Circulation Res., 12, 375 -378.
Гл. 13. Микрососуды большого круга 421
существенно зависит от конкретного сосудистого русла. На мно-
гих препаратах брыжейки обнаружено, что этот угол составляет
30—60°, тогда как в скелетной мышце он часто бывает прямым.
Диаметр самой большой артериолы составляет 50—100 мкм.
По мере ее разветвления он уменьшается и на уровне отхождения
метартериол составляет около 30 мкм. Метартериолы в своем на-
чале имеют диаметр порядка 20 мкм и ответвляются от артериолы
на расстоянии около 600 мкм друг от друга. Подсчитано, что в
m. tenuissimus кошки типичное расстояние от крупной артериолы
до сети капилляров составляет I—2 мм. С уменьшением диаметра
сосуда уменьшается также толщина его стенки. У самых крупных
сосудов, диаметром порядка 100 мкм, толщина стенки составляет
около 20 мкм, а у самых мелких артериол, диаметром 20 мкм,—
около 6 мкм. Таким образом, при уменьшении калибра сосудов в
названных пределах отношение толщины стенки сосудов к их диа-
метру возрастает приблизительно от 0,2 до 0,3.
Система капилляров. Характер ветвления в системе капилля-
ров существенно меняется не только от ткани к ткани, но и в пре-
делах любого конкретного сосудистого русла. Некоторые представ-
ления об организации этих систем дает рис. 13.1. По мере того
как от метартериолы отходят истинные капилляры, гладкие мыш-
цы в ее стенке встречаются все реже (разд. 13.2). Некоторые ка-
пилляры имеют в начальном участке капиллярные сфинктеры. На
этом уровне метартериолы могут как исчезать, так и продолжать-
ся — в виде похожей на капилляр, но несколько большей, чем он,
структуры, не имеющей гладких мышц — основного, или предпоч-
тительного канала.
Прекапиллярный сфинктер состоит из одной или двух гладко-
мышечных клеток, которые обвивают место отхождения капил-
ляра от артериолы. Когда сфинктер расслаблен, его внутренний
диаметр такой же, как и самого капилляра. Такие сфинктеры об-
наружены в стенках микрососудов не всех исследовавшихся обла-
стей; в тех же сосудистых областях, где они выявлены, они не
всегда имеются при входе в каждый капилляр. Прекапиллярные
сфинктеры имеются в языке и брыжейке лягушки; в ее скелетных
мышцах они не обнаружены. Полагают, что в некоторых сосуди-
стых областях функцию сфинктеров выполняют периваскулярные
мышцы, окружающие наиболее мелкие артериолы (см. разд. 13.2.)
Вопрос о том, существуют ли в действительности основные
каналы, вызвал большой интерес. Это обусловлено тем, что, дей-
ствуя вместе с прекапиллярными сфинктерами, такие каналы мо-
гут в значительной мере управлять кровотоком и транскапилляр-
ным обменом в пределах одной микроциркуляторпой единицы. На-
пример, если бы все прекапиллярные сфинктеры закрылись, кровь
направлялась бы преимущественно в основной канал, имеющий
низкое сопротивление, и единица была бы в сущности зашунти-
рована. Такое представление о механизме шунтирования единицы
422 Часть II. Механика кровообращения
Рис. 13.2. Микрофотография m. cremaster крысы. Организация сети сосудов вы-
явлена в результате внутривенного введения мелкодисперсных частиц сажи.
Можно видеть, что главные артерии и вены идут параллельно (вены крупнее
артерий). Мышечные волокна располагаются двумя слоями, направленными при-
близительно под прямым углом друг к другу, а капилляры проходят параллельно
мышечным волокнам. [Smaje, Zweifach, Intaelietta (1970). Micropressures and
capillary filtration coefficients in single vessels of the cremaster muscle of the
rat, Microvasc. Res., 2, 99.]
Гл. 13. Микрососуды большого круга 423
несколько отличается от более старой идеи о том, что за шунтиро-
вание капиллярного русла ответственны артерио-венозные анасто-
мозы. Строение стенки таких анастомозов и стенки мелких арте-
риол и венул должно быть сходным, т. е. в ней должно быть неко-
торое количество гладких мышц. Однако артерио-венозные шунты
выявлены только в некоторых системах микрососудов, а именно в
сосудистом русле кожи и подслизистой оболочки желудка. Вообще
существует много артерио-артериальных и вено-венозных анасто-
мозов, которые делают возможным эффективное перераспределе-
ние потока крови между микроциркуляторными единицами, но ни-
коим образом не регулируют движение крови в пределах единицы.
Примеры таких анастомозов показаны на рис. 13.20.
Во многих сосудистых областях капиллярная сеть извилиста
и имеет множество поперечных связей (см. рис. 13.20). Однако
в скелетной мышце расположение капилляров значительно более
единообразно. На рис. 13.2 показана организация капиллярной си-
стемы m. cremaster крысы. Мелкие артерии и вены идут здесь
параллельно друг другу, артериолы отходят под прямыми углами,
и капиллярные сети расположены так, что капилляры параллельны
мышечным волокнам. На рис. 13.2 видны два слоя капилляров,
идущих параллельно слоям мышечных волокон, причем последние
лежат примерно под прямым углом друг к другу. Расположение
капилляров скелетных мышц (параллельно мышечным волокнам)
очень хорошо иллюстрирует рис. 13.3, на котором большинство во-
локон направлено перпендикулярно плоскости рисунка и можно
видеть, что капилляры параллельны волокнам.
Схема капиллярной сети m. cremaster крысы представлена на
рис. 13.4. Следует особо отметить, что на венозном конце капил-
ляры имеют больший диаметр, чем на артериальном, но он всегда
меньше номинального диаметра эритроцита. В настоящее время
общепризнано, что диаметр капилляров меньше диаметра эрит-
роцитов, которые должны проходить через них, а также что арте-
риальный конец капилляра является самой узкой частью сосуди-
стого русла, ибо дальше диаметр капилляра увеличивается. Так, в
m. tenuissimus кошки диаметр капилляров увеличивается от арте-
риального конца к венозному примерно от 4,7 до 5,9 мкм, тогда
как диаметр эритроцитов у кошки составляет 6,0 мкм. Капилляры
крыла летучей мыши имеют средний внутренний диаметр 3,7 мкм,
а диаметр ее эритроцитов равен 6,4 мкм. Длина капилляров m. cre-
master крысы составляет примерно 600 мкм, a m. tenuissimus кош-
ки— около 1000 мкм. В той и другой мышце множество межка-
пиллярных поперечных соединений, расстояние между которыми
около 200 мкм.
Система венул. На своем венозном конце капилляры сли-
ваются обычно попарно, образуя сначала посткапиллярные, а за-
тем собирательные венулы. Последние также постепенно сливают-
ся, образуя более крупные венулы и в конечном счете вены. Та-
424 Часть II. Механика кровообращения
Рис. 13.3. Микрофотография поперечного среза скелетной мышцы, полученная
прн помощи интерференционного электронного микроскопа. Вокруг мышечных
волокон густая сеть капилляров (показано стрелками). Слева продольный срез
мышечных волокон (/), а также сопровождающие эти волокна капилляры.
(С любезного разрешения д-ра G Gabella, University College, London.)
Рис. 13.4. Идеализированная схема капиллярной сети m. cremaster крысы. Диа-
метр капилляра у артериолярного конца 5,5 мкм, у венулярного — 6,1 мкм;
длина капилляров — 615 мкм Расстояние между капиллярами 34 мкм; расстоя-
ние между поперечными межкапиллярными соединениями 210 мкм. Плотность
капилляров 1300 мм-2; площадь поверхности капилляров 244 см2 на 1 см3 мыш-
цы; средний диаметр эритроцитов крысы 7,5 мкм. [Smaje, Zweifach, Intaglietta
(1970), Micropressures and capillary filtration coefficients in single vessels of
the cremaster muscle of the rat, Microvasc. Res., 2, 105.)
Гл. IS. МикрососуВы большого круга 428
кого вида дихотомическое слияние, хотя и очень распространено,
все же не является универсальным. Например, в m. tenuissimus
венозная сеть заканчивается одной большой венулой, к которой
с небольшими интервалами присоединяются более мелкие венулы.
Схема на рис. 13.1 подразумевает, что все капилляры, происходя-
щие из одной артериолы, сливаются в одну венулу, хотя это не
всегда так, особенно в трехмерных сосудистых областях. Размеры
венул различных сосудистых областей приведены в табл. 13.1.
Лимфатическая система. Кроме кровеносной системы суще-
ствует отдельная, но связанная с ней лимфатическая система,
функцией которой является дренирование межклеточного (интер-
стициального) пространства (разд. 13.2). Как разъясняется далее
(разд. 13.8), жидкость выходит из капилляров на артериальном
конце капиллярной системы и затем частично всасывается обратно
(реабсорбция) на ее венозном конце. Избыток жидкости, который
не реабсорбируется, попадает в лимфатические капилляры. Они
представляют собой тонкостенные «слепые» мешочки, распределен-
ные по всему межклеточному пространству, но особенно часто
встречающиеся вокруг собирательных венул. Лимфатические ка-
пилляры соединяются вместе и образуют собирательные лимфати-
ческие сосуды; они в свою очередь также сливаются в более круп-
ные лимфатические сосуды, которые идут рядом с венулами и ве-
нами. Из них образуются главные лимфатические стволы, откуда
через грудной проток лимфа попадает в подключичную вену на
шее. Таким образом лимфа возвращается в кровеносную систему
большого круга. На протяжении всей лимфатической системы (во
всех сосудах, кроме капилляров) имеются многочисленные кла-
паны, расположенные так, что лимфа может двигаться только по
направлению к грудному протоку.
Поскольку наблюдать лимфатические капилляры и собиратель-
ные лимфатические сосуды in vivo очень сложно, сведения о строе-
нии их сети весьма скудны. Чтобы обнаружить их, прибегают
обычно к введению в межклеточное пространство или непосред-
ственно в лимфатические капилляры (при помощи микропипеток)
красителей, а также мелкодисперсных частиц, например суспензий
сажи. Показано, что лимфатические капилляры брыжейки у кошки
и кролика представляют собой уплощенные мешочки шириной
40—60 мкм и толщиной 5—6 мкм.
Суммарная объемная скорость (расход) лимфы существенно
зависит от интенсивности мышечной работы, потребления пищи и
жидкости, а также от многих других факторов. У взрослого здо-
рового человека она составляет около 150 см3-ч-1.
13.2. Строение микрососудов
Общие сведения об относительном количестве различных тка-
ней, образующих три основных типа микрососудов, приведены в
табл. 13.2. Эти числа приблизительны, так как относительное ко-
426 Часть II. Механика кровообращения
Таблица 13.2
Приблизительный тканевой состав микрососудов, %
Сосуд
артериола капилляр венула
Эндотелий 10 100 20
Эластическая ткань 10
Мышечная ткань 60 20
Соединительная ткань 20 60
личество каждого компонента меняется с изменением размеров
сосудов. Об организации различных компонентов таблица не дает
никакого представления, и, чтобы получить его, необходимо рас-
смотреть сосуды каждого типа отдельно. Стенки всех сосудов
имеют сложное строение и не могут считаться изотропными. Во
всех сосудах самый внутренний клеточный слой — это слой эндоте-
лиальных клеток; во всех сосудах, кроме капилляров, можно рас-
познать гладкомышечные клетки. В последнее время при помощи
специальных методов окрашивания удалось показать, что обра-
щенная в просвет поверхность эндотелия выстлана особым веще-
ством. По-видимому, это очень тонкий слой (толщиной примерно
5—10 нм) мукополисахаридного комплекса, и именно это веще-
ство, а не эндотелий, образует истинную внутреннюю поверхность
кровеносных сосудов.
Артериолы. Артериолы можно рассматривать как очень мелкие
артерии. Строение их стенки подобно рассмотренному в гл. 12 для
артерий, хотя они, естественно, лишены питающих сосудов (vasa
vasorum). Послойная структура стенки одинакова во всех арте-
риолах, но относительное количество компонентов с уменьшением
размеров сосудов постепенно изменяется. На рис. 13.5 представ-
лена электронная микрофотография поперечного среза артериолы,
на которой, как и в случае стенки артерии, видны три основных
концентрических слоя ткани.
Внутренняя оболочка содержит эндотелиальные клетки, базаль-
ную мембрану и некоторое количество соединительной ткани. Эн-
дотелиальные клетки расположены в один слой близко друг к
другу (рис. 13.6). Смежные клетки отделены узкими межклеточ-
ными щелями, идущими от обращенной к просвету сосуда поверх-
ности к базальной мембране. Вблизи конца межклеточной щели,
обращенного к просвету, клетки образуют тесный контакт, или
соединение. Как поясняется чуть ниже, такое соединение в арте-
риолах имеет характер плотного контакта. В обычных физиоло-
гических условиях слой эндотелиальных клеток прикреплен, по-ви-
димому, непосредственно к базальной мембране, но под действием
механического напряжения или при чрезмерной утечке белков из
плазмы он может от нее отделиться. Базальная мембрана пред-
Рис. 13.5. А. Электронная микрофотография артериолы пищевода крысы. Эндо-
телиальные клетки (/) полностью окружены единственной гладкомышечной клет-
кой (2); 3 — коллагеновые волокна; 4 — эритроцит. Б. Электронная микрофото-
графия среза артериолы сердца крысы. Видны два слоя гладкомышечных клеток
средней оболочки (2). 5 — просвет сосуда; 6 — фибробласт наружной оболочки;
1 — эндотелиальная клетка внутренней оболочки. (С любезного разрешения
д-ра G. Gabella, University College, London.)
428 Часть IL Механика кровообращения
Рнс. 13.6. Электронная микрофотография замороженного скола эндотелиальных
клеток капилляра сердца. Видны межклеточные соединения (/) и впячивання по-
верхности (2), образованные открывающимися на ней пузырьками. [Leak (1971).
Frozen-fractured images of blood capillaries in heart tissue, J. ultrastruct. Res.,
35, 127—146.]
ставляет собой, по-видимому, коллагеновую сеть; впрочем, точную
форму и механические свойства этой сети еще предстоит уста-
новить. Когда внутреннюю поверхность сосуда изучают на препа-
рате, фиксированном в нерастянутом состоянии, она имеет сильно
извилистую форму. Если же фиксацию осуществляют при физио-
логических значениях давления, поверхность оказывается гладкой.
Эндотелиальные клетки артериол всех размеров выглядят оди-
наково и имеют толщину порядка 0,1—0,3 мкм, за исключением
области расположения ядра, где они утолщаются до 2—4 мкм. Все
клетки содержат пузырьки, о строении которых говорится на
стр. 439, а о функции — на стр. 508. Кроме того, в цитоплазме
эндотелиальных клеток имеются микрофибриллы. Показано, что
они содержат сократительные белки, подобные сократительным
белкам мышечных клеток, и потому, возможно, несут сократитель-
ную функцию.
Средняя оболочка артериол значительно меняется с измене-
нием их калибра. Гладкомышечные клетки этой оболочки распо-
лагаются по окружности или спирально. Как и в артериях, отдель-
ные мышечные клетки имеют форму веретен длиной 30—40 мкм
при ширине 5 мкм. В более крупных артериолах (50—100 мкм)
можно увидеть два или три слоя гладких мышц, но в сосудах диа-
Гл. 13. Микрососуды большого круга 429
метром менее 50 мкм такой слой один. В еще более мелких
артериолах количество гладких мышц в этом единственном слое
продолжает уменьшаться, а в метартериолах они и вовсе редки.
Слой гладких мышц в средней оболочке расширяясь образует
кольцо вокруг места отхождения капилляров. Когда такое кольцо
распознается четко, его описывают как прекапиллярный сфинктер.
Коллагеновые волокна обычно располагаются среди мышечных
клеток продольно, и с уменьшением количества последних число
таких волокон также уменьшается.
В артериолах диаметром более 30 мкм эндотелиальные клетки
отделены от мышечных базальной мембраной, но в более мелких
сосудах мышечные клетки через щели в этой мембране часто об-
разуют плотное соединение с эндотелиальными клетками. С умень-
шением размера сосудов такое явление встречается чаще. Парал-
лельно артериолам идут тонкие нервные волокна; иногда они об-
разуют соединения с гладкомышечными клетками средней обо-
лочки; чаще всего это наблюдается в самых мелких артериолах.
Наружная оболочка состоит главным образом из коллагеновых
волокон и небольшого числа эластических, сплетающихся в рых-
лую сеть. Внешнюю границу этой оболочки выявить трудно, так
как стенка сосуда постепенно сливается с рыхлой соединительной
тканью межклеточного пространства.
Капилляры. Когда терминальные артериолы делятся на капил-
лярную систему и основные каналы, строение стенки существенно
меняется. Стенка капилляра состоит просто из одинарного слоя
эндотелиальных клеток, окруженного базальной мембраной, кото-
рая, расщепляясь, охватывает встречающиеся иногда клетки, назы-
ваемые перицитами (рис. 13.7 и 13.8). Полагают, что эти клетки
могут превращаться в гладкомышечные.
Края эндотелиальных клеток в стенке капилляра расположены
близко друг к другу, с различной степенью перекрывания (рис. 13.6
и 13.7). Обычно на любом поперечном срезе имеется одна или две
эндотелиальные клетки, окружающие капилляр. В этих эндоте-
лиальных клетках, как и в эндотелиальных клетках более круп-
ных сосудов, в изобилии имеются пузырьки. Представление о трех-
мерной структуре капилляра дает рис. 13.8. Здесь можно видеть
эндотелиальный слой с перекрывающимися клетками, тесную связь
клеток с базальной мембраной и расщепление мембраны, окру-
жающей перицит.
Строение эндотелиального слоя зависит от исследуемой обла-
сти. Различают три основных типа эндотелия, которые показаны
на рис. 13.9. В эндотелии непрерывного типа клетки обычно приле-
жат плотно, соприкасаясь на значительном протяжении (рис. 13.7).
В сосудах поперечнополосатых мышц эндотелиальные клетки мо-
гут быть совсем плоскими, с тонкой периферической частью. В дру-
гих сосудах, например в посткапиллярных венулах, эти клетки
кубические и образуют толстый слой.
Рис. 13.7. Л. Электронная микрофотография капилляра пищевода крысы и окру-
жающей его поперечнополосаюй мышцы. Б. При большем увеличении показан
участок стенки вблизи области, обозначенной на верхнем рисунке звездочкой,
1 — базальная мембрана; 2— фибробласт; 3 — коллагеновые волокна; 4 — пери-
цит; 5 — эритроцит; 6 — эндотелиальная клетка с ядром; 7—волокно попереч-
нополосатой мышцы; 8 — митохондрии; 9 — пиноцнтозные пузырьки. (С любез-
ного разрешения д-ра G. Gabella, University College, London.)
Гл. 13. Микрососуды большого круга 431
Рис. 13.8. Трехмерная схема строения капилляра мышцы серддца с заключенными
в нем эритроцитами (/). Изображены слой эндотелиальных клеток (2), базаль-
ная мембрана (3) и перицит (4). Схема построена по трехмерным рельефным
изображениям, полученным с реплик фиксированных и нефиксированных препа-
ратов, изготовленных методом замораживания — травления. Обе поверхности
эндотелиального слоя — как обращенная в просвет сосуда, так и соприкасающаяся
с соединительной тканью — содержат пиноцитозные пузырьки (5). Расположение
клеток показано как с поверхности, так и в разрезе. Края клеток заходят друг
за друга и образуют межклеточные соединения. Базальная мембрана (3) на про-
тяжении поверхности эндотелия и примыкающего к нему перицита (4) непрерыв-
на. [Leak (1971). Frozen-fractured images of bllood capillaries in heart tissue, J.
ultrastruct. Res., 35, 127—146.]
432 Часть //. Механика кровообращения
Рис. 13.9. Классификация капилляров в соответствии со строением их эндотелия.
Выделено три основных типа — капилляры с непрерывным, окончатым и преры-
вистым эндотелием — ив каждом случае представлено два основных варианта.
Деталей показано мало, так как для разных органов они сильно различаются;
фактически можно сказать, что почти каждому органу присущ свой тип капил-
лярных сосудов. Схемы построены по данным, полученным в основном на млеко-
питающих. А, Б. Непрерывный эндотелий; в нем нет различимых отверстий; ва-
риант плоского эндотелия (А) обнаружен в капиллярах поперечнополосатых
мышц, миокарда, центральной нервной системы, гладких мышц органов пище-
варительной и половой системы, а также подкожной и жировой ткани. Вариант
кубического эндотелия (Б) типичен для посткапиллярных венул лимфатических
узлов и вилочковой железы; близкий к нему эндотелий обнаружен, кроме того,
в крупных артериях, находящихся в состоянии сокращения. В, Г. Окончатый
эндотелий; видны межклеточные окна (показаны стрелками): либо закрытые
(В), как в капиллярах эндокринных желез, сосудистого сплетения, ресничного
тела и ворсинок кишечника, либо открытые (Г), как в капиллярах почечных клу-
бочков. Д, Е. Прерывистый эндотелий имеет межклеточные щели. Такие сосуды
называют также «синусоидами»; они типичны для печени, костного мозга (Д)
и селезенки (В); детали строения эндотелия капилляров в каждом из этих слу-
чаев различны. [Majno (1965). Ultrastructure of the vascular membrane. In: Hand-
book of physiology, Section 3: Circulation, vol II, American Physiological Society,
Washington D. C.]
Клетки окончатого эндотелия настолько тонки, что противопо-
ложные поверхности их мембраны тесно прилежат друг к другу
и образуют небольшие круглые области, называемые диафраг-
мами, или окнами1). Они имеют толщину приблизительно 25 нм и
поперечник порядка 0,1 мкм. В эндотелии этого типа клетки обыч-
но еще плотно соединены друг с другом. Такой эндотелий обнару-
жен в стенке капилляров тонкого кишечника морской свинки
(рис. 13.10), сосудистого сплетения головного мозга и ресничного
тела глаза. Кроме того, окна имеются в эндотелии капилляров
почечных клубочков где, как полагают, они должны быть откры-
тыми.
') Окна представляют собой не сквозные отверстия, в области слияния двух
мембран в одну. — Прим, ред,
Рис. 13.10. А. Электронная микрофотография капилляра с окончатым эндотелием
из слизистой оболочки подвздошной_кишки морской свинки. Б. Детали строения
окна. 1—клетки всасывающего эпителия; 2—коллагеновые волокна; 3 — нерв;
4 — гладкомышечная клетка; 5 — окна; 6 — эндотелиальная клетка; 7—фибро-
бласт; 8 — просвет сосуда;' 9 — базальная мембрана. (С любезного разрешения
д-ра Q. Gabella, University College, London.)
434 Часть II Механика кровообращения
Единичные перициты
толщина стенки 0,5 мкм
Перициты, образующие
сплошной слой,
толщина стенки 1,5 мкм
Несколько слоев гладко-
мышечных клеток,
толщина стенки 2 0 мкм
Примитивная гладкомышечная клетка,
толщина стенки 1,7 мкм
Рис 13 11 А Схема системы венул Цифры (/ — 5) и стрелки указывают место-
положение срезов, представленных на Б и В Б Схема эндотелия и находящихся
снаружи от него клеток в венулах разных размеров В Рисунки с электронных
микрофотографий стенок венул различного размера, иллюстрирующие действи-
тельное расположение клеток [Rhodm (1968) Ultrastructure of mammalian ve-
nous capillaries, venules and small collecting veins J ultrastruct Res, 25, 452—
500]
Еще одним, третьим типом эндотелия является прерывистый,
характерный для капилляров печени, селезенки и костного мозга.
В этом случае клетки эндотелия отделены друг от друга ясно
различимыми щелями, а базальная мембрана также оказывается
прерывистой
Венулы. Хотя посткапиллярные венулы крупнее приносящих
к ним кровь капилляров, строение их в принципе точно такое же.
Единственное существенное различие заключается в том, что в ве-
нулах перициты образуют вокруг эндотелия почти непрерывный
слой (рис 13 11) В собирательных венулах диаметром 30—50 мкм
имеется сплошной слой перицитов с рассеянными между ними
клетками, которые считают примитивными гладкомышечными.
Гл 13 Микрососуды большого круга 435
В
456 Часть II Механика кровообращения
В венулах такого размера можно наблюдать наружный слой фиб-
робластов
В венулах диаметром 50—100 мкм эндотелий и базальная мем-
брана окружены одним или двумя сплошными слоями мышечных
клеток, образующих непрерывную среднюю оболочку Мышцы
навиваются приблизительно по цилиндрической спирали, но рас-
полагаются не столь регулярно, как в артериолах Наружная обо-
лочка, состоящая из фибробластов и коллагена, с увеличением
размера сосудов становится все более выраженной
Лимфатические сосуды. Лимфатические капилляры во многих
отношениях сходны по строению с капиллярами системы большого
круга Стенки их состоят из одинарного слоя эндотелиальных кле-
ток, окруженного базальной мембраной, и лишены гладких мышц
Эндотелиальные клетки здесь также содержат пузырьки, но в их
цитоплазме больше микрофибрилл, чем в клетках капилляров
большого круга Хотя смежные эндотелиальные клетки лимфати-
ческих капилляров и перекрываются на несколько микрон, рас-
стояние между ними меняется от нуля — в редких точках непо-
средственного соприкосновения — до 0,5 мкм
Наиболее примечательная особенность лимфатических капил-
ляров заключается в том, что их базальная мембрана не сплош-
ная и нити коллагеновых и эластических волокон проходят через
нее из межклеточного пространства к мембране эндотелиальных
клеток Представление о трехмерной структуре лимфатического
капилляра дает рис 13 12 Полагают, что фиксирующие нити ока-
зывают влияние на доставку жидкости в лимфатические капил-
ляры возможно, сокращение нитей приводит к более широкому
раскрытию щелей между клетками эндотелия Если же, напротив,
открытие щелей обеспечивается сокращением цитоплазматических
микрофибрилл, то фиксирующие нити могут способствовать ста-
билизации положения стенки капилляра
Собирательные лимфатические сосуды имеют одинарный слой
эндотелиальных клеток, окруженный непрерывной базальной
мембраной Щели между эндотелиальными клетками фактически
отсутствуют зазор составляет не больше 5 нм Здесь имеется окру-
жающий мембрану слой гладкомышечных клеток, и снаружи от
мышечного слоя лежат пучки коллагеновых волокон и небольшое
количество фибробластов
Собирательные лимфатические сосуды и все более крупные
протоки имеют клапаны, состоящие из парных створок, берущих
начало на противоположных сторонах стенки лимфатического со-
суда Створки клапанов не имеют гладких мышц, но цитоплазма
отдельных эндотелиальных клеток содержит множество микро
фибрилл, которые способны выполнять сократительную функцию
Соединения между эндотелиальными клетками. Характер свя-
зи между соседними клетками эндотелия имеет отношение как к
механической прочности эндотелиального слоя, так и к его прони-
Гл 13 Микрососуды большого круга 437
Рис 1312 Трехмерная схема строения лимфатическою капилляра, реконструиро
ванная по серии электронных микрофотографий Показаны связи лимфатического
капилляра с окружающими его структурами межклеточного пространства Видно,
что фиксирующие нити берут начало на поверхности эндотелиальной клетки и
тянутся среди пучков коллагеновых волокон, эластиновых волокон и клеток со
седних тканей, обеспечивая прочное соединение стенки лимфатического капил-
ляра с окружающей его соединительной тканью Обозначены нерегулярно появ
яяющиеся элементы базальной мембраны и коллагеновые волокна [Leak, Burke
(1968) Ultrastructure studies on the lymphatic anchoring filaments J Cell Biol,
36, 129—149]
цаемости и потому заслуживает рассмотрения Щели между клет-
ками очень узки, в связи с чем развитие представлений о строе-
нии межклеточных соединений стало возможным лишь после по-
явления электронного микроскопа.
Мембраны смежных клеток в основном параллельны и разде-
лены межклеточным пространством, или межклеточной щелью,
ширина которой примерно 15—20 нм Хотя достоверных сведений
о природе вещества, заполняющего эти щели, нет, полагают, что
438 Часть II. Механика кровообращения
Рис. 13.13 Схема эндотелиального слоя, показывающая в поперечном сечении
межклеточную щель и соединительный комплекс между соседними клетками.
Эндотелиальная клетке
zonula occludens
(плотный контакт)
Соединительный
комплекс
Базальная мембран
macula occludens
(точечное соединение)
Рис. 13 14 Схема расположения соединительных комплексов между эндотелиаль-
ными клетками.
Гл. 13. Микрососуды большого круга 439
это, возможно, мукополисахариды. Кроме того, поскольку ширина
щели весьма постоянна, допускают, что по всей длине ее между
клетками действует какая-то сила сцепления. Неизвестно, однако,
обусловлено ли возникновение этой силы сьойствами вещества,
заполняющего щель, или взаимодействием между мембранами.
Имеются также локальные области более прочного скрепления —
соединительные комплексы.
Соединительные комплексы, состоящие из ряда специализиро-
ванных структур, обычно располагаются ближе к внутренней по-
верхности смежных клеток, на расстоянии от нее, равном при-
мерно одной трети длины межклеточной щели (рис. 13.13). В этой
области мембраны соседних клеток сближаются очень сильно и
образуют непроницаемую зону, которая преграждает путь диффу-
зии молекул вдоль межклеточной щёли. Иногда комплекс обра-
зует вокруг боковых поверхностей клеток непрерывную соедини-
тельную полосу — запирающую зону (zonula occludens), а иногда
в этой полосе есть разрывы, т. е. имеются лишь запирающие пятна
(macula occludens) (рис. 13.14). Непрерывное соединение обес-
печивает образование вокруг клетки эффективной непроницаемой
перемычки, препятствующей прохождению молекул через щель,
отчего такое соединение называют плотным контактом. С другой
стороны, в случае прерывистого, или «точечного», соединения путь
для диффузии молекул остается открытым.
Прежде полагали, что плотный контакт существует во всех
эндотелиальных слоях, за исключением эндотелия капилляров,
однако в настоящее время такое представление не получило под-
тверждения.
Пиноцитозные пузырьки. Выше было отмечено, что в эндо-
телиальных клетках сосудов большого круга, а также лимфатиче-
ских сосудов имеются пузырьки. Полагают, что они играют роль
в переносе через клетки ряда веществ, в частности крупных моле-
кул. Такой процесс называют пиноцитозом. Он будет рассмотрен
позднее. Здесь мы ограничимся кратким описанием строения и
распределения пузырьков. Как можно видеть из рис. 13.7, пузырь-
ки имеются в клетке в большом количестве, и притом как свобод-
ные, т. е. в цитоплазме клетки, так и прикрепленные к клеточной
мембране. В последнем случае они могут открываться наружу.
Стенки пузырьков состоят, по-видимому, из того же вещества, что
и клеточная мембрана. Форма свободных пузырьков близка к
сферической, их диаметр составляет 60—80 нм. Пузырьки, при-
крепленные к клеточной мембране, обычно открываются наружу
через шейку длиной около 25 нм и шириной 10—25 нм. На каж-
дую клетку приходится приблизительно 500 пузырьков, и их объем
соответствует примерно 25% всего объема ее цитоплазмы, а на
каждый квадратный микрон клеточной поверхности приходится
р среднем около 120 прикрепленных пузырьков.
440 Часть II. Механика кровообращения
Межклеточное пространство. Межклеточное пространство —
это зона, окружающая кровеносные сосуды и клетки всех тканей.
В нем каким-то сложным, пока еще плохо изученным образом раз-
мещены коллагеновые и эластиновые волокна, погруженные в
жидкость. Полагают, что эта жидкость является гетерогенной
коллоидной системой (разд. 9.3), состоящей из двух компонентов,
находящихся в равновесии друг с другом. Один компонент ближе
к гелю, другой напоминает по свойствам разбавленный золь.
Сильно агрегированная гелевая фаза состоит из мукополисахари-
дов, в частности из гиалуроновой кислоты, и по большей части
находится в контакте со структурными элементами межклеточного
пространства. Богатый водой золь подвижен и содержит неагреги-
рованные растворенные вещества. Он обладает большей способ-
ностью растворять, а следовательно, и переносить водораствори-
мые вещества в пределах межклеточного пространства. Полагают,
что распределение этих двух компонентов в межклеточном про-
странстве непрерывно меняется, и вследствие этого выделить оп-
ределенные области, в которых межклеточная жидкость имела бы
какой-то конкретный состав, невозможно.
13.3. Статические механические свойства микрососудов
Статические механические свойства микрососудов, естественно,
зависят от их строения, и поэтому неудивительно, что эксперимен-
тальные данные свидетельствуют о разнообразии механических
свойств этих сосудов. Однако, поскольку малые размеры и высокая
чувствительность мелких сосудов к различным воздействиям чрез-
вычайно затрудняют измерения, знания о свойствах этих сосудов
все еще очень ограничены и частично представляют собой умоза-
ключения, основанные на косвенных данных. Упругие свойства как
артериол, так и венул довольно хорошо объясняются исходя из
соотношения компонентов их стенки, тогда как природа растяжи-
мости капилляров понята в настоящее время не столь хорошо.
В гл. 7 уже указывалось, что соотношение напряжение — де-
формация для артерий соответствует поведению вязкоупругих ма-
териалов. Это справедливо также и для микрососудов, хотя из-за
экспериментальных сложностей мало что известно об их фактиче-
ских вязкоупругих свойствах. Поэтому в данном разделе мы об-
ратимся только к упругим свойствам мелких сосудов.
Упругие свойства артериол. В гл. 7 (разд. 7.3) мы рассмотрели
статику упругой трубки и показали, что окружное напряжение
в стенке большинства кровеносных сосудов сжимающее, а не
растягивающее. Это обусловлено, во-первых, тем, что давление
внутри сосудов несколько больше, чем снаружи, а во-вторых,
относительно большой толщиной стенки сосудов. Кроме того, от-
мечалось, что обычно мы имеем дело не с абсолютной величиной
напряжения, а с ее изменениями. Так, например, представляет
Гл. 13. Микрососуды большого круга 441
2ftsz
Р=—•
интерес изменение напряжения в стенке при некотором изменении
давления в сосуде. Чтобы установить, какое напряжение необхо-
димо для уравновешивания данного растягивающего давления
(определенного по отношению к атмосферному давлению), можно
использовать закон Лапласа [уравнение (7.6)]. Величина добавоч-
ного напряжения s', возникающего в трубке с толщиной стенки h
и внутренним диаметром d под действием растягивающего давле-
ния р, определенного по отношению к атмосферному давлению,
дается соотношением
(13.1)
Произведение hs' характеризует полное натяжение стенки.
Сопоставим полное натяжение и напряжение в стенке для круп-
ных и мелких артерий, рассчитав эти величины. Рассмотрим со-
суды собаки — бедренную артерию и артериолу диаметром 50 мкм.
Диаметр и толщина стенки артерии составляют примерно 0,4 см и
0,04 см, а среднее давление в ней около 1,30-104 Н-м-2 (100 мм
рт. ст.). Толщина стенки артериолы равна примерно 20 мкм, т. е.
этот сосуд является относительно толстостенным (для артериолы
h/d = 0,4, а для артерии — только 0,1); давление в нем примерно
равно 0,9-104 Н-м-2 (70 мм рт. ст.).
Из уравнения (13.1) находим, что полное натяжение в стенке
бедренной артерии составляет 26 Н-м-1, в то время как в стенке
артериолы оно равно всего 0,22 Н-м-1. Значения добавочного на-
пряжения в стенке этих сосудов, однако, гораздо ближе друг к
другу (6,5-104 Н-м-2 для артерии и 1,1-104 Н-м-2 для артериолы).
В стенке артериолы напряжение, вероятно, развивается в базаль-
ной мембране и в средней оболочке сосуда, которые, как известно,
менее податливы, чем эндотелий и наружная оболочка. Таким об-
разом, поскольку толщина базальной мембраны и средней оболоч-
ки составляет половину всей толщины стенки артериолы, оценку
значения возникающего в них напряжения следует удвоить, т. е.
эта величина равна 2,2-104 Н-м-2. Далее, в такой толстостенной
трубке напряжение не может быть распределено равномерно по
всей несущей нагрузку части стенки. В изотропной упругой трубке
напряжение в самых внутренних слоях стенки будет примерно на
20% больше среднего значения, т. е. составит около 2,7-104 Н-м-2.
Хотя такие расчеты могут дать лишь приблизительное пред-
ставление о величине окружного напряжения в сосудистой стенке,
они все же указывают на значительное сходство условий в круп-
ных и мелких сосудах.
Соотношение напряжение — деформация определено для арте-
риол лишь в нескольких случаях, так как такие эксперименты
технически чрезвычайно сложны. Зависимость площади просвета
сосуда от величины растягивающего давления нелинейна и по-
хожа на соответствующую зависимость для артерий. С увеличе-
442 Часть II. Механика кровообращения
нием растяжения артериола становится жестче и эффективный
модуль Юнга растет. Измерения на артериолах брыжейки лягуш-
ки показали, что при диаметре, превышающем начальный на
15%, и при нулевом трансмуральном давлении касательный мо-
дуль Юнга составляет 6-104 Н-м-2, а при растяжении 45% он по-
вышается до 12-104 Н-м-2. Для коллагена модуль Юнга примерно
в 10000 раз больше. Эти данные как будто предполагают, что кол-
лаген в стенке сосудов не принимает на себя существенной части
прикладываемой нагрузки. Однако при этом не учитывается тот
факт, что столь высокая жесткость присуща коллагеновым волок-
нам лишь при условии действия напряжения параллельно их оси.
В сосудах же эти волокна направлены преимущественно про-
дольно, и лишь небольшая их часть испытывает значительное осе-
вое напряжение, поэтому оценить вклад коллагена в упругость со-
суда трудно.
В упругом поведении артериол очень важная роль принадле-
жит активному усилию (разд. 11.3), развиваемому их гладкими
мышцами, потому что оно способно значительно изменять как
диаметр, так и жесткость сосуда. В последнее время влияние ак-
тивного усилия на соотношение напряжение — деформация было
изучено in vivo на сосудах брыжейки лягушки. Сначала артериолу
растягивали в радиальном направлении, повышая до различного
уровня давление внутри нее, и измеряли диаметр (d) при расслаб-
ленных гладких мышцах. Затем при разных уровнях начального
растяжения вызывали при помощи норадреналина максимальное
сокращение мышц сосуда и определяли уменьшение диаметра (А).
Как можно видеть из рис. 13.15, ответ сосуда чрезвычайно сильно
зависит от начальной длины его мышц. Так, например, при уве-
личении растягивающего давления от низких значений до средних
способность артериолы активно суживаться заметно возрастает,
но при более высоких начальных напряжениях эта способность
вновь уменьшается. Следует отметить, что максимальный ответ
артериол наблюдается при начальных напряжениях, соответствую-
щих нормальному физиологическому уровню, а именно (1—2) •
• 104 Н-м-2. Таким образом, в обычных условиях диаметр артериол
близок к оптимальному и они способны отвечать на вазоактивные
стимулы изменением диаметра на 40—50%• Свойство, иллюстри-
руемое графиком на рис. 13.15, подобно обнаруженным для многих
других типов мышц, включая мышцы более крупных артерий мы-
шечного типа.
Характер зависимости интенсивности активного сужения сосу-
дов от длины их мышц важен в связи с вопросом о возможности
активного закрытия кровеносных сосудов. В течение ряда лет по-
лагали, что в ответ на стимул данной величины гладкие мышцы
развивают некоторое постоянное по величине активное усилие, не
зависящее от диаметра сосуда. В таком случае при медленном сни-
жении давления внутри сосуда должно наступить такое состояние,
Гл. 13. Микрососуды большого круга 443
Начальное тангенциальное напряжение 10 *. Н- м~2
Рис. 13.15. Зависимость интенсивности активного сужения артериолы брыжейки
лягушки от начального напряжения. Такая зависимость соответствует кривой
длина — напряжение при активном состоянии гладких мышц. Поведение арте-
риолы подобно поведению скелетной и сердечной мышц (см. рис. 11.9). Указаны
стандартные ошибки и число исследованных сосудов. [Gore (1972). Wall stress:
a determinant of regional differences in response of frog microvessels to norepi-
nephrine. Am. J. Physiol., 222, 82—91.]
когда сумма активного усилия и упругой восстанавливающей силы
в стенке сосуда превысит растягивающее давление и сосуд за-
кроется. Давление, при котором, как утверждали, это должно про-
исходить, назвали «критическим давлением закрытия». Такое пред-
ставление было полезным для развития соображений о взаимо-
связи активных сил и сил давления. Но теперь, принимая во вни-
мание рассмотренные выше данные, его необходимо пересмотреть,
поскольку с уменьшением растягивающего давления уменьшается
и активное усилие, противостоящее давлению. Сказанное не озна-
чает, что сосуды не могут закрыться ни при каких обстоятель-
ствах — спазм сосудов несомненно существует.
Механические свойства капилляров. При наблюдении за капил-
лярами под микроскопом можно видеть, что они в отличие от ар-
444 Часть II. Механика кровообращения
териол не пульсируют. Однако если перекрыть венозный конец
капилляра, то отчетливо видно, как эритроциты, оказавшиеся
выше места пережатия, движутся вперед и назад в соответствии
с фазами сердечного цикла и, постепенно сближаясь, сдвигаются
к перекрытому концу. Как будет показано далее, медленное дви-
жение клеток обусловлено фильтрацией жидкости из сосуда в ок-
ружающие ткани, а быстрое, колебательное движение, как пола-
гают, возникает в результате очень небольшого растяжения капил-
ляра во время систолы. Измерения такого движения клеток и ко-
лебаний давления выше места пережатия капилляра позволили
определить соотношение напряжение — деформация для капил-
ляра и рассчитать эффективный модуль Юнга, который оказался
равным 3-105 Н-м-2.
Трансмуральное давление в капиллярах обычно составляет
около 0,3-104 Н-м-2 (25 мм рт. ст.). Соответственно для капил-
ляра с внутренним диаметром 7 мкм полное натяжение в стенке
равно приблизительно 1-10-2 Н-м-1. Если считать капилляр про-
сто трубкой, плавающей в межклеточной жидкости, то это натя-
жение должно возникать исключительно в эндотелии и в базаль-
ной мембране. Предположим, что оно приходится лишь на внут-
реннюю и наружную мембраны эндотелиальных клеток (толщина
каждой мембраны 10 нм). Тогда эффективный модуль Юнга мем-
браны должен был бы составить примерно 1,3-107 Н-м-2. Эта ве-
личина много больше значения модуля Юнга, обычного для кле-
точных мембран, и потому маловероятно, чтобы упомянутые мемб-
раны были ответственны за поддержание натяжения в стенке ка-
пилляра. Если натяжение приходится на базальную мембрану
(толщина ее 20 нм), то ее модуль Юнга тоже должен составлять
примерно 1,3-107 Н-м-2. Расчетный модуль Юнга базальной
мембраны почечных канальцев, которая представляется весьма
сходной с базальной мембраной капилляров большого круга (но
несколько толще), оказался порядка (0,7—1,0)-107 Н-м-2. По-
этому возможно, что поддержание натяжения стенки капилляров
обеспечивает именно базальная мембрана. Предположив, что пол-
ное натяжение принимает на себя вся стенка капилляра (ее тол-
щина составляет приблизительно 0,7 мкм) и что она состоит из
вещества типа геля, мы получили бы величину модуля Юнга для
этой стенки примерно 4-105 Н-м-2.
Однако для объяснения механических свойств капилляров была
предложена и другая гипотеза. Согласно этой гипотезе, капилляр
следует рассматривать не как трубку, а как туннель в межкле-
точной среде. В таком случае упругие свойства капилляра опреде-
ляются упругими свойствами среды. Эту модель называют «тун-
нель в геле». Приведем результаты математического анализа рас-
тяжимости такого туннеля. Растяжимость (£)) капилляра (радиу-
сом /?) определяется как (l/A) (dA/dp), где А — площадь попе-
речного сечения капилляра, р — растягивающее давление [см.
Гл. 13. Микрососуды большого круга 445
уравнение (12.8)]. Анализ показывает, что связь растяжимости
капилляра-туннеля с растяжимостью капилляра-трубки приблизи-
тельно описывается соотношением
Г>тУв Л(1 +о)£т
где £т и Ес — модули Юнга для стенки трубки и для материала
окружающей среды, h — толщина стенки капилляра, о—коэффи-
циент Пуассона для окружающего материала. Если предположить,
что упругие свойства капилляра и окружающего его материала
одинаковы и что ткань является изотропной и несжимаемой
(о = 0,5), то для капилляра типичных размеров растяжимость
капилляра-туннеля составит примерно 1/3 растяжимости капил-
ляра-трубки.
Исходя из результатов измерения растяжимости капилляров,
можно полагать, что для геля, окружающего капилляр-туннель,
эффективный модуль Юнга должен быть порядка 1,5-104 Н-м~2.
Таким образом, материал межклеточной среды мог бы служить
для капилляра солидной опорой. Для выявления различий между
моделями «туннель» и «трубка» необходимы дальнейшие исследо-
вания. Возможно, в различных тканях поддержание формы капил-
ляров обеспечивается обоими механизмами в разной степени.
Упругие свойства венул. Посткапиллярные венулы, строение
которых сходно со строением капилляров, по-видимому, также яв-
ляются довольно жесткими сосудами. С другой стороны, в более
крупных венулах имеются гладкие мышцы, и они, вероятно, более
растяжимы. Однако подробных данных о соотношении напряже-
ние— деформация для таких венул мало. При низких трансму-
ральных давлениях очень небольшие приращения давления могут
приводить к значительному увеличению объема сосудов, но, по
всей вероятности, это обусловлено в основном раскрытием спав-
шихся сосудов. По оценкам в венулах содержится примерно 25%
всего объема крови. Поэтому полагают, что площадь их попереч-
ного сечения существенно влияет на распределение объемов крови
в сердечно-сосудистой системе. (Подробно механика спадения со-
судов рассматривается в гл. 14.)
Измерения растяжимости системы венул (при нулевом давле-
нии в артериолах) показали, что повышение перфузионного давле-
ния в венах от величины, при которой кровь не движется (т. е. от
нуля), до 0,4-104 Н-м-2 (30 мм рт. ст.) сопровождается увеличе-
нием диаметра этих сосудов более чем на 90% Затем с повыше-
нием давления до 0,9-104 Н-м-2 (70 мм рт. ст.) диаметр растет
незначительно. В других опытах на постоянном, нулевом уровне
поддерживали давление в венах, а изменяли перфузионное давле-
ние в артериолах. Оказалось, что диаметр венул при этом также из-
меняется линейно, но лишь до 130% его исходной величины, что соот-
ветствует повышению давления в артериолах от нуля до 2-104 Н-
446 Часть П. Механика кровообращения
• м-2 (150 мм рт. ст.). Таким образом, влияние давления в венах
на объем венул действительно очень велико: при повышении этого
давления от нуля до 0,13-104 Н-м-2 (10 мм рт. ст.) объем венул
возрастал на 200%, а повышение его до 0,4• 104 Н-м-2 (30 мм
рт. ст.) вызывало увеличение объема относительно исходной вели-
чины (при нулевом давлении) на 350%- Дальнейшее повышение
давления уже мало изменяло объем венул (приблизительно на
20%), что объясняется значительным увеличением в этих усло-
виях их жесткости.
13.4. Давление в системе микрососудов
Распределение давления. При многочисленных попытках полу-
чить представительные сведения о давлении в капиллярах и в при-
водящих и собирательных сосудах было произведено множество
отдельных измерений для сосудов разных диаметров. Разброс
опубликованных данных чрезвычайно велик, что обусловлено
двумя основными причинами. Во-первых, функция сосудов оди-
накового размера в разных органах и даже в одном и том же
сосудистом русле не обязательно в точности одинакова. Во-вто-
рых, давление в сосудах периодически изменяется, и эти измене-
ния бывают весьма значительными. Колебания давления могут
возникать как под действием местных факторов, так и в резуль-
тате изменений центрального артериального и венозного давления.
При наблюдении за движением крови в микрососудах наиболее
поражает, пожалуй, то, что в каждый момент времени приблизи-
тельно в четверти сосудов кровоток либо отсутствует, либо
замедлен.
В последнее время были предприняты обширные исследования
распределения давления в сосудистом русле брыжейки кошки, что
позволило получить четкое представление о профиле давления в
пределах одного русла микрососудов. (Учитывая возможность
своеобразия механических свойств сосудистого русла брыжейки,
распространять относящиеся к нему результаты на другие си-
стемы микрососудов следует с осторожностью.) По размерам со-
суды брыжейки можно разделить на пять основных групп. Прежде
всего это довольно неоднородная группа артерибл диаметром при-
мерно от 50 до 20 мкм. Далее идут обладающие высокой сокра-
тительной способностью прекапиллярные сосуды. Обе эти группы
сосудов составляют подводящую систему. За капиллярами непо-
средственно следуют не обладающие сократительной способностью
посткапиллярные сосуды, которые являются в сущности широкими
капиллярами диаметром от 10 до 20 мкм. За ними следуют более
крупные, обладающие сократительной способностью собиратель-
ные венулы (20—50 мкм).
Измерения на всех группах сосудов показали, что в сосудах
близкого размера давление обычно неодинаково, причем различие
Гл. 13. Микрососуды большого круга 447
Диаметр сосуда, мкм
Рис. 13.16. Распределение давления в сосудах брыжейки кошки, имеющих диа-
метр меньше 60 мкм. Проведена плавная кривая, которая наилучшим образом
проходит через точки, соответствующие средним значениям давления; для каж-
дой группы сосудов указано стандартное отклонение давления. Средние значе-
ния давлений вычислены для групп сосудов, диаметр которых лежит в диапазо-
не ±5 мкм от обозначенных на оси абсцисс величин. Числа в скобках указы
вают число исследованных сосудов в каждой группе. [Zweifach (1974). Quanti-
tative studies of microcirculatory structure and function. I. Analysis of pressure
distribution in the terminal vascular bed in cat mesentery. Circulation Res, 34,
843—857. С разрешения American Heart Association Inc.]
может достигать (0,26 — 0,33)-IO4 Н-м-2 (20 — 25 мм рт. ст.). Та-
кое различие, однако, не было результатом кратковременных, пре-
ходящих колебаний давления, таких как пульсовые: давление в
одном и том же сосуде оставалось фактически постоянным в те-
чение 2—3 мин, если не считать небольших пульсовых колебаний.
За более длительные периоды времени оно изменялось примерно
лишь на (0,04—0,05)-104 Н-м-2 (3—4 мм рт. ст.) даже в тех слу-
чаях, когда в соответствующей области наблюдались значительные
местные изменения кровотока. На рис. 13.16 показано распреде-
ление давления в сосудах брыжейки, диаметр которых меньше
60 мкм. Диапазон значений давления в самых крупных артерио-
лах несколько шире, чем в более дистальных сосудах. Хотя наи-
большее падение давления, как видно из этого рисунка, происхо-
дит в сосудах, предшествующих капиллярам (оно составляет при-
мерно 70% всего перепада давления), распределение градиента
давления по различным участкам сосудистого русла имеет не-
сколько другой характер. Как показано на рис. 13.17, градиент
давления возрастает по мере уменьшения размера сосудов, что со-
ответствует выводам любой теории движения вязкой жидкости
(гл. 5).
448 Часть II. Механика кровообращения
Артериолы
Венулы
1,2
i 1,0
•—
X
7 0,8
х
i °’6
ф
Среднее значение |
= разброс
—г
4015
~|------г-
6015 40±5
—1—
712
2014 20±4
Диаметр сосуда, мкм
Рис. 13.17. Градиенты давления в микрососудах, измеренные при помощи двух
микрозондов. Зонды удалены друг от друга на максимально возможное рас-
стояние, допускаемое ветвлением сосудов. В артериолах градиент давления изме-
ряли для сегментов длиной 1500—2500 мкм; в венулах обычно удавалось произ-
водить измерения для более длиннных сегментов — 2000—3000 мкм. Для капил-
ляров расстояние между зондами, как правило, составляло 200—350 мкм. В каж-
дой выделенной группе сосудов значения диаметров лежат в пределах ±5 мкм
от выбранных центральных значений; только для капилляров этот интервал со-
ставляет ±2 мкм. [Zweifach (1974). Quantitative studies of microcirculatory
structure and function, I. Analysis of pressure distribution in the terminal vascular
bed in cat mesentery. Circulation Res., 34, 843—857. С разрешения American
Heart Association Inc.]
Данные, представленные на рис. 13.16, получены на кошках,
центральное артериальное давление у которых лежало в обычных
пределах, т. е. (1,31—1,85)-104 Н-м-2 (101—142 мм рт. ст.). Ана-
логичные измерения были выполнены также на двух других груп-
пах кошек — с гипертензией, т. е. с давлением (1,85—2,54)-104 Н-
• м~2 (142—192 мм рт. ст.), и с гипотензией (0,78—1,31) -104 Н-м-2
(60—100 мм рт. ст.). Соответствующие данные представлены на
рис. 13.18. У кошек с гипертензией падение давления на мелких
артериолах и прекапиллярных сосудах было больше, чем у живот-
ных с нормальным артериальным давлением, а у кошек с гипо-
тензией— меньше. Соответственно давление в капиллярах и пост-
капиллярных сосудах у животных всех трех групп оказалось близ-
Гл. 13. Микрососуды большого круга 449
Диаметр сосудов, мкм
Рис. 13.18. Распределение давления в микрососудах у кошек с повышенным и
пониженным артериальным давлением. В группе животных с гипертензией раз-
брос данных больше. Обратите внимание на быстрое сближение давлений в пре-
капиллярных сосудах (диаметром 15—20 мкм). Давление в посткапиллярных
сосудах животных обеих групп (кошек с гипертензией 12, кошек с гипотензией
8) фактически одинаково [Zweifach (1974). Quantitative studies of microcircula-
tory structure and function, I. Analysis of pressure distribution in the terminal
vascular bed in cat mesentery. Circulation Res., 34, 843—857. By permission of
the American Heart Association Inc.)
ким: разность их средних значений составила всего (0,04—0,06) •
•104 Н-м-2 (3—5 мм рт. ст.). Кроме того, было обнаружено, что
в ответ на местное нанесение вазоактивных веществ наибольшее
изменение давления происходит в мелких артериолах и в прека-
пиллярных сосудах; давление в капиллярах и в венулах изме-
няется значительно слабее. Эти наблюдения со всей определен-
ностью свидетельствуют о том, что изменения центрального арте-
риального давления лишь очень слабо отражаются на давлении в
капиллярах и что давление здесь существенно зависит от местных
факторов.
Еще один установленный в этих опытах факт состоит в том,
что примерно в 10% венул давление было иногда выше, чем ожи-
далось [примерно 0,9-104 Н-м-2 (70 мм рт. ст.) вместо обычных
0,4-104 Н-м-2 (30 мм рт. ст.)]. Можно было бы думать, что по-
вышение давления отражает прекращение движения крови, вы-
званное, например, закупоркой венулы крупными лейкоцитами. Но
это маловероятно, поскольку такая закупорка скорее могла бы
возникнуть выше по течению. Следовательно, эти данные указы-
вают на возможность существования шунтов (подобных основ-
450 Часть II. Механика кровообращения
ным каналам), по которым кровь течет в обход системы капил-
ляров.
В распределении давления по совокупности капилляров обна-
ружены две характерные особенности. Различие среднего давле-
ния в капиллярах разных участков брыжейки достигало 0,13-
•104 Н-м~2 (10 мм рт. ст.), и столь же сильно оно различалось
в капиллярах одной локальной области. Однако в каждом конк-
ретном капилляре давление оставалось весьма постоянным на
протяжении длйтельных промежутков времени: в капиллярах с
высоким средним давлением, достигающим 0,5-104 Н-м-2 (40 мм
рт. ст.), как и в сосудах с низким давлением, вплоть до 0,26-
• 104 Н-м-2 (20 мм рт. ст.), соответствующий средний уровень дав-
ления сохранялся в течение нескольких минут. Как и следовало
ожидать, давление в капиллярах, отходящих от проксимальной ча-
сти артериол, было обычно выше, чем в капиллярах, начинаю-
щихся дистальнее.
