


























































































































































































































































































































































Text
В.В. IxopoBiiiigj*
С.П. Зуихина
в современную биологию
J о'."' VB
и дендрологию
КОРОВИН
ВЛАДИМИР ВЛАДИМИРОВИЧ
ЗУИХИНА
СОФЬЯ ПАРМЕНОВНА
доктор биологических наук, профессор, преподаватель кафедры селекции, генетики и дендрологии МГУЛ. Автор более 100 научных публикаций, в их числе 2 монографии, 12 учебников и учебных пособий. Научные интересы - анатомия растений, аномальные структурные изменения стебля, бота-
кандидат сельскохозяйственных наук, доцент, профессор кафедры селекции, генетики и дендрологии МГУЛ. Автор более 40 научных и учебнометодических публикаций. Научные интересы - анатомия растений, дендрология, систематика.
ническое древесиноведение, теория эволюции.
Московский государственный университет леса
В.В. КОРОВИН, С.П. ЗУИХИНА
ВВЕДЕНИЕ В СОВРЕМЕННУЮ БИОЛОГИЮ И ДЕНДРОЛОГИЮ
Допущен УМО по образованию в области лесного дела в качестве учебного пособия для студентов высших учебных заведений, обучающихся по направлениям "Лесное хозяйство и ландшафтное строительство" и "Лесоинженерное дело"
Издательство Московского государственного университета леса Москва - 2010
2
УДК 57 630*17; 634. 942 (075.8)
Разработано в соответствии с Государственным образовательным стандартом ВПО 2000 г. для направления подготовки 250 401 на основе примерной программы дисциплины "Биология" для специальности 260 100 Лесоинженерное дело, а также для специальности 250 201 направления подготовки 260 400 Лесное хозяйство и 260 500 Садово-парковое и ландшафтное строительство.
Рецензенты: д.б.н. М.Г. Романовский, главный научный сотрудник Учреждения РАН Институт лесоведения;
д.б.н. О.В. Чернышенко, зав. кафедрой ботаники Московского государственного университета леса.
Компьютерный набор и графика авторов
Авторы - Владимир Владимирович Коровин, профессор
Софья Парменовна Зуихина, доцент;
Коровин В.В.
Введение в современную биологию и дендрологию: учебное пособие /В.В. Коровин, С.П. Зуихина. - М.: ГОУ ВПО МГУЛ, 2010. - 343 с.
Издание представляет собой учебное пособие для студентов лесных ВУЗов, обучающихся по специальностям: лесное хозяйство, ландшафтное строительство и лесоинженерное дело.
В разделах, посвященных вопросам общей биологии, основное внимание уделено царству растений. Дендрологическая часть издания, ориентированная на будущих лесоинженеров, построена по флористическому принципу. В содержание учебного пособия кроме упомянутых разделов, включены главы, касающиеся лесорастительного районирования и основ почвоведения. В приложениях даны дополнительные сведения по систематике растений.
УДК 57 630*17; 634. 942 (075.8)
©В.В. Коровин, С.П. Зуихина © ГОУ ВПО МГУЛ, 2010
3
СОДЕРЖАНИЕ
СОДЕРЖАНИЕ...........................................................3
ВВЕДЕНИЕ.............................................................8
КРАТКИЙ ИСТОРИЧЕСКИЙ ОЧЕРК РАЗВИТИЯ БИОЛОГИИ........................12
КЛЕТОЧНОЕ СТРОЕНИЕ ЖИВЫХ ОРГАНИЗМОВ.................................21
Химический состав клетки..........................................21
Строение клетки...................................................26
Сравнение прокариот с эвкариотами.................................27
Строение растительной клетки......................................28
Плазматическая мембрана (плазмалемма)...........................29
Ядро............................................................29
Пластиды........................................................30
Фотосинтез......................................................36
Митохондрии.....................................................40
Происхождение митохондрий и хлоропластов........................41
Вакуоли.........................................................42
Рибосомы........................................................43
Эндоплазматический ретикулум....................................44
Аппарат Гольджи.................................................46
ДЕЛЕНИЕ КЛЕТКИ......................................................47
Клеточный цикл....................................................47
Митоз.............................................................49
Мейоз.............................................................51
АНАТОМИЯ ДРЕВЕСНЫХ РАСТЕНИЙ.........................................54
Краткая история анатомии растений.................................54
Ткани высших растений.............................................56
Эпидерма........................................................57
Перидерма.......................................................58
Паренхима.......................................................58
Колленхима......................................................59
Склеренхима.....................................................59
Ксилема.........................................................59
Флоэма..........................................................59
СТРОЕНИЕ И ФУНКЦИИ СТЕБЛЯ...........................................60
Первичное строение стебля.........................................61
Прокамбий.......................................................63
Вторичное строение стебля.........................................64
Строение травянистого стебля....................................64
Строение стебля древесного растения.............................65
Камбий..........................................................65
Вторичные проводящие ткани........................................70
Древесина хвойных растений......................................70
Древесина лиственных древесных растений.........................72
Строение клеточных оболочек древесины...........................76
Восходящий ток..................................................79
Луб хвойных растений............................................83
Луб лиственных древесных растений...............................84
Нисходящий ток (транспорт ассимилятов)..........................88
СТРОЕНИЕ ЛИСТА......................................................90
Строение листа двудольного растения...............................90
4
Строение хвои..........................................................92
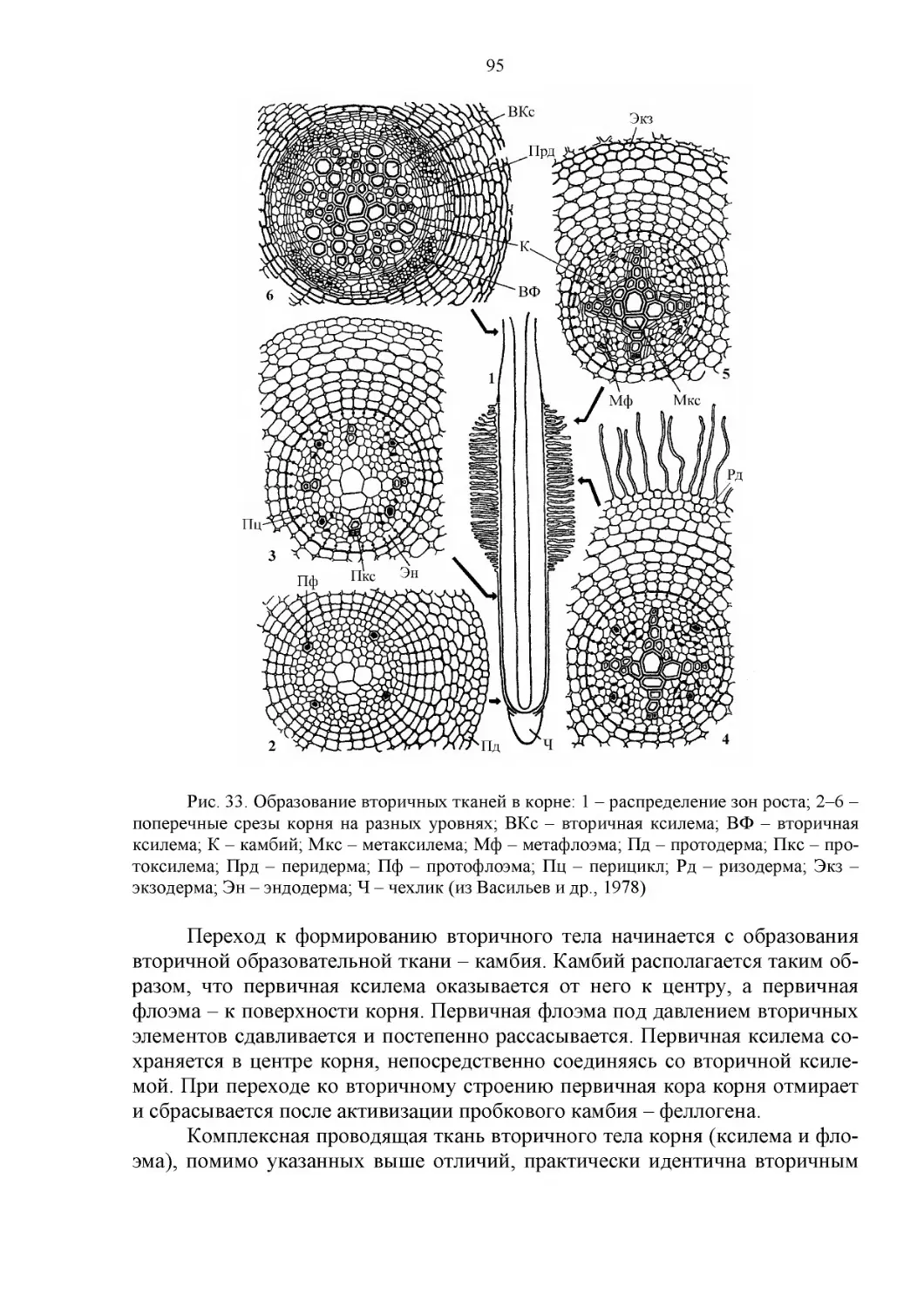
СТРОЕНИЕ КОРНЯ...........................................................94
Первичное строение корня.............................................94
Вторичное строение корня.............................................94
МОРФОЛОГИЯ РАСТЕНИЙ......................................................96
Побег, стебель.........................................................97
Лист..................................................................102
Корень................................................................105
Стробилы хвойных и цветки покрытосеменных.............................110
Соцветие..............................................................114
Плод..................................................................115
ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ НАСЛЕДОВАНИЯ ПРИЗНАКОВ..........................117
Изменчивость..........................................................117
Законы Грегора Менделя................................................120
Наследственность......................................................124
ПУТИ ЭВОЛЮЦИИ...........................................................133
Современное представление о происхождении Земли.......................133
Теории возникновения жизни на Земле...................................135
Современный дарвинизм или синтетическая теория эволюции...............138
Критика синтетической теории эволюции и другие эволюционные учения....140
ПОНЯТИЕ ВИДА В БИОЛОГИИ.................................................149
СИСТЕМАТИКА РАСТЕНИЙ....................................................153
РЕЛЬЕФ И ПОЧВЫ..........................................................157
Рельеф................................................................158
Почвы.................................................................160
Структура почвы.....................................................160
Химизм почв.........................................................161
Органическое вещество...............................................161
Содержание воздуха и воды...........................................162
Образование почвы.....................................................162
Климат и выветривание...............................................162
Материнская порода..................................................163
Топография..........................................................163
Почвообитающие организмы............................................164
Влияние человека....................................................164
Время...............................................................165
Типы почв...........................................................165
Лесные почвы..........................................................165
ПРИРОДНЫЕ ЗОНЫ РОССИИ...................................................167
Почвы и растительность природных зон..................................167
Растительность различных природных зон................................168
Растительность Арктических пустынь..................................169
Растительность Тундры...............................................169
Растительность Лесотундры...........................................169
Растительность Тайги................................................169
Растительность Зоны смешанных лесов.................................169
Растительность Зоны широколиственных лесов..........................169
Растительность Зоны лесостепи.......................................170
Растительность Степной зоны.........................................170
Влияние на растительность провинциальности, или долготной зональности.170
Растительность Субатлантического сектора............................171
Растительность Внутриматерикового сектора...........................171
5
Растительность Тихоокеанского сектора..............................171
Описание природных зон России........................................171
Зона тундры........................................................172
Зона тайги.........................................................174
Зона смешанных лесов...............................................175
Зона широколиственных лесов........................................176
Степная зона России................................................177
Некоторые особенности северной лесной зоны...........................178
ТЕМНОХВОЙНЫЕ ЛЕСА......................................................182
Основные лесообразующие породы.......................................182
Род ель Plcea Dietr..................................................182
Ель европейская Picea cibies (L.) Karst............................183
Ель сибирская -Р. obovata Ledeb....................................187
Типы ельников......................................................190
Род пихта - Abies Mill...............................................193
Пихта сибирская - Abies sibirica Ledeb.............................194
Сосна кедровая сибирская - Pinus sibirica (Rupr) Mayr................198
Типы кедровников...................................................201
Деревья второй величины, кустарники, травяной напочвенный покров.....204
СОСНОВЫЕ ЛЕСА..........................................................205
Род сосна - Pinus L..................................................206
Сосна обыкновенная -Р. sylvestris L................................207
Типы сосновых насаждений...........................................211
ЛИСТВЕННИЧНЫЕ ЛЕСА.....................................................215
Род лиственница - Larix Mill.........................................216
Лиственница сибирская -L. Sibirica Ledeb...........................217
Лиственница даурская - L. gmelinii (Rupr.) Rupr....................222
Типы лиственничных насаждений......................................223
МЕЛКОЛИСТВЕННЫЕ ЛЕСА...................................................225
Род береза - Bdtula L................................................225
Береза повислая - В. pendula Roth..................................227
Береза пушистая - В. pubescens Ehrh................................230
Род тополь - Populus L...............................................231
Осина, тополь дрожащий - Р. tremula L..............................231
Типы леса с участием березы и осины................................234
Род ольха - Alnus L..................................................234
Ольха черная (клейкая) - A. glutinosa Gaertn.......................235
Ольха серая - A. incana Moench.....................................237
Типы ольшаников....................................................238
ЕВРОПЕЙСКИЕ ШИРОКОЛИСТВЕННЫЕ ЛЕСА......................................239
Род- Qudrcus L.......................................................240
Дуб черешчатый - О. robur L........................................244
Спутники дуба........................................................247
Род клен - Acer L....................................................247
Клен остролистный - Acer platanoides L.............................248
Род ясень - Fraxinus L...............................................251
Ясень обыкновенный -Fraxinus excelsior L...........................251
Род липа - Tilia Juss................................................252
Липа мелколистная - Tilia corddta Mill.............................253
Род граб - Carpinus L................................................254
Граб обыкновенный - Carpinus betulis L.............................255
Род вяз (ильм) - Ulmus L.............................................256
6
Вяз гладкий -Ulmus Icevis Pall......................................258
Вяз голый (вяз шершавый, ильм горный) - U. glabra Huds. (U sccibra Mill.; U montdna With.)......................................................259
Вяз граболистный (вяз листоватый, берест, карагач) - U. carpinifolia Rupr. ex Suchow.
(U. folidceae Gilib.)...............................................259
Вяз приземистый - U. pumila L.......................................260
Типы широколиственных лесов...........................................260
ЛЕСА СЕВЕРНОГО КАВКАЗА..................................................261
Важнейшие лесообразующие породы.......................................263
Пихта кавказская - Abies nordmanniana (Stev.) Spach.................263
Ель восточная - Picea orientalis (L.) Link..........................265
Дуб скальный - Quercuspetraea (Mattuschka) Liebl..................266
Бук восточный - Fdgus Orientalis Lipsky.............................267
Каштан посевной - Castanea sativa Mill..............................271
Клен ложноплатановый (клен белый, явор) - Acer pseudoplatdnus L...272
Клен полевой -Л. campestre L........................................273
Орех грецкий - Jiiglans regia L.....................................274
Тополь белый (серебристый, белолистка) - Populus alba L.............276
Тополь черный (осокорь) - Populus. nigra L..........................278
ЛЕСА ДАЛЬНЕГО ВОСТОКА.................................................278
Сосна кедровая корейская - Pinus koraiensis Sieb. et Zucc...........289
Ель аянская - Picea ajanensis Fisch...............................290
Пихта белокорая - Abies nephrolepis (Trautv.) Maxim.................292
Пихта цельнолистная - Abies holophylla Maxim........................292
Тис остроконечный - Taxus cuspiddta Siebold et Zucc. ex Endl........294
Береза ребристая (желтая) - Betula costdta Trautv. и другие дальневосточные виды березы..............................................................296
Дуб монгольский - Quercus mongolica Fish, ex Ledeb..................297
Орех маньчжурский - Jiiglans mandshurica Maxim......................298
Ясень маньчжурский -Fraxinus mandshurica Rupr.......................299
Виды рода липа - Tilia Mill.........................................300
Виды рода клен -Acer L..............................................301
Виды рода ильм - Ulmus L............................................302
Чозения толокнянколистная - Chosenia arbutifolia (Pall.) A. Skvorts.303
Бархат амурский - Phellodendron amurense Rupr.......................304
ПРИЛОЖЕНИЯ..............................................................306
Приложение 1............................................................307
Приложение 2............................................................309
Приложение 3............................................................311
ЦАРСТВО ГРИБЫ.........................................................311
Основные свойства и систематика грибов..............................311
Хозяйственное значение грибов.......................................314
ЦАРСТВО РАСТЕНИЯ......................................................315
ВОДОРОСЛИ ALGAE.....................................................316
ЛИШАЙНИКИ LICHENES..................................................317
ВЫСШИЕ РАСТЕНИЯ.....................................................319
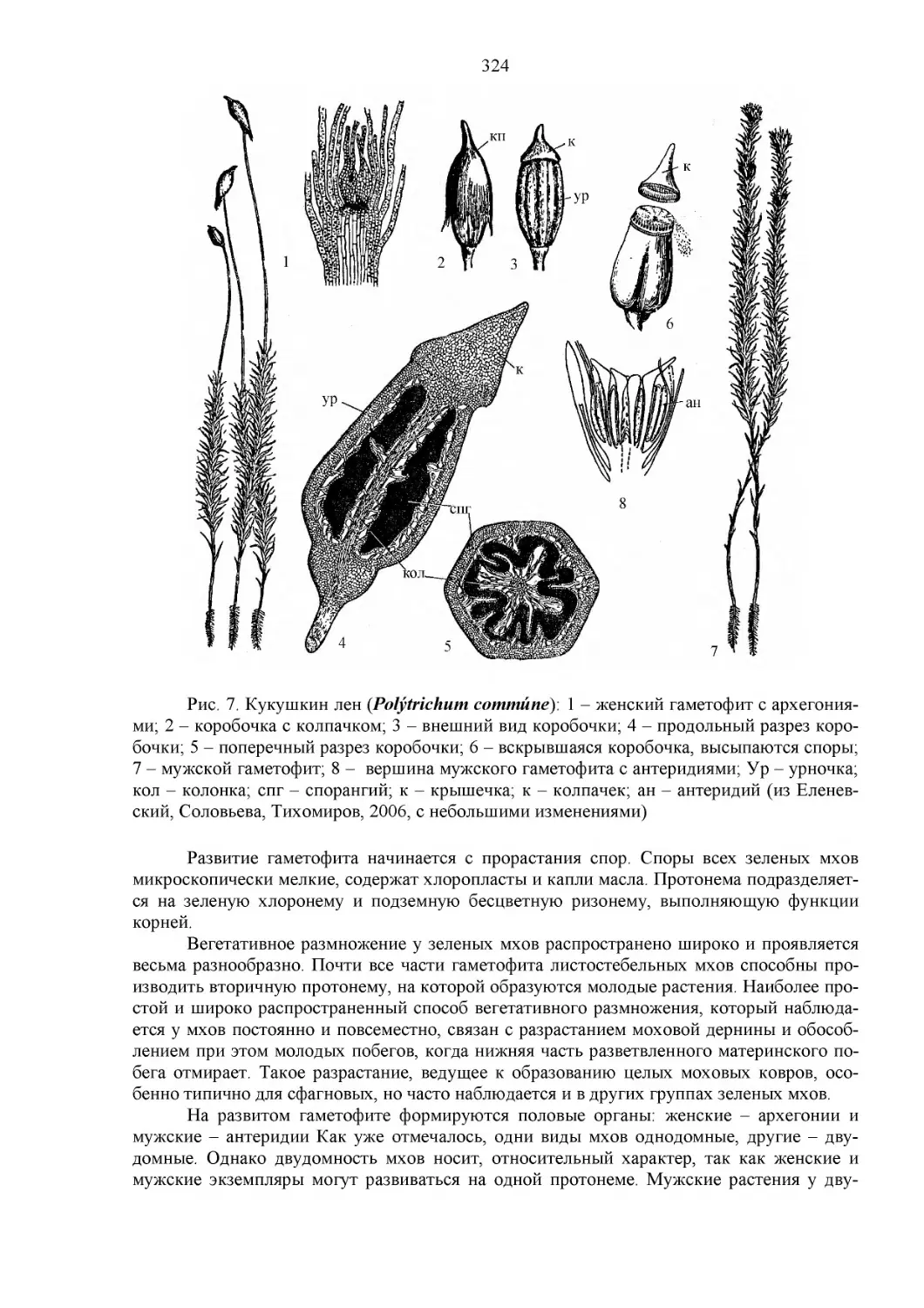
ОТДЕЛ МОХОВИДНЫЕ (BRIOPHYTA)........................................320
ОТДЕЛ ПЛУНОВИДНЫЕ (LYCOPHYTA).......................................325
ОТДЕЛ ХВОЩЕВИДНЫЕ (EQUISETOPHYTA)...................................325
ОТДЕЛ ПАПОРОТНИКООБРАЗНЫЕ (PTERIDOPHYTA)............................327
ОТДЕЛ ГОЛОСЕМЕННЫЕ (PINOPHYTA)......................................328
7
ОТДЕЛ ПОКРЫТОСЕМЕННЫЕ ИЛИ СНЕТКОВЫЕ РАСТЕНИЯ (ANGIOSPERMAE ИЛИMAGNOLIOPHYTA}....................331
СПИСОК ИСПОЛЬЗОВАННОЙ И РЕКОМЕНДУЕМОЙ ЛИТЕРАТУРЫ......335
ТЕРМИНОЛОГИЧЕСКИЙ СЛОВАРЬ.............................338
8
ВВЕДЕНИЕ
В программах обучения студентов по специальностям “Лесное хозяйство”, “Ландшафтное строительство” и “Лесоинженерное дело” дисциплина, соответствующая понятию “современная биология” отсутствует. Две первые программы предусматривают очень краткий курс ботаники, курсы почвоведения, лесной генетики, лесной фитопатологии, лесной энтомологии, краткий курс биологии лесных птиц и зверей и курс дендрологии. Перечисленные дисциплины, исключая в большинстве случаев дендрологию, традиционно включаются в учебники общей биологии, однако содержания учебников общей биологии они далеко не исчерпывают. Например, вопросы цитологии в лесных вузах рассматриваются слишком поверхностно, в курсе анатомии растений недостаточно уделено внимания камбию, строению одревесневших клеток стебля. В предусмотренных программами биологических дисциплинах совершенно отсутствует изложение ведущего вопроса современной биологии - теории эволюции, почти не уделяется внимания видообразованию, изучению зональности и лесорастительного районирования.
Настоящее учебное пособие предназначено восполнить имеющиеся в программах, входящих в компетенцию биологии, пробелы и способствовать восприятию студентами современной биологии в целом1.
Что же касается будущих специалистов лесоинженерного дела, то для них курс биологии введен сравнительно недавно - тринадцать лет это не большой срок, чтобы считать ту или иную дисциплину в числе традиционных для специалистов данного профиля. Однако появление этой новой для лесозаготовителей дисциплины явилось не случайностью, а вполне закономерным явлением. Сейчас представляется странным, почему курса биологии в программе лесоинженерной подготовки до сих пор не было? Ответ может быть таким - леса в России всегда было много, и дай-то Бог вырубить то, что доступно, и благополучно вывезти - остальное не столь важно - лес и сам растет! И так было до совсем еще недавнего времени. Однако производительность лесозаготовительной техники в последние десятилетия существенно увеличилась, соответственно возросли объемы лесозаготовок и площади вырубок. Стало очевидным, что лесная промышленность прогрессирует несравненно быстрее, чем живая природа способна адекватно реагировать на этот прогресс (здесь слово «прогресс» желательно поставить в кавычки).
Одним словом, рубить лес мы умеем гораздо лучше, чем его выращивать, а ведь лес - это «легкие нашей планеты»! И если лет 50 назад человечество (за исключением немногих, умевших смотреть вперед, ученых) еще не задумывалось о том, чем все живое на нашей планете будет дышать через несколько десятилетий, то сейчас этот страшный вопрос возник как апокалип
1 Настоящее учебное пособие не включает крупный раздел общей биологии - зоологию. Это связано в пер-вую очередь с общей ориентацией лесного образования на царство растений, а также с ограниченным объемом запланированного издания.
9
тический предвестник заслуженной расплаты за так называемый технический прогресс. Не будет леса или будет его недостаточно, и ближайшим нашим потомкам при настоящем темпе развития просто дышать будет нечем. И не только чистый воздух мы потеряем, но и лишимся многого того, без чего пока обходиться не можем, например, древесины, сенокосов, грибов, дикорастущих ягод, лекарственного сырья и многого другого. И вообще - может ли житель нашей страны представить весну без пения птиц, населяющих наши леса?
Но все это с одной стороны, с другой - лес нельзя не рубить, рубка спелого леса - это та же уборка урожая. Однако аналогия с сельским хозяйством здесь далеко не полная. В сельском хозяйстве тот, кто сеет или сажает, сам убирает урожай, он же заботится и о восстановлении плодородия почвы. А вот создателю лесных культур, особенно если речь идет об основных лесообразующих породах, редко удается дожить до «сбора урожая», то есть до рубки этого рукотворного леса. Ну а если и доживет, то «убирать урожай» будет не он, а совсем другие, не имевшие к посадке леса и уходу за ним никакого отношения. Что же касается восстановления плодородия почвы, то этот вопрос просто никак не рассматривался. Так было долгое время, но, будем надеяться, - не навсегда.
В последнее время при новой, еще не устоявшейся, системе ведения лесного хозяйства наблюдается тенденция передачи участков леса в аренду с условием, что арендатор после рубки и вывоза леса обязуется очистить и закультивировать лесосеку с соблюдением всех лесоводственных правил. Правила же ведения лесного хозяйства основываются на знании биологии.
Но не только лесовосстановление требует знания основ биологии, сами лесозаготовительные работы в процессе их планирования и осуществления должны проводиться на основе этих знаний. Например, расположение волоков или путей вывоза хлыстов и сортиментов без учета рельефа и почвенных условий может привести к эрозии почвы и тем самым предельно осложнить лесовосстановительные работы, а может быть и сделать их невозможными. Плохо очищенные от порубочных остатков лесосеки становятся причиной лесных пожаров и источниками распространения патогенных грибов и вредных для леса насекомых. Вместе с тем биологически грамотное проведение лесозаготовок позволит в очень короткие сроки превращать лесосеки в лесопокрытую площадь и вносить тем самым свой вклад в решение сложнейшей задачи «неистощительного пользования лесом».
Сказанное выше убеждает нас в том, что познание основ биологии, особенно разделов этой науки, имеющих отношение к жизни леса, должно быть фундаментом как лесохозяйственного, так и лесоинженерного образования.
Термин «биология» (слово, образованное двумя греческими словами: bios - жизнь и logos - учение, понятие) предложен в 1802 независимо друг от друга двумя учёными - французом Ж. Б. Ламарком и немцем Г. Р. Тревира-нусом. Современная биология представляет собой комплекс наук о живой
10
природе. В компетенцию этого комплекса входят все проявления жизни: строение и функции живых существ и их природных сообществ, их распространение, происхождение и развитие, связи друг с другом и с неживой природой. Задачи этой обширной и многоплановой науки состоят в изучении всех биологических закономерностей, раскрытии сущности жизни и её проявлений с целью познания и управления ими.
В настоящее время биология объединяет множество самостоятельных хорошо развитых, располагающих собственной методологией, наук. Можно привести неполный перечень этих наук с приблизительной хронологической последовательностью их возникновения.
Одними из первых сложились науки о животных - зоология и о растениях - ботаника, а также анатомия и физиология человека - основа медицины. В последствии возникли другие крупные разделы биологии, выделяемые по объектам исследования: микробиология - наука о микроорганизмах, гидробиология - наука об организмах, населяющих водную среду, и т.д. Внутри биологии сформировались более узкие дисциплины: в пределах зоологии - изучающие млекопитающих - териология, птиц - орнитология, пресмыкающихся и земноводных - герпетология, рыб - ихтиология, насекомых - энтомология, клещей - акарология, моллюсков - малакология, простейших - протозоология; внутри ботаники:- изучающая водоросли - альгология, грибы - микология, лишайники - лихенология, мхи - бриология, деревья и кустарники - дендрология и т.д. Подразделение дисциплин иногда идёт ещё глубже. Многообразие организмов и распределение их по группам изучают систематика животных и систематика растений. Биологию можно подразделить на неонтологию, изучающую современный органический мир, и палеонтологию - науку о вымерших животных (палеозоология) и растениях (палеоботаника).
Другой аспект классификации биологических дисциплин - по исследуемым свойствам и проявлениям живого. Форму и строение организмов изучают морфологические дисциплины. Образ жизни животных и растений и их взаимоотношения с условиями внешней среды - экология; изучение разных функций живых существ - область исследований физиологии животных и физиологии растений; предмет исследований генетики - закономерности наследственности и изменчивости; закономерности поведения животных -этология; закономерности индивидуального развития изучает эмбриология или в более широком современном понимании - биология развития; закономерности исторического развития - эволюционное учение.
Каждая из названных дисциплин делится на ряд более частных (например, морфология - на функциональную, сравнительную и др.). Одновременно происходит взаимопроникновение и слияние разных отраслей биологии, с образованием сложных сочетаний, например гисто-, цито- или эмбриофизиология, цитогенетика, эволюционная и экологическая генетика и др. Анатомия животных изучает строение органов и их систем макроскопически; микроструктуру тканей изучает гистология (в ботанике это анатомия), клеток -цитология, а строение клеточного ядра - кариология. В то же время и гисто
и
логия, и цитология, и кариология исследуют не только строение соответствующих структур, но и их функции и биохимические свойства.
Можно выделить дисциплины, связанные с использованием определённых методов исследования, например биохимию, изучающую основные жизненные процессы химическими методами и подразделяемую на ряд разделов (биохимия животных, растений и т.п.), биофизику, вскрывающую значение физических закономерностей в процессах жизнедеятельности и также подразделяемую на ряд отраслей. Сформировались дисциплины, изучающие живые объекты на разных уровнях их организации: молекулярная биология, цитология (клетки), гистология (ткани), популяционно-видовая систематика, биогеография, популяционные направления в генетике и экологии и так далее. Важное место в биологии занимают как теоретические, так и практические направления исследований, резкую границу между которыми трудно провести, т.к. любое теоретическое направление неизбежно связано (прямо или косвенно, в данный момент или в будущем) с выходами в практику.
Из перечисленного множества относящихся к биологии наук мы в данном курсе обратимся лишь к некоторым. В первой части учебника рассматривается ряд общих вопросов, относящихся к основам современной биологии, и частности из анатомии и морфологии растений, позволяющие студентам понять строение и функции лесообразующих растений. Во второй части изложены основы дендрологии с флористическим подходом, т.е. виды древесных растений-лесообразователей здесь подобраны не по систематическим признакам, а по представительству лесообразующих пород в том или ином регионе.
В настоящем учебном пособии, в его первой части, посвященной вопросам собственно общей биологии, допущена вынужденная односторонность - основное внимание уделено растениям, а биология животных практически не рассматривается. Это оправдывается ограничением объема данного издания, а также и спецификой учебных заведений, для которых и предназначено учебное пособие.
12
КРАТКИЙ ИСТОРИЧЕСКИЙ ОЧЕРК РАЗВИТИЯ БИОЛОГИИ
История биологии уходит своими истоками в древность. Надо полагать, что первые наблюдения живой природы и логические выводы из этих наблюдений были сделаны задолго до возникновения письменности. В глубокой древности представления о природе носили мистический и умозрительный характер хотя и были попытки получения сведений путем наблюдения. Биология же как наука в современном ее понимании берёт начало в странах Средиземноморья (Древний Египет, Древняя Греция). Потребность в знаниях о живой природе ограничивалась в основном сферой возделывания растений, разведения домашних животных и медициной. Так в Месопотамии (4-3 тысячелетие до ной эры) были известны (или выведены) культурные сорта растений (пшеница, ячмень, рожь, рис, плодовые, конопля, хлопок), выведены верховые породы лошадей и ослов, породы крупного рогатого скота и овец. Был известен прием получения мула путем скрещивания лошади и осла. Значит уже в те далекие времена имела место весьма результативная практическая селекция растений и животных.
Большой вклад в развитие биологии как науки, судя по сохранившимся письменным источникам, внесли Гиппократ (ок. 460 - ок. 370 до н. э.), Аристотель (384-322 до н. э.) и Гален (ок. 130- ок. 200). В средние века накопление биологических знаний диктовалось в основном интересами медицины. Растения изучались преимущественно в связи с их лекарственными свойствами. Вскрытия человеческого тела были запрещены, и преподававшаяся по Галену анатомия была в действительности анатомией животных, главным образом свиньи и обезьяны.
Первые ботанические труды средневековья были комментариями к сочинениям античных учёных: Теофраста, Диоскорида и Плиния Старшего. В дальнейшем появляются оригинальные «травники» - перечни лекарственных растений с их кратким описанием и изображением. Растения делили на деревья, кустарники и травы. Лишь итальянский ботаник А. Чезальпино сделал попытку (1583) создания классификации на основе строения семян, цветков и плодов. У Чезальпино имеются зачатки учения о метаморфозе2, а также впервые вводятся понятия «род» и «вид».
Примерно в то же время были составлены многотомные энциклопедии: «История животных» швейцарского учёного К. Геснера (1551-1587) и серия монографий в 13 томах (1599—1616) итальянского учёного У. Альдрованди. Появились описания «заморских» животных, основанные на наблюдениях в природе и на посещении далёких стран.
Блестящие успехи анатомии в эпоху Возрождения были связаны с внедрением анатомирования человеческого тела в практику как преподавания, так и исследования.
С созданием микроскопа (17 в.) возможности изучения живых существ расширились и углубились. Плеяда блестящих микроскопистов открывает клеточное строение растений (английский учёный Р. Гук (1665); итальянский
2 Метаморфоз - превращение одного в другое. Например, превращение листа в колючку.
13
- М. Мальпиги (1675-1679); английский - Н. Грю (1671-1782)); мир микроскопических существ исследуют голландец - А. Левенгук, (1673); Мальпиги (1669) изучает строение и развитие насекомых.
Примерно в это же время существенно уточняется строение тела многих животных, обнаруживается движение крови в капиллярных сосудах, устанавливаются половые различия у животных. Но одновременно возникают и многие заблуждения в роде теории преформизма, согласно которой в спермин (по мнению других - в яйцеклетке) содержится миниатюрный зародыш будущего организма.
В 17-начале 18 веков явно просматривается тенденция проникнуть в «план сотворения» живой природы, найти связи между отдельными группами организмов. Искусственные системы растений попытались построить английский учёный Дж. Рей, описавший в своей «Истории растений» (1686-1704) свыше 18 тыс. растений, сгруппированных в 19 классов, и французский ботаник Ж. Турнефор, распределивший их по 22 классам (1700). Дж. Рей определил на доступном в то время уровне знаний понятие «вид» и дал классификацию позвоночных, основанную на анатомо-физиологических признаках (1693).
Всеобъемлющую для своего времени «Систему природы» (1735), основанную на признании неизменности изначально сотворённого мира, предложил шведский натуралист Карл Линней (1707-1778). Свою систему растений, названную им «сексуальной», он построил, исходя из числа тычинок и других признаков цветков. Его классификация животных была в то время наиболее естественной и строилась с учётом их внутренних особенностей. Линней выделил класс млекопитающих, в который он правильно включил китов, а также человека, отнесённого им вместе с обезьянами к отряду приматов. Огромная заслуга Линнея - введение бинарной номенклатуры с двойным наименованием (по роду и виду) каждой формы растений и животных.