Для каждого данного капилляра удавалось выделить несколь-
ко характерных периодов изменения давления. Прежде всего это
колебания в ритме сокращений сердца, которые наблюдались по
всей длине капилляра. Амплитуда их составляла примерно (0,01—
0,02)-104 Н-м~2 (1—2 мм рт. ст.) (рис. 13.19), но при расслабле-
нии прекапиллярного сфинктера она могла увеличиваться при-
мерно вдвое. Распространение таких колебаний по микрососудам
обсуждается ниже. Вторая форма — нерегулярные изменения дав-
ления длительностью 15—20 с; амплитуда их составляла (0,04—
0,07)-104 Н-м-2 (3—5 мм рт. ст.). Изменения давления третьего
типа были более значительны и продолжались дольше: величина
их составляла примерно 0,13-104 Н-м"2 (10 мм рт. ст.), а поддер-
живались они в течение 5—8 мин, после чего давление за 2—3 мин
возвращалось к исходному.
Хотя в действительности локальное давление меняется во вре-
мени, из рис. 13.19, на котором приведены значения давления
в различных точках капилляра, можно установить, что по длине
капилляра давление постепенно падает. Чтобы измерить давление
в капилляре, в его боковую ветвь вводили микропипетку; ветвь
при этом закупоривалась и локальное давление в капилляре мож-
но было измерять, не вызывая каких-либо местных нарушений
кровотока или его остановки.
Попытки создать математические модели, предсказывающие
распределение давления в микрососудах, к большим успехам не
привели. Это связано, по-видимому, с двумя основными причи-
нами. Во-первых, трудно устанавливать соответствие между зна-
чениями давления, предсказываемыми моделью и измеряемыми
в опытах, ибо в последних распределение давления обладает боль-
шой изменчивостью. Во-вторых, во всех моделях кровь рассмат-
ривается как однородная ньютоновская жидкость, а условия тече-
ния всегда считаются пуазейлевскими. Как уже говорилось в гл. 10,
Гл. 13. Микрососуды большого круга 451
Рис. 13.19. Реконструкция по микрофотографиям капилляра от артериолы до со-
бирательной венулы. Справа приведены записи давления в указанных на микро-
фотографиях точках капилляра и соответствующие средние значения давления.
Ширина капилляров 7,5—9 мкм. Обратите внимание на сохранение пульсовых
колебаний давления на всем протяжении капилляра. Записи давления осуществ-
ляли через боковые ветви, движение кров'и в подводящем капилляре при этом
не нарушалось. [Zweifach (1974). Quantitative studies of microcircuiatory struc-
ture and function, II. Direct measurement of capillary pressure in splanchnic me-
senteric vessels. Circulation Res., 34, 858—866. С разрешения American Heart
Association Inc.]
462 Часть II. Механика кровообращения
Диаметр сосудов, мкм
Рис. 13.20. А. Схема типичного участка брыжейки кошки. Такие участки ограни-
чены относительно крупными артериями и венами, идущими попарно. Диаметр
сосудов указан в микрометрах Б. Микрофотография типичной единицы в пре-
делах участка, капиллярная сеть которого ограничена парами артерий и вен. В.
Схематическое представление сети сосудов, изображенной ва микрофотографии
Б. Г. Расчетное распределение давления в сосудах модели единицы В — непре-
рывная кривая, методика расчета описана в тексте. Заштрихованная область —
диапазон давлений, соответствующий результатам измерения in vivo для 150 раз-
личных единиц. [Lipowski, Zweifach (1974). Network analysis of microcirculation
of cat mesentery. Microvascular Res,, 7, 73—83.]
Гл 13. Микрососуды большого круга 4ВЗ
вти допущения, вероятно, неприемлемы, что подтверждается ниже
в данной главе (разд. 13.6). Результаты одной попытки модели-
рования представлены на рис. 13.20. На нем показан типичный
участок брыжейки кошки (Д). Лежащие рядом пары артерий и
вен делят брыжейку на части, которые рассматриваются как тре-
угольные участки. Внутри каждого такого участка имеется мно-
жество единиц, ограниченных парами более мелких артерий и вен
(Б). Такие единицы можно схематически представить так, как это
изображено на рис. В: здесь все сосуды считаются прямыми труб-
ками, связанными друг с другом. На основании данных о значе-
ниях давлений и скоростей движения крови в ограничивающих
сосудах и в предположении, что течение во всех сосудах сети яв-
ляется пуазейлевским, было рассчитано распределение давления
в такой системе микрососудов. Оно показано на рис. Г. Это рас-
пределение можно сравнить с полученным экспериментально на
большом числе таких единиц (заштрихованная область). Выяв-
ленное в опытах распределение весьма сходно с распределением,
изображенным на рис. 13.16, теоретическое же значительно отли-
чается от него. Согласно расчетной кривой, резкое падение дав-
ления должно происходить именно на уровне капилляров, тогда
как эксперимент выявляет наибольшее падение давления на уров-
не сосудов, непосредственно предшествующих капиллярам. Это
расхождение может быть обусловлено влиянием различных фак-
торов; очевидным недостатком теории является игнорирование
аномальных вязких свойств крови, проявляющихся при движении
по мелким сосудам. Эти свойства обсуждаются далее.
Распространение пульсовых колебаний давления. Рис. 13.19 по-
казывает, что обусловленные сокращениями сердца колебания дав-
ления сохраняются на всем протяжении русла микрососудов, хотя
становятся значительно слабее, чем в более крупных артериях.
Подробнее эти колебания изучали в опытах, в которых давление
измеряли одновременно на входе в капилляр и на выходе из него.
Обнаруженный при этом сдвиг фаз между пульсовыми колеба-
ниями давления на разных концах капилляра подтверждает, что
пульсовые колебания распространяются от артериального конца
капилляра к его венозному концу. Если известно расстояние ме-
жду точками, в которых регистрируется давление, то по величине
сдвига фаз можно определить скорость распространения пульсо-
вых колебаний по капилляру (при частоте сокращений сердца
2,5 Гц сдвиг фаз между пульсовыми изменениями давления в
мелких артериолах и в мелких венулах составляет 25°). К сожа-
лению, величина этого расстояния в обсуждаемой работе не ука-
зана, но можно предположить, что она должна быть порядка
2 000 мкм. При таком расстоянии расчетная скорость распростра-
нения пульсовых колебаний давления составляет примерно
7,2 см-с-1. Прохождение пульсовых колебаний давления по ка-
пилляру сопровождается уменьшением их амплитуды примерно на
464 Часть П. Механика кровообращения
30% > что при экспоненциальном затухании (разд. 8.3) эквивалент'
но затуханию 83% на 1 см.
Поскольку влияние инерционных свойств крови на механиче-
ские явления в микрососудах пренебрежимо мало, механизм рас-
пространения пульсовых колебаний давления по микрососудам от-
личен от механизма распространения этих колебаний в артериях.
Как указывалось в разд. 12.2, в случае артерий возвращающая
упругая сила в их стенке уравновешивается инерционными си-
лами, возникающими в крови, и пульсовые колебания давления в
этих условиях распространяются как волны давления. Данный
механизм обеспечивает распространение пульсовых колебаний дав-
ления до мелких артерий и крупных артериол, где параметр
Уомерсли а еще близок к единице. Поэтому такие колебания со-
храняются и на входе в более мелкие сосуды, где а < 1. В микро-
сосудах упругим колебаниям стенки препятствуют вязкие, а не
инерционные силы в жидкости. Когда стенка мелкого сосуда ло-
кально растягивается под действием высокого давления, возни-
кающие в ней упругие силы стремятся вернуть ее обратно в по-
ложение равновесия, выталкивая при этом жидкость в лежащую
ниже по течению область с более низким давлением. Это приво-
дит к растяжению стенки сосуда и повышению давления в новой
области, так что те же явления повторяются снова ниже по ходу
сосуда, и т. д. Однако в отличие от того, что происходит в арте-
риях, в микрососудах вязкие силы не дают стенке сосуда при
движении ее после растяжения проскочить положение равновесия.
Поэтому колебания оказываются сильно «задемпфированными»,
подобно колебаниям простого маятника в очень вязкой жидкости
(разд. 8.3). Поскольку наличие вязкого сопротивления приводит
к диссипации механическй энергии, амплитуда пульсовых коле-
баний в микрососудах быстро уменьшается.
Если считать, что кровь является ньютоновской жидкостью, то
этот процесс можно проанализировать в рамках общей теории
распространения волн в упругих трубках при очень малом значе-
нии параметра а (эта теория описана в гл. 12, а результаты ана-
лиза представлены на рис. 12.36). Согласно указанной теории, ско-
рость с распространения колебаний давления синусоидальной
формы равна
1 .( <о у/2
с==4<Ч1йг}
где d — диаметр капилляра, D — его растяжимость, а —угловая
частота пульсовых колебаний, р—вязкость крови. Полученная по
этой формуле скорость распространения волны, имеющей частоту
2,5 Гц, по капилляру диаметром 10 мкм составляет примерно
10 см-с-1. Кроме того, из теории следует, что амплитуда пульсо-
вых колебаний давления должна очень быстро снижаться -— при-
мерно на 99,8% на длине волны (Л = 2л в отличие от случая с
Гл. 13. Микрососуды большого круга 468
высоким значением а, описанного в разд. 12.7, когда k = л д/2 /а),
что эквивалентно затуханию примерно 80% на 1 см. Расхождение
между теоретическими результатами и приведенными выше ре-
зультатами измерений обусловлено чрезмерными упрощениями
при теоретическом рассмотрении, в частности пренебрежением вяз-
коупругими свойствами ткани и неньютоновскими свойствами
крови.
13.5. Давление в межклеточном пространстве
Рассматривая статические механические свойства различных
кровеносных сосудов, мы выяснили, каким образом диаметр и
площадь просвета сосуда связаны с эффективным растягивающим
давлением (трансмуральным давлением). Однако на практике
давление внутри сосуда измеряют по отношению к атмосферному
и соответственно считают (хотя специально это обычно не огова-
ривается), что давление снаружи сосуда, т. е. давление в межкле-
точном пространстве равно атмосферному. В случае малого транс-
мурального давления это допущение оказывается существенным, и
потому его следует тщательно проанализировать.
Для измерения давления внутри тканей использовали самые
разные методы, которые привели к неоднозначным результатам.
Наиболее простой способ — введение в ткань тонкой иглы — обыч-
но дает значения давления, близкие к атмосферному, и долгое
время считали, что эти результаты достоверны. Но в действитель-
ности такой способ измерения неприемлем, поскольку введение
иглы приводит к повреждению и смещению тканей.
Более современные способы измерения обычно дают отрица-
тельные значения давления, т. е. меньше атмосферного. Точное
значение в каждом случае зависит от примененного метода, вида
животного и конкретной области измерения. Чаще всего эти значе-
ния лежат в пределах от 0 до —0,9-103 Н-м-2 (от 0 до —10 см.
вод. ст.).
К настоящему времени наиболее разработаны два основных
метода измерения тканевого давления. Целесообразно кратко опи-
сать их, чтобы дать представление о сложности интерпретации
результатов таких экспериментов. Первый метод заключается в
следующем. Под кожу вживляют небольшую полую капсулу (диа-
метром около 1 см) из пористого материала и оставляют ее в
ткани примерно на три недели. За это время поверхность капсулы
покрывается рыхлой соединительной тканью, а внутреннее про-
странство заполняется жидкостью. Затем в середину капсулы
вводят тонкую иглу и через нее измеряют давление в жидкости.
Обычно оно оказывается отрицательным. Этот метод основан на
предположениях о том, что, во-первых, ко времени измерения ме-
жду содержимым капсулы и окружающим ее межклеточным про-
странством устанавливается равновесие, а во-вторых, что осмоти-
466 Часть 11. Механика кровообращения
ческое давление заполняющей капсулу жидкости равно осмоти-
ческому давлению в межклеточном пространстве. Будь это так,
гидростатическое давление в капсуле и в окружающей ее ткани
было бы одинаковым. Однако состав жидкости и структура веще-
ства внутри капсулы и в межклеточном пространстве, по-види-
мому, различны, и потому предположение о равенстве осмотиче-
ских давлений внутри и снаружи капсулы спорно. Таким образом,
результаты этих измерений не следует принимать безоговорочно.
Второй метод сводится к введению в межклеточное простран-
ство ватного фитилька. Несколько прядей ваты помещают в про-
свет узкой полиэтиленовой трубочки так, чтобы примерно 1 см
фитилька выступал из нее. Другой ее конец соединяют с датчиком
давления и через него всю трубочку заполняют изотоническим
солевым раствором. Тот конец трубочки, в котором закреплен фи-
тилек, вводят под кожу через иглу, которую затем удаляют, оста-
вив трубочку в ткани. Полагают, что волокна фитилька через мель-
чайшие каналы обеспечивают гидростатическую связь между тру-
бочкой и межклеточным пространством. По прошествии часа из-
меряемое давление более или менее стабилизируется. Считают, что
это давление соответствует давлению в тканевой жидкости. Как и
при измерении методом вживления капсулы, давление оказывается
отрицательным.
Возможной причиной того, что давление в тканевой жидкости
ниже атмосферного, считают наличие в межклеточном простран-
стве неполностью гидратированного геля, который состоит из очень
крупных молекул (таких, как молекулы гиалуроновой кислоты) и
способен поглощать воду. Отнимая ее у жидкой фазы (золя), геле-
вая фаза набухает. При этом концентрация в золе тех белковых
молекул, которые слишком велики, чтобы проникнуть в гель, по-
вышается, и осмотическое давление в ткани возрастает. Если вода,
поступающая в межклеточное пространство из сосудов, не уда-
ляется из него постоянно, то объем последнего увеличивается
до тех пор, пока гель не окажется полностью гидратированным.
Тогда между золем и плазмой установится осмотическое равнове-
сие и тканевое давление увеличится до атмосферного или станет
несколько выше него. Дегидратация обеспечивается удалением из
тканей воды через лимфатическую систему и ее обратным по-
ступлением в кровь через венулы. Следовательно, в нормальных
условиях осмотическое давление межклеточной жидкости несколь-
ко превышает осмотическое давление плазмы. Если трубочка, вве-
денная в межклеточное пространство, содержит солевой раствор с
таким же коллоидным осмотическим давлением, как и плазма, то
вода будет стремиться уходить из трубочки. С другой стороны,
тканевой гель не может проникнуть в нее, и в результате будет
регистрироваться отрицательное давление.
Давление в лимфатических капиллярах измеряли микропипет-
ками, и оно оказалось чуть выше атмосферного. Поскольку эндо-
Гл. 13. Микрососуды большого круга 467
телий этих капилляров не образует непрерывного слоя, гидроста-
тически они свободно связаны с окружающей средой. Такие до-
воды использовали для обоснования представления о том, что
давление в лимфатических капиллярах отражает давление в меж-
клеточном пространстве. Однако мы уже видели (разд. 13.2), что
клетки прерывистого эндотелия взаимно перекрываются. Это поз-
воляет сторонникам представления об отрицательном давлении в
тканях утверждать, что гидростатическая непрерывность между
межклеточным пространством и лимфатическими капиллярами
возникает лишь тогда, когда межклеточное пространство подвер-
гается действию сжимающих сил, обусловленных, например, со-
кращением скелетных мышц. В этом случае гидростатическое дав-
ление в ткани становится положительным и превышает давление
в лимфатических сосудах. Предполагается, что при этом фиксирую-
щие нити, показанные на рис. 13.12, открывают лимфатические ка-
пилляры и удерживают их в таком состоянии, благодаря чему ста-
новится возможным отток жидкости в эти капилляры.
Итак, представляется вероятным, что существующие методы
измерения тканевого давления в действительности измеряют осмо-
тическое, а не истинно гидростатическое давление. Для измерения
гидростатического тканевого давления требуется зонд, заполнен-
ный жидкостью с таким же коллоидным осмотическим давлением,
каким характеризуется тканевой золь, причем размеры этого
зонда должны быть меньше, чем размеры заполненных жид-
костью пространств в ткани. Таким образом, всем известным ме-
тодам измерения тканевого давления присущи недостатки, но боль-
шая часть наблюдений свидетельствует о том, что давление в
межклеточном пространстве несколько ниже атмосферного,
13.6. Движение жидкости в моделях микрососудов
и в относительно крупных микрососудах
При любом анализе движения крови в микрососудах необхо-
димо принимать во внимание наличие в ней форменных элементов.
Действительно, рассматривая принципы измерения вязкости крови
(гл. 10), мы выяснили, что при протекании крови в каналах, раз-
мер которых сопоставим с размерами эритроцитов, поведение ее
оказывается аномальным. Наши представления об этом явлении,
как и о многих других механических явлениях в системе микро-
сосудов, опираются в значительной мере на результаты исследо-
вания движения суспензий искусственных частиц или цельной кро-
ви в стеклянных трубках. Особенности течения крови в микросо-
судах определяются эритроцитами, так как концентрация их в
крови значительно превышает концентрацию других форменных
элементов. В связи с этим дальнейшее изложение будет отно-
ситься в основном к поведению в потоке эритроцитов. Установ-
лено, что во многих отношениях движение крови аналогично ДВ1|-
458 Часть 11 Механика кровообращения
жению суспензии, содержащей частицы, которые имеют форму
сфер, дисков и палочек. Поэтому сначала мы рассмотрим движе-
ние таких суспензий и, постепенно усложняя картину, будем пе-
реходить от суспензии с одиночными частицами к суспензиям с
большим числом частиц и от трубок, размер которых значительно
превышает размер частиц, к узким трубкам, в которых частицы
движутся в один ряд. Во всех случаях мы будем иметь дело с по-
токами в таких трубках или сосудах, для которых число Рей-
нольдса обычно мало, т. е. меньше единицы, а скорость сдвига на
стенке может достигать 1000 с-1.
Движение одиночных частиц в потоке с очень низкой скоро-
стью течения. Прежде всего мы рассмотрим поведение одиночных
сферических частиц, палочек и дисков, взвешенных в ньютонов-
ской жидкости, имеющей такую же плотность, как и эти частицы,
и движущейся предельно медленно в длинной прямой трубке, для
которой число Рейнольдса меньше единицы. Такой случай яв-
ляется простейшим, так как ни взаимные влияния частиц, ни
столкновения между ними при этом невозможны. Когда никаких
частиц в жидкости нет, то ниже начального участка поток имеет
характерный для пуазейлевского течения параболический профиль
скорости, и градиент скорости, или скорость сдвига G в точках
потока (положение их характеризуется радиусом г), линейно уве-
личивается от нуля — для точек, лежащих на оси, до максимума —
для точек около стенки (см. рис. 13.21 и разд. 5.1).
Движение сферических частиц. Если мы введем в поток неболь-
шую недеформируемую сферическую частицу с нейтральной пла-
вучестью, то увидим, что она, во-первых, движется вместе с пото-
ком и, во-вторых, вращается с постоянной угловой скоростью. Ско-
рость перемещения частицы по трубке несколько меньше скорости
и, которую имела бы жидкость в точке, совпадающей с центром
частицы в отсутствие таковой. Эта разность (называемая ско-
ростью скольжения) растет с увеличением размера частицы. Вра-
щение обусловлено характером распределения напряжений сдвига
по поверхности частицы и происходит так, как показано на
рис. 13.21 и ранее на рис. 10.2. Кроме того (рис. 13.21), на сфери-
ческую частицу действуют различные по величине нормальные к
ее поверхности напряжения, так что на одной части поверхности
она подвергается сжатию, а на другой — растяжению. При очень
низких скоростях потока твердая сферическая частица движется
вдоль линий тока; перемещения ее поперек потока не возникают.
Поведение в таком же потоке капли жидкости, не смешиваю-
щейся с остальной жидкостью (т. е. податливой сферы), очень
сильно отличается от поведения твердой сферической частицы.
Поскольку нормальное к поверхности капли напряжение в разных
ее точках различно, она деформируется и приближается по своей
форме к эллипсоиду (рис. 13.22). При равновесии силы давления
₽ жидкости, стремящиеся деформировать каплю, уравновешивают-
Гл 13 Микрососуды большого круга 459
ся силами поверхностного натяжения, которые стремятся восстано-
вить сферическую форму. Тщательное наблюдение за поверхностью
капли показывает, что она вращается в том же направлении, что
и твердая сфера, причем вместе с поверхностными слоями вра-
щается, как показано на рис. 13.22, и все содержимое капли. Кро-
Рис. 13.21. Поведение твердой сферы радиуса Ь, имеющей нейтральную плаву-
честь в пуазейлевском потоке в трубке радиуса а Сфера движется вдоль труб-
ки, сохраняя свое исходное радиальное положение г, и одновременно враща-
ется в указанном направлении Кроме того, по всей поверхности сфера испыты-
вает действие различных по величине нормальных к ее поверхности напряжений;
в результате одна часть ее поверхности подвергается сжатию, а другая — растя-
жению
ме того, из любого исходного положения капля перемещается
радиально по направлению к оси трубки (рис. 13 23). Скорость
радиального перемещения максимальна вблизи стенки; по мере
продвижения капли к оси трубки она уменьшается. Теоретически
Рис. 13 22. Деформация и вращение
капли жидкости в потоке, распреде-
ление скорости сдвига в котором та-
кое же, как в изображенном на
рис 13 21 пуазейлевском потоке По-
казано также вращение жидкости
находящейся внутри капли
показано, что такое радиальное перемещение обусловлено дефор-
мируемостью капли, характером распределения скорости сдвига
в жидкости и наличием стенки. При уменьшении деформируемости
капли уменьшается и скорость ее радиального перемещения.
С увеличением размера частицы (по отношению к радиусу трубки)
и с увеличением скорости потока (и соответственно с увеличением
разности скоростей сдвига на противоположных сторонах капли)
эта скорость возрастает. Понятно, что в потоке, в котором линей-
ная скорость во всех точках одинакова, скорость сдвига во всех
точках равна нулю. Поэтому капля в таком потоке не будет дефор-
мироваться и, следовательно, не будет радиально перемещаться.
460 Часть II. Механика кровообращения
Рис. 13.23. Микрофотографии, демонстрирующие деформацию и радиальное пе-
ремещение взвешенной капли; регистрирующая камера движется вдоль трубки
со скоростью, соответствующей скорости движения капли. А — капля в состоя-
нии покоя вблизи стенки. Б, В, Г — последовательное изменение радиального
положения капли и деформация ее в эллипсоид при движении жидкости по
трубке вверх. Обратите внимание на перемещение капли от стенки трубки к ее
оси. Скорость перемещения и степень деформации капли по мере перемещения
уменьшаются, поскольку снижается местная скорость сдвига в потоке. Диаметр
капли 600 мкм; диаметр трубки 0,8 см; отношение диаметра трубки к диаметру
частицы равно 13. [Mason, Goldsmith (1969). The flow behaviour of particulate
suspensions In: Circulatory and respiratory mass transport (ed. Wolstenholme),
J. A. Churchill, London !
Движение палочек и дисков. Движение жестких палочек и
дисков несколько сложнее, чем движение жестких или жидких
сфер. Перемещение таких частиц вдоль трубки также сопровож-
дается их вращением, поскольку, как показано на рис. 13.24, даже
когда ось палочки или плоскость диска параллельны направлению
потока, скорости движения жидкости у противоположных сторон
частицы не одинаковы. Однако угловая скорость вращения таких
частиц уже не постоянна. Силы, вызывающие их вращение, уве-
личиваются с ростом угла <р между осью палочки или плоскостью
диска и направлением потока, и поэтому как палочки, так и диски
при движении вдоль трубки «кувыркаются». Большую часть пе-
риода вращения они находятся в положении, в котором их длин-
Гл. 13. Микрососуды большого круга 461
ная ось почти параллельна направлению потока, а в остальной
части цикла быстро переворачиваются.
Как показано на рис. 13.24, гибкие палочки и диски испыты-
вают, кроме того, деформации, обусловленные их попеременным
сжатием и растяжением. Как и деформируемые жидкие капли, они
также передвигаются к оси трубки. Жесткие палочки и диски при
низкой скорости потока не совершают такого перемещения.
Рис. 13.24. Вращение палочек и дисков в потоке со сдвигом. Обратите внимание,
что гибкие палочки и диски во время той части цикла их вращения, когда они
подвергаются сжатию, могут деформироваться.
Движение одиночных частиц в потоке с высокой скоростью те-
чения. При более высоких числах Рейнольдса (больше единицы),
т. е. когда становится значимым действие инерционных сил, де-
формируемые частицы по-прежнему перемещаются к оси трубки.
Но в этом случае в радиальном направлении перемещаются и
твердые частицы, хотя и не всегда от стенки трубки к ее оси.
Поведение твердых частиц в таких условиях называют приосе-
вым эффектом, или эффектом Сегре — Зильберберга, по именам
исследователей, впервые сообщивших о нем. Частицы, находив-
шиеся сначала около стенки трубки, сдвигаются к оси, а частицы,
находившиеся вблизи оси, — к стенке. И те и другие стремятся за-
нять равновесное положение, находящееся от оси на расстоянии
приблизительно 0,6п, где а — радиус трубки. Скорость перемеще-
ния частиц зависит от их радиального положения, уменьшаясь по
мере приближения к положению равновесия. С увеличением числа
Рейнольдса и размера частиц (относительно радиуса трубки) эта
скорость повышается. Механизмы, лежащие в основе этого явле-
ния и определяющие движение частиц, сложны и связаны как с
инерцией жидкости, так и с взаимодействием частиц со стенкой.
Характер движения твердых и деформируемых частиц при
очень низких и при несколько более высоких скоростях течения
изображен на рис. 13.25. Из него ясно, что, за исключением случая
движения твердых частиц в потоке с очень низкой скоростью тече-
ния, всегда имеет место смещение частиц от стенки трубки.
Движение одиночных эритроцитов при пуазейлевском течении.
Движению изолированных эритроцитов в плазме присущи свойства
движения как твердых, так и деформируемых частиц. Когда ско-
рость течения в трубке достаточно низка, так что местная ско-
462 Часть П. Механика кровообращения
Медленное вязкое течение
Вязкое течение при
наличии инерции
! К
Деформируемые твердые]
к®"ли ' частиць1
Ц ©iP I
Деформируемые ' твердое
капли частицы
Перемещение
к оси трубки
Ото тствие
радиальных
перемещений
Направление
течения
Перемещение
к оси трубки
Перемещением
I в двух
нш1равлениях
Положение!
равновесия ;
Положения равновесия
Перемещение к оси трубки
Приосевой эффект
Рис. 13.25. Схемы перемещения твердых и деформируемых частиц в трубке: сле-
ва — при очень низких числах Рейнольдса, справа — при числах Рейнольдса, су-
щественно больших единицы; вверху — продольный разрез трубки по диаметру,
внизу — поперечный разрез. На нижней части рисунка показано равновесное
положение частиц, когда их перемещение в поперечном сечении трубки обуслов-
лено деформацией частиц (слева) и инерцией жидкости — приосевой эффект
(справа). [Goldsmith (1971). Red cell motions and wall interactions in tube flow.
Fedn Proc. Fedn Am. Socs Exp. Biol., 30, 1578—1588.]
рость сдвига G вблизи поверхности клеток не превышает примерно
20 с-1, клетки вращаются подобно жестким дискам и сохраняют
при этом свою двояковогнутую форму. Они переворачиваются та-
ким же образом и с той же периодичностью, как это теоретически
предсказывается для жестких дисков того же диаметра. Кроме
того, как и жесткие диски, они вращаются вокруг своей малой
оси (рис. 13.26). При столь низких скоростях сдвига приблизи-
тельно 50% времени клетки проводят в положении, в котором
отклонение их больших осей от направления течения не превы-
шает ±20°; смещения к оси трубки в этих условиях не наблю-
дается.
Гл. 13. Микрососуды большого круга 463
При скорости сдвига больше 20 с-1 поведение эритроцитов с
обычными свойствами начинает постепенно отклоняться от пове-
дения жестких дисков. Клетки проводят больше времени в поло-
жении, близком к обозначенному на рис. 13.26 цифрой 3. Когда
скорость сдвига превышает 100 с-1, подавляющее большинство
клеток находится именно в этом положении. Тщательное наблюде-
ние за мембраной эритроцита позволяет предположить, что она
движется вокруг содержимого клетки. Согласно результатам оп-
Эритроциты человека
Жесткие диски
Рис. 13.26. Поведение взвешенных жестких дисков и эритроцитов человека в пуа-
зейлевском потоке со скоростью сдвига меньше 20 с-1. На рисунке приведены
выполненные по микрофотографиям изображения вращающихся частиц при раз-
личной ориентации их за время, равное периода полного переворота. Направ-
ление вращения указано стрелками. Наибольшую угловую скорость частицы
имеют в положениях 1 и 5, когда направление их собственной оси вращения (она
обозначена штриховой линией в положении 3) совпадает с направлением потока.
[Goldsmith (1971). Red cell motions and wall interactions in tube flow. Fedn
Proc. Fedn Am. Socs exp. Biol., 30, 1578—1588].
ределения размеров клетки, последняя деформируется, но двояко-
вогнутая форма ее сохраняется. При столь высоких скоростях
сдвига наблюдается также радиальное перемещение клеток от
стенки трубки, скорость которого возрастает с увеличением ско-
рости сдвига. При очень высоких скоростях сдвига, порядка
5000 с-1, возникает эффект Сегре — Зильберберга, но положение
равновесия эритроцитов оказывается ближе к оси трубки, чем
ожидаемое для жестких дисков.
При скоростях сдвига меньше чем примерно 50 с_| в потоке
могут образовываться монетные столбики (гл. 10). Они ведут
себя так же, как и гибкие палочки, довольно быстро перемещаясь
в радиальном направлении. По мере увеличения скорости сдвига
столбики постепенно разрушаются возникающими при их враще-
нии силами растяжения и сдвига.
Изучено также поведение эритроцитов, жесткость которых "по-
вышена путем обработки их глутаральдегидом. При всех скоро-
стях сдвига такие клетки ведут себя как жесткие диски
(рис. 13.27) и не совершают радиальных перемещений до тех пор,
пока скорости сдвига не окажутся достаточно высокими для воз-
никновения эффекта Сегре — Зильберберга.
464 Часть П. Механика кровообращения
Средняя по сечению скорость сдвига. с~
Рис. 13.27. Зависимость относительного количества эритроцитов [обычных (О)
и с повышенной путем обработки глутаральдегидом жесткостью (•)], ориенти-
рованных в потоке так, что угол между их главной осью и направлением потока
ие превышает 20°, от скорости сдвига. Горизонтальная прямая — независящая от
скорости сдвига ориентация, полученная расчетным путем для жестких дисков,
период вращения которых при заданной скорости сдвига равен периоду перево-
рота эритроцитов человека. [Goldsmith (1971). Red cell motions and wall in-
teractions in tube flow. Fedn Proc. Fedn Am. Socs exp. Biol., 30, 1578—1588.]
Мы уже выяснили, что профиль скорости жидкости, подвер-
гающейся при протекании по длинной прямой трубке действию
пульсирующего градиента давления, зависит от величины пара-
метра Уомерсли а. В микрососудах а обычно меньше 0,08 (см.
табл. I), поэтому можно ожидать, что в отсутствие частиц в лю-
бой момент времени будет иметь место квазистационарное тече-
ние. Но движение жидкости, содержащей частицы, весьма сложно,
и было бы неосмотрительно предполагать, что явления, характер-
ные для стационарного потока, будут наблюдаться также и в пуль-
сирующих потоках. Тем не менее из тех немногих модельных ис-
следований, в которых сопоставляли поведение искусственных час-
тиц и эритроцитов, следует, что описанные выше явления действи-
тельно наблюдаются также и в пульсирующих потоках.
Движение концентрированных суспензий искусственных частиц
и эритроцитов. Эксперименты показывают, что если концентрация
частиц в протекающей по трубке суспензии превышает всего0,5%,
то частицы начинают взаимодействовать друг с другом и возни-
кают явления, похожие на столкновения частиц. Это постепенно
Гл. 13. Микрососуды большого круга 465
изменяет характер движения частиц и профиль скорости. Кроме
того, эритроциты деформируются в таком потоке значительно
сильнее, чем в разбавленной суспензии.
Для наблюдения за движением отдельных частиц и эритро-
цитов в концентрированных суспензиях разработаны специальные
методы. Например, большинство частиц можно сделать практиче-
ски прозрачными, так подобрав материал для частиц и состав
жидкости, чтобы коэффициент преломления света в них был оди-
наков, и добавить в суспензию несколько светонепроницаемых
частиц. Для наблюдения за движением эритроцитов небольшое
количество их вводят в суспензии теней эритроцитов, способ при-
готовления которых описан в гл. 10.
Чтобы понять, как ведут себя концентрированные суспензии,
следует учесть влияние как концентрации суспензии с, так и от-
носительного размера частиц b/а (где а — радиус трубки, Ь —
радиус частицы, рис. 13.21). Когда в качестве частиц используют
относительно небольшие жесткие сферы и концентрация их в сус-
пензии не превышает примерно 5%, профиль скорости движения
сфер (построенный по результатам измерения скоростей этих час-
тиц) близок к параболическому. По мере увеличения относитель-
ного размера жестких сфер или их концентрации профиль ско-
рости частиц становится все более плоским. Для гибких и жидких
частиц с такими же относительными размерами и при такой же их
концентрации уплощение профиля скорости оказывается меньше.
Это различие в поведении частиц иллюстрирует рис. 13.28. В верх-
ней части рисунка приведены профили скорости крупных и мел-
ких жестких сфер, а также крупных жидких капель; во всех трех
случаях концентрация частиц составляет примерно 30%. Можно
видеть, что при такой концентрации профиль скорости крупных
жестких сфер плоский и область поршневого течения, в которой
все сферы движутся с одинаковой продольной скоростью, прости-
рается поперек трубки почти до ее стенок. Более мелкие жесткие
сферы не образуют столь широкой области поршневого течения.
Сопоставив поведение жестких сфер и жидких капель, можно убе-
диться, что в случае крупных жидких капель профиль скорости их
движения уплощается меньше, чем в случае жестких сфер вдвое
меньшего диаметра.
Для жестких частиц профиль скорости определяется только
концентрацией суспензии и относительным размером частиц, от
средней скорости потока он не зависит. Напротив, в суспензиях
жидких капель и гибких частиц профиль их скорости зависит от
средней скорости потока; степень сглаженности его с увеличением
этой скорости уменьшается.
В нижней части рис. 13.28 приведены профили скорости для
суспензии теней эритроцитов и жестких дисков при близких раз-
мерах и концентрациях частиц. Для профиля скорости движения
дисков характерна более широкая область поршневого течения.
466 Часть II. Механика кровообращения
Рис. 13.28. Профили скорости
взвешенных искусственных частиц
и теней эритроцитов, движущихся
по прямой трубке при очень низ-
ких числах Рейнольдса. Значения
локальных скоростей и (г) отнесе-
ны к скорости в центре трубки
иц; это означает, что профили по-
строены в таком масштабе, что
скорость в центре трубки всегда
равна единице. В верхней части
рисунка представлены профили
скорости для крупных (1; b/а =
= 0,12) и мелких (2; b/а = 0,06)
жестких сфер и для крупных ка-
пель жидкости (3; Ь/а = 0,12) при
одинаковой во всех трех случаях
концентрации частиц. Внизу изо-
бражены профили скорости для
суспензий теней эритроцитов и
жестких дисков; размеры частиц
и их концентрации примерно оди-
наковы. Профили скорости теней
эритроцитов построены для двух
разных средних скоростей тече-
ния й. [Goldsmith (1972). The
flow of model particles and blood
cells and its relation to thrombo-
genesis. In: Progress in hemosta-
sis and thrombosis. С разрешения
Grime and Stratton Inc., New
York.)
Кроме того, видно, что в случае суспензий теней эритроцитов с
увеличением средней скорости потока эта область уменьшается.
При более высоких скоростях профиль скорости для теней эрит’
роцитов близок к параболическому.
Механизм, лежащий в основе такого различия поведения час-
тиц, чревычайно сложен. Однако очевидно, что это различие обу-
словлено разной деформируемостью капель и клеток. В концент-
рированных суспензиях эритроцитов сильная деформация их мемб-
раны наблюдается уже при низких скоростях сдвига — порядка
3 с-1, тогда как деформация отдельных взвешенных в жидкости
эритроцитов происходит при значительно больших скоростях
сдвига — около 100 с-1.
Необходимо подчеркнуть, что профили скорости движения час-
тиц не обязательно отражают профиль скорости течения самой
жидкости, особенно вблизи стенки. В случае суспензии жестких
сфер, в которой профиль скорости движения частиц является пло-
ским по всей трубке, это очевидно: вблизи стенки скорость жид-
кости должна быстро уменьшаться, а непосредственно у стенки
ста!?овиться равной нулю в силу условия прилипания (разд. 4.3).
Однако по крайней мере для суспензий жестких частиц профиль
Гл. tS. Микрососуды большого круга 46?
скорости жидкости определяется профилем скорости частиц и не
зависит от средней скорости течения. Это следует из того факта,
что «вязкость» суспензий жестких частиц не зависит от средней
скорости течения и определяется только размером частиц и их
концентрацией. Поскольку вязкость таких суспензий не зависит
от скорости сдвига, их называют «квазиньютоновскими». Суспен-
зии капель указанным постоянством вязкости не обладают, так
как в них профили скоростей движения капель зависят от сред-
ней скорости течения. Вероятно, последнее справедливо также и
для профиля скорости течения жидкой фазы суспензии.
Относительно радиального перемещения частиц в концентри-
рованных суспензиях имеются только качественные данные: сооб-
щали о небольшом снижении концентрации частиц вблизи стенки
и об эффекте Сегре — Зильберберга. В случае движения суспензий
эритроцитов с нормальным гематокритом положение оказывается
более сложным. В потоках с достаточно высокими скоростями
сдвига эритроциты движутся как отдельные клетки, а вблизи стен-
ки, по-видимому, существует тонкий слой жидкости, не содержа-
щий клеток. Однако при более низких скоростях сдвига клетки
стремятся собираться в монетные столбики и толщина свободного
от клеток слоя значительно увеличивается. Такого увеличения сле-
дует ожидать, так как для более крупных частиц силы, вызываю-
щие их радиальное перемещение, оказываются больше.
Вязкость цельной крови. Наблюдения за движением концент-
рированных суспензий искусственных частиц и эритроцитов позво-
ляют качественно понять влияние скорости сдвига на вязкость
цельной крови (гл. 10, рис. 10.16). При самых низких скоростях
сдвига большая часть эритроцитов объединяется, образуя монет-
ные столбики. Эти столбики длинные, обладают определенной"же-
сткостью и, переплетаясь друг с другом, образуют единую струк-
туру, которая делает поведение крови похожим на поведение твер-
дого тела. Когда напряжения сдвига превышают предел текучести
такого «твердого тела» (разд. 10.7), монетные столбики изгибают-
ся, частично разрушаются и связующая решетка распадается. При
дальнейшем увеличении скорости сдвига длина монетных столби-
ков постепенно уменьшается, и в конце концов в суспензии ос-
таются лишь несвязанные друг с другом эритроциты. Сопоставле-
ние изменений формы гибких палочек в концентрированных и раз-
бавленных их суспензиях показывает, что даже при низких ско-
ростях сдвига палочки, находящиеся в концентрированных сус-
пензиях, искривляются гораздо сильнее. Таким образом, можно
предположить, что в цельной крови разрушение монетных стол-
биков происходит при значительно более низких скоростях сдвига
(1 с-1), чем в их разбавленных суспензиях. Поэтому быстрое
уменьшение вязкости цельной крови при скоростях сдвига около
1 с-1 можно объяснить разрушением монетных столбиков и умень-
шением их размеров.
468 Часть II. Механика кровообращения
Уменьшение вязкости цельной крови, наблюдаемое при ско-
ростях сдвига примерно между 10 и 100 с-1, является, вероятно,
результатом постепенного изменения формы отдельных эритро-
цитов. Как отмечено выше, обычные эритроциты, находящиеся
в концентрированных суспензиях их теней, сильно деформируются
при очень низких скоростях сдвига — около 3 с-1. Наконец, при
очень высоких скоростях сдвига (порядка 5000 с-1) одиночные
эритроциты начинают вести себя так, как если бы они были твер-
дыми частицами (стр. 465). Однако на кривой зависимости вяз-
кости цельной крови от скорости сдвига плато наблюдается уже
при скоростях сдвига, превышающих примерно 150 с-1. Это позво-
ляет думать, что в концентрированной суспензии эритроциты ве-
дут себя подобно твердым частицам и при таких относительно низ-
ких скоростях сдвига.
Радиальные блуждания эритроцитов. Во время движения по
трубке концентрированных суспензий эритроцитов клетки постоян-
но сталкиваются друг с другом и отдельные эритроциты хаоти-
чески перемещаются по весьма замысловатым траекториям. Такое
поведение эритроцитов изучали, наблюдая за их движением в сус-
пензиях теней. Оказалось, что величина радиальных хаотических
смещений эритроцита зависит от его радиального положения в
трубке (рис. 13.29).
Измеряя радиальные смещения эритроцитов, можно определить
их коэффициент диффузии (разд. 9.1). Для примера, показанного
на рис. 13.29, коэффициент диффузии изменяется от 3-10~8 см2-с-1
вблизи оси трубки до 1,5-10~7 см2-с_1 при радиальном положении
г = 0,7а. Это существенно больше значения коэффициента диф-
фузии (порядка 4-10~’° см2-с-'1), которое можно было бы ожи-
дать, если бы эритроциты смещались в результате броуновского
движения, т. е. из-за столкновений с молекулами жидкости. Инте-
ресно отметить, что коэффициент диффузии эритроцитов близок к
расчетному коэффициенту свободной диффузии макромолекул
в плазме: при 37 °C типичное значение его для альбумина состав-
ляет 1 • 10~7 см2-с-1.
Радиальные блуждания эритроцитов сопровождаются переме-
щением окружающей их плазмы и других форменных элементов
крови. Изучение радиальных блужданий жестких сфер диаметром
2 мкм (моделирующих тромбоциты) показало, что в движущейся
суспензии эритроцитов они смещаются даже больше, чем сами
эритроциты (рис. 13.29). При непродолжительном наблюдении
коэффициенты диффузии таких сфер и эритроцитов, находящихся
в одном и том же радиальном положении, приблизительно совпа-
дают. Однако при измерении за более длительное время коэффи-
циент диффузии микросфер оказывается большим, так как они
переходят из области влияния одних эритроцитов в область влия-
ния других. Таким образом, можно видеть, что присутствие эрит-
Гл. IS. Микрососуды большого круга 469
Рис. 13.29. Изменение во времени радиального положения обычных эритроцитов-
меток (вверху) и микросфер, изготовленных из поливинилового латекса (внизу),
при движении их в концентрированной суспензии теней эритроцитов, текущей
по стеклянной трубке диаметром 76,5 мкм со средней скоростью 0,015 см-с-1.
Размер частиц указан на рисунке. Точки, в которых наблюдалось соприкосно-
вение эритроцита и микросферы со стенкой, отмечены надписью «удар». Микро-
сфера после соприкосновения со стенкой прилипла к ней; г)а — отношение ра-
диальной координаты частицы к радиусу трубки. [Goldsmith (1971). Red cell
motions and wall interactions in tube flow. Fedn Proc. Fedn Am. Socs exp.
Biol., 30, 1578—1588.]
470 Часть II. Механика кровообращении
роцитов значительно повышает эффективную подвижность тромбо-
цитов в потоке крови.
Пристеночный слой. Из рассмотренного выше описания динами-
ческих процессов следует, что как при постоянном, так и при пуль-
сирующем течении крови по трубке клетки стремятся отойти от
стенки и сконцентрироваться в средней части потока. В резуль-
тате у самой стенки трубки образуется слой жидкости, обеднен-
ной клетками, — пристеночный слой. Этот слой не является пол-
ностью свободным от клеток, так как высокая концентрация их
Рис. 13 30. Образование свободного от клеток
слоя вследствие невозможности размещения
сфер на расстоянии от стенки трубки, меньшем
радиуса сферы.
в средней части трубки и перемешивание за счет радиального
блуждания стремятся выталкивать клетки в эту область, где они
соударяются со стенкой (рис. 13.29).
Отметим, что по чисто геометрическим причинам свободный
от клеток пристеночный слой образуется даже в отсутствие тече-
ния. Рассмотрим равномерно упакованный блок сфер (рис. 13.30)
и окружность, представляющую стенку трубки, содержащей эти
сферы. Очевидно, что те сферы, для которых расстояние от центра
до стенки оказывается меньше их радиуса, не могут поместиться
в трубке, и это приводит к уменьшению концентрации сфер вблизи
стенок.
В движущейся крови толщина пристеночного слоя зависит как
от средней скорости течения, так и от отношения радиуса клеток
к радиусу трубки. Поскольку наблюдать за потоком через стенки
сосуда технически очень сложно, сколько-нибудь успешных пря-
мых измерений толщины этого слоя in vivo провести не удава-
лось. Однако измерения в стеклянных трубках диаметром по-
рядка 100 мкм показывают, что его толщина составляет примерно
2—4 мкм. Влияние этого свободного от клеток слоя заключается
в создании вокруг суспензии, движущейся в средней части трубки,
оболочки из жидкости (плазмы) с низкой вязкостью. Физиологи-
ческое значение пристеночного слоя в системе микрососудов может
быть существенным, и его необходимо изучать.
Динамический гематокрит. Измерив гематокрит цельной крови,
находящейся в резервуаре, и гематокрит той же крови, вытекаю-
щей из него по трубке, мы обнаружим, что в трубке он ниже. Это
явление известно давно. Как показано дальше, оно частично объ-
ясняет изменение вязкости крови в узких трубках.
Гл. 13. Микрососуды большого круга 471
Наблюдаемое снижение гематокрита обусловлено наличием
свободного от клеток слоя, ибо взвешенные в плазме эритроциты
движутся вместе с ней в центральной части трубки с относительно
большой скоростью, а плазма движется не только вместе с эрит-
роцитами, но и у стенки, где скорость ее перемещения мала. Это
явление имеет место независимо от вида профиля скорости. В ре-
зультате среднее время прохождения данного отрезка трубки для
эритроцитов оказывается меньшим, чем для плазмы. Если бы ве-
личина динамического гематокрита была такой же, как его ста-
Обогащенная
плазмой
кровь
Рис. 13.31. Эффект отделения плазмы в мелком ответвлении.
тическая величина на входе в трубку, то на конце трубки концент-
рация эритроцитов должна бы была увеличиваться! В действи-
тельности динамический гематокрит, измеренный в любой доста-
точно узкой трубке, всегда меньше статического. Поэтому, хотя
время прохождения через трубку отдельного эритроцита меньше,
чем время прохождения плазмы, общее число эритроцитов, прохо-
дящих через трубку за определенное время, поддерживается на
соответствующем уровне.
Отделение плазмы. Представим себе тонкую ветвь, отходящую
от более крупного микрососуда (рис. 13.31). Весьма вероятно, что
кровь в такую ветвь будет поступать относительно медленно и со-
держание плазмы в ней окажется больше, чем в питающем ее
крупном микрососуде. Произойдет это потому, что в мелкий сосуд
поступает в основном та кровь, которая находится в крупном со-
суде около стенки и содержит больше плазмы. Это явление назы-
вается отделением плазмы.
Эффект Фореуса — Линдквиста. Фореус и Линдквист измеряли
кажущуюся вязкость крови в трубках различного диаметра — при-
мерно от 0,1 см до 10 мкм. Они проводили опыты при скоростях
сдвига, достаточно высоких для того, чтобы не могли образовы-
ваться монетные столбики и чтобы вязкость не зависела от ско-
рости сдвига (разд. 10.7).
Оказалось, что значение кажущейся вязкости не зависит от
диаметра трубки, если он несколько больше 0,1 см. Однако для
трубок меньшего диаметра по мере уменьшения последнего кажу-
щаяся вязкость снижается (рис. 13.32). Даже в трубках, диаметр
472 Часть //. Механика кровообращения
которых близок к размерам недеформированного эритроцита, этот
эффект выражен очень сильно.
Из эффекта Фореуса — Линдквиста следует, что напряжение
сдвига около стенки с уменьшением диаметра трубки снижается.
Напомним (гл. 5), что градиент давления, вызывающий движе-
ние жидкости по трубке, уравновешивается напряжением сдвига,
которое обусловлено действием стенки на жидкость. В свою оче-
редь напряжение сдвига определяется как произведение скорости
сдвига в жидкости, находящейся вблизи стенки, на местную вяз-
Рис. 13.32. Зависимость отношения кажущейся вязкости цельной крови к кажу-
щейся вязкости плазмы от радиуса трубки при движении по узкой цилиндриче-
ской трубке. [Haynes (I960). Physical basis of the dependence of blood viscosity
on tube radius. Am. J. Physiol., 198, 1193—1200.]
кость жидкости. В движущейся суспензии частиц, каковой являет-
ся кровь, вязкость пристеночного слоя ниже, но в то же время
скорость сдвига в этом слое больше, чем при наличии в нем час-
тиц, а потому напряжение сдвига у стенки может увеличиться. Сле-
довательно, сразу невозможно точно сказать, в какую сторону из-
менится в результате напряжение сдвига у стенки. В трубках с
большим диаметром относительная толщина свободного от кле-
ток слоя мала, в результате чего согласование профилей скорости
в этом слое и в ядре потока происходит на сравнительно неболь-
шом расстоянии. Однако в узких трубках относительная толщина
пристеночного слоя больше и согласование профилей скорости
может происходить на большем расстоянии. Таким образом, ско-
рость сдвига вблизи стенки может быть не так велика, как это
предполагалось. Второй фактор, влияющий на напряжение сдви-
га,— это динамический гематокрит, который снижается с умень-
шением размеров трубки или сосуда. В результате кажущаяся
рязкость также падает.
Гл. 13. Микрососуды большою круга 473
Профили скорости в сосудах. Измерить характерные скорости
кровотока и профили скорости в очень мелких сосудах оказалась
еще сложнее, чем измерить давление в них. Это обусловлено глав-
ным образом сложностью наблюдения за движением отдельных
эритроцитов в сосудах, более крупных, чем капилляры. Кроме
того, рассматривая под микроскопом сеть микрососудов in vivo,
нельзя не заметить изменчивость кровотока в любом из ее сосу-
дов. На протяжении некоторого периода времени течение может
быть равномерным, но затем внезапно замедлиться или вообще
остановиться. Такие изменения скорости движения крови не соот-
ветствуют изменениям давления в сосуде. Вероятно, этого и сле-
дует ожидать, поскольку в таких сетях давление и кровоток в од-
ном сосуде зависят от значений этих величин в примыкающих
сосудах.
В капиллярах, где удается наблюдать за движением отдельных
эритроцитов, скорость кровотока оценивают исходя из линейной
скорости эритроцитов, считая, что в таких сосудах имеют место
условия поршневого течения. Типичные результаты измерения ско-
рости движения крови в капиллярах приведены в табл. 13.1. Срав-
нительно недавно появилась возможность измерять скорость кро-
вотока и в несколько более крупных сосудах. Для этого исполь-
зуют фотометрический метод, причем сосуд освещают через две
расположенные близко друг к другу щели. Одно время этот метод
применяли для измерения не только средних скоростей, но и про-
филей скорости в просвете сосуда и обнаружили, что последние
являются плоскими. Однако не так давно было установлено, что
фотометрический метод непригоден для таких измерений и чрез-
мерно уплощает профили скорости. В других опытах надежно до-
казано, что профиль скорости в протекающей через узкие трубки
цельной крови близок к параболическому при любых значениях
гематокрита. Близкие результаты получены для суспензий теней
эритроцитов (рис. 13.28). Таким образом, по-видимому, эритро-
циты гибки настолько, что при любых значениях параметров, ко-
торые реализуются in vivo, в условиях in vitro устанавливается
приблизительно параболический профиль скорости. Поэтому до
тех пор, пока в опытах на сосудах не будут получены достаточно
надежные прямые данные, разумно полагать, что, за исключением
капилляров, во всех микрососудах профиль скорости близок к па-
раболическому. Но так как пристеночный свободный от клеток
слой может оказывать заметное влияние на значение скорости
сдвига вблизи стенки, было бы неверным рассчитывать падение
давления вдоль сосуда исходя из профиля скорости в его средней
части (ядре).
Условия движения крови вблизи разветвления сосудов неиз-
вестны. Тем не менее сомнительно, чтобы на расстоянии от об-
ласти ветвления, превышающем радиус сосуда, профили скорости
существенно отличались от параболического, поэтому наличие ветв-
474 Часть 11. Механика кровообращения
ления мало сказывается на характере потока. Последнее связано
с тем, что в мелких сосудах числа Рейнольдса значительно меньше
единицы, а в гл. 5 было показано, что для ньютоновских жидкостей
при таких числах Рейнольдса длина начального участка имеет
порядок диаметра трубки.
13.7. Движение крови в капиллярах
Капилляры млекопитающих имеют диаметр от 10 до 3 мкм,
так что клетки крови, особенно эритроциты, должны проходить
через них «гуськом». Чтобы пройти через капилляр, диаметр ко-
торого меньше диаметра эритроцита (7—8 мкм), последний дол-
жен деформироваться. Наблюдения показывают, что обычные эрит-
роциты внутри мелких капилляров принимают самую разную фор-
му. Одни из них изгибаются наподобие купола парашюта
(рис. 13.33,Л), другие свертываются в трубочку и продвигаются
по сосуду вперед торцом (рис. 13.33,Б). Как мы видели, обычные
эритроциты способны значительно деформироваться, не меняя при
этом ни своего объема, ни площади поверхности (разд. 10.4). Из-
мененные же клетки, которым придали большую жесткость или
почти сферическую форму, утрачивают способность к столь силь-
ной деформации и потому не могут проходить через самые мелкие
сосуды.
В специальных экспериментах создавали условия, при которых
эритроциты проходили через мембраны, имеющие поры различных
размеров. Оказалось, что обычные эритроциты могут проходить, не
разрушаясь через поры диаметром больше 3 мкм, но при протал-
кивании через поры меньшего диаметра они разрушаются
(рис. 13.34). Клетки с искусственно повышенной жесткостью или
принявшие под действием осмотического давления сферическую
форму (как то, так и другое делает их менее гибкими) не могут
проходить через поры диаметром меньше 8 мкм, не подвергаясь
гемолизу.
Описана болезнь крови, именуемая наследственным сфероцито-
зом, при которой эритроциты имеют сферическую форму и диаметр
около 6 мкм. У таких больных гемолиз резко усилен, и хотя обра-
зование эритроцитов у них увеличено в несколько раз, оно не ком-
пенсирует их потерю в результате разрушения, и развивается ане-
мия. Удаление селезенки в этих случаях приводит к значитель-
ному улучшению. Причина этого, вероятно, заключается в том, что
при прохождении через капилляры паренхимы селезенки эритро-
цитам приходится «протискиваться» через очень узкие щели — до
3 мкм. Нормальные клетки могут проходить через них без чрез-
мерного гемолиза, точно так же, как они проходят через микро-
поры близкого диаметра в искусственном фильтре. Но патологи-
чески измененные сферические клетки не способны к этому.
Рассматривая механику движения эритроцитов в узких капил-
лярах, мы можем выделить две ситуации в зависимости от того,
Гл. 13. Микрососуды большого круга 475
Рис. 13.33. Микрофотографии эритроцитов, движущихся в узких капиллярах.
А — эритроциты, имеющие форму купола парашюта; Б — эритроцит, свернувший-
ся в «трубку» и движущийся вперед торцом. [Skalak, Branemark (1969). Defor-
mation of red blood cells in capillaries. Science, N. Y., 164, 717—719.]
насколько легко эритроциты входят в эти сосуды. Первая — когда
диаметр капилляров заметно превышает размеры клеток. Клетки
могут входить в такие трубки, не изменяя своей формы (эту си-
туацию называют случаем положительного зазора), и, проходя по
одной вдоль сосуда, деформируются незначительно или не дефор-
мируются вовсе. Давление в плазме в этом случае примерно оди-
наково во всех точках поперек капилляра (но есть более или ме-
нее постоянный продольный градиент давления, обеспечивающий
движение крови) и едва ли способно существенно изменять форму
клеток. Поэтому, анализируя динамику такого движения, дефор-
мируемостью клеток можно пренебречь. Вторая, более распростра-
ненная ситуация возникает в капиллярах, диаметр которых мень-
ше диаметра эритроцитов (отрицательный зазор). В этом случае,
чтобы войти в капилляр, клетки должны деформироваться, а при
продвижении по капилляру форма их изменяется еще сильнее.
Чтобы клетка деформировалась, в тонком слое жидкости, отде-
ляющем ее от стенки капилляра, должно развиться местное высо-
кое давление. Величина его зависит от упругих свойств клетки
(а также от упругих свойств стенки капилляра, хотя она значи-
476 Часть //. Механика кровообращения
Рис. 13.34. Полученные при помощи сканирующего электронного микроскопа ми-
крофотографии эритроцитов до (Д) и после (5) прохождения их через сито из
поликарбоната с диаметром пор 2,2 мкм. На Л многие клетки выглядят сфериче-
скими, потому что они уже частично вошли в поры. [Chien, Luse, Bryant (1971).
Hemolysis during filtration through micropores: a scanning electron microscopic
and hemorheologic correlation. Microvasc. Res., 3, 197—198.]
Гл. 13. Микрососуды большого круга 477
тельно жестче, чем клетка). В целом характер течения опреде-
ляется движением в этих узких областях высокого давления. Ука-
занное движение можно проанализировать при помощи методов,
используемых в гидродинамической теории смазки (в общих чер-
тах они описаны ниже). Рассмотрим обе ситуации.
Положительный зазор. В этом случае детали формы и простран-
ственного расположения клеток, а также их упругие свойства не
имеют существенного значения. Основные особенности такого те-
чения можно уяснить, рассмотрев простую модель, в которой эрит-
роциты заменены жесткими сферами, равномерно и симметрично
движущимися одна за другой по цилиндрической трубке (рис. 13.35).
Рис. 13.35. Линии тока и профили скорости при движении по жесткой трубке
жидкости и твердых сфер. Наблюдатель движется вместе со сферами, и потому
они представляются ему неподвижными, а стенки трубки — движущимися.
[Wang, Skalak (1969). Viscous flow in a cylindrical tube containing a line of
spherical particles. J. Fluid Meehan, 38, 88.]
При такой упрощенной геометрии течение может быть описано
аналитически. Мы приведем численные результаты этого анализа
и обсудим их физический смысл. Поскольку инерционными силами
можно пренебречь, основным параметром, определяющим дви-
жение, является отношение радиуса сфер b к радиусу трубки а.
Характер зависимости от этого параметра скорости движения
сфер относительно жидкости можно проиллюстрировать на двух
крайних примерах: когда отношение b/а очень мало и когда оно
равно единице. В первом случае, который физиологически нереа-
лен, сферы не влияют существенно на течение жидкости, которое
является пуазейлевским. Поскольку сферы находятся на осевой
линии сосуда, скорость их перемещения вдвое превышает сред-
нюю скорость движения жидкости. При увеличении отношения
b/а отношение скорости движения частиц к средней скорости по-
тока уменьшается, приближаясь к единице при b/а, стремящемся
к единице. В этом втором предельном случае частицы полностью
перекрывают просвет трубки, жидкость не может проходить ме-
жду ними и стенкой и движется по трубке «порциями». За исклю-
478 Часть II. Механика кровообращения
чением этого предельного случая, частицы всегда движутся бы-
стрее жидкости (что приводит, как было показано выше, к умень-
шению динамического гематокрита). Последнее означает, что по
отношению к частицам жидкость как бы просачивается назад, а
это приводит к возникновению в жидкости, движущейся по сосуду
между частицами, замкнутого движения (рис. 13.35).
Другой важный результат теории заключается в том, что отно-
шение перепада давления, необходимого для проталкивания по
узкой трубке суспензии сфер, к перепаду давления, который необ-
ходим для перемещения с такой же скоростью одной только жид-
кой фазы, растет с увеличением отношения b/а. Например, когда
Ь/а = 0,9 и сферы соприкасаются друг с другом (в этом случае
при заданном отношении b/а падение давления оказывается наи-
большим), отношение перепадов давлений достигает величины,
равной примерно двум. Иными словами, полный перепад давле-
ния, необходимый для проталкивания по трубке жидкости со сфе-
рами, примерно вдвое превышает перепад давления в случае дви-
жения только одной жидкости. Это означает, что кажущаяся вяз-
кость суспензии приблизительно в два раза превышает кажущую-
ся вязкость ее жидкой фазы.
Если перенести эти результаты на движение крови в капилля-
рах, то можно ожидать, что вязкость крови здесь будет примерно
вдвое больше вязкости плазмы. В действительности известно, что
вязкость цельной крови, измеренная вискозиметром с большим
рабочим объемом, т. е. «усредненная» по всей пробе, примерно в
три раза превышает вязкость плазмы (разд. 10.7). Следовательно,
эффект Фореуса — Линдквиста имеет место даже при больших
значениях отношения b/а (вплоть до 0,9) (рис. 13.32).
Отрицательный зазор. В этом случае эритроциты должны про-
ходить по капиллярам, более узким, чем сами клетки. Чтобы
клетка вошла в такую трубку, ее надо сжать — независимо от
того, будет она затем перемещаться или нет. Когда клетка нахо-
дится в состоянии покоя, она «вжимается» в стенку сосуда или
по крайней мере в выстилающий ее мукополисахаридный слой.
При движении же под действием градиента давления клетка
должна скользить по стенке, чему препятствуют силы трения (или
вязкие силы). Повседневный опыт учит, что, коль скоро движение
началось, силы трения между скользящими друг по другу поверх-
ностями можно значительно уменьшить, если ввести между ними
слой жидкости, т. е. смазки. Наиболее убедительная теоретическая
модель движения эритроцитов в узких капиллярах — модель, в ко-
торой вокруг эритроцитов образуется такой смазочный слой. По-
этому мы опишем здесь в общих чертах основные свойства указан-
ной модели и изложим наиболее важные вытекающие из нее фи-
зиологические следствия.
Как уже отмечалось, эритроциты, движущиеся по узким капил-
лярам, принимают самую разную форму, однако какая из них
Гл. 13. Микрососуды большого круга 479
встречается наиболее часто — неизвестно. Анализ движения для
всех этих форм весьма сложен: единственная конфигурация, для
которой разработана приближенная количественная теория, — это
симметричная относительно оси конфигурация парашюта. Опре-
деляющие движение клеток физические принципы во всех случаях
одинаковы, но особенности, например, распределения давления
вокруг эритроцита различны.
Силы упругости деформированной клетки стремятся «вдавить»
ее в стенку сосуда. Поэтому для того, чтобы между клеткой и
стенкой сохранялся зазор, в смазочном слое плазмы должно раз-
виваться достаточно высокое давление. Это одно из трех основ-
ных условий, которые необходимо иметь в виду, объясняя меха-
нику движения в капилляре. Согласно второму условию, жидкость
внутри смазочного слоя должна двигаться таким образом, чтобы
градиент давления и вязкие силы в нем были уравновешены.
Третье условие заключается в том, что действующие на клетку
силы давления должны находиться в равновесии с препятствую-
щими ее перемещению силами вязкости.
Анализируя движение плазмы в смазочном слое и эритроци-
тов, удобно считать, что «наблюдатель» движется с той же ско-
ростью U, что и клетка (рис. 13.36). Тогда клетка покажется ему
неподвижнейшая стенка капилляра — перемещающейся назад со
скоростью U. Такой подход удобен тем, что, только когда мы
мысленно движемся вместе с клеткой, давление и толщина слоя
представляются постоянными во времени.
Рассмотрение течения в смазочном слое показывает, что в слу-
чае отрицательного зазора также должно происходить обратное
движение жидкости относительно эритроцитов (утечка). Если бы
утечки не было, то профиль скорости был бы везде линейным
(рис. 13.36, А) и, поскольку в области X слой жидкости тоньше,
чем в области Y, расход плазмы в области X был бы меньше.
В несжимаемой жидкости это невозможно, и, следовательно, про-
филь скорости не может быть везде линейным. Как показано на
рис. 13.36,5, профиль должен иметь еще одну составляющую, а
это в свою очередь подразумевает существование в слое обрат-
ного, направленного против движения эритроцита (т. е. положи-
тельного) градиента давления. Если Q — это расход плазмы с уче-
том такого отклонения профиля скорости от линейности, а х —
расстояние, измеренное вдоль стенки капилляра слева направо,
то, согласно условию баланса сил в слое жидкости, градиент дав-
ления dp/dx в точке, где толщина слоя равна h, должен быть
равен сумме двух составляющих, одна из которых положительна
и пропорциональна Q/h\ а другая отрицательна и пропорцио-
нальна —и/№. Градиент давления описывается уравнением
dp 12Н<2 6jxC7 (\оо\
dx fts
480 Часть II. Механика кровообращения
Рис. 13.36. Диаграмма, показывающая необходимость обратного движения жид-
кости в капиллярах при прохождении по ним эритроцитов (движение обращено:
клетки покоятся, стенка капилляра движется). Если профиль скорости везде
линеен (Л), то расход жидкости оказывается в разных сечениях неодинаковым,
что невозможно. Наложение параболической составляющей (5) сохраняет нераз-
рывность потока, но требует наличия соответствующего градиента давления.
Рис. 13.37. Приблизительное распределение давления в тонком смазочном слое
плазмы между эритроцитом и стеикой капилляра (Д) и изменение формы эри-
троцита (Б). Штриховая линия па А—приблизительное распределение давления
в сосуде в отсутствие клеток.
Гл. 13. Микрососуды большого круга 481
где |i — вязкость плазмы. К этому уравнению приводит подроб-
ный анализ условий баланса сил, который называется «теорией
смазки».
Теперь можно качественно описать распределение давления
(см. рис. 13.37). Давление в области, значительно удаленной от
клетки вверх по течению, р+, превышает давление в области, уда-
ленной от клетки вниз по течению, р_, так как средний градиент
давления должен, как и в отсутствие клеток, способствовать
движению крови (т. е. быть отрицательным). На значительном
удалении от клетки толщина смазочного слоя увеличивается, и, со-
гласно уравнению (13.2), градиент давления здесь должен быть
отрицательным (поскольку с ростом h величина Q/Л3 убывает зна-
чительно быстрее, чем U/№). Но в той области, где клетка, если
бы она не была деформирована сосудом, имела бы максимальный
диаметр, т. е. вблизи точки X на рис. 13.37, давление должно быть
наибольшим, потому что именно здесь клетка сильнее всего сдав-
ливается. Несколько выше области максимального давления гра-
диент давления положителен, и для согласованйя с отрицатель-
ным градиентом давления на большом удалении от клетки где-то
в промежуточной области (в точке Z) должен быть минимум дав-
ления. Таким образом, распределение давления должно иметь вид,
изображенный на рис. 13.37, А. Из уравнения (13.2) следует, что
минимальной толщине смазбчного слоя соответствует максималь-
ный положительный градиент давления, и потому этот слой должен
быть тоньше всего в точке Y (рис. 13.37,Б), расположенной выше
области наименьшей толщины слоя в отсутствие движения.
Эта качественная картина подтверждается расчетами, осно-
ванными на подробных данных об упругих свойствах эритроцитов.
Кроме того, вычисления привели к выводам, которые в принципе
можно проверить в физиологических экспериментах. Эти выводы
таковы.
1. При данном общем (для капилляра) градиенте давления
расход плазмы с эритроцитами значительно меньше расхода при
течении одной только плазмы (в противоположность случаю с по-
ложительным зазором).
2. Зависимость скорости движения эритроцитов от общего гра-
диента давления нелинейна-, при стремлении градиента давления
к нулю она очень быстро уменьшается (пропорционально квад-
рату градиента давления).
3. При положительном зазоре толщина смазочного слоя отно-
сительно нечувствительна к скорости движения клетки.
4. Если зазор отрицателен, то толщина смазочного слоя со сни-
жением скорости течения (или общего градиента давления) умень-
шается; когда градиент давления стремится к нулю, толщина слоя
уменьшается пропорционально общему градиенту давления.
Из последнего вывода следует, что, когда градиент давления или
скорость течения становятся достаточно малыми, смазывающий
482 Часть II. Механика кровообращения
слой плазмы оказывается настолько тонким (по толщине в нем мо-
жет укладываться всего несколько молекул), что он уже больше не
действует как непрерывный слой жидкости и теряет свои смазоч-
ные свойства. Слой как бы «заедает», и течение останавливается.
Чтобы преодолеть «сухое» трение и возобновить движение, тре-
буется значительно больший градиент давления, чем для поддер-
жания равномерного движения.
В настоящее время подтвердить все эти выводы при помощи
измерений в физиологических экспериментах не представляется
возможным. Известно, что сопротивление движению крови по ка-
пиллярам превышает величину, предсказываемую для ньютонов-
ской жидкости, вязкость которой равна вязкости цельной крови,
измеренной в большом объеме. Это явление противоположно эф-
фекту Фореуса — Линдквиста, наблюдаемому в трубках большего
диаметра, чем диаметр капилляров, и соответствует первому из
сформулированных выше выводов. Кроме того, в модельных экс-
периментах подтверждена нелинейность соотношения между гра-
диентом давления и скоростью перемещения эритроцитов, но фи-
зиологические опыты in vivo дают противоречивые результаты.
В частности, предсказанное явление «заедания» смазочного слоя
в них, по-видимому, не наблюдается. Одно из возможных объяс-
нений этого факта основано на том, что плазме приписывают не-
ньютоновские свойства. Последние могут быть важными, по-
скольку по расчетам «заедание» должно происходить при тол-
щине смазочного слоя меньше 0,5 мкм — величине, сравнимой с
диаметром наиболее крупных белковых молекул плазмы. (Воз-
можно, необходимо учитывать также свойства слоя мукополиса-
харидов, выстилающего поверхность эндотелия капилляров.)
Другое объяснение — несоответствие предположений, сделанных
относительно упругих свойств эритроцитов, действительности.
Отметим, что как высокое давление в смазочном слое, так и
вращательное движение в «порциях» плазмы, заключенных между
эритроцитами, могут оказывать влияние на перенос веществ ме-
жду кровью и стенкой капилляров.
13.8. Массоперенос в системе микрососудов
Довольно долго считали, что кровь и ткани обмениваются ве-
ществами только на уровне капилляров. Однако теперь известно,
что этот процесс протекает не в столь ограниченной зоне: в боль-
шинстве систем микрососудов он осуществляется как в капилля-
рах, так и в посткапиллярных венулах. Поэтому, рассматривая в
данном разделе процессы переноса веществ в кровь и из крови,
протекающие в обоих типах сосудов, мы, не указывая часто, о ка-
ких сосудах идет речь, будем называть их капиллярами. Обсужде-
ние общих вопросов теории массопереноса читатель может найти
в гл. 9.
Гл. 13. Микрососуды большого круга 483
Перенос веществ через стенку капилляров обычно не является
активным. Это утверждение основано на том, что различные яды
(скажем, цианид калия) не оказывают прямого влияния на ско-
рость переноса. Различают два основных механизма переноса ве-
ществ: движение вместе с жидкостью под действием градиента
гидростатического давления (конвективный перенос) и транска-
пиллярный диффузионный обмен молекулами, обусловленный раз-
личием концентраций этих молекул по разные стороны стенки
сосудов. Через стенку проходят молекулы самых разных видов
и размеров, включая молекулы воды, растворенных газов (таких,
как кислород и двуокись углерода), ионы с низким молекуляр-
ным весом (например, Na+ и НСОГ)» а также органические мо-
лекулы. Диапазон размеров органических молекул, если характе-
ризовать его значениями молекулярного веса, простирается от ве-
личин меньше 60 (мочевина) до нескольких миллионов (некото-
рые липопротеиды). Ниже мы рассмотрим различные пути и ме-
ханизмы переноса веществ.
Хотя два основных механизма переноса — конвективный и
диффузионный — взаимно независимы, результирующие скорости
переноса вещества тем и другим механизмом связаны между со-
бой, поскольку разности давлений и концентраций, приводящие
эти механизмы в действие, связаны через осмотические эффекты.
Последнее обусловлено тем, что стенка сосуда аналогична в дан-
ном случае полупроницаемой мембране.
Фильтрация и реабсорбция воды в отдельных капиллярах.
В XIX столетии считалось, что переход воды из крови, в межкле-
точное пространство обусловлен одной лишь разностью гидроста-
тического давления по разные стороны стенки капилляров. Роль
в этом процессе сил всасывания, обусловленных наличием в плаз-
ме белков и противодействующих силам давления, выявил опять-
таки Старлинг — он сделал это открытие в 1896 г. Старлинг пола-
гал, что стенки капилляров проницаемы для воды, электролитов
и очень небольших молекул, находящихся в плазме, но не для
белковых молекул. Следовательно, выход из плазмы воды и элек-
тролитов (т. е. фильтрация их) должен приводить к нарушению
осмотического равновесия и к возникновению противодействующей
фильтрации разности осмотических давлений. Таким образом, если
гидростатическое давление в капиллярах н в ткани обозначить че-
рез рк и рт, а коллоидное осмотическое давление в плазме и в тка-
нях— через Пл и Пт, то полная обусловливающая фильтрацию
сила Дрф будет определяться как
АРФ = (Рк - рт) - (Пп - Пт). (13.3)
Когда величина Дрф положительна (т. е. когда разность гидро-
статических давлений превышает разность осмотических давле-
ний), происходит фильтрация жидкости из капилляров. Когда же
величина Дрф отрицательна, жидкость переходит из межклеточ-
484 Часть II. Механика кровообращения
ного пространства в капилляры. По предположению Старлинга,
в межклеточном пространстве гидростатическое давление рт, а
также коллоидное осмотическое давление Пт всюду одинаковы (не
зависят от координат). Поэтому он считал, что на артериальном
конце капилляра гидростатическое давление в капилляре превы-
шает онкотическое давление в плазме и в результате здесь про-
исходит фильтрация жидкости из сосуда (рис. 13.38). На веноз-
ном конце капилляра гидростатическое давление падает, онкоти-
ческое давление в плазме оказывается выше гидростатического, и
Рис. 1338. Схема типичного капилляра, иллюстрирующая фильтрационно-реаб-
сорбционную гипотезу Старлинга. Гидростатическое давление в капилляре рк
снижается по его длине; приведено значение онкотического давления в капилля-
ре Пп. [Landis, Pappenheimer (1963), Exchange of substances through the capil-
lary walls In: Handbook of physiology, Section 2 Circulation, vol. II (eds Ha-
milton, Dow), pp. 961—1034, American Physiological Society, Washington D. C ]
в результате происходит обратное поступление жидкости (реаб-
сорбция) *). В какой-то точке капилляра отсутствуют как фильт-
рация, так и реабсорбция, поскольку здесь силы, вызывающие эти
противоположные процессы, равны. Кроме того, Старлинг пола-
гал, что в нормальных условиях фильтрация жидкости несколько
превышает ее реабсорбцию, и это следует принимать в расчет,
рассматривая процессы образования и движения лимфы.
Спустя 31 год, в 1927 г., Лендис представил эксперименталь-
ные данные, подтверждающие гипотезу Старлинга. Наблюдая в
микроскоп за отдельными капиллярами брыжейки лягушки, он
пережимал их тонкой стеклянной палочкой (рис. 13.39) и обнару-
жил, что, когда капилляр пережимался вблизи венозного конца,
эритроциты выше области пережатия совершали колебательные
движения в соответствии с ударами сердца и медленно скапли-
*) Как Старлинг, так и последующие исследователи уделяли сравнительно
мало внимания процессу движения жидкости по самому межклеточному про-
странству, хотя именно это движение позволяет осуществляться реабсорбции и
оттоку в лимфатические сосуды. По-видимому, оно сходно с фильтрацией в по-
ристой среде. — Прим, ред.
Гл. 13. Микрососуды большого круга 485
вались вблизи места пережатия. Когда пережимался артериаль-
ный конец, часто удавалось наблюдать, как эритроциты ниже ме-
ста пережатия выходят из капилляра в венулу. Эти наблюдения
толковались как свидетельство справедливости гипотезы Старлин-
га, поскольку при пережатии венозного конца капилляра во всем
капилляре устанавливалось давление, почти равное давлению в
артериоле, и это приводило к фильтрации плазмы. При пережа-
тии артериального конца в капилляре устанавливалось давление,
практически равное давлению в венулах, и обычно в итоге преоб-
ладало поступление в капилляр межклеточной жидкости. Лендис
полагал, что эритроцит плотно «пригнан» к просвету капилляра,
перекрывая его так, что плазма не обтекает клетку. Кроме того,
Рис. 13 39 Определение коэффициента фильтрации (Лгидр = Лпдр/Др) мето-
дом пережатия капилляра При пережатии возникают колебательные движения
эритроцитов в такт сокращениям сердца и происходит постепенное скопление их
вблизи пережимающей палочки За время t эритроцит, находившийся в самом
начале изображенного отрезка капилляра, смещается на расстояние L — L'. Вну-
трений диаметр капилляра d.
он считал, что в поперечном сечении капилляры являются круг-
лыми. Поэтому, измерив скорость движения эритроцитов по капил-
ляру, он мог непосредственно оценить скорости фильтрации и ре-
абсорбции, отнесенные к единице площади стенки капилляра. Эти
величины он сопоставил со средним гидростатическим давлением
в капиллярах, которое измерял, прокалывая их боковые ветви
микропипеткой. Результаты этих исследований представлены на
рис. 13.40. Коэффициент фильтрации Кгидр = ]ГК№/Ьр (расход в
расчете на единицу времени, площади и перепада давлений; см.
разд. 9.6.) определяется здесь как тангенс угла наклона прямой,
проходящей через нанесенные точки; он оказался равным прибли-
зительно 5,6-10~5 мкм3 • мкм-2 • с-1 - Н-1 • м2.
В следующих сериях опытов вызывали повреждение сосудистой
стенки, воздействуя на нее спиртом, создавая условия аноксии
либо прекращая кровоснабжение. Тангенс угла наклона в этом
случае значительно увеличивался, но давление, при котором не
происходит ни фильтрации, ни реабсорбции (т. е. давление, при
котором экспериментальная прямая пересекает ось абсцисс), оста-
валось прежним. Это давление соответствует точке, в которой пе-
репад на стенке капилляра гидростатического давления в точности
486 Часть II. Механика кровообращения
Рис. 13.40. Соотношение между потоком жидкости через стенку капилляра и
давлением в капилляре; измерения производили на отдельных капиллярах бры-
жейки лягушки. Тангенс угла наклона проведенной прямой равен коэффициенту
фильтрации жидкости, который одинаков для процесса фильтрации жидкости
из капилляра и для обратного поступления ее в капилляр. Давление, при кото-
ром эта прямая пересекает ось абсцисс, соответствует эффективному коллоид-
ному осмотическому давлению плазмы. [Landis (1927). Micro-injection studies of
capillary permeability II: The relation between capillary pressure and the rate at
which fluid passes through the walls of single capillaries, Am. J. Physiol., 82,233.]
уравновешивается перепадом коллоидного осмотического давле-
ния. Если в действительности гидростатическое и коллоидное ос-
мотическое давления в ткани близки к нулю, то давление, при
котором экспериментальная прямая пересекает ось абсцисс, соот-
ветствует среднему значению коллоидного осмотического давле-
ния в плазме.
В дальнейшем метод Лендиса был несколько видоизменен, с
тем чтобы применить его для исследования сосудов млекопитаю-
щих, у которых диаметр капилляров меньше, чем у лягушки. В на-
стоящее время коэффициенты фильтрации в капиллярах измерены
Гл. 13. Микрососуды большого круга 487
для многих тканей животных разных видов. Некоторые из этих
данных приведены в табл. 13.3. В ряде случаев измеряли также
коэффициенты фильтрации в венулах, где они оказались больше,
чем в капиллярах. Кроме того, было замечено, что как в скелет-
ных мышцах, так и в брыжейке фильтрация жидкости из плазмы
осуществляется на всем протяжении капилляра, а в брыжейке
кошки она может идти даже в венулах, диаметр которых дости-
гает 20—30 мкм. Таким образом, точка равновесия между фильт-
рацией и реабсорбцией не обязательно должна находиться в сред-
ней области капилляра — она может лежать и в венуле. В бры-
жейке такой точки может вообще не существовать: в этом случае
реабсорбция отсутствует и вся жидкость, фильтрующаяся из плаз-
мы, удаляется через лимфатическую систему.
Таблица 13,3
Коэффициенты фильтрации в капиллярах 2)
Ткань Коэффициент /гидр» МКМ8’МКМ“‘2*С“’,*Н“11*М2
Отдельные капилляры
Брыжейка лягушки Сальник кролика (4 - 16) 10-5
артериальный конец (2 - 8) • 10-5
венозный конец (16 - 25) • Ю-8
М. cremaster крысы Брыжейка кошки3 1 • io-5
капилляры (10 - 30) - io-8
собирательные венулы (30 - 50) • 10-5
Капиллярные русла
Предплечье человека 1 • ю-8
Задняя конечность собаки 2,5 • 10-8
Задняя конечность кошки (3,5 - 5,2) 10-8
Сердпе кролика 8,6 • 10-8
Легкие собаки 0,22 • 10-8
0 Отметим, что при вычислении коэффициентов фильтрации для капиллярного русла
использованы предполагаемые значения площади поверхности капилляров, приходящейся
иа единицу веса тканн
2) По Michel (1972) Flows across the capillary wall. In: Cardiovascular fluid dynamics
ed. Bergel), vol II, Academic Press, London.
3) По Verinder (1974) Pressure and permeability studies in single vessels of mammalian
microvasculature, Ph. D thesis, University of London.
Изменение величины коэффициента фильтрации по ходу мик-
рососудов означает, что гипотезу Старлинга в ее исходной про-
стой форме нельзя использовать для моделирования обменных
процессов сразу во всей сети сосудов, в которых происходит пере-
нос веществ. Тем не менее положение точки равновесия можно
рассчитать из уравнения (13.3), так как в этой точке
Рк (Пп Пт) -f- рт.
488 Часть II. Механика кровообращения
ь А Исходные условия
к 4ОГ
20 -
10 -
0 -
О
£ о
40 р
30 -
Сужение сосудов
Рис. 13.41. Изменение фильтрации и реабсорбции жидкости, вызываемое измене-
нием артериального и венозного давления, гидравлического сопротивления со-
судов и коллоидного осмотического давления плазмы. и рв — давление на
артериальном и венозном концах капилляра; Пп — коллоидное осмотическое дав-
ление плазмы. Штриховые линии указывают исходные уровни фильтрации и
реабсорбции, которые на А представлены сплошными линиями [Friedman (1971).
Microcirculation. In: Physiology (ed. Selkurtt), pp. 259—273, Little Brown and
Co., Boston.]
Коллоидное осмотическое давление в плазме составляет прибли-
зительно 3,0-103 Н-м-2 (30 см вод. ст.), а сумма гидростатиче-
ского и коллоидного осмотического давления в тканях, как было
сказано ранее (разд. 13.5), равна примерно —0,5-103 Н-
• м-2(—5 см вод. ст.). Следовательно, в точке равновесия гидро-
статическое давление в капиллярах должно быть около 2,5-103 Н-
•м~2 (25 см вод. ст.). Это меньше приводившегося ранее
Гл. 13. Микрососуды большого круга 489
(разд. 13.4) типичного значения давления в капиллярах, но соот-
ветствует давлению, которое может существовать в венулах.
Влияние изменения гидростатического давления в капиллярах
на фильтрацию и реабсорбцию иллюстрирует рис. 13.41. На
рис. 13.41, Л отражено исходное состояние. При сужении прека-
пиллярных сосудов (Б) давление в капиллярах снижается, в ре-
зультате чего фильтрация уменьшается, а реабсорбция жидко-
сти из межклеточного пространства увеличивается. Расширение
сосудов (В) оказывает обратное влияние. При повышении веноз-
ного давления (Г) давление в капиллярах также повышается, и
это способствует фильтрации жидкости из них. Влияние дегидра-
тации (приводящей к увеличению коллоидного осмотического дав-
ления в плазме) и гипопротеинемии (снижения коллоидного осмо-
тического давления в плазме) можно видеть из рис. 13.41, Д и Е
соответственно.
Давление в капиллярах и фильтрация воды в препаратах целых
органов. Окклюзионный метод Лендиса можно использовать лишь
для изучения процессов фильтрации и реабсорбции в отдельных
капиллярах плоских сосудистых областей, таких, как сосудистое
русло брыжейки или гп. cremaster. Для сосудистых областей мышц
конечностей и целых органов необходимы другие методы. Одним
из них является изогравиметрический метод, при котором через
сосуды изолированного органа пропускают кровь или плазму и не-
прерывно регистрируют его вес. В стационарных условиях вес
препарата постоянен, что соответствует изогравиметрическому со-
стоянию. Суммарное количество жидкости, переходящей из крови
в ткань и обратно, в этих условиях равно нулю. При изменении
кровотока вес препарата также изменяется, что можно предотвра-
тить, изменив соответствующим образом центральное венозное
давление рв.
Если допустить, что сопротивление движению крови /?в между
капиллярами и центральными венами не зависит от расхода крови
Q, то давление в капиллярах рк можно определить из соотношения
Рк = Рв + RB’ Q- (13.4)
Нанеся на график значения расхода крови, получающиеся в изо-
гравиметрических условиях при различных значениях централь-
ного венозного давления, можно оценить величину рк; она равна
венозному давлению при нулевом расходе (рис. 13.42). Линейность
показанной на этом рисунке зависимости давления от кровотока
подтверждает предположение о том, что сопротивление (Лв) сосу-
дов, расположенных между капиллярами и центральными венами,
постоянно. Представленные на рисунке данные двух опытов, в ко-
торых использовалась плазма с различным коллоидным осмоти-
ческим давлением, подтверждают вытекающее из гипотезы Стар-
линга положение о близости значений гидростатического давления
в капиллярах (рк) и онкотического давления в плазме Пц.
490 Часть П. Механика кровообращения
Расход крови в изогравиметрических условиях, мл . мин-»
Рис. 13.42. Соотношение между расходом крови и давлением в венах изолирован-
ной задней конечности кошки. Представлены две серии данных, полученные при
разных значениях коллоидного осмотического давления плазмы. [Pappenheimer,
Soto-Rivera (1948). Effective osmotic pressure of the plasma proteins and other
quantities associated with the capillary circulation in the hind limb of cats and
dogs, Am. J. Physiol., 152, 479.]
Изогравиметрический метод можно использовать также для
оценки скорости фильтрации в капиллярах сосудистого русла це-
лого органа. Повышение венозного давления при неизменном кро-
вотоке приводит к внезапному резкому возрастанию веса препа-
рата, после чего вес продолжает непрерывно, но более медленно
увеличиваться. Считают, что начальное быстрое увеличение веса
обусловлено заполнением венозных емкостных сосудов, а последую-
щее медленное нарастание — суммарной фильтрацией жидкости из
сосудов в ткани. Скорость фильтрации, отнесенная к единице веса
ткани, пропорциональна величине, на которую давление в капил-
лярах превышает изогравиметрическое давление, рассчитанное из
уравнения (13.4). Наклон кривой вес — время отражает скорость
фильтрации в расчете на единицу веса органа и разности давле-
ний, но не величину коэффициента фильтрации, так как площадь,
через которую осуществляется фильтрация, остается неизвестной.
Измерения скорости фильтрации произведены для целого ряда
тканей; результаты некоторых из них представлены в табл. 13.4.
Из нее видно, что через окончательный эндотелий капилляров ки-
щечника фильтрация осуществляется значительно легче, чем через
Гл. 13. Микрососуды большого круга 491
Таблица 13.4
Скорость фильтрации в капиллярах с эндотелием разных типов
Объект исследования Тип эндотелия Скорость фильтрации, Г«с“^Н^-м2 иа 1 кг ткани
Задняя конечность
КОШКИ Непрерывный 18•10~4
крысы »» 40 • 10—4
Сердце кролика » 30 • 10“4
Легкие собаки и 2 • 10“4
Кишечник кошки Окоичатый 2 • 10—2
Почка кошки И 20•10“2
Ч Данные взяты из источников, указанных в справочнике Altman, Dittmer (ed ) (1971).
Biological handbooks. Circulation and respiration, pp. 502—503, Federation of American
Societies of Experimental Bicloqy.
непрерывный эндотелий стенки капилляров скелетных мышц, хотя
плотность капилляров на единицу веса в кишечнике меньше, чем
в мышцах. Коэффициент капиллярной фильтрации для конкрет-
ного препарата можно рассчитать, если известна отнесенная к еди-
нице веса ткани площадь поверхности капилляров. Некоторые
оценки значений коэффициента фильтрации в разных тканях при-
ведены в табл. 13.3. Вычисленные таким образом коэффициенты
всегда оказываются значительно меньше, чем полученные более
прямыми методами для отдельных капилляров. Однако этому, ве-
роятно, не стоит удивляться, поскольку посмертные морфо-
метрические подсчеты площади поверхности капилляров мало что
говорят о соотношении числа закрытых и функционирующих ка-
пилляров в условиях, более близких к физиологическим. Всякое
завышение данных о площади поверхности капилляров приводит
к снижению расчетного коэффициента фильтрации. С другой сто-
роны, исследования на одиночных капиллярах сопряжены с хи-
рургическим обнажением ткани, введением канюль и нагреванием
при освещении изучаемого под микроскопом объекта. Все эти воз-
действия могут увеличивать проницаемость капилляров.
Зависимость онкотического давления плазмы от концентрации
белков. Как следует из уравнения (13.3),скорость фильтрации во-
ды из плазмы зависит как от гидростатического, так и от осмоти-
ческого давления в плазме и в межклеточном пространстве. По
мере выхода из плазмы воды и небольших молекул концентрация
белков в ней увеличивается. Возникающее в результате этого по-
вышение онкотического давления плазмы можно определить, обра-
тившись к закону Вант-Гоффа [уравнение (9.13)], согласно кото-
рому для разбавленных растворов зависимость между осмотиче-
ским давлением П и молярной концентрацией с имеет следующий
вид:
П = с/?Т.
492 Часть II. Механика кровообращения
Риа. 13.43. Зависимость осмотического давления плазмы от концентрации в ней
белков и осмотического давления растворов альбумина и у-глобулина от их кон-
центрации. Штриховая линия — теоретическая зависимость осмотического давле-
ния раствора альбумина от его концентрации, получаемая из закона Вант-Гоффа.
[Landis, Pappenheimer (1963) Exchange of substances through the capillary walls.
In: Handbook of physiology. Section 2: Circulation vol. II (eds. Hamilton and
Dow), pp. 961—1034, American Physiological Society, Washington D. C.]
Здесь T — абсолютная температура, R — универсальная газовая
постоянная. Однако такой расчет дает заниженное значение при-
роста осмотического давления, так как при увеличении концент-
рации белков с в плазме осмотическое давление в ней растет не-
пропорционально с. Это можно видеть из рис. 13.43, где представ-
лена зависимость онкотического давления плазмы от концентра-
ции белков. Здесь же приведена зависимость осмотического давле-
ния раствора альбумина от его концентрации и такая же зависи-
мость, рассчитанная из уравнения Вант-Гоффа для разбавленных
растворов. Причины такого поведения плазмы весьма сложны.
Частично оно определяется иммобилизацией воды крупными мо-
лекулами, а также электростатическим взаимодействием молекул.
Доказательство существования в стенке капилляров фильтрую-
щих пор. При данных значениях перепада гидростатического или
Гл. 13. Микрососуды большого круга 493
осмотического давления перенос воды и водорастворимых мелких
молекул через стенку капилляра оказывается значительно больше,
чем через мембрану эритроцитов. Поэтому уже давно предпола-
гали, что такие вещества проходят сквозь стенку капилляров че-
рез узкие поры. Представление о том, что нерастворимые в жирах
вещества могут фильтроваться или диффундировать только через
очень небольшую часть стенки капилляров, также предполагает
существование особых заполненных водой пор или щелей, прони-
зывающих выстилающий сосуды эндотелий.
Ранее (разд. 13.2) было отмечено, что между эндотелиальными
клетками имеются длинные щели, ширина которых, за исключе-
нием узких зон, где соседние мембраны почти сливаются друг с
другом, составляет примерно 15—20 нм. Как показывают элект-
ронно-микроскопические исследования, когда зона слияния яв-
ляется прерывистой (по типу macula occludens), между соседними
областями слияния имеются овальные зазоры, или щели, шириной
всего около 8 нм. Такие щели, следовательно, создают наибольшее
ограничение фильтрации и диффузии между клетками. Кроме того,
отмечалось, что капилляры с окончатым эндотелием могут иметь
строение, обеспечивающее эффективную диффузию через круглые
оконца, а капилляры с прерывистым эндотелием имеют большие
межклеточные щели, которые, очевидно, оказывают переносу ве-
ществ очень низкое сопротивление.
Считают, что фильтрация воды и небольших водорастворимых
молекул осуществляется в основном именно через межклеточные
щели.
Диффузия через стенку капилляров. Фильтрация и реабсорбция
могут осуществляться только через заполненные водой щели в
•стенке капилляров. Но диффузия молекул в принципе возможна
и через мембрану эндотелиальных клеток. Так могут диффунди-
ровать, вероятно, вещества, хорошо растворимые в жирах, напри-
мер дыхательные газы. По-видимому, их молекулы диффундируют
в сущности через всю поверхность мембраны.
Однако более крупные молекулы, а также молекулы веществ,
нерастворимых в жирах, вероятно, не способны столь свободно
диффундировать через эндотелий. Тем не менее для таких веществ
стенка капилляров значительно более проницаема, чем мембрана
эритроцитов. Кроме того, через стенку капилляров хотя и плохо,
но все-таки проходят высокомолекулярные вещества, скажем аль-
бумин.
Напомним, что в разбавленных растворах скорость диффузии
J химически свободного вещества определяется, согласно закону
Фика, соотношением
/=-ол#' ' <13-6>
где Ас/Ах— градиент концентрации вещества в направлении пе-
реноса, А — площадь, через которую осуществляется диффузия,
494 Часть П. Механика кровообращения
D — коэффициент диффузии. Это уравнение можно переписать в
более удобном виде (разд. 9.5):
/ = Р-ДмДс, (13.6)
где Р = D/Ах. Такая запись удобнее потому, что невозможно оп-
ределить точное расстояние, на котором изменяется концентрация
вещества. Здесь Дм — площадь поверхности мембраны, Р называют
коэффициентом проницаемости или просто проницаемостью.
Для ряда жиронерастворимых молекул определены коэффи-
циенты проницаемости капилляров скелетных мышц. Некоторые
данные такого рода приведены в табл. 13.5. Из таблицы видно,
что с увеличением молекулярного веса веществ и радиуса молекул
проницаемость значительно уменьшается. Снижение проницаемо-
сти естественно, так как при увеличении молекулярного веса
уменьшается коэффициент диффузии. Но из табл. 13.5 видно так-
же, что с увеличением молекулярного веса уменьшается и отноше-
ние проницаемости к коэффициенту диффузии. Это позволяет вы-
сказать предположение об уменьшении площади, через которую
может осуществляться диффузия, об увеличении длины диффу-
зионного пути или об увеличении сопротивления диффузии по ка-
ким-либо другим причинам. Если длина диффузионного пути при-
мерно соответствует толщине эндотелиальных клеток и не зависит
от размера молекул, то эффективная площадь диффузии жироне-
растворимых веществ существенно меньше действительной пло-
щади поверхности капиллярной стенки.
Таблица 13.5
Проницаемость капилляров скелетной мышцы для жиронерастворимых
соединений 4
Соединение Молекуляр- ный вес Радиус, им Р х ю’> см-с—1 D X 10е. см2 с*-1 (₽/£>) X Ю3. см-1
Н2О 18 0,15 0,54 34 17,2
Мочевина 60 0,26 0,26 19,5 13,5
Глюкоза 18Q 0,37 0,090 9,0 10,0
Сахароза 342 0,48 0,050 7.0 6.9
Рафиноза 504 0,57 0,039 6,4 6,0
Инулин 5500 1,30 0,005 2,5 2,6
Миоглобин 17000 1,90 0,0004 1,7 0,3
Сывороточный альбумин 67000 3,60 <ю-в 0,85 0
О Данные взяты из следующих источников.
Landis, Pappenhelmer (1963). Exchange of substances through the capillary walls. In Handbook
of physiology (ed. Hamilton), Section 2, vol. II, American Physiological Society, Washington
D. C. Renkin (1959). Capillary permeability and transcaplflary exchange In relation to molecu-
lar size. In: The microcirculation: factors Influencing exchange of substances across capillary
wall. University of Illinois Press.
Обозначив площадь мембраны, через которую на самом деле
может осуществляться перенос данного вещества, символом А,
Гл. 13. Микрососуды большого круга 495
уравнение (13.6) можно переписать в виде
/ = (РЛ)-Дс,
или
РА=^- (13.7)
В большинстве исследований по экспериментально получен-
ным значениям I и Дс определяют в сущности именно произведе-
ние РА для различных молекул. Если для двух типов молекул раз-
ного размера одинакова площадь мембраны,через которую может
осуществляться их диффузия, то сравнение величин РА прямо ука-
зывает на соотношение проницаемостей мембраны для этих моле-
кул. Однако, как будет показано дальше, такое предположение
не всегда выполняется.
Методы измерения коэффициентов проницаемости. Для измере-
ния проницаемости стенки сосудов разработано несколько методов.
Все они очень изящны, но сопряжены с различными эксперимен-
тальными и расчетными трудностями. Результаты произведенных
разными методами измерений проницаемости в одной и той же
ткани для молекул одного и того же вещества заметно различают-
ся. Существенно различаются и величины, полученные для различ-
ных тканей И хотя это расхождение может отражать истинное
различие свойств эндотелиальных клеток, выстилающих сосуды
этих тканей, полагают, что оно обусловлено также и недостат-
ками методов измерения.
Чтобы определить проницаемость сосуда, необходимо знать не
только общее количество жидкости, переходящей из крови в ткань,
но и разность концентраций вещества по разные стороны стенки
сосуда. Поскольку перенос вещества из крови в ткань и движение
крови по капилляру происходят одновременно, разность концентра-
ций по разные стороны стенки капилляра и количество жидкости,
проходящей сквозь нее, изменяются вдоль капилляра. В каждой
точке капилляра эти величины измерить невозможно, поэтому при-
ходится оценивать разность концентраций, усредненную по всей
длине сосуда. В свою очередь выбор соответствующей средней ве-
личины зависит от сложности модели, описывающей процессы об-
мена между кровью и тканью.
Кроме того, необходимо принимать во внимание, что в неко-
торых условиях значительное влияние на процессы массопереноса
может оказывать скорость кровотока как таковая, и его нельзя
путать с влиянием, обусловленным изменением проницаемости
стенки. Рассмотрим случай, когда диффундирующие из крови в
ткань молекулы вводятся в артериальную кровь, подходящую к
капилляру. По мере продвижения крови по капилляру концентра-
ция в ней этих молекул-меток будет постепенно падать, как это
показано на рис. 13 44. Если проницаемость стенки для данных
молекул высока, а способность ее накапливать такие молекулы
496 Часть II Механика кровообращения
Рис. 13.44. Снижение по мере прохождения крови по капилляру концентрации в
ней трех веществ. Стенка капилляра наиболее проницаема для вещества I и наи-
менее проницаема для вещества III. В случаях I и II равновесие между кровью
и тканями достигается раньше, чем кровь покидает капилляр; это соответствует
условиям «ограниченного потоком» массопереноса. В случае III равновесие не
достигается, и такой массоперенос называют «диффузионно ограниченным». [Mi-
chel (1672). Flow acrpss the capillary wall. In: Cardiovascular fluid dynamics
(ed. Bergel), vol. II, Academic Press, London ]
низкая, то концентрация сначала будет быстро спадать (кривая/),
а затем, по достижении равновесия, по ходу капилляра меняться
не будет. Кривая // относится к случаю, когда равновесная кон-
центрация молекул в крови и в ткани устанавливается как раз на
конце капилляра, а кривая /// — к случаю, когда длина капилляра
слишком мала, чтобы такое равновесие могло установиться. По-
нятно, что, поскольку скорость уменьшения количества молекул-
меток в крови равна произведению расхода крови на разность кон-
центраций этих молекул на артериальном и венозном концах ка-
пилляра (са — Св), в случаях / и II количество перешедших из ка-
пилляра в ткань молекул одинаково и больше, чем в случае III.
Если кровоток снизить, то в случаях I и II определенная выше
скорость перехода молекул из крови в ткань уменьшится прямо
пропорционально снижению кровотока. В случае /// эта скорость
тоже уменьшится, но в значительно меньшей степени, так как уве-
личение времени пребывания крови в капилляре, а соответственно
и времени, в течение которого молекулы могут переходить из кро-
ви в ткань, приведет к увеличению артерио-венозной разности кон-
центраций. Повышение кровотока приводит к увеличению скоро-
сти перехода молекул из крови в ткань во всех случаях, но в наи-
большей степени это сказывается в случае /, а в наименьшей-—
Гл. 13. Микрососуды большого круга 497
в случае III. Такие различия зависимости от кровотока скорости
перехода веществ из крови в ткань привели к понятиям «ограни-
ченного потоком» и «диффузионно ограниченного» массопереноса.
Поскольку в случаях / и II перенос можно увеличить, лишь повы-
сив кровоток, их называют ограниченными потоком. В случае ///
диффузионное равновесие не достигается, и потому такой случай
называют диффузионно ограниченным.
При изучении проницаемости необходимо производить измере-
ния при разных значениях кровотока, чтобы убедиться в том, что
переход исследуемого растворенного вещества в ткани не ограни-
чен потоком в указанном выше смысле. В случаях I и II артерио-
венозная разность концентраций одинакова, и любые вычисления
средней по длине капилляра разности концентраций по обе сто-
роны стенки сосуда, использующие величину артерио-венозной
разности концентраций, в обоих случаях приведут к одинаковому
результату, т. е. дадут одно и то же значение проницаемости. Ме-
жду тем очевидно, что в случае / проницаемость стенки выше, чем
в случае II.
Неопределенности, связанные с ограничением массопереноса
потоком, привели к тому, что большинство исследований проницае-
мости относится к жиронерастворимым молекулам, а определить
проницаемость для мелких молекул веществ, хорошо растворимых
в жирах, оказалось невозможно.
Применение в физиологических исследованиях радиоактивных
изотопов позволило использовать для измерения произведения
проницаемости на эффективную площадь весьма изящный метод
разведения индикатора. Этот метод заключается в том, что радио-
активные молекулы, которые заведомо остаются в крови, и мече-
ные другим способом молекулы, способные проходить через стенку
капилляров, вводят вместе одной небольшой порцией в мелкую
артерию и анализируют пробы крови, оттекающей по венам ис-
следуемого капиллярного русла. Сравнение отношения концент-
рации изотопов в пробах венозной крови и во введенной смеси
показывает, какая доля обмениваемых молекул-меток прошла
через стенки капилляров. Результаты эксперимента, в котором
определяли проницаемость стенок для сахарозы, а в качестве не-
обмениваемой внутрисосудистой метки использовали альбумин,
представлены на рис. 13.45. Концентрация сахарозы в оттекаю-
щей крови растет медленнее и достигает меньшей величины, чем
концентрация альбумина, так как при прохождении крови через
капилляры часть молекул сахарозы переходит во внесосудистое
пространство. Через некоторое время после достижения макси-
мальных значений концентраций кривые пересекаются. Это обу-
словлено тем, что большая часть альбумина к этому времени уже
покидает капилляры, а концентрация сахарозы в венозной крови
поддерживается относительно высокой, поскольку теперь саха-
роза поступает в капилляры из тканей, где ее концентрация выше,
498 Часть II. Механика кровообращения
Время, с
Рис. 13 45 Относительные концентрации в венозной крови меченых альбумина
и сахарозы после одновременного введения их в артерию. Уровни радиоактив-
ности отнесены к максимальной радиоактивности альбумина.
Если предположить, что первоначально в каждой точке капил-
ляра количество молекул, переходящих из крови в ткань, пропор-
ционально их локальной концентрации в крови (концентрация ме-
ченой сахарозы в тканях исходно равна нулю), то концентрация
сахарозы в крови должна падать по ходу капилляра экспонен-
циально. Можно показать, что в этом случае средняя концентра-
ция ее в капиллярах с определяется соотношением
С = (св — с«)/1п
Гл. 13 Микрососуды большого круга 499
где са и св — концентрация сахарозы на артериальном и венозном
концах капиллярного русла соответственно. Значения этих кон-
центраций можно получить прямо из максимумов кривых на
рис. 13.45: максимум кривой для альбумина дает концентрацию
сахарозы на артериальном конце капилляров, максимум кривой
для сахарозы — ее концентрацию на венозном конце. Поскольку
начальная концентрация меченой сахарозы в ткани равна нулю,
разность Ас средних концентраций по обе стороны стенок сосудов
равна с. Скорость перехода сахарозы из крови в ткань опреде-
ляется соотношением
J = Q (са — Св),
где Q — кровоток в капиллярном русле. Подстановка Ас = с и вы-
ражения для J в уравнение (13.7) дает
РА = . . = Q In (ca/cB).
(са — св)/1п (Са/Св) ' а' в/
Значительные преимущества метода разведения индикатора
заключаются в его простоте, а также в том, что он позволяет изу-
чать проницаемость в естественных условиях деятельности сер-
дечно-сосудистой системы. Вместе с тем применение этого метода
связано с введением весьма сильного допущения: предполагается,
что все сосуды исследуемого русла имеют одинаковую проницае-
мость и отношение площади поверхности, через которую осуществ-
ляется массоперенос, к кровотоку Q для всех сосудов также оди-
наково. Однако даже простое наблюдение за функционирующим
капиллярным руслом заставляет усомниться в справедливости
этого предположения, а любое отклонение от однородности пара-
метров по сосудистому руслу ведет к тому, что вычисленное зна-
чение проницаемости оказывается заниженным; чтобы преодолеть
эту трудность, проницаемость измеряют при возможно более вы-
соких скоростях кровотока Предполагается, кроме того, что моле-
кулы-метки обоих типов распределяются в капиллярном русле
одинаково. Хотя на первый взгляд это кажется вполне правдопо-
добным, в действительности такая ситуация вовсе не обязательна.
Дисперсия молекул в движущейся крови зависит в какой-то мере
от их диффузионной способности, и, поскольку молекулы двух ука-
занных типов отличаются друг от друга по размеру, следует ожи-
дать, что вследствие дисперсии по Тэйлору (разд. 12.9) распреде-
ление их по сосудам будет несколько различаться.
Другой подход к определению проницаемости заключается в
сравнении концентрации тех или иных молекул в плазме и лимфе.
Как отмечалось в данной главе, в стационарном состоянии боль-
шая часть воды и малых молекул, вышедших из крови на арте-
риальном конце капилляров, возвращается на их венозном конце
в кровь, но оставшаяся часть фильтрующихся веществ покидает
межклеточное пространство через лимфатические сосуды. Концент-
БОО Часть II. Механика кровообращения
рация электролитов в плазме и в лимфе очень близка, но кон-
центрация белков различна: в плазме она больше, чем в лимфе.
Кроме того, как показано в табл. 13.6, в плазме и в лимфе раз-
личны и относительные концентрации разных белков. Глобулин,
молекулы которого значительно крупнее, чем молекулы альбу-
мина, по-видимому, с большим трудом проходит через стенку ка-
пилляров одних органов, чем других.
Таблица 13.6
Общее содержание белков и отношение концентраций альбумина
и глобулина в плазме и в лимфе,
оттекающей от разных областей тела1'
Общее содержание Отношение концентраций
белков, г на 100 см3 альбумина и глобулина
Плазма 6,3 1,43
Лимоа из грудного протока 4,0 1,50
Лнмоа из скелетных мышц 2,3 1,54
Лимоа из кишечника 3,8 1,50
Лимфа нз печени 5,3 1,48
По Courtice (1972). The chemistry of lymph. In: Lymph vessel system (ed Heessen),
Springer Verlag, Berlin and Heidelberg.
Молекулы известного размера вводят в вену и следят за кон-
центрацией их в плазме и в лимфе. (При использовании радиоак-
тивных молекул были предприняты попытки измерять концентра-
цию вещества-метки непосредственно в межклеточном простран-
стве.) В стационарном состоянии суммарная скорость удаления
через стенку капилляров растворенного в крови вещества равна
скорости оттока его через лимфатические сосуды. Выход молекул
из капилляров обусловлен как фильтрацией, так и диффузией. Од-
нако исследование проницаемости для относительно крупных мо-
лекул показало, что концентрация их в лимфе сл значительно
меньше, чем в плазме. Поэтому можно предположить, что сум-
марная фильтрация их очень мала вследствие высокого сопротив-
ления прохождению крупных молекул через поры, и, следователь-
но, переход их через стенку капилляров осуществляется в основ-
ном за счет диффузии. В таком случае
==: (^^4) (^-п £л)>
где 7л — объемная скорость течения лимфы, сп — концентрация мо-
лекул-меток в плазме, РА — произведение (для этих молекул) про-
ницаемости на площадь, определенное в соответствии с уравне-
нием (13.7). Использование в качестве средней разности концент-
раций по обе стороны стенки капилляра величины (сп — сл) осно-
вано, во-первых, на том, что скорость суммарного обмейа в этом
случае очень низка, и потому артерио-венозная разность концепт-
Гл. IS. Микрососуды большого круга 501
Рис. 13.46. Произведение проницаемости капилляров иа площадь диффузии для
молекул поливинилпирролидона разного размера, определенное по отношению их
концентраций в плазме и в лимфе и по расходу лимфы. Обратите внимание, что
при низких значениях молекулярного веса проницаемость по мере повышения по-
следнего снижается, а при больших его значениях становится постоянной. [Ren-
kin (1964). Transport of large molecules across capillary walls, Physiologist, 7,
20.]
раций очень мала по сравнению с их абсолютными значениями, а
во-вторых, на допущении, что сопротивление диффузии оказывает
только стенка капилляра, и концентрация молекул в’ тканевой
жидкости и в лимфе одинакова.
Таким способом измерена проницаемость стенки капилляров
для целого ряда веществ, в частности для полисахаридов (напри-
мер, для декстранов) с различными молекулярными весами и для
полимера поливинилпирролидона, мол. вес которого лежит в диа-
пазоне от 10000 до 200000 и выше. Оказалось, что при изменении
молекулярного веса произведение проницаемости на площадь ме-
няется так, как показано на рис. 13.46. В области низких значений
молекулярного веса проницаемость снижается с его ростом; в об-
ласти же более высоких значений проницаемость перестает за-
висеть от размера молекул. Это позволяет предположить, что ме-
ханизмы прохождения через мембраны капилляров крупных и
мелких молекул различны.
Рассмотрим теперь три основных предположения, лежащих в
основе этого метода определения проницаемости капилляров. 1.
Предполагается, что доля молекул, проходящих через стенку ка-
пилляра посредством фильтрации, от общего количества вышед-
ших из крови молекул пренебрежимо мала. Это предположение,
по-видимому, выполняется, поскольку при повышении скорости
оттока лимфы измеренное значение проницаемости увеличивается
лишь незначительно. 2. Разность концентраций молекул в области,
602 Часть И. Механика кровообращения
непосредственно примыкающей к наружной стенке капилляра, и
в лимфатических капиллярах также предполагается пренебрежимо
малой. Хотя для относительно крупных молекул эта разность мо-
жет несколько отличаться от нуля, для более мелких молекул
она, по-видимому, ничтожно мала, так что это предположение, ве-
роятно, вполне приемлемо. 3. Третье предположение заключается
в том, что концентрация молекул в лимфе отражает концентрацию
их в тканевой жидкости, и это весьма сомнительно. Если прони-
цаемость различных капилляров рассматриваемого сосудистого
русла не вполне одинакова, то лимфа должна состоять преиму-
щественно из воды и молекул, вышедших из наиболее проницае-
мых капилляров, а локальные концентрации белка в. тканевой
жидкости должны быть относительно высокими. Таким образом,
состав тканевой жидкости будет характеризоваться некоторым
«спектром», а состав оттекающей лимфы будет отражать средний
состав ультрафильтратов из всех капилляров.
В одном из наиболее ранних методов определения проницае-
мости использовали изолированный препарат задней конечности.
В жидкость, поступающую в капиллярное русло, добавляли моле-
кулы растворимого вещества, что вызывало переход воды из тка-
ней конечности в плазму. Для предотвращения этого перехода
увеличивали давление в капиллярах, повышая венозное давление
до тех пор, пока вес конечности не возвращался к исходному
уровню, на котором и поддерживался. Необходимый для этого
прирост давления в капиллярах равен разности осмотических дав-
лений по обе стороны их стенки, обусловленной присутствием до-
бавленного вещества. Скорость перехода J этого вещества через
стенку капилляра оценивали по произведению артерио-венозной
разности концентраций на кровоток. Среднюю разность концентра-
ций по обе стороны сосудистой стенки оценивали по изменению
осмотического давления АП, используя закон Вант-Гоффа [урав-
нение (9.14)]. Подставив соответствующие величины в уравнение
(13.7), получим
рл=/(дпда.