Искусственная система Линнея не удовлетворяла многих ботаников, пытавшихся найти «естественную» систему растений, в соответствии с их сходством и «сродством». Французский ботаник Б. Жюсьё осуществил её (1759) лишь в виде насаждений в Королевском саду в Трианоне (Версаль), а французский учёный М. Адансон пытался создать естественную систему семейств растений (1763). Завершил эти попытки французский ботаник А. Л. Жюсьё в своём труде «Роды растений, расположенные по естественным порядкам» (1789). Враждебную позицию по отношению ко всяким системам, в том числе и к системе Линнея, занял французский натуралист Ж. Бюффон. Его «Естественная история», 36 томов которой он успел опубликовать (1749-1788), включает описание не только животных и человека, но и минералов и историю прошлого Земли. Бюффон искал единства в плане строения животных, строил догадки о прошлом животного мира и пытался объяснить сходство близких форм их происхождением друг от друга. Научные поиски Бюффона были началом современных представлений об эволюции в мире живой природы.
В области физиологии дыхания много сделали английский учёный Дж. Пристли, показавший (1771-1778) в опытах на растениях, что они выделяют газ, способствующий горению и необходимый для дыхания животных, а также французские - А. Лавуазье, П. Лаплас и А. Сеген, выяснившие свойства кислорода в окислительных процессах и роль его в дыхании и образовании животного тепла (1787-1790). Роль солнечного света в способности зелёных листьев выделять кислород, используя углекислый газ из воздуха, установили голландский врач Я. Ингенхауз (1779), швейцарские учёные Ж. Сенебье (1782) и Н. Соссюр (1804). В конце 18 в. начинают широко изучать вещества, выделяемые из тканей животных и растений, закладывая тем самым основы будущей органической химии.
Немецкий ботаник ИГ. Кёльрёйтер (1733-1806), он же русский академик, окончательно доказал наличие пола у растений, а своими работами по гибридизации показал участие в оплодотворении и развитии как яйцеклеток, так и пыльцы растений (1761 и
14
позже). В конце века итальянский учёный Л. Спалланцани провёл точные опыты, опровергшие возможность самозарождения (до него считалось, что живое может в любое время зародиться из неживого). Здесь следует уточнить, что самозарождение, видимо, имело место, но процесс этот мог завершиться только на заре возникновения живой материи в археозойской эре примерно 3-3.5 миллиарда лет назад.
Идеи исторического развития органического мира всё настойчивее возникают во 2-й половине 18 в. Немецкий философ, математик, физик Г. В. Лейбниц (1646-1716), по приглашению Петра I работавший на благо России, провозгласил принцип градации живых существ и предсказал существование переходных форм между растениями и животными. Бюффон (1749, 1778) построил смелую гипотезу об истории Земли, длительность которой он исчислял в 80-90 тыс. лет и делил на 7 периодов; он утверждал, что лишь в последние периоды появляются на Земле растения, затем животные и, наконец, человек. Бюффон допускал превращение одних форм в другие под влиянием климата, почвы и питания. Французский физик Пьер Мопертюи (1750) обратил особое внимание на изменчивость организмов и высказывал догадки о роли элиминации форм, не приспособленных к существованию. Все это постепенно основывало фундамент для грядущего создания теории эволюции. Интересны умозрительные, но на удивление правильные для того времени, высказывания М.В. Ломоносова (1763) о развитии живой материи.
Вторая половина 18 века была буквально пропитана духом зарождения научно обоснованной теории эволюции жизни на Земле. Помимо упомянутых нами ране известных ученых, множество других, в разной степени известных или почти забытых биологов, философов и мыслителей, высказывали свои эволюционные идеи. Кроме фантастических или курьезных в теперешнем понимании измышлений, в то время были высказаны эволюционные воззрения, не потерявшие научной ценности и в наши дни.
В числе если не забытых, то, по крайней мере, мало упоминаемых, был английский врач, поэт и натуралист Эразм Дарвин (1731-1802), дед столь известного Чарльза Дарвина, опубликовавший научный трактат «Зоономия» (1794-1796). Этот трактат явился по-существу синтезом накопившихся к тому времени знаний в области эволюции живой природы. Он впервые сказал о «миллионах веков» эволюции жизни вместо 80-90 тыс. лет по Бюффону, отметил существование борьбы между близкими видами и наследование полезных уклонений от нормы. Он предвосхитил основные научные выводы своего внука. Современники и потомки не придали должного значения трудам Э. Дарвина потому, что считали его не ученым, а поэтом, он и научные труды создавал в виде поэм.
В 19 веке французский учёный Ж. Б. Ламарк (1744-1829) эволюционно истолковал «лестницу существ», нарисовав в «Философии зоологии» (т. 1-2, 1809) путь совершенствования живых существ от низших к высшим, совершающийся на основе внутреннего, присущего организмам стремления к прогрессу (принцип градации). Внешняя среда, согласно его представлениям, вызывает отклонения от «правильной» градации и определяет приспособление видов к условиям существования либо прямым воздействием (растения и низшие животные), либо через упражнение и неупражнение органов в связи с изменением привычек (животные с нервной системой).
15
Теорию Ламарка критиковали многие, в том числе основоположник сравнительной анатомии и палеонтологии животных французский учёный Ж. Кювье (кстати, Ламарка во многом несправедливо критикуют и в наши дни).
Для объяснения исторической смены живых форм и исчезновения многих из них французский зоолог Ж. Кювье (1769-1832) выдвинул учение о катастрофах, претерпеваемых органическим миром под влиянием геологических катаклизмов (1825). Многое из этой теории не отвергается и в наше время.
Французский учёный Э. Жоффруа Сент-Илер (1772-1844) основал учение о «единстве плана строения» животных, которое он в дальнейшем объяснял общностью их происхождения. По его представлениям, эволюционные изменения происходят внезапно в результате прямых воздействий внешней среды; особенно резкие изменения претерпевают животные в эмбриональный период (последнее утверждал и Ламарк). Эти идеи нашли отражение и дальнейшее развитие во взглядах русского учёного, хранителя зоологического музея Московского университета К.Ф. Рулье (1814-1858), значительно углубившего их. Что же касается Жоффруа Сент-Иллера (Жоффруа не имя, а первая фамилия), то он помимо прочего является автором первого учебника общей биологии (1840) и родоначальником науки об уродствах.
Наибольшее влияние на биологию натурфилософского3 направления, корни которой уходят в начало 18 в., оказали ученые Германии. Немецкие философы и естествоиспытатели также обосновывали учение о единстве «плана строения» организмов. Так, немецкий писатель, великий поэт, мыслитель и естествоиспытатель И. В. Гёте (1749-1832) утверждал существование «идеи органа» и типов «прарастения» и «праживотного» (1782-1817); Л. Окен считал, что в основе строения и развития всех живых существ лежит «пузырёк» или инфузория (1805). Наиболее плодотворной идеей немецких натурфилософов был принцип параллелизма между онтогенезом4 и филогенезом5 (К. Кильмейер, 1793; И. Меккель, 1811), ставший впоследствии отправной точкой при формулировке биогенетического закона. Существо этого закона было сформулировано в 1866 г. Э. Геккелем, и состоит оно в том, что онтогенез всякого организма есть сжатое и краткое повторение филогенеза данного вида.
Идея развития организмов нашла подлинное научное подтверждение в эмбриологических исследованиях русских академиков X. И. Пандера (1817) и К. М. Бэра (1827), в обосновании Бэром принципов сравнительной эмбриологии (1828-1837) и в создании немецким биологом Т. Шванном (1839) единой для всего органического мира клеточной теории. Учение о единстве клеточного строения всех живых существ сыграло огромную роль в развитии биологии.
Большие успехи были достигнуты в середине 19 в. в области физиологической химии, главным образом, благодаря трудам немецкого учёного, члена-корреспондента Российской АН, Ю.Либиха (1803-1873) и французского - Ж. Б. Буссенго (1802-1887), которые установили особенности питания растений и его отличие от питания животных, сформулировав принцип круговорота веществ в природе. Либих разделил все вещества, входящие в состав живых существ, на белки, жиры и углеводы, выяснил многие химические процессы обмена веществ, в том числе образование жиров из углеводов.
Работы французского учёного Л. Пастера (раскрытие роли микроорганизмов в процессах брожения (1857-1864)), имевшие выдающееся значение для медицины, пищевой промышленности и сельского хозяйства, позволили окончательно опровергнуть учение о самозарождении организмов (1860-1864). В дальнейшем он показал роль микроорганиз
3 Натурфилософия - философия природы, умозрительное (без эксперимента) истолкование природы в ее целостности.
4 Онтогенез - индивидуальное развитие, совокупность изменений, претерпеваемых организмом от зарождения до конца жизни.
3 Филогенез - процесс исторического развития мира организмов, их видов, родов, семейств, порядков, клас-ссов. отделов, царств.
16
мов в инфекционных заболеваниях животных и человека, разработал меры борьбы против бешенства и сибирской язвы с помощью защитных прививок. Природу процессов брожения, вызывавшую споры между сторонниками физико-химического (Либих) и микробиологического (Пастер) её объяснения, окончательно раскрыл немецкий учёный Э. Бухнер, выделив из дрожжевых грибов фермент зимазу (1897). Этим было положено начало новой науке о ферментах - энзимологии. Русский врач Н.П. Лунин доказал (1881) наличие в пищевых продуктах витаминов, позже названных так польским учёным К. Функом (1912). В конце 19 в. были достигнуты первые успехи в изучении химии белков и нуклеиновых кислот. Принципиальное значение для установления круговорота азота, серы и железа в природе имело обнаружение русским микробиологом С. Н. Виноградским (1887-1891) бактерий, способных образовывать путём хемосинтеза (открытого Виноградским) органические вещества из неорганических. Основоположник вирусологии Д. И. Ивановский открыл новую форму организации живого - вирусы (1892).
Крупнейшим событием 19 века был выход с свет в 1859 году всемирно известной книги Чарльза Дарвина (1809-1882) «Происхождение видов...». По значению для человечества и популярности ее ставили на второе место после Библии. Дарвин дал опирающееся на собственные наблюдения и огромное число фактов из биогеографии, палеонтологии, сравнительной анатомии эмбриологии обобщенное доказательство эволюционного развития органического мира.
Основная идея «Происхождения видов» состоит в том, что внутривидовая изменчивость организмов и численная избыточность потомства создает обширное поле деятельности для естественного отбора. Естественный отбор элиминирует (исключает) из размножающейся части популяции6 наименее приспособленные к конкретным условиям окружающей среды особи. Успешно размножаются только приспособленные. Среда с течением времени меняется, соответственно меняются и виды.
Учение Дарвина внедрило в мышление биологов исторический подход ко всем явлениям жизни. Это способствовало разработке ряда новых направлений в биологии: в эволюционной сравнительной анатомии (немецкий учёный К. Гегенбаур (1826-1903), эволюционной эмбриологии (русские биологи А. О. Ковалевский (1840-1901), И. И. Мечников (1845-1916)), эволюционной палеонтологии (В. О. Ковалевский (1842-1883)). На этой же основе был сформулирован биогенетический закон (немецкие учёные Ф. Мюллер, 1864; Э. Геккель, 1866 и позже) и разработан ряд филогенетических обобщений. С развитием эволюционного учения огромный размах получили зоо- и фитогеография (английские учёные Ф. Склетер (1829-1913) и А. Уоллес (1823-1913), русские - А.Н. Северцов (1866-1936) и А. Н. Бекетов (1825-1902), немецкие - А. Гризебах (1814-1879) и А. Энглер (1844-1930), датский - Э. Варминг (1841-1924) и мн. др.). Кстати, А. Уоллес одновременно с Ч. Дарвином и независимо от него, на одном и том же заседании Королевского ботанического общества, изложил собственную, принципиально совпадающую со взглядами Дарвина, теорию естественного отбора. Большую роль в пропаганде дарвинизма сыграли в Англии Т. Гексли (1825-1895), в Германии Э.
6 Популяция - совокупность особей одного вида, обладающих общим генофондом и занимающих опреде-ленную, не разобщающую их, территорию. Генофонд - совокупность генов, которые имеются у особей данной популяции или вида.
17
Геккель (1834-1919). В России крупнейший вклад в пропаганду и развитие эволюционной теории внесли К. А. Тимирязев (1843-1920) и целая плеяда сравнительных анатомов, эмбриологов, палеонтологов.
Учение о естественном отборе быстро получило самое широкое признание. Однако невыясненность закономерностей изменчивости и наследственности служила источником расхождений в толковании факторов эволюции. К концу 19 века возникли различные направления неодарвинизма, неоламаркизма, а также новые научные направления, не признающие ведущей роли естественного отбора в процессе эволюции.
Попытки раскрыть механизмы наследственности умозрительно, предпринятые многими учеными, в том числе и Ч. Дарвином, не увенчались успехом. Лишь чешскому монаху и одновременно ученому-биологу Г. Менделю удалось установить основные закономерности наследственности (1865). Однако его работа сначала осталась незамеченной, и лишь успехи цитологии и эмбриологии подготовили её переоткрытие (1900) и правильную оценку в 20 в. Первым шагом в этом направлении было раскрытие тонких процессов распределения хромосом при клеточном делении - митозе (французский биолог А. Шнейдер, 1873; русский - И. Д. Чистяков, 1874; польский - Э. Страсбур-гер, 1875; немецкий - В. Флемминг, 1882, и др.). Далее были выяснены процессы оплодотворения, созревания гамет и явление редукции хромосом сначала у животных (немецкий биолог О. Гертвиг, 1875; бельгийский - Э. ван Бенеден, 1875-1884; немецкий - Т. Бовери, 1887-1888), а затем и у растений (русский - И. Н. Горожанкин, 1880-1883; русский - С. Г. Навашин, 1898; французский - Л. Гиньяр, 1899).
20 век характеризуется развитием новых биологических дисциплин и подъёмом исследований в «классических» отраслях биологии, в том числе на основе дальнейшей специализации или интеграции старых разделов. Особенно интенсивно развиваются в 20 веке физиология животных и растений, биохимия, эмбриология, цитология, генетика, иммунология, эволюционное учение, экология, учение о биосфере, а также микробиология, вирусология, паразитология и многие другие отрасли биологии.
Отправным пунктом развития генетики стал менделизм, подкрепленный рядом обобщений, в том числе мутационной теорией голландского учёного X. де Фриза (1901— 1903), сыгравшей, несмотря на ошибочность многих положений, важную роль в подготовке синтеза генетики и теории эволюции. Были разработаны понятия ген, генотип, фенотип (датский учёный В. Иогансен, 1909), обоснована хромосомная теория наследственности (американские учёные Т. X. Морган (1866-1945), А. Стёртевант (1891-1970) и др.). Важное методологическое значение приобрёл вопрос о причинах возникновения наследственных изменений - мутаций. Доказательства влияния на мутационный процесс физических, а затем и химических факторов (русские учёные Г. А. Надсон и Г. С. Филиппов, 1925, В. В. Сахаров, 1932, и др. и особенно американские учёные Г. Дж. Мёллер, 1927, Л. Стедлер, 1928, и др.) опровергли автогенетические7 концепции генетиков, подчёркивавших исключительно самопроизвольный характер возникновения мутаций.
На основе внедрения в биологию методов физики, химии, математики и др., а также успехов в области познания структуры белков, закономерностей их синтеза, передачи
7 Автогенез - учение, объясняющее эволюцию организмов действием только внутренних факторов.
18
и осуществления наследственных факторов расширяется круг исследований на молекулярном уровне. Расшифрована последовательность расположения аминокислот свыше чем в 200 белках, выяснены их вторичная структура и способ укладки полипептидных нитей в молекуле белка.
Значительные успехи достигнуты в физиологии и биохимии животных: учение об условных рефлексах, разработанное И. П. Павловым (1849-1936); бурное развитие нейрофизиологии; изучение физиологии и биохимии мышечного сокращения; выделение и всестороннее исследование ферментов, определяющих направление и скорость различных процессов биосинтеза, и осуществление с их помощью синтеза гормонов (инсулин и др.), витаминов, ферментов (рибонуклеаза и др.) и иных биологически активных веществ. Физиология растений добилась успехов в познании химии фотосинтеза, в изучении участвующих в нём пигментов и прежде всего хлорофилла, который удалось искусственно синтезировать. Заметными были успехи в изучении роста и развития растений, например выделены и частично синтезированы некоторые гормоны роста (ауксины, гиббереллины, ретарданты и др.).
Многие исследования, в том числе и русских биологов, имели не только теоретические, но и важное прикладное значение, например для медицины или сельского хозяйства. Таковы учение о трансмиссивных заболеваниях и природной очаговости Е. Н. Павловского (1884-1965), капитальные труды по паразитологии В. А. Догеля (1852-1922), В. Н. Беклемишева (1890-1962) и К. И. Скрябина (1878-1972), закон гомологических рядов в наследственной изменчивости и учение о центрах происхождения культурных растений Н. И. Вавилова(1887-1943)и мн. др.
В 1920-1930-х гг. возникло существенное оживление в развитии теории эволюции - был осуществлен синтез дарвинизма и генетики. Начало этому положили многочисленные исследования в области генетики популяций (русские ученые: Н.К. Кольцов (1872- 1942), Ю.А. Филипченко (1882-1930), С. С. Четвериков (1915, 1926); русский, уехавший в 1927 году в США, Ф.Г. Добржанский (1900-1975); американский - С. Райт (1921-1932); английские - Дж. Б. С. Холдейн (1924-1932) и Р. Фишер (1928-1930)). Работами перечисленных исследователей и их коллегами были выявлены «факторы микроэволюционных процессов», таких как мутации, отбор, изоляция, миграция, дрейф генов и др. Правда, в дальнейшем стало понятным, что в этих случаях правильнее вести речь не об эволюционных изменениях, а об изменении частоты аллелей8.
Изучение генетики популяций способствовало разработке теории факторов эволюции (И. И. Шмальгаузен (1884-1963), Н.В. Тимофеев-Рессовский (1900-1981) и др.), обоснованию главных типов эволюции и вычленению из них в качестве основных - ароморфо-зов9 и ид иоадаптаций10 (А. Н. Северцов (1866-1936)), развитию представлений о темпах и формах эволюции.
Большие успехи достигнуты в изучении закономерностей образа жизни организмов и их связи со средой обитания, т. е. в экологии как особей и популяций, так и сложных сообществ (биоценозов и экосистем). Выявлены закономерности связи условий среды с распределением организмов в пространстве и времени; особенности сложной структуры популяций и биоценозов; факторы, определяющие динамику численности популяций, и дру
8 Аллель - одно из возможных состояний гена в данном локусе. Например: доминантный аллель или рецессивный аллель; дикий (неизмененный) аллель или мутантный аллель.
9 Ароморфоз - усложнение организации и функций организмов в процессе их эволюции.
10 Идиоадаптация - частное приспособление к условиям существования.
19
гие фундаментальные зависимости. Созданы концепции трофических11 уровней, цепей питания, жизненных форм, экологических ниш, биологической продуктивности и связанных с ней понятий и представлений. Крупнейшим достижением биологии является создание советскими учёными В.И. Вернадским (1863-1945) биогеохимии и учения о биосфере (1926) и В.Н. Сукачевым (1880-1967) - биогеоценологии, которые составят научную основу взаимоотношений человечества со средой своего обитания - биосферой Земли. Кстати, В.Н. Сукачев был первым заведующим кафедрой дендрологии и систематики в Московском лесотехническом институте.
Развитие упомянутых и, других важных направлений современной биологии было подготовлено в мире и в нашей стране научной деятельностью множества выдающихся биологов, имена которых просто невозможно перечислить в этом кратком экскурсе в историю биологии.
Однако развитие биологии в нашей стране отмечено не только периодами успехов и открытий. В науку грубо и бестактно вмешивалась политика. В 1936 и 1939 имел место ряд острых дискуссий по методологическим проблемам теоретической биологии. В ходе этих дискуссий путем «давления сверху» была практически запрещена генетика и некоторые направления теории эволюции. Группа учёных, возглавляемая Т.Д. Лысенко и поддерживаемая представителями власти, отстаивала ошибочные, механистические взгляды на природу наследственности, видообразования, естественного отбора, органической целесообразности и др. Эти взгляды были декларированы как развитие научного наследия выдающегося советского селекционера И. В. Мичурина (в действительности Мичурин не имел к этому отношения) и названы «мичуринской биологией» и «творческим дарвинизмом». После печально известной сессии ВАСХНИЛ (1948) обстановка особенно обострилась, исследования ряда направлений общей биологии полностью прекратились. Всё это создало почву для распространения непроверенных фактов и гипотез (учение о неклеточном «живом веществе», скачкообразное «порождение» видов, «превращение» вирусов в бактерии и др.). Отрицательную роль сыграли также дискуссии по физиологии (Объединённая сессия АН и АМН СССР, 1950), по эволюционной морфологии (1953). Всё это сильно затормозило развитие в СССР генетики, эволюционного учения, цитологии, молекулярной биологии, физиологии, эволюционной морфологии, систематики и других отраслей биологии.
Постепенная нормализация положения началась в октябре 1964, когда были предприняты меры по восстановлению и развитию современного генетического и других направлений (созданы соответствующие институты, организовано Всесоюзное общество генетиков и селекционеров, усилена подготовка специалистов в этих областях).
В это же время в развитии биологии происходили весьма важные события. Американский физик и химик, иностранный член РАН, Л.К. Полинг заложил основы структурного анализа белков (1946-1950) и предложил модель спирального строения полипептидной цепи. Основываясь на открытиях Полинга, в 1953 году американец Д.И Уотсон совместно с англичанином Ф. Криком выяснили пространственную организацию молекулы ДНК, что позволило определить основные функции и свойства этой молекулы - носителя генетической информации. Открытие Уотсона и Крика, удостоенное Нобелевской премии, повлекло за собой ряд крупнейших открытий во многих областях современной биологии.
11 Трофия от греч. питание. Трофический уровень - совокупность организмов, объединяемых типом пита-
НИЯ.
20
Во второй половине XX века и начале XXI основные достижения отмечены в области эволюции органического мира, в молекулярной биологии, иммунологии, генетике.
Буквально до последнего времени бурно развивалась популяционная биология или генетика популяций, изучающая так называемые микроэволюционные процессы (мутации, изоляцию, миграции, разные типы изменчивости, отбор). Исследования проводились с модельными популяциями животных и растений в лабораториях, иногда в полевых условиях. Результаты этих работ привели Ф. Добржанского к убеждению, что «эволюция есть изменение генетической структуры популяций». По генетике популяций накопилась громадная по объему труднообозримая литература (Ф. Айала, 1984; Ю.П. Алтухов, 1983; Н.П. Дубинин и Я.Л. Глембоцкий, 1967; Р. Левонтин, 1978; Л. Меттлер и Т. Грегг, 1972; Н.В. Тимофеев-Рессовский, А.В. Яблоков и Н.В. Глотов, 1973; С.С. Шварц, 1980 и многие другие широко известные публикации).
Популяционно-генетические и селекционные исследования проводились и с лесными древесными растениями (Б. Линдквист, 1954; Линдгрен с соавторами, 1997, 1999 и др; Д.; Намкунг, 1981; Дж Райт, 1978; Э. Ромедер, Г. Шенбах, 1962 и другие). Важную роль в этом направлении исследований сыграли отечественные «лесные» селекционеры и генетики (В.Н. Сукачев, 1934, 1939, 1952; Л.Ф. Правдин, 1964, 1975; А.В. Альбенский, 1969; М.М. Вересин, 1946, 1963 и др.; В.А. Драгавцев, 1997; А.И. Ирошников, 1974, 1978 и др.; Ю.Н. Исаков; 1991, 1999; С.А. Мамаев, 1973; А.Е. Проказин, 1996 и др.; В.М. Роне, 1980; А.П. Царев, 1985, 1998) в их числе сотрудники МЛТИ (МГУЛа): А.Я. Любавская, 1978, 1982; С.П. Погиба, 1988, 2000, 2002; М.Г. Романовский, 1994, 1997, 2002; А.В. Чудный, 1981; А.С. Яблоков, 1949, 1952).
Изучение популяций позволило глубже понять структуру вида, его подвижность во времени и сложную организацию. Однако объяснить процессы макроэволюции с позиций популяционной генетики не удалось, так как микроэволюционные процессы сводятся в основном к изменению частоты генов в популяциях и не могут служить точной моделью процессов видообразования.
В настоящее время можно с уверенностью говорить, что крупнейшие завоевания биологии конца XX и начала XXI веков лежат в русле углубления наших представлений о процессах эволюции живой материи. Большой вклад в понимание этих процессов внесли иммунология и иммуногенетика - науки об иммунитете у животных и растений. Иммунологами в частности было установлено, что имеет место так называемый горизонтальный перенос генов (перенос генов между организмами не связанный с процессом размножения, т. е. с вертикальным переносом).
С открытием горизонтального переноса генов между разными видами и даже царствами живых организмов (в последнее время получено множество доказательств такой возможности) ситуация изменилась, и эволюция предстает в совершенно ином свете. Получается, что любое "удачное изобретение" одного из видов становится доступным и может быть заимствовано всеми остальными. Биосфера теперь представляется единой информационной средой, в которой вирусы и различные мобильные генетические элементы распространяют информацию примерно так же, как в человеческом обществе благодаря устной и письменной речи, а в последнее время и благодаря интернету, достижения и открытия одних людей становятся известными другим и могут ими использоваться (http://www.macroevolution.narod.ru/latgentrans.htm).
21
Обобщая сказанное выше, можно выделить важнейшие вехи в истории биологии: принципы классификации живой природы (К. Линней), клеточное строение живых организмов (Т. Шванн и М. Шлейден), теория естественного отбора (Ч. Дарвин), законы наследственности и наследования (Г. Мендель), хромосомная теория наследственности (А. Вейсман и Т.Г. Морган), расшифровка двойной спирали ДНК (М. Вилкинс, Дж. Уотсон, Ф. Крик) и генетического кода (М. Ниренберг), пространственной структуры белка (Дж. Кендрью и М. Перутц), последовательности аминокислот в молекуле белка (Ф. Сенгер), регуляции биосинтеза белка (Ф. Жакоб и Ж. Моно) и явления обратной транскрипции - переноса информации от РНК к ДНК (Г. Темин и Д. Балтимор). К этому следует добавить и обобщения о биосферной организации жизни (В.И. Вернадский, В.Н. Сукачев)), а также формирование популяционного мышления и учения о микроэволюции (Т. До-бржанский, Н.В. Тимофеев-Ресовский, Н.И. Вавилов, Дж. Хаксли, Э. Майр, Дж. Холдейн).
Биология продолжает бурно развиваться, и в мире науки и практики у нее, вне сомнения, великое будущее.
КЛЕТОЧНОЕ СТРОЕНИЕ ЖИВЫХ ОРГАНИЗМОВ
Химический состав клетки
Клетки организмов содержат около 90 химических элементов, значение для жизни 24 из них точно установлено. Роль остальных пока неизвестна. По доле участия в строении организмов химические элементы можно разделить на три группы.
В первую группу входят О (65-70 %), С (15-18 %), Н (8-10 %) и N (1,5-3 %). Перечисленные элементы составляют около 97 % массы условно усредненной живой клетки и их называют макроэлементами. Макроэлементы входят в состав белков, углеводов, липидов и множества других органических веществ.
Вторую группу составляют Р, S, С1 и металлы: К, Са, Mg, Na и Fe. На их долю приходится около 3 % массы клетки. Магний является ядром молекулы хлорофилла; железо - ядром молекулы гемоглобина; сера и фосфор вместе с макроэлементами вносят основной вклад в построение основных жизненно важных органических соединений, поэтому их называют биоэлементами.
Третья группа объединяет остальные элементы. Их не более 0,1 % вещества клетки, однако некоторые из них, присутствуя в живой клетке в тысячных долях процента, относятся к жизненно важным. Так, цинк входит в состав более, чем ста различных ферментов; соединения меди и бора необходимы для нормального развития растений. Медь, кроме того, является ядром молекул вещества (гемалимфы), выполняющего у насекомых функции крови. Роль некоторых элементов третьей группы пока еще не выявлена.
Организмы обладают способностью избирательно поглощать из окружающей среды и накапливать в себе определенные вещества. Так кислорода
Т1
в почве содержится 49 %, а в растениях 70 %; кремния в почве - 33 %, а в растениях - 0,15 %. В почве и водоемах очень мало ионов К и много ионов Na в клетке же, наоборот.
Все химические соединения, присутствующие в организмах, образуют два больших класса: неорганические и органические вещества.
Неорганические вещества. Самое распространенное неорганическое вещество в живых существах - вода. В многоклеточных организмах вода составляет до 80 % массы. Она обладает своеобразными, и до сих пор не до конца изученными, физико-химическими свойствами. Каждая молекула воды связана с несколькими соседними такими же молекулами водородными связями. В результате формируется сложная сеть связанных молекул. Водородные связи не позволяют молекулам воды независимо двигаться, поэтому вода при температуре от 0-100 °C - жидкость, а не газ. Значительное сцепление молекул воды между собой и с молекулами других веществ позволяет воде легко перемещаться вверх по проводящим элементам ксилемы и переносить в виде растворов питательные вещества.
Вода обладает высокой теплоемкостью и выполняет в организмах терморегулирующую функцию. При испарении через устьица и с поверхности растений вода защищает последние от перегрева.
Будучи хорошим растворителем, вода способствует поступлению необходимых для жизни веществ в клетки в виде растворов. Молекулы воды электростатическим воздействием расщепляют молекулы различных соединений на анионы и катионы и тем самым обеспечивают протекание многих реакций, свойственных живым клеткам.
Перечисленные свойства воды позволяют предполагать, что жизнь на нашей планете возникла в водной среде.
Большая часть прочих неорганических соединений присутствуют в организмах в виде растворимых и нерастворимых солей, роль которых в метаболизме живых существ весьма разнообразна.
Органические вещества. К важнейшим для жизни органическим веществам относятся белки, нуклеиновые кислоты, углеводы и жиры. В составе органических соединений обязательно присутствует углерод.
Белки представляют собой полимерные соединения, состоящие из аминокислот12. Молекулы белков можно представить в виде очень длинных цепей, каждое звено которых является аминокислотой. Молекулярный вес разных белков колеблется в больших пределах и достигает десятков тысяч а.е.м.13. По строению и свойствам белки очень многообразны, но все они образованы двадцатью аминокислотами. При этом одни белки содержат все 20 аминокислот, другие - несколько меньше, есть содержащие менее половины. Общее же число аминокислот (с повторностями) в одной крупной молекуле белка может достигать 300-500. Свойства белков определяются последова
12 Аминокислоты - органические соединения, в молекуле которых одновременно содержатся карбоксильные и аминные группы.
13
а.е.м. - атомная единица массы
23
тельностью и повторяемостью аминокислот в "цепочке", а также структурой (пространственной организацией молекул).
Структура белков сложна и для каждого белка специфична. Несколько упрощая действительность, форму молекулы белка можно представить в виде цепочки аминокислот, которая скручена в спираль, а спираль в свою очередь уложена и скручена в клубок или глобулу. Есть белки, состоящие из нескольких глобул.
Белки в клетках организмов выполняют множество очень важных функций. Они входят в состав так называемых биологических мембран14; являются ферментами - катализаторами метаболических процессов; регулируют функции клеток, выступая в роли гормонов; осуществляют двигательные функции; участвуют в активном переносе веществ сквозь клеточные мембраны; оповещают организм об изменении внешних воздействий; играют решающую роль в проявлении иммунитета; выполняют запасающую функцию у растений.
Углеводы - органические соединения, в состав которых кислород и водород входят в тех же пропорциях, что и в состав воды (Cn(H2O)m), отсюда и название. В растительных клетках доля углеводов может составлять до 2/3 сухой массы, в животных - значительно меньше. В организмах они выполняют энергетическую, запасающую и строительную функции. Углеводы подразделяют на моносахариды (простые сахара) и полисахариды (сложные сахара).
Моносахариды - бесцветные сладкие вещества, хорошо растворяются в воде. Их общая формула СП(Н2О)П, п = 3+9. Моносахариды с п > 4 могут образовывать циклические формы. Наиболее важная из пентоз (п = 5), рибоза, входит в состав рибонуклеиновой кислоты (РНК), а дезоксирибоза - в состав дезоксирибонуклеиновой кислоты (ДНК). Большое значение в энергетическом снабжении организмов имеют гексозы (п = 6): глюкоза, фруктоза и галактоза.
Полисахариды образуются путем соединения моносахаридов в линейные или разветвленные полимеры так называемой гликозидной связью. Самыми простыми полисахаридами являются дисахариды. Наиболее известна сахароза - тростниковый или свекловичный сахар. В ее состав входят остатки двух простых сахаров - глюкозы и фруктозы. Сахароза является первичным продуктом фотосинтеза и транспортной формой углеводов в растениях.
С увеличение числа полимерных звеньев растворимость полисахаридов уменьшается, сладкий вкус пропадает. Самыми важными полисахаридами, составленными из молекул глюкозы, являются гликоген, крахмал и целлюлоза.
Полисахарид - крахмал в клетках растений, гликоген в клетках животных и грибов являются источниками глюкозы и выполняют функции основных запасающих веществ.
14 Биологические мембраны - белково-липидные структуры молекулярных размеров, до 100 А толщиной, образующие плазматическую мембрану (плазмалемму) и ограничивающие внутриклеточные образования (ядра, митохондрии, пластиды и пр.).
24
Целлюлоза или клетчатка тоже представляет собой полисахарид на основе глюкозы. Это высокополимерное соединение, до 10 тыс. остатков. Целлюлоза обладает высокой химической стойкостью и механической прочностью. Волокно целлюлозы превосходит по прочности стальную проволоку того же диаметра. Древесина - почти чистая целлюлоза, из нее построены стенки (оболочки) клеток растений.
Липиды - разнообразные по строению жироподобные вещества, почти нерастворимые в воде. Наиболее распространенные липиды: воски, нейтральные жиры, фосфолипиды и стероиды.
Воски - сложные эфиры жирных кислот и спиртов - покрывают листья и побеги многих растений. Они защищают от ультрафиолета и механических повреждений; регулируют водный баланс, не пропуская внутрь лишнюю воду. Пчелы и шмели используют воск из своих восковых желез для постройки сот.
Нейтральные жиры - соединения, к классу которых относятся большинство животных и растительных жиров. Основная функция жиров - энергетическая. При расщеплении жиров на СО2 и Н2О выделяется вдвое больше энергии, чем при расщеплении углеводов. Однако энергия углеводов выделяется быстрее, так что последние являются не "стратегическим", а "оперативным" запасом.
При метаболизации 1 кг жира образуется более 1 литра воды. Благодаря запасам жира в горбах верблюды могут не пить 10-12 суток, а медведи, ежи, сурки во время зимней спячки обходятся без воды несколько месяцев. У многих растений жиры содержатся в семенах, они необходимы проростку в качестве начального источника энергии.
Фосфолипиды входят в состав клеточных мембран. По строению они сходны с нейтральными жирами, только вместо одной из жирных кислот - у них остаток фосфорной кислоты.
Стероиды выполняют в организмах регулирующие функции, к ним относятся некоторые гормоны, витамин D, а также холестерин - компонент мембран клеток животных.
Нуклеиновые кислоты ДНК и РНК. Нуклеиновые кислоты хранят наследственную информацию организмов, так что жизненную важность этих соединений трудно переоценить. Дезоксирибонуклеиновая кислота (ДНК) находится в ядрах клеток и небольших количествах - в митохондриях и хлоропластах (об этих органеллах в дальнейшем изложении).
ДНК состоит из двух параллельных полимерных цепей, скрученных в двойную правозакрученную спираль (одна вокруг другой и обе вокруг общей оси. Каждая цепь ДНК - полимер, мономерами которого являются нуклеотиды (до 100 млн. нуклеотидов). В составе нуклеотида - одно из четырех азотистых оснований: аденин (А), тимин (Т), гуанин (Г) или цитозин (Ц), а также моносахарид дезоксирибоза и остаток фосфорной кислоты.
Четыре вида нуклеотидов различаются только азотистыми основаниями. В полинуклеотидной цепи ДНК молекулы дезоксирибозы связаны прочными ковалентными связями с остатками фосфорной кислоты соседних нук
25
леотидов. Две цепи ДНК соединяются друг с другом посредством водородных связей между азотистыми основаниями.
В соответствии со строением молекул азотистых оснований наблюдается следующая закономерность: основание А связывается только с Т, а Г -только с Ц. Нуклеотиды как бы дополняют друг друга, образуя так называемые комплементарные пары: А-Т и Г-Ц.
Если известен порядок следования нуклеотидов в одной цепи ДНК, то по принципу комплементарности можно установить порядок нуклеотидов в другой цепи. Комплементарность цепей позволяет молекуле ДНК точно реплицироваться (от лат. replicatio - повторение) в две идентичные дочерние молекулы. Процесс удвоения (хромосом) лежит в основе размножения всех организмов.
Рибонуклеиновая кислота РНК. Молекулы РНК большинства организмов представляют собой одиночную цепь нуклеотидов, сходную по строению с отдельной цепью ДНК. Только вместо дезоксирибозы РНК включает другой моносахарид - рибозу, а вместо тимина (Т) - урацил (У). Наследственную информацию, хранящуюся в ДНК, реализуют именно РНК. Их молекулы синтезируются на соответствующих участках одной из нитей ДНК по принципу комплементарности. РНК переносят информацию о строении белков от хромосом к месту синтеза и непосредственно участвуют в сборке белков.
Различают три основных типа РНК.
Матричные РНК (мРНК) служат матрицей для синтеза белков. Они содержат информацию (другое их название - "информационные РНК", и-РНК) о первичной структуре синтезируемых белков и переносят информацию о строении белков из ядра в цитоплазму к рибосомам, синтезирующим белки из аминокислот.
Рибосомные РНК (р-РНК) входят в состав рибосом, определяют их устройство и функционирование. На долю р-РНК приходится 85 % РНК клетки.
Транспортные РНК (т-РНК) имеют малые размеры, в их состав входят 70-100 нуклеотидов. Они доставляют аминокислоты к рибосомам в процессе синтеза белков.
Аденозинтрифосфорная кислота (АТФ). Энергия, выделяемая при расщеплении углеводов и липидов, запасается в молекулах АТФ - универсальном аккумуляторе энергии. У растений АТФ образуется в основном в процессе фотосинтеза. Использование АТФ позволяет организмам легко и быстро высвобождать и накапливать энергию.
По строению АТФ сходна с адениловым (А) нуклеотидом, входящим в состав ДНК, только вместо одного остатка фосфорной кислоты (фосфата) в состав АТФ входят три остатка. Клетки не могут содержать кислоты в заметных количествах - только их соли. Поэтому фосфорная кислота входит в АТФ в виде остатка (вместо ОН-группы кислоты имеется О”).
Под действием специальных ферментов молекула АТФ подвергается гидролизу, то есть присоединяет Н2О и расщепляется с образованием адено-
26
зиндифосфорной кислоты (АДФ). Этот процесс называют дефосфорилированием (потерей фосфата):
АТФ + Н2О АДФ + Н3РО4
Эта реакция обратима: АДФ может присоединять фосфат (фосфорилироваться) и переходить в АТФ, аккумулируя энергию органических соединений, полученных при фотосинтезе или с пищей. При переходе от АТФ к АДФ теряется значительно меньше энергии, чем приобретается при обратном процессе. Таким образом АТФ играет в клетках центральную роль универсального биологического аккумулятора энергии.
Молекулы АТФ синтезируются в митохондриях и хлоропластах, они обеспечивают энергией все процессы жизнедеятельности.
Строение клетки
Важнейшей принципиальной особенностью организмов является их клеточное строение. Из клеток состоят все животные, растения, грибы, микроорганизмы, исключение составляют только вирусы, но и они не способны жить и размножаться вне клетки, только внутри клеток вирусы проявляют себя как живые объекты.
Клетки - структурные и функциональные единицы всего живого. Мельчайшие организмы состоят из одной клетки; самые крупные - из миллиардов, каждая из которых выполняет определенную функцию и относительно независима. Идея о том, что все организмы построены из клеток, стала одним из наиболее важных теоретических достижений в истории биологии, поскольку создала единую основу для изучения всех живых существ. На клеточном уровне даже наиболее отдаленные виды весьма схожи по строению и биохимическим свойствам. Клеточная теория была сформулирована в начале XIX в. задолго до создания Дарвином теории эволюции, но эти две фундаментальные концепции тесно связаны между собой. В сходстве клеток мы улавливаем отзвук длительной эволюции, связывающей современные организмы, включая и растения, и нас с вами, с первыми клеточными структурами, возникшими на Земле миллиарды лет назад.
Существует великое множество различных типов клеток. Тело человека состоит более чем из ста типов клеток. В чайной ложке прудовой воды обнаруживается не один одноклеточный организм, а небольшой пруд, населяет несколько сотен их видов.
Вместе с тем растения состоят из клеток, и внешне, и по внутреннему строению, совершенно непохожих на клетки человеческого организма, а у насекомых имеются типы клеток, не встречающиеся ни у растений, ни у позвоночных, клетки грибов существенной отличаются от клеток всех ранее перечисленных. Таким образом, важная черта клеток - их разнообразие.
Другой, не менее существенной чертой организации клеток является их сходство. Каждая живая клетка представляет собой замкнутую и отчасти независимую единицу, окруженную плазматической мембраной, или плазма-леммой (часто называемой просто клеточной мембраной), которая контроли
27
рует транспорт веществ и обеспечивает структурную и биохимическую обособленность клетки от окружающей среды. У растений и бактерий поверх плазмалеммы формируется клеточная стенка или оболочка. Внутреннее пространство клетки заполнено цитоплазмой, которая у большинства клеток содержит ряд органелл и растворенные или суспензированные вещества. Кроме того, каждая живая клетка содержит ДНК, в которой закодирована генетическая информация. Принцип организации генетического кода одинаков у всех организмов, будь то бактерия, дуб или человек.
Все организмы можно разделить на две основные группы или надцарства: прокариоты, т. е. не имеющие ядра (pro на греческом - до, karion - ядро) и эвкариоты или эукариоты (ёи - хорошо, полностью), характеризующиеся наличием хорошо выраженного ядра. При этом надцарство прокариоты является одновременно и царством наравне с царствами эвкариотов (подробнее об этом будет сказано в разделе «Систематика»), Прокариоты в основном одноклеточные или нитчатые организмы, а эвкариоты преимущественно многоклеточные. Однако разделение представителей надцарств по этому признаку несколько условно - есть одноклеточные эвкариоты, например, эвгленовые водоросли (Euglenophytd), а некоторые прокариоты имеют тенденцию образовывать скопления взаимодействующих между собой одноклеточных организмов, что в какой-то мере напоминает многоклеточный организм. Предполагается, что эвкариоты произошли от прокариот. Однако, это не единственная точка зрения - можно, например, допустить, что они имели общих предков.
Сравнение прокариот с эвкариотами
К прокариотам относятся бактерии и сине-зеленые водоросли; к эвка-риотам - грибы, зеленые растения (включая остальные водоросли) и животные. Прокариоты отличаются от эвкариот по множеству признаков. Самые существенные различия показаны в таблице 1.
Таблица 1.
Основные различия между прокариотами и эвкариотами
Показатель Прокариоты Эвкариоты
Г енетический Нет истинного ядра и хромосом, Линейные молекулы ДНК, связан-
аппарат нет ядрышка. Замкнутая, условно кольцеобразная, нить ДНК частично ограничена от цитоплазмы так называемым нуклеоидом. ные с белками, и РНК образуют хромосомы внутри ядра. Ядро отделено от протоплазмы мембраной, включает ядрышко.
Органеллы Органелл мало, они не имеют оболочек (двойных мембран). Эндоплазматического ретикулума нет. Органелл много. Некоторые (ядро, митохондрии, хлоропласты) окружены двойной мембраной. Присутствует эндоплазматический ретикулум, к которому могут присоединяться рибосомы.
Фотосинтез Хлоропластов нет. Фотосинтез происходит в специализированных Фотосинтез происходит в хлоропластах, содержащих специальные
28
мембранах, не имеющих специальной упаковки.
Размеры кле- Диаметр в среднем составляет 0,5-ток 5 мкм.
Форма Одноклеточные или нитчатые.
мембраны, уложенные в ламеллы или граны.
Диаметр до 200 мкм, объем клетки как правило в 1000-10000 раз больше, чем у прокариот.
Одноклеточные, нитчатые или истинно многоклеточные.
Разделение эвкариотических клеток на компартменты (обособленные части) осуществляется с помощью мембран. Мембраны клеток различных организмов, видимые в электронный микроскоп, выглядят трехслойными. Два темных слоя толщиной около 25 ангстрем15 каждый разделены более светлым слоем толщиной около 35 ангстрем. Для обозначения мембраны с трехслойной организацией употребляют термин элементарная мембрана.
Строение элементарной мембраны до сих пор уточняется. Последние исследования выявили многие структурные особенности этого весьма сложного образования.
Строение растительной клетки
Растительная клетка состоит, как правило, из более или менее жесткой клеточной оболочки и протопласта (Рис. 1).
Рис. 1. Обобщенная схема строения растительной клетки, составленная по данным изучения разных растительных клеток с помощью электронного микроскопа: 1 - аппарат Гольджи (в данном случае две диктиосомы); 2 - свободно расположенные рибосомы; 3 -хлоропласты; 4 - межклеточные пространства; 5 - полирибосомы (несколько связанных между собой рибосом); 6 - митохондрии; 7 - лизисомы; 8 - гранулированная эндоплазматическая сеть; 9 - гладкая эндоплазматическая сеть; 10 - микротрубочки; 11 - пластиды; 12 - пластиды, проходящие сквозь поры в оболочке; 13 - клеточная оболочка; 14 - яд-
15
Ангстрем - внесистемная единица длины, равная 0,1 нанометра (нм).
29
рышко; 15, 18 - оболочка ядра; 16 - поры в ядерной оболочке; 17 - плазмалемма; 19 -гиалоплазма; 20 - тонопласт; 21 - вакуоли; 22 - ядро (Из Н.И. Арронет, 1974)
Протопласт состоит из цитоплазмы и ядра. В цитоплазме находятся органеллы (такие, как рибосомы, микротрубочки, пластиды и митохондрии) и мембранные системы (эндоплазматический ретикулум и диктиосомы). Их можно подробно рассмотреть только в электронный микроскоп. Кроме того, цитоплазма включает в себя цитоплазматический матрикс, или основное вещество, в которое погружены органеллы и мембранные системы. Цитоплазма, как мы об этом уже говорили, отделена от клеточной оболочки плазматической мембраной или плазмалеммой, которая представляет собой так называемую элементарную мембрану. В отличие от большинства животных клеток в цитоплазме растительных клеток содержатся одна или несколько вакуолей. Вакуоли - это пузырьки, заполненные жидкостью и окруженные элементарной мембраной, называемой тонопластом.
В живой растительной клетке основное вещество находится в постоянном движении. Можно видеть, как органеллы и другие включения вовлекаются в это движение, называемое током цитоплазмы, или циклозом. Циклоз прекращается в мертвых клетках. Он, несомненно, облегчает передвижение веществ в клетке и обмен ими между клеткой и окружающей средой. Однако основная функция циклоза пока неизвестна.
Коротко остановимся на строении и функциях отдельных органоидов или органелл.
Плазматическая мембрана (плазмалемма)
Среди многочисленных мембран клетки в плазмалемме или плазматической мембране ясно выражена трехслойность (темный - светлый - темный слои). Для растительных клеток обычны инвагинации (впячивания) плазма-леммы (на приведенной выше схеме они не отражены).
Плазматическая мембрана выполняет следующие функции: участвует в обмене веществ между клеткой и окружающей средой; у растений координирует синтез и сборку целлюлозных микрофибрилл клеточной оболочки; передает гормональные и внешние сигналы, контролирующие рост и дифференцировку клеток.
Ядро
Ядро - часто наиболее заметная структура в цитоплазме эукариотической клетки. Ядро выполняет важные функции: контролирует жизнедеятельность клетки, определяя, какие белки и в какое время должны синтезироваться; хранит основную генетическую информацию и передает ее дочерним клеткам в процессе клеточного деления.
Ядро эвкариотической клетки окружено двумя элементарными мембранами, образующими ядерную оболочку (рис. 2). Она пронизана многочисленными порами диаметром от 30 до 100 нм, видимыми в электронный
30
микроскоп. Поры - это не просто отверстия в оболочке: они имеют сложную структуру. Наружная мембрана ядерной оболочки в некоторых местах объединяется с эндоплазматическим ретикулумом (о нем позже). Ядерную оболочку можно рассматривать как специализированную, локально дифференцированную часть эндоплазматического ретикулума.
В ядре, окрашенном специальными красителями, используя световой микроскоп, можно различить тонкие нити и глыбки хроматина и нуклеоплазму или основное по объему вещество ядра. Хроматин состоит из ДНК (дезоксирибонуклеиновая кислота), связанной с большим количеством специальных белков - гистонов. В процессе клеточного деления хроматин все более уплотняется и в конце концов собирается в хромосомы. Генетическая информация у прокариот и эвкариот закодирована в молекулах ДНК (рис. 3). Содержание ДНК в одной клетке эвкариот значительно выше, чем в бактериальной клетке. У бактерий молекулы ДНК свободно располагаются в цитоплазме, а у эвкариот они организованы в хромосомы, находящиеся в ядре.
Перед делением клетки двойные нити ДНК соединяются с белками (гистонами), сжимаются, скручиваются и превращаются в хромосомы. Считается, что каждая хромосома включает одну молекулу ДНК.
Организмы различаются по числу хромосом в соматических (неполовых) клетках. Например, пустынный однолетник Haplopappus gracilis имеет 4 хромосомы; капуста - 20; подсолнечник - 34; большинство видов сосны и ели - 24; береза повислая - 28, Береза пушистая - 56; пшеница - 42; человек - 46; а один из видов папоротника из рода Ophioglossum - около 1250. Половые клетки, или гаметы, имеют только половину числа хромосом, характерного для соматических (греч. soma - тело) клеток организма. Число хромосом в гаметах называют гаплоидным (одинарным), в соматических клетках -диплоидным (двойным). Клетки, имеющие более двух наборов хромосом, называют полиплоидными.
Под световым микроскопом в ядре можно различить сферические структуры - ядрышки. В каждом ядре имеется одно или несколько ядрышек, которые заметны в неделящихся ядрах. В ядрышках синтезируются рибосомные РНК.
Пластиды
Наряду с вакуолями и клеточной оболочкой пластиды - характерные компоненты растительных клеток. Каждая пластида окружена собственной оболочкой, состоящей из двух элементарных мембран. Внутри пластиды различают мембранную систему и более или менее гомогенное вещество -строму. Зрелые пластиды обычно классифицируют на основании содержащихся в них пигментов.
31
Рис. 2. Интерфазное ядро в клетке корневой меристемы чернушки посевной Nigella saliva: В - вакуоли; Ядр - ядрышко; ФК - фибриллярный компонент ядрышка; Хн - хроматин; КО - оболочки двух соседних клеток. Ув. 25 000 раз. (Из Козубов, Данилова, 1972.)
32
Рис. 3. Часть молекулы ДНК (условно деспирализованная плоскостная модель). Пунктирами обозначены водородные связи между комплементарными парами азотистых оснований. Согласно модели, разработанной Дж. Уотсоном и Ф. Криком (1953), молекулы ДНК представляют собой две правозакрученные вокруг общей оси спиральные полинуклеотид ные нити (цепи) с шагом спирали 34 ангстрема (А), содержащие 10 нуклеотидов на виток и расположенные на расстоянии 18 А друг от друга. Противолежащие основания в цепях образуют за счет водородных связей так называемые комплементарные пары АТ и ГЦ. Таким образом последовательность оснований в одной цепи однозначно определяет последовательность оснований в другой (комплементарной) цепи молекулы (из Биологический энциклопедический словарь, 1989)
Хлоропласты, в которых протекает фотосинтез, содержат хлорофиллы и каротиноиды. Хлоропласты растений обычно имеют форму диска диаметром от 4 до 5 мкм. В одной фотосинтезирующей клетке листа может находиться 40-50 хлоропластов; в квадратном миллиметре листа - около 500 000.
33
В цитоплазме хлоропласты обычно располагаются параллельно клеточной оболочке (рис. 4, 5).
34
Рис. 4. Фрагмент хлоропласта в клетке мезофилла листа кукурузы (Zea mays). ОП -оболочка пластиды; Гр - граны тилакоидов; Тл - тилакоиды; Р - рибосомы; ОГ - осмие-фильные глобулы; Кр - крахмальное зерно в хлоропласте. Ув. 100 000 раз (из Козубов, Данилова, 1972)
Внутренняя структура хлоропласта довольно сложна. Строма16 пронизана развитой системой мембран, имеющих форму плоских пузырьков, называемых тилакоидами. Тилакоиды состоят из собственно фотосинтетических мембран, представляющих собой бимолекулярные липидные слои и мозаично вкрапленные в них липопротеидопигментные комплексы, образующие фотохимически активные центры, и содержат также специальные компоненты, участвующие в транспорте электронов и образовании аденозинтрифосфата (АТФ). Часть хлоропласта, находящаяся между тилакоидами, - строма, содержит ферменты, катализирующие темновые реакции фотосинтеза (например, биосинтез углеводов и белков). В строме откладывается образуемый при фотосинтезе крахмал. Каждый тилакоид, так же как и оболочка хлоропласта, состоит из двух мембран. Считается, что тилакоиды образуют единую систему. Как правило, тилакоиды собраны в стопки - так называемые граны, напоминающие столбики монет. Тилакоиды отдельных гран связаны друг с другом ламеллами стромы, эти же образования называются межгранными тилакоидами. Хлорофиллы и каротиноиды встроены в тилакоидные мембраны. Хлоропласты зеленых водорослей и растений часто содержат зерна крахмала и мелкие липидные (жировые) капли. Крахмальные зерна - это временные хранилища продуктов фотосинтеза. Они могут исчезнуть из хлоропластов растения, находящегося в темноте всего лишь 24 ч, и появиться вновь уже через 3-4 ч после переноса растения на свет.
Хлоропласты имеют собственные ДНК, РНК, рибосомы, синтезирующие белки, и обладают некоторой генетической автономностью, но находятся под общим контролем ядра. Фотосинтезирующие бактерии и большинство водорослей не имеют хлоропластов. Фотосинтетический аппарат большинства водорослей представлен специализированными внутриклеточными органеллами - хроматофорами, а фотосинтезирующих бактерий и сине-зелёных водорослей - тилакоидами (мембраны их содержат пигмент бактериохлорофилл или бактериовиридин, а также другие компоненты реакций фотосинтеза), погруженными в периферические слои цитоплазмы.
Хлоропласты - полуавтономные органеллы, они в некотором отношении напоминают бактерии. Например, хлоропласты имеют собственные рибосомы, которые у них, как и у бактерий, на одну треть меньше рибосом эв-кариот.
16
Строма - коллоидная белковая масса, заполняющая внутренние полости пластид.
35
Рис. 5. Строение хлоропласта. А - продольный разрез через хлоропласт: 1 - граны, образованные стопками ламелл; 2 - бимембранная оболочка; строма (матрикс); 4 - ламел-лы стромы; 5 - капли жира. Б - трехмерная схема расположения гран и ламелл: 1 - граны; 2 - ламеллы (Из Н.И. Арронет, 1974)
Синтез белка на рибосомах бактерий и хлоропластов подавляется антибиотиком хлорамфениколом, не оказывающим подобного действия в клетках эукариот. Кроме того, и бактерии, и хлоропласты имеют один или несколько нуклеоидов17 - светлых, лишенных гран участков, содержащих тяжи ДНК. ДНК хлоропластов и бактерий организована сходным образом, а именно - не окружена мембраной, не связана с гистонами и обычно существует в кольцевой форме.
Хлоропласты могут считаться основными клеточными органеллами растений, поскольку первыми стоят в цепи преобразования солнечной энергии, в результате которого мы получаем пищу и топливо. В хлоропластах протекает не только фотосинтез. Они участвуют и в синтезе аминокислот и жирных кислот, служат хранилищем временных запасов крахмала.
Хромопласты (от греч. chroma - цвет) - пигментированные пластиды. Многообразные по форме хромопласты не имеют хлорофилла, но синтезируют и накапливают каротиноиды, которые придают желтую, оранжевую и красную окраску цветкам, старым листьям, плодам и корням. Хромопласты могут развиваться из зеленых хлоропластов; последние при этом теряют хлорофилл и внутренние мембранные структуры и накапливают каротиноиды. Это происходит, например, при созревании многих фруктов.
Лейкопласты - непигментированные пластиды. Некоторые синтезируют крахмал (амилопласты), другие, по-видимому, способны к образованию разных веществ, в том числе липидов и белков. На свету лейкопласты могут превращаться в хлоропласты.
17 Нуклеоиды - оптически неразличимые эквиваленты ядер, свойственны прокариотам.
36
Пропластиды - мелкие бесцветные или бледно-зеленые недифференцированные пластиды, которые находятся в меристематических (делящихся) клетках корней и побегов. Они являются предшественниками других, более дифференцированных пластид - хлоропластов, хромопластов и амилопластов.
Для пластид характерны относительно легкие переходы от одного типа к другому. Пластиды размножаются делением надвое, и в этом отношении они также напоминают бактерии. В меристематических клетках время деления пропластид приблизительно совпадает со временем деления клеток. Однако в зрелых клетках большая часть пластид образуется в результате деления зрелых пластид в промежутках между делениями клетки.
Фотосинтез
Рядом ученых, изучавших фотосинтез, приводились весьма образные выражения, суть которых сводилась к тому, что сидя у камина или затапливая печку, мы согреваемся солнечным теплом давно прошедших дней. И это не просто образное выражение, а факт18.
В отличие от человека и животных все зеленые растения и часть бактерий способны синтезировать органические соединения из неорганических. Такой тип обмена веществ называется автотрофным (от греч. autos сам + tro-phe пища). В зависимости от вида энергии, используемой автотрофами для синтеза, их делят на две группы. Фотоавтотрофы используют энергию солнечного света, а хемоавтотрофы - химическую энергию, высвобождающуюся при окислении ими различных неорганических соединений.
Зеленые растения являются фотоавтотрофами. Их хлоропласты содержат хлорофилл, позволяющий осуществлять фотосинтез (от греч. photos свет + synthesis соединение) - преобразование энергии солнечного света в энергию химических связей синтезируемых органических соединений. Молекулы хлорофилла поглощают красную и синюю часть спектра солнечного излучения, а зеленая составляющая достигает сетчатки наших глаз. Поэтому большинство растений мы видим зелеными.
Для осуществления фотосинтеза растения поглощают из атмосферы СО2, а из водоемов и почвы - Н2О, соли азота и фосфора. Итоговое уравнение фотосинтеза выглядит довольно просто:
6СО2 + 6Н2О энергия света С6Н12О6 (глюкоза) + 602, но всем хорошо известно, что при смешивании углекислого газа и воды глюкоза не образуется. Фотосинтез - сложный многоступенчатый процесс, для прохождения которого необходимы солнечный свет, хлорофилл, ряд ферментов, энергия АТФ и молекулы-переносчики. Выделяют две фазы фотосинтеза - световую и темновую.
Световая фаза. Солнечные фотоны, передавая свою энергию молекуле хлорофилла, переводят молекулу в возбужденное состояние: ее электроны, получая дополнительную энергию, переходят на более высокие орбиты. От
18
Далее текст и рисунки раздела из Вертьянов, 2006.
37
рыв таких возбужденных электронов происходит значительно легче, чем невозбужденных. Молекулы-переносчики захватывают их и по цепи реакций перемещают на другую сторону мембраны тилакоида.
В темновые реакции
Рис. 5а. Световая фаза фотосинтеза (из Вертьянов, 2006)
Потери электронов восполняются хлорофиллом в результате фотолиза - расщепления воды под действием света на протоны и молекулярный кислород О2 (катализирует этот сложный процесс специальный марганецсодержащий ферментный комплекс):
2Н2О 4Н+ + 4е + О2, где е - электрон.
Молекулярный кислород диффундирует сквозь мембраны тилакоидов и выделяется в атмосферу. Протоны неспособны к проникновению че-
рез мембрану и поэтому остаются внутри.
В результате снаружи мембраны накапливаются электроны, доставленные молекулами-переносчиками с возбужденных молекул хлорофилла, а внутри - протоны, образовавшиеся в результате фотолиза; еще часть прото
нов доставляется молекулами-переносчиками из стромы за счет энергии возбужденных электронов. На мембране возникает разность потенциалов.
Рис. 56. Фотосинтез на мембранах тилакоидов хлоропластов (из Вертьянов, 2006)
В мембраны тилакоидов хлоропласта, так же как и во внутренние мембраны митохондрий, встроены молекулы фермента АТФ-синтазы, осуществляющие синтез АТФ. Аналогичным образом в молекулярной структуре АТФ-синтазы имеется канал, через который могут проходить протоны. При достижении на мембране критической разности потенциалов протоны, влекомые силой электрического поля, продвигаются по каналу АТФ-синтазы, затрачивая энергию на синтез АТФ.
Электроны от возбужденного хлорофилла передаются по цепи переносчиков на кофермент НАДФ (никотин-амид-аденин-динуклеотид с фосфа
38
том) и восстанавливают его до НАДФН («надф-аш»), донором протонов служит вода.
Таким образом, во время световой фазы фотосинтеза происходят следующие ключевые процессы: выделение в атмосферу свободного кислорода, синтез АТФ и восстановление НАДФ до НАДФН. Протекание дальнейших реакций может происходить и в темноте (за счет запаса энергии АТФ и НАДФН), потому носит название темновой фазы.
Темновая фаза. Реакции этой фазы происходят в строме хлоропласта за счет энергии АТФ при участии НАДФН, образовавшихся в световой фазе, и комплекса ферментов. Молекулы СО2 восстанавливаются до глицеральдегид-3-фосфата, из которого синтезируется глюкоза (а затем и другие углеводы):
СО2 глицеральдегид-3-фосфат C6Hi2O6 (глюкоза).
Для образования одной молекулы глюкозы требуется 18 АТФ (отдающих энергию и переходящих в АДФ) и 12 НАДФН (отдающих протоны и электроны, восстанавливающих СО2 и переходящих в НАДФ). Реакции темновой фазы носят название цикла Кальвина. Для получения одной молекулы глюкозы необходимо 6 оборотов цикла, после каждого оборота все ферменты и молекулы-переносчики возвращаются в исходное состояние.
Кроме глюкозы из глицеральдегид-3-фосфата могут синтезироваться жирные кислоты, аминокислоты и пр. Углеводы и жирные кислоты далее транспортируются в лейкопласты, где из них формируются запасные питательные вещества - крахмал и жиры.
С наступлением темноты растения продолжают процесс фотосинтеза, используя запасенные на свету соединения. Когда этот запас исчерпывается, прекращается и фотосинтез. В темное время суток растения напоминают по типу обмена веществ животных: они поглощают кислород из атмосферы (дышат) и окисляют при его помощи запасенные днем питательные вещества. На дыхание растения используют в 20-30 раз меньше кислорода, чем выделяют в атмосферу в процессе фотосинтеза.
Ежегодно растительность планеты дает 200 млрд, т кислорода и 150 млрд, т органических соединений, необходимых человеку и животным. Энергия химических связей этих соединений (за счет которой и живут животные) значительно превышает количество тепла, выделяющегося при сжигании всем населением планеты горючих полезных ископаемых.
Хемосинтез. Большинство бактерий лишены хлорофилла. Некоторые из них являются хемоавтотрофами: для синтеза органических веществ они используют не энергию света, а энергию, высвобождающуюся при окислении неорганических соединений. Такой способ получения энергии и синтеза органических веществ назвали хемосинтезом (от греч. chernia химия). Явление хемосинтеза открыто в 1887 г. русским микробиологом С. Н. Виноградским.
Нитрифицирующие бактерии окисляют аммиак, образующийся при гниении органических остатков в почве или в водоемах, до азотистой кислоты и далее - азотистую до азотной.
39
Водородные бактерии окисляют молекулы водорода (до Н20), образующиеся в почве в результате гликолиза органических остатков другими микроорганизмами.
Железобактерии используют энергию, высвобождающуюся при окислении двухвалентного железа до трехвалентного (закисные соли до окисных).
Серобактерии обитают, как правило, в болотах и «питаются» сероводородом. Окисляя сероводород, они получают необходимую для жизнедеятельности энергию. В результате накапливается сера, при ее окислении до серной кислоты высвобождается еще часть энергии. Огромное количество серобактерий обитает в Черном море, воды которого со стометровой глубины насыщены сероводородом.
Гетеротрофный тип обмена веществ. Человек и животные не способны синтезировать необходимые для жизнедеятельности органические вещества из неорганических и вынуждены получать их с пищей. Такие организмы называют гетеротрофными (от греч. heteros другой). К гетеротрофам относятся также грибы и большинство бактерий. Органические вещества разлагаются гетеротрофами на простые углеводы, аминокислоты, нуклеотиды, из которых далее синтезируются необходимые макромолекулы. Часть веществ расщепляется до конечных бедных энергией молекул (СО2, Н20), а высвобождающаяся энергия используется для жизнедеятельности. Некоторое количество энергии рассеивается в виде тепла, поддерживающего температуру тела.
У ряда фотосинтезирующих бактерий, одноклеточных водорослей миксотрофное (от греч. mixis смешение) питание. На свету они фотосинтезируют, а темноте переходят к питанию органическими веществами.
Способность зеленых растений к фотосинтезу определила и сформировала биосферу Земли, дав пищу всем гетеротрофным организмам, а главное (или первое), дав им возможность дышать. Считается, что до появления зеленых растений в атмосфере Земли не было, или было очень мало, молекулярного кислорода. Человек и все остальные животные были и остаются абсолютно зависимыми от чистоты и насыщенности кислородом воздуха, а лесные массивы мира плюс фотосинтезирующий планктон водной среды с полным основанием можно считать «легкими планеты».
Наряду с фотосинтезом на Земле совершаются примерно равноценные по масштабам, но противоположные по направлению процессы окисления органических веществ и восстановленного углерода при горении топливных материалов (каменный уголь, нефть, газ, торф, дрова и т.п.), при расходовании органических веществ живыми организмами в процессе их жизнедеятельности (дыхание, брожение), в результате которых образуются полностью окисленные соединения - углекислый газ и вода, и освобождается энергия. Затем с помощью энергии солнечной радиации углекислый газ, вода снова вовлекаются в процессы фотосинтеза. Таким образом, энергия солнечного света, используемая при фотосинтезе, служит движущей силой колоссального по размерам круговорота на Земле таких элементов, как углерод, водород, кислород. В этот круговорот включаются и многие другие элементы, напри
40
мер, N, S, P, Mg, Ca. За время существования Земли благодаря фотосинтезу важнейшие элементы и вещества прошли уже много тысяч циклов полного круговорота.
В предшествующие эпохи условия для фотосинтеза на Земле были более благоприятны в связи с сильным перевесом восстановительных процессов над окислительными. Постепенно огромные количества восстановленного углерода в органических остатках оказались захороненными в недрах Земли, образовав громадные залежи горючих ископаемых. В результате этого в атмосфере сильно снизилось относительное содержание углекислого газа (до 0,03 объёмных %) и повысилось содержание кислорода, что существенно ухудшило условия для фотосинтеза.
Следствием появления на Земле мира фотосинтезирующих растений и непрерывного новообразования ими больших количеств богатых энергией органических веществ явилось развитие мира гетеротрофных организмов (бактерий, грибов, животных, человека) - потребителей этих веществ и энергии. В результате (в процессе дыхания, брожения, гниения, сжигания) органические соединения стали окисляться и подвергаться разложению в таких же количествах, в каких образуют их высшие растения, водоросли, бактерии. На Земле установился круговорот веществ, в котором сумма жизни на нашей планете определяется масштабами фотосинтеза. В текущем геологическом периоде (антропогеновом) размеры фотосинтетической продуктивности на Земле, вероятно, стабилизировались. Однако в связи с бурно возрастающим использованием продуктов фотосинтеза основным её потребителем - человеком - приходится думать о предстоящем истощении горючих ископаемых, пищевых, лесных ресурсов и т.п. Фотосинтетическая мощность современной растительности недостаточна для регенерации атмосферы: растительность Земли не способна полностью усваивать весь углекислый газ (относительное содержание его в атмосфере за последние 100 лет медленно, но неуклонно возрастает), дополнительно поступающий в окружающую среду в результате бурно возрастающих масштабов добычи и сжигания горючих ископаемых.
При этом потенциальная фотосинтетическая активность растений используется далеко не полно. Проблема сохранения, умножения и наилучшего использования фотосинтетической продуктивности растений - одна из важнейших в современном естествознании и практической деятельности человека.
Митохондрии
Как и хлоропласты, митохондрии окружены двумя элементарными мембранами. Внутренняя мембрана образует множество складок и выступов, называемых кристами, которые значительно увеличивают внутреннюю поверхность митохондрии (рис. 6). Митохондрии обычно мельче, чем пластиды, имеют около половины микрометра в диаметре и очень разнообразны по длине и форме.
41
Рис. 6. Строение митохондрии. А - продольные разрезы митохондрий из разных частей растения: 1 - наружная мембрана; 2 - внутренняя мембрана, образующая кристы; 3 - кристы; 4 -матрикс. Б - трехмерная схема, часть митохондрии условно срезана; обозначения те же (из Н.И. Арронет, 1974)
В митохондриях осуществляется процесс дыхания, в результате которого органические молекулы расщепляются с высвобождением энергии и передачей ее молекулам АДФ (аденозиндифосфата), благодаря чему по
следний подзаряжается до АТФ (аденозинтрифосфата), основного резерва энергии всех эвкариотических клеток. Как правило, митохондрии плохо вид
ны под световым микроскопом, но их можно хорошо рассмотреть с помощью электронного микроскопа. Большинство растительных клеток содержит сотни и тысячи митохондрий, их число в одной клетке определяется потребностью клетки в АТФ.
С помощью прерывистой съемки можно увидеть, что митохондрии находятся в постоянном движении. Они поворачиваются, изгибаются, перемещаются из одной части клетки в другую, а кроме того, сливаются друг с другом и делятся. Митохондрии обычно собираются там, где нужна энергия.
Если плазматическая мембрана активно переносит вещества из клетки или в клетку, то митохондрии располагаются вдоль поверхности мембраны. У подвижных одноклеточных водорослей митохондрии, как правило, скапливаются у оснований жгутиков и, по-видимому, поставляют энергию, необходимую для их движения.
Митохондрии, подобно пластидам, являются полуавтономными органеллами, содержащими компоненты, необходимые для синтеза собственных белков. Внутренняя мембрана окружает жидкий матрикс, в котором находятся белки, РНК (рибонуклеиновые кислоты), тяжи ДНК, рибосомы, сходные с бактериальными, и различные растворенные вещества. ДНК существует в виде кольцевых молекул, располагающихся в одном или нескольких нуклеоидах.
Происхождение митохондрий и хлоропластов
На основании сходства бактерий с митохондриями и хлоропластами эвкариотических клеток высказывалось предположение, что митохондрии и хлоропласты произошли от бактерий, которые нашли «убежище» в более
42
крупных гетеротрофных19 клетках - предшественниках эвкариот. Правда в это убежище они попадали, вероятнее всего, не "по собственной воле", а вследствие поглощения гетеротрофными клетками. Бактерии, имеющие возможность использовать молекулярный кислород для окисления питательных веществ и использовать энергию света, придавали эти полезные свойства более крупным клеткам, которые с такими «помощниками» имели явное преимущество перед своими современниками и, несомненно, вскоре размножились за их счет. Все ныне живущие эвкариоты, за очень малым исключением, содержат митохондрии, а все автотрофные эвкариоты содержат также хлоропласты. По-видимому, они были приобретены в результате независимых случаев симбиоза (симбиозом называется тесное объединение двух или более организмов, которое может быть, но не обязательно, полезно для каждого). Крупные и более сложные клетки эвкариот, по-видимому, защищали свои симбиотические органеллы от неблагоприятных внешних воздействий. В результате эвкариоты смогли захватить сушу и кислые воды, в которых отсутствуют прокариотические цианобактерии (или сине-зеленые водоросли), но в изобилии живут эвкариотические зеленые водоросли.
Однако существуют и другие точки зрения. Согласно одной из них при историческом становлении прокариот и органелл эвкариотических организмов реализовалась общая для них закономерность (гомологичность, параллелизм) развития.
Вакуоли
Вакуоли - это свойственные растениям ограниченные мембраной участки клетки, заполненные жидкостью, которая называется клеточным соком. Они окружены тонопластом, или вакуолярной мембраной.