Этот метод кажется очень привлекательным, но основным его
недостатком является введение предположения, что среднюю раз-
ность концентраций по обе стороны стенки капилляра можно рас-
считать непосредственно из закона Вант-Гоффа. В действительно-
сти этот закон для многих крупных молекул не выполняется при
всех концентрациях, исключая очень низкие (стр. 491). Кроме
того, он предполагает, что мембрана функционирует как идеаль-
ный полупроницаемый барьер, препятствующий проникновению че-
рез него любых молекул растворенных веществ. Тот факт, что
мембрана лишь ограничивает, но не предотвращает выход молекул
из сосудов, приводит к снижению осмотического давления
(разд. 9.7).
Гл. 13. Микрососуды большого круга 503
Величина коэффициента осмотического отражения о [уравне-
ние (9.15)] зависит как от гидродинамических, так и от диффу-
зионных факторов, оказывающих влияние на движение молекул
в каналах мембраны, но теории, которая могла бы удовлетвори-
тельно оценить значимость этих влияний, в настоящее время не
существует. Оценка коэффициентов отражения является одним из
наиболее важных вопросов современной физики мембран.
Полагают, что величина о зависит от отношения коэффициен-
тов ограниченной диффузии растворенного вещества (£>i) и раство-
рителя (Дг) следующим образом:
Эффективный коэффициент диффузии можно определить как
Д = DAKaxJAncT, (13.9)
где Д — коэффициент свободной диффузии, Дкаж и ДИст — кажу-
щаяся и истинная площади, через которые осуществляется диффу-
зия. Объединяя уравнения (13.8) и (13.9), получаем
’“('-^Г-к)- <l3J0>
где Лкаж, 1 и Лкаж, 2 — кажущиеся площади диффузии соответствен-
но для растворенного вещества и растворителя; истинные пло-
щади, через которые могут диффундировать растворенное веще-
ство и растворитель, предполагаются одинаковыми. В настоящее
время считают, что в действительности площади участков мембра-
ны, через которые осуществляется перенос воды и большинства
жиронерастворимых молекул, вероятно, не одинаковы, и потому
уравнение (13.10) нельзя считать удовлетворительным представле-
нием для коэффициента осмотического отражения.
Путь диффузии через стенку капилляров. Для установления
пути диффузии через стенку капилляра предпринимались иссле-
дования двоякого рода. В одном случае в просвет капилляра вво-
дили нерастворимые в жирах молекулы известного размера, ко-
торые можно обнаружить при помощи электронного микроскопа,
и позволяли им диффундировать через стенку капилляра. Затем
ткань фиксировали, приготовляли препараты для микроскопиче-
ского изучения и определяли локализацию и концентрацию этих
молекул в стенке и в межклеточном пространстве. Второй подход
использует описанные выше исследования проницаемости как ос-
нову для суждения о существовании пор определенного размера
и затем устанавливает связь размеров этих пор с теми структу-
рами стенки, которые удается идентифицировать при помощи
электронного микроскопа,
504 Часть II. Механика кровообращения
Показано, что в случае капилляров, имеющих эндотелий непре-
рывного типа, молекулы средней величины (пероксидаза хрена,
мол. вес около 40000) проникают во внесосудистое пространство
путем пиноцитоза и не обнаруживаются в больших количествах в
межклеточных щелях. Более мелкие молекулы (цитохромперок-
сидаза, мол. вес около 12000) выходят из капилляров быстрее; эти
молекулы обнаруживают в межклеточных щелях, и концентрация
их здесь уменьшается с увеличением расстояния от поверхности,
обращенной к просвету. Основываясь на этих и других аналогич-
ных наблюдениях, полагают, что межклеточные щели способны
пропускать относительно мелкие молекулы, но препятствуют диф-
фузии крупных.
Результаты таких исследований согласуются с представленной
на рис. 13.46 зависимостью проницаемости от молекулярного веса
соединений. При больших значениях молекулярного веса (превы-
шающих примерно 40000) проницаемость в широких пределах не
зависит от размера молекул.
Теория эквивалентных пор, разработанная Паппенхаймером.
Всюду в этой главе мы неизменно полагали, что фильтрация и
диффузия сквозь стенку капилляров осуществляются через поры.
Эта концепция была выдвинута довольно давно, но только в
1951 г. Паппенхаймер и его сотрудники предложили количествен-
ную модель диффузии молекул через поры, размеры которых срав-
нимы с размерами диффундирующих молекул. Эта теория дает
очень удобное описание потока через мембрану, и с ее помощью
можно получить точные предсказания относительно строения
мембраны.
Теория Паппенхаймера позволяет оценить величину коэффи-
циента ограниченной диффузии по отношению к коэффициенту
свободной диффузии исходя из кажущейся и истинной площадей
пор, как это определено уравнением (13.9). Предполагается, что
причиной уменьшения эффективного коэффициента диффузии
(в сравнении с коэффициентом свободной диффузии) является
уменьшение эффективной площади пор, обусловленное различием
формы пор и молекул, препятствующим входу последних в поры,
а также большее гидродинамическое сопротивление движению
молекул через ограниченное пространство, чем через неограничен-
ную среду.
Так, например, если молекула радиуса а входит в пору с ра-
диусом г, не касаясь ее стенок, то истинная площадь диффузии
Дист равна п(г— а)2, или
= = (13.11)
Оценить увеличение сопротивления' движению молекул в узких
порах очень сложно, так как оно зависит не только от относи-
тельных размеров, но и от формы молекулы, от распределения
Гл. 13. Микрососуды большого круга 505
заряда на ее поверхности, от ее гибкости и от многих других фак-
торов. Некоторые из этих обстоятельств (например, влияние фор-
мы и гибкости) частично принимаются во внимание при опреде-
лении эквивалентного радиуса молекул исходя из формулы Сток-
са — Эйнштейна (9.6). Если обусловленное трением сопротивление
свободному движению обозначить через f, а сопротивление при
прохождении сквозь пору — через /огр, то с хорошей точностью вы-
полняется следующее соотношение:
4 = 1 - 2,10(^)+2,09(4)’ - O.9S(^)’. (13.12)
Согласно уравнению Нернста (9.4),
^огр f
(13.13)
Подставив (13.11), (13.12) и (13.13) в уравнение (13.5), получим
1----Dff (1 -Ж1 -2-‘° (т) +2’09 (Я-°-95 (т)1-
(13.14)
Таким образом, если для некоторого физиологического объекта
измерено произведение площади на проницаемость [уравнение
13.7)] для определенного ряда молекул различных размеров, то,
используя уравнение (13.14), можно оценить эквивалентные раз-
меры пор, через которые осуществляется диффузия.
На рис. 13.47 результаты вычислений, основанных на теории
пор, сопоставлены с экспериментальными данными, полученными
на препарате задней конечности. Как видно из рисунка, с увели-
чением размера молекул проницаемость падает, причем характер
этого падения очень близок к предсказанному по уравнению (13.14)
в предположении, что поры круглые и имеют радиус 4 нм.
Такое согласие эксперимента и теории очень обнадеживает,
особенно если учесть, что ширина межклеточных щелей, как пока-
зывают электронно-микроскопические исследования, имеет как раз
такой порядок. Однако столь близкое соответствие получается не
всегда, и, чтобы прийти к четким выводам о структуре диффузион-
ного пути в различных тканях, необходимы дальнейшие обстоя-
тельные исследования.
Путь переноса воды через стенку капилляров. Построенная
Паппенхаймером теория пор позволяет оценить их размер в стен-
ках капилляров исходя из данных о скорости фильтрации воды.
Если поры имеют радиус порядка 4 нм, то их радиус значительно
больше радиуса молекул воды, и, следовательно, есть основания
считать, что течение воды через такие поры можно описать при
помощи формулы Пуазейля [уравнение (5.1)]. Справедливость
этого предположения подтверждается результатами исследования
506 Часть II. Механика кровообращения
Рис. 13 47. Проницае-
мость капилляров задней
конечности кошки и со-
баки для молекул раз-
личного радиуса а. Экс-
периментальные точки
ложатся на кривую,
предсказываемую теори-
ей для ограниченной
диффузии молекул через
поры радиусом 4 нм.
[Michel (1972). Flow
across the capillary wall.
In: Cardiovascular fluid
dynamics (ed. Bergel),
vol. II, Academic Press,
London.]
переноса воды через искусственные мембраны с порами таких раз-
меров. Если при эффективной обусловливающей фильтрацию силе
(Др — ДП) [уравнение (13.3)] скорость фильтрации равна Л-идр, то
f __ пПг< (Др - ДП)
•'гидр— 8fi • дх
где п — число пор радиуса г в мембране толщиной Дх. Если Ав —
площадь пор, через которую могут проходить молекулы воды, то
Ав = ппг2
и
4идр = -gjj- д^ (ДР — ДП),
ИЛИ
. (13.15)
Дх г2 (Др — ДП) ' '
Таким образом, зная величину радиуса пор (ее определяют по ре-
зультатам измерения коэффициента фильтрации воды изограви-
метрическим методом), можно получить представление о площади
пор, отнесенной к единице длины пути воды через мембрану. Кроме
того, используя этот же метод, можно найти для молекул разного
размера величину эффективной площади пор, отнесенной к еди-
нице длины диффузионного пути:
А _ JRT
Lx £>ДП'
(13.16)
Гл. 13. Микрососуды большого круга 507
Если предположить, что площадь пор Ав, через которую может
проходить вода, равна истинной площади пор Ап, допускающей
диффузию растворенных веществ, то, разделив (13.15) на (13.16)
и сравнив результат с (13.14), можно получить уравнение, связы-
вающее радиус пор с экспериментально определяемыми величи-
нами. Решение этого уравнения дает значение эффективного ра-
диуса пор, равное 14 нм, что существенно больше значений, полу-
чаемых другими методами, и расхождение данных невозможно
отнести на счет ошибок экспериментов.
Это расхождение удается, однако, объяснить, отбросив пред-
положение о том, что вода способна проходить лишь через те поры,
которые пропускают и жиронерастворимые молекулы. Если при-
нять радиус этих пор равным 4 нм, то можно подсчитать, что че-
рез них проходит только половина молекул воды, а остальная
часть использует другие пути.
Перенос крупных молекул. Долгое время полагали, что для
молекул, более крупных, чем молекулы альбумина, стенка капил-
ляров непроницаема. Однако результаты исследований, подобные
приведенным на рис. 13.46, показали, что для таких молекул стен-
ка имеет хотя и очень низкую, но все-таки конечную проницае-
мость, и значение ее фактически не зависит от размера молекул.
Цель ряда электронно-микроскопических исследований заключа-
лась в том, чтобы выявить в стенке капилляров крупные лоры,
способные пропускать такие молекулы. Эту роль могут играть
окна в эндотелии некоторых капилляров. Полагают, что перенос
крупных молекул осуществляется также цитоплазматическими пу-
зырьками, имеющимися в стенках любых капилляров. Экспери-
ментально установлено, что через стенки капилляров задней ко-
нечности могут проникать пластмассовые микросферы диаметром
до 30—70 нм.
Если перенос таких крупных молекул и частиц осуществляется
через большие поры, то проницаемость капилляров для этих мо-
лекул и частиц должна меняться пропорционально их диффузион-
ной способности. Исследование проницаемости сосудистого русла
печени и кишечника для молекул декстрана, диффузионная спо-
собность которых уменьшается пропорционально квадратному
корню из их молекулярного веса, выявило линейное снижение
проницаемости с увеличением квадратного корня из молекуляр-
ного веса. Эндотелий капилляров в этих органах действительно
является окончатым. Аналогичные исследования проницаемости
сосудистого русла скелетных мышц, капилляры которого имеют
непрерывный эндотелий, не обнаружили такого четкого соотноше-
ния. Возможно, проницаемость в этом случае не столь сильно за-
висит от диффузионной способности молекул, и перенос их осу-
ществляется при помощи пузырьков, а не через неидентифициро-
ванные поры.
S08 Часть И. Механика кровообращения
В 1953 г. Палад впервые высказал предположение, что пере-
носчиками молекул через эндотелий могут служить пузырьки.
С тех пор предпринимались некоторые попытки понять работу та-
кого механизма и оценить его значение. Мы уже упоминали, что
мембраны, ограничивающие пузырьки, — это, насколько можно
установить, точно такие же мембраны, как и те, которые ограни-
чивают клетки, и что находят как свободные пузырьки, располо-
женные в цитоплазме клетки, так и прикрепленные к ее наруж-
ной мембране. Поскольку размер пузырьков мал, изучать их по-
ведение очень трудно — все надежды приходится возлагать на
электронную микроскопию.
Выявлено три существенных процесса, связанных с поведением
пузырьков: движение пузырьков внутри клетки, прикрепление их
к поверхности клетки и отделение от нее и обмен их содержимого
с кровью или с тканевой жидкостью.
Полагают, что внутри клетки пузырьки перемещаются благо-
даря броуновскому движению, обусловленному столкновениями их
с молекулами содержащихся в цитоплазме веществ. Но даже если
допустить, что плотность пузырьков совпадает с плотностью цито-
плазмы, оценить фактическую диффузионную способность пузырь-
ков очень трудно, поскольку неизвестна фактическая вязкость ци-
топлазмы. Более того, реальный диапазон перемещений пузырьков
не может быть таким же, каким он был бы в среде, не имеющей
границ. Расстояние между противоположными мембранами клет-
ки составляет лишь 0,3—0,4 мкм, т. е. 4—5 диаметров пузырьков
(примерно 70 нм). Тем не менее, используя элементарную кине-
тическую теорию, можно сделать вывод, что соударения с окру-
жающими молекулами быстро разгоняют пузырек до скорости
около 30 см-с-1. Поскольку вязкое торможение таких мелких час-
тиц велико, пузырек возвращается в состояние покоя за малые
доли секунды.
Еще менее понятны процессы, связанные с прикреплением пу-
зырьков к стенке клетки и отделением от нее. Если пузырек, дви-
жущийся внутри клетки, подходит достаточно близко к ее наруж-
ной мембране (скажем, на расстояние 2—3 нм), то он попадает в
область действия вандерваальсовых сил, которые притягивают его
к мембране. При этом цитоплазма, заполняющая пространство ме-
жду пузырьком и мембраной, должна вытесниться. По мере сбли-
жения необходимые для вытеснения жидкости силы увеличивают-
ся, но растут также и вандерваальсовы силы. Когда пузырек под-
ходит к наружной мембране клетки очень близко, форма ее изме-
няется, как это показано на рис. 13.48. Затем мембраны пузырька
и клетки сливаются и образуют перемычку, которая в конце кон-
цов разрывается и дает содержимому пузырька выход во внекле-
точное пространство (рис. 13.49). Остатки перемычки хорошо
видны на рис. 13.50. Механизм отделения пузырька неясен, ио,
если отделение произошло, он снова может свободно перемещать-
Гл. 13. Микрососуды большого круга 509
Рис. 13.48. Изгиб мембраны
эндотелиальной клетки при
достаточном приближении к
ней пузырька. [Palade,
Bruns (1968). Structural mo-
dulations of plasmalemmal
vesicles, J. Cell Biol., 37,
633—649.]
ся благодаря броуновскому движению. Пузырек может вновь
прикрепиться к наружной мембране клетки — в той же точке или
поблизости от нее, а может переместиться к отдаленной точке,
скажем у противоположной стороны клетки. Типичное время, в те-
чение которого пузырек находится внутри клетки в свободном
состоянии и прикрепленным к ее мембране, оценивается величи-
нами порядка 5 и 3 с соответственно.
«Загрузка» и «разгрузка» пузырька осуществляются благо-
даря диффузии. Таким образом, пузырьки обеспечивают перенос
как воды и веществ с малым молекулярным весом, так. и крупных
молекул. Однако из-за относительно малой скорости этот путь не
играет большой роли для молекул, способных проникать через
межклеточные щели. Максимальный размер молекул, которые в
состоянии переносить пузырьки, определяется возможностью про-
хождения этих молекул через шейку пузырька, т. е. ими могут
быть молекулы липопротеидов с диаметром около 50 нм. Моле-
кулы чуть меньшего размера проникают в пузырек, но диф-
фузия их через шейку затруднена, Поэтому проницаемость стенки
510 Часть II. Механика кровообращения
| 70 нм 11 70 нм || 70 нм |
Рис. 13.49. Схематическое изображение последовательных стадий слияния мем-
бран пузырька и клетки и разрыва перемычки. [Palade, Bruns (1968). Structural
modulations of plasmalemmal vesicles, J. Cell Bioll, 37, 633—649.]
Рис. 13.50. Вид пузырька вскоре после разрушения перемычки. Четко виден оста-
ток перемычки в середине шейки пузырька. [Palade, Bruns (1968). Structural
modulations ox plasmalemmal vesicles, J. Cell Biol., 37, 633—649J
Гл. 13. Микрососуды большого круга 611
капилляров для молекул, переносимых пузырьками, может до не-
которой степени зависеть от коэффициента диффузии этих мо-
лекул.
СПИСОК ЛИТЕРАТУРЫ
Crone С., Lassen N. A. (eds). (1970). Capillary permeability, Munksgaard, Co-
penhagen.
Fung Y. C. (1968). Biomechanics, its scope, history, and some problems of conti-
nuum mechanics in physiology, Appl. Meehan. Rev., 21, 1—20.
Gaehtgens P., Uekermann U. (1971). The distensibility of venous microvessels,
Pflfig. Arch. ges. Physiol., 330, 206—216.
Landis E. M., Pappenheimer J. R. (1963). Exchange of substances through the ca-
pillary walls. In: Handbook of physiology, Section 2: Circulation, Vol. II (eds.
W. F. Hamilton and P. Dow), pp. 961—1034, American Physiological Society,
Washington, D. C.
Lightfoot E. N. (1974). Transport phenomena and living systems, Wiley-Intersci-
ence, London. Имеется перевод: Лайтфут Э. Явления переноса в живых си-
стемах. М., Мир, 1977.
Johnson Р. С. (1973). The microcirculation and local humoral control of the cir-
culation. In: Cardiovascular physiology (ed. A. C. Guyton), M. P.T., London.
Majno 0. (1965). Ultrastructure of the vascular membrane. In: Handbook of phy-
siology (eds W. R. Hamilton and P. Dow), Section 2: Circulation, Vol. II,
pp. 2293—2375, American Physiological Society, Washington, D. C.
Michel С. C. (1972). Flows across the pacillary wall. Cardiovascular fluid dyna-
mics, Vol. II (ed. D. H. Bergel), Chapter 17, Academic Press, New York.
Middleman S. (1972). Transport phenomena in the cardiovascular system, Wiley-
Interscience, London.
Pappenheimer J. R., Renkin E. M., Borrero L. M. (1951). Filtration, diffusion and
molecular sieving through peripheral capillary membranes, Am. J. Physiol.,
167, 13—46.
Snashall P. D., Lucas J., Guz A., Floyer M. A. (1971). Measurements of intersti-
tial «fluid» pressure by means of a cotton wick in man and animals: an ana-
lysis of the origin of the pressure, Clin. Sci., 41, 35—53.
Weinbaum S., Caro C. G. (1976). A macromolecule transport model for the arte-
rial wall and endothelium, J. Fluid Meeh., 74, 611—40.
Wolstenholme G. E. IF. (ed.). (1969). Circulatory and respiratory mass transport,
Churchill, London.
Zweifach B. W. (1974). Quantitative studies of microcirculatory structure and func-
tion, Circulation Res., 34, 834—866.
Глава 14
ВЕНЫ БОЛЬШОГО КРУГА
Сведения о механике движения крови в венах значительно ме-
нее обширны, чем о механике кровотока в артериях. Между тем
фактически вся кровь, изгоняемая левым желудочком, должна воз-
вращаться в правое предсердие через вены1)- В нормальных ус-
ловиях в венах содержится почти 80% всего объема крови сосу-
дистой системы большого круга, и в регуляции минутного объема
сердца вены играют значительную роль. Вот почему изучение
механики вен столь необходимо.
Венозная система напоминает артериальную в том смысле, что
она тоже представляет собой древовидно ветвящуюся сеть сосу-
дов. Главными стволами являются полые вены, которые соеди-
няются друг с другом и впадают в сердце. Однако в некоторых
отношениях венозная система принципиально отличается от арте-
риальной: во-первых, как можно видеть из рис. 12.11, в нормаль-
ных условиях давление в венах значительно ниже, чем в артериях,
находящихся на том же уровне относительно сердца, и может
быть даже ниже атмосферного давления (например, в венах, рас-
положенных выше уровня сердца); во-вторых, венозные сосуды
имеют более тонкие стенки, и при изменении давления в физио-
логических пределах диапазон изменения их растяжимости значи-
тельно больше, чем для артерий; далее, кровь движется в венах
от периферии к сердцу, и скорость притока крови в вены опреде-
ляется разностью давлений в артериях и в венах и сопротивле-
нием находящихся между ними микрососудов; наконец, во многих
венах имеются клапаны, препятствующие обратному течению кро-
ви Все эти различия приводят к тому, что распределение давле-
ния и скорости кровотока в венозной системе и в артериальной
оказываются совершенно разными, хотя многие определяющие их
физические факторы, например распространение и отражение волн,
качественно одинаковы.
Эта глава начинается с описания анатомической организации
и гистологического строения венозных сосудов, после чего рас-
сматриваются действующее на вены трансмуральное давление и
упругая реакция их на изменения этого давления. Затем обсуж-
*) Артериальная кровь, поступающая к легким через сосудистое русло брон-
хов, возвращается в левое предсердие, в нормальных условиях ее объем состав-
ляет очень малую долю (меньше 5%) минутного объема сердца.
Гл. 14. Вены большого круга 513
Даются наблюдаемые в венах динамические флуктуации давления
и скорости кровотока и определяющие их физические факторы.
Специальный раздел посвящен интересным явлениям, которые мо-
гут происходить при протекании крови через сосуд, способный спа-
даться. В заключение рассматриваются некоторые вопросы меха-
ники венозной системы большого круга как единого целого.
14.1. Анатомическая организация
Вены большого круга берут начало от венул, т. е. сосудов,
к которым сходятся капилляры, и заканчиваются там, где полые
вены впадают в правое предсердие. От этой общей схемы строе-
ния венозной системы имеются отклонения: есть области, где кровь
протекает последовательно через две системы микрососудов (на-
пример, в системе воротной вены1) и в почках), и области, где су-
ществуют артерио-венозные анастомозы, полностью шунтирующие
капилляры. Тем не менее венозную систему можно представить
сетью сосудов, которые по мере приближения к сердцу объеди-
няются и увеличиваются в диаметре, в то время как полная пло-
щадь поперечного стечения соответствующих сосудов при этом
уменьшается. Емкость венозной системы значительно больше, чем
артериальной. Это обусловлено как большим числом венозных
сосудов, так и тем, что на каждом конкретном уровне вены, отво-
дящие кровь от данного сосудистого русла, обычно оказываются
крупнее, чем соответствующие приносящие кровь артерии. Способ-
ность венозных сосудов значительно изменять свою емкость опре-
деляется их большей растяжимостью, а также особенностями дей-
ствия мышц, расположенных в стенке сосудов и окружающих эти
сосуды.
Относительно деталей распределения объема крови по веноз-
ной системе имеются некоторые разногласия, что связано со слож-
ностью изучения этого распределения. Как мы видели в гл. 13,
наиболее обстоятельно изучены двумерные, расположенные в про-
зрачных для света тканях сосудистые русла. Пример тому — кры-
ло летучей мыши; размеры и площади поперечного сечения раз-
личных кровеносных сосудов этого русла даны в табл. 12.1. Из
таблицы следует, что при переходе от капилляров к венулам об-
щая площадь поперечного сечения сосудов увеличивается, но при
переходе к еще более крупным Сосудам уменьшается: площадь ве-
нул превышает площадь мелких вен в 4,6 раза, а крупных — в 26
раз. Площадь венул превышает также и площадь артериол — при-
мерно в 25 раз. Различие емкости венул, мелких вен и крупных
’) Здесь кровь, оттекающая из ЖеЛуДка, киТпеЧника, Поджелудочной железы.
Селезенки и Желчного пузыря, поступает в Воротную вену, откуда направляется
Непосредственно в печень, которая имеет свою собственную систему мйкрососу-
дов Отсюда через печеночные вены кровь проходит в нижнюю полую вену.
5t4 Часть tl. Механика Кровообращения
вен, естественно, не столь велико, как различие суммарной пло-
щади их поперечного сечения, так как наиболее мелкие сосуды
являются также и наиболее короткими.
Результаты измерения параметров сосудистого русла сальника
собаки оказались, однако, существенно иными. Сообщалось, что
в этом русле полная площадь поперечного сечения венул превы-
шает полную площадь артериол лишь в 4,5 раза. Было бы весьма
Желательно подтвердить эти данные другими методами, например
измерив скорость крови в различных сосудах (при увеличении
полной площади поперечного сечения скорость крови должна про-
порционально уменьшаться). Но, Насколько нам известно, такие
измерения не производились ни для крыла летучей мыши, ни для
сальника собаки. Тем не менее результаты измерения скорости
крови в сосудах сальника кошки согласуются с данными о соот-
ношении площадей поперечного сечения в сосудах сальника со-
баки. Наиболее вероятная причина расхождения результатов из-
мерения площади сосудов у летучей мыши и у собаки заключает-
ся в том, что относительные размеры венозной и артериальной си-
стем различных органов могут быть у разных животных неодина-
ковы.
Стенки вен значительно тоньше, чем стенки артерий. Напри-
мер, при физиологических значениях трансмурального давления
отношение толщины стенки к диаметру для вен составляет обычно
0,01—0,02, а для артерий большого круга — 0,06—0,08 (табл. I).
О строении венозного русла известно слишком мало, чтобы де-
тально характеризовать конусность отдельных венозных сосудов
(если она вообще существует) или говорить об отношении площа-
дей и об углах ветвления в области соединений даже самых круп-
ных венозных сосудов.
Исходя из данных о размерах вен, а также из результатов ги-
стологических исследований строения их стенки венозные сосуды
разделяют на 5 групп: посткапиллярные венулы, венулы, мелкие
вены, средние вены и крупные вены. Последние две группы иногда
объединяют в одну. Строение стенки посткапиллярных венул (диа-
метр 8—30 мкм) и венул (диаметр 100—200 мкм) было подробно
описано в гл. 13 и здесь рассмотрено лишь вкратце. Переход от
капилляров к венам совершается постепенно: элементы соедини-
тельной ткани появляются в стенках сосудов уже вблизи конца
капилляров, а гладкомышечные клетки обнаруживаются чуть
дальше. При объединении нескольких капилляров образуются
посткапиллярные венулы диаметром около 20 мкм. Их стенка со-
стоит из слоя эндотелия, окруженного тонким слоем продольно
расположенных коллагеновых волокон и фибробластов. В вену-
лах диаметром 40—50 мкм между эндотелием и соединительной
тканью появляются отдельные циркулярные гладкомышечные
клетки. В венах диаметром 200—300 мкм расположенные таким
образом гладкие мышцы образуют непрерывный слой, а наружная
Гл, 14. Вены большого круга 515
оболочка становится более толстой, и, помимо продольно ориен-
тированных коллагеновых волокон, в ней обнаруживаются волок-
на, расположенные хаотически. Мышечная оболочка мелких вен
(диаметром до 0,2 см) многослойная, причем отдельные мышеч-
ные слои отделяются друг от друга рыхлой коллагеновой тканью.
Именно на этом уровне выявляются три характерных для стенок
кровеносных сосудов слоя: внутренняя, средняя и наружная обо-
лочки (разд. 12.1). Стенка еще более крупных вен сходна по
строению со стенкой артерий. Основные различия заключаются в
том, что средняя оболочка вен значительно тоньше, а наружная
их оболочка, состоящая преимущественно из коллагеновых воло-
кон, представляет собой по объему основную часть стенки. Кроме
того, в стенке вен слабо развита внутренняя эластическая мембра-
на. У вен тех частей тела, которые при обычной позе оказываются
ниже уровня сердца (например, вены нижних конечностей), про-
является тенденция к относительному утолщению средней оболоч-
ки. Обусловлено ли это тем, что они постоянно подвергаются дей-
ствию сравнительно высокого трансмурального давления, неиз-
вестно.
В стенке вен диаметром выше примерно 0,1 см имеются питаю-
щие их сосуды (vasa vasorum — сосуды сосудов). Они отходят от
расположенных поблизости мелких артерий большого круга и про-
никают в наружную оболочку вен, где образуют обширную капил-
лярную сеть. Эта сеть простирается вглубь стенки почти до внут-
ренней оболочки и соединяется с венулами в наружной оболочке.
Вены богато иннервированы симпатическими .волокнами. В обыч-
ных условиях расположенные в стенке вен гладкие мышцы нахо-
дятся в состоянии активного напряжения (т. е. тонуса). Перерезка
нервов или устранение влияния нервных импульсов теми или
иными веществами приводит к расширению вен вследствие умень-
шения напряжения в их стенке. Такое расширение может значи-
тельно увеличить площадь поперечного сечения вены.
Многие вены средних размеров, особенно вены конечностей,
снабжены клапанами, которые препятствуют движению крови в
обратном направлении. (У человека в тех венах, по которым кровь
оттекает от органов брюшной полости, за исключением воротной
вены, а также в полых венах клапанов нет.) Створки клапанов —
обычно их две — состоят из содержащей эластиновые волокна сое-
динительной ткани, покрытой эндотелием. Вблизи каждой створ-
ки со стороны, лежащей ниже по течению (расположенной ближе
к сердцу), обычно имеется карман, т. е. выпячивание стенки. Пред-
полагается, что механическая функция карманов такая же, как и
синусов Вальсальвы, расположенных позади каждой створки аор-
тального клапана (механика его рассмотрена в разд. 11.5): они
обеспечивают стабильное открывание и плавное закрывание кла-
панов. Подробно действие венозных клапанов не изучено.
516 Часть //. Механика кровообращения
14.2. Трансмуральное давление и статические упругие свойства
Когда испытуемый лежит в горизонтальном положении на
спине, давление в находящихся на уровне сердца венозных сосу-
дах, измеренное по отношению к атмосферному давлению, меняет-
ся примерно от 2,0-103 Н-м-2 (20 см вод. ст.) в венулах до одной
трети этой величины в полых венах, хотя в грудной полости из-за
дыхательных колебаний давление может быть и ниже атмосфер-
ного (см. рис. 14.20). Разность этих давлений заставляет кровь
двигаться по венам обратно к сердцу. Поскольку в обычных усло-
виях давление снаружи вен близко к атмосферному, приведенные
значения давления в венах характеризуют также трансмуральное
давление, которое стремится их растянуть. Когда испытуемый пе-
реходит из горизонтального положения в вертикальное, избыточ-
ное давление рКЗб (относительно гидростатического давления;
разд. 12.2), которое определяет среднюю величину кровотока в
венах, остается в сущности прежним, тогда как трансмуральное
давление сохраняется неизменным только на уровне сердца. Вены
ног при этом наполняются оттекающей из микрососудов кровью,
до тех пор, пока давление в них не увеличится примерно на
104 Н-м~2 (1. е. на величину гидростатического давления, созда-
ваемого столбом крови высотой 1 м). В то же время в венах шеи
или поднятой руки давление значительно уменьшается и стано-
вится ниже атмосферного. Как мы увидим дальше, растягивающее
вены ног трансмуральное давление можно уменьшить, если на-
прячь окружающие их скелетные мышцы и тем самым повысить
наружное (для вен) давление. Это приведет к уменьшению пло-
щади их поперечного сечения, а следовательно, и к уменьшению
емкости, и кровь из этих вен переместится к сердцу. Вены, рас-
положенные выше сердца, обычно спадаются (разд. 12.2), и их
невозможно раскрыть никакими мышечными усилиями.
Таким образом, физиологические пределы изменения площади
поперечного сечения и формы вен значительно больше, чем арте-
рий, круглое поперечное сечение которых при таких же изменениях
трансмурального давления изменяет свою площадь на относи-
тельно небольшую величину. Возможность спадения вен имеет
целый ряд важных механических последствий, которые рассмот-
рены в следующих трех разделах. Здесь мы опишем (в той мере,
в какой это известно) изменения площади поперечного сечения,
формы и растяжимости вен, сопровождающие постепенное измене-
ние трансмурального давления (в физиологически нормальном
диапазоне). Все упоминаемые ниже измерения производились на
удаленных из организма и утративших тонус сегментах крупных
вен, длину которых поддерживали такой же, какой она была в
условиях организма. In vivo реакции должны быть количественно
существенно иными, поскольку из-за сокращения гладких мышц
вены становятся менее растяжимыми; тем не менее активность
Гл. 14. Вены большого круга 517
гладких мышц не меняет в значительной степени их сопротивление
спадению, так как стенки вен очень тонки.
Наглядное представление об имеющихся данных относительно
упругих свойств сегментов вен дает рис. 14.1, построенный по ре-
зультатам измерений и исходя из тех выводов, которые можно сде-
лать из этих измерений. На рисунке обобщено все, что известно
о статическом поведении вен, и он принципиально важен для по-
нимания последующего материала этой главы. Кривые построены
не по непосредственным данным измерений для одного сегмента, и
их следует рассматривать как некие схемы. Они отражают изме-
нение ряда параметров в зависимости от величины трансмураль-
ного давления (ртм). Этими параметрами являются:
А. Изменение площади поперечного сечения (Л—Ло), отнесен-
ное к площади поперечного сечения Ло при нулевом трансмураль-
ном давлении; эта величина представляет собой безразмерную
меру изменения площади, и потому ее можно использовать при
сравнении результатов, полученных для разных .сосудов. В диапа-
зоне значений трансмурального давления Л^-Ю3 Н-м-2 эта кри-
вая непосредственно отражает результаты измерений, произведен-
ных на полой вене собаки.
Б. Аналогичное отношение для периметра I поперечного сече-
ния. Центральная часть этой кривой (в диапазоне ±0,5-103 Н-м-2)
построена по данным, представленным на рис. 14.2,5 и рис. 14.1, Л;
для трансмурального давления, превышающего 1,5-103 Н-м-2,дан-
ные взяты из рис. 14.3,Л; остальная часть кривой проведена так,
чтобы получить плавное сопряжение указанных выше участков.
В. Отношение длин большой и малой осей поперечного сече-
ния, а/b- поперечное сечение принимает форму эллипса лишь ког-
да оно начинает терять форму круга; на конечных же стадиях
спадения оно не эллиптическое, и потому это отношение не опре-
делено. Кривая построена по результатам ряда измерений.
Г и Д. Два параметра, каждый из которых является мерой
растяжимости сосуда. В разд. 12.3 растяжимость была опреде-
лена как относительное изменение площади (АЛ/Л), вызванное
небольшим изменением трансмурального давления (Артм):
ДрТМ
Таким образом, величина D показывает, насколько легко измене-»
ние трансмурального давления изменяет площадь поперечного се-
чения: сосуд с низкой растяжимостью является очень жестким, и
наоборот. Именно это определение растяжимости используется в
уравнении (12.9) для скорости распространения волн давления.
В артериях изменение трансмурального давления в обычных фи-
зиологических пределах сопровождается не слишком сильным из-
менением площади поперечного сечения. Поэтому при замене в
знаменателе формулы (14.1) площади Л некоторой фиксированной
(14.1)
518 Часть II. Механика кровообращения
Рис. 14.1. Графики, дающие представление об упругих свойствах сегментов вены.
По оси абсцисс везде отложено трансмуральное давление р™; за единицу его
принята величина 103 Н-м-2 (10 см вод. ст.). По осн ординат отложены сле-
дующие величины. А. Изменение площади поперечного сечения (А — Ао), отне-
сенное к площади поперечного сечения при трансмуральном давлении, равном
нулю (Ао). Б. Изменение периметра (Z — /о), отнесенное к периметру при транс-
муральном давлении, равном нулю (/о). В. Отношение длин большой и малой
осей поперечного сечения, а/b; это отношение вводится только когда сечение
имеет форму круга или эллипса. Г. «Псевдорастяжимость» Do, определяемая
уравнением (14.2) и равная наклону кривой А. Д. Действительная растяжи-
мость D, определяемая уравнением (14.1); Максимум этой кривой при траисму-
Гл. 14. Вены большого круга 519
(14.2)
площадью, например площадью поперечного сечения артерии при
растягивающем давлении 13,3-103 Н-м-2 (100 мм рт. ст.)—такую
замену обычно производят для удобства — в значение D вносится
лишь очень небольшая ошибка. Однако для вен замена величины
А фиксированной площадью приводит к значительной ошибке, так
как в этом случае при изменении трансмурального давления в
пределах физиологического диапазона площадь меняется очень
сильно. Тем не менее некоторые исследователи вместо площади А
используют фиксированную площадь, а именно площадь попереч-
ного сечения вены Ло при трансмуральном давлении, равном нулю,
и определяют таким образом «псевдорастяжимость» Do:
D - 1 АЛ
0 Ло Дртм
Чтобы проиллюстрировать значительное различие между этими
двумя характеристиками растяжимости, построены графики как
для Do (рис. 14.1,Г), так и для D (рис. 14.1, Д). Обе кривые полу-
чены пересчетом из приведенной на рис. 14.1, А кривой изменения
площадей. Более полезной следует считать нижнюю кривую
(рис. 14.1, Д).
Рассмотрим, что происходит с веной при постепенном сниже-
нии трансмурального давления от высокого (для вен) значения,
превышающего 7-103 Н-м-2 (70 см вод. ст.). При таком давлении
поперечное сечение вены имеет форму круга (а/6=1), и вена
относительно жесткая, поскольку растяжимость ее мала, а модуль
Юнга велик. Это, по-видимому, обусловлено тем, что ответ вены
на изменение давления определяется в таких условиях в основном
коллагеновыми волокнами ее стенки. При снижений трансму-
рального давления примерно до 1,5-103 Н-м-2 сечение сосуда
остается круглым, а растяжимость его меняется незначительно;
площадь поперечного сечения уменьшается медленно, а пери-
метр — еще медленнее (так как пока сечение круглое, его пери-
метр пропорционален квадратному корню из площади). Когда
трансмуральное давление снижается от 1,5-103 до 1-103 Н-м-2,
сечение все еще остается круглым, но сосуд становится несколько
более растяжимым, вероятно, потому, что механические свойства
его стенки при этом начинают определяться уже не столько кол-
лагеном, сколько эластином (как и для артерий; разд. 7.2). Когда
ральном давлении +0,5-103 Н-м~2 соответствует максимальному наклону кри-
вой А; максимум при отрицательном трансмуральном давлении обусловлен очень
малыми значениями А Над кривой А изображена форма поперечного сечения
вены при соответствующих значениях трансмурального давления (Часть данных
взята из работы Moreno, Katz, Gold, Reddy (1970) Mechanics of distension of
dog veins and other very thin-walled tubular structures, pp 1069—r 1079, Circulation
Res 27, с любезного пазрешения American Heart Association Inc. и из работы
Attincfer (1969) Wall Properties of Veins, pp. 253—261, Trans on Bio-med Eng.,
BME-16 Institute of Electrical and Electronics Engineers Inc., New York.]
620 Часть It. Механика кровообращения
трансмуральное давление падает ниже аНО3 Н-м-2 (но еще пре-
вышает 0,5-103 Н-м-2), сечение становится слегка эллиптическим,
площадь его уменьшается быстрее, а растяжимость сосуда про-
должает расти. Периметр также продолжает уменьшаться, хотя
величина его теперь уже не строго пропорциональна квадратному
корню из площади, так как поперечное сечение сосуда перестало
быть круглым. При этом основной вклад в уменьшение площади,
а следовательно, и в изменение растяжимости сосуда все еще вно-
сит именно уменьшение периметра, а не изменение формы попе-
речного сечения. При снижении трансмурального давления до
0,5-103 Н-м-2 и дальше поперечное сечение сосуда становится су-
щественно эллиптическим, и изменение его формы дает все боль-
ший вклад в изменение площади и растяжимости. Растяжимость
достигает максимума при трансмуральном давлении 0,5-103 Н-м-2,
когда наступает резкое изменение площади поперечного сечения.
Наклон кривой для площади, а следовательно, и псевдорастяжи-
мость £>0 после этого уменьшаются, потому что изгиб стенки со-
суда в местах ее наибольшей кривизны (см. ниже) все более
затрудняется. Такой изгиб должен произойти перед тем, как при
отрицательном трансмуральном давлении, от —1-103 до —2-
•103 Н-м-2 (от —10 др —20 см вод. ст.), сосуд окончательно спа-
дется. Поскольку по мере спадения сосуда площадь поперечного
сечения его становится очень малой, при трансмуральном давле-
йии около —0,5-Ю3 Н-м-2 действительная растяжимость D дости-
гает еще одного максимума, после чего, при переходе к состоянию
окончательного спадения, она неравномерно убывает до очень низ-
ких значений. (В этом диапазоне давлений точки имеют большой
разброс, и потому неравномерность хода кривой может быть час-
тично обусловлена неточностью измерений малой площади и ее
изменения.)
На последней стадии спадения сосуда периметр его почти не
меняется. Когда конечное состояние уже достигнуто, поперечное
сечение имеет форму силуэта гантели, т. е. представляет собой
два маленьких, почти круглых боковых канала, разделенных сплю-
щенной частью вены. Согласно некоторым гистологическим дан-
ным, при спадении сосудов внутренняя поверхность их сморщи-
вается и боковые каналы могут при этом полностью закрыться.
Важно помнить, что весь процесс спадения вен происходит при
трансмуральном давлении приблизительно от —103 Н-м-2 до
103 Н-м-2, т. е. в середине физиологического диапазона давлений.
Результаты исследования упругих свойств кровеносных сосудов
часто представляют через эффективный касательный модуль Юнга
Е, определяемый при растяжении сосуда по окружности. Когда
сосуды круглые, величина Е связана с растяжимостью и отноше-
нием толщины стенки к диаметру (h/d) соотношением
1 Eh
D d
(14.3)
Гл. 14. Вены большого круга 521
[ср. с уравнением (7.36)]. Это соотношение показывает, что если
стенки сосуда достаточно тонки, то и при большом значении мо-
дуля Юнга он может иметь очень высокую растяжимость. Если
поперечное сечение сосуда не является круглым, то определять мо-
дуль Юнга с помощью уравнения (14.3) нельзя, потому что в этом
случае он непостоянен по сечению сосуда (в окружном направле-
нии). Однако модуль Юнга остается важным параметром, харак-
теризующим легкость спадения сосуда (см. ниже), и если две
трубки одинакового размера имеют разные модули Юнга, то по-
ведение их при спадении оказывается различным. Поэтому, на-
пример, не следует распространять на поведение вен те законо-
мерности, которые выявляются при наблюдении за свойствами ре-
зиновых трубок с близкими диаметром и толщиной стенок. Мо-
дуль Юнга для резины равен примерно 2,1 • 106 Н-м-2, в то время
как типичное значение его для вен, когда поперечное сечение их
круглое [вычисляется по уравнению (14.3)], — около 5-Ю4 Н-м-2.
Различие поведения вены и тонкостенной трубки из латекса
при снижении трансмурального давления от значений, превышаю-
щих 2-103 Н-м-2, до более низких, чем —103 Н-м-2, отображено
на рис. 14.2. На рис. 14.2, Л графически представлены соотноше-
ния давление — площадь, а на рис. 14.2,5 — соотношения между
площадью и периметром. При больших трансмуральных давле-
ниях максимальная площадь поперечного сечения трубки из ла-
текса меньше максимальной площади вены и такая трубка значи-
тельно менее растяжима. Сечение трубки из латекса остается
круглым при снижении трансмурального давления почти до нуля,
после чего оно, как и в случае вены, становится эллиптическим,
а затем и гантелеобразным. Однако периметр такой трубки при
спадении остается почти постоянным, и изменение площади попе-
речного сечения связано только с изменением его формы. Более
того, скорость изменения площади трубки из латекса максимальна
при отрицательном значении ртм (около —0,25-Ю3 Н-м-2), тогда
как для вен она достигает максимума при положительном р™
(около +0,5-103 Н-м-2). Изменение наклона кривых на
рис. 14.2, Л (т. е. псевдорастяжимости) также различается: для
вены наклон меняется менее резко, чем для трубки. Это обуслов-
лено тем, что даже после того, как начинает меняться форма по-
перечного сечения вены, изменение ее периметра все еще продол-
жает влиять на изменение площади.
В следующей главе мы покажем, что артерии и вены малого
круга, которые, подобно венам большого круга, являются тонко-
стенными гибкими сосудами, при низких, но еще физиологических
значениях трансмурального давления ведут себя приблизительно
так же, как вены большого круга (а не как трубки из латекса).
Интересно сравнить упругие свойства трех основных типов круп-
ных кровеностных сосудов: вен большого круга, артерий малого
круга и артерий большого круга. С этой целью на рис. 14.3,4 и §
522 Часть II. Механика кровообращения
Рис. 14.2. А. Соотношения давление—площадь для сегмента rpmw ы ппс
из латекса: площадь, как н па рис. 14.1, А. характеризуется отношением ее изменения‘uffi
иой площади; давление представлено как трансмуральное давление рти. Внутренний" днаме™
вены н трубки равен 1.2 см, а отношение толщины стенки к /ь/л. А „
однако модуль Юнга для трубки в 40 раз'больше, тем^ля йМ?Т1г,,<’иОда;
относительного изменения периметра вены и трубки из латекса от опи^. ’ Ь' Зависим°оть
площади нх поперечного сечения. Крестики-вена*-гХуго^ки-^Х^1ЬНОГО нэ“?не«ия
Kata Gold. Reddy (1970) Mechanics of distension of dog vems and otfc^hKW??0'
etrutturea. Circulation R.s , 2?, Ю75, 1076, 1079, до разр^ен^ТтеП^Щ
Гл. 14. Вены большого круга 523
показаны результаты измерений, произведенных для сосудов со-
баки в диапазоне давлений, в котором поперечное сечение их
остается круглым. На рис. 14.3, Л приведена зависимость диа-
метра сосудов, отнесенного к их диаметру при трансмуральном
давлении 103 Н-м-2 (10 см вод. ст.), от трансмурального давле-
ния. Графики зависимости наклона этих кривых от величины
трансмурального давления приведены на рис. 14.3, Б; значения на-
клона здесь пересчитаны в значения эффективного касательного
модуля Юнга для стенки сосудов, который связан с растяжи-
мостью сосудов уравнением (14.3). Заштрихованные области на
обоих графиках указывают диапазоны значений трансмураль-
ного давления, реализующиеся в физиологических условиях. Если
считать эти результаты представительными для всех таких сосу-
дов, то можно сделать два вывода:
1. В нормальном для каждого типа сосудов диапазоне давле-
ний легочные артерии имеют такой же модуль Юнга, как и арте-
рии большого круга, но из-за меньшего отношения толщины стен-
ки легочных артерий к диаметру (табл. I) растяжимость этих
артерий примерно в семь раз больше; растяжимость вен большого
круга приблизительно в три раза превышает растяжимость легоч-
ных артерий, так как модуль Юнга для этих вен составляет около
одной трети модуля Юнга для легочных артерий, а отношение
h/d для тех и других сосудов примерно одинаково.
2. При равных значениях трансмурального давления во всем
исследованнном диапазоне давлений вены оказываются более же-
сткими, чем легочные артерии, которые в свою очередь более
жесткие, чем артерии большого круга. Эти результаты могут быть
обусловлены тем, что у верхней границы физиологически нормаль-
ного диапазона давлений в связи с возрастанием при больших
трансмуральных давлениях роли коллагена происходит резкое
увеличение жесткости сосудов. Рассматривая поведение артерий
(разд. 12.2), мы уже выяснили, что при повышении трансмураль-
ного давления равные приращения его вызывают постепенно уве-
личивающиеся приращения площади до тех пор, пока при этом не
начнет возрастать жесткость сосуда Давление, требующееся для
разрыва вены, во много раз превышает обычные значения транс-
мурального давления и составляет примерно 5-105 Н-м-2 (5 атм.).
Это установил еще в 1733 г. английский священник Хейлс
(1677—1761) в одном из своих многочисленных опытов по изуче-
нию большого круга кровообращения у животных.
Поскольку материал стенки сосуда несжимаем, при растяже-
нии сосуда стенка должна становиться тоньше. Соответствующие
количественные данные для средних и крупных вен, а также для
других сосудов представлены на рис. 14.4. Из этого рисунка вид-
но, что при увеличении трансмурального давления отношение
толщины стенки к диаметру сосуда (h/d) уменьшается до по-
стоянной величины, составляющей для вен большого круга и ар-
524 Часть 11. Механика кровообращения
d/di
Рис. 14.3 А. Зависимость диаметра различных сосудов собаки от трансмураль-
ного давления ртм (диаметр d отнесен к диаметру rfj при трансмуральном дав-
лении 103 Н-м-2 = 10 см вод. ст.). Заштрихованные области указывают физио-
логические диапазоны значений трансмурального давления для артерий большого
круга, легочных артерий, а также для вен большого круга, расположенных на
уровне сердца • — полая вена, И — яремная вена, О — легочная артерия, □ —
нисходящая аорта, Д — сонная артерия.
терий малого круга примерно 0,01—0,02, чго во много раз меньше
аналогичной величины для артерий большого круга.
Все приведенные выше данные получены на вырезанных сег-
ментах крупных вен. In vivo на соотношение между трансмураль-
ным давлением и растяжимостью вен может оказывать влияние нх
тонус, а также окружающие ткани. Однако о влиянии этих фак-
торов известно очень мало, поскольку in vivo измерять трансму-
ральное давление в венах и исследовать при этом их форму крайне
сложно. Известно, что и в физиологических условиях поперечное
сечение крупных вен большого круга может становиться не круг-
лым. Это показано методом рентгенографии в двух плоскостях для
вен мышц голени людей, находившихся в вертикальном положе-
нии. Рентгенограммы получали после местного внутривенного вве-
Гл 14. Вены большого круга 525
Рис. 14.3 Б. Зависимость эффективного касательного модуля Юнга Е от транс-
мурального давления р™ для различных сосудов. Заштрихованные области — то
же, что на А. Ц — нижняя и верхняя полые вены, А — яремная вена, • — ле-
гочная артерия, □ — нисходящая аорта, О — сонная артерия. fAttinger (1969).
Wall properties of veins, pp. 257—258, Trans, on Bio-med. Eng., BME-16, Institute
of Electrical and Electronics Engineers Inc., New York.]
дения рентгено-контрастного вещества — сначала при неподвиж-
ной конечности и трансмуральном венозном давлении около 7-
• 103 Н-м-2, а затем после значительного снижения этого давле-
ления, вызванного сокращением мышц голени (что приводит к вы-
давливанию крови из вен; см. разд. 14.5). В первом случае сред-
ние значения двух диаметров вены составляли 0,61 и 0,47 см, а
во втором — 0,15 и 0,06 см. Могут ли мелкие вены большого круга
принимать при низких значениях трансмурального давления фор-
му, при которой их сечение существенно отличается от круглого,
неизвестно.
Сопротивление стенки трубки изгибу. Очевидно, что изгиб стен-
ки является важной особенностью поведения кровеносных сосудов
при низких значениях трансмурального давления. Как мы убеди-
лись, вены и резиновые трубки с одинаковыми размерами ведут
себя в этом отношении существенно по-разному. Чтобы объяснить
такое различие, нужно разобраться в механике процесса изгиба-
ния. Предположим, что стенка трубки изгибается на всем своем
протяжении одинаково. В этом случае достаточно рассмотреть от-
резок трубки, образующий тонкостенный элемент в форме кольца
526 Часть II. Механика кровообращения
Рис. 14.4. Зависимость отношения толщины стенки различных сосудов к их диа-
метру (hid) от трансмурального давления р1М. Обратите внимание на значитель-
ное изменение этого отношения при низких растягивающих давлениях. Заштри-
хованные области — то же, что на рис 14.3 А. • — нисходящая аорта, □ — сон-
ная артерия, О-—легочная артерия, А—яремная вена, Ц—верхняя полая
вена. [Attinger (1969) Wall properties of veins, p. 257. Trans, on Bio-med. Eng.,
BME-16, Institute of Electrical and Electronics Engineers Inc., New York.]
Б
Рис. 14.5. А. Часть трубки, имеющая форму кольца. Внутренний диаметр d,
толщина стенки h, ширина вдоль оси трубки Ь. ВС — элемент кольца, который
можно приближенно считать неизогнутым, но длина которого значительно боль-
ше, чем Ь и h. Б. Выделенная часть трубки после того, как сжатие привело к
ее спадению. На концах, отмеченных буквой А, изгибающий момент имеет наи-
большую величину, а радиус кривизны — наименьшую.
Гл. 14. Вены большого круга 527
(рис. 14.5,Л). Толщину кольца в радиальном направлении обо-
значим через h, а ширину вдоль оси трубки — через Ь. Когда под
действием сжимающего трансмурального давления кольцо дефор-
мируется, оно принимает форму, подобную изображенной на
рис. 14.5, Б; наибольшую кривизну его стенка имеет у точек, по-
меченных буквой А.
После того как кольцо «сплющивается», точный анализ стано-
вится очень сложным. Но можно провести приближенный анализ,
рассмотрев поведение элемента кольца ВС, длина которого доста-
точно мала, чтобы этот элемент можно было считать практически
прямым, но в то же время достаточно велика по сравнению
с параметрами его поперечного сечения (ft и Ь). Такой элемент
напоминает балку с постоянным поперечным сечением, и для опи-
сания его поведения можно использовать элементарную теорию
изгиба однородной балки, закрепленной на одном конце
(рис. 14.6, А). Сжимающие силы, действующие на стенку трубки,
можно представить в виде равномерно распределенной вдоль
балки и направленной вниз силы, заставляющей балку сгибаться.
Еще более простой (но эквивалентный) случай — действие силы на
балку в одной точке (рис. 14.6,Б). Степень изгиба зависит от ве-
личины силы F и от расстояния между точкой приложения силы
и местом закрепления балки, точнее от момента силы относительно
места закрепления, т. е. от произведения Fx, которое называют
изгибающим моментом. (Изгибающий момент, создаваемый рав-
номерно распределенной силой, в месте закрепления балки равен
2~.PL2, где Р — сила, приходящаяся на единицу длины балки, а
L — длина балки.)
Рассмотрим небольшой элемент балки, обозначенный на
рис. 14.6 буквой X. При изгибе верхняя часть такого элемента
растягивается относительно центральной линии, а нижняя сжи-
мается (рис. 14.6,В). Следовательно, сопротивление балки изгибу
определяется сопротивлением ее материала растяжению и сжа-
тию, которое характеризуется модулем Юнга Е (разд. 7.1). Кроме
того, при данном Е балка будет сгибаться тем легче, чем меньше
ее толщина h. Это обусловлено тем, что необходимая для достиже-
ния определенного радиуса кривизны балки (Р, рис. 14.6, В) сте-
пень растяжения верхней части элемента X и сжатия его нижней
части оказывается при малых h меньше, чем при больших. Со-
противление балки изгибу зависит также от ее ширины Ь, но эта
зависимость менее существенна. Все выше сказанное можно выра-
зить одним уравнением для радиуса кривизны Р, который являет-
ся удобной мерой степени изгиба, вызываемого заданным изги-
бающим моментом Fx:
1 _ Fx
К Fl '
(14.4)
528 Часть П. Механика кровообращения
Рис 14 6 Прямая балка длиной L, толщиной h и шириной Ь, закрепленная на
одном конце Балка изгибается под действием силы, направленной вертикально
вниз А Балка до изгиба Сила Р равномерно распределена по ее верхней по-
верхности: изгибающий момент в точке закрепления равен -^PL2. Б. Балка до
изгиба Сосредоточенная в точке сила Р приложена на расстоянии х от места
закрепления изгибающий момент равен Fx. Обратите внимание на недеформи-
рованный элемент X. В. Изогнутая балка Элемент X деформирован — растянут
сверху и сжат снизу. Радиус кривизны балки R
Это уравнение показывает, что степень изгиба прямо пропорцио-
нальна изгибающему моменту и обратно пропорциональна модулю
Юнга Е. Кроме того, она обратно пропорциональна постоянной /,
которая определяется только геометрией поперечного сечения бал-
ки (т. е. ее толщиной h и шириной Ь). В случае балки прямо-
угольного сечения величина / дается соотношением
1 = -^Ыг3, (14.5)
которое, как и ожидалось, показывает, что сопротивление балки
изгибу значительно сильнее зависит от Л, чем от Ъ.