Молодая растительная клетка обычно содержит многочисленные мелкие вакуоли, которые увеличиваются в размерах и сливаются в одну вакуоль по мере старения клетки. В зрелой клетке до 90% ее объема может быть занято вакуолью, тогда как цитоплазма в виде тонкого периферического слоя прижата к клеточной оболочке. Заполняя большую часть клетки «дешевым» вакуолярным содержимым, растения экономят потребляющую азот «дорогую» цитоплазму, в то время как ее поверхность остается обширной. В основном увеличение размера клетки происходит за счет роста вакуоли. В результате этого возникает тургорное давление и поддерживается упругость ткани, в чем и заключается одна из важных функций вакуоли и тонопласта.
Современные исследования (Гамалей, 2004; см. список литературы), говорят о том, что вакуоли тесно связаны с эндоплазматической сетью и возникают первоначально как местное расширение участка эндоплазматического ретикулума. Выяснено, например, что химический состав содержимого микротрубочек ретикулума и основной вакуоли (полости) ситовидных трубок во флоэме практически идентичен. Вакуоли увеличиваются за счет по
19 Гетеротрофные организмы - получающие углерод и пр. вещества за счет других организмов, противопоставляются автотрофным зеленым (фотосинтезирующим) растениям.
43
вышения внутреннего давления, возникающего при накапливании продуктов фотосинтеза.
Основной компонент клеточного сока - вода с растворенными в ней олигосахарами (первичные продукты фотосинтеза, остальные, в том числе растворимые белки, варьируют в зависимости от типа растения и его физиологического состояния.
Тонопласт, являющийся по-существу продолжением мембран, образующих микротрубочки эндоплазматической сети, играет важную роль в активном транспорте и накоплении в вакуолях некоторых ионов. Концентрация ионов в клеточном соке, таким образом, может быть выше, чем в окружающей цитоплазме. При высоком содержании некоторых веществ в вакуолях могут образоваться кристаллы. Особенно часто встречаются кристаллы оксалата кальция, имеющие различную форму. Обычно содержимое вакуолей слабокислое, иногда - очень кислое, как в плодах лимона.
Вакуоли - места накопления различных метаболитов (продуктов метаболизма, жизнедеятельности), таких, как запасные белки. Они удаляют из цитоплазмы и ядовитые вторичные продукты метаболизма.
В вакуолях часто откладываются пигменты. Голубой, фиолетовый, пурпурный, темно-красный и пунцовый цвета придают растительным клеткам пигменты из группы антоцианов. В отличие от большинства других растительных пигментов антоцианы легко растворяются в воде и содержатся в клеточном соке. Они определяют красную и голубую окраску многих овощей (редис, турнепс, капуста), фруктов (виноград, сливы, вишни) и множества цветов (васильки, герани, дельфиниумы, розы и пионы). Иногда эти пигменты маскируют в листьях хлорофилл, как, например, у декоративного клена Шведлера (краснолистная форма клена остролистного). Антоцианы окрашивают осенние листья в ярко-красный цвет. Они образуются в холодную солнечную погоду, когда в листьях прекращается синтез хлорофилла. По мере разрушения имеющегося хлорофилла проявляются антоцианы. В листьях, где эти пигменты не образуются, после разрушения хлорофилла становятся заметными желто-оранжевые каротиноиды хлоропластов. Наиболее ярко окрашены листья холодной ясной осенью.
Вакуоли участвуют и в разрушении макромолекул, и в круговороте их компонентов в клетке. Отдельные органеллы, такие, как рибосомы, митохондрии и пластиды, могут попадать в вакуоли и там разрушаться. По этой переваривающей активности вакуоли сравнимы с лизисомами - органеллами, которые встречаются в клетках животных.
Рибосомы
Рибосомы - маленькие частицы всего около 17-23 нм в диаметре, состоящие примерно из равных количеств белка и РНК. В рибосомах аминокислоты соединяются с образованием белков; они наиболее многочисленны в цитоплазме клеток с активным метаболизмом. Рибосомы располагаются в
44
цитоплазме клетки свободно или же прикрепляются к эндоплазматическому ретикулуму (рис. 7). Обнаруживают их и в ядре.
Как указывалось выше, пластиды и митохондрии содержат рибосомы, подобные таковым у прокариот. Рибосомы, активно участвующие в синтезе белка, образуют комплексы, называемые полирибосомами, или полисомами. Клетки, синтезирующие белки в больших количествах, имеют обширную систему полисом. Полисомы часто прикреплены к наружной поверхности оболочки ядра.
Рис. 7. Шероховатый или гранулярный эндоплазматический ретикулум: Р - рибосомы на поверхности трубочек и цистерн эндоплазматического ретикулума; ГЭР - гранулярный эндоплазматический ретикулум; Д - диктиосомы; ПГ - пузырьки Гольджи, отделившиеся от краев диктиосом (из Козубов, Данилова, 1972)
Эндоплазматический ретикулум
Эндоплазматический ретикулум (эндоплазматическая сеть) - это сложная трехмерная мембранная система неопределенной протяженности. В разрезе любой фрагмент эндоплазматического ретикулума выглядит как две
45
элементарные мембраны с узким прозрачным пространством между ними. Форма и протяженность эндоплазматического ретикулума зависят от типа клетки, ее метаболической активности и стадии дифференцировки. Например, в клетках, секретирующих или запасающих белки, эндоплазматический ретикулум имеет форму плоских мешочков, или цистерн, с многочисленными рибосомами, связанными с его внешней поверхностью. Эндоплазматический ретикулум, несущий рибосомы, называют шероховатым эндоплазматическим ретикулумом (см. рис. 4). Полисомы и шероховатый эндоплазматический ретикулум - основные места синтеза белка. Напротив, клетки, которые секретируют липиды, имеют обширную систему трубочек, на которых нет рибосом. Эндоплазматический ретикулум, не имеющий рибосом, называют гладким эндоплазматическим ретикулумом (рис. 7). Гладкий эндоплазматический ретикулум обычно имеет трубчатую форму. Шероховатый и гладкий эндоплазматические ретикулумы могут присутствовать в одной и той же клетке. Как правило, между ними имеются многочисленные связи.
Рис. 8. Органеллы в клетке мезофилла березы повислой (Betula penduld). Электронная микрофотография. Ув. 70 000. Эр - элементы эндоплазматического ретикулума; Д -диктиосомы; Мт - митохондрия; Хл - часть хлоропласта (из Козубов, Данилова, 1972)
Эндоплазматический ретикулум функционирует как коммуникационная система клетки. На электронных микрофотографиях можно видеть, что он связан с внешней оболочкой ядра. Фактически эти две структуры образу
46
ют единую мембранную систему, в которой по последним данным, первична эндоплпзматическая сеть. Когда ядерная оболочка во время деления клетки разрывается, ее обрывки напоминают фрагменты эндоплазматического ретикулума.
Ранее было сказано, что эндоплазматический ретикулум являет собой систему транспортировки веществ, в первую очередь сахаров, а также белков и липидов, в разные части клетки. Кроме того, эндоплазматические ретикулумы соседних клеток соединяются через цитоплазматические тяжи - плаз-модесмы, - которые проходят сквозь клеточные оболочки. В парах пор контактирующих живых паренхимных клеток, а также между члениками ситовидных трубок, внутри плазмодесменного тяжа всегда находится трубочка эндоплазматического ретикулума. Таким образом трубочки и цистерны ретикулума вместе с вакуолями образуют единую сеть, пронизывающую и заполняющую все живые контактирующие между собой клетки.
Эндоплазматический ретикулум - основное место синтеза клеточных мембран. По-видимому, в некоторых растительных клетках здесь образуются мембраны вакуолей и микротелец, а также цистерны диктиосом.
Аппарат Гольджи
Термин аппарат Гольджи используется для обозначения всех диктиосом, или телец Гольджи, в клетке. Диктиосомы - это группы плоских, диско-видных пузырьков, или цистерн, которые по краям разветвляются в сложную систему трубочек. Диктиосомы в клетках высших растений обычно состоят из четырех - восьми цистерн, собранных вместе (рис. 7, 8).
Обычно в пачке цистерн различают формирующуюся и созревающую стороны. Мембраны формирующихся цистерн по структуре полностью сходны с мембранами эндоплазматического ретикулума, а мембраны созревающих цистерн - с плазматической мембраной. По существу аппарат Гольджи является частью эндоплазматической сети.
Диктиосомы участвуют в секреции, а у большинства высших растений - в образовании клеточных оболочек. Полисахариды клеточной оболочки, синтезируемые диктиосомами, накапливаются в пузырьках, которые затем отделяются от созревающих цистерн. Эти секреторные пузырьки мигрируют и сливаются с плазматической мембраной; при этом содержащиеся в них полисахариды встраиваются в клеточную оболочку. Продукты, накапливающиеся в диктиосомах, не всегда синтезируются ими. Некоторые вещества образуются в других структурах, например в других участках эндоплазматического ретикулума, а затем транспортируются в диктиосомы, где видоизменяются перед секрецией. Хороший пример подобных веществ - гликопротеины (углеводно-белковые соединения), важный строительный материал клеточной оболочки. Белковая часть синтезируется полисомами шероховатого эндоплазматического ретикулума, углеводная - в диктиосомах, где обе части объединяются, образуя гликопротеины.
47
В настоящем кратком обзоре строения клетки рассмотрены далеко не все органеллы и структурные компоненты. Мы заострили внимание лишь на наиболее важных, выполняющих самые ответственные функции. На самом деле клетка значительно сложнее и к настоящему времени еще далеко не полностью изучена. Без малейшего преувеличения можно сказать, что самая простая живая клетка значительно сложнее и более рационально организована, чем, скажем, самый совершенный современный компьютер.
ДЕЛЕНИЕ КЛЕТКИ
Клеточный цикл
Все новые клетки возникают в результате деления уже существующих клеток надвое. Если делится одноклеточный организм, то из старого организма образуются два новых. Многоклеточный организм начинает свое развитие с одной-единственной клетки; все его многочисленные клетки образуются затем путем многократных клеточных делений. Эти деления продолжаются в течение всей жизни многоклеточного организма.
Новообразованные клетки обычно становятся способными к делению лишь после некоторого периода роста. Кроме того, делению должно предшествовать удвоение клеточных органелл; в противном случае в дочерние клетки попадало бы все меньше и меньше органелл. Некоторые органеллы, например хлоропласты и митохондрии, сами воспроизводятся делением надвое; клетке достаточно иметь хотя бы одну такую органеллу, чтобы затем образовать их столько, сколько ей требуется. Каждой клетке необходимо также иметь вначале какое-то количество рибосом, чтобы использовать их для синтеза белков, из которых затем можно построить новые рибосомы, эндоплазматический ретикулум и многие другие органеллы.
Перед началом клеточного деления ДНК клетки должна реплицироваться (дуплицироваться) с очень высокой точностью, поскольку ДНК несет в себе информацию, необходимую клетке для синтеза многочисленных и строго определенных белков (рис. 9).
Если какая-нибудь дочерняя клетка не унаследует полный набор заключенных в ДНК инструкций, то она может оказаться не в состоянии синтезировать все те белки, которые могут ей потребоваться. Чтобы этого не случилось, ДНК должна реплицироваться очень точно и каждая дочерняя клетка при клеточном делении должна получить ее копию. Только после этого начинается собственно деление клетки - митоз. Все выше сказанное свершается в процессе, который называется клеточным циклом (рис. 10).
48
Деление ядра
Деление цитоплазмы
Телофаза
Деление цитоплазмы
Рис. 9. Дупликация двойной спирали ДНК. Спираль родительской ДНК раскручивается (вверху), и комплементарные (соединенные строго определенным образом: аденин с тимином, цитозин с гуанином) цепи (нити) отделяются одна от другой в результате разрыва слабых водородных связей между комплементарными основаниями (нуклеотидами). Сами нити ДНК не разрываются, благодаря прочности их фосфатных связей между линейно расположенными нуклеотидами. Теперь каждая нить служит матричной цепью. Далее происходит спаривание между нуклеотидами (основаниями) каждой из двух разделившихся нитей (матричных цепей) и свободными нуклеозидтрифосфатами. Последние связываются между собой фосфатными (фосфодиэфирными) связями, образуя новые полинуклеотидные цепи. Таким
образом получаются две идентичные молекулы ДНК В разделении родительских цепей и
образовании новых полинуклеотидных цепей участвует фермент ДНК-полимераза (затемненный круг на рисунке) и другие ферменты (из Грин, Стаут, Тейлор, 1990)
Рис. 10. Клеточный цикл, состоящий из интерфазы (Gi, S и G2) и митоза (М). Gi - интенсивные процессы синтеза. Образование митохондрий, хлоропластов и других органелл. Ядрышко продуцирует РНК, образуются рибосомы. Клетка синтезирует белки. Рост клетки. S - репликация ДНК. Синтез белковых молекул, называемых гистонами, с которыми связы-
вается каждая нить ДНК. Каждая хромосома превращается в две хроматиды. G2 - интенсивные процессы синтеза. Деление митохондрий и хлоропластов. Увеличение энергетических запасов. Образование веретена деления. М - митоз. Деление ядра, включающее четыре условно выделяемые фазы. С - равномерное распределение органелл и цитоплазмы между дочерними клетками (из Грин, Стаут, Тейлор, 1990)
Повторяемость клеточных циклов у разных организмов в разных органах не одинакова. Например: бактерии могут, размножаясь, повторять клеточные циклы до тех пор, пока им хватает питательного субстрата; клетки слизистой оболочки млекопитающих делятся в течение всей жизни животного; клетки камбия у древесных растений в зоне умеренного климата активно делятся в начале вегетационного периода, затем частота делений уменьшается, зимой камбиальные клетки не активны, а весной деления вновь активизируются.
49
Митоз
Митоз - это деление ядра, приводящее к образованию двух дочерних ядер, в каждом из которых имеется точно такой же набор хромосом, как в родительском ядре. Поскольку за делением ядра обычно следует клеточное деление, термин «митоз» нередко употребляют в более широком смысле, имея в виду и сам митоз, и клеточное деление, которое за ним следует.
Митозу, как нам уже известно, должно предшествовать удвоение хромосом, происходящее в интерфазе клеточного цикла. Удвоившаяся хромосома состоит из двух одинаковых половинок (хроматид), соединенных при помощи особой структуры, которую называют центромерой. Эти две хроматиды превращаются в обособленные хромосомы лишь к середине митоза, когда центромера делится и их уже больше ничто не связывает.
Удвоение хромосом происходит в интерфазе, т. е. в период между делениями. В это время вещество хромосом распределено по всему ядру в виде рыхлой массы. Между удвоением хромосом и началом митоза проходит обычно некоторое время (стадии Gi, S и G2, рис. 10).
Митоз представляет собой непрерывную цепь событий, но, для того чтобы удобнее было его описывать, биологи делят этот процесс на четыре стадии (фазы) в зависимости от того, как выглядят в это время хромосомы в световом микроскопе (рис. 11).
1. Профаза - стадия, на которой появляются первые указания на то, что ядро собирается приступить к митозу. Вместо рыхлой массы ДНК и белка в профазе становятся ясно видны нитевидные удвоившиеся хромосомы. Такая конденсация хромосом - весьма нелегкая задача: это примерно то же самое, что свернуть тонкую двухсотметровую нить так, чтобы ее можно было втиснуть в цилиндр диаметром 1 мм и длиной 8 мм. По большей части в профазе ядрышко и ядерная мембрана исчезают.
2. Метафаза - стадия подготовки к делению. Для нее характерно завершение образования митотического веретена, т.е. каркаса из микротрубочек. Каждая удвоившаяся хромосома в области центромеры прикрепляется к микротрубочке и направляется к середине веретена. Все хромосомы собираются в средней части клетки в одной плоскости, формируется так называемая «серединная пластинка».
3. Анафаза - стадия, на которой центромеры, наконец, делятся и из каждой удвоившейся хромосомы образуются две отдельные, совершенно идентичные хромосомы. Разделившись, эти идентичные хромосомы движутся к противоположным концам, или полюсам, митотического веретена; однако, что именно приводит их в движение, пока неясно. В конце анафазы у каждого полюса находится полный набор хромосом.
4. Телофаза - последняя стадия митоза. Хромосомы начинают раскручиваться, снова превращаясь в рыхлую массу ДНК и белка. Вокруг каждого набора хромосом вновь появляется ядерная мембрана. Телофаза обычно сопровождается делением цитоплазмы, в результате которого образуются две клетки, каждая с одним ядром. В животных клетках клеточная мембрана пережимается посередине и в конце концов разрывается в этой точке, так что
50
получаются две отдельные клетки. У растений в цитоплазме посередине клетки возникает перегородка, а затем каждая дочерняя клетка строит возле нее со своей стороны клеточную стенку.
Рис. 11. Митотическое деление клетки в кончике корешка лука (Allium сера). Слева (прописные буквы) - рисунки, сделанные с использованием микроскопа, справа (строчные буквы) - схемы.
А - интерфаза (состояние клетки между циклами деления). Хромосомы дес-пирализованы. Б - профаза. Хромосомы хорошо различимы и состоят из двух хроматид каждая. Клетка готова к делению. В -метафаза. Оболочка ядра растворяется, хромосомы располагаются в экваториальной плоскости клетки, появляются нити веретена, прикрепляющиеся к центромерам хромосом. Г - анафаза. Каждая хромосома расщепляется вдоль на две хроматиды, последние расходятся к противоположным полюсам клетки. Д - телофаза. Хромосомы деспирализуются и становятся мало заметными. Формируются ядра, ограниченные каждое своей оболочкой; появляются ядрышки; исчезают нити веретена; в середине делящейся клетки образуется фрагмопласт, формирующий оболочку, которая, разрастаясь от центра к периферии, делит клетку на две. Е - цитокинез. Завершение образования оболочек у двух дочерних клеток; ядра принимают интерфазный вид.
Далее весь цикл деления повторяется (из Козубов, Данилова, 1972).
При помощи факторов (веществ), нарушающих митоз, можно получать тетраплоидные клетки, т.е. клетки с числом хромосом, вдвое большим, чем в исходной (диплоидной) клетке. Одним из таких факто-
ров является колхицин - вещество, экстрагируемое из однодольного растения - безвременника (Colchicum). Колхицин связывается с белком микротрубочек и препятствует образованию веретена. Вследствие этого дочерние хромосомы отделяются одна от другой, но не делятся на две группы, распределяемые по разным клеткам, так что возникает ядро с удвоенным по сравнению с нормальным числом хромосом. Если обработать побег какого-нибудь растения колхицином, а затем дать этому растению зацвести и завязать семена, то получаются тетраплоидные семена. Тетраплоидные растения часто бывают
51
крупнее и мощнее исходного родительского растения; многие сорта культурных растений - фрукты, овощи и цветы - это именно тетраплоиды, либо возникшие естественным путем, либо полученные искусственно.
Вместе с тем далеко не все тетраплоиды - рекордсмены. Так, береза пушистая (56 хромосом) по отношению к березе повислой (28 хромосом) -тетраплоид, однако эти два вида не различаются существенно по средним таксационным показателям.
Значение митоза состоит прежде всего в поддержании генетической стабильности всех клеток организма и увеличении числа клеток в процессе роста и регенерации тканей. У одноклеточных организмов митоз служит механизмом бесполого размножения.
Митоз - процесс деления соматических клеток (от греч. soma - тело), применительно к растениям такие клетки называют вегетативными. Деление половых (репродуктивных) клеток с образованием гамет, участвующих затем в оплодотворении, называется мейозом.
Мейоз
Мейоз (рис. 12) принципиально отличается от митоза тем, что в результате деления образуются клетки с одинарным (гаплоидным) набором хромосом, в отличие от всех вегетативных (или соматических), обладающих в норме двойным (диплоидным) набором.
Число хромосом в гаплоидном наборе всегда в два раза меньше, чем в соматических (диплоидных) клетках. Число хромосом принято обозначать буквой п, количество ДНК в хромосомном наборе - буквой с.
Следовательно, в соматических клетках хромосомный набор обозначается 2п2с, в половых клетках - 1п1 с.
Мейоз осуществляется путем двух, следующих одно за другим клеточных делений. I мейотическое деление протекает следующим образом. Профаза начинается спирализацией хромосом. Они видны в виде тонких слабо окрашивающихся нитей. Затем гомологичные хромосомы сближаются, и каждая точка одной хромосомы совмещается с соответствующей точкой другой гомологичной хромосомы. Процесс тесного и точного сближения гомологичных хромосом в мейозе называется конъюгацией (рис. 13).
В процессе конъюгации гомологичные хромосомы сближаются и удерживаются рядом благодаря образованию каждой хроматидой нитей толщиной 1,5-2,0 нм, растущих по направлению к одной из хроматид второй (гомологичной) хромосомы. Нити на конце утолщены (петли ДНК). Утолщения нитей двух противостоящих несестринских хроматид соединяются наподобие застежки «молния». Благодаря образованию таких мостиков гомологичные хромосомы могут долго находиться в сближенном состоянии. Во время профазы между конъюгированными хромосомами может происходить обмен гомологичными участками - кроссинговер (рис. 13). К концу профазы гомологичные хромосомы разъединяются в области центромер, оставаясь соединенными в области плеч.
52
цовских по отношению к ядру А) хромосом,
Рис. 12. Схема поведения хромосом «условной» клетки при мейозе.
А - изображено диплоидное ядро клетки, содержащее 3 пары хромосом (1 и 1' - первая пара гомологичных хромосом, 2 и 2' - вторая пара, 3 и 3' - третья пара). 2п = 6. Происходящие от материнского организма 3 хромосомы - светлые {цифры без штриха} парные им гомологичные хромосомы, происходящие от отцовского организма, зачернены {цифры со штрихом} Г омологичные хромосомы обозначены общим номером. Каждая хромосома состоит из двух хроматид. Удвоения хромосом перед первым делением не происходит. Б - слияние гомологичных хромосом. На этой стадии гомологичные хромосомы обмениваются между собой отдельными участками. В результате происходит некоторое перераспределение материнского и отцовского наследственного материала между хромосомами (кроссинговер). В -образуются нити веретена, прикрепляющиеся к хромосомам, исчезает оболочка ядра. Г - гомологичные хромосомы расходятся к противоположным полюсам клетки, у полюсов оказывается по одной гомологичной хромосоме из каждой пары, общее число хромосом у каждого полюса вдвое меньше, чем в исходном ядре А. Д -образуются два ядра с гаплоидным набором хромосом в каждом; в одно из них попало больше "бабушкиных" (светлых, материнских по отношению к ядру А) и меньше "дедушкиных" (зачерненных, от-в другое - наоборот; таким образом, новые
ядра не вполне тождественны друг другу по составу своего наследственного вещества; их различие обусловлено также и кроссинговером, происходящим с хромосомами на стадии В. Е и Ж - митотическое деление каждого из гаплоидных ядер (в интерфазе второго деления хромосомы удваиваются). Д: продольное расщепление каждой хромосомы, расхождение хромосом к полюсам, образование двух гаплоидных ядер из каждого ядра Д. В итоге появилось 4 клетки с гаплоидными ядрами вместо одной клетки с диплоидным набором хромосом (из Арронет, 1974)
В метафазе I деления, в период созревания, конъюгированные хромосомы располагаются по экватору клетки. В этот момент спирализация хромосом достигает максимума. Центромеры обращены к полюсам.
53
Рис. 13. Схема кроссинговера во время мейоза: 1 - две гомологичные хромосомы, каждая из которых состоит из двух сестринских хроматид; 2 - гомологичные хромосомы сплелись друг с другом; 3 - каждая хромосома расщепляется на составляющие ее хроматиды; хромосомы начинают отталкиваться друг от друга и расходиться, но в некоторых точках (на рисунке в четырех) они остаются связанными; в центральной петле сдвоены сестринские хроматиды, а в обеих крайних петлях - не сестринские; точки, где возможен кроссинговер, показаны стрелками; 4 - хроматиды обменялись участками, и хромосомы, претерпев как бы взаимную гибридизацию, расходятся; 5 - две разошедшиеся, но уже "гибридные" ( в отличие от
показанных на 1) хромосомы (из Арронет, 1974)
В анафазе гомологичные хромосомы расходятся к полюсам. Следовательно, из каждой пары гомологичных хромосом в дочернюю клетку попадает только одна. Число хромосом уменьшается в два раза и образовавшиеся клетки содержат одинарный, гаплоидный набор хромосом. Поэтому первое мейотическое деление носит название редукционного (от лат. «reduction -возвращение). Следует помнить, что в период созревания вступают диплоидные клетки, удвоившие количество ДНК еще в периоде размножения (в процессе митоза). Их хромосомный набор можно обозначить как 2п4с. После расхождения в анафазе I деления хромосомы остаются удвоенными. В результате редукционного деления клетки содержат гаплоидное число хромосом и двойное количество ДНК (1п2с\ которое приводится в соответствие с хромосомным набором во время следующего, II мейотического деления. В анафазе этого деления к полюсам расходятся хроматиды и образующиеся клетки получают хромосомный набор, равный 1п1с.
От митотического деления соматических клеток мейоз отличается рядом особенностей.
1. Профаза I мейотического деления занимает очень большой промежуток времени. Во время профазы гомологичные хромосомы конъюгируют и могут обмениваться участками (кроссинговер, (рис. 13)).
2. В метафазе I мейотического деления по экватору клетки располагаются не отдельные хромосомы, а пары конъюгированных хромосом.
3. Между I и II мейотическими делениями не синтезируется ДНК.
4. Силы отталкивания в мейозе проявляются в области центромер, в то время как в митозе вначале разъединяются плечи хроматид.
Биологическая роль мейоза, также как и митоза, заключается в поддержании постоянства хромосомного набора, свойственного данному виду организмов. При оплодотворении - слиянии половых клеток - в зиготе восстанавливается двойной набор хромосом. В то же время, мейоз обеспечивает и варьирование клеток у потомков.
54
АНАТОМИЯ ДРЕВЕСНЫХ РАСТЕНИЙ
Краткая история анатомии растений
Следуя тексту из учебника ботаники для университетов, написанного заслуженным профессором МГУ им. М.В. Ломоносова Л.И. Лотовой, о первых этапах развития анатомии растений можно сказать ниже следующее.
Основы учения о тканях были заложены в XVII веке, когда в научных исследованиях стали использовать микроскоп, предшественником которого, по-видимому, был оптический инструмент, сконструированный Кеплером (1571-1630), дававший увеличенное обратное изображение объекта.
Первым открытием в мире растений, сделанным с помощью микроскопа было открытие клетки англичанином Робертом Гуком (1633-1703). Рассматривая под микроскопом собственной конструкции срез бутылочной пробки, он обнаружил в ней структуру, напоминающую пчелиные соты. Эти соты Гук назвал клетками.
Открытие клетки стимулировало изучение внутреннего строения растений - их анатомии (от греч. anatome - рассечение). Основоположники этой науки - итальянский врач Марчелло Мальпиги (1628-1694) и англичанин, секретарь Лондонского Королевского общества Неймия Грю (1641-1712). Работая независимо один от другого, оба исследователя одновременно (1617) представили монографии по анатомии растений.
М. Мальпиги описал внутреннее строение стебля, ветвей, листьев. Он обнаружил устьица, спиральные сосуды, которые были названы им трахеями по аналогии с органами дыхания насекомых. Заслуга М. Мальпиги состоит также в доказательстве нисходящего тока веществ, осуществляемого корой.
Н. Грю изучил строение корня, расположение проводящих тканей в осевых органах растений и обосновал положение о том, что все органы растений имеют всегда определенное, типичное для них строение. Н. Грю ввел в литературу термин «ткань», считая ее сложным сплетением тонких нитей, похожим на кружево.
Признавая клеточное строение всех органов растений, оба ученых не знали, однако, как устроена клетка. Лишь в 1812 г., применив метод мацерации (разрыхление ткани с разделением ее на отдельные клетки) И. Молден-гауэр (1766-1827) доказал индивидуальность каждой клетки, а в 1830 г. Ф. Мейен (1804-1840) сформулировал понятие о клетке как о пространстве, ограниченном оболочкой, которую он назвал мембраной. Эти исследования имели важное значение для дальнейшего изучения тканей.
В 1807 г. Г.Ф. Линк (1767-1851) ввел в литературу понятие о паренхиматических и прозенхиматических клетках. Первую группу составляют клетки шаровидные (изодиаметрические) или слегка призматические, вторую -более или менее длинные, с заостренными концами. Ткани, состоящие из
55
этих клеток, стали называть соответственно паренхимой и прозенхимой. (Кстати, этими терминами пользовался и Н. Грю).
Ботаники второй половины XIX века А. де Бари (1831-1888) и Ф. ван Тигем (1839-1914) называли тканью совокупность клеток одинакового строения, независимо от того, разбросаны они в теле растения или составляют компактные группы.
Классифицируя ткани, Ф. ван Тигем обращал внимание на наличие в клетках живого содержимого. По этому признаку он разделил ткани на живые и мертвые. К последним он отнес ткани, выполняющие в растении опорную роль и участвующие в проведении веществ. Условность такого разделения тканей очевидна: мертвые ткани на ранних стадиях развития состоят из живых клеток, а клетки живых тканей со временем могут терять содержимое и переходить в разряд мертвых.
Ю. Сакс (1832-1897) различал в растениях покровную, пучковую (проводящие пучки, например, в листьях) и основную ткани. Этой классификацией пользуются и в настоящее время. Неудобство ее состоит в том, что в одну ткань часто попадают клетки, разные не только по строению, но и по выполняемым функциям. Так, пучковую ткань составляют клетки, проводящие воду и растворы органических веществ, а также клетки с толстыми одревесневшими оболочками, играющие механическую роль. Еще большее число функционально разных клеток объединяет основная ткань. Нередко к тому же участки этой ткани разрознены и топографически (по своему расположению) и не составляют непрерывную систему. Все это усложняло определение термина «ткань» и создавало трудности в отношении классификации тканей.
Наиболее удачное определение тканей, которым пользуются и в настоящее время, предложено в конце XIX века Ф. Габерландтом (1826-1878). Ткань - это устойчивый комплекс клеток, обладающих одним или несколькими сходными признаками: физиологическими, морфологическими, топографическими и общностью происхождения. Каждый из этих признаков может быть использован для классификации тканей, но, учитывая, что только согласованное функционирование разных тканей может обеспечить осуществление всех присущих любому организму жизненных процессов, наиболее целесообразна их группировка по анатомо-физиологическому принципу. Ткани и локальные структуры, выполняющие одинаковые функции, Ф. Га-берландт объединил в 9 систем, выделив покровную, механическую, проводящую и прочие, наблюдаемые у растений системы.
Интерес ученых к анатомии растений и темпы развития этой науки в течении времени были разными. Саму анатомию породило, как уже было сказано, изобретение микроскопа. По мере совершенствования этого прибора появлялись новые открытия. Однако публикации основных работ великого шведского ученого, основателя подлинно научной систематики живых организмов Карла Линнея (1735, 1751), отвлекло внимание большинства ботаников от микроскопа и направило их творческие поиски на систематизацию мира растений в основном по внешним морфологическим признакам.
56
Дальнейшее развитие анатомия растений получила только в начале 19 века. Английский ботаник Р. Броун в 1831 обнаружил в клетке ядро, что вместе с работами немецкого ботаника М. Шлейдена сыграло большую роль в создании клеточной теории, творцом которой был немецкий биолог Т. Шванн (1839). С этого времени всё большее внимание уделяется изучению содержимого растительной клетки, названной чешским учёным Я. Э. Пуркине протоплазмой (1839). Ряд свойств протоплазмы изучил и описал немецкий учёный X. Моль (1846). Физиологический принцип в изучении строения растений применил немецкий ботаник Г. Габерландт. Большой вклад в анатомию растений сделали французский биолог Ф. Э. ван Тигем, немецкие биологи А. де Бари, К. Негели, К. Санио, И. Ханштейн, С. Швенденер и др. Значительную роль в развитии этой науки сыграли работы русских ботаников И. В. Баранецкого, С. П. Костычева, В. Р. Заленского, В. Ф. Раздорского, В. Г. Александрова, А.А. Яценко-Хмелевского, Л.И. Лотовой и др.
Весьма важную роль сыграло изобретение электронного микроскопа. Этот прибор позволил уточнить многие представления о строении растений, полученные ранее с помощью светового микроскопа. Особо важную роль электронный микроскоп сыграл в цитологии - науке о клетке.
Ткани высших растений
Тело растения, как мы это уже выяснили, состоит из морфологически четко очерченных единиц - клеток, каждая из которых заключена в свою собственную клеточную оболочку и соединена с другими клетками посредством межклеточного цементирующего вещества. В этой связанной воедино массе отдельные группы клеток различаются между собой либо в структурном, либо в функциональном, либо в том и другом отношениях.
По широко распространенной классификации ткани сосудистого растения сгруппированы на основе их топографической непрерывности в три системы тканей: покровную, проводящую и основную.
Покровная система включает эпидерму - первичный наружный защитный слой, покрывающий тело растения, и перидерму - защитную ткань, которая замещает эпидерму главным образом в тех частях растения, которые претерпевают вторичные утолщения. Проводящая система состоит из двух типов проводящих тканей - флоэмы, или луба (проведение питательных веществ), и ксилемы, или древесины (проведение почвенных растворов).
Основная система включает ткани, которые в определенном смысле составляют основу растения, но в то же время обнаруживают различную степень специализации. Главными из тканей, относящихся к основным, являются: паренхима во всем ее разнообразии, колленхима - толстостенная опорная ткань, родственная паренхиме, и склеренхима - главная опорная ткань с толстыми твердыми, часто одревесневшими стенками.
В пределах одного растения различные ткани распределены характерным образом в зависимости от того, в какой части растения они находятся и к какому виду принадлежит данное растение. По существу, характер их рас
57
пределения сходен в том отношении, что проводящая ткань всегда бывает погружена в основную, а покровная ткань составляет наружный защитный слой тела растения. Главные изменения в характере распределения тканей связаны с относительным положением проводящей и основной тканей. У двудольных, например, проводящая ткань стебля образует полый цилиндр, а основная ткань находится внутри цилиндра (сердцевина) и вокруг него - между проводящими и покровными тканями (первичная кора). В листе проводящая ткань представляет собой проводящий пучок, состоящий из ксилемы и флоэмы, погруженный в основную ткань, дифференцированную здесь как мезофилл. В корне проводящий цилиндр может не содержать сердцевины, но первичная кора в нем присутствует.
Клетки и ткани растения обычно развиваются из зиготы - оплодотворенной яйцеклетки - через промежуточные стадии развития, осуществляемые в зародыше. Эмбриональная стадия, однако, не прекращается полностью после того, как зародыш разовьется во взрослое растение. Растения обладают уникальным свойством неограниченного роста, обусловленным присутствием в них эмбриональных тканевых зон - меристем, в которых образование новых клеток продолжается в то время, когда другие части растений уже достигли зрелого состояния. Меристемы, находящиеся на кончиках корней и верхушках побегов - апикальные меристемы, - образуют клетки, производные которых дифференцируются в новые части корня и побега. Этот рост называется первичным. Тело растения, сформировавшееся в результате первичного роста, представляет собой первичное тело растения.
У многих растений стебли и корни нарастают в толщину путем добавления к первичному телу проводящих тканей. Такое утолщение обеспечивается камбием и называется вторичным ростом. Обычно вторичный рост сопровождается также появлением перидермы, которую формирует особая меристема - феллоген или пробковый камбий. Камбий и феллоген получили название - латеральные (боковые) меристемы, вследствие того, что они расположены параллельно боковым поверхностям стебля и корня.
Далее приведем краткую характеристику типов клеток и тканей.
Эпидерма
На первых этапах роста эпидерма образует непрерывный слой на поверхности первичного тела растения. Эпидермальные клетки, составляющие основную часть эпидермы, разнообразны по форме, но чаще они бывают таблитчатыми. К эпидермальным клеткам относятся также замыкающие клетки устьиц и различные трихомы (одноклеточные или много клеточные выросты эпидермальных клеток), в том числе и корневые волоски. Главная отличительная особенность эпидермальных клеток надземных частей растения - наличие кутикулы20 на наружных стенках клеток. Эпидерма обеспечивает механическую защиту и участвует в регулировании транспирации и газообмена.
20 Кутикула - слой жирового вещества. Кутина, покрывающего сплошной пленкой поверхность надземных органов многих растений (главным образом листьев, стеблей, некоторых плодов).
58
В стеблях и корнях, характеризующихся вторичным ростом, эпидерма обычно замещается перидермой.