Гл. 14. Вены большого круга 529
Эти результаты можно использовать для качественного описа-
ния изгиба стенки трубки. В этом случае изгибающий момент в
любой точке поперечного сечения стенки трубки пропорционален
сжимающему трансмуральному давлению, но, кроме того, он за-
висит от локальной кривизны стенки. У концов большой оси по-
перечного сечения, где радиус кривизны минимален (в точках А
на рис. 14.5,5), он оказывается наибольшим. Различие в поведе-
нии вены и трубки из латекса (рис. 14.2) неудивительно, если
принять во внимание уравнение (14.4). Модуль Юнга исследован-
ной трубки был в 40 раз больше, чем модуль Юнга вены (2,1-
•106 Н-м-2 против 5-104 Н-м-2), в то время как толщина их стен-
ки и исходный диаметр были одинаковы. Поэтому произведение
EI для трубки было в 40 раз больше. Это объясняет как большее
сопротивление трубки изгибу — об этом свидетельствует тот факт,
что поперечное сечение ее остается круглым до тех пор, пока
трансмуральное давление не станет отрицательным, — так и мень-
шую растяжимость ее, пока поперечное сечение остается круг-
лым. Большим значением произведения EI для трубки из латекса
обусловлено и то, что после начала изгибания периметр ее не ме-
няется, в то время как периметр вены продолжает уменьшаться.
Детальное объяснение механизма возникновения гантелеобразной
формы весьма сложно. Один простой вывод, который можно сде-
лать, состоит в том, что трубка не может закрыться полностью, по-
скольку это предполагало бы нулевой радиус кривизны на концах
поперечного сечения и, следовательно, бесконечно большой изги-
бающий момент [уравнение (14.4)]. В действительности, однако,
из-за сморщивания внутренней поверхности эти концевые участки
могут быть фактически закрыты.
14.3. Динамика движения крови в крупных венах
В венах, как и в артериях, давление и кровоток меняются во
времени, а поскольку вены растяжимы, по ним распространяются
волны давления Распространение волн давления по упругим со-
судам мы уже рассмотрели (гл. 12) и теперь продолжим обсуж-
дение этого вопроса, принимая во внимание особые свойства вен:
малую толщину их стенок, высокую растяжимость и наличие кла-
панов. Чтобы проверить справедливость выводов теории распро-
странения волн для вен и предсказать характер движения крови
в них, нужны экспериментальные данные о распределении давле-
ния и его изменениях во времени на протяжении венозной си-
стемы. Мы приводили примеры записи волн давления в венуле
(рис. 13.19) и в полой вене (рис. 11.29) и поэтому можем рассчи-
тать полное падение давления в венозной системе. Но для проме-
жуточных ее областей данных почти нет. Еще меньше известно о
форме волн кровотока. Эти весьма скудные данные приведены
ниже, но их недостаточно для столь полного описания колебаний
530 Часть И. Механика кровообращения
Гл 14 Вены большого круга 531
давления и потока в венах, какое было дано в гл. 12 для артерий.
Предполагаемый характер распределения скоростей течения крови
в венах мы тоже в состоянии рассмотреть лишь очень кратко.
Существует несколько возможных причин непостоянства давле-
ния и потока в венах: 1) наличие артериальных пульсовых волн,
передаваемых к венулам через артериолы и капилляры, где они
очень сильно затухают (разд. 13.4); 2) сокращения правых камер
сердца, передаваемые к периферии по крупным венам в направле-
нии, противоположном направлению движения крови; 3) дыха-
тельные движения, которые могут очень сильно влиять на веноз-
ный кровоток из-за колебаний давления снаружи сосудов, находя-
щихся в грудной полости; 4) действие мышц, вблизи которых про-
ходят вены. Колебания очень большой амплитуды могут возникать
при нарушениях работы сердца, например при недостаточности
трехстворчатого клапана и при поперечной блокаде проводящей
системы сердца (стр. 533). Кроме того, в венах возможны само-
возбуждающиеся колебания. Ввиду их необычных механических
свойств такие колебания рассмотрены особо в разд. 14.4.
Наблюдаемые формы волн давления и кровотока. Записи волн
давления, зарегистрированных у человека в четырех местах между
правым предсердием и плечевой веной, представлены на рис. 14.7.
Все записи произведены в течение короткого промежутка времени,
поэтому вполне можно считать, что они правильно отображают
именно изменение формы волны при прохождении ее по данному
участку венозного русла. В верхней полой вене колебания выра-
жены очень сильно, их амплитуда составляет примерно 700 Н-м-2,
а форма почти совпадает с формой волн в правом предсердии. Не
может быть никаких сомнений в том, что они обусловлены пуль-
сирующим характером работы правых камер сердца. В подклю-
чичной вене форма волн давления несколько изменена, а ампли-
туда их значительно ниже — она составляет примерно 250 Н-м-2.
Здесь можно видеть и определенное отставание по фазе: пиковые
значения давления в подключичной вене достигаются позднее, чем
в верхней полой вене. Это служит свидетельством того, что волна
Рис 14 7 Волны давления, зарегистрированные последовательно при помощи
одного и того же зонда в четырех следующих друг за другом участках венозной
системы человека Места регистрации- верхняя полая веиа (Л), подключичная
вена (Б), подкрыльцовая вена (В), плечевая вена (Г). Давление измерялось
относительно атмосферного; испытуемый лежал на спине, так что все места из-
мерения находились приблизительно на одном уровне Обратите внимание на
различие средних значений давления в этих четырех участках Для совмещения
всех четырех записей давления по времени за начало отсчета времени приняты
зубцы одновременно производившихся записей ЭКГ Это позволяет выявить
сдвиг фаз между волнами давления в полой и в подключичной венах. Волны
давления «а», «с» и «V» представлены также иа рис 14 8 и обсуждаются в тексте
в связи с этим рисунком (С любезного разрешения д-ра G. М. Miller, Brompton
Hospital, London)
532 Часть 11. Механика кровообращения
давления распространяется от сердца к периферии (скорость рас-
пространения не вычислялась, так как расстояние между местами
регистрации давления неизвестно). В подкрыльцовой вене наблю-
дается дальнейшее изменение формы волны: в ней отсутствуют не-
которые высокочастотные колебания. При продвижении зонда, ис-
пользуемого для регистрации давления, вдоль этой вены выяв-
ляются резкие изменения формы волн давления, которые почти не-
сомненно связаны с наличием клапана (см. ниже). В плечевой вене
амплитуда колебаний очень мала — меньше 100 Н-м-2. Однако мы
уже видели (рис. 13.19), что в венулах брыжейки кошки ампли-
туда колебаний давления составляет примерно 400 Н-м-2, т. е.
приблизительно в 30 раз меньше амплитуды колебаний давления
в артериях большого круга. В гл. 13 было показано, что эти коле-
бания скорее всего представляют собой «следы» пульсовых коле-
баний давления в артериях, оставшиеся после затухания их в ар-
териолах и капиллярах. При некоторых условиях, например в слу-
чае особо высокой амплитуды пульсовых колебаний давления в
артериях у больных с резко выраженной недостаточностью аор-
тального клапана, в мелких периферических венах обнаруживают-
ся более сильные колебания давления такого типа. При этом за-
тухание волн в системе микрососудов оказывается недостаточным,
чтобы уменьшить пульсовые колебания до обычного уровня.
Следует еще раз подчеркнуть, что волны, изображенные на
рис. 14.7, были зарегистрированы в различных участках одного
конкретного венозного пути. По всей вероятности, распределение
давления вдоль разных путей в венах может различаться значи-
тельно сильнее, чем в артериях. Для такого суждения имеются
два основания: первое — наличие в венах клапанов, которые, как
мы покажем, должны оказывать очень большое влияние на пуль-
совые колебания давления; второе — тот факт, что в зависимости
от величины трансмурального давления в некоторых путях вены,
вероятно, должны находиться в спавшемся состоянии. Растяжи-
мость таких вен, а следовательно, и скорость распространения по
ним волн будут иными, чем для полностью открытых вен, и потому
спавшиеся вены представляют собой еще и объект, сильно отра-
жающий волны.
Форма волны скорости крови в полых венах человека изобра-
жена на рис. 14.8. Как видно из рисунка, в течение каждого
цикла сокращений сердца происходят два основных колебания
скорости крови, причем оба колебания оказываются совершенно
не в фазе с колебаниями давления. Во время сокращения правого
предсердия, соответствующего волне «а» на записи давления в
нем, кровоток в полых венах сильно уменьшен и его направление
на короткое время может даже измениться на обратное. Затем
скорость крови в этих венах увеличивается, и нарастание ее про-
должается во время волны «с» и сокращения желудочков. Макси-
мальная скорость достигается во время систолы желудочков. По-
Гл. 14. Вены большого Круга 533
Рис. 14.8 Совмещенные по времени волны давления и скорости крови на входе
в правые камеры сердца и на выходе из них; диаграмма составлена по данным
ряда различных измерений. Верхние кривые — скорость крови в легочной арте-
рии (/), нижней полой вене (II) и верхней полой вене (III)', затем идут записи
давлений в легочной артерии и в правых камерах сердца — в правом предсер-
дии (/V) и правом желудочке (V), а ниже—фонокардиограмма (V/) и ЭКГ
(VII). Последние записи приведены для привязки остальных записей по времени.
Т/3 — трехстворчатый клапан закрыт; Л/О—легочный клапан открыт; Л/3 — ле-
гочный клапан закрыт; Т/О — трехсгворчатый клапан открыт. [Wexler, Bergel,
Gabe, Makin, Mills (1968). Velocity of blood flow in normal human venae cavae.
Circulation Res., 23, p. 352. С любезного разрешения American Heart Association
Inc.]
вторно увеличение скорости наблюдается в начале диастолы же-
лудочков, во время спада «у» давления в правом предсердии, ко-
торый следует за волной «V» и возникает при открытии атрио-вент-
рикулярных клапанов. Если кровь может поступать через трех-
створчатый клапан обратно в правое предсердие или если прове-
дение возбуждения по проводящей системе сердца нарушено, так
что предсердия сокращаются во время систолы желудочков, когда
атрио-вентрикулярные клапаны закрыты (так называемая бло-
када сердца), колебания кровотока в полой вене могут быть зна-
534 Часть II. Механика кровообращения
чительно большими. Форму волн кровотока в периферических ве-
нах пока еще не регистрировали.
Распространение волн в венах. Эксперименты, в которых пуль-
совые колебания давления регистрировались бы одновременно в
двух участках крупной вены, весьма немногочисленны, поэтому
сведений о скорости распространения и о затухании пульсовой
волны в венах очень мало. Те данные, которыми мы располагаем,
показывают, что скорость распространения пульсовой волны давле-
ния в краниальной полой вене собаки составляет от 0,5 до
3,0 м-с-1. Почти такой же разброс получается и при определении
скорости распространения разных частей (вершины, подножия и
т. д.) пульсовой волны давления у одного и того же животного.
Этот разброс, вероятно, связан либо с изменением венозного дав-
ления во время прохождения волны (разд. 12.5), либо с отраже-
нием волн, которое изменяет кажущуюся скорость их распростра-
нения (разд. 12.3). В нашем распоряжении есть еще только два
источника информации: во-первых, оценки, основанные на данных
о механике распространения волн в артериях, и, во-вторых, ре-
зультаты одной группы опытов на брюшном отделе каудальной
полой вены собаки. В этих опытах на одном участке сосуда искус-
ственно создавали короткую серию высокочастотных синусоидаль-
ных волн давления, а на другом участке эти волны регистриро-
вали. Как и в аналогичных опытах на артериях (разд. 12.4), из-
мерения проводили лишь для тех волн, которые появлялись в точ-
ке регистрации раньше, чем к ней возвращались первые отражен-
ные волны.
Из приближенной теории, представленной в общих чертах в
гл. 12 (разд. 12.3), следует, что волны давления малой амплитуды
распространяются по упругой трубке со скоростью с, определяе-
мой уравнением (12.9):
с = (рО)“1/2, (14.6)
где р — плотность крови, a D — растяжимость сосуда [см. уравне-
ние (14.1)]. При растягивающем давлении порядка ±0,5-103 Н-
•м-2 растяжимость грудного отдела полой вены собаки макси-
мальна и составляет примерно (2—2,5) • 10_3 м2-Н-1 (рис. 14.1, Д),
а при больших значениях трансмурального давления она много
меньше (при ртм = 3-103 Н-м-2 растяжимость равна примерно
4-10-5 м2-Н-1 и при дальнейшем увеличении ртм уменьшается).
При больших отрицательных значениях трансмурального давле-
ния растяжимость также становится очень малой: при ртм = —2-
-103 Н-м-2 она равна 10-4 м2-Н-1. Плотность крови составляет
примерно 103 кг-м-3, и, таким образом, предсказываемая теорией
скорость распространения волн давления меняется от 5 м-с-1 или
больше при высоком трансмуральном давлении до минимального
значения около 0,6 м-с-1 при спадении вены; в условиях полного
спадения вены эта скорость снова возрастает примерно до 3 м-с-1.
Гл. 14. Вены большого круга 535
Рис. 14.9. Зависимость скорости волны с низкоамплитудных синусоидальных волн
давления в каудальной полой вене собаки от трансмурального давления рТм-
Разные значки соответствуют волнам разной частоты, квадратики — 30 Гц, тре-
угольники 40 Гц, кружки —50 Гц. [Anhker, Wells, Ogden (1969). The transmis-
sion characteristics of large and small pressure waves in the abdominal vena cava,
p, 271, Trans on Bio-med. Eng., BME-16, Institute of Electrical and Electronics
Engineers Inc, New York.]
Еще один вывод можно сделать из данных, представленных на
рис. 14.3, Б и 14.4. Подставив в уравнение (14.3) значения модуля
Юнга, приведенные на рис. 14 35, и значение отношения толщины
стенки к диаметру, равное 0,01 (рис. 14.4), можно оценить растя-
жимость вены, а затем из уравнения (14.6) определить скорость
волны. Получаемые таким способом значения лежат в интервале
приблизительно от 1 м-с-1 при трансмуральном давлении 0,5-
-103 Н-м-2 до 9 м-с-1 при трансмуральном давлении 104 Н-м-2.
Эти расчеты показывают, что при низких трансмуральных дав-
лениях (меньше, чем примерно 3-103 Н-м-2) скорость волны в ве-
нах ниже, чем в крупных артериях. Но когда трансмуральное
давление в венах приближается к 104 Н-м-2 (как в стопе стоя-
щего человека), скорость волны в вене может достичь такого же
значения (и даже больше), как в соответствующей артерии, хотя
давление в артерии превышает давление в вене примерно на
104 Н-м-2. (Этот вывод, возможно, имеет чисто теоретическое зна-
чение, поскольку, как мы видели, в вены конечностей пульсации
обычно не передаются.)
Регистрацию высокочастотных синусоидальных волн давления
с малой амплитудой осуществляли в брюшном отделе полой вены
собак при трансмуральных давлениях от 0,5-103 Н-м-2 до 2,5-
-103 Н-м-2. Результаты, полученные для одной собаки при раз-
личных частотах, показаны на рис. 14.9; видно, что в этом диапа-
зоне давлений скорость распространения волны меняется от 2 м-
’С-1 до 6 м-с-1. Для разных собак разброс данных рказдлся рерь-
536 Часть II. Механика кровообращения
ма большим: при трансмуральном давлении 103 Н-м-2 скорость
волны менялась от значений, меньших 1 м-с-1, почти до 3 м-с-1.
Хотя использованные частоты (20 Гц или выше) были слишком
велики по сравнению с физиологически возможными, эти значения
скорости волн охватывают именно тот диапазон, который теорети-
чески предсказан для данного трансмурального давления. При по-
нижении частоты измеренные значения скорости волны несколько
снижались. На сосудах, форма поперечного сечения которых су-
щественно отличалась от круглой, подобные опыты не проводились.
Поэтому данных, позволяющих проверить теоретический вывод о
быстром изменении скорости волны при изменении трансмурально-
го давления в диапазоне очень малых его значений, не существует.
В рассмотренных выше опытах короткие серии высокочастот-
ных колебаний использовали специально для того, чтобы избежать
искажений, вызываемых отражением волн. Но in vivo отражения
неизбежно возникают всюду, где изменяются свойства сосуда. Это
могут быть места соединения с другими венами, клапаны, а также
области, в которых меняется трансмуральное давление, а следо-
вательно, и площадь поперечного сечения и растяжимость (приме-
ром служит область входа вены в грудную полость, где она может
спадаться). Экспериментальные работы, в которых исследовали
бы отражение и прохождение пульсовой волны в каком-либо из
такого рода мест, весьма немногочисленны, и мы знаем о веноз-
ном русле слишком мало, чтобы пытаться делать какие-нибудь
четкие выводы. Из наблюдений известно, однако, что в сред-
ней по калибру вене руки давление фактически не пульсирует
(рис. 14.7), и остается постоянным даже при кашле, когда цен-
тральное венозное давление испытывает флуктуации, достигающие
1,3-103 Н-м-2 (100 мм рт. ст.). Это означает, что волна почти пол-
ностью отражается раньше, чем достигает вен среднего калибра
(как и в артериях, эффекты, обусловленные вязкостью и вязко-
упругостью, не могли бы на таких расстояниях вызвать столь
сильное затухание волны; см. разд. 12.6). Полное отражение, ве-
роятно, обусловлено наличием клапанов, так как у больных с не-
полноценными клапанами [у которых может быть, кроме того, и
варикозное расширение вен (разд. 14.5)] в периферических венах
часто можно зарегистрировать кратковременные изменения давле-
ния, возникающие при повышении давления в грудной или в брюш-
ной полости (например, при кашле).
Можно умозрительно рассмотреть некоторые механизмы, су-
щественные для отражения волн от области венозного клапана.
Опыты на модели показали, что, когда пульсовая волна давле-
ния приходит со стороны сердца к уже закрытому клапану, створ-
ки клапана, упруго прогибаясь, выпячиваются, но количество кро-
ви, протекающее через него в обратном направлении, пренебре-
жимо мало. Поэтому через клапан передается лишь малая часть
Энергии волны, а большая ее часть отражается, как в трубке с
Гл. 14. Вены большого круга 537
закрытым концом (разд. 12.4). Когда же пульсовая волна давле-
ния приходит к открытому клапану, через который в это время
кровь движется по направлению к сердцу, положение оказывается
более сложным. Сначала должно измениться направление гра-
диента давления, а затем начать закрываться клапан, по-види-
мому, под действием тех же механизмов, которые приводят к за-
крытию аортального клапана (разд. 11.5). Может оказаться, что
времени, в течение которого градиент давления направлен в об-
ратную сторону, недостаточно для полного закрытия клапана до
того, как направление (знак) градиента давления изменится еще
раз и он опять будет направлен к сердцу. Таким образом, в отли-
чие от условий отражения от места соединения сосудов (разд. 12.4)
особенности процессов в области венозного клапана, а именно ха-
рактер отражения волны и характер движения крови по обе сто-
роны клапана, зависят от временных параметров пульсовой волны,
а следовательно, и от частоты следования пульсовых волн.
При экспериментальном определении скорости волны в венах
амплитуда синусоидальных колебаний давления была меньше
0,2-103 Н-м'2. В большинстве случаев это составляло лишь не-
большую долю от среднего трансмурального давления, и регистри-
руемые волны оставались синусоидальными. Однако амплитуда
нормальной пульсовой волны может вдвое превышать указанную
величину, а при некоторых патологических состояниях быть еще
больше. Кроме того, в отдельных венах среднее значение трансму-
рального давления лежит в пределах ±0,5-103 Н»м-2. При таких
давлениях вены должны быть особенно растяжимы, и при измене-
нии давления на 0,4-103 Н-м-2 их форма и площадь поперечного
сечения будут меняться очень сильно. Тогда прохождение пульсо-
вой волны давления вызовет значительные изменения поперечного
сечения сосуда. В то же время максимальная скорость крови, рав-
ная примерно 0,25 м-с-1 (рис. 14.8), будет составлять почти по-
ловину расчетной скорости волны (0,6 м-с-1 — см. стр. 534). Как
было выяснено при обсуждении вопроса о волнах давления в ар-
териях (разд. 12.5), оба этих обстоятельства могут привести к
тому, что в распространении волн важную роль будут играть нели-
нейные эффекты.
Наиболее примечательное проявление нелинейности состоит в
увеличении крутизны фронта волны давления. При определенных
условиях он может превратиться в резкий скачок давления, ана-
логичный фронту ударной волны, и прохождение его будет сопро-
вождаться быстрыми изменениями поперечного сечения сосуда.
Прямым доказательством существования в венозной системе та-
ких «ударов» служит регистрация в венах тонов типа «пистолет-
ного выстрела». Такие тоны можно прослушивать в бедренной и
яремной венах у больных с пороком трехстворчатого клапана.
Тоны возникают во время сокращения правого желудочка, когда
значительная часть крови из его полости возвращается у таких
538 Часть 11. Механика кровообращения
Рис. 14.10. Изменение формы искусственно созданных волн давления с частотой
7,5 Гц при прохождении их по брюшному отделу полой вены собаки. На каждой
паре записей представлены волны одной исходной амплитуды, причем нижние
записи произведены на 4 ем дистальнее, чем верхние, и при большем коэффи-
циенте усиления. Амплитуды волн указаны слева от записей Обратите внимание,
что по мере увеличения амплитуды волны все более искажаются. [Anliker, Wells,
Ogden (1969). The transmission characteristics of large and small pressure waves
in the abdominal vena cava, p. 268, Trans on Bio-med. Eng., BME-16, Institute
of Electrical and Electronics Engineers Inc., New York.]
больных в правое предсердие и в полые вены; это приводит к
сильному увеличению амплитуды венозного пульса (ср. с арте-
риальными «пистолетными выстрелами», описанными в разд. 12.5).
Увеличение крутизны фронта волн давления в венах также иссле-
довали путем создания в полой вене собаки искусственных высо-
кочастотных волн давления. Волны регистрировали в двух местах,
удаленных друг от друга на 4 см; амплитуду накладываемых ко-
лебаний меняли от 0,46-103 до 1,3-103 Н-м-2 (3,5—10 мм рт. ст.);
частота колебаний равнялась 7,5 Гц. Результаты экспериментов
показаны на рис. 14.10. Несинусоидальный характер волн, а также
постепенное увеличение крутизны фронта по мере продвижения их
по вене очевидны. Увеличение крутизны фронта происходит не-
смотря на значительное затухание, проявляющееся в уменьшении
амплитуды волн, регистрируемых на некотором удалении от ис-
точника. Другое возможное проявление нелинейности заключается
в том, что разные по частоте составляющие волн градиента дав-
ления, профиля скорости и скорости крови могут уже не быть не-
зависимыми, а их взаимодействие приведет к изменению средних
значений этих величин (разд. 8.5). Последнее означает, что оцен-
Гл. 14. Вены большого круга 539
ки средней скорости кровотока, сделанные на основании данных о
средней величине градиента давления, могут оказаться не соответ-
ствующими действительности. Однако никакими эксперименталь-
ными данными по этому вопросу мы не располагаем; в артериях
такой эффект, вероятно, несуществен.
Обратимся, наконец, к вопросу о затухании волн давления
в венах. Как и всякие другие свойства волн, затухание здесь изу-
чено для искусственно созданных высокочастотных волн, и, как
можно видеть из рис 14.10, оно весьма значительно. Измерение
Рис. 14.11. Затухание низкоамплшудных волн давления в каудальной полой веие
собаки. По оси ординат отложено отношение амплитуды волны а к амплитуде
ее по в некотором фиксированном месте; по оси абсцисс — расстояние от этого
фиксированного места до расположенного дистальнее места измерения (х), де-
ленное на длину волны (X). Прямая на этом графике, построенном в полулога-
рифмическом масштабе, определяется уравнением а/йо = е~kxl/* при k = 1,05.
Разные значки относятся к волнам разных частот. Трансмуральное давление со-
ставляло (1,0—1,5)-103 Н-м-2. [Anliker, Wells, Ogden (1969). The transmission
characteristics of large and small pressure waves in the abdominal vena cava,
p. 271, Trans, on Bio-med. Eng., BME-16, Institute of Electrical and Electronics
Engineers Inc, New York.]
амплитуды малых синусоидальных волн давления в двух местах,
удаленных друг от друга на различные расстояния, показывает,
что, как и в артериях (разд. 12.6), в венах амплитуда волн умень-
шается с расстоянием экспоненциально. Изучение затухания волн
с различными частотами показало, что для частот больше 20 Гц
степень затухания в расчете на длину волны (т. е. уменьшение
амплитуды волны при прохождении ею расстояния, равного одной
длине волны) не зависит от частоты (см. рис. 14.11). Иными сло-
вами, амплитуда волны а равна аое~кх1к, гдех— расстояние вдоль
сосуда, ао — амплитуда волны при х = 0, X — длина волны, k —
постоянная. Для полой вены собаки значение k лежит в пределах
1,0—2,5 (затухание составляет 63—92% на длине волны), а для
аорты (см. рис. 12.37)—в пределах 0,7—1,0 (затухание 50—63%
на длине волны). Таким образом, степень затухания высокочас-
640 Часть II. Механика кровообращения
тотных волн в полой вене больше, чем в аорте. При частоте серд-
цебиений 2 Гц пульсовая волна давления в вене, распространяю-
щаяся со скоростью 2 м-с-1, на расстоянии 30 см (что примерно
соответствует длине брюшного отдела полой вены собаки) будет
затухать на 20—30% • Такое затухание не может быть целиком
обусловлено вязкостью крови, потому что, как и в случае артерий,
значение параметра а в таких опытах велико (разд. 12.3) и потери
энергии в вязких пограничных слоях относительно малы. Следова-
тельно, в действительности затухание должно быть обусловлено
главным образом тем, что стенка сосудов обладает вязкоупругими,
Рис. 14.12. Проявление гистерезиса в аенах. Кривая давление — объем для сег-
мента каудальной полой вены собаки при синусоидальном изменении объема с
частотой 5 Гц. Во время увеличения объема давление в сегменте вены выше,
чем во время уменьшения, части кривой, отражающие увеличение и уменьшение
объема, помечены стрелками, направленными соответствующим образом. (На
графике вместо площади поперечного сечения указан объем, поскольку длина
сегмента веиы неизвестна). [Attinger (1969). Wall properties of veins, p. 256,
Trans, on Bio-med. Eng., BME-16, Institute of Electrical and Electronics Engineers
Inc., New York.]
а не чисто упругими свойствами. Вены проявляют заметные вяз-
коупругие свойства также при несколько более низких (чем в опы-
тах по измерению постоянной k) частотах. Например, при синусои-
дальном изменении объема сегмента вены с частотой 5 Гц (значе-
ние а в этих условиях еще велико — около 14) был обнаружен
гистерезис (разд. 7.2): во время увеличения объема давление в
сегменте было выше, чем при таких же значениях объема во время
его уменьшения (рис. 14.12).
Картины течения крови и профили скорости в венах. Средняя
скорость крови в каудальной полой вене собаки составляет 10—
20 см-с-1, а диаметр вены — около 1 см (табл. I), так что среднее
число Рейнольдса лежит между 250 и 500, а максимальное со-
ставляет примерно 700. В более мелких венах число Рейнольдса,
конечно, меньше. Другим важным безразмерным параметром яв-
ляется зависящий от частоты параметр Уомерсли а (см. разд. 5.6).
Наибольшее значение этот параметр имеет в случае полой вены —
при частоте сокращений сердца 2 Гц он равен примерно восьми.
Для более мелких вен его значение меньше.
Зная эти два параметра, мы можем представить в общих чер-
тах характер течения крови в крупных венах в условиях, когда их
Гл. 14. Вены большого круга 541
поперечное сечение остается более или менее круглым. 1. Мы по-
лагаем, что течение должно быть ламинарным, потому что даже
пиковое число Рейнольдса в этом случае много меньше его кри-
тического значения (примерно 2300) для стационарного течения.
2. Длина начального участка для среднего (по времени) течения
крови в наиболее крупных венах должна быть большой, потому
что соответствующее среднее число Рейнольдса достаточно велико
(предполагается, что средние скорости не зависят от колебатель-
ных составляющих, а это может быть неверно — см. разд. 12.8).
Однако длина начального участка не будет превышать длину со-
суда, как это имеет место в случае аорты. Эта длина в каудаль-
ной полой вене собаки в соответствии с уравнением (5.4) равна
7,5—15 см. Наличие вторичных течений в потоке при входе в вену
может привести к некоторому снижению этих оценок из-за повы-
шения интенсивности поперечного перемешивания слоев жидкости,
движущихся более медленно и более быстро. В результате тече-
ние может развиться быстрее. Поэтому, даже если в области входа
потока из приводящих вен в рассматриваемый сосуд имеются тон-
кие пограничные слои, средняя составляющая течения должна
почти полностью развиться раньше, чем достигнет сердца, т. е. на
расстоянии от входа менее 30 см. 3. Поскольку значение а доста-
точно велико, «колебательные» пограничные слои у стенки сосуда
останутся тонкими (разд. 5.6) и пульсирующее течение вблизи оси
сосуда (в ядре потока) будет иметь плоский профиль скоростей
в отличие от выпуклого профиля полностью развитого среднего
течения. 4. В соответствии с уравнением (12.28) длина начального
участка для нестационарной составляющей течения будет неболь-
шой — около 4,5 см.
Помимо влияния клапанов, о котором известно очень мало,
важным отличием венозного потока от артериального является то,
что первый направлен от более мелких сосудов к более крупным.
Поэтому течение в области соединения венозных сосудов отли-
чается по характеру от наблюдаемого в области разветвления ар-
териальных сосудов (разд. 12.8). В области соединения два по-
тока объединяются таким образом, что у профиля скорости (в пло-
скости соединения) сразу возникает провал в центре. Это обуслов-
лено тем, что в каждом подводящем сосуде точка, где скорость
течения максимальна, не лежит на оси объединяющего сосуда. Оба
потока поворачиваются, при этом возникают вторичные движения
(разд. 5.8), приводящие к образованию двух пар вторичных вих-
рей. Именно такая картина получается при пропускании дыма че-
рез модель бифуркации (рис. 14.13Д). Исследование в опытах на
моделях профиля скорости в общей трубке показывает, что погра-
ничные слои здесь остаются тонкими и, за исключением области,
расположенной вблизи места соединения потоков, профиль скоро-
сти почти плоский (рис. 14.13S). По-видимому, это обусловлено
характером перестройки продольного течения за счет влияния вто-
S42 Часть П Механика кровообращения
Относительной расстояний ho радиусу
Гл. 14. Вены большого круга 543
ричных движений. Поведение нестационарных потоков в области
таких соединений не изучено. Нам неизвестны какие-либо систе-
матические исследования профиля скорости даже в наиболее круп-
ных венах. Однако сквозь тонкую стенку нижней полой вены,
вниз по потоку от места слияния подвздошных вен, вторичные
движения видны вполне отчетливо.
14.4. Течение в трубках, способных спадаться
В условиях, когда сосуд может спадаться и его поперечное
сечение становится существенно не круглым, наблюдается ряд ин-
тересных явлений, а связь между градиентом давления и крово-
током очень сильно усложняется. Поэтому течению в спадающих-
ся трубках посвящен специальный раздел.
При очень низком трансмуральном давлении в вене свойства
потока крови становятся весьма необычными. Обнаружено, что,
когда давление в сосуде меньше, чем снаружи, снижение давления
внутри него на выходе мало влияет (или даже не влияет вообще)
на скорость кровотока. Такой же эффект можно наблюдать на
модели — при протекании воды по тонкостенной резиновой трубке.
Это явление широко изучалось, правда, большинство работ вы-
полнено не на кровеносных сосудах, а на тонкостенных трубках
из латекса. Ввиду различия механических свойств вен и таких
трубок (рис. 14.2) переносить результаты модельных исследований
на реальные сосуды следует с осторожностью. Мы начнем с опи-
сания опыта на модели, а затем разберем возможные объяснения
его результатов. В заключение мы обсудим вопрос о том, где
именно в системе кровообращения могут возникать такие явления
и наблюдаются ли они в действительности.
Опыты на модели. Установка для исследований схематически
изображена на рис. 14.14. Участок способной спадаться трубки
прикреплен по концам к двум жестким трубкам такого же диа-
метра и помещен в камеру, давление в которой рк можно изменять
контролируемым образом. По трубке течет вода, и статическое
давление измеряется в точках, расположенных выше и ниже спо-
Рис. 14 134 Вид с торца общей трубки модели симметричной бифуркации при
установившемся течейии по ней воздуха, поступающего из подводящих трубок.
Картина течения визуализирована путем введения в воздух дыма Видны две
пары вторичных вихрей Число Рейнольдса для потока в общей трубке равно 700.
[Schroter, Sudlow (1969) Flow patterns in models of the human bronchial air-
ways, Respiration Physiology, 7, pp 341—355, North-Holland Publishing Co, Am-
sterdam ]
Рис 1413Б Профили скорости в двух плоскостях общей трубки модели сим-
метричной бифуркационной системы, состоящей из четырех поколений подводя-
щих трубок Воздух протекает из подводящих трубок в общую равномерно
Число Рейнольдса для потока в общей трубке равно 500 Обратите внимание,
насколько плоским является профиль и как тонок пограничный слой.
544 Часть II. Механика кровообращения
собной спадаться трубки. Эти давления обозначены соответственно
через pi и р2. За местом отвода для измерения давления р2 вода
вытекает в атмосферу через сопротивление фиксированной вели-
чины (выходное сопротивление), так что давление р2 превышает
атмосферное давление на величину, пропорциональную расходу
жидкости Q. Расход можно изменять, регулируя либо высоту рас-
положения резервуара, из которого вода поступает в систему, либо
величину сопротивления между резервуаром и отводом для из-
мерения давления pi (входного сопротивления). Кроме того, рас-
ход воды можно регулировать, изменяя выходное сопротивление.
Атмосфера
Рис 14 14 Схема установки для изучения движения жидкости в трубке, способ-
ной спадаться Трубка присоединена к жестким трубкам такого же диаметра и
помещена в закрытую камеру Давление в камере (рк) можно изменять незави-
симо от давления в трубке, предусмотрена возможность измерения давления до
и после способного спадаться сегмента (pi и рг). Расход жидкости в системе (Q)
можно менять, регулируя либо сопротивление на выходе системы, открываю-
щемся в атмосферу, либо сопротивление на входе, перед соединением системы с
питающим ее резервуаром Жидкость движется справа палево
Последствия изменения расхода разными способами несколько
различаются и будут рассмотрены дальше.
Результаты, на которых мы остановимся прежде всего, были
получены при фиксированном давлении в камере. Расход снижали,
увеличивая входное сопротивление. Результаты представлены
в виде графика зависимости перепада давления (pi — р2) от рас-
хода Q (рис. 14.15,Л). Числа вдоль кривой соответствуют номе-
рам фотографий на рис. 14.15,5, на которых показано состояние
(вид сбоку) способного спадаться участка трубки на разных ста-
диях опыта (поток направлен справа налево). Подчеркнем, что
для преодоления вязкого сопротивления течению жидкости р\
должно быть всегда больше р2. Результаты отчетливо разделяются
на три категории.
1. Если расход достаточно велик для того, чтобы Давление на
выходе из трубки р2 было больше, чем давление в камере рк, то
давление внутри способного спадаться участка везде оказывается
больше рк и трубка остается почти круглой (фотографии 1—6 на
рис. 14.15,5). При этом в любом участке трубки течение пуазей-
Гл. 14. Вены большого кру->а 545
Направление
Рис* 14.15. А. Зависимость перепада давления pi—рз) от расхода Q в модели, подобной пока-
занной на рис. 14 14. Изменения Q вызывали, меняя входное сопротивление, выходное сопро
тивленне и давление в камере (рк=3,3 • 103 Н • м“2) поддерживали постоянными. Числа
вдоль кривой—порядковые номера фотографий, приведенных иа Б. Три режима течения,
обозначенные цифрами I, II и III, описаны в тексте. Б. Вид сбоку на способный спадаться
сегмент трубки на разных стадиях опыта, номера соответствуют помеченным числами точкам
графика на А. Течение направлено справа налево [А и В—из Conrad (1969). Pressure-flow
relationships in collapsible tubes, pp 284—295, Trans, on Blo-med. Eng., BME-16, Institute of
Electrical and Electronics Engineers Inc , New York 1 В. Три кривые, аналогичные кривой А,
которые получены при трех разных значениях выходного сопротивления; давление в камере
поддерживали постоянным (рк=3,9 • 10s Н • м“2). Каждая кривая —непрерывная запись,
полученная при постепенном увеличении расхода; в некоторых случаях при режиме тече-
ния II возникали самовоз буж дающиеся колебания, что видно из кривых. Аналогичные кривые
получаются и в том случае, когда выходное сопротивление остается постоянным, а давление
в камере изменяетси jKatz Chen, Moreno (1969) Flow through a collapsible tube. В lophys. J., ®,
pp. 1261-1279 Имеется перевод. См. кн.: Гидродинамика кровообращения, М., Мир, 1970,
с. Hi—130 I Г. Зависимость расхода Q от давлении на выходе р2, отнесенного к давлению
в камере рк. Когда р2 меньше Рк, расход не зависит от р2. [Holt (1969). Flow through collap-
sible tubes and through in situ veins, pp 274 —283, Тгапз. on Blo-med. Eng., BME-16, Institute
of Electrical and Electronics Engineers Inc., New York. J
546 Часть 11. Механика кровообращения
левское, сопротивление почти постоянное и в соответствии с зако-
ном Пуазейля [уравнение (5.1)] падение давления пропорцио-
нально расходу. Это и отображает правая, обозначенная цифрой I,
часть кривой на рис. 14.15, Л. Сопротивление не является абсо-
лютно постоянным, так как растяжимость трубки хотя и мала,
но все-таки отлична от нуля (рис. 14.2,Л), и при снижении рас-
хода диаметр трубки несколько уменьшается.
2. Когда расход меньше некоторого критического значения,
давление на выходе из трубки оказывается ниже давления в ка-
мере. В этом случае при очень небольшом дальнейшем снижении
расхода трансмуральное давление на выходе способной спадаться
трубки становится отрицательным, и трубка начинает менять фор-
му своего поперечного сечения и спадаться. Поскольку растяжи-
мость трубки в этом случае велика (рис. 14.2,Л), площадь попе-
речного сечения при снижении трансмурального давления быстро
падает. По мере спадения трубки сопротивление течению быстро
растет, и перепад давлений, необходимый для поддержания рас-
хода (который постепенно снижается), также резко увеличивает-
ся. Это отражает средняя часть кривой II на рис. 14.15, Л. Посте-
пенное спадение трубки иллюстрируют фотографии 7—16 на
рис. 14.15,5. Сначала (7 и 8) трубка сужается вблизи выхода (где
давление внутри трубки минимально и близко к р2) и частично
спадается. По мере дальнейшего уменьшения расхода область спа-
дения трубки расширяется в направлении входа (9—16), пока в
конце концов не спадается весь гибкий участок (16). На этой стадии
давление на входе щ оказывается приблизительно равным давле-
нию в камере рк. Если в это время менять выходное сопротивле-
ние, меняя тем самым давление р2 независимо от давления pi, из-
менение расхода оказывается небольшим. Но если вместо этого
менять давление в камере, то изменяется степень спадения труб-
ки, а в соответствии с этим и расход. Фактически при таких воз-
действиях расход Q оказывается приблизительно пропорциона-
лен разности давлений щ — рк, а не щ — р2.
3. Наконец, при спадении всего участка (давление на входе
меньше давления в камере) поперечное сечение принимает уже
описанную в разд. 14.2 довольно жесткую гантелеобразную форму.
Поэтому при дальнейшем снижении расхода никакого дополни-
тельного изменения поперечного сечения не происходит (фотогра-
фии 17—20) и сопротивление течению снова становится постоян-
ным, в результате чего перепад давления снижается пропорцио-
нально расходу (участок кривой III на рис. 14.15,А). Однако из-за
очень малого просвета каналов, через которые должна протекать
жидкость, сопротивление оказывается в 10—100 раз больше, чем
до спадения.
Если на какой-либо стадии течение полностью остановить, пе-
рекрыв отток на выходе, то по всей трубке внутреннее давление
станет одинаковым и величина его будет определяться уровнем
Гл. 14. Вены большого круга 547
расположения находящегося на входе резервуара. Если при этом
Pi окажется меньше, чем давление в камере, трубка останется
спавшейся, но если pi превысит давление в камере, то трубка
опять раскроется.
Каковы бы ни были числа Рейнольдса в потоке через трубку,
последовательность наблюдаемых событий оказывается качествен-
но одной и той же. Но при достаточно больших числах Рейнольдса
возникает еще одно интересное явление: при определенных усло-
виях, когда значения давления в камере и выходного сопротивле-
ния, а также уровень положения резервуара поддерживаются по-
стоянными, в системе могут возникать самовозбуждающиеся коле-
бания. Указанные условия предопределяют некоторое постоянное
значение расхода, ио если оно лежит в пределах участка II
(рис. 14.15, Л), то возникают спонтанные колебания поперечного
сечения трубки, перепада давления и расхода. (При наличии та-
ких колебаний график на рис. 14.15, Л отражает средние значения
расхода и перепада давления.) Если сопротивление на входе уве-
личить, понизив тем самым средний расход, то частота колебаний
увеличится. В опытах, из описания которых взят рис. 14.15, на-
блюдались колебания с частотой в пределах от 0,1 до 1 Гц. Чем
определяется значение этой частоты, не вполне ясно; возможные
механизмы возникновения колебаний обсуждаются ниже.
Следует еще раз подчеркнуть, что все результаты, представ-
ленные на рис. 14.15, Л, были получены при фиксированных зна-
чениях выходного сопротивления и давления в камере. При изме-
нении любой из этих величин получается иная кривая. Все такие
кривые имеют сходную форму и сливаются-в области I, т. е. когда
трубка полностью открыта. Но если уменьшить давление в камере
или увеличить выходное сопротивление, то наклон кривой на
участках II и III уменьшится, так же как и максимум перепада
давления (рис. 14.15,В). Отсюда ясно, что перепад давления, не-
обходимый для поддержания заданного значения среднего рас-
хода, зависит от давления в камере и выходного сопротивления.
Если сопротивления на входе и на выходе поддерживаются по-
стоянными, а давление в камере повышается начиная от значения,
равного атмосферному, то как расход, так и перепад давлений
Pi — Pz не изменяются до тех пор, пока давление в камере не пре-
высит давление на выходе. После этого расход начинает падать
пропорционально величине р\ — р2, и это падение продолжается
до тех пор, пока трубка не спадется полностью; тогда расход
становится небольшим, но более или менее постоянным. Аналогич-
ным образом, если снижать выходное сопротивление при фиксиро-
ванных значениях давления в камере (выше атмосферного) и со-
противления на входе, давление на выходе будет сначала падать,
а следовательно, перепад давления pi — р2 и расход будут расти.
Но стоит давлению на выходе упасть ниже давления в камере,
как трубка начнет спадаться, расход перестанет расти и в конеч-
548 Часть II. Механика кровообращения
ном счете примет значение, определяемое величиной pi — рк
(рис. 14.15, Г).
Механизмы. Легко понять, почему сопротивление гибкой труб-
ки при спадении увеличивается. Действительно, спадение трубки
сопровождается уменьшением площади ее поперечного сечения,
так что при фиксированном расходе средняя скорость увеличи-
вается, и соответственно вязкое напряжение сдвига на стенке,
«сдерживающее» движение жидкости, возрастает. Площадь по-
верхности, на которой действует это напряжение сдвига, остается
прежней, потому что периметр трубки почти не меняется. Напря-
жение сдвига увеличивается еще и вследствие сплющивания труб-
ки, так как при этом в самой узкой ее части возникают высокие
скорости сдвига. Таким образом, заданный расход может поддер-
живаться лишь за счет увеличения перепада давления. Представ-
ление о том, насколько должен увеличиться перепад давления,
можно получить из теории полностью развитого вязкого течения
в прямой трубке эллиптического сечения, большая и малая оси
которого равны 2а и 2Ь. В этом случае отношение градиента дав-
ления к расходу (сопротивление течению) определяется формулой
я., g2 + j>2 Н4 7\
8,1 2as*3rt ’ I14-')
где у,— вязкость жидкости. Если поперечное сечение круглое
(а = 6), то коэффициент (а2 + &2)/2а363 равен 1/а4 и уравнение
(14.7) сводится к закону Пуазейля [уравнение (5.1)]. Влияние на
этот коэффициент уменьшения площади эллипса при сохранении
постоянным его периметра иллюстрирует рис. 14.16, Л. График за-
висимости сопротивления трубки с эллиптическим сечением от
площади сечения сопоставлен на рисунке с таким же графиком
для круглой трубки. Видно, что по мере уменьшения площади
сечения рост сопротивления, обусловленный увеличением эллип-
тичности, все более и более ускоряется. Заметим, что уменьшение
отношения малой оси к большой (b/а) до 0,75 приводит к увели-
чению сопротивления трубки всего на 10% (рис. 14.16,5).
Эти теоретические выводы получены в предположении, что
трубка с эллиптическим сечением имеет неизменную по длине фор-
му. В случае местного сужения, во всяком случае при больших
числах Рейнольдса (около 100 и выше), начинают действовать
другие факторы и приведенные результаты могут оказаться не-
верными. В таких условиях поток, ускоренный при прохождении
через суженный участок, в области, где площадь начинает снова
увеличиваться, может отделиться от стенок трубки, что приведет
к возникновению ниже по потоку течения, похожего на струю
(разд. 5.7). Появление такого отрывного течения всегда сопровож-
дается большими потерями энергии и большим перепадом давле-
ния, превышающими те, которые обусловлены непосредственно
вязкостью жидкости и рассматривались ранее. Рост сопротивле-
Гл. 14. Йены большого круга 549
Рис. 14.16 Зависимость сопротивления прямой трубки с поперечным сечением
в форме эллипса, имеющим постоянный периметр, от площади ее поперечного
сечения (А) и от отношения малой главной оси к большой (6/а) (В). Безразмер-
ный параметр R — отношение сопротивления трубки с сечением в форме эл-
липса к сопротивлению той же трубки при а = о (те когда сечение круглое);
А/Ао — отношение площади поперечного сечения эллиптической трубки к пло-
щади ее поперечного сечения при а = b Таким образом, для круглой трубки
A!A^ = 1, b/а = 1, R = 1. На А сравниваются две кривые — для трубки с се-
чением в форме эллипса и для трубки с круглым сечением такой же площади
(периметры сечений, естественно, разные). С приближением площади поперечного
сечения к нулю сопротивление обеих трубок становится бесконечно большим, но
когда площадь А становится меньше примерно 0,5 До (b/а меньше, чем при-
мерно 0,25), сопротивление трубки с эллиптическим сечением превышает сопро-
тивление трубки с круглым сечением больше чем вдвое
ния, наблюдаемый в опытах и соответствующий участку II кривой
на рис. 14.15, Л, может быть частично обусловлен явлением от-
рыва.
Для течений с большими числами Рейнольдса, характерными
для большинства экспериментов (и соответствующими условиям
в крупных венах), полный теоретический анализ соотношения ме-
жду давлением и расходом в настоящее время провести не удает-
ся. Можно прибегнуть лишь к очень приближенному анализу,
предположив, что ниже области сужения поток «отделен» от сте-
нок (имеет вид струи), и, следовательно, восстановления давле-
ния не происходит (рис. 14.17). В этом случае давление в области
сужения равно давлению pz на выходе, в то время как площадь
поперечного сечения А2 здесь много меньше, чем площадь Л! в об-
ласти измерения входного давления. Поэтому средняя скорость
движения жидкости в месте сужения (0]А2) значительно больше,
чем в вышележащей части трубки (Q/А). Пренебрегая разностью
давлений между областью измерения входного давления и об-
ластью сужения, обусловленной непосредственно вязкостью жид-
кости, можно, воспользовавшись уравнением Бернулли (4.6), запи
550 Часть П. Механика кровообращения
сать соотношение
(14.8)
где р — плотность жидкости. Все величины в этом уравнении,
кроме площади А2 в области сужения, измеряются в опыте; зна-
чение Л2 можно оценить либо по фотографиям, аналогичным при-
веденным на рис. 14.15,5, либо по известному трансмуральному
давлению р2 — рк, использовав для этого кривую растяжимости
трубки (рис. 14.1 или 14.2). Тогда уравнение (14.8) можно про-
верить экспериментально. Такая проверка показала достаточно хо-
рошее согласие теории и опыта.
Рис. 14.17. Отрыв потока от стенок трубки за областью сужения при больших
числах Рейнольдса.
Объяснить возникновение колебаний значительно труднее. Для
описания одного из предполагаемых механизмов также исполь-
зуется уравнение Бернулли. Суть дела заключается в следующем.
Допустим, что на выходном конце трубки давление несколько
ниже, чем в камере, так что трубка начинает спадаться. Из-за
инерции жидкости в начале процесса сужения трубки расход не
уменьшается сразу, поэтому скорость в области сужения увеличи-
вается, и, согласно уравнению Бернулли, давление здесь падает.
Это снижение давления увеличивает разность давлений в камере
и в трубке, в результате чего спадение трубки ускоряется. Когда
площадь просвета трубки становится очень малой, сопротивление
движению оказывается очень высоким, что обусловлено как непо-
средственным влиянием вязких сил в области сужения, так и от-
рывом потока от стенок и возникновением турбулентности ниже
места сужения. В результате расход уменьшается, а давление не-
посредственно перед областью сужения повышается Это высокое
давление заставляет спавшуюся часть трубки снова открыться,
после чего давление на входе опять падает и весь цикл повто-
ряется,
Гл. 14. Вены большого круга 551
Такое объяснение механизма колебаний кажется весьма прав-
доподобным, но не может считаться исчерпывающим, поскольку
не учитывает влияния инерции, связанной с местными ускорением
и замедлением движения жидкости в каждом цикле. При наблю-
даемых частотах колебаний значение параметра а должно быть
достаточно большим (от 4 до 13), и инерцией в этом случае пре-
небрегать нельзя. Теории, учитывающей это, пока нет. Если бы
такую теорию удалось создать, она могла бы описывать и возник-
новение в трубке «ударных волн» (разд. 12.5). Вероятно, необ-
ходимо учитывать также диссипацию энергии, обусловленную вяз-
коупругими‘свойствами стенки трубки.
Физиологическое свидетельство: тоны Короткова. Ранее мы уже
говорили, что при отрицательном трансмуральном давлении вены
спадаются; в качестве примера упоминались вены на шее стоя-
щего человека. Однако не было проведено, вероятно, ни одного
эксперимента, в котором измеряли бы одновременно трансмураль-
ное давление, площадь поперечного сечения и расход крови в вене
в области ее спадения или вблизи этой области, равно как нет
никаких сведений о наблюдениях в венах самовозбуждающихся
колебаний. Тем не менее давно известно о возникновении высоко-
частотных колебаний в артериях большого круга — колебаний, ве-
роятно, подобных тем, которые наблюдаются в опытах на моде-
лях. Это тоны Короткова.
При измерении артериального давления с помощью надуваемой
манжетки давление в ней обычно сначала повышают до уровня,
заведомо превосходящего максимальное (систолическое) давле-
ние, так что артерия спадается и кровоток в ней прекращается.
Затем, поместив фонендоскоп над артерией дистальнее манжетки
и постепенно снижая давление, прослушивают возникающие в ар-
терии звуки. На определенном этапе появляются тоны Короткова,
совпадающие по времени с сокращениями сердца. Ангиографиче-
скими и ультразвуковыми методами показано, что появление то-
нов совпадает с возобновлением течения крови через спавшийся
участок артерии. По мере постепенного снижения давления в ман-
жетке длительность тонов Короткова во время каждого сокраще-
ния сердца увеличивается, потому что пережимаемый участок ар-
терии оказывается открытым в течение все большего промежутка
времени. Когда давление в манжетке приближается к диастоли-
ческому давлению в артерии, тоны становятся более глухими, а
затем и вовсе исчезают. Точно не установлено, что именно — при-
глушение или исчезновение тонов — соответствует диастолическому
давлению, но по международному соглашению принято считать,
что приглушение. Если амплитуды разных частотных составляю-
щих тонов изобразить так, как это сделано на рис. 14.18, то можно
увидеть, что до того, как тоны становятся более глухими, значи-
тельный вклад в них дают составляющие с частотой вплоть до
180 Гц. Но как только тоны приглушаются, амплитуда всех со-
Б52 Часть II. Механика кровообращения
Рис. 14.18. Зависимость амплитуды высокочастотных составляющих тонов Ко-
роткова от частоты Верхняя кривая—давление в манжетке, промежуточное
между систолическим и диастолическим; средняя кривая — тоны стали более
глухими, давление в манжетке приблизительно равно диастолическому давлению,
нижняя кривая — давление в манжетке ниже диастолического. [McCutcheon,
Rushmer (1967). Korotkoff sounds: an experimental critique Circulation Res., 20,
154. С разрешения American Heart Association Inc]
ставляющих с частотой от 60 до 180 Гц значительно уменьшается.
Вводя в артерии сжимаемой манжеткой конечности специаль-
ные вещества под давлением, равным артериальному, можно полу-
чить слепки артерий. По слепкам видно, что форма поперечного
сечения артерий в этом случае заметно отличается от круглой.
Как площадь, так и периметр сечения значительно уменьшены, и
это напоминает поведение сегментов вен при сжатии. На внут-
ренней поверхности сосудов обнаруживаются продольные Складки.
Это говорит о том, что артерии большого круга, так же как и
вены, спадаются, когда трансмуральное давление близко к нулю
или отрицательно. Поэтому представляется весьма правдоподоб-
ным, что тоны Короткова есть следствие рассмотренных выше са-
мовозбуждающихся колебаний ’).
*) Спонтанные колебания в способной к спадению трубке, о которых шла речь
выше и которые упоминаются здесь, представляют собой следствие неустойчиво-
сти в механической системе «поток — стенка» Одиако возникновение тонов Ко-
роткова имеет, по-видимому, принципиально иной нелинейный механизм: перио-
дическое прохождение волн раскрытия сосуда с крутым фронтом и высокочастот-
ными колебаниями за ним (см Григорян С С, Саакян Ю 3, Цатурян А. К.
О механизме генерации звуков Короткова. — ДАН СССР, 1980, 251, № 3, 570—
574). — Прим. ред.
Гл. 14. Вены большого круга 553
14.5. Механика венозного русла
От изложения свойств вен мы перейдем теперь к рассмотрению
целого венозного русла отдельных органов н влияния на течение
крови внешних воздействий, таких, как сокращение скелетных
мышц и дыхание. Поскольку трансмуральное давление в веноз-
ных сосудах низкое и растяжимость их при таком давлении ве-
лика, объем венозной системы очень чувствителен к воздействиям,
изменяющим трансмуральное давление. Кроме того, соответствую-
щие им изменения кровотока трудно предсказать из-за нелиней-
ности механических свойств вен. Поэтому мы можем только каче-
ственно охарактеризовать поведение системы и указать, насколько
наши выводы согласуются с физиологическими наблюдениями.
Поднятие руки над уровнем сердца. Когда венозное русло
(например, руки) расположено приблизительно на уровне сердца,
среднее давление в русле повсюду составляет около 0,5-103 Н-м-2,
и, вероятно, ни один сосуд не находится в спавшемся состоянии.
При поднятии руки трансмуральное давление уменьшается и вены
спадаются, но чтобы это произошло, заключенный в них объем
крови должен уменьшаться. [Уменьшение объема обусловлено как
резким кратковременным отливом крови из венозного русла руки,
так и небольшим преходящим снижением притока крови. Послед-
нее связано с тем, что при спадении вен площадь их поперечного
сечения уменьшается, а следовательно, увеличивается импеданс
венозной системы (об импедансе см. разд. 12.4 и 12.10). Более
существенным является, вероятно, отлив крови, так как импеданс
крупных вен и сердца остается значительно меньшим, чем импе-
данс системы микрососудов.] Поскольку входной импеданс арте-
риальной системы руки, определяющийся вязкоупругими свой-
ствами артерий и сопротивлением системы микрососудов, при под-
нятии руки фактически не меняется, в сущности неизменным ос-
тается и средний расход в артериях. Поэтому расход в венозном
русле также остается примерно постоянным. В результате как
средняя скорость движения крови, так и скорость сдвига в венах
увеличиваются. Когда кровь отливает из спадающихся вен руки,
скорость движения крови увеличивается больше всего в участках
венозного русла, расположенных ближе к сердцу. Вследствие
этого скорость сдвига в венах, расположенных ближе к средцу,
на короткое время возрастает сильнее, чем в более удаленных
от него венах.
Сокращение скелетных мышц. Можно ожидать, что при умерен-
ном непрерывно поддерживаемом сокращении скелетных мышц
конечности, находящейся на уровне сердца или ниже, трансму-
ральное давление в венах этих мышц, как и при поднятии конеч-
ности, снизится и на состояние венозного русла мышц такое сокра-
щение будет оказывать аналогичное влияние. Однако мощное род
554 Часть II. Механика кровообращения
держивающееся сокращение скелетных мышц конечности спо-
собно привести к прекращению кровоток^ в этих мышцах, ве-
роятно, из-за сдавливания и полного пережатия сначала вен, а за-
тем, возможно, и артерий.
Перемежающиеся сокращения скелетных мышц конечности
оказывают более сложное воздействие на движение крови по их
венозному руслу, чем умеренное непрерывное сокращение. Эффект
зависит от ряда факторов, в том числе от силы и частоты сокра-
щений, объема крови в соответствующем венозном русле, наличия
в венах клапанов и от времени, необходимого для заполнения
этого русла после его опустошения. Последствия таких сокраще-
ний можно наглядно продемонстрировать на венах ноги здоро-
вого человека, измеряя давление в них (по отношению к атмо-
сферному; в этом случае измеряемое давление приблизительно
равно трансмуральному) в состоянии покоя и при произвольных
повторяющихся сокращениях мышц голени.
Когда испытуемый лежит горизонтально, среднее давление
в вене на уровне лодыжки равно примерно 0,7-Ю3 Н-м-2
(5 мм рт. ст.). Если после этого перевести испытуемого в верти-
кальное положение, но так, чтобы он при этом не совершал актив-
ных движений, давление постепенно повысится до нового постоян-
ного значения — около 11,9-103 Н-м-2 (85 мм рт. ст.). Повышение
давления происходит постепенно из-за того, что венозные клапаны
предотвращают обратное движение крови, и наполнение веноз-
ного русла осуществляется только за счет притока крови из си-
стемы микрососудов.
Если теперь испытуемый начинает повторно напрягать мышцы
ног, как при ходьбе (рис. 14.19), то давление в вене во время пер-
вого «шага» повышается, а затем, при расслаблении мышц, па-
дает. С каждым последующим шагом давление продолжает па-
дать и в конечном счете достигает значения в пределах (2—4) • 103
Н-м-2. Когда сокращения мышц прекращаются, давление в вене
постепенно повышается до исходного. Измерение полного объема
конечности в этих условиях показывает, что повторяющиеся со-
кращения мышц приводят к его уменьшению вследствие снижения
объема крови в венах. Следовательно, если интервал между со-
кращениями значительно превышает время, необходимое для но-
вого наполнения венозного русла, — это время меняется прибли-
зительно от 50 с для покоящейся конечности до 5 с для интен-
сивно работающей, — то такие сокращения не могут устойчиво по-
низить местное давление в венах (или объем содержащейся в них
крови).
Сокращения, совершающиеся настолько часто, что за время
между ними венозное русло не успевает заполняться до исходного
уровня, оказывают на условия движения крови в соответствующих
венах влияние иного рода. Поскольку скорость притока крови в
сосудистое русло определяется разностью давлений в артериальч
Гл 14. Вены большого круга 555
Рис 14 19 Максимальные и минимальные значения давления в вене на уровне
лодыжки у испытуемого, находящегося в вертикальном положении Сначала
испытуемый неподвижен, затем шагает по тредбану, а в конце опыта спокойно
стоит. I — покой, // — первый шаг; III— второй шаг; /V —третий шаг, V — пя-
тый шаг, VI — седьмой шаг, VII — остановка тредбана, VIII — последний шаг.
[Pollack, Wood (1949) Venous pressure in the saphenous vein at the ancle in
man during exercise and changes in posture. J. appl. Physiol, 1, 656]
ной и венозной системах и величиной периферического сопро-
тивления, снижение среднего по времени венозйого давления при-
ведет к увеличению притока крови, а также ее оттока из рассмат-
риваемого сосудистого русла через вены к сердцу, причем крово-
снабжение возрастет пропорционально увеличению разности дав-
лений. Однако в обычных условиях венозное давление настолько
ниже артериального, что существенного увеличения притока крови
при достаточно частых, но слабых сокращениях не происходит,
Следует иметь в виду, что, хотя избыточное давление в вене ло-
дыжки у неподвижно стоящего человека составляет 12-Ю3 Н-м~2,
избыточное давление в расположенной рядом артерии равно при-
мерно 24-103 Н-м-2, так что разность давлений в артериях и в ве-
нах все еще составляет примерно 12-103 Н-м~2. Частые слабые
сокращения приводят в действительности к колебаниям кровотока
в венах и, возможно, к изменению распределения кровотока, так
как одни вены могут оказываться сжатыми, а другие — растяну-
тыми. При определенных частотах сокращений отток крови из пе-
режимаемых вен по направлению к сердцу будет происходить
только во время сокращения мышц, потому что в период их рас-
слабления вены вновь наполняются. Но, поскольку средняя по вре-
мени величина расхода крови через сосудистое русло конечности
изменяется незначительно, а средний по времени объем крови в
венах уменьшается, средняя скорость движения крови в венах
556 Часть II Механика кровообращения
должна возрасти, а среднее время ее пребывания в них (отноше-
ние объема венозного русла к расходу) — уменьшиться
Отсюда можно сделать несколько важных выводов У людей,
стоящих длительное время неподвижно, гидростатическое давле-
ние в венах нижних конечностей несомненно выше и эти сосуды
растянуты сильнее, чем у тех, кто попеременно напрягает мышцы
голени, как при ходьбе. При неполноценности венозных клапанов
сокращения мышц голени не приводят к столь эффективному сни-
жению трансмурального давления в венах и уменьшению их диа-
метра, которое присуще здоровым людям. Обычным последствием
неполноценности венозных клапанов является образование в венах
тромбов. Предполагают, что длительное растяжение вен способ-
ствует патологическому (варикозному) их расширению.
Ранее (гл. 10) уже упоминалось, что тромбы в венах голени
образуются преимущественно у больных, прикованных к постели.
Никаких объяснений этому факту нет, хотя известно, что тромбы
образуются реже, если больные производят упражнения — сокра-
щения мышц голени, или если голени подвергать ритмическому
внешнему сжатию (без изменения среднего по времени крово-
тока) *). Это наводит на мысль, что образование тромба связано
с ослаблением пульсаций венозного кровотока, с уменьшением
средних по времени значений скорости крови в венах и скорости
сдвига и с увеличением времени пребывания крови в венах го-
лени. Однако никакой причинной связи между этими механиче-
скими факторами и образованием тромбов не выявлено* 2).
Важно иметь в виду, что повторяющиеся сокращения мышц го-
лени оказывают на давление крови в венах и на кровоток в них
качественно одинаковое влияние независимо от того, функциони-
руют ли венозные клапаны нормально, имеет ли место их непол-
ноценность или они вообще отсутствуют. Чтобы показать это, рас-
смотрим заполненную жидкостью горизонтальную растяжимую
трубку, центральная часть которой подвергается быстрому сжа-
тию. Как мгновенный, так и средний по времени оттоки жидкости
из обоих концов трубки будут одинаковы лишь в том случае, если
на обоих концах одинаков импеданс. Поэтому, если с одной сто-
роны трубки имеется полноценный или частично полноценный кла-
пан, а с другой — растяжимая камера (такая, как правое пред-
сердие), то ббльшая часть жидкости будет направляться к камере,
*) Измерения показали, что повторяющееся один раз в минуту сжатие тка-
ней конечности в течение 5 с при скорости нарастания внешнего давления
IO3 Н м2 с-1 не оказывает влияния на средний по времени кровоток, но заметно'
снижает число случаев образования тромбов в глубоких венах
2) Заметим, что данные, полученные в условиях модельных экспериментов
с движущейся кровью, показывают, что почти каждая из составляющих процес-
са образования тромба (агрегация тромбоцитов, высвобождение из них АДФ
и т. д —см разд. 10 4, 10 5) сложным образом зависит от характеристик тече-
ния, в частности от уровня скоростей сдвига н наличия отрывных зон с замкну-
тыми траекториями. — Прим. ред.
Гл. 14. Вены большого круга 557
импеданс которой ниже. В случае вен результат будет таким же,
даже если клапаны совершенно неполноценны или отсутствуют,
потому что импеданс приводящих сосудов намного больше импе-
данса расположенных ближе к сердцу вен и правого предсердия.
До сих пор мы имели дело лишь с небольшими участками ве-
нозного русла — анализ их поведения без учета остальной части
сосудистого русла был оправдан, так как происходящие в этих
участках изменения не оказывают заметного влияния на состояние
системы кровообращения в целом Но если такие изменения затра-
гивают значительную часть венозной системы, то они могут сущест-
венно изменить условия возвращения крови по венам в сердце, а
это, как было показано в гл 11, может повлиять на производитель-
ность сердца В свою очередь его производительность влияет на кро-
воток в самом венозном русле. К явлениям такого рода приводят
изменения давления в легких — различные дыхательные пробы.
Дыхательные пробы. Достоверных данных о влиянии дыхатель-
ных проб на венозную систему очень мало, а мнения разных ис-
следователей по этому вопросу несколько расходятся Поэтому мы
остановимся на нем лишь вкратце и почти целиком ограничимся
результатами измерений, проводившихся на полой вене Сле-
дует помнить, что дыхательные пробы могут сильно влиять также
на условия течения крови в артериях большого круга; это влияние
опосредуется как через изменение количества крови, возвращаю-
щейся по венам в сердце, так и через воздействие на механические
процессы в артериях, находящихся в грудной полости Рефлектор-
ным путем дыхательные пробы могут изменять также частоту
сердцебиений и состояние периферического сосудистого русла.
Влияние дыхания на вены и на приток по ним крови в сердце
привлекает внимание исследователей уже больше двухсот лет.
Выдающийся швейцарский анатом и физиолог Альбрехт фон Хал-
лер (1703—1788) полагал, что расширение грудной клетки во вре-
мя вдоха и связанное с этим снижение давления в грудной поло-
сти присасывают кровь из вен вне этой полости в сосуды, находя-
щиеся внутри нее, и в сердце Выдох, по его представлениям,
оказывает противоположное действие повышение давления в груд-
ной полости приводит к уменьшению объема находящихся здесь
вен и к увеличению давления в них, так что скорость поступле-
ния к ним крови из вен, находящихся вне грудной полости (их со-
стояние при дыхании не меняется), уменьшается Именно это и
происходит при спокойном дыхании. Однако другие дыхательные
движения могут приводить к более сложным последствиям.
События, сопровождающие глубокий задержанный вдох, совер-
шаемый при закрытой голосовой щели (проба Мюллера), отобра-
жены на рис 14.20. Альвеолярное и внутриплевральное давления
уменьшаются, а следовательно, уменьшается и давление в правом
предсердии и в грудном отделе полой вены. Однако в брюшной
полости давление резко повышается из-за сокращения брюшных
558 Часть II. Механика кровообращения
ЭКГ
Рис. 14 20. Изменение давления в правом предсердии, давления в нижней полой
вене н скорости крови в ней во время пробы Мюллера Верхняя запись — ЭКГ.
Датчики для измерения давления и скорости крови в нижней полой вене распо-
ложены ниже диафрагмы, т е. в брюшном отделе полой вены. Вертикальная
прямая нанесена так, чтобы показать, что во время пробы максимум волны
кровотока совпадает по времени с максимумом волны давления в предсердии.
[Wexler, Bergel, Gabe, Makin, Mills (1969) Velocity of blood flow in normal
human venae cavae Circulation Res., 23, 356. С разрешения American Heart
Association Inc ]
мышц и опускания диафрагмы, и это повышение передается в
брюшной отдел нижней полой вены. Поэтому разность давлений
в брюшном и грудном отделах полой вены значительно увеличи-
вается и движение крови в ее брюшном отделе резко ускоряется.
Но такое повышение скорости крови оказывается лишь кратковре-
менным: позднее она остается только слегка увеличенной. Это обу-
словлено тем, что давление в брюшной полости превышает давле-
ние в ближайшей к сердцу части брюшного отдела полой вены
(т. е. в той ее части, которая расположена тотчас ниже диаф-
рагмы), и она в этой области спадается. Поэтому включаются в
работу механизмы, описанные на стр. 543—551 (хотя самовоз-
буждающихся колебаний не возникает), и кровоток в брюшном от-
деле полой вены оказывается независящим от давления в правом
предсердии, пока давление в нем остается ниже давления в брюш-
ной полости. Запись скорости крови в брюшном отделе полой вены,
приведенная на рис. 14.20, показывает, что фаза колебаний этой
Гл. 14. Вены большого круга 559
скорости, обусловленных сокращениями сердца, отличается от
обычной: максимум скорости крови (направленной к сердцу) до-
стигается во время систолы предсердий, а не желудочков. Этот
факт еще раз подчеркивает, что давление в правом предсердии не
оказывает влияния на кровоток в брюшном отделе полой вены.
Происхождение колебаний скорости крови в данном случае не-
ясно. На более поздних стадиях пробы давление в предсердии по-
вышается, а в брюшном отделе полой вены падает; скорость крови
увеличивается, и в конечном счете восстанавливаются ее нормаль-
ные пульсовые колебания. Предположительно это объясняется тем,
что испытуемый не может больше поддерживать высокое давле-
ние в брюшной полости или низкое давление в' грудной полости,
так что вена оказывается уже не спавшейся.
Вообще об изменениях давления в брюшной полости при ды-
хании известно мало. Предполагают, что в условиях спокойного
дыхания перед опусканием диафрагмы при вдохе или одновре-
менно с ним мышцы брюшной стенки расслабляются, так что дав-
ление в брюшной полости при вдохе остается постоянным или не-
сколько снижается. В условиях глубокого дыхания мышцы брюш-
ной стенки, по-виднмому, во время вдоха частично сохраняют ак-
тивное напряжение и опускание диафрагмы приводит, как и в
случае пробы Мюллера, к повышению давления в брюшной по-
лости. В результате скорость движения крови в венах, располо-
женных дальше от сердца, чем вены брюшной полости (например,
в бедренных венах), уменьшается, а кровь, находящаяся в венах
брюшной полости, выталкивается в вены грудной полости1).
Проба Вальсальвы, во время которой испытуемый совершает
при закрытой голосовой щели усиленный выдох и таким образом
повышает давление как в брюшной полости, так и в грудной по-
лости, также оказывает влияние на механику движения крови в ве-
нах. В этом случае в тех участках крупных вен, где они входят в
грудную и в брюшную полости, кровоток падает до нуля. Он не
восстанавливается до тех пор, пока приток крови из системы
микрососудов в эти вены не повысит давление в них до уровня,
превышающего давление в венах, которые находятся в грудной и
в брюшной полостях. Таким образом, проба Вальсальвы приво-
дит к значительному уменьшению притока крови по венам к пра-
’) При некоторых заболеваниях, например при фиброзном утолщении пери-
карда (констриктивном перикардите), или когда в полости перикарда скапли-
вается избыточная жидкость, сердце не может растягиваться свободно. Когда
больной делает глубокий вдох, центральное венозное давление (т. е. давление в
правом предсердии) повышается и вены на шее становятся растянутыми. В нор-
мальных условиях повышение давления в правом предсердии приводит к увели-
чению производительности правого желудочка, что может компенсировать уве-
личение притока крови в полость грудной клетки Но у названных больных такая
компенсация иногда бывает невозможна. В медицине это явление известно под
названием «симптом Куссмауля?»,
860 Часть II. Механика кровообращения
вому предсердию, а следовательно, и к уменьшению количества
крови, перекачиваемого правыми камерами сердца. Это, в свою
очередь, приводит к уменьшению производительности левых камер
сердца. Но до нуля производительность левого желудочка не па-
дает, так как при этой пробе условия движения крови по сосу-
дам, находящимся в грудной и в брюшной полостях, не изменяют-
ся. Кровоснабжение мозга обычно поддерживается на уровне, до-
статочном для сохранения у испытуемого сознания. Это обеспечи-
вается за счет рефлекторного сужения периферических сосудов,
хотя давление в артериях большого круга все-таки падает.
Глава 15
МАЛЫЙ КРУГ КРОВООБРАЩЕНИЯ
В малом круге кровообращения вся кровь, поступающая из
правого желудочка, доставляется по легочным артериям в капил-
ляры альвеол, а затем по легочным венам возвращается в левое
предсердие. Легкие имеют еще одно сосудистое русло, значительно
меньшее, — бронхиальное. Оно берет начало из грудной аорты и
обеспечивает легкие артериальной кровью из большого круга.
Кровь, оттекающая от легких по этому руслу, поступает в веноз-
ную систему большого круга. Между микрососудами бронхиаль-
ного сосудистого русла и малого круга кровообращения сущест-
вуют анастомозы.
Между малым кругом кровообращения и большим имеется ряд
существенных различий. Прежде всего малый круг — это система
низкого давления и низкого сопротивления движению крови. Сред-
нее по времени избыточное давление в легочных артериях состав-
ляет всего около 2-103 Н>м~2 (15 мм рт. ст., или 20 см вод. ст.),
т. е. приблизительно одну шестую от величины избыточного давле-
ния в артериях большого круга. Тем не менее расход крови в ма-
лом круге такой же, как и в большом. Другое отличие состоит в
том, что стенка легочных артерий значительно тоньше стенки арте-
рий большого круга. Кроме того, сосудистое русло легких, по-ви-
димому, не имеет регионарной специализации. Наконец, при обыч-
ных условиях регуляция сосудов малого круга вазомоторной си-
стемой, как полагают, играет незначительную роль: в отличие от
артерий и вен большого круга существенных активных изменений
просвета этих сосудов не происходит.
Основная функция легких — осуществление газообмена, т. е.
обмена кислорода и двуокиси углерода, между кровью и воздухом.
Однако через мембрану альвеолярных капилляров диффундирует
любой газ, если различны его парциальные давления в крови ка-
пилляров малого круга и в альвеолах. К важным в практическом
отношении газам относятся газообразные средства для наркоза,
например закись азота. Определение скорости поглощения закиси
азота из вдыхаемой смеси газов ныне используют для измерения
мгновенного расхода крови в капиллярах малого круга (разд. 15.4
и рис. 15.17). Поглощение окиси углерода в индикаторных дозах
применяют для определения диффузионной способности легких ц
Объема крови в капиллярах малого круга (разд. 15.2),
562 Часть II. Механика кровообращения
Помимо своей основной функции, сосуды малого круга благо-
даря их податливости участвуют в некоторых механических про-
цессах. Мы видели, что'при натуживании, кашле и других состоя-
ниях, сопровождающихся повышением давления в грудной и в
брюшной полостях, приток крови к правому предсердию по венам
большого круга, находящимся вне этих полостей, на короткое вре-
мя прерывается (разд. 14.5). В такие моменты приток крови к ле-
вому предсердию, а следовательно, и в артерии большого круга
обеспечивается за счет крови, оттекающей из сосудов малого кру-
га. Поскольку приток крови в малый круг в это время снижен,
объем содержащейся в его сосудах крови уменьшается. Кроме
того, сосуды малого круга обеспечивают затухание колебаний дав-
ления и кровотока, создаваемых сокращением правого желудочка
и левого предсердия; эти противоположные по фазам и направ-
лению распространения колебания могли бы препятствовать дви-
жению крови в сосудах малого круга (разд. 15.3 и рис. 15.14).
Можно указать здесь еще две важные функции малого круга
кровообращения. Во-первых, его сосуды действуют как фильтр для
удаления из циркулирующей крови частиц, появляющихся в ре-
зультате тех или иных патологических процессов. Так, эмбол —
оторвавшийся кусочек тромба, образовавшегося в венах большого
круга или в правых камерах сердца, попадая в сосуды малого кру-
га, застревает в них. Такое явление не относится к категории же-
лательных, однако, если эмбол достаточно мал, никаких суще-
ственных нарушений при этом не происходит и он впоследствии
растворяется под действием фибринолитического механизма
(разд. 10.5). События развиваются совершенно по-иному, если
такой эмбол попадает в сосудистое русло столь небольшого, но в
высшей степени специализированного органа, как головной мозг.
Другая очень важная роль малого круга связана с его способ-
ностью влиять на химический состав крови: при прохождении кро-
ви через легкие определенные вещества удаляются из нее или до-
бавляются к ней. Достаточно, например, однократного прохожде-
ния крови через сосуды малого круга, чтобы значительная часть
содержащегося в ней сосудосуживающего вещества ангиотензин I
превратилась в его активную форму—ангиотензин II, сосудорас-
ширяющее вещество брадикинин почти полностью инактивиро-
валось, а серотонин (5-окситриптамин, 5-ОТ) — один из ами-
нов, играющий важную роль в функции тромбоцитов и дея-
тельности центральной нервной системы, — был удален из
крови.
Как мы покажем, наибольшее влияние на протекающие в ма-
лом круге механические процессы оказывают механические свой-
ства легких и степень наполнения их воздухом. Именно поэтому мы
посвящаем кровообращению в малом круге целую главу, в кото-
рой значительное внимание уделено влиянию на него механики
легких,
Гл. 15. Малый круг кровообращения 56
15.1. Анатомическая организация
Малый круг кровообращения. Артериальная система малой
круга начинается от легочного клапана (на выходе из правой
желудочка сердца). За створками этого клапана легочный ствол
как и аорта, образует расширения — синусы. У человека длина ле
точного ствола 5—6 см, а диаметр 2,5—3,0 см. Как показано н<
рис. 15.1, легочный ствол, отходя от сердца, направляется ввер:
и назад, располагаясь под дугой аорты; здесь он разделяется и;
Безымянная артерия
Правая безымянная вена
Дуга аорты -----------
Правая легочная артерия
Верхняя полая вена
Нижняя полая вена
Правые легочные вены
Правое предсердие-
Трехстворчатый клапан
Митральный клапан
Аортальный клапан
Левый желудочек
Легочный клапан
Межжелудочковая перегородка
Правый желудочек
Диафрегма
Брюшная аорта
Левая общая сонная артерия
Левая подключичная артерия
Левая безымянная вена
Левая легочная артерия
Левые легочные вены
Легочной ствол
Рис. 15.1. Схема сердца и больших сосудов человека (вид спереди). Стрелки
показывают направление движения крови. [Pauchet, Dupret (1937). Pocket atlas
of anatomy (3-rd edn), Oxford University Press.] Более простая схема движения
крови через сердце показана на рис. 111.
правую и левую легочные артерии. [У собаки легочный ствол ко-
роче (3—4 см) и уже (1,4—1,7 см в диаметре).] Легочные арте-
рии направляются соответственно к правому и левому легкому, но,
прежде чем войти в ткань органа, сосуды делятся на несколько
ветвей, которые снабжают кровью отдельные доли легких. В об-
ласти входа в легкие — у их ворот, а также дальше по ходу вет-
вей артерии окутаны оболочками из соединительной ткани; они
представляют собой продолжение плевры — соединительноткан-
ной оболочки, охватывающей легкие и окружающей бронхи. Ле-
гочные артерии и бронхи прободают эту оболочку лишь при входе
в собственно ткань легких, уже на уровне терминального возду-
хоносного пространства.
На рис. 15.2 показан изготовленный из отвердевающего поли-
мера слепок артериального дерева малого круга человека. Со-
суды диаметром меньше 0,08 см удалены, чтобы показать более
крупные артерии. Характер ветвления такой же, как для артерий
и вен большого круга, в том смысле, что делители потока отно-
сительно острые и ветвление в разных областях происходит по-
В64 Часть II. Механика кровообращения
Таблица 15.1
Число и размеры сосудов артериальной системы
малого круга кровообращения человека
Порядок Число ветвей Диаметр, см Длине, см Ud Общее число конечных ветвей
17 1,00 3,000 9,05 3,00 3,00 • 10®
16 3,00 1,483 3,20 2,16 1,00 • 10®
15 8,00 0,806 1,09 1,35 3,02 • Ю7
14 2,00 • 10 0,582 2,07 3,56 1,38 • 107
13 6,60 • 10 0,365 1,79 4,90 3,98 • 10®
12 2,03- 102 0,209 1,05 5,02 1,16-10®
11 6,75 • 102 0,133 0,66 4,96 3,47- 10®
10 2,29 • 103 0,085 0,47 5,54 8,92 • 104
9 5,86 103 0,053 0,32 6,05 4,81 • 104
8 1,76 • 104 0,035 0,21 6,00 1,60-104
7 5,26 • 104 0,022 0,14 6,36 5,36 103
6 1,57 • 106 0,014 0,09 6,43 1,79 • Ю3
5 4,71 • 105 0,009 0,07 7,78 5,98 -102
4 1,41 • 10е 0,005 0,04 8,00 2,00 -102
3 4,23 • 10е 0,003 0,03 10,00 6,66 • Ю
2 1,27 -107 0,002 0,02 10,00 2,37 • 10
1 3,00 • 10® 0,001 0,01 10,00 1,00
) Singhal et at, (1973), Circulation Res , 33 190-197.
разному. Например, легочный ствол делится на две примерно оди-
наковые легочные артерии (они видны в центре слепка), тогда как
большая часть остальных ветвлений осуществляется за счет от-
хождения небольших артерий от более крупных. Ветвление такого
характера можно видеть и на рис. 15.3, где показана перифери-
ческая часть слепка; диаметр центрального сосуда этой части
слепка около 0,07 см.
Описать столь разветвленную систему — задача не простая.
Можно расположить ветви по поколениям (по порядку их образо-
вания), начиная счет от центрального сосуда; другой способ — клас-
сифицировать их по порядкам, начиная от самых периферических
сосудов. Для сосудов малого круга характерна значительная асим-
метричность ветвлений, и близкие по диаметру сосуды могут по-
являться в разных поколениях. Поэтому первый способ не позво-
ляет получить правильного представления. Второй способ (он под-
робно излагается в работе, указанной в сноске к табл. 15.1), хотя
он и более сложен, дает по крайней мере возможность отнести
сосуды близкого диаметра к одному порядку. Использование этого
метода для классификации данных, полученных при обмерах слеп-
ков, подобных показанному на рис. 15.2 и 15 3, позволило рассчи-
тать некоторые средние величины, характеризующие артериальное
дерево малого круга человека (табл. 15.1 и 15.2). Как следует цз
табл. 15.1, для более крупных артерий отношение длины к диа-
Гл. 15. Малый круг кровообращения 565
Рис 15 2 Артериальная система малого круга кровообращения Чтобы выявить
общий характер ветвления артерий, ветви диаметром меньше 0,08 см удалены.
[Cumming, Henderson, Horsfield, Singhal (1969). The functional morphology of
the pulmonary circulation. In: The pulmonary circulation and interstitial space (ed,
Fishman, Hecht), University of Chicago Press.]
метру составляет от 1,35 до 3,0, а для более мелких оно оказы-
вается больше и достигает 6—10. Полная длина среднего пути от
начала легочного ствола до капилляров составляет примерно 20 см.
Полная площадь поперечного сечения артериального русла, как
видно из табл. 15.2, сначала несколько уменьшается, так что сред-
няя скорость движения крови в первых двух ветвях, следующих
за легочным стволом, должна увеличиться, но после этого она сни-
жается. Среднее число Рейнольдса на протяжении всего артери-
ального дерева должно уменьшаться, потому что уменьшается
произведение скорости на диаметр. Отношение площадей (отно-
шение суммарной площади поперечного сечения ветвей к площади
поперечного сечения приносящего сосуда) при разделении легоч-
ного ствола на правую и левую легочные артерии как у чело-
века, так и у собаки равно примерно 0,8.
Характер ветвления прекапиллярных сосудов малого круга от-
личается от рассмотренного выше: от одного сосуда диаметром
15—25 мкм отходит ряд капилляров, которые располагаются в пе-
регородках альвеол и на их поверхности. По-видимому, один пре-
ббб Часть II. Механика кровообращения
Рис. 15.3. Небольшая ветвь артериального дерева малого круга кровообращения,
на примере которой виден характер ветвлений. [Cumming, Henderson, Horsfietd,
Singhal (1969). The functional morphology of the pulmonary circulation. In: The
pulmonary circulation and interstitial space (ed. Fishman, Hecht), University of
Chicago Press.]
капиллярный сосуд снабжает капиллярами несколько соседних
альвеол; как осуществляется такое снабжение — последовательно
или параллельно, — неизвестно.
Принято считать, что капилляры малого круга, как и капил-
ляры большого круга, представляют собой цилиндрические труб-
ки. Однако измерения показали, что длина их сравнима с диамет-
ром, а в некоторых случаях даже меньше диаметра. Кроме того,
оказалось, что они обычно уплощенные, а не круглые. Поэтому
правильнее рассматривать находящуюся в альвеолярной перего-
родке кровь как слой жидкости, движущейся между почти парал-
лельными альвеолярными мембранами, которые удерживаются на
определенном расстоянии друг от друга достаточно часто располо-
женными распорками из соединительной ткани (рис. 15.4). В лег-
ких кошки, строение которых подробно изучено, такие распорки
обычно занимают около 10% всей площади канала в межальвео-
лярной перегородке, внутри которого движется слой крови.
Стенка артерий малого круга намного тоньше стенки близких
по диаметру артерий большого круга. Отношение толщины стенки
к диаметру (h/d) для легочного ствола собаки равно примерно
Гл. 15. Малый круг кровообращения 567
Величины, ) рассчитанные по данным таблицы 15.1 Таблица 15.2
Порядок Площадь поперечного сечения, см2 Общая площадь поперечного сечения, см2 Общий объем, см3 Кумулятив- ный объем, см3 Скорость см-с-1
17 7,07 7,07 63,97 63,97 11,3
16 1,73 5,18 16,58 80,55 15,4
15 5,10- 10“’ 4,08 4,45 85,00 15,8
14 2,66 • 10“ 5,32 11,01 96,01 13,8
13 1,05 • 10~* 6,91 12,36 108,37 10,2
12 3,43 10“* 6,96 7,31 115,68 9,0
11 1,39 • 10“* 9,38 6,19 121,87 6,7
10 5,67 • 10, 12,99 6,09 127,96 4,2
9 2,17- 10“" 12,69 4,01 131,97 5,9
8 9,68 • 10~* 16,99 3,57 135,54 4,5
7 3,94 • 10“] 20,71 2,86 138,40 3,6
6 1,50 10“* 23,54 2,14 140,54 3,2
5 5,81 • 10““ 27,38 1,78 142,32 2,7
4 2,29- 10. 32,31 1,42 143,74 2,3
3 9,08 10. 38,37 1,Н 144,85 2,0
2 3,46 • 10“° 43,85 0,88 145,73 1,8
1 1,33 • 10 398,20 5,18 150,91 0,2
*) Значения площади поперечного сечения даны для одной ветвн. Общую площадь
поперечного сечения сосудов каждого порядка ветвления определяли как суммарную пло-
щадь всех ветвей данного порядка. Общий объем — суммарный объем всех ветвей каждого
порядка. Кумулятивный объем—сумма общих объемов от высшего порядка к низшему.
Скорость крови определяли как среднюю прн расходе в малом круге 80 мл-с“1. [Singhal et
al. (1973). Circulation Res., 33, 190-1971.
0,01, что сравнимо co значениями для вен, но не артерий боль-
шого круга: для аорты, например, оно составляет примерно 0,07
(табл. I). Гистологически также выявляются примечательные раз-
личия. Артерии малого круга, начиная от легочного ствола и вплоть
до сосудов диаметром 0,1 см, относят к артериям эластического
типа, потому что их средняя оболочка состоит в основном из эла-
стической ткани с небольшим количеством гладких мышц и кол-
лагена. На этом уровне происходит постепенный переход к арте-
риям мышечного типа (10—11-й порядки ветвления, табл. 15.1), и
сосуды диаметром 0,1—0,01 см относят к сосудам мышечного типа,
поскольку их средняя оболочка построена преимущественно из
мышц. В большом круге такой переход происходит уже в крупных
артериях. Например, коронарные и бедренные артерии, имеющие
диаметр несколько миллиметров и отходящие соответственно от
аорты и от общих подвздошных артерий, относятся к артериям
мышечного типа.
Еще одной характерной особенностью сосудистого русла ма-
лого круга является то, что в нем нет сосудов, аналогичных мел-
ким артериям мышечного типа большого круга и его артериолам.
Например, толщина стенки артерии малого круга диаметром
568 Часть П. Механика кровообращения
Рис. 15.4. Трехмерная схема строения межальвеолярной перегородки в легких
собаки, показывающая заключенный в этой перегородке канал, который состоит
из сосудистого пространства (капиллярного ложа) и межкапиллярных «распо-
рок». Эпителиальный слой альвеол откинут назад, чтобы показать соединительно-
тканный матрикс перегородки. Коллагеновые волокна, окружающие стенку ка-
пилляров, сходятся иа распорке. [Rosenqust et al. Microvascular Research, 1973,
5, 199—212.]
60 мкм составляет всего лишь 8 мкм (Л/d = 0,13), и она состоит
только из тонкой средней оболочки. Для соответствующих по диа-
метру артерий большого круга отношение Л/d равно 0,25—0,4, так
как средняя оболочка их толстая. Более того, в стенках «арте-
риол» малого круга, диаметр которых 30 мкм, гладкомышечные
клетки не выявляются, тогда как в средней оболочке артериол
большого круга они несомненно имеются. Трудно представить себе,
чтобы артериолы малого круга, в такой мере лишенные мышечных
клеток, могли бы интенсивно суживаться. Вазомоторные реакции,
которые выявляются в артериальной системе малого круга, осу-
ществляются, вероятно, в основном сосудами диаметром 0,01—
0,1 см, обладающими хорошо развитой средней оболочкой.
Тканевой барьер, отделяющий в альвеолах кровь от воздуха,
чрезвычайно тонок: обычно толщина его составляет около 0,2 мкм
(рис. 15.5). Он состоит из клеток, которые выстилают альвеолы
(альвеолярный эпителий) и капилляры (сосудистый эндотелий);
эти клетки отделены друг от друга узким межклеточным простран-
ством. Вблизи ядер клеток толщина тканевого барьера может до-
стигать 10 мкм. Эндотелий сосудов малого круга представляет
собой продолжение эндотелиальной выстилки всей остальной сер-
Гл. 15. Малый круг кровообращения 569
Рис. 15.5. Электронная микрофотография альвеолярио-капнллярного барьера в
области, где он очень тонок. Слева находится эритроцит. Стрелками отмечены
пять клеточных мембран: эритроцита (/), эндотелия капилляра (2) и эпителия
альвеол (3). Между эндотелиальными н эпителиальными клетками имеется узкое
межтканевое пространство (4). Видны также пиноцитозные пузырьки (5). [Wei-
bel (1969). The ultrastructure of the alveolarcapillary membrane of barrier. In:
The pulmonary circulation and interstitial space (ed, Fishman, Hecht), University
of Chicago Press.]
570 Часть II. Механика кровообращения
дечно-сосудистой системы. Альвеолярная сторона тканевого барье-
ра, т. е. поверхность альвеолярного эпителия, покрыта слоем, со-
стоя'щим из лецитина, возможно связанного с белками. Этот слой
называется легочным сурфактантом. Он играет важную роль в ме-
ханике легких и оказывает влияние на трансмуральное давление
в капиллярах малого круга.
Из альвеолярных капилляров кровь поступает в посткапилляр-
ные сосуды, а затем в венулы и вены малого круга1). Вены ма-
лого круга проходят в перегородках, разделяющих дольки легких,
и в отличие от артерий этого круга большую часть своего пути
идут вдали от бронхов. Легочные вены покидают каждое легкое
через его ворота и впадают в левое предсердие, обычно по две
вены от каждого легкого (рис. 15.1). Клапанов в этих венах нет.
Стенка легочных вен, как и вен большого круга, тонкая (табл. I).
В стенке мелких вен малого круга (диаметром 0,01—0,1 см) со-
держатся одиночные эластические волокна и соединительная ткань;
гладких мышц в ней почти нет. В стенке вен среднего размера
имеется некоторое количество таких мышц и эластических воло-
кон; наиболее крупные легочные вены, хотя и обладают средней
оболочкой, состоящей из гладкомышечных клеток, в основном об-
разованы толстой наружной оболочкой, построенной из коллаге-
новых волокон.
Сведения о сосудах, питающих стенку артерий малого круга
(vasa vasorum), крайне ограничены. Некоторые из этих артериаль-
ных веточек отходят от артерий бронхов, хотя по крайней мере у
кролика имеются также добавочные веточки, отходящие от коро-
нарных артерий. Наиболее крупные питающие сосуды (диаметром
50—100 мкм) проходят в наружной оболочке артерий малого круга,
тогда как в средней оболочке встречаются лишь более мелкие пи-
тающие веточки. Проникают ли капилляры во внутреннюю обо-
лочку, неизвестно. Вены, выносящие кровь из стенки легочных ар-
терий, идут параллельно снабжающим эту стенку артериям и впа-
дают в ближайшие вены большого круга. Артерии и вены малого
круга, а также бронхи окружены лимфатическими каналами; по
ним осуществляется отток лимфы также из межклеточного про-
странства, окружающего терминальные воздухоносные пути.
Кровоснабжение бронхов. Обычно каждое легкое снабжает
пара бронхиальных артерий. Они ответвляются от аорты и входят
в соответствующее легкое вместе с его главным бронхом. На про-
тяжении всего этого пути обе артерии, многократно связанные
друг с другом сосудами-перемычками, отдают ветви к плевре и в
конце концов делятся, обеспечивая питание всех бронхов. Сосуди-
*) Между артериями и венами малого круга существуют, по-видимому, ка-
кие-то прямые связи, в обход альвеолярных капилляров Сообщают, что такие
анастомозы обнаруживаются вблизи терминальных воздухоносных пространств и
имеют диаметр вплоть до 500 мкм Их физиологическое значение не установлено.
Гл. 15. Малый круг кровообращения 571
стая система бронхов простирается до терминальных бронхиол и
доставляет кровь в vasa vasorum легочных артерий (вплоть до
артерий диаметром 0,1 см), а также к перибронхиальным нервам.
Вены бронхов впадают в верхнюю полую вену и в меньшем чис-
ле— в вены малого круга.
Методом инъекций установлено существование в нормальных
легких прекапиллярных анастомозов, соединяющих сосудистое
русло бронхов с малым кругом. Диаметр этих сосудов составляет
от 20 до 200 мкм, и обнаруживаются они главным образом в об-
ласти мелких бронхов. Физиологическая роль их неясна, хотя
предполагается, что они поддерживают кровоснабжение неболь-
ших областей легких, которые временно не вентилируются или
оказываются спавшимися.
15.2. Трансмуральное давление и статические упругие свойства
сосудов
В этом разделе мы рассмотрим статические упругие свойства
сосудов малого круга, а также силы, которые на них действуют.
Реальное трансмуральное давление, действующее на артерии ма-
лого круга, с трудом поддается измерению in situ по двум причи-
нам. Первая заключается в том, что из-за гидростатических влия-
ний давление внутри артерий меняется в зависимости от располо-
жения их в легких. Вторая причина связана с тем, что реальное
значение давления, действующего на эти сосуды снаружи, зависит
от механических свойств легких, от давления в грудной полости и
от степени наполнения легких. Поэтому необходимо рассмотреть
подробно те факторы, которые определяют величину этих давле-
ний, а также упругие свойства стенки сосудов малого круга.
Давление внутри сосудов. Малый круг кровообращения являет-
ся системой низкого давления: у здорового человека среднее избы-
точное давление (превышение над атмосферным давлением) в пра-
вом желудочке и в крупных легочных артериях составляет при-
мерно 2-103 Н-м-2 (15 мм рт. ст., или 20 см. вод. ст.), хотя при
мышечной работе оно может повышаться примерно до 4-103 Н-
• м~2 (рис. 15.11). Среднее избыточное давление в легочных венах
составляет примерно половину указанной величины; это свиде-
тельствует о том, что падение давления в системе микрососудов
малого круга гораздо меньше, чем в системе микрососудов боль-
шого круга. У стоящего человека размер легких по вертикали
составляет около 30 см, причем артерии входят в них примерно
посередине. Градиент гидростатического давления в артериях ма-
лого круга составляет примерно 0,1 «103 Н-м~2 на 1 см, так что
у верхушки легкого давление оказывается примерно на 1,5-103 Н-
•м-2 (15 см вод. ст.) ниже среднего избыточного давления в круп-
ных легочных артериях. Точно так же у основания легких давле-
ние оказывается примерно на 1,5-103 Н-м-2 (15 см вод. ст.) выше.
872 Часть 11. Механика кровообращения
При движении крови падение давления между главными легоч-
ными артериями и капиллярами составляет примерно 0,7-103 Н-
• м-2. Таким образом, в обычных условиях среднее давление в мик-
рососудах верхней части легких оказывается ниже атмосферного.
Давление вне сосудов. Кровеносные сосуды малого круга мож-
но разделить на две группы: I) мелкие сосуды, расположенные
в альвеолярной перегородке и вокруг нее, их называют альвеоляр-
ными сосудами-, 2) более крупные, экстра-альвеолярные сосуды,
которые в свою очередь можно подразделить на интра- и экстра-
паренхимальные сосуды, т. е. на расположенные внутри и вне соб-
ственно ткани легких (хотя мы и используем такие обозначения,
в действительности, как отмечено выше, более крупные сосуды ле-
жат в соединительнотканных оболочках, т. е., строго говоря, не
в самой паренхиме легких). Основания для разделения сосудов на
указанные две группы не анатомические, а функциональные: на со-
суды этих двух групп действует разное по величине внешнее дав-
ление. Это можно показать на удаленных из тела легких, заполняя
их артерии и вены керосином — жидкостью, которая не может
войти в более мелкие сосуды из-за поверхностного натяжения.
Глубина проникновения керосина в сосудистое русло определяет-
ся равновесием между перепадом гидростатических давлений и по-
верхностным натяжением на поверхности раздела керосина и вы-
тесняемой им жидкости. Вычисления показывают, что при раз-
ности гидростатических давлений 2-Ю3 Н-м-2 граница раздела
вода — керосин должна быть на уровне сосудов диаметром 50 мкм.
Ёсли наполнить легкие воздухом, повысив давление его в альвео-
лах (без изменения давления на поверхности легких или в резер-
вуаре с керосином, т. е. произвести «наполнение при положитель-
ном давлении»), то объем сосудов, заполненных керосином, уве-
личится. Однако в случае, когда все сосудистое русло заполнено
водным раствором полигклюкина, такое же воздействие приводит
к меньшему увеличению объема сосудов или даже к его умень-
шению. Это можно объяснить только уменьшением объема мелких
сосудов. Чтобы разобраться в механизме такого явления, надо
выяснить, какие факторы определяют величину давления, дей-
ствующего на сосуды разного калибра снаружи.
Рассмотрим сначала экстра-альвеолярные интрапаренхималь-
ные сосуды (рис. 15.6,А). По ряду причин давление, действующее
на них со стороны окружающих сосуды тканей, т. е. оболочек
(обозначим его Рос), не равно давлению газа в альвеолах раЛьв-
Во-первых, если вообще соседние альвеолы заполнены воздухом,
то их мембраны действуют на оболочку, окружающую околососу-
дистое пространство; их растяжение равносильно снижению ральв на
величину, которую можно назвать «давлением упругого растяже-
ния» Рупр. раст.- Однако влияние этого давления частично компен-
сируется действием сил упругого восстановления самой околосо-
судистой оболочки (за вычетом давления, которое оказывают на
Гл. 15. Малый круг кровообращения 573
Б
Рис. 16.6. Схемы, поясняющие влияние
различных факторов иа давление внутри
сосудов рВс А Экстра-альвеолярный ни-
трапаренхимальный сосуд, окруженный
ОКОЛОСОСУДИСТЫМ пространством (Рос —
давление в этом пространстве), которое
ограничено оболочкой. Эта оболочка рас-
тягивается мембранами альвеол
(Pynp раст — фактическая величина соот-
ветствующего давления, направленного
наружу) и отталкивается назад к сосу-
ду действующими в одном направлении
Ральв — давлением в альвеолах и
рупр восст — собственным давлением вос-
становления вместе с давлением, обус-
ловленным действием нерегулярных со-
единений между околососуднстой обо-
лочкой и стенкой сосуда. Б. Схема, пояс-
няющая тот факт, что околососудистое
пространство является, за исключением
нерегулярных мест соединения околосо-
судистой оболочки со стеикой сосуда,
продолжением полости плевры, давление
Рпл в которой близко по величине к око-
лососудистому давлению рм (см. текст).
В. Альвеолярный сосуд: давление в око-
лососудистом пространстве приблизи-
тельно равно давлению в альвеолах; не-
большое различие обусловлено слабым.
Направленным наружу натяжением стен-
ки сосуда альвеолярной мембраной и
слоем сурфактанта (рсурф). '
574 Часть II. Механика кровообращения
(16.1)
стенку сосуда снаружи любые упругие связи, соединяющие ее с
околососудистой оболочкой). Это давление мы обозначим рупр.восст..
Таким образом,
Рос = Ральв Рупр. раст. Ч" Рупр •ВОССТ
Если бы легкие были однородными, то все альвеолы наполнялись
бы воздухом одинаково и на каждой поверхности паренхимы лег-
ких, а также на наружной поверхности плевры давление упругого
растяжения было бы одинаковым (рис. 15.6). В статических усло-
виях при любом объеме легких раЛьВ всюду одинаково, потому что
все воздухоносные пути сообщаются между собой. По этой же
причине давление в полости плевры рпл, чрезвычайно узкой щели
между двумя листками плевры — висцеральным, покрывающим
легкие, и париетальным, выстилающим грудную клетку, — отли-
чается от рос лишь в том случае, если давление упругого восста-
новления околососудистой оболочки рупр. восст. отличается от сум-
марного давления восстановления обоих листков плевры. Для
экстрапаренхимальных сосудов рос равно рпл, так как в этом
случае околососудистым пространством является плевральная
щель.
Когда легкие раздуваются под действием положительного дав-
ления, увеличивается ральв и еще сильнее увеличивается рупр. раст.,
что обусловлено нелинейными упругими свойствами альвеолярных
мембран, которые по мере их растяжения становятся все более
жесткими. В конечном счете раздувание легких приводит к паде-
нию рос, и поэтому, если давление внутри данного сосуда остается
неизменным, трансмуральное давление возрастает и объем сосуда
также возрастает. Таково объяснение наблюдений в опытах с
введением керосина в сосуды легких, удаленных из тела.
Тот факт, что механические свойства легких, находящихся
внутри грудной клетки, неоднородны, не влияет на сделанные
выше выводы. Например, из-за того, что давление на поверхности
плевры (давление на поверхности легких) по вертикали неодина-
ково, объем альвеол в верхушках легких оказывается больше
объема альвеол у основания (изменение давления на поверхности
плевры по вертикали определяется в первую очередь характером
распределения веса легких между поддерживающими их струк-
турами грудной клетки). Вертикальный градиент давления на по-
верхности легких составляет примерно 25 Н-м-2 (0,25 см вод. ст.)
на 1 см. Это соответствует лишь четверти того значения градиента
гидростатического давления, которое можно было бы ожидать,
если бы все части плевральной щели свободно сообщались между
собой (в действительности это условие не выполняется). Можно
думать, что примерно такой градиент по вертикали должен быть
для рос, но даже на уровнях, различающихся по высоте на 30 см
(как для легких стоящего человека), полное изменение давления
в окружающем сосуды пространстве не превышает 0,75-103 Н-м~2
Гл. 1В. Малый круг кровообращения 575
(7,5 см вод, ст.), тогда как изменение внутрисосудистого давле-
ния составляет 3-103 Н-м2 (30 см вод. ст.).
Обратимся теперь к альвеолярным сосудам (рис. 15.6,В).
Можно считать, что кровь в этом случае движется между двумя
слоями эндотелия капилляров; эти слои удерживаются на опреде-
ленном расстоянии друг от друга распорками, покрытыми эндо-
телием (рис. 15.4). Эндотелий капилляров окружен тонким слоем
соединительной ткани, затем следуют альвеолярный эпителий и
тончайший слой сурфактанта, выстилающий альвеолы. Давление в
пространстве, окружающем альвеолярные сосуды, почти совпадает
с давлением в альвеолах: небольшое различие обусловлено только
натяжением самих мембран альвеол, направленным наружу,
и слоем сурфактанта (рСурф). Такое заключение подтверждают
опыты на удаленных из тела легких, заполненных жидкостью и
погруженных в жидкость: когда давление в их артериях оказы-
валось равным давлению в альвеолах, кровоток в легких прекра-
щался. В этих условиях влияние сил поверхностного натяжения
на границе раздела сурфактант — жидкость, так же как и верти-
кальных градиентов трансмурального давления и давления в со-
судах, сводится практически к нулю. Если же изолированные лег-
кие помещали в воздушную среду и раздували воздухом, кровоток
в них прекращался только при равенстве артериального давления
венозному даже при условии, что венозное давление было не-
сколько ниже давления в альвеолах. Правда, каков вклад сурфа-
ктанта в величину трансмурального давления в этих условиях, не-
известно.
Если легкие очень медленно раздуваются in situ за счет рас-
ширения грудной клетки, то ральв остается фактически равным
атмосферному давлению, тогда как рпл, а следовательно, и рос сни-
жаются. В этом случае именно снижение рос, в соответствии с урав-
нением (15.1), компенсирует повышение рупр. раст, вызываемое рас-
тяжением мембран альвеол. Снижение рос приводит к увеличению
трансмурального давления в экстра-альвеолярных сосудах. Но,
поскольку ральв не меняется, трансмуральное давление в альвео-
лярных сосудах меняется мало или не меняется вовсе, и потому
они не спадаются, как в случае раздувания легких при положи-
тельном давлении. Однако ввиду того, что альвеолярная перего-
роцка при этом растягивается, просвет их все-таки уменьшается.
Рассмотрим, что происходит, когда давление внутри какого-
нибудь кровеносного сосуда снижается, а значения остальных дав-
лений поддерживаются постоянными. Снижение давления в сосуде
приведет к падению трансмурального давления. Если оно станет
отрицательным, что в обычных условиях возможно вблизи вер-
хушки легкого, то может произойти спадение сосуда: в случае
альвеолярных сосудов именно это и имеет место. Площадь попе-
речного сечения экстра-альвеолярных сосудов может несколько
уменьшиться, но само это уменьшение приведет к местному растя-
676 Часть II. Механика кровообращения
жению мембраны альвеол. В результате рупр. раст. в этой области
повысится; рос при этом уменьшится [в соответствии с уравнением
(15.1)] и будет препятствовать спадению сосуда. Это важное
явление взаимозависимости означает, что в условиях, когда
легкие наполнены воздухом, их паренхима фактически уменьшает
растяжимость находящихся в ней кровеносных сосудов (или воз-
духоносных путей). Количественно степень такого уменьшения для
артерий малого круга пока еще не определяли. Это ограничивает
ценность каких бы то ни было предсказаний, которые можно сде-
лать, например, для скорости распространения пульсовой волны
по артериям малого круга при разной степени наполнения легких
воздухом.
Упругие свойства. Свойства удаленных из легких артерий и вен
лучше всего рассматривать, сравнивая их со свойствами вен боль-
шого круга и принимая во внимание сведения, изложенные в гл. 14,
в особенности представленные на рис. 14.1. Как и вены, артерии
малого круга при трансмуральном давлении (2,0—2,5)-103 Н-м-2
(20—25 см вод. ст.) имеют круглое поперечное сечение, но когда
трансмуральное давление снижается до 0,5-103 Н-м-2 (5 см вод. ст.),
сечение их перестает быть круглым. Это относится также к сосу-
дам целых легких кролика, удаленных из грудной клетки. По-
этому можно ожидать, что сосуды легких способны менять свою
форму указанным образом и in vivo. Упругие свойства сегмента
крупной артерии легких собаки (в том диапазоне изменений транс-
мурального давления, в котором она сохраняет примерно круглое
сечение) сопоставлены с упругими свойствами вен и артерий боль-
шого круга собаки на рис. 14.3. При физиологических значениях
трансмурального давления модуль Юнга этой артерии близок к
модулю Юнга артерий большого круга, но она более растяжима,
потому что стенка ее значительно тоньше стенок сопоставимых
по диаметру артерий большого круга. Вместе с тем по сравнению
с венами большого круга указанная артерия менее растяжима.
Это обусловлено тем, что, хотя стенки легочной артерии и вен
большого круга сравнимы по толщине, модуль Юнга стенки ле-
гочной артерии выше. При одинаковых значениях трансмураль-
ного давления наиболее растяжимыми оказываются артерии боль-
шого круга, а наименее — вены большого круга.
На крупных венах малого круга подобных исследований не
производили. Но упругие свойства средних и мелких артерий и
вен малого круга собаки (диаметр 0,08—0,36 см) определены. Для
этого осуществляли перфузию удаленных из грудной клетки лег-
ких; диаметр сосудов измеряли рентгенографическим методом.
Усредненные данные о диаметре сосудов при различных давлениях
показаны на рис. 15.7. Из рисунка видно, что при изменении внут-
рисосудистого давления в диапазоне (1,0—3,5) -103 Н-м-2 (10—
35 см вод. ст.), когда сечение сосудов, как можно ожидать, почти
круглое, артерии, диаметр которых больше 0,16 см, увеличиваются
Гл 15 Малый круг кровообращения 577
а
200 г
100
200
150
150
Диаметр сосудов
2800 - 3600 мч •
100*-
200 Г
d150 -
| 100 L
а 200 г
200
150
Диаметр сосудов
150
100
0
Среднее давление, см вод ст
150
1200 - 1600 мкм
*---•/
«£^>.£0--$ II
йЬо-^гоо мкм
ГЛ—р
10 20 30 40 50
Среднее давление, см вод ст
100 L
0
Рис. 15 7 Соотношение давление — диаметр для легочных артерий (7) и вен (77)
собаки. Кривые объединены попарно в соответствии с диаметром сосудов, зна-
чения которого приведены в процентах по отношению к его значению при вну-
трисосудистом давлении 8—10 см вод ст Давление наполнения легких —
10 см вод ст [Maloney, Rooholamini, Wexler (1970). Pressure-dtameter relations
of small blood vessels in isolated dog lung. Microvasc Res., 2, 1—2.]
в диаметре почти линейно, тогда как для более мелких артерий
эта зависимость линейна лишь в начале указанного диапазона, а
потом рост замедляется. Для вен изменение диаметра на единицу
изменения давления значительно меньше; кроме того, в отличие
от артерий диаметр средних и относительно крупных вен (диамет-
ром более 0,12 см) с увеличением давления стремится достичь
некоторого постоянного уровня.
На основании некоторых данных, представленных на рис. 15.7,
рассчитаны средние значения растяжимости D [уравнение (14.1)].
При внутрисосудистом давлении (1,0—2,25) -103 Н-м-2 (10—
22,5 см вод. ст.) растяжимость артерий составляет от 0,35-10-3 м2.
•Н-1 для самых крупных сосудов до 0,88-10-3 м2-Н-1 для самых
мелких Растяжимость наиболее крупных вен равна при тех же
давлениях примерно 0,17-10-3 м2-Н-1, а наиболее мелких — около
0,35-10-3 м2-Н-1. При растяжимости 0,35-10-3 м2-Н-1 расчетное
значение скорости распространения пульсовой волны по артериям
малого круга [уравнение (14.6)] составляет 1,7 м-с-1, что сопоста-
вимо с величинами, получаемыми при измерении этой скорости
для легочного ствола собаки, — около 2,5 м-с-1 (см. табл. I).
Упругие свойства стенки артерий малого круга изучали также
путем измерения кажущейся скорости распространения искус-
ственно создаваемых низкоамплитудных волн давления (для лег-
ких кролика). На путях различной длины — от легочного ствола
до разных точек артериального дерева малого круга — опреде-
ляли средние значения скоростей Были получены следующие дан-
ные: расстояние 2 см —скорость 0,55 м-с-1, расстояние 3,2 см —
скорость 0,62 м-с-1, расстояние 5,5 см —скорость 0,80 м-с-1. Как
578 Часть II. Механика кровообращения
Рис. 15.8. Зависимость числа эритроцитов, приходящихся на 10 мкм длины аль-
веолярной перегородки, от перфузионного давления. Поскольку давление в ар-
териях легких повышается в направлении сверху вниз примерно на 1 см вод. ст.
(102 Н-м~2) на 1 см, величина перфузионного давления соответствует расстоя-
нию от рассматриваемого участка легкого до его верхушки, и в области вер-
хушки легкого оно равно нулю. Венозное давление низкое. Приведены резуль-
таты измерений при значениях транспульмонального давления 1,0-10s Н-м“2
(кривая •) и 2,5-103 Н-м 2 (кривая X). Обратите внимание на то, что при
транспульмональном давлении 1,0-10s Н-м-2 число эритроцитов по мере удаления
от верхушки увеличивается быстро, а при наполнении легких большим объемом
воздуха (транспульмональиое давление 2,5-10s Н-м 2)—значительно медлен-
нее. [West, Glazier, Hughes, Maloney (1969). Pulmonary capillary flow, diffusion
ventilation and gas exchange. In: Ciba Foundation Symposium on Circulatory and
Respiratory Mass Transport, pp. 256—272, J. and A. Churchill, Edinburgh.]