Перидерма
Перидерма состоит из пробки (феллемы), пробкового камбия (феллоге-на) и феллодермы (рис. 14).
Рис. 14. Перидерма и ее расположение по отношению к проводящей ткани на поперечном (А) и продольном (Б) разрезах части стебля (схема): 1 - зона непроводящего луба, в которой закладывается перидерма; 2 - слои перидермы; 3 - проводящая флоэма; 4 -камбий; 5 - ксилема; 6 - лучи (из Лотова, 2007)
Феллоген образует на своей внешней стороне пробку, а на внутренней - феллодерму. Феллодерма, состоящая из паренхимных клеток, либо образуется в небольшом количестве, либо вообще отсутствует. Клетки пробки имеют обычно таблитчатую форму, плотно сомкнуты, в зрелом состоянии лишены протопластов и имеют опробковевшие стенки. Опробковение происходит вследствие отложения в оболочках клеток суберина - смеси производных жирных кислот. Суберин делает пробку водонепроницаемой. Полости пробковых клеток березы заполняются белым веществом - бетулином, по составу близким суберину.
Паренхима
Паренхимные клетки образуют сплошную ткань в первичной коре стебля и корня и в мезофилле листа. Они встречаются также в виде вертикальных тяжей и лучей в проводящих тканях. По происхождению они первичны в коре, сердцевине и листе, в проводящих тканях они могут быть первичными или вторичными. В типичном случае паренхимные клетки представляют собой живые клетки, способные к росту и делению. Они разнообразны по форме, часто имеют вид многогранника, но могут быть также звездчатыми или сильно удлиненными. Для паренхимных клеток травянистых растений характерны первичные оболочки, но встречаются и вторичные. Паренхимные клетки листа и молодых побегов имеют хлоропласты и связаны с процессом фотосинтеза, они выполняют и многие другие функции.
59
Паренхимные клетки в проводящей ткани древесных растений выполняют функции запасания и проведения продуктов метаболизма.
Колленхима
Клетки колленхимы образуют тяжи, или непрерывные цилиндры, вблизи поверхности первичной коры в стеблях и черешках и вдоль жилок вегетативных листьев. В корнях она обнаруживается редко. Колленхима представляет собой живую ткань, близко родственную паренхиме, фактически ее обычно и рассматривают как форму паренхимы, специализированную в качестве опорной ткани молодых органов.
Склеренхима
Склеренхимные клетки либо образуют сплошную ткань, либо располагаются в виде небольших групп или поодиночке среди других клеток. Склеренхимные клетки имеют толстые вторичные оболочки, часто одревесневшие, и в зрелом состоянии обычно лишены протопластов. Различают два типа склеренхимных клеток - склереиды и волокна. По своей форме склереиды варьируют от многогранных до удлиненных и могут быть разветвленными. Волокна, как правило, представляют собой тонкие удлиненные клетки.
Ксилема
Клетки ксилемы образуют сложную как в структурном, так и в функциональном отношении ткань, которая вместе с флоэмой проходит, не прерываясь, по всему телу растения. Эта ткань состоит из нескольких типов клеток и служит для проведения воды с растворенными в ней минеральными веществами, отложения питательных веществ в запас и для опоры. По своему происхождению ксилема может быть первичной или вторичной. Первичная ксилема - продукт деятельности первичных образовательных тканей, вторичная - образована камбием. Вторичную ксилему обычно называют русским синонимом - древесина. Основными проводящими воду элементами являются трахеиды и членики сосудов. Отдельные членики смыкаются конец в конец, образуя сосуды. Запасные вещества откладываются в паренхимных клетках древесины, которые расположены вертикальными рядами, а также в лучевой паренхиме вторичной ксилемы. Механические элементы представлены волокнами и склереидами.
Флоэма
Флоэма представляет собой сложную ткань, состоящую из нескольких типов клеток. Эта ткань вместе с ксилемой проходит, не прерываясь, по всему телу растения и по своему происхождению может быть первичной или вторичной. Флоэма является опорной, проводящей питательные вещества и запасающей тканью. Главными проводящими элементами служат ситовидные клетки и членики ситовидных трубок, причем в зрелом состоянии и те, и другие лишены ядер. Членики ситовидных трубок соединяются конец в конец, образуя ситовидные трубки, и связаны с паренхимными клетками, назы
60
ваемыми клетками-спутниками. Другие паренхимные клетки флоэмы располагаются вертикальными рядами. Вторичная флоэма также содержит паренхиму в виде лучей. Опорные клетки представлены волокнами и склереидами.
Пространственно ксилема - основная водопроводящая ткань сосудистых растений объединена с флоэмой - тканью, проводящей пластические вещества. Обе ткани вместе именуются сосудистой, или проводящей, тканью (или тканями). Комплекс ксилемы и флоэмы образует непрерывную проводящую систему, проходящую через все органы растения, включая все ответвления стебля и корня.
По происхождению и развитию целесообразно различать первичные и вторичные проводящие ткани. Первичные ткани дифференцируются в процессе развития первичного тела растения. Меристемой, обеспечивающей формирование первичных проводящих тканей, является прокамбий. Вторичные проводящие ткани образуются в течение второй важной стадии развития растения, когда в связи с латеральным (боковым) приростом новых тканей происходит увеличение толщины осевых частей растения - стебля и корня -и их крупных ответвлений. Это достигается в результате активности камбия. Вторичный рост отсутствует у небольших по размерам однолетних двудольных и у большинства однодольных растений.
СТРОЕНИЕ И ФУНКЦИИ СТЕБЛЯ
Согласно представлениям, сложившимся в морфологии, вегетативное тело сосудистых растений состоит из трех органов: листа, стебля и корня. Однако это подразделение довольно условно и в первую очередь очевидна условность в разделении стебля и листа, так как большинство тканей у них тесно связаны между собой. В результате многие морфологи и анатомы объединяют эти органы в одном понятии - побег, означающем облиственный стебель. И все-таки, несмотря на тесную связь между стеблем и листом у большинства высших растений эти два органа хорошо отличаются друг от друга, что требует их отдельного рассмотрения.
У очень многих растений стебель характеризуется ветвистостью, т.е. наряду с главной осью возникают ответвления - ветви. В подавляющем большинстве случаев анатомическое строение ветвей не отличается от строения главной оси и все они включаются в общее понятие - стебель. Этот термин в разговорной речи, а иногда и в научной литературе применяется только к травянистым растениям, а главную ось древесных растений, лишенную листьев, называют стволом. Тем не менее, ствол вместе с ветвями - тоже стебель.
Стебель обладает длительным ростом в длину за счет верхушечной меристемы. В первичной меристеме конуса нарастания стебля различают два комплекса клеток (рис. 15). На поверхности его находится туника, состоящая из одного или нескольких слоев клеток, делящихся в основном периклиналь-но (параллельно поверхности); под туникой расположен многослойный корпус, клетки которого делятся в различных направлениях. Из клеток туники
61
формируется эпидермис, а клетки корпуса дифференцируются в остальные постоянные ткани стебля. Если туника состоит из нескольких слоев клеток, внутренние слои их дают начало также первичной коре.
Рис. 15. Почка и верхушечная меристема побега. А - схема продольного разреза вегетативной почки: 1 - апекс, включающий апикальную меристему; 2 - зачатки листьев; 3 - зачатки пазушных почек; 4 - тяжи прокамбия. Б -продольный разрез верхушки побега гороха (Pisuin)c верхушечной (апикальной) меристемой: 1 - антиклинальное деление в одной из клеток апикальной меристемы; 2 - инициальный слой корпуса; 3 - двухслойная туника; 4 - листовой примордий (от слова "прима" - первый); 5 - корпус; 6 - сердцевинная меристема (из Лотова, 2007 с добавлением)
Анатомическое строение стебля тесно связано с выполняемыми им функциями. Различают первичное и вторичное строение стебля.
Первичное строение стебля
Первичное тело формируется в результате дифференциации клеток первичной меристемы конуса нарастания. Проводящие пучки образуются из особых участков меристемы - прокамбия. Прокамбий возникает вследствие того, что некоторые меристематические клетки конуса нарастания начинают делиться преимущественно в продольном направлении (периклинально, т.е. параллельно поверхности), в то время как деление их поперечными перегородками замедляется. В результате появляются вытянутые, делящиеся клетки, заполненные густым зернистым содержимым, которые могут быть расположены в виде отдельных тяжей или сплошного цилиндра. В первичном строении стебля травянистых однодольных и двудольных растений имеются существенные различия.
Однодольные растения. Характерной особенностью однодольных растений являются закрытые проводящие пучки, такие пучки не содержат собственной образовательной ткани вследствие чего сами пучки и стебли не могут утолщаться. Наблюдающийся у некоторых пальм рост стебля в толщину осуществляется не за счет камбия, а за счет специального кольца утолщения, которое образуется из клеток основной паренхимы.
Проводящие пучки у однодольных растений на поверхности поперечного среза стебля расположены по спирали.
62
В большинстве случаев стебель у однодольных растений состоит из следующих тканей: эпидермиса, склеренхимы и основной паренхимы, в которой проходят проводящие пучки. В стебле однодольных растений отсутствует четкое разделение на первичную кору и осевой, или центральный, цилиндр, так как кора очень плохо выражена и в виде узкой полосы расположена на периферии стебля. Проводящие пучки обычно окружены кольцом склеренхимы.
У некоторых однодольных растений (кукуруза, пальмы) из каждого листа в стебель выходит большое число пучков, которые распределяются по всему стеблю. Стебель большинства злаков (ржи, пшеницы, ячменя и др.) представляет собой соломину. На поверхности соломины находится эпидермис, за ним расположена склеренхима, состоящая из толстостенных клеток. Между эпидермисом и склеренхимой находятся участки хлорофиллоносной паренхимы. Далее к центру расположена паренхимная ткань, состоящая из крупных клеток, оболочки которых могут одревесневать. Среди клеток паренхимы проходят закрытые проводящие пучки. В центре стебля находится крупная полость, к которой примыкают клетки основной ткани.
Первичное строение сохраняется у однодольных растений в течение всей жизни, на всем протяжении стебля.
Двудольные растения. У двудольных растений разделение стебля на эпидермис, первичную кору и центральный цилиндр, как правило, довольно четко выражено. Первичная кора в основном состоит из паренхимной ткани, в клетках которой часто содержатся хлоропласты. В состав первичной коры входят также механические ткани - колленхима и склеренхима. Колленхима обычно залегает непосредственно под эпидермисом, а склеренхима располагается несколько ближе к центру стебля в виде сплошного кольца (тыква) или отдельными участками (подсолнечник).
Глубже лежащие клетки паренхимы бесцветны, так как хлоропластов не содержат. Самый внутренний слой первичной коры представлен эндодермой, которая часто дифференцируется в крахмалоносное влагалище. В клетках его содержатся многочисленные мелкие крахмальные зерна, которые не потребляются даже при значительном недостатке питательных веществ. Некоторые авторы считают, что крахмалоносное влагалище обусловливает вертикальное положение стебля.
Центральный цилиндр стебля начинается с перицикла, непосредственно примыкающего к эндодерме. В перицикле многих растений могут зарождаться придаточные корни, придаточные почки, вторичные меристемы. К центру от перицикла располагаются проводящие пучки, состоящие из флоэмы, ксилемы и пучкового камбия.
В стебле двудольных растений при первичном строении наблюдаются следующие анатомические особенности, отличающие его от стеблей однодольных. У двудольных кора четко отделяется от сердцевины; у однодольных основная ткань представлена паренхимой без разделения на кору и сердцевину. У двудольных проводящие пучки располагаются кольцом, прилегающим изнутри к эндодерме; у однодольных пучки разбросаны в тоще ос-
63
новной ткани. Проводящие пучки у двудольных открытые, т. е. содержат камбий и имеют возможность ограниченного вторичного роста; у однодольных пучки закрытые, камбий в них отсутствует.
Первичное строение стебля у двудольных растений наблюдается только в наиболее молодой части стебля - под верхушечной почкой, а затем постепенно заменяется вторичным строением.
Прокамбий
У семенных растений развитие стеблевого прокамбия инициируют зачатки листьев. В основании молодого листового зачатка образуется небольшой очаг делящихся клеток. Мелкие клетки, образовавшиеся в результате этих делений, постепенно вытягиваются в длину, становятся прозенхимны-ми, заостренными на концах, что характерно для клеток прокамбия. В дальнейшем, заложившийся в виде тяжа прокамбий развивается как акропетально - к верхушке листового бугорка, так и базипетально - вниз по первичной коре стебля, которую он пересекает под некоторым углом к его продольной оси и постепенно приближается к так называемому образовательному кольцу. В основании каждого листового зачатка у голосеменных и двудольных покрытосеменных растений закладывается от одного до пяти - семи (редко больше) прокамбиальных тяжей, каждый из которых дает начало одному проводящему пучку, состоящему из флоэмы и ксилемы. Совокупность всех пучков называют листовым следом. Иногда следом называют каждый из этих пучков.
Удлиняющиеся тяжи прокамбия, соответствующие будущим пучкам листовых следов, приблизившись внутренними концами к образовательному кольцу, стимулируют деление его клеток, формирующих стеблевой прокамбий. Таким образом стеблевой прокамбий оказывается непосредственным продолжением прокамбия листового. В образовательное кольцо входят про-камбиальные тяжи из листовых зачатков разного возраста.
Появление в образовательном кольце отдельных тяжей прокамбия приводит к его разделению на будущие пучковые и межпучковые зоны. Клетки последней дифференцируются в паренхимные клетки сердцевинных лучей, соединяющих сердцевину стебля с первичной корой.
Из тяжей прокамбия образуются проводящие пучки, включающие первичную флоэму и первичную ксилему. При этом клетки прокамбия дифференцируются в ткани проводящего пучка.
У голосеменных и двудольных покрытосеменных растений еще до окончания дифференциации прокамбия в первичные проводящие ткани, из среднего слоя его клеток вычленяются клетки камбия. При этом клетки прокамбия делятся периклинально, то есть тангентальными перегородками, параллельными поверхности стебля. В результате этих делений образуются характерные для камбия более или менее таблитчатые в поперечном разрезе клетки, расположенные в один слой.
Клетки камбия, делясь периклинально, отчленяют от себя наружу и внутрь клетки, дифференцирующиеся в элементы вторичных флоэмы и ксилемы. Начавшееся вторичное утолщение стебля некоторое время идет одно
64
временно с продолжающейся дифференциацией клеток прокамбия в элементы первичных проводящих тканей. После ее окончания увеличение объема проводящих тканей осуществляется только камбием.
Производные прокамбия (ксилема и флоэма) называются первичными, производные камбия - ксилема (древесина) и флоэма (луб) - вторичными проводящими тканями.
Вторичное строение стебля
Строение травянистого стебля
Вторичное строение появляется в стеблях двудольных растений в результате деятельности вторичной меристемы. Переход ко вторичному строению начинается с появления межпучкового камбия (рис. 16), начало которому дают клетки паренхимной ткани сердцевинных лучей, расположенных между пучками. Межпучковый камбий соединяется с пучковым камбием, в результате чего получается сплошное деятельное кольцо образовательной ткани. Участки межпучкового камбия образуют новые проводящие пучки, постепенно все пучки соединяются вместе, вследствие чего возникают сплошные слои флоэмы и ксилемы. В центре стебля из клеток основной ткани формируется сердцевина, а после ее разрушения, образуется полость. На этом развитие стебля заканчивается, так как у растений умеренных широт травянистый стебель однолетний и отмирает в конце вегетационного периода, не достигнув большой толщины.
Внутр и пучковый камбий
Межпучковый камбий—
Эпидермальный волосок---------—
(у подсолнечника многоклеточный)
Кутикула------------
(образована из кутина)
Эпидермис —---------
(однослойный)
Кора (в наружных Г Колленхима-^ клетках могут И
быть хлоро пласты) ^ПаРснхима ц /
Проводящий пучек Б
Й &
с с
’S >4 И >4
>Х
4 X Т
Кутикула
(как у двудольных)
Эпидермис
(как у двудольных)
Гиподерма
(колленхима, в наружных клетках могут быть хлоро пласты)
Основная ткань
(паренхима, нет деления на кору и сердцевину)
Сосудистые пучки (многочисленные, разбросанные, к центру крупнее)
Эндодерма__________"
(толщиной в одну клетку; клетки могут содержать крахмал)
Сердцевина-(паренхима)
Перицикл
(волокна склеренхимы)
Флоэма (ситовидные трубки, клетки-спутницы, паренхима, волокна) Внутри пучковый камбий Метаксилема
(зрелые сосуды ксилемы, волокна и паренхима)
Поперечный разрез проводящего пучка
Протоксилема эндархная, т. е. находится ближе к центру, чем метаксилема; сосуды с кольчатыми и спиральными утолщениями
Перицикл или обкладка----
проводящего пучка (склеренхима, окружающая пучек)
Фл оэ м а----------------
(только ситовидные трубки и клетки-спутницы) Метаксилема —— (два больших сосуда) Протоксилема
Поперечный разрез проводящего пучка
Лизигенный канал (образовался в результате разрушения протоксилемы)
Рис. 16. Отличия анатомического строения стебля у типичных представителей травянистых двудольных (А) и однодольных (Б) растений на примере подсолнечника
65
(Helianthus annuus) - двудольное и кукурузы (Zea mays) - однодольное (из Грин Н., У. Стаут, Д. Тейлор, 1990)
Строение стебля древесного растения
Стебли древесных растений, как правило, имеют непучковое строение. Древесный стебель в отличие от травянистого является многолетним и может достигать возраста в десятки, сотни и даже тысячи лет. В связи с этим утолщение древесного стебля продолжается неопределенно долго. Анатомические особенности строения древесного стебля заключаются в формировании на его поверхности перидермы, которая сменяет эпидермис, и в образовании у растений умеренной зоны в той или иной мере выраженных годичных колец в древесине.
В древесном стебле обычно выделяют кору, камбий (рис. 17 (2, 2а)), древесину (ксилему) и сердцевину.
Камбий
Делясь тангентально, то есть параллельно поверхности органа, клетки прокамбия, имеющие многоугольные очертания поперечных сечений, образуют клетки камбия.
Рис. 17. Сектор выпила из четырехлетнего стволика сосны (Pinus sylvestris). П - плоскость поперечного среза; Р -плоскость радиального среза; Т - тангентальная (тангенциальная) поверхность, кора (вторичная флоэма с перидермой) на рисунке от-делилена по камбию; 1 -флоэма (луб); 2 - камбий; 2а - освобожденная от коры поверхность камбиального слоя; 3 - поздняя часть годичного прироста древесины; 4 - вертикальные смоляные ходы; 5 - прирост четвертого года; 6 - лучи; 7 - ранняя часть годичного прироста древесины; 8 -граница годичного прироста; 9 - прирост третьего года; 10 - прирост второго года; 11 - прирост первого года; 12 - сердцевина; 13 -
66
поверхность ствола (наружная часть перидермы), перидерма; 14 - флоэмные (лубяные) лучи (из Molisch, Hofler, 1965)
Слой клеток, возникших непосредственно из клеток прокамбия, называют инициальным слоем камбия. Клетки последнего, делясь периклиналь-21 но , отчленяют наружу и внутрь такие же прозенхимные клетки, располагающиеся на поперечных срезах радиальными рядами. Самые молодые производные инициального слоя, примыкающие к нему с двух сторон и сохраняющие способность к дальнейшим тангентальным делениям, называют материнскими клетками флоэмы (луба) и материнскими клетками ксилемы (древесины). Инициальный слой камбия вместе с материнскими клетками будущих вторичных проводящих тканей составляют камбиальную зону (рис. 18).
Клетки, образованные материнскими клетками луба и древесины, постепенно разрастаются в радиальном направлении и затем дифференцируются в элементы вторичной древесины и вторичного луба. Так как клетки инициального слоя камбия отчленяют материнских клеток древесины больше, чем материнских клеток луба, и первые делятся чаще, то и древесины в многолетних органах всегда бывает больше, чем луба.
Созревающая флоэма
Радиально расширяющиеся клетки флоэмы
Делящиеся клетки флоэмы
Камбиальные инициали
Делящиеся клетки ксилемы
Радиально расширяющиеся клетки ксилемы
Созревающие клетки ксилемы
21 Периклинальные деления - деление клеток в плоскости, параллельной поверхности ткани или органа, приводящее к образованию двух смежных слоев.
67
10 10 10 10 10
и И И И И
12 12 12 12 12
Рис. 18. Схема камбиальной зоны и прилегающих к ней клеток в стебле древесного растения при формировании флоэмы и ксилемы в плоскости поперечного среза (из Лотова, 2007, с изменениями)
У древесных растений инициальный слой камбиальной зоны состоит из двух типов клеток: длинных, прозенхимных, называемых веретеновидными инициалями, и коротких, изодиаметрических, расположенных на продольных срезах в один или несколько слоев разной толщины. Эти клетки называют лучевыми инициалями. Комплексы лучевых инициалей на продольных тан-гентальных срезах, сделанных в плоскости, параллельной поверхности ствола или ветви, имеют разные очертания. Вследствие синхронных делений всех клеток каждого комплекса внутрь и наружу от инициального слоя образуются клетки, наращивающие лубяную или древесинную части лубо-древесиных лучей. Поэтому протяженность лучей в радиальном направлении с возрастом стебля увеличивается.
Веретеновидные клетки камбия у разных древесных пород имеют разную длину. У хвойных она варьирует от 2 до 5 мм. У лиственных древесных пород длина веретеновидных клеток колеблется от 0,1 до 4,5 мм. По сравнению с длиной ширина клеток очень мала, размеры их поперечника не превышают обычно 20-30 мкм, а поперечники лучевых инициалей обычно еще меньше.
По мере утолщения ствола длина окружности камбиальной зоны увеличивается не только путем растяжения клеток в тангентальном направлении, но и вследствие их делений. Таким образом, окружность ствола с возрастом увеличивается, а число клеток на единицу длины окружности остается постоянным за счет регулярно происходящих делений. Плоскость, в которой происходят такие деления, называемые антиклинальными", под тем или иным углом пересекает ось стебля.
Очень длинные веретеновидные клетки делятся перегородками, расположенными косо к продольной оси клетки. Такое деление называют антиклинальным ложно-поперечным (рис. 19). В результате возникают две клетки, расположенные одна под другой, при этом длина каждой из них немного больше половины длины исходной клетки. Образовавшиеся клетки затем удлиняются путем интрузивного роста, внедряясь между соседними. Так как интенсивность и продолжительность роста каждой из этих клеток разные, обычно одна из них по размерам превышает другую, поэтому их концы находятся на разных уровнях. Такой камбий называют неярусным. Он характерен для голосеменных и многих покрытосеменных растений, например, для яблони, груши, березы (рис. 20 А).
22
Антиклинальные деления - деления в плоскости перпендикулярной поверхности органа.
68
Рис. 19. Антиклинальные ложнопоперечные (а, б, в) и антиклинальное радиальное (г) деления веретеновидных клеток камбия: А -деление веретеновидных клеток неярусного камбия; Б -изменение угла наклона перегородок при ложнопоперечном делении, связанное с эволюционным укорочением веретеновидных клеток; В - деление веретеновидных клеток ярусного камбия. Камбиальные клетки показаны в плоскости тангенталь-ного среза (из Лотова, 2007)
Считается, что в процессе эволюции веретеновидные клетки камбия укорачивались, а вместе с этим изменялись и углы наклона перегородок при их делениях (рис. 19 Б). В коротких инициа-лях они ориентированы радиально, таким образом, деление становится строго продольным и приводит к образованию двух равных по размерам клеток, расположенных одна рядом с другой (рис. 19 В), Концы клеток в этом случае находятся почти на одном уровне. Такой камбий называют ярусным (рис. 20 А). Он характерен для ясеня, робинии лжеакации и других лиственных по
род.
Рис. 20. Ярусный (А - белая акация) и неярусный (Б - груша) камбий на тангенталь-ных срезах: 1 - веретеновидные инициали: 2 -лучевые инициали (из Лотова, 2007)
Деления, приводящие к образованию новых веретеновидных инициа-лей, происходят довольно часто. Появление новых инициалей в камбиальном слое приводит к формированию новых
радиальных рядов клеток вторичных древесины и луба.
Увеличение с возрастом числа лучевых инициалей тоже связано с делением веретеновидных клеток. Если при ложно-поперечном делении перегородка смещена к одному из концов клетки, то меньшая из образовавшихся клеток становится лучевой инициалью (рис. 21 А). В дальнейшем она может делиться поперечно, образуя многослойный луч, а также продольно, при
этом луч становится многорядным.
69
Рис. 21. Образование лучевых инициалей из веретеновидных: А - отделение от конца веретеновидной клетки; Б - отделение сбоку веретеновидной клетки; В - сегментация веретеновидной клетки; Г - элиминация веретеновидной клетки; 1 - веретеновидные инициали; 2 - лучевые инициали (из Лотова, 2007)
Новые лучевые инициали могут отчленяться от веретеновидной сбоку (рис. 21 Б), у некоторых растений они возникают вследствие сегментации, то есть многократных поперечных делений веретеновидной клетки (рис. 21 В). При этом нередко возникают сразу несколько комплексов лучевых инициалей, начало каждому из которых может дать любая из образовавшихся путем сегментации клеток. Наконец, появление новых лучевых инициалей иногда оказывается следствием элиминации веретеновидных, то есть значительного уменьшения их длины из-за отмирания концевых участков, обусловленного сдавливанием их соседними клетками или какими-либо другими причинами (рис. 21 Г). При этом оставшаяся часть веретеновидной клетки превращается в лучевую инициаль, претерпевающую в дальнейшем, как и во всех отмеченных выше случаях, поперечные и продольные деления. Как правило, из вновь возникших лучевых инициалей сначала формируются однослойные однорядные лучи, затем они становятся многослойными, а у некоторых растений, например, у дуба, бука, черемухи, и широкими, многорядными. Некоторые из образовавшихся лучевых инициалей отмирают.
Все сказанное позволяет рассматривать камбий как структурно сложную меристему. Ее основное отличие от прокамбия состоит в том, что элементы вторичных проводящих тканей образуются не из клеток инициального слоя, то есть собственно клеток камбия, а из их производных; элементы же первичных проводящих тканей дифференцируются непосредственно из клеток прокамбия. Преобладание в камбии делений, происходящих в танген-тальных плоскостях, обусловливает расположение на поперечных срезах всех клеток камбиальной зоны и клеток, приступивших к дифференциации, правильными радиальными рядами.
70
Вторичные проводящие ткани
Древесина хвойных растений
Далее рассматриваются микроскопические особенности строения стеблей, относящиеся преимущественно к древесным растениям умеренной зоны Евразии.
Основные элементы древесины хвойных пород - трахеиды и лучи. Трахеиды осуществляют восходящий ток воды и растворенных в ней солей, а паренхимные клетки лучей выполняют функцию проведения и запасания продуктов метаболизма23. У хвойных встречаются также тяжевая паренхима и смоляные ходы, окруженные обкладкой из паренхимных клеток, а в лучах -лучевые трахеиды, по которым вода перемещается в радиальных направлениях (см. рис. 17). Для правильного описания анатомических особенностей древесины необходимо рассмотрение трех срезов: поперечной, радиальной и тангенциальной ориентации.
На поперечных срезах трахеиды расположены радиальными рядами (рис. 22 А), представляющими собой продолжение таких же рядов камбиальной зоны. Между рядами трахеид проходят однорядные древесинные лучи. В пределах годового прироста хорошо выражены различия между трахеидами ранней (или весенней) и поздней (или летней) древесины. Ранние трахеиды в очертании прямоугольные, реже - многоугольные, они более тонкостенные и широкопросветные, чем поздние; радиальные стенки ранних трахеид имеют окаймленные поры, расположенные чаще в один вертикальный ряд. К одному из отверстий окаймленной поры обычно прижат торус (рис. 22 Б), предотвращающий попадание воздуха из поврежденной трахеиды в соседнюю трахеиду, заполненную водой. Торус, однако, характерен не для всех хвойных. Трахеиды поздней древесины сплюснуты в радиальном направлении. Поры в их стенках немногочисленные, довольно мелкие, иногда они встречаются на тангенциальных стенках.
23
Метаболизм - весь комплекс реакций, протекающих в организме. Близкое понятие - обмен веществ.
71
Рис. 22. Строение древесины сосны на поперечном (А), радиальном (Б) и танген-тальном (В) срезах; Обозначения: а - апертура поры; к.т - концы трахеид; ок - окаймление поры; окц - оконцевые поры в полях перекреста; о.п - окаймленные поры; п.л - паренхимные клетки лучей; п.о - паренхимная обкладка смоляного канала; с.к - смоляной канал; см - смола; т - торус; т.л - лучевые трахеиды; т.п.д - трахеиды поздней древесины; т.р.д - трахеиды ранней древесины; э.к - эпителиальные клетки (из Лотова, 2007)
Переход от ранней древесины к поздней у сосны, например, постепенный, у ели и некоторых других хвойных пород - более резкий. Различие в строении трахеид поздней древесины предыдущего и ранней древесины последующего годовых приростов определяет наличие между ними визуально четких границ. Благодаря этим границам возможен подсчет возраста стебля на уровне поперечного спила.
Ранние и поздние трахеиды различаются не только структурно, но и функционально. Первые участвуют, главным образом, в проведении воды с растворенными в ней минеральными веществами, вторые, помимо функции проведения растворов, играют опорную роль, так как специальных механических элементов в древесине хвойных нет.
Трахеиды составляют продольную (осевую) систему древесины, что хорошо видно на радиальном срезе (рис. 22 Б). Они представляют собой клетки длиной от 2 до 5 и более мм со слегка суженными и обычно тупыми окончаниями. На их радиальных стенках находятся окаймленные поры. У большинства хвойных очертания окаймлений пор округлые или овальные, расположены они одна под другой, составляя чаще один, иногда два продольных ряда, как у сосны, пихты, ели (рис. 22 Б, 22 В). На радиальных срезах поры видны в плане.
Древесинную паренхиму, составляющую, наряду с трахеидами, продольную систему, называют тяжевой, потому что ее клетки собраны в однорядные тяжи; конечные клетки каждого тяжа на продольных срезах имеют клиновидные очертания. Паренхимные клетки часто заполнены буроватым содержимым, их стенки имеют простые поры. Тяжевая паренхима характерна не для всех хвойных. Ее довольно много у кипариса, можжевельника, у которых она располагается более или менее диффузно по годовому слою. Очень мало ее у лиственницы, практически нет у сосны. Отсутствие или слабое развитие тяжевой паренхимы у этих пород сочетается с наличием у них смоляных ходов, вокруг которых имеются обкладки из паренхимных клеток. Смоляные ходы обычно приурочены к поздней древесине годового прироста (рис. 22 А), но встречаются и в ранней. Они представляют собой длинные межклетники, выстланные вытянутыми вдоль эпителиальными клетками, участвующими в образовании смол, растворяющихся в эфирном масле, также синтезируемом этими клетками.
Способность к смолообразованию - очень важная биологическая особенность хвойных. Входящие в состав смолы терпеноидные соединения, сходные по некоторым свойствам с дубильными веществами, усиливают сопротивляемость растений инфекции, повышают их морозоустойчивость. Не случайно поэтому хвойные, произрастающие в северных широтах, накапли
'll
вают больше смолы, чем растения южных стран. При поранении стволов на их поверхности выступает живица. Застывая на воздухе, она способствует «заживлению» ран. Живица служит сырьем для получения канифоли и скипидара.
У отечественных хвойных семейства сосновые встречаются лучи, у которых все клетки лежачие, но разные в функциональном отношении. Внутренние клетки луча запасающие. В их смежных с продольными трахеидами стенках образуются по 2-3 крупные поры с плохо заметным окаймлением, как у корейской сосны, либо по одной простой поре, размеры которой чуть меньше ширины радиальной стенки соседней трахеиды. Такие поры, называемые оконцевыми, характерны для сосны обыкновенной (рис. 22 Б, приложение 2). Сверху и снизу паренхимные клетки обрамлены лучевыми трахеидами, которые могут иметь разные очертания. Внутренняя поверхность их стенок часто неровная из-за наличия зубчатых выростов. Контакты между лучевыми и продольными трахеидами осуществляют мелкие окаймленные поры (рис.22 Б).
Однорядные древесинные лучи хвойных обычно многослойные, но высота их меньше длины продольных трахеид. Среди лучей, свойственных сосновым, наряду с однорядными могут быть и более высокие многорядные веретеновидные лучи со смоляными ходами (рис. 22 В). У пород с лучевыми смоляными ходами всегда имеются и продольные смоляные ходы, как у сосны, ели, лиственницы.
Зоны контакта трахеид с запасающими клетками лучей называются полями перекреста. Число пор и их форма в полях перекреста у представителей разных таксонов хвойных бывают весьма специфичными и служат надежным признаком для установления родовой или даже видовой принадлежности (Приложение 2).
Древесина лиственных древесных растений
Состав элементов, слагающих древесину лиственных древесных пород, более разнообразен, чем у хвойных (рис. 23).
У подавляющего большинства лиственных деревьев трахеиды, хотя и имеются, но основные водопроводящие элементы представлены сосудами (трахеями), а у некоторых пород трахеид вообще нет. Наряду с сосудами и трахеидами продольную систему древесины слагают специализированные механические элементы - древесинные волокна, называемые также волокнами либриформа (присутствуют не всегда), и тяжевая паренхима, более обильная, чем у хвойных. Древесинные лучи состоят только из паренхимных клеток. Сосуды - наиболее широкопросветные элементы древесины - лучше других специализированы для проведения почвенных растворов. Они состоят из сравнительно коротких клеток - члеников сосудов. Для более длинных члеников характерны лестничные перфорации конечных стенок, как у березы, ольхи, лещины (рис. 23 Г). Простые перфорационные пластинки характерны для более коротких члеников, как у дуба, ясеня, белой акации. Окайм
73
ленные поры на боковых стенках члеников сосудов обильные. По расположению они могут быть супротивными, очередными и беспорядочными. Последние два типа поровости боковых стенок члеников сосудов встречаются наиболее часто. У липы, лещины, бересклета и некоторых других пород стенки сосудов, наряду с порами, имеют спиральные утолщения, образованные третичными оболочками. Торуса в окаймленных порах лиственных растений нет.
На поперечных срезах годовых приростов древесины сосуды расположены поодиночке или собраны в группы разных размеров и очертаний. В поперечном сечении они округлые, овальные или угловатые. По особенностям распределения сосудов в годовом приросте различают два типа строения древесины: рассеяннососудистый (береза, ольха, липа) и кольцесосудистый (дуб, ясень, вяз). В первом случае сосуды распределены по годовому приросту более или менее диффузно, хотя в ранней древесине они более обильные и широкопросветные, чем в поздней (рис. 24). Во втором случае в ранней древесине сосуды очень широкие, их просветы можно иногда видеть невооруженным глазом, как у ясеня, дуба. В поздней древесине сосуды немногочисленные, их просветы в несколько раз меньше, чем у сосудов ранней древесины. Между этими типами строения имеются переходные варианты.
Пространство между сосудами занято остальными элементами продольной системы: трахеидами, древесинными волокнами, продольной (осевой, тяжевой) паренхимой.
Трахеиды представляют собой длинные прозенхимные клетки с окаймленными порами, расположение которых варьирует у разных пород. Иногда трахеиды приурочены к сосудам и к поздней древесине годового прироста, в этом случае в древесине присутствует либриформ.
Различают сосудистые и волокнистые трахеиды. Первые отличаются от сосудов меньшим диаметром просветов и отсутствием конечных стенок с перфорационными пластинками; вторые - характеризуются более мелкими щелевидными порами и по строению приближаются к либриформу. Поры чаще располагаются на радиальных стенках трахеид, но могут быть и на тан-гентальных.
У некоторых растений, например у дуба, полости сосудов, реже - трахеид, закупориваются тилами. При этом паренхимная клетка (чаще лучевая паренхима), примыкающая к сосуду, врастает в его полость через пору, растягивая замыкающую пленку, и образует пузыревидный или мешковидный вырост. В ряде случаев он отделяется от паренхимной клетки, делится, увеличивая число тилл. Молодые тиллы живые, с ядром, цитоплазмой, часто они содержат крахмал. Впоследствии у некоторых растений тиллы склерифици-руются, превращаясь в каменистые клетки. Тиллы могут формироваться очень рано. У белой акации, например, уже в первый год жизни сосуды могут затилловываться, однако, у большинства растений тиллообразование связано с появлением ядровой древесины.