и в случае артериального дерева большого круга, кажущаяся ско-
рость волны явно увеличивается к периферии.
Приведенные выше результаты относятся к артериям и венам
малого круга. Мы должны рассмотреть также упругие свойства
альвеолярных капилляров, которые образуют в альвеолярной пе-
регородке канал. Капилляры альвеол растяжимы. Установили это
путем подсчета эритроцитов в альвеолярных капиллярах на по-
лученных методом быстрого замораживания препаратах изоли-
рованных перфузируемых легких. Число эритроцитов сопостав-
ляли со значением перфузионного давления непосредственно пе-
ред замораживанием (рис. 15.8).
При значении растягивающего легкие давления (транспульмо-
нального давления) 1,0-103 Н-м-2 (10 см вод. ст.) повышение пер-
Гл. 15. Малый круг кровообращения 579
фузионного давления приблизительно от 0 до 3,5 • 103 Н • м~2 со-
провождалось почти линейным увеличением числа эритроцитов
примерно от 0,4 до 1,5 на 10 мкм длины альвеолярной перего-
родки. При транспульмональном давлении 2,5-103 Н-м~2 число
эритроцитов, отнесенное к единице длины перегородки, увеличи-
валось с ростом перфузионного давления значительно медленнее.
Обнаружено также независимое влияние объема легких на раз-
меры альвеолярных капилляров. На рис. 15.9 приведены микро-
фотографии гистологических срезов быстрозамороженных кусоч-
ков легкого. В обоих случаях кусочки брали из области, располо-
женной на 32—35 см ниже того уровня, где давление в венах в
точности равно давлению в альвеолах, так что трансмуральное
давление в капиллярах в обоих случаях одинаково, но давление
в альвеолах в случае, соответствующем рис. 15.9, Л, равно 1,0-
•103 Н-м-2 (10 см вод. ст.), а рис. 15.9,Б — 2,5-Ю3 Н-м-2
(25 см вод. ст.). В соответствии с более высоким транспульмональ-
ным давлением в случае Б размеры альвеол больше, а число эрит-
роцитов в альвеолярной перегородке резко уменьшено. Такие из-
менения, вероятно, обусловлены большим натяжением стенок аль-
веол при большем объеме легких [растяжение листа упругого ма-
териала в двух направлениях—в плоскости листа — приводит к
уменьшению размера в третьем направлении (разд. 7.1)].
Для прямого измерения размера канала в альвеолярной пере
городке в сосуды легких вводили при разных давлениях жидкий
силоксановый каучук, обладающий низкой вязкостью, и давали
ему затвердеть. Оказалось, что толщина альвеолярного канала уве-
личивается с ростом перфузионного давления приблизительно ли-
нейно, и при увеличении объема легких наклон соответствующей
прямой (пропорциональный растяжимости) уменьшается
(рис. 15.10). Мы уже знаем, что in situ размеры альвеол изме-
няются в зависимости от их положения в легких по высоте. По-
этому можно ожидать соответствующих различий и в размерах
альвеолярных капилляров.
Объем крови в малом круге. Вопрос об объеме крови в сосудах
малого круга и о распределении этого объема между различ-
ными сосудами крайне важен как для физиологии кровообраще-
ния, так и для клинической медицины.
Объем крови в сосудистом русле малого круга и в составляю-
щих его артериях, капиллярах и венах оценивают разными мето-
дами. Наиболее распространенный метод заключается во введении
индикатора в легочный ствол и последующем анализе проб крови
из левого предсердия для выявления момента поступления сюда
индикатора. Таким путем можно рассчитать среднее время про-
хождения крови по сосудам малого круга. Если в то же самое
время каким-либо независимым методом измерить также минут-
ный объем сердца, то можно определить объем сосудистого русла
малого круга, так как он является произведением этих двух вели-
580 Часть II. Механика кровообращения
Гл. 15. Малый круг кровообращения 581
чин. У здорового человека эта величина составляет 0,5 л, или
10% объема циркулирующей крови. Объем сосудистого русла ма-
лого круга собаки оценивается величиной около 0,2 л.
Объем крови в артериях малого круга собаки измеряли in vivo
видоизмененным методом разведения индикатора. Измеренный та-
ким способом объем этих артерий в среднем составляет около
50 см3, т. е. примерно одну четверть полного объема крови в
малом круге. Один из вариантов этого метода позволил опреде-
лить также объем вен малого круга собаки; он оказался прибли-
зительно равным объему артерий. Следовательно, объем капилля-
ров малого круга составляет примерно половину полного объема
крови в этом сосудистом русле.
Объем слепка артериального дерева малого круга человека
составляет примерно 150 мл, причем около 80% этой величины
приходится на несколько сотен сосудов, диаметр которых больше
0,13 см. Остальная часть приходится на миллионы более мелких
сосудов. Однако к этим данным следует относиться осторожно, так
как вполне возможно, что измерения на мелких сосудах неточны.
Подобные сведения о венах малого круга отсутствуют.
Объем крови в капиллярах малого круга определяют in vivo
при помощи метода, основанного на измерении диффузионной спо-
собности легких по отношению к окиси углерода. Эта величина
зависит от позы испытуемого (см. ниже), но у здорового человека,
находящегося в горизонтальном положении, составляет примерно
100 мл. Если допустить, что оба приведенных результата, относя-
щихся к объему артериального русла малого круга и объему его
капилляров, верны, то можно считать, что вены малого круга че-
ловека содержат примерно половину всего объема крови, находя-
щегося в малом круге. Отметим также, что если считать минут-
ный объем сердца человека равным 5 л-мин-1, то время прохож-
дения эритроцитов через капилляры легких составит всего около
1 с. Как показывают вычисления, при тяжелой мышечной работе
это время не превышает 0,5 с.
Объем крови в сосудах малого круга может изменяться в зави-
симости от физиологических условий. Измерения, произведенные
на человеке, лежащем горизонтально на спине, показывают, что
при поднимании ног производительность сердца, давление в ле-
гочной артерии и в легочной вене, а также полный объем крови
в малом круге увеличиваются. Если перевести испытуемого из
горизонтального положения в вертикальное (его мышцы при этом
Рис. 15.9. Гистологические срезы быстро замороженных кусочков перфузируемых
легких собаки. Показано независимое от трансмурального давления в капиллярах
влияние транспульмонального давления на капилляры альвеол. Описание иссле-
дования приведено в тексте. [Glazier, Hughes, Maloney, West (1969). Measure-
ments of capillary dimensions and blood volume in rapidly frozen lungs J appl
Physiol., 26, 65—76.]
582 Часть II. Механика кровообращения
Рис. 15.10. Зависимость средней толщины слоя крови от трансмурального давле-
ния в капиллярном канале альвеолярной перегородки. Сосуды малого круга
кошки заполнили жидким силоксановым каучуком низкой вязкости; это вещество
отвердевает в сосудах без усадки. В диапазоне трансмурального давления
(0,6—2,7)-10s Нм"2 зависимость приблизительно линейна. При нулевом транс-
муральном давлении (экстраполяция) толщина слоя составляет 4,28 мкм. [Sobin,
Fung, Trewer, Rosenquist (1972) Elasticity of the pulmonary alveolar microvascu-
lar sheet in the cat. Circulation Res., 30, 440—450.]
не должны напрягаться), производительность сердца падает, дав-
ление в легочной артерии и в левом предсердии несколько сни-
жается и полный объем крови в малом круге уменьшается на
50—100 мл. Однако, как показано на рис. 15.11, когда испытуемый
сам поднимается из горизонтального положения в вертикальное,
давление в легочной артерии существенно не изменяется. Это гово-
рит о том, что тонус вен, расположенных ниже уровня сердца, уве-
личивается в достаточной мере, чтобы не допустить излишнее скоп-
ление в них крови.
На рис. 15.11 показано также влияние мышечных упражнений
на давление в легочной артерии. Когда испытуемые спокойно
стояли, среднее давление в легочной артерии составляло 15 мм
рт. ст. (2,0-103 Н-м-2), а при ходьбе со скоростью 4,8 км-ч~’ по
тредбану (наклон 16:100) это давление повышалось примерно до
30 мм рт. ст. (4,0-10s Н-м~2). Потребление О2 при этом составляло
в среднем 2,35 л-мин-1 (при 0,35 л-мин-1 в покое), 8 минутный
объем сердца достигал 17,5 л-мин~* (при 5,1 л-мин-1 в покое).
Гл 15 Малый круг кровообращения 583
Рис. 15.11. Среднее давление в легочной артерии (10 мм рт. ст = 1,36-10’Н-м~2)
у 24 здоровых испытуемых. Приведены индивидуальные и средние (±а) зна-
чения в условиях, когда испытуемый лежит горизонтально на спине, спокойно
стоит и когда он идет со скоростью 4,8 км-час-1 по горизонтальному тредбану
и по наклонному с наклоном к горизонтальной плоскости 8:100 и 16:100.
[Damato, Galante, Smith (1965). Haemodynamic response to treadmill exercise
in normal subjects. J. appl. Physiol., 21, 959—966.]
Ни давление в легочных венах, ни полный объем крови в малом
круге в этих исследованиях не измеряли. Однако другие иссле-
дования показали, что мышечная работа едва ли вызывает какие-
либо изменения давления в левом предсердии, хотя объем крови в
малом круге при этом увеличивается. Например, у здоровых взрос-
лых испытуемых во время мышечной работы, сопровождающейся
увеличением потребления О2 от 0,14 л-мин-^м-2 до 0,49 л-мин-1-
•м-2 (работа менее тяжелая, чем в упомянутых выше исследо-
ваниях), давление в легочной артерии повышалось в среднем от
13,9 мм рт. ст. до 17,3 мм рт. ст. [(1,9—2,4)-103 Н-м~2], а объем
крови в малом круге возрастал в среднем от 0,28 л-м~2 до 0,35 л-
584 Часть 11. Механика кровообращения
400г
300
200
100
о -------------1-----------1-----------1------------1
1,0 2,0 3,0 4,0
Давление в легочной артерии « W3 Н • м-2
Рис. 15.12. Зависимость полного объема крови в сосудах перфузируемых легких
собаки от давления в легочной артерии, которое повышается за счет увеличения
притока крови. Давление в левом предсердии поддерживается постоянным:
0,5-Ю3 Н-м-2 — светлые значки, 3.3-103 Н-м 2 — темные значки. [Maseri, Cal-
dini, Harward, Joshi, Permutt, Zierler (1972). Determinants of pulmonary vascular
volume. Circulation Res., 31, 218—228.]
• m—2 (значения потребления Ог и объема крови в малом круге оп-
ределялись в расчете на единицу площади поверхности тела).
Исследования на легких собаки подтвердили данные о том,
что именно повышение давления в легочной артерии в первую оче-
редь ответственно за увеличение объема сосудистого русла малого
круга при увеличении расхода крови в нем. Так, при перфузии
легких in situ насосом в условиях, когда давление в легочной ар-
терии н в левом предсердии можно было изменять независимо
друг от друга, полный объем крови в малом круге не изменялся,
если расход снижали за счет повышения давления в левом пред-
сердии, а давление в легочной артерии поддерживали постоянным.
Однако, как видно из рис. 15.12, если постоянным поддерживали
давление в левом предсердии, а давление в легочной артерии по-
вышали за счет увеличения расхода крови, полный объем крови
в малом круге увеличивался. При давлении в левом предсердии
5 см вод. ст. и давлении в альвеолах, превышающем на всем
протяжении легких давление в их венах (зона II, разд. 15.4), обу-
словленное повышением давления в артериях увеличение объема
составляло 12 мл на 1 см вод. ст. Когда давление в левом пред-
Гл. 15. Малый круг кровообращения 585
сердии составляло 33 см вод. ст. и давление в венах малого круга
на всем протяжении легких превышало давление в альвеолах
(зона Ш, разд. 15.4), эта величина была равной 7 мл на
1 см вод. ст. Для объяснения такого различия было выдвинуто
предположение, что в первом случае повышение давления приво-
дит к раскрытию большего числа ранее закрытых каналов. К во-
просу о раскрытии дополнительных каналов мы вернемся ниже.
Не следует упускать из виду, что в подобных исследованиях
изменение механического ответа сосудов малого круга отчасти
обусловлено активным изменением тонуса их гладких мышц. Ни-
каких данных об активном изменении растяжимости артерий ма-
лого круга при мышечной работе нет. Однако при перфузии лег-
ких собаки с помощью насоса, создающего пульсирующий поток
крови, было обнаружено, что раздражение симпатических нервов,
идущих к легким, приводит к уменьшению растяжимости артерий
малого круга без изменения сопротивления его сосудов.
Объем крови, содержащейся в капиллярах малого круга, также
изменяется в зависимости от физиологических условий. Воздей-
ствия, которые приводят к перемещению крови в сосуды, находя-
щиеся в грудной полости, а именно переход испытуемого из вер-
тикального положения в горизонтальное или повышение давления
в специальном костюме, охватывающем живот и ноги испытуемых,
увеличивают объем крови в капиллярах легких в среднем от 70 до
100 мл. Следует, однако, помнить о том, что способность капил-
ляров легких растягиваться ограничена. Например, дальнейшее
изменение положения тела испытуемого — наклон вниз головой под
углом 60° к горизонтальной плоскости, — хотя и увеличивает транс-
муральное давление в сосудах легких, не вызывает дополнитель-
ного прироста объема крови в их капиллярах. Мышечная работа
также приводит к увеличению объема крови в капиллярах ма-
лого круга: при максимальной нагрузке он может вырасти больше
чем в 2 раза. Как и в опытах, результаты которых представлены
на рис. 15.12, это сопровождается повышением давления в легоч-
ной артерии, но не в легочной вене.
На объем крови в капиллярах малого круга влияет также
объем легких: у здоровых испытуемых при максимальном вдохе он
обычно несколько меньше, чем после спокойного выдоха. По-види-
мому, это можно связать с уже упомянутыми наблюдениями на
удаленных из тела легких, когда увеличение транспульмонального
давления приводило к уменьшению числа эритроцитов в перего-
родках альвеол.
Возможны два механизма увеличения объема крови в капил-
лярах легких при повышении давления в легочной артерии: растя-
жение уже открытых капилляров и раскрытие ранее спавшихся
капилляров. Известно, что у стоящего человека в состоянии покоя
капилляры в области верхушки легкого обычно спадаются, и кро-
воток через них очень мал, но при мышечной работе, когда дав-
586 Часть 11. Механика кровообращения
Перфузионное давление * 10‘3 н м 2
Рис. 15 13 Зависимость числа открытых капилляров на единицу длины альвео-
лярной перегородки от давления в легочной артерии. Результаты подсчета на
гистологических препаратах быстрозамороженных кусочков легких собаки. [War-
rell, Evans. Clarke, Kingaby, West (1972) Pattern of filling in the pulmonary
capillary bed. J. appl. Physiol, 32, 346—356 [
ление в легочной артерии повышается, кровоток в них увеличи-
вается. Результаты гистологических исследований быстро заморо-
женных кусочков легких собаки указывают на то, что раскрытие
новых капилляров в этой области играет очень важную роль,
когда давление в альвеолах ниже давления в артериях малого
круга, но выше давления в его венах (разд. 15.4).
Как видно из рис. 15.13, при повышении давления в легочной
артерии число открытых капилляров на единицу длины альвеоляр-
ной перегородки быстро увеличивается примерно от 25 мм-1 при
давлении 1 см вод. ст. до 55 мм-1 при давлении 12 см вод. ст., од-
нако дальнейшее повышение давления изменений почти не вызы-
вает. Не приведенные здесь результаты, полученные в этом же
исследовании, показывают, что когда перфузионное давление под-
нимается выше 12 см вод. ст., длина альвеолярной перегородки,
занимаемая каждым капилляром, резко возрастает. Последнее сви-
детельствует о том, что теперь происходит растяжение капилля-
ров, а не увеличение числа открытых капилляров. Эти условия
соответствуют положению в области верхушки легких стоящего
человека: в этой области легких имеется резерв нераскрытых ка-
пилляров, который может быть использован при повышении дав-
ления в артериях малого круга, что и происходит при мышечной
работе.
Г л. 15. Малый круг кровообращения 587
Чтобы определить относительный вклад в увеличение объема
крови, содержащейся в капиллярах малого круга, их растяжения
и раскрытия, нужно знать механизм раскрытия ранее закрытых
сосудов, значение давления, необходимого для раскрытия, и рас-
тяжимость этих капилляров при различных трансмуральных дав-
лениях. Предполагали, что закрываться способны артериолы лег-
ких и что при повышении давления в артериях малого круга они
действительно раскрываются. На самом деле имеющиеся данные
указывают на то, что существенно закрытие капилляров, а не арте-
риол. Даже если давление, необходимое для раскрытия отдель-
ных участков капилляров, очень мало, из-за различия в длинах
путей и сопротивлениях сосудистого русла выше областей, в кото-
рых находятся закрытые капилляры, для раскрытия всех сосудов
может потребоваться очень высокое артериальное давление. Этот
вопрос изучен путем моделирования на ЭВМ.
15.3. Динамика течения крови в крупных сосудах малого круга
В этом разделе мы рассмотрим формы волн давления и ско-
рости крови в крупных артериях и венах малого круга кровооб-
ращения, а также условия распространения, затухания и отраже-
ния волн давления в этих сосудах с учетом уже обсуждавшихся
их упругих свойств. В общих чертах мы познакомимся также с
тем немногим, что известно о характере течения в них крови и о
профилях скорости.
Форма волн. Типичные формы волн давления и расхода, реги-
стрируемые в легочном стволе собаки, приведены на рис. 15.14.
Здесь же показаны формы волн расхода в крупной легочной ве-
не и давления крови в левом предсердии. Среднее по времени зна-
чение давления в легочной артерии равно примерно 2,0-103 Н-м~2
(15 мм рт. ст.), а амплитуда пульсовых колебаний этого давления
составляет примерно 1,0-103 Н-м-2, т. е. 50% средней величины,
что значительно превышает относительную величину пульсовых
колебаний давления в артериях большого круга (разд. 12.2). Фор-
ма этих волн давления не отличается существенно от формы пуль-
совых волн давления в аорте. Но в несколько менее крупных арте-
риях большого и малого круга формы волн давления различаются:
в первом случае эта форма существенно изменяется с расстоянием,
тогда как во втором явных изменений не происходит. Это могло бы
быть обусловлено тем, что из-за очень малой длины легочных ар-
терий пульсовая волна возникает во всех этих артериях почти
одновременно. Однако такое предположение приводит к значи-
тельным ошибкам при расчете зависимости входного импеданса
артерий малого круга от частоты (см. гл. 12 и ниже в данном раз-
деле) .
Относительно происхождения волн давления и расхода в венах
малого круга (рис. 15.14) имеются разногласия. Вопрос заклю-
588 Часть 11. Механика кровообращения
Рис. 15.14. Записи давления (Р) и расхода крови (Q) в сосудах малого круга
бодрствующей собаки в состоянии покоя. Обозначения: ЛА — легочная артерия;
ЛВ — легочная вена; ЛП — левое предсердие; ЭКГ — электрокардиограмма в
первом отведении Рлв и Q/Jg зарегистрированы на расстоянии в пределах 2 см
от левого предсердия. Острые обращенные вниз пики на записях Слв и пики
меньшей величины на записях ФЛА приходящиеся на фазу диастолы, — ЭКГ-сиг-
налы. Вертикальные линии — отметки времени 0,02 с. [Milnor (1972). Pulmonary
haemodynamics. In: Cardiovascular fluid dynamics (ed. Bergel), vol. 2, pp. 299—
340. Academic Press, New York.]
чается в том, порождаются ли они главным образом явлениями,
происходящими в правом желудочке, и достигают венозного русла,
распространяясь через систему микрососудов малого круга, или
обусловлены ретроградным влиянием левого предсердия и желу-
дочка. Данные, полученные при искусственной перфузии легких
непульсирующим потоком крови, но при сохранении нормального
соединения их вен с левым предсердием, подтверждают представ-
ление о том, что в естественных условиях источником волн дав-
ления в этих венах являются левые камеры сердца: в легочных
венах в этих условиях наблюдаются волны давления, совпадающие
по форме с обычными волнами давления в левом предсердии. Но
когда легочные вены отделяли от левого предсердия и легкие пер-
фузировали через артерии пульсирующим потоком, изливающийся
из этих вен поток также оказывался пульсирующим. Кроме того,
если легочную вену пережать и регистрировать в ней давление
выше области пережатия, то in vivo можно наблюдать колебания
давления в этой вене, напоминающие колебания давления в легоч-
Гл. 15. Малый круг кровообращения 589
ной артерии, но несколько запаздывающие во времени. В таком
случае среднее по времени давление в вене, естественно, повы-
шается до уровня, характерного для давления в легочных арте-
риях, и, следовательно, растяжимость вены значительно умень-
шается. Эти наблюдения позволяют думать, что, хотя поток крови,
поступающий из системы микрососудов легких в их вены, сохра-
няет пульсовые колебания, он практически не вызывает колебаний
давления в венах, поскольку их растяжимость при низких значе-
ниях трансмурального давления (которые физиологически нор-
мальны) велика. Прижизненные микроскопические наблюдения за
микрососудами поверхности легких, проводимые сквозь тонкий ли-
сток висцеральной плевры, показали, что кровоток в этих сосу-
дах, включая прекапиллярные сосуды и альвеолярные капилляры,
является пульсирующим. Изучение процесса перехода в кровь че-
рез легкие закиси азота — газа, растворимость которого в крови
очень высока, — подтверждает эти данные. На рис. 15.15 схемати-
чески показано изменение среднего и пульсового давления, а так-
же среднего и пульсового расхода крови в сосудистом русле ма-
лого круга. К рассмотрению пульсирующего течения крови в ка-
пиллярах малого круга (рис. 15.17) мы обратимся позднее.
Средняя скорость движения крови в легочном стволе прибли-
зительно такая же, как в аорте, так как диаметр обоих сосудов
примерно одинаков, а все количество крови, выбрасываемое серд-
цем, проходит через каждый из них. Однако пиковое значение
скорости движения крови в легочном стволе примерно вдвое мень-
ше, чем в аорте (приблизительно 0,7 м-с-1 вместо 1,2 м-с-1).
Распространение волн. Скорость распространения пульсовой
волны в артериях малого круга измеряли у представителей раз-
ных видов, в том числе и у человека, но данные для каждого от-
дельного вида весьма скудны. Для человека типичное значение
составляет примерно 1,75 м-с-1, а для собаки (табл. I)—2,5 м-с-1.
Как уже отмечено (разд. 15.2), полученные для собаки резуль-
таты довольно хорошо соответствуют величинам, предсказывае-
мым теоретически на основании измерений растяжимости легочной
артерии в статических условиях. Измерения скорости распростра-
нения пульсовой волны у больных с легочной гипертонией, для
которой характерно значительное увеличение толщины и жестко-
сти стенки сосудов малого круга, дали значение около 4,8 м-с-1.
При распространении по сосудистому руслу малого круга вол-
ны, несомненно, затухают, особенно на уровне микрососудов (см.
рис. 15.15). Однако количественных данных о скорости затухания
в конкретных сосудах нет, поскольку для сосудов малого круга
не проводили опытов с созданием коротких серий высокочастотных
колебаний, как это было сделано для артерий и вен большого
круга (разд. 12.5 и 14.3). Создавали, правда, волны более низкой
частоты, но измерения коэффициента затухания таких волн на
протяжении артерий нескольких порядков ветвления не позволили
590 Чат II. Механика кровообращения
Рис. 15.15. Среднее и пульсовое давление (10 мм рт. ст. = 1,36-103 Н-м-2) и
расход крови на усредненном пути через сосудистое русло малого круга крово-
обращения человека. Обратите внимание на изменение масштаба длины в точ-
ках, соответствующих 0,5 см. [Milnor (1972). Pulmonary haemodynamics. In: Car-
diovascular fluid dynamics (ed. Bergel), vol. 2, pp. 299—340, Academic Press, New
York.]
различить влияний, обусловленных отражением волн и вязкими
свойствами стенок артерий или вязкостью крови.
Хотя просмотр записей волн давления и расхода не позволяет
выявить каких-либо существенных изменений их формы при рас-
пространении по артериям малого круга, анализ Фурье (разд. 8.6)
все же свидетельствует об изменении состава гармоник этих волн.
Как и в случае артерий большого круга, это означает, что либо
в системе имеются отраженные волны, либо она существенно
нелинейна. Есть основания полагать, что нелинейные эффекты не-
велики: хотя скорость распространения волн по легочному стволу
меньше, чем по аорте, отношение пиковой скорости к скорости рас-
пространения волны здесь не больше, чем в аорте. Вместе с тем
следуёт ожидать, что отражения должны возникать на любом
плохо согласованном ветвлении сосудов (т. е. на таких ветвле-
ниях, для которых при сохранении постоянным отношения h/d
отношение площадей не близко к 1; см. разд. 12.4). Как и в слу-
чае артерий большого круГа, fro$§$HbiM й Ьрддающймся Измерению
Гл. 15. Малый круг кровообращения 59 1
показателем наличия отражений, который к тому же указывает и
на область их возникновения, служит входной импеданс сосуди-
стого русла малого круга. Он определяется для каждой частот-
ной составляющей как отношение измеренных в одном и том же
месте легочного ствола значений давления и расхода крови (точ-
нее, их комплексных значений; более полное определение входного
импеданса, его модуля и фазы дано в разд. 12.10).
На рис. 15.16 приведены значения входного импеданса арте-
рий малого круга, вычисленные на основании измерений, которые
производили на наркотизированных собаках со вскрытой грудной
клеткой; вверху показан модуль импеданса (в зависимости от час-
тоты), внизу — фаза. Значения модуля снижаются от высоких при
низкой частоте до минимума при частоте 3—4 Гц, когда фаза им-
педанса становится равной нулю. Для этой частоты легочный ствол
является областью узла (т. е. областью, в которой амплитуда волн
давления минимальна, а амплитуда волн расхода максимальна).
Такой результат предполагает, что на расстоянии четверти длины
волны от легочного ствола имеет место частичное отражение типа
отражения от закрытого конца. Если скорость распространения вол-
ны равна 2,5 м-с-1, то при частоте 4 Гц четверть длины волны со-
ставляет примерно 16 см. Для крупных собак это примерно равно
средней длине пути до микрососудов. Мысль о том, что здесь должна
быть область такого отражения, вполне приемлема. Наличие вто-
рого максимума модуля импеданса при частоте, примерно вдвое
превышающей ту, при которой модуль импеданса минимален, под-
тверждает справедливость представления о расположении обла-
стей отражения на уровне микрососудов: легочный ствол оказы-
вается областью пучности для волн, частота которых вдвое выше
(а длина вдвое меньше), чем тех волн, для которых легочный
ствол является областью узла.
Различие вида кривых входного импеданса артериальной си-
стемы малого круга кровообращения (рис. 15.16) и большого
круга (рис. 12.29) объясняется, по всей вероятности, относитель-
ной симметричностью артериального дерева малого круга и не-
симметричностью артериального дерева большого круга: в послед-
нем длина пути к передней части тела собаки меньше, чем к зад-
ней (см. рис. 12.31).
Величину входного импеданса малого круга кровообращения
кролика определяли расчетным путем, предполагая, что все арте-
рии малого круга одновременно растягиваются пульсовой волной
и одновременно же спадаются [как в случае модели упругого ре-
зервуара (разд. 12.2)]. Это предположение может показаться при-
емлемым, так как в малом круге длина артериального пути до
микрососудов значительно меньше длины пульсовой волны (при-
мерно в 4 раза при обычной частоте сокращений сердца). ОднЙко
пбйученный с помощью таких расчетов модуль импеданса ока-
зался для всех частот ниже измеренного, причем минимальное его
592 Часть II. Механика кровообращения
Гл. 15. Малый круг кровообращения 593
значение, близкое к нулю, пришлось на резонансную частоту около
4 Гц. По-видимому, это является результатом пренебрежения мно-
жественными отражениями, возникающими на пути между легоч-
ным стволом и системой микрососудов.
Значение модуля входного импеданса на нулевой частоте —
это мера сопротивления, которое оказывает сосудистое русло ма-
лого круга стационарному (не пульсирующему) потоку крови. Эта
величина очень сильно зависит от степени наполнения легких воз-
духом, а механизм, который определяет зависимость, весьма сло-
жен. Поэтому мы отложим его рассмотрение до следующего раз-
дела, в котором приведем также краткий анализ сопротивления
альвеолярных капилляров.
Картины течения крови. Подробности течения крови в легочных
артериях почти не изучены. Исследования последнего времени
позволяют полагать, что в легочном стволе профиль скорости от-
носительно плоский; для участка, расположенного столь близко
к выходу из правого желудочка, этого и следовало ожидать. Дей-
ствительно, можно полагать, что во всех крупных артериях (и ве-
нах) малого круга входной профиль довольно плоский, потому
что число Рейнольдса здесь велико, значение параметра а боль-
шое, а сосуды короткие, и, следовательно, на каждом очередном
делителе потока образуются новые тонкие пограничные слои, ко-
торые никогда не заполняют просвет сосудов. Кривизна сосудистой
стенки вблизи бифуркаций, по-видимому, достаточно мала, чтобы
предотвратить отрыв потока. Кроме того, можно ожидать, что, как
и в артериях большого круга, вблизи бифуркаций изменяются на-
пряжения сдвига на стенке. Однако для подробного изучения кар-
тин течения крови в ветвях сосудов малого круга имеющиеся в
настоящее время датчики скорости крови слишком грубы: тре-
буются более миниатюрные датчики. Для легочного ствола собаки
среднее число Рейнольдса равно примерно 600, а в пике волны
расхода — около 3000; значение параметра а (при частоте сокра-
щений сердца 2 Гц) близко к 15. Если для предсказания харак-
тера течения в легочном стволе (турбулентное оно здесь или ла-
минарное) использовать тот же критерий, что и для течения в
аорте (см. рис. 12.50), то при указанных значениях числа Рей-
нольдса и параметра а результат такого предсказания попадает
как раз на границу между ламинарным и турбулентным тече-
Рис. 15.16. Входной импеданс артерий малого круга наркотизированной собаки
со вскрытой грудной клеткой. Точки — средние значения для 29 животных, вер-
тикальные отрезки — средние ошибки средних величин. Модуль импеданса сни-
жается до минимума при частоте 3—4 Гц; примерно при такой же частоте фаза
его пересекает нулевую линию. При частоте около 6 Гц модуль импеданса имеет
второй максимум. [Milnor, Bergel, Bargainer (1966). Hydraulic power associated
with pulmonary blood flow and its relation to heart rate. Circulation Res., 19,
467—480.]
сл
О
Часть 11. Механика кровообращения
Гл. 15. Малый круг кровообращения 595
нием. Никаких данных, указывающих на наличие турбулентности
в легочном стволе, не измененном патологическим процессом, пока
не получено.
15.4. Сопротивление сосудов малого круга
Течение крови в капиллярном канале альвеолярных перегоро-
док. Между движением крови в капиллярах альвеол и в капиллярах
большого круга имеются два важных различия. Первое связано
с особым расположением капилляров в альвеолах: они образуют
здесь узкий канал с податливыми стенками, которые удержи-
ваются на определенном расстоянии друг от друга распорками
(рис. 15.4); вторая особенность — наличие хорошо заметных пуль-
совых колебаний движения крови в альвеолярных капиллярах
(рис. 15.17). Однако в двух основных отношениях движение крови
в капиллярах того и другого типа одинаково: число Рейнольдса
в обоих случаях мало (при обычной скорости движения крови в
капиллярах альвеол 0,005 м-с-1 и характерной толщине слоя кро-
ви в альвеолярной перегородке при нулевом трансмуральном дав-
лении (ho = 4 мкм) число Рейнольдса равно примерно 0,005), и
значение параметра а, даже для наиболее высоких еще существен-
ных гармоник пульсовой волны, также мало (для частоты 10 Гц
а = 0,0002). Это означает, что, во-первых, инерционные свойства
жидкости оказывают на течение крови в капиллярах пренебре-
Рис. 15.17. Записи, иллюстрирующие пульсирующий характер кровотока в ка-
пиллярах малого круга. А и Б. Записи давления в герметически закрытой жест-
кой камере (метод общей плетизмографии тела), в которой сидит испытуемый;
записи сделаны в тот период, когда испытуемый задержал дыхание при объеме
легких, соответствующем состоянию спокойного выдоха; голосовая щель у испы-
туемого открыта. На этих фрагментах приведены также записи ЭКГ. До за-
держки дыхания испытуемый дышал воздухом (Д) или смесью кислорода с
очень хорошо растворимой в крови закисью азота N2O (Б). А. Происходит не-
большое равномерное повышение давления из-за нагревания воздуха (смещение
линии вниз соответствует увеличению давления; изменения давления преобразо-
ваны в изменения объема при калибровке); заметны небольшие колебания дав-
ления, обусловленные такими факторами, как сжатие газа при сокращениях
сердца. Б. Давление в плетизмографе падает, процесс носит колебательный ха-
рактер; период колебаний совпадает с периодом сокращений сердца. (Стрелками
отмечены моменты времени, в которые путем прокачивания через плетизмограф
соответствующей газовой смеси давление в нем восстанавливали до исходного,
равного атмосферному.) Для определения мгновенных значений расхода крови
в капиллярах легких рассчитывали значения мгновенной скорости поглощения
закиси азота, вычитая по точкам контрольную кривую А из кривой 5; затем
пересчитывали эти значения в мгновенные значения расхода крови, вводя в ка-
честве масштабного коэффициента значение растворимости закиси азота в крови
при измеренном значении парциального давления этого газа в альвеолах.
В. Представлены полученные таким образом значения мгновенного расхода крови
в капиллярах легких у здорового испытуемого в состоянии покоя (/) и после вы-
полнения мышечной работы (//). Штриховые линии — средние значения расхода
крови; внизу — ЭКГ. [Lee, Dubois (1955). Pulmonary capillary blood flow in man.
J. clin. Invest., 34, 1380—1390.]
696 Часть II. Механика кровообращения
жимо малое влияние и для любой точки течения градиент давле-
ния уравновешен вязкими силами; во-вторых, течение квазиста-
ционарно в том смысле, что в любой момент времени во всех час-
тях слоя крови в альвеолярной перегородке волны расхода крови
и градиента давления совпадают по фазе. Для ньютоновской жид-
кости в трубке или плоском канале с жесткими стенками при та-
ких условиях расход крови через систему всегда был бы прямо
пропорционален перепаду давления на ней. В системах с упру-
гими стенками это не так, потому что ширина канала, которая
определяет его сопротивление потоку, зависит в этом случае от
локального давления, и, следовательно, сопротивление уже нельзя
считать не зависящим от градиента давления, вызывающего тече-
ние. Как и в микрососудах большого круга, на сопротивление ка-
пилляров альвеол влияет также наличие взвешенных в плазме
форменных элементов крови.
Разработана приближенная теория квазистапионарного тече-
ния крови в капиллярах альвеол, основанная на результатах не-
которых опытов на моделях и на следующих упрощающих допу-
щениях.
А. Как в теории, так и при исследованиях на моделях кровь
рассматривается как однородная ньютоновская жидкость, вязкость
которой примерно в четыре раза превышает вязкость воды; при-
сутствие в ней эритроцитов, образующих концентрированную сус-
пензию, не принимается во внимание явно. [Для оценки влияния
эритроцитов можно было бы провести анализ с использованием
теории смазки, как это было сделано для капилляров большого
круга (разд. 13.7), однако такой анализ пока не производился.]
Б. Наличие распорок на начальном этапе не учитывается.
В. Предполагается, что толщина слоя крови в альвеолярной
перегородке h всюду линейно зависит от величины трансмураль-
ного давления (рвн. с. — Ральв) •
Й Й0 V (Рвн. с. Ральв)> (15.2)
где у и йо — постоянные. Справедливость этого уравнения под-
тверждена измерениями толщины слоя крови в альвеолярных пе-
регородках собаки и кошки; распорки в изучавшихся гистологи-
ческих препаратах, естественно, имелись. Измерения на препара-
тах легких собаки показали, что уравнение (15.2) справедливо
в диапазоне трансмуральных давлений от нуля (или чуть выше)
приблизительно до 2,5-103 Н-м-2 (25 см вод. ст.). Кроме того,
было показано, что «податливость» у зависит от величины транс-
пульмонального давления; это связано с тем, что изменение транс-
пульмонального давления приводит к изменению степени растя-
жения мембраны альвеол. При транспульмональном давлении
103 Н-м~2 (10 см вод. ст.) значение у составляет примерно 1,2-
•10-3 мкм на 1 Н-м-2, тогда как при транспульмональном давле-
нии 2,5-103 Н-м-2 (25 см вод. ст.) оно равно примерно0,8-lO^3 мкм
Гл. 15. Малый круг кровообращения 597
на 1 Н-м~2 (см. также рис. 15.8). Когда давление в альвеолах
превышает давление в сосудах, уравнение (15.2) перестает вы-
полняться, потому что в этом случае капилляры альвеол быстро
спадаются и толщина слоя крови в канале альвеолярной перего-
родки падает от значения ho почти до нуля.
Теоретический анализ течения крови в альвеолярном канале
проводили исходя из уравнения (15.2) и двух основных принци-
пов, определяющих течение жидкости в узкой щели. Согласно этим
принципам, в случае квазистационарного течения несжимаемой
жидкости градиент давления всюду уравновешивается вязкими
силами, и в любой данный момент времени расход жидкости через
Рис. 15.18. Схема, поясняющая обозначения, используемые при анализе течения
жидкости в двумерном слое.
все сечения щели должен быть одинаков, так как жидкость ни-
куда не исчезает и ниоткуда не добавляется. Это точно те же
принципы, которые использовались при рассмотрении движения
эритроцитов в капиллярах большого круга при помощи теории
смазки (разд. 13.7). Результаты лучше всего проиллюстрировать
на примере течения жидкости в двумерном слое (рис. 15.18). Дву-
мерность слоя подразумевает, что никаких изменений толщины его
или скорости движения жидкости в направлениии, перпендикуляр-
ном плоскости рисунка, не происходит. Предполагается, что дав-
ление в слое жидкости, т. е. внутри такого «сосуда» (рвн. с_), опре-
деляется давлением на его артериальном и венозном концах — со-
ответственно ра и рв. Давление вне «сосуда» — это альвеолярное
давление раЛьв. Толщина слоя h меняется по его длине х и на ар-
териальном конце равна ha. Основные результаты анализа таковы.
1. Если давление в альвеолах меньше давления в венах малого
круга (ральв < рв < ра), то канал, в котором заключен рассмат-
риваемый сЛой жидкости, остается открытым, а изменение h в за-
висимости от х описывается уравнением
h4 = /it-₽x, (15.3)
где 0 — коэффициент, который зависит от величин ра и рв, а также
от длины слоя I в направлении движения жидкости. Уравнения
698 Часть II. Механика кровообращения
(15.2) и (15.3) означают, что выражение, в которое величина
рВн с. входит в четвертой степени, с изменением х меняется ли-
нейно. Такой результат совершенно отличен от формулы для тече-
ния жидкости в прямой жесткой трубке или в узком канале, ог-
раниченном двумя жесткими плоскостями, когда с изменением
расстояния х линейно меняется именно само давление рвн. с.- В рас-
сматриваемом случае расход жидости Q прямо пропорционален
величине р/, и из уравнения (15.3) следует:
Q = (15.4)
где hB — толщина слоя жидкости на венозном конце, а С — по-
стоянная, которая не зависит ни от толщины слоя, ни от давления.
2. Если канал, в котором заключен рассматриваемый слой
жидкости, остается повсюду открытым, то сопротивление движе-
нию жидкости через него, равное перепаду давления, деленному
на расход крови, и обозначаемое символом /?, определяется соот-
ношением
1 йа - лв
P=CV
(15.5)
ha — ha
где у и С — те же постоянные, что и в уравнениях (15.2) и (15.4).
Этот результат несомненно изменится, если будет учтено наличие
распорок. Один из способов, позволяющих оценить их влияние,
заключается в том, чтобы найти обусловленный наличием таких
распорок коэффициент увеличения сопротивления для случая дви-
жения слоя жидкости между жесткими плоскостями, а затем ис-
пользовать тот же самый коэффициент для случая податливых
плоскостей. Согласно результатам опытов на моделях и теорети-
ческого анализа, значение этого коэффициента примерно равно 3.
При отсутствии данных, которые могли бы быть получены в ис-
следованиях течения крови в реальных условиях, т. е. в канале
альвеолярной перегородки или в более точной его модели с по-
датливыми стенками, при определении сопротивления из уравне-
ния (15.5) следует применять именно этот поправочный коэффи-
циент.
3. Определяемое уравнением (15.3) изменение толщины слоя
жидкости h с расстоянием х оказывается наиболее быстрым на
венозном конце этого слоя. Это обусловлено тем, что скорость та-
кого изменения, d/i/dx, равна —р/4/i3, т. е. тем больше, чем
меньше h, и, значит, она максимальна при наибольших х. Изме-
нение h на венозном конце происходит особенно быстро, если дав-
ление в альвеолах лишь чуть-чуть меньше венозного давления р3,
так как в этом случае величина h3 близка к ho. При очень неболь-
шом дополнительном увеличении давления в альвеолах венозный
конец канала полностью спадается и расход жидкости Q стано-
вится равным Cha, т. е. не зависит больше от условий на венозном
Гл. 15 Малый круг кровообращения 599
конце. По мере дальнейшего повышения давления в альвеолах
толщина открытой части канала уменьшается и расход жидкости
падает приблизительно пропорционально (ра— ральв)4, что сле-
дует из уравнения (15.2). Наконец, если давление в альвеолах
превысит артериальное давление, то течение полностью прекра-
тится. Этот процесс очень близок к процессу, рассмотренному
нами при изучении явления спадения вен (разд. 14.2), но в дан-
ном случае инерционные свойства жидкости не играют какой-либо
роли в механизме спадения, и потому самовозбуждающиеся ко-
лебания не возникают.
Измерить толщину слоя крови в альвеолярной перегородке
in vivo, когда кровь движется, фактически невозможно, и поэтому
многие тонкие вопросы этой теории нельзя проверить непосред-
ственно. Тем не менее можно проверить (по крайней мере каче-
ственно) следствия теории относительно сопротивления, а кроме
того, оценить величину объема крови в капиллярах. Последнее
важно вот почему: вывод теории о наибольшей скорости измене-
ния толщины слоя жидкости на венозном конце [вытекающий из
уравнения (15.3)] приводит к заключению, что объем содержа-
щейся в капиллярах крови пропорционален ра— Ральв и факти-
чески не зависит от венозного давления. Как уже отмечалось
(разд. 15.2), это наблюдается в опытах.
Из приведенного выше анализа следует, что в зависимости от
соотношения величин ра, рв и ральв состояние капилляров в аль-
веолярных перегородках и закономерности движения крови в них
принимают одну из трех возможных форм.
Зональное распределение кровотока. С некоторых пор стали
признавать, что распределение кровотока в легких зависит от
относительных уровней артериального (ра), венозного (рв) и
альвеолярного (ральВ) давлений, и позднее мы приведем некото-
рые свидетельства в пользу такого представления. Как мы уже
знаем (см. гл. 14), течение жидкости по сосудам, способным спа-
даться (например, по венам), сильно зависит от местного транс-
мурального давления. В случае капилляров малого круга, как и
в случае вен, можно различать три режима или три зоны.
I. Если давление в альвеолах выше и венозного, и артериаль-
ного давления, то сосудистое русло оказывается закрытым и тече-
ния крови не происходит.
II. Если давление в альвеолах ниже артериального, но выше
венозного, сосудистое русло оказывается спавшимся частично и
расход крови в нем зависит от разности давлений ря — ральв.
III. Если давление в альвеолах ниже и артериального, и веноз-
ного (а давление в артериях, естественно, выше, чем в венах), то
все альвеолярные капилляры полностью открыты и расход крови
зависит от разности давлений ра — рв.
У здорового человека, находящегося в вертикальном положе-
нии в состоянии покоя, могут существовать все три режима, по-
600 Часть II. Механика кровообращения
тому что, как уже объяснялось, давление в сосудах легких имеет
вертикальный гидростатический градиент, тогда как давление в
альвеолах примерно одинаково и близко к атмосферному. Поэтому
у верхушки легкого давление крови относительно низкое и выпол-
няются условия первого режима; эту область легких называют
зоной I. Для средней части легкого выполняются условия второго
режима и, соответственно, эту часть называют зоной II. Наконец,
в нижней части легкого, в зоне III, венозное давление превышает
давление в альвеолах1).
Основание Верхушка
I I '« I
10 20
в ^альв
Основание ^Верхушка
______J___•«—!___
10 20
Расстояние, см Расстонние. см
“ ₽апьв
'Х’***Ц, ₽в=₽апьв
Основание Верхушка
_______I______I__л
10 20
Расстояние, см
В
Рис. 15.19. Распределение расхода крови в изолированных перфузируемых легких
собаки в опыте с использованием изотопа ксенона [,33Хе] (Объяснения см. в
тексте.) [West, Dollery, Naimark (1963). Distribution of blood flow in isolated
lung: relation to vascular and alveolar pressures J. appl. Physiol., 19 713—724.]
Очевидно, что положение уровней перехода одной зоны в дру-
гую изменяется в зависимости от давления в альвеолах. При на-
полнении легких воздухом под положительным давлением боль-
шая часть легких относится к зоне I, тогда как при быстром про-
извольном вдохе давление в альвеолах оказывается ниже атмо-
сферного и зона II расширяется. При повышении давления в ар-
териях малого круга, например при выполнении мышечной работы
или при легочной гипертонии, следует ожидать еще больших изме-
нений: в этих условиях зона II охватывает все легкие.
Существование указанных зон подтверждено рядом способов,
например путем сопоставления числа эритроцитов в альвеолярных
капиллярах на гистологических препаратах легких собаки, полу-
ченных методом быстрого замораживания во время перфузии при
разных уровнях давления в сосудах и в альвеолах (рис. 15.8).
Подтверждения получены также в опытах на изолированных пер-
фузируемых легких и в исследованиях на здоровых испытуемых.
На рис. 15.19 показано влияние артериального, альвеолярного
и венозного давления на распределение по вертикали кровотока
в изолированных перфузируемых легких собаки. В этих опытах
использовали изотоп ксенона (133Хе), который вводили с физиоло-
’) Опыты показывают, что на самом деле следует выделить еще одну, чет-
вертую область — зону IV, которая находится вблизи основания легкого Ско-
рость движения крови в этой области очень сильно зависит от объема легких;
ниже мы подробнее рассмотрим этот вопрос.
Гл. 15. Малый круг кровообращения 601
гическим раствором в легочную артерию. Когда раствор попадает
в капилляры легких, из-за большой разности парциальных дав-
лений ксенона в крови и альвеолярном воздухе, а также из-за
низкой растворимости в крови большая часть ксенона переходит
из крови в альвеолярный воздух. Как только это произойдет, вен-
тиляцию останавливают и производят сканирование легких с по-
мощью счетчика. При соответствующей калибровке уровень ра-
диоактивности каждого участка легких отражает величину мест-
ного кровотока.
На рис, 15.19, Л показано распределение кровотока в условиях,
когда давление в альвеолах равно атмосферному, а наполнение
легкого осуществляется при отрицательном давлении 1-Ю3 Н-м-2
(10 см вод. ст.). У основания легкого артериальное давление со-
ставляет 3,2-103 Н-м-2 (32 см вод. ст.), вертикальный размер лег-
кого 26 см, и нижняя граница располагается несколько выше ре-
зервуара, в который оттекает кровь из вен, так что венозное дав-
ление на всем протяжении легкого ниже атмосферного. Кровоток
в верхней части легкого при этом близок к нулю, а при переме-
щении области измерения сверху вниз он увеличивается прибли-
зительно линейно. На рис. 15.19, Б показано влияние на распре-
деление кровотока снижения давления в легочной артерии до
1,6-103 Н-м'2 (16 см вод. ст.). В этих условиях на высоте 16 см
от основания легкого артериальное давление равно давлению в
альвеолах, и снижение кровотока до нуля происходит примерно
на этом уровне. В случае, представленном на рис. 15.19, В, давле-
ние в артериях нижней части легкого составляет 2,55-103 Н-м-3
(25,5 см вод. ст.), а венозный резервуар поднят на 19 см выше
уровня основания. В тех областях легкого, где венозное давление
превышает давление в альвеолах, кровоток увеличивается в на-
правлении сверху вниз постепенно, тогда как в верхушке легкого,
где давление в альвеолах выше венозного, изменение кровотока по
вертикали оказывается значительно более крутым. Из всех этих
данных следует, что: 1) в зоне I кровоток отсутствует
(рис. 15.19, Б); 2) в зоне II кровоток круто возрастает сверху вниз
(рис.15.19, Б и Б) 3) в зоне III увеличение кровотока в направле-
нии сверху вниз невелико (рис. 15.19,Б). Такие же результаты
дали исследования на человеке.
Чтобы понять, почему расход крови в зонах II и III увеличи-
вается по мере удаления от верхушки легкого и почему скорость
такого изменения кровотока в этих двух зонах различна, следует
выяснить, какая разность давлений определяет в каждом случае
расход крови и какие факторы определяют сопротивление сосудов
легкого в той и другой зоне.
В зоне II расход крови определяется разностью давлений
Ра — Ральв, и, ПОСКОЛЬКУ Значение Ральв по всей высоте легкого в
сущности одинаково, тогда как значение ра увеличивается от вер-
хушки легкого к основанию примерно на 1 см вод. ст. (0,1-103 Ц*
602 Часть 11. Механика кровообращения
• м-2) на 1 см, разность давлений, вызывающая движение крови,
возрастает на такую же величину (см. сноску на этой странице). Со-
противление сосудов легкого в этой зоне определяется давлением
в артериях и в капиллярах; давление в венах здесь не играет су-
щественной роли. Так как артериальное давление от верхушки
легкого к основанию возрастает, можно ожидать постепенного рас-
тяжения артерий, соответствующего местному трансмуральному
давлению и упругим свойствам сосудов. Таким образом, сопротив-
ление движению крови в артериальных сосудах будет умень-
шаться от верхушки легкого к основанию, хотя точно предсказать
соотношение между диаметром сосудов и их сопротивлением в
этом случае невозможно из-за сложного характера течения. Транс-
муральное давление в альвеолярных капиллярах по мере удале-
ния от верхушки также возрастает; в результате увеличивается
число открытых капилляров и растягиваются уже функционирую-
щие капилляры, что приводит к снижению сопротивления капил-
лярного русла. Итак, все изменения, происходящие с сосудами при
увеличении давления, определяющего расход крови, способствуют
возрастанию расхода в направлении от верхушки легкого к его ос-
нованию.
В зоне III расход крови определяется разностью давлений
Ра — Рв, и, поскольку изменение обеих этих величин по высоте лег-
ких одинаково (они увеличиваются сверху вниз примерно на 1 см
вод. ст. на 1 см), эта разность оказывается независящей от вы-
соты1). Сопротивление сосудистого русла в этой зоне опреде-
ляется сопротивлением артерий, капилляров и вен. Все замеча-
ния, касающиеся размеров и сопротивления артерий в зоне II,
справедливы также для артерий и вен в зоне III. Поэтому можно
предполагать, что сопротивление движению крови через эти со-
суды и через капилляры по мере удаления от верхушки будет
падать. Для капилляров преобладающим изменением, как пола-
гают, является их растяжение, хотя пока еще невозможно экспе-
риментальными методами отличить влияние их растяжения от
влияния раскрытия ранее не функционировавших капилляров.
Эти различия механических явлений в зонах II и III дают од-
нозначное объяснение более быстрого увеличения расхода крови
по мере удаления от верхушки легких в зоне II. В следующем раз-
деле мы рассмотрим влияние объема легких на относительные
сопротивления экстра-альвеолярных и альвеолярных сосудов.
Влияние свойств легких. Раньше (разд. 15.2) мы уже выяс-
нили, каким образом плевральное давление и объем легких
влияют на экстра-альвеолярные сосуды, а давление в альвеолах и
*) Это не совсем верно, поскольку распределение давления вдоль сосудов
легких зависит от сопротивления как артериальных, так и венозных сосудов,
а также и от сопротивления капилляров. Поэтому распределение давления и рас-
пределение расхода крови оказываются взаимосвязанными.
Гл. 15. Малый круг кровообращения 603
Расстояние см
Рнс. 15.20 Зависимость удельного кровотока (на альвеолу), выраженного в про-
центах от величины, которую следовало бы ожидать при равномерной перфузии
всех альвеол, от расстояния между рассматриваемым участком и верхушкой
легкого при различной степени наполнения легких воздухом: / — заполнение всей
жизненной емкости легких, II — функциональной остаточной емкости, /// — оста-
точного объема Точки соответствуют средним значениям для левого н правого
легкого всех испытуемых Обратите внимание иа уменьшение кровотока у осно-
вания легких, сильнее выраженное при малых объемах [Hughes, Glazier, Malo-
ney, West (1968) Effect of lung volume on the distribution of pulmonary blood
flow in man Respiration Physiol, 4, 58—72 ]
объем легких — на альвеолярные капилляры. Теперь обратимся к
влиянию механики легких на распределение кровотока и сопро-
тивления по сосудистому руслу малого круга.
Упомянутые выше исследования на стоящем человеке, под-
тверждающие существование трех зон, проводились при довольно
больших объемах легких. Однако когда для подобных же иссле-
дований применили несколько более совершенные методы и осу-
ществили эти исследования для всего физиологического диапазона
объемов легких, обнаружились ранее неизвестные особенности по-
ведения кровотока в малом круге (рис. 15.20). При заполнении
воздухом всей жизненной емкости легких у их основания выяв-
ляется небольшая область, в которой теперь кровоток падает в
вертикальном направлении. При уменьшении объема легких до
величины их функциональной остаточной емкости эта область
увеличивается, а при еще меньшем объеме легких, равном их
остаточному объему, кровоток в области верхушки оказывается
больше, чем у основания, т. е. распределение кровотока оказы-
вается обратным тому, которое возникает при заполнении возду-
хом всей жизненной емкости легких. Объяснить это исходя из про-
664 Часть it. Механика кровообращений
стой модели, описанной выше, невозможно. Полагают, что такое
изменение распределения кровотока в легких связано с влиянием
их веса, которое затрудняет наполнение воздухом участков у ос-
нования, а также с повышенным давлением в пространствах, ок-
ружающих экстра-альвеолярные сосуды этих участков. Это ведет
к уменьшению просвета таких сосудов, в результате чего возрас-
тают их сопротивления кровотоку. С таким объяснением согла-
суются данные о том, что объем альвеол в заполненных воздухом
изолированных легких на всем их протяжении относительно по-
стоянен.
Обратимся далее к результатам, полученным в опытах, в кото-
рых изолированное перфузируемое легкое наполняли воздухом,
снижая внешнее давление (ниже уровня атмосферного), а давле-
ние в альвеолах поддерживали равным атмосферному. Разность
между артериальным и венозным давлением, а также между арте-
риальным давлением и давлением в альвеолах в этих опытах
можно было устанавливать независимо от давления в камере и
от объема легкого. При разных значениях объема легкого изме-
ряли расход крови в его сосудах и вычисляли их сопротивление.
Как показано на рис. 15.21, при увеличении объема легкого сопро-
тивление его сосудистого русла сначала падало и достигало ми-
нимального значения, когда легкое было наполнено воздухом уме-
ренно (до 100—150 мл). При дальнейшем наполнении легкого
сопротивление его сосудистого русла возрастало до значения, при-
мерно вдвое превышающего минимальное.
Выше мы отметили, что наполнение легкого воздухом в таких
условиях приводит к растяжению экстра-альвеолярных сосудов
Полагают, что этим объясняется начальное снижение сопротивле-
ния сосудистого русла легких при увеличении их объема. Кроме
того, как уже говорилось, увеличение объема легких в таких усло-
виях, увеличивая натяжение стенок альвеол, приводит к сжатию
альвеолярных капилляров; этим предположительно объясняют по-
вышение сопротивления сосудистого русла легких при дальнейшем
увеличении их объема. Таким образом, подразумевается, что при
малых объемах легких основной вклад в сопротивление их сосу-
дистого русла вносят экстра-альвеолярные сосуды, а при больших
объемах — альвеолярные.