74
Рис. 23. Строение древесины березы пушистой на поперечных (А, Б), тангенталь-ном (В) и радиальном (Г) срезах: в.т - волокнистые трахеиды; д.л - древесинные лучи; д.п - диффузная тяжевая (осевая) паренхима; л.и - лучевая паренхима; и.и - перфорационные
75
пластинки с лестничными перфорациями; с - сосуды; т.п - терминальная паренхима в поздней древесине на границе годичного прироста (из Коровин, Курносов, 2004)
Клетки
Рис. 24. Схематические изображения рассеяннососудистой (А) и кольцесосудистой (Б) древесин на поперечных срезах (из Braune, 1967)
Древесинные волокна, или волокна либриформа, как правило, длиннее трахеид. В поперечном сечении они чаще всего многоугольные, но в поздней древесине, в которой их больше, чем в ранней, они нередко сплюснуты в радиальном направлении. Стенки древесинных волокон имеют простые щелевидные поры, в паре пор щели скрещены. Иногда в смежных стенках волокон с другими элементами древесины пор нет.
древесинной паренхимы отличаются от остальных элементов
продольной системы многочисленными простыми порами, а также наличием в их полостях зернистого содержимого и запасного крахмала.
Расположение продольной древесинной паренхимы в годовых приростах имеет важное диагностическое значение. Паренхиму, ассоциированную с сосудами, называют паратрахеальной (от греч./щга - возле). Если она полностью окружает сосуд, говорят о вазицентрической (от греч. vas - сосуд) паренхиме. Различают также крыловидную, сомкнуто-крыловидную и другие варианты паратрахеальной паренхимы, не образующей полной обкладки вокруг сосуда.
Продольную паренхиму, не связанную с сосудами, называют апотрахе-альной (от греч. аро - без). Существует несколько вариантов такой паренхимы: диффузная паренхима, одиночные клетки которой распределены более или менее равномерно по годовому приросту (рис. 23 А); метатрахеальная паренхима, расположенная на поперечных срезах древесины короткими тан-гентальными тяжами; терминальная паренхима, приуроченная к наружным слоям поздней древесины (рис. 23 А, Б). В древесине могут сочетаться разные типы расположения паренхимы.
Древесинные лучи лиственных растений на поперечных срезах либо ровные, строго радиальные, либо изогнутые, огибающие широкие сосуды. Наряду с 1-5-рядными лучами, как у ольхи, березы, ивы, у лиственных древесных пород встречаются и широкие, многорядные (до 30 и более клеток) лучи, характерные для дуба, бука, платана. Лучевая и осевая паренхима образует единую сообщающуюся систему. В заболонной части ствола эти клетки сохраняют живой протопласт, выполняя функцию запасания продуктов метаболизма.
Соотношение между разными элементами, слагающими древесину, а также толщина, пористость и степень одревеснения их оболочек определяют физико-механические свойства этой ткани, в том числе - значения удельной массы, показатели прочности и твердости. Исключительно твердую и красивую древесину имеет парротия персидская (Parrotia persicd), которую назы
76
вают железным деревом, виды рода дальбергия (Dalbergia sp.). Неповторимой по красоте текстуры и весьма прочной является древесина карельской березы и капов ореха грецкого, дуба монгольского, березы пушистой. Очень декоративна древесина видов клена с текстурой "птичий глаз". Твердая, тяжелая древесина с удельной массой 1,42 свойственна бакауту (Guaiacum sp.). Тяжелая древесина с удельной массой 1,17 характерна для эбенового дерева (Diospyros sp.). Древесина бальсы (Ochroma pyramidale) легче пробки (удельная масса - 0,12) и имеет высокую плавучесть. Из ее бревен был сделан плот Кон-Тики, на котором в 1947 г. норвежский ученый и путешественник Тур Хейердал со спутниками пересекли Тихий океан от побережья Перу до островов Полинезии. Еще легче (удельная масса - 0,116) древесина анноны болотной (Annona glabra).
Строение клеточных оболочек древесины
Свойства древесины - основной продукции леса - зависят от ряда показателей, в числе которых качественные и количественные характеристики клеточных оболочек, слагающих древесину, занимают одно из ведущих мест. Строение клеточных оболочек видоспецифично. Например, лучшие бочки для производства вина и коньяка получаются из древесины дуба скального, хорошие - из дуба черешчатого и в разной степени плохие - из североамериканских видов дуба; древесина клена остролистного твердая, мало деформируется при изменении влажности, высоко ценится как сырье для множества изделий, в том числе - музыкальных инструментов; древесина же клена ясенелистного мягкая и не имеет практической значимости; древесина сосны кедровой сибирской отличается от древесины сосны обыкновенной, красивым цветом, возможностью полировки и рядом прочих важных показателей. Все эти различия определяются в первую очередь микроструктурными и химическими особенностями клеточных оболочек, свойственными тем или иным видам древесных растений.
Клеточную оболочку - принадлежность растительной клетки - следует отличать от плазмалеммы, ограничивающей протопласт и присутствующей в клетках всех живых организмов. Плазмалемма растительной клетки координирует синтез и сборку целлюлозных микрофибрилл клеточной оболочки. Клеточная же оболочка служит как бы футляром, окружающим клетку и придающим последней прочность и специфическую форму.
Мы уже говорили о том, что диктиосомы (весь набор диктиосом одной клетки называется аппаратом Гольджи), структурно и функционально связанные с эндоплазматическим ретикулумом, участвуют в секреции полисахаридов, а у большинства высших растений и в образовании клеточных оболочек. Полисахариды клеточной оболочки, синтезируемые диктиосомами, накапливаются в пузырьках, которые затем отделяются от созревающих в диктиосомах цистерн. Эти секреторные пузырьки перемещаются к плазмалемме и сливаются с плазматической мембраной; при этом содержащиеся в них полисахариды встраиваются в формирующуюся на поверхности плазмалеммы
77
клеточную оболочку. Синтез целлюлозы, гемицеллюлозы и пектинов происходит под контролем плазмалеммы.
Сформировавшиеся клеточные оболочки (или стенки) выполняют механическую функцию, т.е. служат опорой растительному организму и сообщают ему необходимую жесткость. С этим связана и другая важная их функция: они ограничивают поступление в клетку воды, от избытка которой протопласт мог бы попросту лопнуть. Вода из-за разности осмотического давления стремится проникнуть в клетку. Клетка, поглощая воду, набухает и давит на клеточную стенку. Именно это гидростатическое давление поддерживает тургор растительных тканей. Увеличение размеров клеток, обусловленное поступлением воды, может продолжаться лишь до тех пор, пока клеточные стенки сохраняют свою растяжимость.
Первичная клеточная стенка (т. е. тот слой клеточной оболочки, который формируется первым) состоит из переплетения микрофибрилл, погруженных в основное вещество, или матрикс. Сами микрофибриллы построены из полисахарида целлюлозы, а матрикс - из других полисахаридов и гликопротеидов. Первичные оболочки контактирующих клеток вместе с полисахаридом пектином, скрепляющим примыкающие друг к другу клетки, образуют так называемую срединную пластинку. Срединные пластинки скрепляют соседние клетки в единое целое. В процессе завершения дифференциации проводящей ткани первичные оболочки вместе с пектиновой межклеточной прослойкой пропитываются лигнином, причем степень лигнификации срединных пластинок значительно выше, чем вторичных оболочек.
Пока продолжается рост клетки, первичная клеточная стенка остается тонкой и эластичной, сохраняя способность сильно растягиваться. Правда, и в период удлинения клетки может происходить некоторое утолщение клеточной стенки, однако обычно оно происходит уже после того, как клетка достигнет своих максимальных размеров. В это время между первичной клеточной оболочкой и плазматической мембраной откладывается вторичная клеточная оболочка и формируются пары пор. Вторичная оболочка по своему строению неоднородна, в ней различаются три слоя. Каждому из трех слоев свойственна своя ориентация микрофибрилл целлюлозы. Микрофибриллы соседних слоев оказываются скрещенными, что существенно повышает прочность клеток древесины (рис. 25).
Рис. 25. Схема строения оболочки клеток вторичной ксилемы: 1 - пектиновая прослойка между оболочками соседних клеток; 2 - так называемая серединная пластинка, объединяющая первичные оболочки прилегающих друг к другу клеток; 3 - вторичная оболочка, состоящая из трех слоев, с разной ориентацией фибрилл целлюлозы (из Яценко-Хмелевский, 1961 с изменениями)
78
У древесных растений клеточные оболочки составляют основную массу древесины. Полости клеток выполняют функцию проведения почвенных растворов и некоторых метаболитов, то есть продуктов жизнедеятельности, от кончиков корней вверх по стволу и ветвям. Эти структурные элементы частично участвуют и в нисходящем проведении некоторых продуктов фотосинтеза и других метаболитов надземной части деревьев. Вторичная оболочка обычно образуется у тех растительных клеток, которые по достижении взрослого состояния теряют протопласт, что и позволяет им выполнять в дальнейшем функции проведения растворов и газообмена.
Основа клеточной оболочки - высокополимерные углеводы. Молекулы целлюлозы, как мы уже отмечали, собраны в сложные пучки (фибриллы), образующие каркас клеточной оболочки, погруженный в ее основу - матрикс, состоящий из гемицеллюлозы и пектинов. В зависимости от типа ткани, в состав которой входит клетка, в клеточной оболочке могут быть и другие органические (лигнин, суберин, воск, белок) и неорганические (соли кальция, кремнезем) вещества. Основной объем материала, составляющего клеточную оболочку элементов древесины, представлен целлюлозой и лигнином. Целлюлоза, как известно, относится к полисахаридам. Сложнее обстоит дело с определением химического состава лигнина. С уверенностью можно сказать, что лигнин относится к ароматическим соединениям и представлен очень крупными полимерными молекулами. Окончательной формулы молекулы лигнина не существует. В целлюлозно-бумажной промышленности при соответствующей обработке древесины от лигнина избавляются как от нежелательного компонента и на предприятиях он накапливается в виде неликвидного отхода производства. В древесинных же клетках лигнин, инкрустируя фибриллы целлюлозы, придает клеточным стенкам прочность и сохранность во времени. Прочность стенок клетки прямо связана со степенью лигнифика-ции. В древесине ствола и ветвей, как мы уже отмечали, сильнее всего лиг-нифицирована срединная пластинка.
У клеток со вторичным утолщением стенок, к каковым относятся клетки вторичной ксилемы древесных растений, в оболочках имеются утонченные участки, называемые порами. Через поры происходит обмен растворенными веществами из клетки в клетку. В понятие пора обычно включают не только канал во вторичной оболочке, но и замыкающий его участок первичной оболочки, который называют замыкающей пленкой поры. Замыкающая пленка пронизана многочисленными плазмодесменными каналами (отверстия, через которые проходили тяжи протопластов (плазмодесмы), соединявшие протопласты соседних клеток).
По особенностям строения выделяют два основных типа пор - простые и окаймленные. У простых пор полость более или менее одинакова по всей глубине поры. Простые поры обычны для паренхимных клеток. Окаймленные поры представляют собой более сложные образования, такие поры характерны для осевых проводящих элементов древесины (сосудов, трахеид). Особенность окаймленных пор заключается в резком сужении полости поры в процессе утолщения вторичной оболочки, так что внешнее отверстие поры
79
гораздо шире, чем внутреннее (рис. 26). При рассмотрении такой поры с поверхности всегда бывают заметны очертания двух отверстий - меньшего, которое соответствует внутреннему отверстию поры, и более крупного, соответствующего внешнему отверстию и как бы окаймляющего очертания меньшего. Это более крупное очертание и называют окаймлением поры. Окаймление нависает над расширенной частью поры. Окаймленные поры трахеид у хвойных имеют так называемый торус - линзообразное утолщение середины замыкающей пленки. Считается, что торус может прижиматься к краю окаймления и таким образом перекрывать пору.
Рис. 26. Окаймленные поры: А -окаймленная пора с включенным округлым отверстием; Б - окаймленная пора с пересекающим щелевидным отверстием: а и б -два взаимно перпендикулярных сечения через пору; 1 - серединная пластинка; 2 -внешний слой вторичной оболочки; 3 - замыкающая пленка поры (серединная пластинка в пределах камеры поры); 4 - торус; 5 - камера поры; 6 - окаймление поры (вид камеры поры в плане); 7 - канал поры; 8 -внутреннее отверстие канала поры; 9 -
внешнее отверстие канала поры; вид на пору сверху (из Яценко-Хмелевский, 1961)
Форма окаймленных пор у различных клеток может быть самой разнообразной. Полость поры может быть большой, значительно превосходящей канал, или маленькой, а очертания ее - округлыми или более или менее вытянутыми. Канал поры может быть цилиндрическим или щелевидным, при этом в одной паре пор направления каналов могут совпадать или находиться под углом друг к другу.
Восходящий ток
Поступление воды и растворенных в ней минеральных веществ осуществляется в растении главным образом по древесине. У древесных растений растворы поднимаются из тонких всасывающих корней на высоту дерева и распределяются по всей кроне, достигая каждого листа. Небольшая доля воды может поступать в растения с поверхности листьев или неодревесневших побегов. Механизмы поглощения воды и особенно подъема ее на высоту дерева весьма сложны и изучены не полностью. Для древесных растений наиболее обстоятельные исследования в этом направлении проведены американскими учеными - П.Д. Крамером и Т.Т. Козловским (см. список литературы), содержание настоящего раздела изложено по их данным.
Транспирация24 - доминирующий процесс в водообмене растений, так как она в значительной степени определяет скорость поглощения и передвижения воды по растению и вызывает водный дефицит. Непрерывное поглощение достаточного количества во
24
Транспирация - испарение воды растением.
80
ды для возмещения ее потери при транспирации имеет существенное значение для роста и даже для выживания растений.
Количество воды, поглощаемой через листья и стебли, невелико, и на рассмотрении этого процесса мы не останавливаемся.
Поглощение через корни. Корневые системы древесных многолетних растений состоят из корней, находящихся на всех стадиях развития, - от тонких, недавно образовавшихся, неопробковевших кончиков диаметром менее 1 мм до старых, одревесневших корней, покрытых толстым слоем коры. Кроме того, корни часто бывают видоизменены из-за наличия микоризных грибов. В результате у разных корней наблюдаются большие различия в проницаемости для воды и солей.
При старении корней эпидерма, корневые волоски и часть первичной коры отмирают вследствие образования перидермы. Раньше предполагали, что вода и растворенные вещества передвигаются от клетки к клетке через вакуоли клеток, расположенных между поверхностью корня и ксилемой. Однако эксперименты показали, что движение воды может в значительной мере происходить по клеточным стенкам. Эта точка зрения находит все большую поддержку. Значительное количество воды у древесных растений поглощается не только через всасывающие участки кончиков корней, но и через старые опробковевшие корни.
Механизмы поглощения воды. Существуют два механизма поглощения воды: активное поглощение, обычно наблюдаемое у слабо транспирирующих растений, и пассивное, которое преобладает у интенсивно транспирирующих растений и обеспечивает большую часть поглощения воды древесными растениями. Необходимо подчеркнуть, что всякое поглощение воды происходит по градиентам водного потенциала - от среды обитания корней до ксилемы. Различие между активным и пассивным поглощением заключается в способе создания этих градиентов.
АКТИВНОЕ ПОГЛОЩЕНИЕ. Корни растений, произрастающих на теплой, хорошо аэрируемой, влажной почве, действуют как осмометры25, когда растения транспири-руют медленно, потому что накопление солей в ксилемном соке повышает его осмотический потенциал, а следовательно, понижает водный потенциал, который становится меньше водного потенциала почвы. Возникающая в результате этого диффузия воды, направленная внутрь, создает корневое давление, благодаря которому происходят гуттация и "плач", наблюдаемый при поранении некоторых растений (например, березы и кленов весной до распускания листьев).
ПАССИВНОЕ ПОГЛОЩЕНИЕ. Когда интенсивность транспирации увеличивается и возникает натяжение в ксилемном соке, начинается массовый приток воды. При этом уносятся соли, накопившиеся в пасоке корней, и уменьшается значение осмотического передвижения воды. Корни становятся пассивной поглощающей поверхностью, через которую вода всасывается благодаря массовому потоку, создаваемому транспирирующимй побегами. Вероятно, что практически все поглощение воды транспирирующими растениями, как древесными, так и травянистыми, происходит пассивно.
Корневое и стеблевое давление. С незапамятных времен люди наблюдали выделение сока из поврежденных растений. На Востоке добывали сок из пальм для приготовления сахара и вина. В Англии и в контитентальной Европе давно проводилась и проводится подсочка березы, а сок использовался для различных целей, в том числе в пивоварении. Интерес к использованию березового сока вновь повысился на Украине. Первые европейцы, побывавшие в Канаде и Новой Англии, заметили, что индейцы делали надрезы на стволах клена и выпаривали сок для получения сахара. В Мексике испанские конкиста
23 Осмомер - прибор, измеряющий осмотическое давление. Осмос - одностороннее проникновение (диффузия) через полупроницаемую мембрану, отделяющую раствор от чистого растворителя или раствора меньшей концентрации. Обусловлен стремлением системы к термодинамическому равновесию и выравниванию концентраций раствора по обе стороны мембраны.
81
доры обнаружили, что туземцы собирали сладкий сок из агавы и сбраживали его для получения спиртного напитка "пульке".
КОРНЕВОЕ ДАВЛЕНИЕ. Такое давление не имеет существенного значения для водопроведения у деревьев умеренной зоны, встречается оно главным образом весной, пока листья не распустились и транспирация невелика. Однако обильный "плач" у березы вишневой наблюдали в Новой Англии и в октябре-ноябре, после листопада.
Потеря сахара при подсочке древесных растений, очевидно, безвредна, так как многие деревья подсачивались десятилетиями без видимых повреждений. Подсчитано, что подсачиваемые клены выделяют менее 10% общего сахара. Эта потеря слишком мала, чтобы иметь значение, если только условия не будут очень неблагоприятными для фотосинтеза в следующее лето. Деревья на бедной или сухой почве обычно дают меньше сока, чем растущие на плодородной, влажной почве.
СМОЛОТЕЧЕНИЕ. Один из наиболее важных примеров выделений, вызываемых поранением, - истечение из сосен живицы, которая служит основой подсочки. Однако живица вытекает из особых смоляных ходов и ее истечение не связано с истечением сока, обсуждавшимся в этой главе.
ФАКТОРЫ, ВЛИЯЮЩИЕ НА ПОГЛОЩЕНИЕ ВОДЫ. Во влажной почве скорость поглощения воды контролируется в основном двумя факторами: интенсивностью транспирации, поскольку она в значительной степени определяет величину водного потенциала в ксилеме корня, и эффективностью деятельности корневых систем как поглощающих поверхностей. При высыхании почвы доступность воды начинает ограничиваться вследствие понижения водного потенциала и гидравлической проводимости. Аэрация и температура почвы, концентрация и состав почвенного раствора также иногда ограничивают поглощение воды.
ПРОТЯЖЕННОСТЬ И ЭФФЕКТИВНОСТЬ РАБОТЫ КОРНЕВЫХ СИСТЕМ. Успешное поглощение любым растением воды и минеральных веществ зависит от протяженности и проницаемости корней У большинства деревьев корневые системы простираются за пределы проекции крон и проникают в почву так глубоко, как это допускают аэрация и физическая структура почвы. В сомкнутых древостоях почва полностью занята корнями на глубину от 25-30 см до нескольких метров.
Эффективность работы корней зависит от размера и проницаемости их поверхности, находящейся в контакте с почвой, и от проницаемости их поверхности. Очевидно, корневые системы с многочисленными мелкими ответвлениями должны быть более эффективными, чем состоящие из небольшого количества крупных, слабо разветвленных корней. Микоризы, по-видимому, повышают эффективность поглощения минеральных веществ благодаря тому, что гифы далеко распространяются в почве, увеличивая таким образом поглощающую поверхность. Они также долго сохраняют активной поглощающую систему более старых корней после их опробковения. Влияние микоризных корней на поглощение воды определить труднее. По сообщению некоторых исследователей, наличие микоризы повышает засухоустойчивость сеянцев древесных растений. Как отмечалось выше, значительное поглощение воды и минеральных веществ происходит через опробковевшие корни, несмотря на их относительно низкую проницаемость.
ПОДЪЕМ КСИЛЕМНОГО СОКА. Существование высоких наземных растений стало возможным только после того, как развилась сосудистая система, обеспечивающая возможность быстрого передвижения воды к транспирирующим побегам. Наземным растениям высотой более 20-30 см трудно существовать без сосудистой системы, за исключением растений, произрастающих в наиболее влажных местообитаниях. Это объясняется тем, что передвижение воды от клетки к клетке путем диффузии происходит слишком медленно для того, чтобы предотвратить обезвоживание надземной массы транспири-рующих растений. На важность этой проблемы для деревьев указывает тот факт, что в жаркий летний день 200 л воды или более может пройти от корней к испаряющей поверхности листьев на высоту 20, 30 или даже 100 м. Исследования показали, что вода, заклю
82
ченная в узких трубках, какими являются элементы ксилемы, обладает очень большой силой сцепления и может подвергаться натяжению.
Недостаточно доказано, что вода может выдержать натяжение, необходимое для того, чтобы притягивать воду к верхушкам высоких деревьев. Теоретически межмолекулярные силы притяжения в воде чрезвычайно велики. Измерения потенциала ксилемного сока, выполненные при помощи барокамеры, показали существование давления до -80 бар, а для преодоления как силы тяжести, так и сопротивления току при передвижении воды к верхушке дерева высотой 100 м достаточно натяжения всего лишь около -20 бар.
ВОДОПРОВОДЯЩАЯ СИСТЕМА. Вода из корней в листья передвигается по специализированной проводящей ткани - ксилеме или древесине, обеспечивающей путь наименьшего сопротивления. Однако, вода должна пересечь ряд слоев живых клеток, чтобы поступить в ксилему корня, а в листьях она опять должна пройти через несколько клеток, прежде чем достигнет испаряющих поверхностей. В ксилеме она передвигается по мертвым элементам. Ксилема, как мы уже установили, состоит из клеток древесной и лучевой паренхимы, волокон, трахеид и сосудов. У хвойных вода передвигается по трахеидам, у покрытосеменных - преимущественно по сосудам, так как они оказывают наименьшее сопротивление току воды. Трахеиды хвойных - это одиночные клетки длиной до 5 мм и диаметром порядка 30 мкм, вода должна проходить через тысячи клеточных стенок, передвигаясь вверх по стеблям. Несмотря на то, что это передвижение облегчается благодаря многочисленным порам в стенках трахеид, сопротивление току воды через трахеиды хвойных и короткие сосуды рассеяннососудистой древесины значительно больше, чем через широкие сосуды кольцесосудистых видов. Сосуды покрытосеменных - это сложные трубчатые структуры, образуемые путем растворения поперечных стенок и протопластов большого количества клеток, расположенных конец к концу. В результате возникают трубки диаметром от 20 до 700-800 мкм и длиной от нескольких метров до величины равной высоте дерева.
СЛОЖНОСТЬ ПРОВОДЯЩЕЙ СИСТЕМЫ Было бы большим упрощением рассматривать ксилему как простое собрание трубок. На самом деле, она нередко образует удивительно сложную систему, в которой пасока проходит неожиданными путями. Например, ксилема организована обычно так, что вода чаще передвигается по спирали, чем вертикально.
Размеры общего поперечного сечения стволов деревьев, участвующего в проведении воды, различны. У многих видов центральная часть ствола превращается в ядро и становится физиологически неактивной. Даже не вся заболонь участвует в проведении восходящего тока, хотя существуют весьма различные мнения по этому вопросу. По крайней мере, у некоторых видов кольцесосудистых деревьев, в том числе у каштана, ясеня и дуба, вода передвигается, по-видимому, по самому наружному годичному кольцу, хотя некоторая часть может проходить по поздней древесине предыдущего года. Характер передвижения растворов у хвойных и рассеяннососудистых древесных растений более сложен вследствие участия в этом процессе большого числа годичных колец. У сеянцев хвойных вплоть до 4-летнего возраста вода передвигается по всем годичным кольцам, причем по самому наружному кольцу менее быстро, чем по второму. Через крупные трахеиды ранней древесины проходит больше воды, чем через более мелкие поздней древесины. У более старых хвойных деревьев существуют, по-видимому, значительные различия в ширине проводящей воду заболони.
ВОДНЫЙ БАЛАНС. Рост древесных и травянистых растений чаще ослабляется вследствие водного дефицита, чем из-за какого-либо другого отдельного фактора. Обширные литературные данные показывают наличие корреляции между ростом растений в высоту и толщину и количеством доступной воды: 70-80 % изменений ширины годичных колец во влажных районах и 90% в сухих местообитаниях можно отнести за счет различий в напряженности водного режима. Степень водного стресса в растениях определяется относительной скоростью поглощения и потери воды, поэтому водный дефицит могут вы
83
зывать или слишком медленное поглощение, или слишком быстрая потеря воды, или чаще всего - сочетание того и другого. Таким образом, изучение факторов, влияющих на поглощение воды и на транспирацию, имеет большое значение, так как способствует пониманию внутреннего водного баланса, который в свою очередь влияет на физиологические процессы и условия, контролирующие быстроту роста.
ОТСТАВАНИЕ ПОГЛОЩЕНИЯ ОТ ПОТЕРИ ВОДЫ. Точно установлено, что содержание воды при солнечной погоде в листьях и стеблях растений часто заметно понижается около полудня. По сообщениям многих исследователей, это проявляется и в уменьшении диаметра ствола.
Содержание воды достигает максимума около восхода солнца, понижается утром и в полдень, повышается после полудня и вечером. Такой режим, характерный, по-видимому, для многих растений при теплой солнечной погоде указывает на то, что стволы деревьев служат местом запасания воды, которая извлекается, когда транспирация превосходит поглощение, и вновь пополняется при противоположной ситуации.
Передвижение воды из древесного ствола отстаёт от её потери листьями. Это отставание указывает на наличие значительного сопротивления току воды через ствол и ветви и по направлению к испаряющим поверхностям листьев. К концу дня, когда температура понижается и устьица закрываются, транспирация быстро уменьшается, но вода продолжает поглощаться до тех пор, пока водный потенциал в растении не повысится приблизительно до величины водного потенциала почвы. На это может потребоваться вся ночь. В случае высыхания почвы, полного восстановления водного потенциала за ночь не происходит, наступает устойчивое увядание. Длительный сильный водный дефицит вызывает отмирание растения вследствие обезвоживания.
Полуденный водный дефицит и временное увядание вызываются тем, что транспирация временно превышает поглощение даже у растений, растущих на почве с достаточной влажностью. Длительное увядание происходит вследствие слабого поглощения воды, вызванного высыханием почвы, а иногда низкой температурой или недостаточной аэрацией, препятствующими возмещению воды, потерянной при транспирации. При усилении водного стресса устьица закрываются, уменьшая расход воды, и деревья с сильно кутини-зированными листьями, могут выдерживать длительные периоды водного стресса с минимальным ущербом.
Луб хвойных растений
Ранее мы установили, что луб, как и древесина, является производным камбия. Обязательные компоненты луба хвойных - ситовидные клетки, осуществляющие транспорт продуктов ассимиляции, а также клетки тяжевой и лучевой паренхимы. Наряду с ними в лубе могут быть волокна и склереиды с толстыми, одревесневающими оболочками, смоловместилища, масляные и слизевые клетки. Разные сочетания этих элементов создают большое разнообразие типов строения луба.
Совокупность всех тонкостенных элементов принято называть мягким лубом, а совокупность толстостенных одревесневших клеток - твердым лубом. Проводящая зона луба у всех древесных пород состоит преимущественно из мягкого луба, в непроводящей может развиваться луб твердый. У сосны и проводящая, и непроводящая зоны сложены только тонкостенными клетками.
Ситовидные клетки составляют большую часть проводящей зоны. На поперечных срезах они более или менее прямоугольные и расположены ра
84
диальными рядами, как и клетки камбиальной зоны, находящиеся с внутренней стороны от них. Ситовидные клетки внешне похожи на трахеиды, но имеют тонкие целлюлозные оболочки.
В проводящей зоне встречаются также клетки тяжевой паренхимы, расположенные диффузно между ситовидными клетками или собранные в короткие либо довольно длинные тангентальные тяжи. Паренхимные клетки обычно заполнены бурым содержимым.
В течение вегетационного периода сначала образуются тонкостенные ситовидные клетки, позднее - клетки с более толстыми стенками. На следующий год под давлением нарастающих изнутри новых слоев вторичных проводящих тканей - производных камбия - прошлогодний луб отодвигается к периферии, составляя внутреннюю часть непроводящей зоны; тонкостенные ситовидные клетки деформируются, сдавливаются, их радиальные стенки становятся извилистыми. Более толстостенные ситовидные клетки противостоят сильной деформации, оставаясь широкопросветными. В многолетнем непроводящем лубе слои этих клеток чередуются со слоями узкопросветных деформированных клеток, соответствующих границам годовых приростов луба. У сосны, ели, лиственницы и других хвойных тяжевая паренхима луба участвует в отложении запасного крахмала и кристаллов оксалата кальция, кубической, ромбоэдрической или призматической форм.
У многих хвойных тяжевая паренхима непроводящей зоны луба подвергается склерификации, при этом оболочки ее клеток сильно утолщаются, становятся многослойными. После одревеснения оболочек и отмирания протопластов клетки превращаются в склереиды, выполняющие механическую функцию.
Лубяные лучи состоят из паренхимных клеток. Клетки имеют крупные ядра, богаты цитоплазматическим содержимым, обычно не содержат крахмала. В непроводящей зоне луба они отмирают вместе с ситовидными клетками. Внешне лубяные лучи похожи на древесинные, обычно они многослойные и однорядные; в веретеновидных лучах встречаются смоляные ходы, формирующиеся уже в камбиальной зоне. В непроводящей зоне в лучах могут развиваться отдельные смолоносные полости.
Луб лиственных древесных растений
Основное отличие луба лиственных деревьев от луба хвойных состоит в строении проводящих элементов. У хвойных они представлены ситовидными клетками, у лиственных - ситовидными трубками, вдоль члеников которых расположены тяжи сопровождающих клеток. Луб включает также тяжевую паренхиму, наиболее обильную в непроводящей зоне. В состав твердого луба могут входить склереиды и волокна. Лубяные, как и древесинные лучи - только паренхимные.
Проводящая зона луба состоит из тонкостенных элементов: ситовидных трубок, тяжевой и лучевой паренхимы. Твердый луб появляется обычно на втором или третьем году жизни ветки.
85
Ситовидные трубки - наиболее широкопросветные элементы, имеющие многоугольные очертания на поперечных срезах. Чаще всего они расположены группами, окруженными клетками тяжевой паренхимы, но нередко составляют довольно длинные тангентальные слои (рис. 27).
Членики ситовидных трубок имеют либо простые, либо сложные ситовидные пластинки подобно перфорационным пластинкам между члениками сосудов в древесине. На поперечных срезах простые ситовидные пластинки обычно видны в плане, сложные - перерезанными, так как они всегда расположены наклонно к продольной оси членика ситовидной трубки (рис. 27 Б). В плане они видны только на радиальных срезах, как и перфорационные пластинки в сосудах древесины. В проводящей зоне луба ситовидные трубки за полнены зернистым содержимым, в непроводящей они пустые. Сопровождающие клетки, прижатые к одной из стенок членика ситовидной трубки, -самые мелкие элементы луба, характеризующиеся плотным содержимым (рис. 27 Б, В, Г). Обычно каждый членик ситовидной трубки ассоциирован с одним тяжом сопровождающих клеток, но иногда их может быть два и более. Число клеток в тяже сильно варьирует. Как и у хвойных, ситовидные элементы функционируют в течение одного вегетационного периода. Исключение составляет липа, у которой живые ситовидные трубки отмечены в 2-5-летних ветвях. Осенью ситовидные пластинки закупориваются мозолистыми телами.
Тяжевая паренхима у лиственных древесных растений двух типов: запасающая, содержащая зерна крахмала, и кристаллоносная, в клетках которой откладывается оксалат кальция в виде одиночных кристаллов (липа, береза, клен, черемуха, дуб), друз (ольха, лещина), кристаллического песка или очень мелких игольчатых кристаллов (ясень). У вяза некоторые клетки тяжевой паренхимы сильно разрастаются, превращаясь в слизевые клетки, особенно много их у вяза приземистого, или мелколистного. К тонкостенным элементам луба принадлежат также млечники, встречающиеся в первичном лубе клена, вторичном лубе бересклета, у которого они содержат гуттаперчу.
Лишь у немногих древесных пород в лубе нет одревесневших элементов. У большинства лиственных деревьев твердый луб в той или иной степени развит. По характеру слагающих его элементов лиственные древесные породы составляют три группы.
У пород первой группы твердый луб представлен только склереидами (рис. 28), возникающими обычно вследствие склерификации клеток тяжевой паренхимы (береза, ольха). Склереиды имеют толстые, твердые оболочки, пронизанные поровыми канальцами; расположены склереиды поодиночке или группами разных размеров и очертаний. У пород второй группы твердый луб состоит только из волокон длиной 0,5-2,5 мм, дифференцирующихся непосредственно из производных веретеновидных клеток камбия. В поперечном сечении они мельче склереид, имеют одно-, двухслойные оболочки с немногочисленными порами. Волокна обычно располагаются группами, составляющими плотные или рыхлые тангентальные тяжи.
Породы третьей группы характеризуются наличием в твердом лубе и волокон, и склереид (ясень, дуб). Чаще всего склереиды развиваются между
86
группами волокон, скрепляя их, но они могут располагаться и диффузно. У клена склерифицируются клетки не только тяжевой, но и лучевой паренхимы.
У большинства лиственных деревьев группы элементов твердого луба ассоциированы с клетками кристаллоносной паренхимы. Дилатация, приводящая к тангентальному разрастанию наружных слоев луба, у лиственных пород может быть, как и у хвойных, диффузной, в которой участвует тяжевая паренхима, и лучевой. Диффузная дилатация свойственна березе, ольхе, лучевая — яблоне, липе. У многих древесных пород, как уже было отмечено, лучевая дилатация сочетается с диффузной.
В дилатационной зоне у деревьев с чешуйчатой коркой периодически закладываются слои феллогена или пробкового камбия. Феллоген образует пробку, а отделенные феллогеном наружные слои луба становятся мертвым лубом, входящим в состав корки.
У хвойных рано формируется корка. Нередко уже на 4-5 году жизни ветки развиваются внутренние перидермы. Пробка, составляющая наружную часть перидермы, у большинства хвойных древесных пород имеет однородное строение, но у сосны, ели, лиственницы в ней можно различить три типа клеток: обычные, со слегка утолщенными стенками, губчатые, с очень тонкими, извилистыми стенками, и толстостенные феллоиды, имеющие одревесневшие стенки. Механически они слабо связаны с тонкостенными клетками, между ними легко возникают трещины, способствующие сбрасыванию корки. У большинства хвойных корка чешуйчатая, у кипариса, некоторых видов можжевельника - кольцевая, у пихты корка не образуется.
Корка у большинства лиственных пород чешуйчатая, но у некоторых видов и гладкокорых форм корка совсем или долго не образуется. У таких растений перидерма остается кольцевой. У растений с чешуйчатой коркой клетки пробки, входящие в состав перидермы, нередко дифференцированы на тонко- и толстостенные, как, например, у платана. Опадению чешуи такой пробки способствуют напряжения, вызываемые различиями в гигроскопичности и прочности разных слоев пробки. У растений с кольцевой коркой ее опадение сопровождается разрывами на длинные лентовидные куски, которые на некоторое время остаются соединенными с более молодой коркой своими концами; впоследствии они сбрасываются.
Практическое значение имеет корка пробкового дуба, произрастающего в юго-западной Европе и Северной Америке. Первый феллоген у него функционирует очень долго, образуя мощную, со временем грубеющую и растрескивающуюся пробку. Ее полностью срезают. Следующий закладывающийся феллоген порождает мягкую упругую пробку, используемую в качестве закупорочного материала. Эту пробку снимают несколько раз обычно через каждые 8-10 лет до достижения деревом 200-летнего возраста.
87
Рис. 27. Строение камбия и вторичной флоэмы (мягкого луба) березы пушистой: А - камбиальная зона; Б - поперечный срез; В - тангентальный срез; Г - радиальный срез; б.с - боковые ситовидные поля; в.и - веретеновидные инициали; д.л.и - делящиеся веретеновидные инициали; к.з - камбиальная зона; л - лучи; с.к - сопровождающие клетки; с.и - ситовидные пластинки; т.п - тяжевая паренхима; с.т - ситовидные трубки (из Коровин, 1970)
88
Рис. 28. Луб и перидерма березы: А - луб и кольцевая перидерма гладкокорых форм; Б - луб и чешуйчатая корка грубокорых форм; д.з - дилатационная зона; м.л -мертвый луб; и.л - непроводящий луб; пк - пробка (годичные слои прироста пробки); и.л - проводящий луб; ск - склереиды (группы каменистых клеток) (из Коровин, 1970)
У некоторых лиственных древесных пород корка образуется только в основании старых стволов, как у березы, ствол которой на большом протяжении покрыт пробкой (берестой), состоящей из чередующихся слоев узко- и широкопросветных клеток, благодаря чему хорошо видны годовые приросты пробки (бересты) (рис. 28 А).
Нисходящий ток (транспорт ассимилятов)26
Транспорт ассимилятов27 осуществляется по флоэме и включает в себя три этапа: загрузку флоэмы в окончаниях тонких жилок листа, передвижение веществ по проводящим элементам флоэмы и разгрузку флоэмы в зонах потребления ассимилятов.
Проводящими элементами флоэмы двудольных служат членики ситовидных трубок, которые, соединяясь конец в конец, образуют ситовидные трубки. Они связаны с особыми паренхимными клетками, называемыми клетками-спутниками, или сопровождающими клетками. Ситовидные трубки с сопровождающими клетками и вертикальные тяжи флоэм
26 Раздел написан по публикации Л.Л. Новицкой (2008) с некоторыми изменениями.
27 Ассимиляты - продукты фотосинтеза.
89
ной паренхимы вместе составляют осевую (вертикальную) систему клеточных элементов, образуемую веретеновидными инициалами камбия. Она пересекается лучевой (поперечной или радиальной) системой, возникающей в результате активности лучевых инициалей. Главными компонентами лучевой системы являются паренхимные клетки лучей. Располагающиеся друг напротив друга флоэмный и ксилемный лучи возникают из общей группы лучевых камбиальных инициалей и в совокупности образуют радиальный луч. Лучевая и осевая паренхима флоэмы и ксилемы в целом представляют собой единую разветвленную паренхимную сеть, отдельные клетки которой объединены между собой посредством многочисленных поро-плазмодесменных28 связей.
Отток ассимилятов (сахаров) из листьев происходит по проводящим элементам -ситовидным трубкам флоэмы. Загрузка сахаров из клеток мезофилла листа в русло дальнего транспорта ассимилятов по флоэме осуществляется в терминальных участках-мелких жилок листа. По структурно-функциональным особенностям окончаний мелких жилок листа растения делятся на «симпластные», или «эндопластные» (с открытой флоэмой) и «апопластные» (с закрытой флоэмой). У симпластных растений ассимиляты из клеток мезофилла доходят до ситовидных элементов флоэмы по непрерывной системе вакуолей и каналов эндоплазматической сети - эндопласту. Эндоплазматические сети отдельных клеток объединяются в единую систему межклеточного транспорта посредством плазмодесм - пор в клеточной оболочке, содержащих трубочку эндоплазматической сети. У апопластных растений плазмодесменные связи между клетками мезофилла и ситовидными трубками флоэмы отсутствуют, поэтому эндопласт листа состоит из двух самостоятельных доменов (частей, отдельных участков), разделенных между собой зонами клеточных оболочек без плазмодесм. Для перехода из одного эндопластного домена в другой ассимиляты должны преодолеть наружные клеточные мембраны пограничных клеток и пространство разделяющих их клеточных оболочек - апопласт.
Транспортными формами ассимилятов (сахаров) являются сахароза и олигосахариды29. Олигосахариды синтезируются на основе сахарозы. По плазмодесмам могут перемещаться как сахароза, так и крупные молекулы олигосахаров. Преодолеть плазматическую мембрану может только сахароза. Поэтому у симпластных растений по флоэме, т. е. в русле дальнего транспорта, транспортируются сахароза и олигосахара, а у апопластных - только сахароза.
Между двумя крайними группами видов с только симпластной или только апопла-стной загрузкой флоэмы существует много промежуточных (комбинированных) вариантов загрузки. Прежде всего сюда относятся листопадные деревья и кустарники с выраженной сезонной ритмикой роста. Для них характерна полуоткрытая флоэма (с небольшим числом плазмодесменных связей между мезофиллом и флоэмой), комбинированный аппарат загрузки и, соответственно, переменный состав флоэмного экссудата30.
Передвижение ассимилятов из кроны к зонам их потребления в стволе происходит внутри ситовидных трубок в продольном направлении. Из трубки в трубку они попадают через систему ситовидных пор на поперечных ситовидных перегородках, отделяющих один членик ситовидной трубки от другого. Обеспечение продуктами фотосинтеза активно делящихся клеток камбиальной зоны, а также всех остальных клеток и тканей коры и древесины, осуществляется за счет латерального (бокового) обмена между ситовидными элементами и другими клетками флоэмы. Такой обмен возможен благодаря наличию на
28 Плазмодесмы - каналы в клеточных оболочках, соединяющие протопласты соседних клеток. Состоят из плазмалеммы, выстилающей канал, слоя цитоплазмы и проходящей сквозь нее трубки эндоплазматического ретикулума. Трубка связывает эндоплазматические сети (ретикулумы) соседних клеток. Плазмодесмы объединяют протопласты растительных клеток в единую систему.
29 Олигосахариды - углеводы, состоящие из 2-10 моносахаридных остатков, связанных гликозидными связями. Олигосахариды с 2-5 моносахаридными остатками являются транспортными формами сахаров у растений.
30 Экссудат - раствор, транспортируемый по сосудам ксилемы (ксилемный экссудат) и ситовидным трубкам флоэмы (флоэмный экссудат).
90
боковых стенках ситовидных трубок большого количества ситовидных полей, представляющих собой совокупности ситовидных пор.
Ситовидные элементы содержат пластиды, аккумулирующие крахмал. Они имеют довольно примитивную структуру: их мембранная система ограничивается двойной оболочкой и несколькими внутренними ламеллами. Число и величина крахмальных зерен в пластидах непостоянны, наиболее распространенная форма - большие скопления мелких зерен.
Запас крахмала в пластидах ситовидных элементов легко подвижен. В частности, при повреждениях флоэмы, вызывающих быстрое образование каллозы, крахмал исчезает из пластид, расходуясь, очевидно, на ее синтез. Кроме того, путем отложения и мобилизации крахмала амилопласты ситовидных трубок могут поддерживать на постоянном уровне концентрацию сахарозы в самих ситовидных элементах. Однако последняя функция является недостаточной и дополняется деятельностью паренхимных клеток проводящей флоэмы, содержащих иногда значительные количества резервной сахарозы. Латеральный выход сахарозы из ситовидных трубок во время далекого транспорта представляет собой результат активной деятельности клеток флоэмы, что составляет один из регуляторных механизмов продольного транспорта в ситовидных трубках. Абсорбируемые клетками паренхимы сахара прежде всего превращаются в них в крахмал, а также размещаются в вакуолях под защитой тонопласта. Таким образом, наличие в проводящей флоэме некоторого количества сахарозы, выведенной из непосредственного участия в транспорте, можно считать обычным явлением. Этот запас находится в состоянии обменного равновесия с сахарозой, движущейся по ситовидным трубкам. Роль запасных резервуаров при этом отводится вакуолям и пластидам клеток флоэмной паренхимы.
Существует биологический предел накопления сахаров, т. е. максимальный уровень концентрации сахарозы в запасающем пространстве клеток.
Размер радиального выхода сахарозы за пределы проводящей флоэмы в большой степени определяется физиологическим состоянием потребляющих тканей. Иногда в клетках флоэмной паренхимы древесных растений может происходить значительная аккумуляция углеводов. В наиболее ярком виде это проявляется при осеннем оттоке сахаров из кроны в ствол, а затем в корни древесного растения. Функцию накопления запасных веществ при этом выполняют паренхимные клетки флоэмы и древесины.
СТРОЕНИЕ ЛИСТА
Строение листа двудольного растения
Лист развивается из зачатка, образующегося на конусе нарастания побега. Верхушечный рост развивающегося листа очень скоро прекращается, и дальнейшее его формирование осуществляется за счет деятельности вставочной меристемы, расположенной в основании листовой пластинки.
Этот орган недолговечен, образован первичными тканями и в отличие от корня и стебля имеет двухстороннее или дорзовентральное (спиннобрюшное) строение, причем верхняя и нижняя стороны в типичных случаях отличаются своими анатомическими особенностями. Рассмотрим типичное строение горизонтально расположенного двухстороннего листа двудольного растения (рис. 29).
91
Рис. 29. Поперечный разрез листовой пластинки георгина через главную жилку (схематическое изображение): 1 - эпидерма; 2 - колленхима; 3 - проводящие пучки; 4 -столбчатый мезофилл; 5 - губчатый мезофилл (на самом деле в зоне губчатого мезофилла межклетников больше и они крупнее); 6 - устьице: 7 - паренхимная обкладка вокруг проводящих пучков; 8 - хлоропласты в клетках мезофилла (их относительные размеры увеличены) (из Иванов, 1929)
Сверху и снизу лист покрыт эпидермисом. Верхний эпидермис имеет, как правило, небольшое количество устьиц и часто покрыт блестящей бесструктурной пленкой - кутикулой. Нижний эпидермис не имеет кутикулы, в нем образуется много устьиц, на нем часто развивается восковой налет или опушение. Между верхним и нижним эпидермисом заключена мякоть листа - мезофилл. Мезофилл состоит из ассимиляционной паренхимы и обычно подразделяется на два типа тканей столбчатую, или палисадную паренхиму, примыкающую к верхнему эпидермису, и губчатую, примыкающую к нижнему эпидермису (рис. 30). Основная функция мезофилла - фотосинтез. Кроме того, в губчатой ткани осуществляются газообмен и транспирация, а также отток ассимилятов в проводящие пучки - "жилки" листа. Пучки листа обычно закрытые. Флоэма обращена в сторону губчатой ткани, т. е. морфологически вниз. В зоне мезофилла часто присутствует колленхима (под эпидермисом) и различной формы склереиды.
92
Рис. 30. Поперечный разрез листа груши: 1 - палисадная паренхима (столбчатый мезофилл); 2 - обкладка пучка; 3 - верхний эпидермис; 4 - продолжение обкладки пучка; 5 - волокна; 6 - ксилема; 7 - флоэма; 8 - губчатая паренхима (губчатый мезофилл); нижний эпидермис. Два проводящих пучка окружены обкладкой, но лишь у большего из них обкладка доходит до верхнего и нижнего эпидермиса. В клетках мезофилла содержатся хлоропласты, за исключением тех из них, где имеются кристаллы. В клетках обкладки хлоропластов мало (на рисунке они не изображены), (из Эсау, 1969)
Листья злаков не имеют дифференциации мезофилла на губчатую и столбчатую ткань, он состоит из однородной ассимиляционной ткани - хлоренхимы, напоминающей губчатый мезофилл двудольных.
Строение хвои
Хвоя (лист хвойных) отличается своеобразным анатомическим строением (рис. 31, 32). Эпидермис ее состоит из толстостенных клеток с небольшим объемом протопласта и обычно покрыт мощным слоем кутикулы. Устьица расположены на дне глубоких бороздок, заполненных зернышками воска.
Под эпидермисом находится слой гиподермы, имеющей клетки с утолщенными и одревесневшими стенками. Мезофилл хвои состоит из ассимиляционной ткани, получившей название складчатой паренхимы. Она представлена клетками, оболочки которых образуют складки, заходящие в полость клетки. Цитоплазма с содержащимися в ней хлоропластами образует обычно в клетках хвои пристенный слой, располагаясь вдоль клеточной обо
лочки.
93
Рис. 31. Строение хвои (листа) сосны черной форма австрийская (Pinus nigra var. austriaca) на поперечном срезе: 1 - смоляные каналы; 2 -складчатый мезофилл; 3 - проводящий пучек (один из двух сближенных):4 - эндодерма; 5 - эпидерма; 6 - склерифици-рованная гиподерма; 7 - устьице; 8 - трансфузионная паренхима (из Molisch, Hofler, 1965)
Такое строение клеток складчатой паренхимы в значительной степени увеличивает ассимиляционную поверхность мезофилла. В центре хвои расположены 1 или 2 проводящих пучка, окруженных трансфузионной паренхимой. Клетки трансфузионной паренхимы проводят воду и органические вещества и осуществляют связь между проводящими пучками и мезофиллом. К пучкам в некоторых местах примыкают толстостенные клетки склеренхимы. Центральная часть хвои отделена от мезофилла кольцом клеток эндодермы, также имеющих утолщенные и одревесневшие оболочки. В мезофилле проходят смоляные ходы, окруженные склеренхимой и выстланные клетками эпителия, выделяющими смолу в полость хода.
Рис. 32. Поперечный разрез хвои пихты белой (Abies alba). 1 - ксилема проводящего пучка; 2 - эпидерма; 3 - мезофилл (хлоренхима); 4 - смоляной канал; 5 - устьица; 6 -
94
флоэма проводящего пучка; 7 - нижняя эпидерма; 8 - эндодерма; 9 - смоляной канал; 10 -хлоренхима; И - склерифицированная гиподерма (из Molisch, Hofler, 1965)
СТРОЕНИЕ КОРНЯ
Первичное строение корня
По анатомическим признакам и функциональным особенностям корень можно разделить на четыре зоны: 1) зона делящихся клеток; 2) зона растяжения; 3) зона специализации и всасывания; 4) зона проведения. Первая зона -апикальная меристема корня. В отличие от конуса нарастания стебля верхушечная меристема корня образует новые клетки в двух противоположных направлениях - кнаружи от кончика корня и внутрь него. Из наружных клеток формируется корневой чехлик, защищающий нежные клетки образовательной ткани от повреждений при внедрении в почву. Из внутренних производных апикальной меристемы формируются все ткани первичного тела корня.
В зоне роста деление клеток первичной меристемы прекращается и начинается их дифференциация, они вытягиваются по длине корня и в них появляются вакуоли. Здесь первичное тело корня удлиняется. Протяженность зоны роста составляет несколько миллиметров.
В зоне специализации, или всасывания, клетки первичной меристемы окончательно дифференцируются в различные ткани - покровную, проводящую, основную и другие, характерные для первичного строения корня. Эпидермис здесь образует корневые волоски, всасывающие из почвы воду с минеральными веществами. Зона специализации обычно имеет длину в несколько сантиметров. Корневые волоски функционируют недолго (10-20 дней) и вскоре отмирают. Вместо них формируются новые корневые волоски на молодом участке корня, выросшем за это время из первичной меристемы конуса нарастания.
Зона проведения занимает всю остальную часть корня - от зоны специализации до корневой шейки - и имеет наибольшую протяженность. Величина этой зоны и характеризует длину корней того или иного растения.
Вторичное строение корня
Первичное строение корня у однодольных растений свойственно не только зоне всасывания, но и зоне проведения, вследствие чего корни их не способны ко вторичному утолщению. Для двудольных растений в зоне проведения характерно вторичное строение корня, обеспечивающее его рост в толщину (рис. 33).
95
Рис. 33. Образование вторичных тканей в корне: 1 - распределение зон роста; 2-6 -поперечные срезы корня на разных уровнях; ВКс - вторичная ксилема; ВФ - вторичная ксилема; К - камбий; Мкс - метаксилема; Мф - метафлоэма; Пд - протодерма; Пкс - протоксилема; Прд - перидерма; Пф - протофлоэма; Пц - перицикл; Рд - ризодерма; Экз -экзодерма; Эн - эндодерма; Ч - чехлик (из Васильев и др., 1978)
Переход к формированию вторичного тела начинается с образования вторичной образовательной ткани - камбия. Камбий располагается таким образом, что первичная ксилема оказывается от него к центру, а первичная флоэма - к поверхности корня. Первичная флоэма под давлением вторичных элементов сдавливается и постепенно рассасывается. Первичная ксилема сохраняется в центре корня, непосредственно соединяясь со вторичной ксилемой. При переходе ко вторичному строению первичная кора корня отмирает и сбрасывается после активизации пробкового камбия - феллогена.
Комплексная проводящая ткань вторичного тела корня (ксилема и флоэма), помимо указанных выше отличий, практически идентична вторичным
96
ксилеме и флоэме стебля. Небольшие различия относятся в основном к количественным показателям.
МОРФОЛОГИЯ РАСТЕНИЙ
В данном разделе приведен обзор только основных терминов и понятий для отделов Голосеменные и Покрытосеменные растения, к которым относятся отечественные виды деревьев и кустарников. Из множества терминов, используемых в морфологических описаниях, здесь приведены лишь главнейшие, относящиеся к следующим частям растения: стебель и почка, лист, корень, соцветие, цветок, плод. Перечисленные части растения и органы характеризуются множеством частных особенностей или проявлений. Следуя в основном тексту Ю.Е. Алексеева с соавторами (см. список литературы) остановимся на основных особенностях морфологии древесных растений (рис. 34).
Рис. 34. Схема строения двудольного растения (побег, семядоли и корень): 1 - верхушечная почка; 2, 9 - междоузлия; 3 - пазушная почка; 4 - придаточные корни; 5, 6 - боковые корни; 7 - главный корень; 8 - узел; 9 - междоузлие; 10 -семядоли (из Strasburger, Noll, Schenck, Schimper, 1967)
После того как семя прорастает, апикальная меристема - верхушечная образовательная ткань побега - начинает формировать в правильной последовательности листья, узлы и междоузлия. Из апикальных меристем, находящихся в пазухах листьев, образуются пазушные побеги, которые в свою очередь дают начало дополнительным пазушным побегам.
В результате такой активности на главном стебле растения развивается система ветвей. Если пазушные меристемы остаются неактивными, то на побеге не образуются ветви, что наблюдается, на-
пример, у многих пальм. Из апикальной меристемы корня развивается главный, первичный, корень. У многих растений из новых апикальных меристем, закладывающихся глубоко в главном корне (эндогенное происхождение), возникают боковые, вторичные, корни, которые в свою очередь дают начало боковым корням следующего порядка. В конечном счете у растения развивается сильно разветвленная корневая система. У некоторых растений, пре
имущественно у однодольных, корневая система взрослого растения состоит из придаточных, адвентивных, корней, возникающих на стебле. Такие корни могут напоминать щетку или кисть (мочковатая корневая система), так как они сходны между собой по длине и форме.
Описанный выше рост представляет собой вегетативную стадию жизни семенного растения. В соответствующее время, определяемое отчасти эндо
97
генным ритмом роста, а отчасти внешними условиями, особенно светом и температурой, вегетативная апикальная меристема определенной части побегов превращается в репродуктивную апикальную меристему, называемую у покрытосеменных флоральной апикальной меристемой, из которой развивается цветок или соцветие. Таким образом, за вегетативной стадией в жизненном цикле растения следует репродуктивная.
Стадия развития, которая заканчивается тем, что все более или менее прямые производные апикальных меристем переходят в зрелое состояние, называется стадией первичного роста. Все тело растения, включая корни, стебли, листья, цветки, плоды и семена, а также системы покровной (эпидерма), основной и проводящей тканей, может образовываться в результате первичного роста. У некоторых двудольных (мелкие однолетники) и большинства однодольных первичный рост наблюдается на протяжении всего жизненного цикла растения. Однако у большей части двудольных и голосеменных обнаруживается и вторичный рост, обусловленный активностью камбия. В результате деятельности этой меристемы увеличивается количество проводящей ткани, что вызывает утолщение оси растения. Образование защитной ткани - перидермы, замещающей эпидерму, также происходит путем вторичного роста. Вторичное отложение проводящих тканей и защитного покрова обеспечивает развитие крупных, сильно разветвленных растений, представленных древесными формами.
Семенное растение и в зрелом состоянии представляет собой непрерывно изменяющийся организм. Благодаря активности апикальных меристем побегов и корней оно сохраняет способность добавлять новые части к своему телу, а благодаря активности латеральных меристем (камбий, феллоген) -увеличивать объем вторичных тканей. Древесные растения продолжают расти практически до самой смерти.
Побег, стебель
Побег является одним из основных органов растений и состоит из стебля, листьев и почек. Такое определение правильно характеризует однолетний побег. В ботанической литературе и разветвленный ствол взрослого дерева часто называют побегом, хотя на нем давно уже нет ни листьев, ни почек. Видимо, правильнее всю надземную вегетативную часть дерева называть стеблем, а побегами - только однолетние приросты стебля.
Первый побег растения появляется из зародышевой почки семени. У многолетних растений годичные побеги формируются из почек, которые являются зачатками побегов и расположены на прошлогодних побегах, а точнее на их узлах. В некоторых случаях побеги формируются из придаточных почек, которые закладываются на корнях, листьях или на междоузлиях побегов. В последних вариантах побеги называются придаточными. К ним относятся и так называемые корневые отпрыски, свойственные некоторым видам деревьев и кустарников (осина, виды ольхи и др.).
98
Побеги состоят из морфологически однородных составляющих - метамеров. Каждый метамер представлен узлом и междоузлием. Узел это место прикрепления листа и пазушной почки к стеблю, а междоузлие - участок стебля между двумя узлами. В стволах сформировавшихся деревьев и в многолетних ветвях следы метамеров исчезают под приростами древесины, луба и корки, однако у некоторых растений они долго остаются хорошо различимыми, например, на стволе сосны обыкновенной до 40-50 лет.
Групповое заложение почек возобновления (придаточных спящих почек) у многих видов древесных растений приводит к образованию своеобразных наростов на стволах, чаще в комлевой части, и (реже) на ветвях. Такие наросты называются капами (рис. 35).
Рис. 35. Схематическое изображение прикорневого капа (рис. В. Коровина)
Капы отличаются от других наростов на стволах тем, что на их поверхности находятся многочисленные почки, апексы которых являются источниками ростовых веществ - ауксинов. Ауксины стимулируют активность камбия. В зоне скопления придаточных почек резко увеличивается радиальный прирост стебля. Эти же росто-
вые вещества вызывают образование новых придаточных почек. Таким образом, скопле-
ние почек запускает своеобразную цепную реакцию - скопление почек вызывает образо-
вание новых почек, а местное увеличение концентрации ростовых веществ стимулирует камбий и приводит к росту наплыва древесины. Некоторые из спящих почек на поверхности капов "просыпаются" и дают укороченные или удлиненные побеги. Такие побеги часто укореняются и образуют дочерние растения.
Капы, особенно прикорневые - формирующиеся в основании ствола - иногда достигают больших размеров (в литературе есть описание капа березы, имеющего диаметр 4 метра). Древесина капов из-за своеобразной свилеватости и густой сети следов спящих почек очень прочна и декоративна, ее используют для изготовления подарочных изделий и дорогой мебели; из-за отдаленного сходства с древесиной карельской березы ее часто путают с последней. Особенно красива древесина капов грецкого ореха.
Целый ряд кустарников формирует новые многолетние стебли из почек возобновления, которые размещаются в основании более старых стволиков ниже уровня почвы или на уровне почвы. Одно из названий таких многолетних стволиков - скелетные стебли, тогда как ежегодно образующиеся новые побеги (годовые приросты) называют годичными побегами. Скелетные стебли по своему положению похожи на порослевые побеги, которые образуются у некоторых деревьев от пня после вырубки или из прикорневых капов. Мно
гие виды деревьев и кустарников сохраняют неизменным положение и направление роста своих стволов в течение всей их жизни. Однако существует своеобразная группа кустарников, их называют стланики, у которых направление роста каждого скелетного стебля изменяется в течение всей его жизни. Не касаясь здесь деталей этого процесса, отметим лишь главное: скелетный стебель у стланика первые годы своей жизни растет наклонно, восходящим образом; несколько позже корни "прижимают" основание побега к почве,
99
подстилке, тогда как верхняя часть стволика продолжает сохранять вертикальное или наклонное положение.
Годичные побеги древесных растений в отличие от побегов трав характеризуются "одревеснением" целого ряда внутренних тканей и, кроме того, появлением, начиная со второго года жизни, особой покровной ткани, приводящим к образованию так называемой корки (в просторечии чаще называемой корой). Каждый годичный побег со временем становится частью многолетней ветви или ствола растения. Форма кроны или ее архитектоника определяются комплексом морфологических особенностей ветвей и побегов, специфичных для каждого вида.
В некоторых случаях наблюдаются своеобразные нарушения в организации кроны, выражающиеся в образовании так называемых "ведьминых метел". Такие аномальные части кроны могут возникать практически у всех древесных растений. По внешним очертаниям "метлы" представляют собой близкие к шаровидным скопления часто ветвящихся коротких равноценных по быстроте роста и характеру ветвления побегов. Подобные аномальные образования часто можно наблюдать в кронах старых сосен, издали они бывают похожими на вороньи гнезда. Изредка эти образования достигают больших размеров (рис. 36) и даже могут заменять собой всю крону.
Независимо от свойственных данному виду морфологических особенностей (характер ветвления, форма кроны) «ведьмины метлы» всегда представляют собой шаровидные образования (крона «ведьминой метлы» с той или иной степенью приближения вписывается в шар, так как ни один из побегов по отношению к другим не является лидирующим). В данном случае имеются в виду только мутантные «метлы», так как во многом сходные с ними «ведьмины метлы» патологического происхождения могут носить черты некоторой специфичности, связанной с особенностями того или иного патогенна.
Дерево с мутантной «ведьминой метлой» или с несколькими такими «метлами» представляет собой химеру - одна корневая система несет генетически различные стебли. Причем эти стебли (в данном случае можно сказать - самостоятельные кроны) находятся в конкурентных отношениях. Мелкие «метлы» на боковых ветвях обычно недолговечны и через 2-4 года после возникновения засыхают, крупные же, образующиеся на вершине дерева или близко к ней, живут довольно долго и в некоторых случаях подавляют основную крону.
Мутантные «метлы» способны образовывать женские стробилы (образование мужских стробилов не наблюдалось) и давать фертильные семена. Учитывая мужскую стерильность мутантных «метел», опыление у них всегда перекрестное.
В семенном потомстве «ведьминых метел» белой (веймутовой) сосны (Pinus stro-bus L.) наблюдалось расщепление нормальных и мутантных особей 1:1, то есть половина растений были корнесобственными «ведьмиными метлами». Корнесобственные «метлы» сосны обыкновенной были найдены в Приобских ленточных борах Алтая. Разумеется, такие находки - редкость, так как эти карликовые растения в естественных условиях совершенно не конкурентоспособны. По утверждению многих ученых корнесобственные "ведьмины метлы" явились исходным материалом для выведения карликовых (шаровидных) декоративных форм древесных растений.
100
Рис. 36. "Ведьмина метла" на сосне обыкновенной. Алтай, Приобские боры (фото В.В. Коровина)
По морфологическим признакам и функциональным особенностям побеги и ветви древесных растений делятся на удлиненные, так называемые "ауксиб-ласты", и укороченные - "брахибласты". Удлиненные побеги обеспечивают прирост дерева в высоту и разрастание кроны. У древесных покрытосеменных растений укороченные побеги формируются из боковых пазушных или придаточных почек, их междоузлия очень короткие (длина междоузлий равна толщине побега или меньше нее). Листьев на укороченных побегах меньше, чем на удлиненных, форма этих листьев часто имеет заметные отличия от формы листьев на удлиненных
побегах. Обычно укороченных побегов больше в нижней, более затененной, части кроны. У молодых быстро растущих деревьев укороченные побеги могут отсутствовать, но начиная с определенного возраста они обязательно появляются. Вместе с тем склонность к образованию укороченных побегов свойственна не всем видам.
У некоторых голосеменных хвойных (сосна, лиственница) наличие укороченных побегов (брахибластов) является обязательным родовым признаком. Однако следует иметь в виду, что брахибласты хвойных не являются прямым аналогом укороченных побегов покрытосеменных.
Побеги развиваются из почек, которые у большинства растений являются пазушными, то есть расположенными на узле побега в пазухе листа. По характеру расположения на побеге различают почки боковые, то есть пазушные, и почки верхушечные. При супротивном листорасположении на одном узле супротивно расположены две боковые почки (рис. 37). У некоторых растений верхушечная почка к концу лета отмирает естественным образом (липа, вяз), поэтому на следующий год "удлинение" стебля осуществляется за счет роста тех боковых почек, которые были расположены около верхушки побега. У некоторых растений наблюдается чередование способов нарастания побегов в зависимости от общего возраста ветви или всего растения, от экологических условий, поломки побегов.
101
Ясень Дуб Бук
Рис. 37. Три типа листорасположения: 1 - супротивное; 2 - очередное спиральное; 3 - очередное двухрядное (из Strasburger, Noll, Schenck, Schimper, 1967)
Почки древесных растений в зависимости от особенностей внешнего строения делят на почки закрытые покрытые почечными че-шуями, и почки открытые, у которых зашита зачатка побега выполняется не почечными че-шуями, а недоразвитыми мелкими зелеными листочками. Последний тип почек встречается
у некоторых древесных растений, например у крушины ломкой. Расположенные в пазухе отдельного листа почки могут быть одиночными в одних случаях или групповыми (множественными) в других (последние почки не следует путать с супротивными почками, которые расположены в пазухах разных листьев). Групповые почки, в свою очередь, подразделяются на сериальные почки (жимолости), которые расположены в ряд вдоль продольной оси побега, и коллатеральные почки, размещенные на узле в ряд, перпендикулярный к продольной оси побега. Побеги, которые развиваются из групповых почек, могут различаться длиной (в том числе укороченные и удлиненные), специализацией (одни вегетативные, другие репродуктивные) и другими признаками. Такие "пучковидные собрания" побегов - характерная отличительная особенность некоторых деревьев и кустарников.
Помимо рассмотренных выше существует целый ряд признаков почек, относящихся к разряду качественных. В частности, почки подразделяются на
почки сидячие, не имеющие удлиненного междоузлия в своем основании, и почки черешчатые, у которых такое междоузлие имеется. Размеры последнего невелики - около 1-2 мм, но по его наличию легко отличаются представители таких родов, как ольха, дерен и др. Различно число почечных чешуй, покрывающих почку, их может быть одна (ива, калина обыкновенная), две (липа, ольха), пять-семь или более. Форма почечных чешуй и их цвет, а также общая форма и абсолютные размеры почки, угол ее наклона к оси побега достаточно разнообразны и одновременно специфичны для отдельных родов и видов древесных растений.
К качественным особенностям почек относится и их различия по ростовой активности - они делятся на ростовые и спящие. Ростовые пазушные почки образуют боковые побеги, за счет них происходит ветвление стебля. Пазушные почки, не образовавшие на следующий год после их появления боковых побегов, могут в течение многих лет (часто десятков лет) оставаться живыми. Такие почки называются спящими. Спящие почки остаются в толще коры или из нее выступают, прирастая каждый год своим основанием на величину радиального прироста стебля. При повреждении кроны дерева спящие почки активизируются и дают нормальные, удлиненные, или укороченные побеги, восстанавливая синтезирующую поверхность кроны. За счет
102
спящих почек в комлевой части ствола (в зоне так называемой "корневой шейки") равно, как и за счет почек прикорневых капов, происходит вегетативное возобновление многих видов древесных растений.
Помимо пазушных почек, на стеблях древесных растений довольно обычны придаточные почки, возникающие в междоузлиях. Такие почки могут оставаться спящими или давать как удлиненные, так и укороченные побеги.
Почка является частью того комплекса структур, которые размещены на узле побега. В него входят следующие элементы: под почкой находится листовой рубец - участок узла, к которому прикрепляется черешок листа, рубец "открывается" после листопада. По краям листового рубца находятся прилистниковые рубцы у тех растений, которые имеют листья с прилистниками. На рубцах можно различать листовые следы - окончания сосудистоволокнистых пучков, которые обычно в виде темных точек размещены на поверхности листового рубца. Наконец, в некоторых редких случаях листовой рубец может находиться на некотором возвышении, которое одновременно и закрывает некоторую часть почки, скрывает ее. Такое возвышение называют листовой подушкой, поскольку она образована частью листового черешка (чубушник, актинидия и др).
Элементами характеристики стеблей древесных растений являются окраска коры ветвей и стволов, структура поверхности побегов (опушенность, наличие "желёзок") и форма их поперечного сечения, характер отслаивания старых участков коры, окраска внутренней "живой" части коры, наличие или отсутствие наплывов и др. Для описания большей части такого рода особенностей специальные термины, как правило, не употребляются.
Лист
Лист - неотъемлемая часть побега, отличающаяся большим разнообразием и одновременно наличием таких признаков, которые специфичны для отдельных родов, семейств и других систематических групп. По продолжительности жизни листа растения подразделяют на листопадные растения (лист живет 1 вегетационный сезон) и вечнозеленые растения (лист живет более одного года). Последнюю группу растений не разделяют далее, хотя в нее входят виды с различной продолжительностью жизни листьев например, 2-3 года (сосны), порядка 10 лет (пихты) и др. Листорасположение имеет три следующие основные формы: листья очередные (расположены по одному на узле побега), листья супротивные - расположены на узле по 2 и супротивно, листья мутовчатые - расположены на узлах по 3 и более. Основными частями листа являются листовая пластинка, черешок и прилистники. Указанные выше основные части листа, и прежде всего его форма и размеры, значительно отличаются, если сравнивать между собой нижние, средние и верхние листья побега, не говоря уже о почечных чешуях, которые, по существу, также являются листьями. Разнообразие листьев в рассматриваемом отношении носит название разнолистности или гетерофиллии. В описаниях отдельных ви
103
дов или родов растений приводятся главным образом характеристики срединных листьев побегов, поскольку они обладают в наибольшей степени видовыми или родовыми признаками.
Узловыми моментами в характеристике листьев являются особенности прикрепления листа к узлу побега, форма и структура листовой пластинки, типы расчленения листа, формы края, основания и верхушки листа, его жилкование.
По особенностям прикрепления листа к узлу побега различают листья сидячие - они не имеют черешка и прикреплены непосредственно основанием пластинки, листья стеблеобъемлющие сидячий лист с расширенным основанием, охватывающим стебель в узле, листья влагалищные имеют трубчатое влагалище различной длины и формы, листья черешковые обладают черешком различной формы. Последний тип листа преобладает у отечественных древесных растений.
В зависимости от общей структуры различают листья простые, состоящие только из одной листовой пластинки, и листья сложные, которые состоят из нескольких листовых пластинок, называемых листочками (рис. 38). Различают несколько типов сложных листьев: тройчатосложные (ракитник, клевер, клен маньчжурский), пальчатосложные (конский каштан, витекс, люпин), непарноперистосложные (ясень, многие рябины), парноперистосложные (карагана, горошек). Наконец, существуют дважды - и трижды сложные листья, у которых основной черешок листа разветвлен соответственно на черешки 2-го и 3-го порядка и листочки прикрепляются к этим разветвлениям (аралия) (рис. 38).
Важнейший признак формы простого листа или листочка сложного основан прежде всего на относительных размерах его длины и ширины, а также на положении максимальной ширины по отношению к его центру -яйцевидные, обратнояйцевидные, овальные, округлые. Названия других форм листьев - треугольные, ромбические, эллиптические, лопатчатые, сердцевидные, почковидные, игольчатые - основаны на сходстве с некоторыми фигурами или предметами (рис. 39).
Кроме простых листьев, у которых пластинка не имеет глубоких разрезов и выемок, выделяют группу расчлененных листьев, у которых пластинка имеет признаки, показанные на рис. 39. Название формы подобных листьев учитывает две особенности: количество выступов и глубину их расчленения, которая сравнивается с шириной одной из продольных половин листовой пластинки.
Лопастным листом считается такой, у которого полупластинка расчленена менее чем наполовину. У раздельного листа она расчленена более чем наполовину, а у рассеченного листа почти до основания, то есть почти до средней жилки листа. Рассеченный лист иногда похож на лист сложный, однако последний принципиально отличается тем, что состоит из нескольких листочков, каждый из которых почти всегда имеет маленький черешок и осенью отпадает самостоятельно от основного черешка листа.
104
Рис. 38. Сложные листья: 1 - тройчатосложный; 2 - пальчатосложный; 3, 4 - парноперистосложные; 5 - непарноперистосложный; 6 - прерывистоперистосложный; 7 -дваждыперистосложный; 8 - Триждыперистосложный
Сравнивая формы листьев разных видов древесных растений, а также формы листьев одного и того же растения, легко заметить проявления как сходства, так и различия. У большинства хвойных жилки погружены в мякоть мезофилла и снаружи незаметны.