Возникает вопрос: приложимы ли результаты этих эксперимен-
тальных исследований к условиям in vivo. Несомненно, что in vivo
объем легких изменяется за счет изменения плеврального давления,
и, как и в опытах на изолированном перфузируемом легком, если
воздухоносные пути сообщаются с атмосферой, то давление в аль-
веолах равно атмосферному при любом не изменяющемся объеме
легких. Однако механика кровообращения в малом круге in vivo
иная, чем в условиях этих опытов: в естественных условиях как
правый желудочек, так и левое предсердие находятся внутри плев-
ральной полости и, следовательно, также подвергаются действию
Гл. 15. Малый круг кровообращения 605
Объем легкого, см3
Рнс. 15 21 Зависимость сопротивления сосудов от объема легкого. Все значения
определены в статических условиях Стрелками показано направление изменения
объема. [Thomas, Griffo, Roos (1961). Effect of negative-pressure inflation of the
lung on pulmonary vascular resistance. J. Appl. Physiol., 16, 451—456.]
внутриплеврального давления. В действительности из-за тесной
связи между сосудами малого круга и большого это различие
оказывается скорее кажущимся. Так, если кровь из сосудов боль-
шого круга перемещается к сердцу (скажем, при поднимании ног),
то давление в артериях и венах малого круга повышается и объем
крови, содержащейся в сосудистом русле малого круга, в том чис-
ле и объем крови в капиллярах малого круга, увеличивается
(разд. 15.2). Поэтому можно думать, что изменение объема лег-
ких in vivo будет приводить к изменению сопротивления их сосу-
дистого русла, подобно тому как это наблюдается в опытах на
изолированном перфузируемом легком. Некоторые эксперимен-
тальные данные подтверждают справедливость такого суждения,
но единого мнения по этому вопросу нет, возможно, потому, что
in vivo все оказывается гораздо сложнее, чем in vitro, и, напри-
мер, изменение позы способно вызывать и другие изменения, ска-
зывающиеся на производительности сердца.
606 Часть tt. Механика кровообращения
СПИСОК ЛИТЕРАТУРЫ
Banister J, Torrance R. W. (1960). The effects of tracheal pressure upon flow:
pressure relations m the vascular bed of isolated lungs, J. exptl Physiol., 45,
352—367
Bates D V, Macklem P. T., Christie R. V. (1971). Respiratory function in disease,
W. В Saunders, Philadelphia, London, Toronto
Benjamin J. J., Murtagh P. S., Proctor D. F., Menkes H. A., Permutt S (1974).
Pulmonary vascular interdependence m excised dog lobes, J. appl. Physiol., 37,
887—894.
Caro C. G. (ed). (1966). Mechanics of the pulmonary circulation In: Advances in
respiratory physiology, Edward Arnold, London
Cumming Q. (1974). The pulmonary circulation. In Cardiovascular physiology,
Physiology Series I, Vol. I, pp 93—122 (eds A. S Guyton and С. E. Jones),
MTP International Review of Science, Butterworths, London and University
Park Press, Baltimore.
Fishman A. P. (1963). Dynamics of the pulmonary circulation. In- Handbook of
physiology, Section 2: Circulation, Vol II (eds W. F. Hamilton and P. Dow),
American Physiological Society, Washington, D C
Fishman A. P., Hecht H. H. (eds). (1969). The pulmonary circulation and intersti-
tial space, University of Chicago Press, Chicago and London
Fung У. C., Sobin S. S. (1972). Pulmonary alveolar blood flow, Circulation Res,
30, 470—490.
Fung У. C., Sobin S. S. (1969). Theory of sheet flow in lung alveoli, J. appl. Phy-
siol., 26, 472—488.
Harris P., Heath D. (1962). The human pulmonary circulation: its form and func-
tion in health and disease, E. & S. Livingstone, Edinburgh and London.
Howell J B. L., Permutt S., Proctor D. F., Riley R. L. (1961). Effect of inflation
of the lung on different parts of pulmonary vascular bed. J. appl. Physiol.,
16, 71—76.
Permutt S., Howell J. B. L., Proctor D F., Riley R. L. (1961) Effect of lung in-
flation on static pressure-volume characteristics of pulmonary vessels, J. appl.
Physiol, 16, 64—70
Spencer H. (1968). Pathology of the Lung, Pergamon Press, Oxford
West J. В (1974). Blood flow to the lung and gas exchange, Anesthesiology, 41,
124—138.
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Аденозиндифосфат (АДФ) 199
Альбумин 179, 188
Альвеолы, капиллярный канал 566
----- давление 596—597
-----движение крови 595—599
------податливость 596—597
------- размеры 579
-------толщина слоя крови 596—
599
Амплитуда колебаний 135—136
Анализ размерностей 74, 103—109,
140, 142, 143. См. также Размер-
ностная однородность
— — метод показателей 107
Анастомозы 296, 405
— артерно-артериальные 423
— артерно-венозные 423, 513
-----в малом круге 570
— вено-венозные 423
Аорта, бифуркация, коэффициент от-
ражения 345
-----площадь поперечного сечения
345
— брюшная, напряжение сдвига на
стенке 409
-----профиль скорости 377—378
— ветвление, коэффициенты и углы
292—296
— восходящая 318
----- входной импеданс 354—355
-----давление 326 См также Дав-
ление в аорте
-------и расход, синфазность из-
менений 319—320
— — мгновенные профили скорости
373
-----напряжение сдвига на стейке
409
--параметр Уомерсли (а) 12—13
--профили скорости 374—376
--Рейнольдса число 12—13
--смешивание 400
-- ускорение крови 318—319
-----форма пульсовых волн давле-
ния 318, 325
-----расхода 318, 325
-----—-------скорости 319
«-грудная, диаметр 12—13, 304—305
Аорта грудная, турбулентность тече-
ния 392—393
— диаметр, изменения в течнне сер-
дечного цикла 317
— Длина начального участка 381—
382
— дуга, профили скорости 374—376
— изгиб 291
-----профиль скорости 382—383
— конусообразное сужение 291—292,
293
— линии тока 278
— нисходящая, профили скорости 376
— область входа 379—382
— отношение толщины стенки к диа-
метру 12—13, 526
— пограничный слой 371, 372—374,
376—377
— профили скорости 372—378
-----•— определяющие их физиче-
ские механизмы 379—389
— профиль средней скорости 372—
373
— пульсовые волны, гармонический
состав 323—324
— радиус 12—13, 293
— Рейнольдса число 12—13, 274
— собаки, анатомия 289—291
----- посмертные слепки 291, 292
— упругие свойства 12—13, 116—117,
118—122, 523, 524, 525
— человека, восходящая, входной им-
педанс 352—353
-----касательный модуль Юнга 307
-----турбулентность течения 393—
394
Аортальный клапан см. Клапан аор-
тальный
Артериальное давление 310
-----влияние силы тяжести 310—311
-----единицы измерения 309
— — способ измерения 309, 551
Артериальное дерево большого круга,
площадь поперечного сечения
12—13, 292—296
— — — — человека, формы пульсо-
вых волн давления и скорости
крови 345
608 Предметный указатель
Артериальное дерево малого круга
563—566
----------площадь поперечного се-
чения 565, 567
— русло большого круга, распреде-
ление давления 314
Артерии бедренные, напряжение сдви-
га на стенке 409
— большого круга 288—415
— ------ влияние гравитации 310
-------- давление кровн н кровоток
309—325
-------- и гидростатическое давление
313—314
---------- избыточное давление 314
--------коэффициенты ветвления 292
—-------напряжение сдвига на стен-
ке 409
-------- трансмуральное давление
309—313
--------углы ветвления 294, 296
— бронхиальные 570—571
— волны давления, влияние нелиней-
ностей на распространение 359—
361
--------затухание 364—367, 368
— ------отражение и прохождение
336—358
-------- скорость распространения
364—365, 368
------ сдвиговые 369
— картины течения крови 370—397
— крупные, параметр Уомерсли (а)
12—13, 330—331
— — профили скорости 370—379
— легочные 563
— легочный ствол 563
--------напряжение сдвига на стен-
ке 409
— малого круга, легочный ствол см.
Артерии, легочный ствол
—-------отношение толщины стенки
к диаметру 12—13, 523—524, 526
•-------отраженные волны 590—591
—-------пульсовые волны, скорость
12—13, 589
--------распространение волн 589—
593
--------Рейнольдса число 12—13,
273—274
-------русло, площадь поперечного
сечення 565, 567
•-------упругие свойства 12—13,
522—523, 524, 525
— образование тромбов 202
•— постстенотическое расширение
396—397
- распространение волн 326—336. См.
Также Артерии, волны давления
Артерии бедренные, распространение
давления 328—329 .
— смешивание и перенос веществ
397—412
— сонные, отношение толщины стен-
ки к диаметру 12—13, 526
---упругие свойства 523, 524, 525
— стенка 116
— — возрастные изменения 305—309
--- накопление липидов 406—407
--перенос веществ 404—412
--- строение 296—303
---толщина 12—13, 303—305
— турбулентность течения 389—397
— ударная волна 361
— упругие свойства 12—13, 118—
119, 301
— форма пульсовой волны расхода,
влияние вязкости крови 362—364
— человека, изменения упругих
свойств с возрастом 306—308
Артериография 294
Артериолы 419, 421, 423, 425, 426
— внутренняя оболочка 426
— гладкие мышцы 426, 428—429
— давление 446—447
— диаметр 12—13, 420, 421
— длина 420
— «критическое давление закрытия»
443
— линейная скорость движения крови
12—13, 420
— напряжение сдвига на стенке 409
— наружная оболочка 429
— параметр Уомерсли (а) 12, 13
— плотность 420
— Рейнольдса число 12—13
— система 419, 421
— соединительная ткань 426
— спазм 443
— средняя оболочка 428—429
— строение 426—429
— терминальные 419
— толщина стенки 12—13, 421
— упругие свойства 440—443
— эластическая ткань 426
— эндотелий 426
— Юнга модуль 442
Атероматозное поражение сосудов,
механизмы 405—412
Атероматозные бляшки 405—406
Атеросклероз 306
Атрно-вентрнкулярный пучок 220
Бернулли теорема 61—64, 83, 90, 93
— уравнение 63, 155, 549
Бифуркация симметричная, движение
жидкости 384—387
Предметный указатель 609
Блуждания эритроцитов радиальные
468—470
Бронхи, кровоснабжение 512, 570—
571
Броуновское движение 160, 185, 508,
509
Вальсальвы проба 559—560
- синус 276, 277, 278, 291, 515
Вант-Гоффа закон 172—173, 491,492,
502
Вектор, определение 23
— скорости 23, 61
— сложение 24
Вена воротная 513
— полая ннжняя, напряжение сдви-
га на стенке 409
— ------передняя, форма волн кро-
вотока 273
--- упругие свойства 524, 525
— яремная, отношение толщины стен-
ки к диаметру 526
--- упругие свойства 524, 525
Венозное русло, механика 553—560
--------влияние дыхания 557—560
----------сокращения скелетных
мышц 553—557
--------возвращение крови к сердцу
557—560
---— изменение положения отно-
сительно уровня сердца 553
Вентури трубка 84
Венулы 423, 425, 434, 436, 445—446,
514
— гладкие мышцы 426
— давление 446—447
— диаметр 12—13, 420
— длина 12—13, 420
— коэффициент фильтрации 487
— напряжение сдвига на стенке 409
— параметр Уомерсли (а) 12—13
— посткапиллярные 423, 445, 482
---строение стенки 514
— Рейнольдса число 12—13
— система 423, 425
— скорость крови 12—13, 420
— собирательные 423
— соединительная ткань 426
— строение 434—436
— упругие свойства 445—446
— число 420
— эластическая ткань 426
— эндотелий 426
Вены см. Вена, Венозное русло, Ве-
нулы
— большого круга 512—560
--------анатомическая организация
513—515
— бронхов 57|
Вены, внутренняя оболочка 515
— волны, распространение 534—540
— картины течения 540—543
— малого круга, анатомическая орга-
низация 570
— ------строение 570
--------форма пульсовых волн дав-
ления и кровотока 587—589
- наружная оболочка 514—515
— отношение толщины стенки к диа-
метру 12—13, 523—524, 526
— печени 513
— растяжимость 517—520, 520—521,
534
— расширение варикозное 536, 556
— содержание воды в стенке 116
— спадение 516, 520, 521, 529, 551,
558
— сравнение механического поведения
с механическим поведением труб-
ки из латекса 521, 522, 529
— средняя оболочка 515
— упругие свойства 517—521
— форма пульсовых волн давления
и кровотока 531—534
Вес 28, 30, 41
Ветвление артерий большого круга
292—296
---- малого круга 563—566
— асимметричное, картины течения
387—388
— вен 514
— симметричное, картины течения
384—387
— сосудов, картины течения 383—389
Вещество 159
— количество 42—43
Виллизнев круг 296
Вискозиметр капиллярный 206—207
— ротационный 206, 207
— с коаксиальными цилиндрами 207—
208
— типа конус — плоскость 207, 208—
209
— типы 206
Вихревая дорожка 97
Вихрь 96, 102, 275, 276
— в аорте 277—278, 278—279
— кольцевой 102
----в левом желудочке 275
Водитель ритма 220
Волна см. Волны
— бегущая 141—142, 149
— длина 143
— затухающая 149. См. также Зату-
хание
— отраженная 150, 151—152. См.
также Отражение
.---перенос энергии 341
вЮ Предметный указатель
Волна падающая 150, 151—152
— прошедшая 152
----коэффициент прохождения 152
— пульсовая см Пульсовые волны
— скорость см Скорость волны
— стоячая 140, 149—151
Волны в артериях 326—336, 368—370
-------отражение н прохождение в
местах ветвления 336—340, 344—
348, 348—351
— •— венах 539—540
------- проявление нелинейности
537—539
-------увеличение крутизны фронта
537—538
---- натянутой струне 139—140
— давления в артериях 144, 313—317,
328—329
------- венах 529—540
— — коэффициент отражения 341
------- пропускания 341
----множественные отражения
348—351
----отражение на одном ветвлении
336—340
----пульсовые см Пульсовые волны
-------форма см Форма пульсовых
волн
---- распространение 328—329
— затухание см Затухание
— звуковые 144
— на поверхности воды 143—144
— простые примеры волнового дви-
жения 139—144
— скорость распространения см Ско
рость волны
— увеличение крутизны фронта 537—
538
Время пребывания крови в венах
555—556
Выталкивающая сила 52
Вязкий след 97
Вязкость 49, 54—56, 70, 75—76, 80—
81
— газов 180
— динамическая 44
— жидкостей и суспензий 180—185
— зависимость от скорости сдвига
181—182
•------температуры 180—181
— кажущаяся 471
----жидкости 206
— кинематическая 44, 76
— крови см Кровь, вязкость
— относительная 184
— плазмы 186—187
— разбавленных суспензий невзаимо-
действующих частиц 183—184
=-----------Тэйлора формула 184
Вязкость разбавленных суспензий не-
взаимодействующих частиц, Эйн-
штейна формула 183—184
— суспензий деформируемых частиц
182, 185, 467
-----жестких частиц 467
— цельной крови 467'—468
------- влияние скорости сдвига
467—468
— эффективная 181—182, 183—184
-----стенки кровеносных сосудов 124
Вязкоупругие свойства 113, 440
----- вен 540
.----способных спадаться трубок
551
-----стенок кровеносных сосудов
122—126
-------------роль гладких мышц
122—123
---------------- коллагена 123
Ушаа 241—242, 268
Газ, определение 16
Гармоники 140, 322—323
Гель 165, 440, 444, 445
Гематокрит 188
’— динамический 470—471
— И движение крови 212
-----предельное напряжение сдвига
крови 210—211
Гемоглобин 189
— и кислородная емкость крови 189
Гемолиз 192, 195—197
— в условиях ламинарного течения
195
--------турбулентного течения 197
Гепарин 201
Гипертензия, влияние на давление в
мнкрососудах 448, 449
Гипертонический раствор 188
Гипотензия, влияние на давление в
микрососудах 448, 449
Гипотонический раствор 188
Гиса пучок 220
Гистерезис 123
— вен 540
Гладкие мышцы 421, 442, 445
----- в артериолах 426, 428—429
----- — — активное усилие 442—443
-------- артериях 300
-------- венулах 426
-------- капиллярах 426
-------- сосудах малого круга 585
----- и вены 514—515
-----кровеносных сосудов и нх мио-
генная реакция 303
------------- упругие свойства и$
ртецок 116, 12?
Предметный указатель 611
Гладкие мышцы, сокращение 301—303
----тонус 301, 585
---- и упругие свойства крове-
носных сосудов 122, 301—303
Гладкомышечные клетки 421, 426,429,
434, 436
----в артериолах 428—429
Гравитационное притяжение 30 См
также Сила гравитационная,
Ускорение силы тяжести
---- влияние на давление в сосудах
310—311
----------систему кровообращения
311—313, 553, 571—572, 581—582,
585, 559—560, 603—604
Гука закон НО—111, 112
---- и свойства стенок кровеносных
сосудов 116, 118
Давление 49
— артериальное 310 См также Арте-
риальное давление
— в альвеолах 557
---- аорте 313—317
----брюшной полости 557—559
----грудной полости 557—559
---- крупных артериях 326—336
----левом желудочке 314—315
----левом предсердии, влияние мы-
шечной работы 583—584
---- легочной артерии 581—583
----------влияние мышечной рабо-
ты 582—584
----лимфатических капиллярах
456—457
----межклеточном пространстве
455—457
---- мнкрососудах 446—455
--------математическое моделирова-
ние 450—453
— внутриплевральное 557
— внутрисосудистое, для сосудов ма-
лого круга 571—572
— в правом предсердии 557—559
----яремных венах 257
— гидростатическое 49—52, 63, 311
— градиент 65
— — «боковой» 88
----и профиль скорости 71—72
----отрицательный 57—58
----положительный 57—58, 95
— датчик, принцип работы и основ-
ные характеристики 146
— динамическое 63
— единицы измерения 44, 51—52
— закрытия сосудов, критическое 443
— коллоидно осмотическое 171, 188
483—484, 485—486
Давление наполнения 257
— околососудистое, для сосудов ма-
лого круга 572—576
— онкотическое 171, 483—484
----плазмы 188, 491—492
— осмотическое 171
----плазмы 187—188
— полное 63
— тканевое 455—457
— тканевой жидкости 455—457
— трансмуральное 118, 126
---- в артериях 309—317
-------- венах 516
-------- микрососудах 455
--------сосудах малого круга 571—
576
---- отрицательное 520
— центральное венозное 257
Движение жидкости см Поток, Те-
чение
----при обтекании сферы 101—102
---- тел 91—102
----------цилиндра 91 —101
------------- и число Рейнольдса
96—101
-------------с некруглым попереч-
ным сечением 99
— — уравнение 56—58
— — через узкое отверстие в диа-
фрагме 86—88
— капель жидкости 458—459
— концентрированных суспензий
искусственных частиц 464—467
--------эритроцитов 464—467
— кровн в венах 529—553
-------- капиллярах альвеол 589
--------большого круга 474—482
----------отрицательный зазор 475,
478—482
----------положительный зазор 475,
477—478
----------теория смазки 477, 479
-------- капиллярном канале альвео-
лярных перегородок 595—599
--------крупных артериальных сосу-
дах большого круга 317—320 См
также Аорта, Артерии
----------сосудах малого круга
587—595
--------относительно крупных мик-
рососудах 457—474
— монетных столбиков 463
— тел в жидкости, вязкое торможе-
ние 508
---------- сопротивление гидроди-
намическое 94, 504—505
-------------лобовое 94, 95, 99
----------------коэффициент 99—
102
612 Предметный указатель
Давление одиночных частиц 458—461
--------при высоких скоростях те-
чения 461
— — эритроцитов при пуазейлевском
течении 461—464
— палочек и дисков 460—461
— по окружности 25—27, 32, 88
— сферических частиц 458—459
— частицы, уравнение 28
— эритроцитов с искусственно повы
шенной жесткостью 463
Демпфирование см. Затухание
Деформация жидкости 53—54
— пластическая 112
— скорость 54
— твердого тела ПО—115
— эритроцита 195
Диастола 223
Дисперсия индикатора в жидкости,
движущейся в трубке 401—404
Диссипация энергии 37, 63—64, 145,
151
Дифференциальное исчисление, тер-
минология 22
Диффузия 159, 160—164
— в капиллярах 482—483, 493—495
— встречная 167
— зависимость от температуры и
вязкости 160—162
— и броуновское движение 160, 162—
163
— и объемный поток 174—178
— коэффициент 44, 161, 163
— несвободная 166
— ограниченная 166, 167—169, 503,
504
— путь транскапиллярного обмена
крупных молекул 507
— через поры и мембраны 166—169
Длина, определение 19—20
Дыхание, влияние на вены 557—560
(dp/d/) max 269—270
Единицы измерения 39—46
-----основные 40—41
Емкость венозной системы 513
Желудочек левый, конечно-диастоли-
ческое давление 257, 258
----напряжение в стенке 246—251
----ориентация мышечных волокон
в стенке 218—220, 249
----работа за удар 263—264
---- сила изгнания 279
---- строение стенки 218—220
----толщина стенки 246
----форма 246—248
----эллипсоидальная модель 245—
246, 248
— напряжение в стенке 244, 244—245,
246, 249, 250
— правый, механизм изгнания крови
252
— — форма 251—252
— рабочая характеристика 265, 266
— сократимость 265—267
— сократительное состояние 265
— сокращение 222
— характеристика 253—260
------- насоса 260—270
Жидкость неньютоновская 56, 206
— несжимаемая 60 См также Мате
риал
— ньютоновская 55, 65, 450
— определение 16
— основы механики 47—64
— тканевая см. Межклеточное про-
странство, жидкость
— элемент 17—18, 47
Жировые полоски 405, 406, 407, 408—
409, 410
Затухание 144—149
— волн 149
---- в артериях 364—368
-------веиах 539—540
— коэффициент 146
— критическое 147
Звук, скорость в жидкости 60
Зенона парадокс 15, 16, 21
Золь 164—165, 440
Зубец дикротический 280, 284, 315
Желудочек 216
— вихрь кольцевой 275
— давление 243
— — (dp/df)max 269—-270
---скорость повышения 268—269
— конечно-диастолическое давление
257, 258, 263
---------- и объем 257
— кривая функционирования 265
Изгиб 115
— балки 527—528
— стенки сосуда 520
-----трубки 130, 131, 525—529
Измерение кровотока методом разве-
дения индикатора 398
---------- красителя 398
Изоволюмический период 223
Изотонический раствор 188
Импеданс в венах 557
— венозной системы 553
Предметный указатель 613
Импеданс входной, модуль 351—352
----- определение 413, 415
----- фаза 351—352
— источника, определение 415
— математическое представление
412—413
— модуль, определение 414
— определение 412
— погонный, определение 414
— согласование 340—342
— терминальный, определение 415
— терминология 412—415
— фаза, определение 414
— характеристический 338
----- определение 414
— эффективный, определение 412—
413, 415
Индикатора дисперсия 401—404
Инерционные силы 72, 75—76, 81—82
Инерция 28, 72, 80, 88
— в трубквх, способных спадаться
550—551
— и колебательное движение 132—
133, 140, 143—144
— элемента жидкости 58
Инотропные влияния, определение
243
Интерстициальное пространство см.
Межклеточное пространство
Каналы в системе микрососудов, ос-
новные 421, 449—450
—---------предпочтительные 421
Капилляры 421, 423, 425, 443—444,
482
— альвеолярные см. Альвеолы, капил-
лярный канал
-----движение крови 589, 595—599
—• — размеры 578—579
— — растяжение и раскрытие 585—
587
----- упругие свойства 578—579
— гладкие мышцы 426
— давление 446—447
— движение крови 443—444, 446,
474—482
— диаметр 12—13, 420, 423
— диффузия 493
— длина 12—13, 420, 423
— лимфатические 436
----- давление 456—457
— малого круга, канал в альвеоляр-
ных перегородках 566
-------морфология 565—566
-------объем содержвщейся крови
581
----------------изменения в зави-
симости от физиологических усло-
вий 585
Капилляры, механические свойства
443—445
— напряжение сдвига на стенке 409
— плотность 420, 424
— площадь поверхности 424
— проницаемость 483
— — для жиронерастворимых моле-
кул 493
---------- — очень крупных молекул 501
------зависимость от молекулярного
веса 494, 501
— Рейнольдса число 12—13
— сеть 422, 423, 424
— система 421—423
----в скелетной мышце 423
— скорость крови 12—13, 420
— соединительная ткань 426
— строение 429—434
— эластическая ткань 426
— фильтрация см. Фильтрация в ка-
пиллярах
— эндотелий 426
— Юига модуль 444
Катетеры 146
Катехоламины 266
Киноангиография 246—247, 274, 277
Клапан аортальный 98, 216—217, 276
----механика закрытия 276—279
— атрио-вентрикулярный 216, 533
— венозный 515, 532
----неполноценный 536, 556—557
----отражение волн 536—537
---- синусы 515
— легочный 216
— митральный 216
----движение крови 274, 275
---- механика закрытия 274—276
— — Рейнольдса число 274
— трехстворчатый 216, 533
Клетки гладкомышечные см. Гладкие
мышцы, Гладкомышечные клетки
— крови 189—200
— эндотелиальные см Эндотелиаль-
ные клетки, Эндотелий
Колебание, амплитуда 135—136
— гармоники 140, 141, 156
— гармоническое простое 138—139
— основное 140, 141, 156
— период 135—136
— со сверхкритическим затуханием
147
— фаза 138, 142
— — опережение и отставание 138—
139
Колебания и волны 132—158. См.
также Волны
— самовозбуждающиеся 547
----в трубках, способных спадаться
547, 550—551
614 Предметный указатель
Колебательное движение отдельной
частицы 132—139
Количество движения 34
— тепла 42, 44
Коллаген 116, 122, 123, 301, 428, 429,
436, 440, 442, 523
— и стенки вен 514, 515, 519
— упругие свойства 116
Коллоидная система межклеточной
жидкости 440
Коллоидное состояние 164
Конвекция 159
— свободная 64
Конусность артерий, влияние на рас-
пространение пульсовых волн
356—358
— вен 514
Концентрация, определение 160
Координаты 19
— точки по отношению к осям 19
Коронарный кровоток 250
Короткова тоны 158, 551—552
Косинус 136—137
Коэффициент диффузии 44, 161, 163
— затухания 146, 149
— массопередачи 165-—166
— отражения 151
---осмотический 173, 503
— фильтрации 171
Кривая напряжение — деформация
119
------- линеаризация 119—120
Кристаллические твердые тела 180
Кровь 179—213. См. также Движение
крови
— вязкость 55, 56, 180—181, 209—213,
467
— — измерение 457
•-- принципы измерения 206—207
— давление см. Артериальное давле-
ние, Давление
— как несжимаемая жидкость 60
— кислородная емкость 189
— клетки см Клетки крови
— механические свойства 204—213
— предельное напряжение сдвига 210
— свертывание 200—201
---в кровеносной системе 201—
203
Кровяные пластинки 189. См также
Тромбоциты
Кручецие, определение 115
Куссмауля симптом 559
Лапласа закон 130, 245, 312, 441
Лейкоциты 189, 190, 198
— механические свойства 198
— размеры 189, 198
Лейкоциты, типы 189, 198
Легкие, альвеолярно-капиллярный
барьер, строение 568—570
— кровоток, распределение по зонам
559—600
— ----------- влияние давления в
альвеолах 600—602
-------------------в артериях и ве-
нах малого круга 600—602
---------------- наполнения легких
воздухом 602—605
— объем, влияние иа сопротивление
сосудов малого круга 604—605
Лимфатическая система 425
Лимфатические сосуды, строение 425,
436
Линеаризация 119, 154—155
Линейность 153—156, 331—332, 334
Линии тока 61
Липопротеиды 188
Малый круг кровообращения 561—
605
—------анатомическая организация
563—570
—------входной импеданс 591—593
—------давление в околососудистом
пространстве 572—576
------- сосудах 571—572
------- трансмуральное 571
—------движение крови в крупных
сосудах 587—595
-------сосудах межаль-
веолярных перегородок 595—599
-------общая характеристика 561—
562
-----— объем крови 579, 581
-----—-------в зависимости от фи-
зиологических условий 581—587
— — —--------методы измерения 579,
581
-------сосуды альвеолярные 572,
575
----- ---- гистология 567—570
----------интрапареихимальные
572—574
----------растяжимость 576—577
---------- упругие свойства 571,
576—579
--------- — экстраальвеолярные 572
----------экстрапаренхимальные
572
Масса, принцип сохранения 59—60
— тела 42, 44
----- как мера инерции 28
— частицы 15—16
Массопередача см. Массопереиос
— коэффициенты 165—166
Массопереиос 159—178, 482—511
— определение 159
Предметный указатель 615
Материал изотропный 116
— несжимаемый 115
— однородный 116
Маятник простой 133—138
----уравнение движения 134
Мегакариоциты 198
Международная система единиц 39,
43—46
Межклеточная щель 426, 436—439,
493, 504, 505
Межклеточное пространство 165, 425,
436, 440, 483
---- давление 455—457
----жидкость 440
— — коллоидная система 440
— соединение 426, 436—439, 493
Мембрана базальная 426, 429, 434,
436, 441, 444
----модуль Юнга 444
— полупроницаемая 483, 502
— эритроцитов 191, 192, 192—194
Метартериола 419, 421, 429
Механика жидкости 47—64
— определение 15
— твердого тела 110—131
Механические модели сердечной мыш-
цы 235
----скелетной мышцы 233—234
Микрососуды большого круга 417—
511
-------давление, влияние гипертен-
зии 448—449
---------- гипотензии 448—449
-------движение крови 457—458,
461—482
— -----массоперенос 482—511
-------механические свойства 440—
446
—------организация русла 419—425
------- перераспределение кровото-
ка 421, 423
-------профили скорости 473—474
------- пульсовые колебания давле-
ния 450
---------------- распространение
453—455
------- распределение давления
446—453
—------------математическое моде-
лирование 450, 453
-------Рейнольдса числа 12—13
-------строение стенки 425—439
—------тканевой состав стенки 426
Микрофибриллы цитоплазматические
428, 436
Миофибриллы гладких мышц 300
— Миокарда 224—225, 226
ЭДитрхондрии 19Q
Митральный клапан см Клапан мит-
ральный
Модель упругого резервуара 326—
328
-------и кровообращение в малом
круге 591
Моенса — Кортевега уравнение 333
Моль, определение 43
Мольная доля 160
Молярная концентрация 160
Молярный поток 160
Монетные столбики 197, 205, 463, 467
-----длина, влияние скорости сдви-
га 212
Мощность 44
Мышечное сокращение изометриче-
ское, определение 232
-----изотоническое, определение 232
----- и теплопродукция 232
----- одиночное 231
----- скорость укорочения 232—233
-----соотношение сила — скорость
для сердечной мышцы см. Сер-
дечная мышца, соотношение
сила — скорость
----------------скелетной мышцы
233—234
----- тетаническое (тетанус) 231
Мышца, активное состояние 230, 235,
236—238
— гладкая см. Гладкие мышцы, Глад-
комышечные клетки
— сердечная см. Сердечная мышца
— скелетная, влияние на вены 553—
557
----- система микрососудов 423
-----соотношение длина — напряже-
ние 228—229, 230
— сосочковая (папиллярная) 216,231,
274
Мюллера проба 557—559
Нагрузка 232
— предварительная 241, 257
— рабочая 259
Напряжение в стенке 244, 244—245,
246, 249, 250
— единицы измерения 44
— нормальное 49, 53, 112, 127—128
— окружное 118, 128—129, 440, 441
— определение 47—49
— продольное 119
— растягивающее 32
— сдвига 55, 115, 472
-----на стенке 70, 409—410
— — предельное 55, 210
— сжимающее 129
— тангенциальное 49, 54
Ндтяжерие 32, Щ—Ц2, 13?
616 Предметный указатель
— окружное в стенке трубки 129
Натяжение поверхностное 32, 44, 143
— струны 140
Начальный участок 65
------в аорте 379—382
----трубке, характер течения
71—72
----длина, в аорте 381—382
----------венах 541
-------при ламинарном течении 75
---------- очень низких числах
Рейнольдса 75
----—-----турбулентном течении 80
Невесомость 33
Нелинейность, проявление в артериях
359—361
---- — венах 537—539
— реальных физических систем 154—
156, 361
Нернста уравнение 163—164, 505
Неустойчивые состояния 131
Нити сухожильные 216, 274
Ньютона второй закон 28—29, 35, 40,
41, 50
— законы движения 15—18, 27—33
Область входа 65. См. также Началь-
ный участок
Обморок 311—312
Оболочка внутренняя артерий 297—
298, 301
---- артериол 426
— наружная артерий 297, 300, 301
— артериол 429
— средняя артерий 297, 298, 300, 301
302
----артериол 428—429, 441
---- вен 515
Объем, единицы измерения 43
• — уточнение термина 43
Осмос 169, 171—173
— обратный 170
Осмотический коэффициент отраже-
ния 173, 503
Отношение толщины стенки к диа-
метру, для артерий 12—13, 305,
526
----------------вен 12—13, 514,526
—---------------кровеносных сосу-
дов 12—13, 118
Отражение волн 149—153
---- в артериях 336—358
------- венах 536—537
— коэффициент 151, 341
— от закрытого конца 342—343
----открытого конца 343—344
— полное 151
— положительное и отрицательное
§42-544
Отражение частичное 151
Отрыв потока 84, 85. См. также По-
граничный слой, отрыв
- в трубках, способных спадаться
548—550
Перенос крупных молекул 507—511
Перераспределение кровотока 421, 423
Перикард 217
Перикардит констриктивный 559
Периметр вен 517—518, 519—520
Перицит 429, 434
Плазма 186
— вязкость 186—187
188
— коллоидно-осмотическое давление
188
— осмотическое давление 187—188
— отделение 471
— состав 179
Плотность 44, 47
Площадь поперечного сечения арте-
риального русла большого круга
292—296
----------— малого круга 565, 567
------- артерий 292—296
------- вен 516—520
-----— сосудов 12—13, 513—514
Пограничный слой 72—75, 94—96
— — в венах 541
-----гидродинамика 72—75, 90—91
-----при нестационарном течении
83
-----диффузионный 175
-----толщина 175—178
----- определение 72
----отрыв 85—86, 95, 100
------- при ламинарном и турбу-
лентном течении 86, 100—101
-----толщина 72, 73—75
Подобие 103—104
— и моделирование 104
Ползучесть 113, 122
Полупроницаемость мембран 168—
169, 172—173
Поток нестационарный 59
— полностью развитый 65
— стационарный 59
Предел прочности при растяжении
116
— текучести 112, 467
Предельное напряжение сдвига 55,
210
Предельный переход 21—22
Предсердие 216
— правое, а- и v-волны 272
— — движение крови 272
Прикрепление кровеносных сосудов
118, 121, 126
Предметный указатель 617
Прилипания условие 56
Принцип сохранения массы 62
-----энергии 36, 61, 62
Приосевой эффект 461. См. также
Сегре — Зильберберга эффект
Пристеночный слой 470, 472
Продольная деформация 111
Проницаемость 170, 483, 493—494
— измерение, изогравиметрический
метод 502
----- метод разведения индикатора
497—499
-----методы 495—503
— произведение проницаемости на
площадь 495, 505
— эндотелия 407—409, 411—412
Проток грудной 425
Прохождения коэффициент 152
Псевдоподии тромбоцитов 199—200
Пуазейлевское течение 66—70, 71, 90,
450, 458, 461
-----и движение одиночных эритро-
цитов 461—464
-----напряжение сдвига 68—69
— — скорость сдвига на стенке 70
Пуазейля закон 66—70, 103, 108, 505,
548
Пуассона коэффициент 114—115, 115,
121, 445
Пузырек 428
— пиноцитозный 439, 507—508
механика 508—511
Пульсовые волны в аорте, гармони-
ческий состав 323—324, 325
-------венах малого круга, проис-
хождение 587—589
-------микрососудах большого кру-
га 450
-----давления в крупных артериях
большого круга 313—317
------- распространение в крупных
артериях большого круга 326—
336
----------в микрососудах большого
круга 453—455
----------— сосудах малого круга
589—593
Пуркине волокна 220
Пучность 150, 343, 346—347
Пучок атрио-вентрикулярный 220
— Гиса 220
Работа 34—35, 44
— отрицательная 232
— сил давления 62
Равновесная поверхностная концен-
трация 173—174
Радиальные блуждания эритроцитов
468—470
— перемещения частиц 458, 459, 461,
467
---- эритроцитов 463
Раздражение тетаническое (электри-
ческое) 231
Размерностная однородность 43,
107—109. См. также Анализ раз-
мерностей
Рассеяние энергии см. Диссипация
энергии
Раствор насыщенный 173
Растяжимость 121
— вей 517—520, 520—521, 534
— сосудов малого круга 576—577
Расход объемный 61
— определение 43
Расширение постстенотическое 396—
397
Резонанс 98, 152—153, 155, 346—347
Рейнольдса число 75, 75—76, 94, 95—
96, 417
----для аорты 12—13, 274
----вен 12—13, 540—541
—-------легочного ствола 12—13, 274
----митрального клапана 274
---- критическое 79, 389
-------- для течения через отверстие
в диафрагме 88
— — определение 75—76
---- переходные значения 79
Релаксация напряжения 113, 122
Рефлекторные влияния на сердце 214
Саркомер, длина 228—230
— поперечные мостики 227
— структура 226—227
Свертывание крови 199—203
----и тромбоз вен 202
— — образование сгустка 200—201
----процесс 199
Сегре — Зильберберга эффект 461,
463, 467
Сердечная мышца 224—243
----активное состояние 230, 235—
238, 239—240, 241
----динамические механические
свойства 230—243
----изометрическое сокращение 235,
235—236, 239
----изотоническое сокращение 239
---- максимальная скорость укоро-
чения (Утах) 241—242, 268
----механические модели 235, 242,
243
----миофибриллы 224—225, 226
----напряжение активное 228—229
--------покоя 228, 234
-------- развиваемое 228
618 Предметный указатель
Сердечная мышца, параллельный уп-
ругий элемент 234, 235, 242
----последовательный упругий эле-
мент 233—234, 238, 243
---- потенциал действия 231
------- покоя 231
----саркомер 226—228
----скорость укорочения 234, 236—
237, 238, 242—243, 255
----сократимость 240, 241, 243,
265—270
----- сократительный элемент 233,
234, 236, 237, 239
----------скорость укорочения 236—
238, 241, 255
----соотношение длина — напряже-
ние 228—230
-------сила — скорость 235, 238,
239, 239—240, 243, 254—255, 267
---- статические механические свой-
ства 228—230
---- строение 224—228
Сердечно-легочный препарат 260
Сердечный цикл 220—224
------ механические явления 222—224
---- электрические явления 220—222
Сердце 214—286 См также Желудо-
чек, Сердечная мышца
— анатомия 216—220
— блокада 533
— водитель ритма 220
— гидродинамические аспекты дея-
тельности 272—282
— клапаны, гидромеханика 274—279.
См. также Клапан
— механические аспекты деятельно-
сти 243—272
— производительность, влияние дыха-
ния 557—560
— рефлекторный контроль 214
— смешивание 398—404
— тоны 282—286
----второй 223, 284
----первый 223, 283, 284—285
— турбулентность течения крови 400
— ударный объем 215, 224
— шумы 286
«Сжатое сечение» 87
Сила 27—28, 44
— возвращающая 132—133, 140
— гравитационная 36—37, 47
— демпфирующая 144, 145
— диссипативная 37
— инерции 58. См. также Инерцион-
ные силы
— консервативная 37
— массовая 47
— объемная 47
— размерность 40—41
Сила трения 54
— центробежная 32
Силы близкодействующие 30, 31, 47
— дальнодействующие 30, 47
Синус 136—137
Систола 223
Скорости градиент 55, 71
— профиль 66
----в аорте 372—376, 377—389
------- венах 541
------- крупных артериях 370—379
------- микрососудах 473—474
----жидкой фазы суспензии 466—
467
---- частиц в суспензиях 465—466
Скорость см Течение, скорость
— волны в аорте собаки 12—13,
360
------- артериях малого круга 12—
13, 577
-------венах 12—13, 534—536, 537
---------- зависимость от давления
534—536
----— кровеносных сосудах, спо-
соб определения 329—330
-------микрососудах 453, 454
------- натянутой струне 142—143
----кажущаяся 333
----«от подножия до подножия»
333
---- собственная 333
— единицы измерения 44
— изменения величины 22
— конечная 204
— крови в микрососудах 12—13, 420
— определение 20—24
— сдвига 55, 70, 90, 418, 472, 473
----на стенке 55, 70, 90—91, 418,
473
— составляющие 22
След 97
Смешивание крови в мелких сосудах
400—401
•------системе кровообращения
398—399
Соединительный комплекс 439
Сокращение желудочков сердца 222
Соотношение давление — площадь для
вей 521—522
----------трубки из латекса 521—
522
---- расход для артерий 309—325
---------- вен 543
----------способных спадаться тру-
бок 544—551
— длина — напряжение 228—230
— сила — скорость 233—234 См так-
же Сердечная мышца, соотноше-
ние сила — скорость
Предметный указатель 619
Сопротивление сосудов малого круга
595—605
----------и объем легких 603—605
— трубок, способных спадаться 546—
551
----эллиптического сечения 548—
549
Сосуд кровеносный крупный, анато-
мия 289—292
—------напряжение сдвига на стен-
ке 408—411
—------перемешивание крови 398—
404
------- прикрепление 118
----отношение толщины стеики к
диаметру 12—13, 523—524
----свойства стенки НО—131
Сосудистое русло крыла летучей мы-
ши, основные параметры 295
--------------- параметры микрососу-
дов и вен 513—514
---------------сальника кошки 514
--------------- собаки 514
Сосуды стенок артерий большого кру-
га 301
-------малого круга 570
---- вен 515
Спадение артерий 551—552
— вен 516, 520, 521, 529, 551, 558
— кровеносных сосудов 131
Среда сплошная 16—17, 47
Старлинга гипотеза 483—484
— закон сердца 260—262
Статика упругой трубки 126—131
Стеноз, сход вихрей 98
Степени свободы 139
Стокса закон 109, 164, 204
Стокса — Эйнштейна радиус 164, 505
Струи образование 286
Струхаля число 99
Сужение в трубках 83—88
Сурфактант легочный 570
Суспензии, вязкость 180—185
— движение в узких трубках 457—
467
Сфигмоманометр 309
Сфинктер прекапиллярный 421, 429
Сыворотка, вязкость 186—187
— состав 186
Твердое тело деформируемое 110
----кристаллическое, структура 180
---- механика 110—131
---- определение 16
Температура 42, 44
Теплота 42. См. также Количество
тепла, Энергия тепловая
Термоанемометрия пленочная 371 *
Течение в трубке 65—91
---------- — ветвящейся 384—388
------ — изогнутой 88—91
------— профили скорости 382—
383
--------прямой 65—88
--------способной спадаться 543—
552
— вторичное 89
-----в венах 541—543
— крови в сосудах см Движение
крови
— ламинарное 79, 86, 389
— нестационарное 80—83. См. также
Поток
-----мгновенные профили скорости
380
— поршневое 465, 473
— пуазейлевское 65—70
----- при наличии колебаний 80
— скорость мгновенная 321
-----объемная (расход) 61, 320
----- средняя 321
----------по времени 321—322
-----терминология 320—322
-----усредненная по ансамблю 322
— стационарное см Поток
— турбулентное 77—80, 86
----- свойства 389—397
— устойчивое 88, 389—397
Тонус сосудов 301
Транскапиллярный обмен 483
-----диффузионно ограниченный 497
-----ограниченный потоком 497
-----путь диффузии 503—504
----------и теория пор Паппенхай-
мера 504—505
---------- перенос воды 505—507
Транспорт активный 169, 482
Трение 31, 37, 54
— коэффициент 31
Тромбоз 201—203
— в артериях 202
-----венах 202—203, 556
----- левом предсердии 203
----- микрососудах 202
Тромбоциты 189, 198—200
— и кровотечение 200
----- свертывание крови 199—201
— коэффициент диффузии 468—470
— псевдоподии 199—200
— размеры 189
Трубка из латекса, соотношение дав-
ление— площадь 521, 522
Турбулентное течение в трубке 77—80
Турбулентность потока в артериях
389—397
----- в сердце 286
Тэйлора дисперсия 499
620 Предметный указатель
Ударная волна в артериальной систе-
ме 361
-------- венах 537
Ударный объем, определение 224
Узел 150, 342—343, 346—347
— атрио-вентрикулярный 220
— колебаний 140, 141
— сино-атриальный 220
Ультрафильтрация 170
Уомерсли параметр (а) 80—83, 330—
331, 417, 454, 464
---в артериях 12—13, 330—331
--------венах 12—13, 540
--------микрососудах 12—13, 417,
454, 464
Упругость 110—131, 440
— вен 516—517
— линейная 112
— определение 110—115
— стенки кровеносных сосудов 116—
126
Ускорение 24—27, 44, 72
— конвективное 58—59
— местное 58—59
— определение 24
— силы тяжести 30
Фагоцитоз 190
Фаза 159. См. также Колебание
Фенна эффект 232
Фибрин 200—201
Фибриноген 179, 188, 197, 199, 205,
210
Фибринолиз 201
Фибробласты в стенках вен 514
Фика закон 160—161, 493
Фильтрация в капиллярах 483
------- влияние гипопротеинемии
488, 489
----------дегидратации 488, 489
----------повышения венозного
давления 488, 489
----------расширения сосудов 488,
489
-------сужения сосудов 488,489
-------коэффициент 485, 487, 491
-------методы изучения, изограви-
метрический 489—490
---Лендиса метод 484—
485, 489
---скорость 490—491
--------зависимость от типа эн-
дотелия 490—491
---капиллярном русле 489
--- отдельных капиллярах
коэффициент 171
— через мембраны 170—171
Фильтрация через поры 492—493
Фореуса-Линдквиста эффект 471—472,
478, 482
Форма пульсовых волн в сосудах ма-
лого круга 587—589
------- давления в артериях малого
круга 587, 588
-----------------крупных артериях
большого круга, анализ 351—356
-------------левом предсердии 587,
588
—---------волна а 221, 223, 272,
530—531, 532
-----------------v 221, 223, 272, 530—
531, 533
-----------------с 530—531, 532
----------и расхода в аорте, гар-
монический состав 323—324
------------------- легочном стволе
587, 588
----------------- скорости крови в ве-
нах большого круга 531—534,
558—559
-------расхода в венах малого кру-
га 587, 588
Франка — Старлинга механизм 262
Фурье анализ 156—158, 322—325
Хиломикроны 179, 188
Ходьба, влияние иа вены 554—556
Хронотропные влияния, определение
243
Центр масс 16
— тяжести 16
Частицы асимметричные 184—185
— деформируемые 182
— и сплошная среда 15—18
— механика 19—38
— определение 15—16
— положение 19—20
---радиус-вектор 23
— твердые, сферические 181—182
Частота 44
— колебаний 137
---в случае стоячих волн 140—141
— основная 323
— собственная 140
------ затухающих колебаний 146
— — незатухающих колебаний 147
— угловая 44, 137
«Шейки» в растянутой проволоке 111,
113
Шмидта число 178
Шумы в сердце 286, 394—395
Предметный указатель 621
Эванс синий и проницаемость эндо-
телия артерий 407
Эластин 122, 123, 301, 440
— в венах 519
— упругие свойства 116—117
Электрокардиограмма 220—222
Эллиптичность вен 517, 524—525
Эндотелиальные клетки 426, 428, 429
----проницаемость 411—412
----соединение 436—439
Эндотелий 297—298, 429, 432, 434,
435, 444
— в артериолах 426
---- венулах 426
---- капиллярах 426
— непрерывный 429, 432
— окончатый 432, 493
— прерывистый 432, 434, 457, 493
— проницаемость 407—409, 411—412
----перенос крупных молекул 411 —
412
Энергия 42, 44
— диссипация 37, 63—64, 145, 151
— запасенная 36
— кинетическая 35, 62
— поглощение 152
— потенциальная 36, 62
— сохранение 36, 61, 144
— тепловая 61
Эритроциты 189—197
— вращение 195
— вязкость 190
— деформация 195
----в суспензии при сдвиговом те-
чении 195
— диаметр 17, 189, 423
— коэффициент диффузии 468—470
— мембрана 191, 192, 192—194
---- сопротивление изгибу 192
Эритроциты, мембрана, сопротивление
растяжению 192
— метаболизм 191
— оседание в крови 204—205
-----скорость (СОЭ) 204—205
— радиальные блуждания 468—470
— свойства при заболеваниях 190—
191, 474
— с искусственно увеличенной жест-
костью 213
— способность деформироваться, гиб-
кость 193, 213
— сферические 187, 193, 213
— тенденция к образованию агрега-
тов 197
— тени 192, 464—465
Эхокардиография 258
Юнга модуль 114, 115
-----артерий большого круга 12—
13, 122, 523, 525
--------малого круга 523, 525
----- артериол 442
-----базальной мембраны 444
-----вен 12—13, 122, 521, 523, 525
-----динамический и вязкоупругие
свойства стенок кровеносных со-
судов 124—125
-----для окружного растяжения 121
--------продольного растяжения 121
----- капилляров 444
-----касательный 119, 121, 307,
520—521
----- коллагена 442
-----межклеточного геля 445
— — мембраны эритроцитов 193
— трубки из латекса 522, 529
-----эффективный 118, 119, 129
Оглавление
Предисловие редакторов перевода ..................................... б
Предисловие авторов к русскому изданию.......................... .... 8
Предисловие........................................................... 9
Благодарности......................................................... 14
Часть I. ОСНОВЫ МЕХАНИКИ
Глава 1. Частицы и сплошные среды....................................15
Глава 2. Механика частиц.............................................19
2.1. Положение частицы...........................................19
2.2. Скорость....................................................20
2.3 Ускорение ...................................................24
2.4. Законы движения Ньютона: масса и сила.......................27
2.5. Количество движения.........................................34
2.6. Работа и энергия............................................34
Глава 3. Единица измерении...........................................39
3.1. Единицы измерения и размерности.............................40
3.2. Масса, длина и время как основные величины..................40
3.3. Неудобство использования силы в качестве основной величины . . 41
3.4. Энергия и теплота . . . '...................................42
3.5. Понятие о количестве вещества...............................42
3.6. Размерностная однородность и совместимость единиц...........43
3.7. Использование понятий объема и расхода в физиологии ..... 43
3.8. Международная система единиц (СИ)...........................43
Глава 4. Основы механики жидкостей . ................................47
4.1. Напряжение..................................................47
4 2 Гидростатическое давление...................................49
4.3 Напряжение в движущейся жидкости; вязкость...................52
4.4. Уравнение движения жидкости................................,56
4.5. Конвективное и местное ускорение............................58
4.6. Сохранение массы . .........................................59
4.7. Теорема Бернулли............................................61
Глава 5. Движение жидкости в трубках и при обтекании тел.............65
5.1. Пуазейлевское течение в трубке..............................65
5.2. Течение на начальном участке трубки.........................71
5.3. Предстчвление о пограничном слое............................72
5.4. Число Рейнольдса............................................75
Оглавление 623
5.5. Турбулентность при течении в трубке............................77
5.6. Нестационарное течение в очень длинной трубке..................80
5.7. Влияние сужений на характер течения в трубке...................83
5.8. Течение в изогнутых трубках....................................88
5 9. Обтекание тел . ...........................................91
Глава 6. Анализ размерностей...........................................103
6 1. Подобие и моделирование ... ...........................104
6.2. Некоторые примеры выбора масштабов в биологических системах 105
6.3. Метод получения размерностно однородных зависимостей между пе-
ременными ........................................................107
Глава 7. Механика твердого тела и свойства стенок кровеносных сосудов ПО
7.1. Свойство упругости.............................................ПО
7.2. Свойства стенок кровеносных сосудов...........................116
7.3. Статика упругой трубки........................................126
Глава 8. Колебания и волны.............................................132
8.1. Простое гармоническое движение...............................132
8.2. Простые примеры волнового движения..........................139
8.3. Затухание....................................................144
8.4. Отражения волн и резонанс...................................149
8.5. Линейность...................................................153
8.6 Фурье-анализ..................................................156
Глава 9. 'Введение в теорию массопереноса...............................159
9.1. Диффузия..................................................160
9.2. Коллоидное состояние......................................164
9.3. Коэффициенты массопередачи................................165
9.4. Диффузия через поры и мембраны............................166
9.5. Проницаемость.............................................170
9.6. Фильтрация через мембраны.................................170
9.7. Осмос.....................................................171
9.8. Простая модель массопереноса..................................173
9.9. Взаимодействие объемного потока и диффузии....................174
Часть II. МЕХАНИКА КРОВООБРАЩЕНИЯ
Глава 10. Кровь......................................................179
10.1. Визкость жидкостей и суспензий.............................180
10.2. Осмотическое давление плазмы...............................187
10.3. Взвешенные элементы........................................188
10.4. Клетки крови...............................................189
10.5. Свертывание крови..........................................200
10.6. Тромбоз.......................... .........................201
10.7. Механические свойства цельной крови.......................204
Глава 11. Сердце.....................................................214
11.1. Анатомия сердца............................................216
11.2. Сердечный цикл.............................................220
11.3. Свойства сердечной мышцы...................................224
11.4. Механическое поведение целого сердца.......................243
11.5 Гидродинамические аспекты деятельности сердца...............272
Список литературы ........................................... 286
624 Оглавление
Глава 12. Артерии большого круга........................... .... 288
12.1. Анатомическая организация и гистологическое строение . . . 289
12.2. Давление и кровоток в артериях большого круга............309
12.3. Распространение волн в артериях..........................326
12.4. Отражение и прохождение волн в местах ветвления артерий . 336
12.5. Влияние нелинейности.....................................359
12.6. Влияние вязкости.........................................362
12.7. Волны других типов.......................................368
12.8. Картины течения крови в артериях.........................370
12.9. Смешивание и перенос веществ в артериях..................397
12.10. Дополнение. Импеданс....................................412
Список литературы ........................................ 415
Глава 13. Микрососуды большого круга...............................417
13.1. Организация русла микрососудов...........................419
13.2. Строение микрососудов....................................425
13.3. Статические механические свойства микрососудов...........440
13.4. Давление в системе микрососудов..........................446
13.5. Давление в межклеточном пространстве.....................455
13.6. Движение жидкости в моделях микрососудов и в относительно
крупных микрососудах..........................................457
13.7. Движение крови в капиллярах............................ 474
13.8. Массопереиос в системе микрососудов......................511
Список литературы ........................................ 482
Глава 14. Вены большого круга......................................512
14.1. Анатомическая организация................................513
14.2. Трансмуральное давление и статические упругие свойства . . .516
14.3. Динамика движения крови в крупных венах..................529
14.4. Течение в трубках, способных спадаться...................543
14.5. Механика венозного русла.................................553
Глава 15. Малый круг кровообращения................................561
15.1. Анатомическая организация................................563
15.2. Трансмуральное давление и статические упругие свойства сосудов 571
15.3. Динамика течения крови в крупных сосудах малого круга '. . 587
15.4. Сопротивление сосудов малого круга.......................595
Список литературы..........................................606
Предметный указатель...............................................607
К. Каро, Т. Педли, Р. Шротер, У. Сид
МЕХАНИКА КРОВООБРАЩЕНИЯ
Ст. научный редактор Е. А. Яиовская Научный редактор Н. Н. Шафрановская
Мл. научный редактор 3. В. Соллертннская Художник Е. К. Самойлов
Художественный редактор Б. Н. Юдкин Технический редактор И. М. Кренделева
Корректор Б. И. Киселева
ИБ № 2395
Сдано в набор 10 03.81. Подписано к печати 29.10.81. Формат 60X901/16. Бумага типограф-
ская № 2. Гарнитура литературная. Печать высокая. Объем бум. л 19,50. Усл.-печ. л 39,00.
Усл. кр.-отт. 39,00. Уч -изд. л. 41,72. Изд. №4/1009. Тираж 5700 экз. Зак. 1089. Цена 5 р. 90 к.
ИЗДАТЕЛЬСТВО „МИР" 129820, Москва, И-110, ГСП, 1-й Рижский пер., 2.
Ленинградская типография Ns 2 головное предприятие ордена Трудового Красного Зна-
мени Ленинградского объединения „Техническая книга" им. Евгении Соколовой Союзлоли-
графпрома при Государственном комитете СССР по делам издательств, полиграфии и книж-
ной торговли. 198052, г. Ленинград, Л-52, Измайловский проспект, 29.