Характеристика листа будет неполной без учета такого признака, как жилкование (рис. 40). У большинства хвойных жилки погружены в мякоть мезофилла и снаружи незаметны. У покрытосеменных древесных растений жилкование может быть перистым (от основания листовой пластинки в разные стороны расходятся несколько основных жилок, почти равных друг другу), параллельным (несколько почти одинаковых по диаметру жилок протянуты от основания до верхушки листовой пластинки параллельными рядами), дуговидное жилкование (отчасти сходно с предыдущим, но жилки не являются прямыми линиями и, кроме того, боковые жилки иногда отходят на разных уровнях от средней жилки).
105
Рис. 39. Форма простых листовых пластинок; 1 - игольчатый лист; 2 - линейный; 3 - продолговатый; 4 - ланцетный; 5 - овальный; 6 - округлый; 7 - яйцевидный; 8 - обратнояйцевидный; 9 - ромбический; 10 - лопатчатый; 11 - сердцевиднояйцевидный; 12 -почковидный; 13 - стреловидный; 14 - копьевидный
Наконец, на листьях хорошо выражены восковой налет (если он имеется) и опушение, характеристика последнего складывается из указания на морфологический тип волоска (волоски простые, ветвистые, звездчатые, железистые, жгучие, чешуйчатые, пузыревидные, двураздельные, прямые или изогнутые), а также на его густоту и характер создаваемой им "поверхности" (войлочное, паутинистое, шелковистое опушение и др.).
Корень
Корень - один из основных вегетативных органов листостебельных растений (за исключением мхов), служащий для прикрепления к субстрату, поглощения из него воды и питательных веществ, первичного превращения ряда поглощаемых веществ, синтеза органических соединений, дальнейшего перемещения их в другие органы растения, а также для выделения некоторых продуктов обмена. У ряда растений корни несут дополнительные функции, например, вместилища запасных веществ, а у корнеотпрысковых растений -органа вегетативного размножения.
Корень - осевой орган растения, по происхождению родственный стеблю. Морфологически он отличается от стебля отсутствием листьев, наличием чехлика и эндогенным ветвлением (возникновением боковых корней во
106
внутренних частях корня - в перицикле31). Типичный корень имеет узкоцилиндрическую или нитевидную форму. Зачаток корня существует уже в зародыше семени, при прорастании которого он развивается в главный корень. Для корня характерно акропетальное32 ветвление (более молодые боковые корни и их зачатки появляются ближе к верхушке корня.).
Рис. 40. Жилкование листьев: 1 - параллельное у злаков; 2 - дуговидное у ландыша; 3 - пальчатое у клена; 4 - сетчатое у ивы; 5 - дихотомическое у гинкго
У многих растений, кроме главного и боковых корней, имеются придаточные. Внешне они не отличаются от боковых и несут те же функции, но обычно развиваются на других органах растения - стеблях, листьях, подземных и надземных видоизменённых побегах (луковицах, клубнях, корневищах и т. д.), а также на старых корнях. Образование придаточных корней определяет возможность вегетативного размножения растений.
Кончик корня (рис. 41) прикрыт корневым чехликом, защищающим его в процессе роста от повреждений твердыми структурными элементами почвы. Корень растет в длину за счёт деления клеток верхушечной (апикальной) меристемы, в так называемой зоне деления, а также за счет значительного роста производных апикальной меристемы в длину в зоне роста, или растяжения. Длина зоны растяжения обычно равна 1-2 мм (иногда до 10 мм). За зоной роста расположена зона поглощения и дифференциации; самый поверхностный слой клеток этой зоны дифференцируется в эпиблему, клетки которой образуют корневые волоски, увеличивающие поверхность поглощения корня в 5-20 раз. По мере роста корня, вследствие перемещения апи
31 Прерицикл - слой клеток первичной меристемы в корнях и иногда в стеблях, окружающий проводящий цилиндр и располагающийся под эпидермой.
32 Акропетальный - направленный к верхушке . Противопоставляется базипетальному - направленному к основанию.
107
кальной зоны в почве, зона всасывания сдвигается в сторону апекса, на ней образуются новые корневые волоски, а старые отмирают.
Рис. 41. Строение кончика корня: 1 - зона поглощения; 2 - зона роста; 3 - корневой чехлик; 4 -верхушечная меристема (апекс) корня; 5 - прокамбий; 6 - корневые волоски; 7 - клетки эпиблемы, образующие корневые волоски (из БСЭ, 1989)
Совокупность корней одного растения называется корневой системой, общую форму и характер которой определяет соотношение роста главного, боковых и придаточных корней. При преобладающем росте главного корня образуется стержневая корневая система (дуб, люпин, хлопчатник). У некоторых древесных растений (сосна) образуется так называемая якорная корневая система - хорошо развитые боковые корни на некотором рас
стоянии от ствола резко уходят в глубину почвы (рис. 42). При слабом росте или раннем отмирании главного корня и преобладающем развитии большого числа придаточных корней образуется мочковатая корневая система (однодольные, из двудольных - виды лютика, подорожника и др.). Корни у хлебных злаков (ржи, пшеницы) проникают на глубину 1-1,5 м, у люцерны -до 10 м, у древесных растений могут углубляться на 10-12 (у дуба иногда до 15 м), хотя обычно твёрдый грунт препятствует проникновению их глубже 3-5 м. Площадь, занятая корневой системой одного растения хлебных злаков, достигает в поперечнике 40-60 см, тыквы - 6-8 м, у древесных растений обычно в 2-3 раза превышает диаметр кроны. Суммарная длина корней однолетнего растения может составлять несколько километров, а общая по
верхность корней, учитывая поверхность волосков, во много раз превышает
поверхность надземных органов.
По расположению в почве в зависимости от среды обитания различают корневые системы специализированные (поверхностные или глубинные) и универсальные, равномерно развивающиеся вширь и вглубь. Например, в северной лесной зоне на подзолистых почвах, часто пересыщенных влагой, плохо аэрируемых, с трудно проницаемой подпочвой, корневая система растений на 90-95% сосредоточена в поверхностных слоях (10-15 см). У ели, например, поверхностная корневая система стала видовым признаком. В зоне полупустынь и пустынь у одних растений имеются только поверхностные кони, которые используют осадки ранней весны либо влагу дождей и конденсационную влагу, оседающую в верхних слоях почвы в ночное время (кактусы). У других растений корни достигают грунтовых вод (у дуба, верблюжьей колючки). Универсальные корневые системы используют в разное время влагу разных горизонтов, вследствие чего растения могут вегетировать
108
всё лето (сосна обыкновенная, береза повислая, саксаул, эфедра). В зонах недостаточного увлажнения ярко проявляется ярусность в расположении корней различных видов растений, произрастающих совместно на одной площади.
Рис. 42. Якорные корни сосны обыкновенной. Макарьевский лесхоз Костромской области (фото В.В. Коровина)
Соотношение между корнями и надземными частями растений также зависит от почвенно-климатических условий. В северной влажной зоне масса корневой системы в 5-10 раз уступает массе надземных частей. Через корни растения вместе с водой поглощают из почвы ионы минеральных солей, а также некоторые продукты жизнедеятельности почвенных микроорганизмов и корневые выделения других растений. Поглощённые корнями соединения азота, фосфора и серы взаимодействуют с притекающими из листьев продуктами фотосинтеза с образованием аминокислот, нуклеотидов и других органических соединений. По сосудам ксилемы поглощенные корнями элементы в форме ионов (калий, кальций, магний, фосфор) и органических молекул (азот, сера) передвигаются в листья и стебли. В корнях синтезируются также алкалоиды (например, никотин), гормоны роста (кинины, гиббереллины) и другие физиологически активные вещества. У некоторых растений, преимущественно из семейства бобовых, на тонких корнях поселяются бактерии,
109
вызывающие образование клубеньков (формируются из паренхимных клеток корня). Клубеньковые бактерии способны поглощать азот из атмосферы.
Корни растений выделяют в почву ионы минеральных солей, аминокислоты, сахара, мочевину, фенольные соединения. У одних растений (например, из семейства паслёновых) корни выделяют также ауксины и вещества типа гиббереллинов, стимулирующие рост растений, у других - студнеобразные вещества, образующие корнезащитные чехлы. Выделения корней способствуют развитию в ризосфере33 почвенных микроорганизмов. С возрастом растения количество выделений и численность ризосферных микроорганизмов уменьшаются. Клубеньковые бактерии играют важную роль в азотном питании растений и повышении плодородия почвы. На поверхности или внутри корневых окончаний у многих древесных и травянистых растений поселяются грибы, образуя так называемую микоризу; в таких случаях корневые волоски на корнях не развиваются.
На корнях многих (корнеотпрысковых) растений образуются придаточные почки, дающие надземные побеги. У ряда растений корни служат местом отложения запасных питательных веществ, такие корни называются корнеплодами (морковь, свекла). У некоторых деревьев тропических лесов от стволов близ их основания отходят досковидные корни, так называемые контрфорсы. В мангровых лесах на болотах и морских побережьях на ветвях древесных растений образуются придаточные так называемые ходульные столбовидные корни - служащие для опоры и питания.
У лазящих лиан (например, у плюща) развиваются корни-прицепки. В тропиках многие эпифиты образуют придаточные воздушные корни, поглощающие воду из атмосферных осадков. У некоторых тропических эпифит-ных орхидей листья и стебли недоразвиты и их вегетативные органы представлены главным образом плоскими зелёными корнями, выполняющими функции ассимиляции. У растений, живущих на бедных кислородом почвах (например, у болотного кипариса, у мангровых деревьев и др.), имеются дыхательные корни, или пневматофоры, верхушки которых расположены над почвой или водой и снабжают подземные органы кислородом. У некоторых пальм и растений семейства мареновых часть горизонтальных корней превращается в защитные колючки. Корни паразитирующих на деревьях растений, например омелы, имеют вид длинных цилиндрических тяжей, расположенных в коре дерева. У растений-паразитов (заразиха, повилика и др.) и полупаразитов (марьянник, погремок и др.) корневая система развивается слабо; при этом окончания некоторых корней внедряются в виде особых присосок (гаусторий) в тело растения-хозяина, высасывая из него питательные вещества. У некоторых водных растений (например, у роголистника, пузырчатки и др.) корни отсутствуют, что связано со специфическими условиями их существования.
Корни многих растений широко используются человеком. Они имеют большое пищевое и хозяйственное значение. Корни, содержащие крахмал,
33
Ризосфера - зона обитания корней.
110
сахара, масла, алкалоиды, гуттаперчу, красящие и др. ценные вещества, применяются в медицине и промышленности. Растения с мощно развитой корневой системой используются для закрепления подвижных песков, оврагов и эродированных почв.
Стробилы хвойных и цветки покрытосеменных
Стробилы и цветки являются специализированными видоизмененными побегами, приспособлеными к половому размножению (рис. 43).
У хвойных растений (рис. 43 А) функции цветков выполняют однополые стробилы, которые входят в собрания стробилов (констробилы или сложные стробилы), неверно называемые иногда "соцветиями" (у не имеющих цветков хвойных, соцветий, разумеется, быть не может). Собрание мужских стробилов - микростробилов, иногда неудачно называемое "мужским колоском" - состоит из отдельных микростробилов на укороченных осях, на которых расположены сильно редуцированные микроспорофиллы - аналоги тычинок. Расположение микроспорофиллов у большинства хвойных спиральное. Форма микроспорофиллов весьма разнообразна. Представителям семейства сосновых, к которым относятся основные лесообразователи северной лесной зоны, свойственна форма, изображенная на рисунках 56, 58, 60, 63.
Женский стробил или макростробил, каковым является и хорошо известная шишка сосны, состоит из центральной оси, на которой сидят кроющие чешуи, в пазухах каждой из которой находится семенная чешуя. Их называют семенными потому, что на своей верхней стороне они несут семязачатки, превращающиеся при созревании шишки в семена. У представителей сосновых семязачатков всегда два. Семенную чешую с семязачатками принято называть мегастробилом.
У покрытосеменных растений в типичных случаях цветок расположен в пазухе кроющего листа, называемого прицветником, а на своей цветоножке несет один или два прицветничка. У многих групп растений, особенно с многоцветковыми соцветиями, прицветники и прицветнички (или только последние) отличаются формой и меньшими размерами по сравнению с обычными листьями побегов или полностью отсутствуют.
Цветок расположен на верхушке цветоножки и состоит из цветоложа, околоцветника, тычинок и пестика (рис. 44). Число и строение всех этих органов очень разнообразно и одновременно специфично для отдельных систематических групп цветковых растений.
Ill
Рис. 43. Сравнительная схема репродуктивной части голосеменных хвойных (на примере сосны обыкновенной - А) и покрытосеменных растений (Б): 1 - прорастающее семя; 2 - побеги с мужскими и женскими стробилами; 3 - мужской и женский стробилы; 4 - микроспорофилл (слева) и семенная чешуя с семязачатком (мегастробил); 5 - пыльца (микроспора); 6 - семенная чешуя во время оплодотворения; 7 - семенная чешуя с созревшими семенами; 8 - покрытосеменное двудольное растение с цветочной почкой; 9 -цветок во время оплодотворения; 10 - плод с семенем; 11 - прорастающее семя (из Stras-burger, Noll, Schenck, Schimper, 1967)
В плане половой специализации различают цветки обоеполые, однополые, имеющие только тычинки или только пестики, и цветки бесплодные, имеющие только околоцветник. Последний тип цветков обычно расположен на периферии соцветия и служит для привлечения насекомых-опылителей (калина).
112
Рис. 44. Цветки с околоцветником (вверху) и без него: 1 - полный обоеполый цветок; 2 - отдельные части цветка (схема); 3 - цветок пеона в продольном разрезе; 4 - обоеполый цветок у белокрыльника; 5 - ложнообоеполый цветок у ясеня (андроцей недоразвит); 6 - однополый тычиночный цветок ивы; 7 - однополый пестичный цветок ивы. ЦЛ -цветоложе; Ч - чашелистики; Л - лепестки венчика; Е - тычинки; П - пестики; ПЛ - прицветный лист
В тех случаях, когда разные экземпляры одного вида растения имеют только мужские или только женские цветки, растение называют двудомным (ива, тополь, облепиха). Если же раздельнополые цветки размещены на одном и том же экземпляре, то растение называют однодомным (береза, лещина, орех, каштан).
Околоцветник цветка в одних случаях может состоять из зеленой чашечки и венчика, в этом случае он имеет название двойной околоцветник. Простой околоцветник имеют те цветки, околоцветник которых состоит из одинаковых по форме, размеру и окраске листочков (вяз, копытень). Обычно цветки с двойным околоцветником имеют относительно небольшую окрашенную в зеленый цвет чашечку и более крупный различной (но не зеленой) окраски венчик. Однако встречаются виды растений, в цветках которых венчик может быть короче чашечки, или чашечка и венчик почти не отличаются по окраске, (купальница, многие мимозовые).
Околоцветник характеризуется рядом качественных параметров. Один из них - наличие или отсутствие срастания между его отдельными элементами. При отсутствии срастания говорят о раздельнолистной чашечке и раздельнолепестном венчике. Чашечку со сросшимися чашелистиками называют спайнолистной, а венчик со сросшимися лепестками - спайнолепестным.
Второй качественный параметр цветка касается его симметрии. Если элементы околоцветника равны по форме и размерам и через цветок можно
из
провести несколько плоскостей симметрии, то цветок считается правильным - актиноморфным (яблоня, чубушник). В других случаях, когда элементы околоцветника не равны по форме и размерам, а через цветок можно провести только одну плоскость симметрии, околоцветник считается неправильным зигоморфным, а его "половины" являются зеркальным отображением друг друга (бобовые, фиалка).
Тычинки (андроцей - мужская часть цветка) состоят из тычиночных нитей и пыльников, которые имеют разные размеры и окраску у отдельных видов. Совокупность тычинок цветка существует в двух формах: у одних видов растений тычинки свободные, у других сросшиеся своими нитями в трубку (бобовые) или сросшиеся только своими основаниями (чозе-ния, ива пурпурная) (рис. 45).
Рис. 45. Схема продольного разреза цветка: 1 - основание околоцветника; 2 - нектарники; 3 - зародышевый мешок с яйцеклеткой и другими его компонентами; 4 - стенка завязи; 5 - тычиночная нить; 6 - пыльник (продольный разрез); 7 - столбик; 8 - рыльце; 9 - пыльца, прорастающая на рыльце; 10 - одна из пыльцевых трубок достигла зародышевого мешка; 11 - пыльник (поперечный разрез)
Взаимоотношения тычинок с другими органами цветка тоже выражаются прежде всего в наличии или отсутствии срастания. Одна из форм подобного срастания заключается в том, что возникает так называемая цветочная трубка, или гипантий. Это чашевидное образование - продукт срастания между собой цветоложа, нижних частей ча-
шелистиков, лепестков и тычиночных нитей (яблоня, шиповник, боярышник). В других случаях тычинки прирастают к трубчатой части спайнолепестного венчика (сирень) или к основанию лепестков у раздельнолепестного
венчика.
В центре цветка находится пестик (гинецей - женская часть цветка), состоящий из трех элементов: завязь - нижняя, расширенная часть пестика, столбик - средняя, обычно узкая часть пестика и рыльце, имеющее разнообразную форму. Пестик состоит из отдельных элементов - плодолистиков, о числе которых можно судить по числу рылец, по числу гнезд в завязи или крупных жилок на ее стенках. Цветок может иметь несколько свободных пестиков, каждый из которых состоит из одного плодолистика (малина, ломонос). В других случаях пестик может состоять из нескольких плодолистиков, которые срастаются между собой различным образом. Различают верхнюю завязь, если тычинки и околоцветник прикрепляются к цветоложу ни-
114
же завязи, а также нижнюю завязь, если остальные органы цветка прикреплены к верхней части завязи. После опыления цветка семяпочки превращаются в семена, а пестик в целом - в плод. Иногда в образовании плода принимают участие: цветоложе, основания лепестков и чашелистиков (яблоня, шиповник, орех). В таком случае плоды называются полуложными.
Соцветие
По характеру расположения цветков на побегах можно выделять цветки одиночные и соцветия (рис. 46), которые представляют собой разветвленный побег с несколькими или многими цветками. В соцветии цветки располагаются в пазухах кроющих листьев или прицветников. На цветоножках цветков находятся прицветнички. Как одиночные цветки, так и соцветия могут быть верхушечными и боковыми. У некоторых растений встречаются оба способа размещения этих структур.
Рис. 46. Схематическое изображение простых (верхний ряд) и сложных (нижний ряд) соцветий; 1 - кисть; 2 - колос; 3 - початок; 4 - зонтик; 5 - головка; 6 - корзинка; 7 -щиток; 8 - монохазий (завиток); 9 - монохазий (извилина); 10 - дихазий; 11 - плейохазий (сложный зонтик); 12 - плейохазий (щитковидная метелка)
Разнообразие соцветий зависит от таких признаков, как длина оси соцветия и цветоножек, наличие или отсутствие прицветников и прицветнич-ков, способности оси соцветия к утолщению в процессе развития или к образованию утолщенного полого вместилища с цветками на внутренней стенке (инжир) и, наконец, от порядка распускания цветков соцветия - центростремительного (снизу вверх, от края к центру) или, наоборот, центробежного.
К числу соцветий с преимущественно центростремительным способом распускания цветков относятся многие простые соцветия: кисть - ось соцветия длиннее цветоножек (черемуха); сережка - похожа на кисть, но цветоножки короткие или отсутствуют (ива); колос - цветоножки отсутствуют,
115
цветки сидячие, часто двухрядные (пшеница); щиток - ось соцветия недлинная, цветоножки разной длины, у нижних цветков длиннее, чем у верхних (груша); зонтик - главная ось соцветия почти не развита, цветоножки прикрепляются почти в одной точке (элеутерококк); головка - ось соцветия расширенная, уплощенная, а цветки сидячие, расположены на ее поверхности и окружены листочками обертки (ромашка).
Центробежный характер распускания цветков свойственен следующим сложным соцветиям: завиток - ось заканчивается верхушечным и одним боковым цветком, от цветоножки которого отходит цветок - ось с цветком 3-го порядка и т. д. (незабудка) и извилина; полузонтик, или дихазий - ось заканчивается верхушечным и 2 боковыми цветками, от цветоножек которых отходят оси с цветками 3-го порядка и т. д. (липа). Кроме вышеназванных соцветий, существуют и более сложно устроенные: метелка, разветвленная иногда многократно (многие виды клена); сложный зонтик - или зонтик из частных зонтиков (аралия); щитковидное соцветие или щитковидная метелка (клен остролистный).
Плод
Плоды различают на основе нескольких морфологических признаков (рис. 47), многие из которых имеют существенное экологическое значение, так как определяют способ распространения плодов и семян. Среди этих признаков наиболее важными являются число семян в плоде, способ вскрывания плода или его естественное разделение на отдельные части, наличие или отсутствие у него сочных тканей, образование особых отростков - шипов, крыльев и др., которые связаны с определенным способом распространения семян и плодов. На основе этих признаков выделяют такие экологобиологические и морфологические группы плодов, как плоды сухие и сочные, одно - или многосемянные, вскрывающиеся или невскрывающиеся и др.
Рис. 47. Некоторые типы сухих плодов и семян: 1 - крылатый орех (береза); 2 - семя сосны с крыловидным придатком; 3 - дробный плод двукрылатка (клен); 4 - крылатый орех (вяз); 5 - крылатый орех (ясень); 6 - стручок (сем. крестоцветные); 7 - боб (сем. бобовые); 8 - листовка (сем. лютиковые); 9 - раскрывшаяся коробочка (осина); 10 - семя осины с пучком волосков. Относительные размеры не соблюдены
116
Основные типы плодов следующие: боб сухой многосемянной одногнездный плод, вскрывающийся двумя створками сверху вниз (семейство бобовые); листовка сухой многосемянной одногнездный плод, вскрывающийся одной продольной щелью (некоторые лютиковые); коробочка сухой многосемянной плод, одногнездный или многогнездный, вскрывающийся створками, зубчиками, крышечкой или дырочками (сем. ивовые, сирень); зерновка сухой односемянный невскрывающийся плод, у которого оболочка плода и покровы семени срастаются между собой (злаки); орех сухой односемянный невскрывающийся плод с кожистым покровом, обычно не приросшим к семенной кожуре (дуб, бук, лещина, береза); крылатка - сухой односемянный плод с крыловидными выростами (вяз, ясень); костянка -плод, у которого внешние покровы сочные, а внутренние деревянистые, образующие косточку с семенем внутри (сливовые); ягода - сочный многосемянной плод (крыжовник, смородина, актинидия).
Кроме этих основных плодов отечественных древесных растений существуют плоды членистые, которые благодаря поперечным перетяжкам распадаются на отдельные фрагменты, заключающие одно семя (софора, копеечник); плоды дробные, которые распадаются на отдельные односемянные фрагменты (клен, мальва); яблоко или яблочко, имеющие кожистые внешнюю и внутреннюю оболочки плода и мясистую, сочную среднюю оболочку (яблоня, рябина); стручок сухой многосемянной 2-гнездный плод, вскрывающийся снизу вверх двумя створками (редька, редиска, капуста). Следует иметь в виду, что встречаются плоды с переходными признаками: например, костянка не с сочной, а мучнистой полусухой мякотью (лох) и др. В тех случаях, если цветок имеет несколько свободных пестиков и каждый из них, естественно, становится плодом, для общего собрания таких плодов используют названия многокостянка (малина, ежевика), многоорешек (шиповник), многолистовка (магнолия, лимонник, лютиковые).
Развивающееся из семяпочки семя находится внутри плода у цветковых (покрытосеменных) растений или расположено открыто, в пазухе чешуевидного листа у голосеменных растений. На семени различают семяножку и рубчик место прикрепления семени. На семени некоторых растений существует мясистый придаток - присемянник (бересклет), или хохолок (тополь, ива) из волосков.
Наконец, существуют и такие образования, как соплодия, которые возникают из нескольких естественно срастающихся плодов соседних цветков, например, у голубой жимолости в отличие от обыкновенной.
Окончательная система классификации плодов до настоящего времени не разработана.
117
ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ НАСЛЕДОВАНИЯ ПРИЗНАКОВ
Изменчивость
Изменчивостью в биологи называется разнообразие признаков и свойств у особей и групп особей любой степени родства. Однако чаще это понятие относится к разнообразию признаков и свойств конкретных организмов внутри вида или популяции.
Изменчивость присуща всем живым организмам, поэтому в природе отсутствуют особи, идентичные по всем признакам и свойствам, даже у однояйцевых близнецов обнаруживаются отличительные признаки. Термин "изменчивость" может употребляться для обозначения способности живых организмов отвечать морфологическими и физиологическими изменениями на внешние воздействия, а также для характеристики преобразований форм организмов в процессе их эволюции.
Различают изменчивость наследственную (генотипическую) и ненаследственную (паратипическую - вызванную прямым воздействием факторов внешней среды); индивидуальную и групповую; прерывистую (дискретную - например, контрастные различия цвета у лепестков венчика цветков) и непрерывную (разная высота одновозрастных деревьев в лесу); качественную и количественную; независимую изменчивость разных признаков и коррелятивную (когда изменение одного признака тесно связано с изменением другого); направленную (определённую, по Ч. Дарвину) и ненаправленную (неопределённую, по Ч. Дарвину); адаптивную (приспособительную) и неадаптивную. При решении общих проблем биологии и особенно проблем эволюции наиболее существенно подразделение изменчивости на наследственную и ненаследственную, а также - на индивидуальную и групповую. Все перечисленные категории изменчивости могут встречаться в наследственной и ненаследственной, групповой и индивидуальной изменчивости. В публикациях последних лет показано, что наследуются не конкретные морфологические и физиологические изменения, их может быть бесчисленное множество, а сама способность к изменчивости. Разумеется, при этом наследственная передача потомству конкретных изменений не исключается.
Наследственная изменчивость обусловлена многими факторами, в том числе обменом фрагментами хромосом при кроссинговере, возникновением разных типов мутаций34 и их комбинаций в последующих скрещиваниях. Изменчивость, обусловленную возникновением мутаций, называют мутационной, а обусловленную дальнейшим комбинированием генов в результате скрещивания - комбинационной.
В индивидуальном развитии организма проявление наследственных признаков и свойств всегда определяется не только основными, ответственными за данные признаки и свойства генами, но и их взаимодействием со
34 Мутации - естественные или вызванные искусственно наследуемые изменения генетического материала, приводящие к изменению тех или иных признаков организма.
118
многими другими генами, составляющими генотип особи, а также условиями внешней среды, в которой протекает развитие организма. Исследованиями последних лет показано, что факторы среды обитания могут менять и наследственные свойства, что в первую очередь относится к организмам на эмбриональной стадии развития (в этом случае материнский организм выступает в качестве среды обитания). Это возвращает нас к положениям Ламарка о наследовании приобретенных в индивидуальном развитии признаков. Такие взгляды Ламарка и его последователей до последнего времени категорически отрицались.
В понятие условно ненаследственной изменчивости входят те изменения признаков и свойств, которые у особей или определённых групп особей вызываются воздействием внешних факторов (питание, температура, свет, влажность и т. д.). Чаще всего такие изменения (модификации) в их конкретном проявлении у каждой особи не передаются по наследству, они развиваются у особей последующих поколений лишь при наличии условий, в которых они возникли. Такая изменчивость называется также модификационной. Например, окраска многих насекомых при низкой температуре темнеет, при высокой - светлеет; однако их потомство будет окрашено независимо от окраски родителей, изменившейся в соответствии с температурой. Существует ещё одна форма условно ненаследственной изменчивости - так называемые длительные модификации, часто встречающиеся у одноклеточных организмов, но изредка наблюдаемые и у многоклеточных. Они возникают под влиянием внешних воздействий (например, температурных или химических) и выражаются в качественных или количественных отклонениях от исходной формы, обычно постепенно возвращающихся к исходному при последующем размножении. Они основаны, по-видимому, на изменениях относительно стабильных цитоплазматических структур. Некоторые современные исследователи считают, что длительные модификации могут приводить к соответствующим им наследственным изменениям, играющим важную роль в процессе эволюции, и приводят весьма убедительные тому примеры.
Между ненаследственной и наследственной изменчивостями существует тесная связь. Ненаследственных (в буквальном смысле) признаков и свойств нет, так как условно ненаследственные изменения являются отражением наследственно обусловленной способности организмов отвечать определёнными изменениями признаков и свойств на воздействия факторов внешней среды. При этом пределы ненаследственных изменений определяются нормой реакции генотипа на условия среды.
Все формы изменчивости изучают как внутри отдельных совокупностей живых организмов, когда исследуют различия признаков отдельных особей (индивидуальная изменчивость), так и при сравнении между собой различных совокупностей особей (групповая изменчивость); в основе любых межгрупповых различий также лежит изменчивость индивидуальная. Даже в пределах близкородственных групп нет абсолютно идентичных особей, которые не различались бы по степени выраженности каких-либо наследственных или условно ненаследственных признаков и свойств. Ввиду сложности
119
организации живых систем, даже у генотипически идентичных (например, однояйцевые близнецы) и развивающихся в практически одинаковых условиях особей всегда можно обнаружить хотя бы незначительные морфофизиологические различия, связанные с неизбежными флуктуациями условий среды и процессов индивидуального развития. Групповая изменчивость включает различия между совокупностями любых рангов - от различий между небольшими группами особей в пределах популяции до различий между царствами живой природы (животные - растения). В сущности, вся систематика организмов построена на сравнительном анализе групповой изменчивости.
Для изучения пусковых механизмов эволюционного процесса особое значение имеют различные формы внутривидовой групповой изменчивости. Большинство видов распадается на подвиды или географические расы. В случае полной изоляции географических форм они могут резко различаться по одному или нескольким признакам. Популяции, населяющие обширные территории и не разделённые резкими изолирующими барьерами, могут (благодаря перемешиванию и скрещиванию) постепенно переходить друг в друга, образуя количественные градиенты по тем или иным признакам (кли-нальная изменчивость). Географическая, в том числе и клинальная, изменчивость в природных условиях - результат действия факторов, приводящих к разделению исходной группы особей в ходе исторического формирования вида на две или несколько групп, различающихся по численным соотношениям генотипов.
В некоторых случаях различия между группами особей в пределах вида не связаны с различиями их генотипического состава, а обусловливаются модификационной изменчивостью (различными реакциями сходных генотипов на разные внешние условия). Так называемая сезонная изменчивость обусловлена влиянием на развитие соответствующих поколений разных погодных условий (например, у некоторых насекомых и травянистых растений, дающих два поколения в год, весенние и осенние популяции различаются рядом признаков). Иногда сезонные формы могут быть результатом отбора разных генотипов (например, рано- и поздноцветущие формы трав на сенокосных лугах: в течение многих поколений устранялись особи, цветущие летом, во время сенокоса). Большой интерес представляет экологическая изменчивость - различия между группами особей одного вида, растущими или живущими в разных местах (возвышенности и низменности, заболоченные и сухие участки и т. д.). Часто такие формы называются экотипами. Возникновение экотипов также может быть результатом как модификационных изменений, так и отбора генотипов, лучше приспособленных к местным условиям.
Наследственной изменчивостью обусловлены различные формы внут-рипопуляционного полиморфизма35. В некоторых популяциях наблюдается сосуществование двух или более ясно различимых форм (например, ели европейской - красношишечные и зеленошишечные формы; у двухточечной божьей коровки почти во всех популяциях встречаются чёрная форма с красными пятнами и красная форма с чёрными пятнами). В основе этого явления могут лежать разные механизмы: неодинаковая приспособленность сосуще
33 Полиморфизм - многоформенность. изменчивость морфологических признаков внутри популяции. Иногда это понятие применяют и к изменчивости функций организмов.
120
ствующих форм к условиям различных сезонов года, повышенная жизнеспособность гетерозигот, в потомстве которых постоянно выщепляются обе гомозиготные формы или другие, ещё недостаточно изученные механизмы. Таким образом, и групповая, и индивидуальная изменчивость включают изменения как наследственной, так и ненаследственной природы.
При этом, повторяем, следует помнить, что абсолютно ненаследственной изменчивости не бывает, так как наследуются не конкретные внутрипо-пуляционные вариации, а сама способность к изменчивости и пределы возможных внутривидовых вариаций.
Независимой изменчивости признаков противопоставляют коррелятивную изменчивость - взаимосвязанное изменение различных признаков и свойств: связь между высотой деревьев и их диаметром; ростом и весом особей (положительная корреляция) или темпом клеточного деления и величиной клеток (отрицательная корреляция). Корреляции могут быть обусловлены чисто генетическими причинами или взаимозависимостями процессов становления определённых признаков и свойств в индивидуальном развитии особей (онтогенетические корреляции), а также сходными реакциями разных признаков и свойств на одни и те же внешние воздействия (физиологические корреляции). Наконец, корреляции могут отражать историю происхождения популяций из смеси двух или более форм, каждая из которых привносит не отдельные признаки, а комплексы взаимосвязанных признаков и свойств (исторические корреляции). Изучение коррелятивной изменчивости имеет важное значение в палеонтологии (например, при реконструкции вымерших форм по отдельным ископаемым остаткам), в антропологии (например, при восстановлении черт лица на основе изучения черепа), в селекции и медицине.
Основные методы изучения изменчивости - описательный и биометрический. Совокупность этих методов позволяет исследовать как паратипическую (вызванную изменением факторов внешней среды), так и генотипическую компоненты общей фенотипической изменчивости. Так, первую можно изучать, сравнивая генотипически идентичные клоны и чистые линии, развивающиеся в разных условиях. Делаются попытки выделить чисто генотипическую изменчивость из общей фенотипической. Считается, что это возможно сделать на основе биометрического анализа гибридного потомства организмов, различающихся какому-либо контролируемому признаку. Однако, и это уже подчеркивалось, чисто паратипической изменчивости быть не может.
Наследственность и изменчивость живых организмов иногда противопоставляют как "консервативное" и "прогрессивное" начала. В действительности же они теснейшим образом связаны. Чёткое представление о категориях и формах изменчивости необходимо в хозяйственной практике и при построении эволюционных схем и теорий. Явления наследственности и изменчивости лежат в основе эволюционного процесса, они же являются исходным материалом в практической селекции растений и животных, при изучении ряда проблем медицинской географии и популяционной антропологии.
Законы Грегора Менделя
Таблица 2
121
Перечень генетических терминов, необходимых для дальнейшего изложения
Термин
Ген.
Аллели.
Локус Г омозигота
Г етерозигота
Фенотип
Г енотип
Доминантный
Рецессивный
Поколение F i
Поколение F2
Объяснение
Основная единица наследственности, определяющая данный признак.
Альтернативные формы одного и того же гена, определяющие альтернативные признаки Местоположение аллеля в хромосоме
Диплоид, содержащий два идентичных аллеля данного гена.
Диплоид, содержащий два разных (контрастирующих) аллеля данного гена.
Физическое или химическое проявление рассматриваемого признака
Имеющиеся у особи аллели в локусе, определяющем данный признак.
Аллель, определяющий фенотип как в гомозиготном, так и в гетерозиготном состоянии Аллель, определяющий фенотип только в гомозиготном состоянии
Первое гибридное поколение
Второе гибридное поколение, полученное от скрещивания особей из F i
Пример
Ген, определяющий цвет семени.
А или а
ЛА или аа
Аа
Желтая горошина, зеленая горошина.
АА, Аа, аа
А
а
В 1866 году вышла в свет краткая работа Грегора Менделя - чешского монаха и образованного для своего времени естествоиспытателя - «Опыты с растительными гибридами». С этого момента и отсчитывает свой возраст наука генетика. Исследованиям Менделя сопутствовала большая удача - он, сам того не подозревая, выбрал такие альтернативные, взаимоисключающие, признаки растений (а опыты он ставил с горохом), которые мы сейчас называем моногенными, контролируемыми парой аллельных генов (пазушные и верхушечные цветки (рис. 48); цветки красного цвета и белые; горошины желтые и зеленые, гладкие и морщинистые). Аллельные - значит, локализованные в одном участке (локусе) ДНК. Это были случаи простого наследования, когда одному гену соответствовал один признак, в природе таких случаев мало.
Рис. 48. Схематическое изображение побегов с цветками: А - цветки пазушные; Б - цветок верхушечный
Далее кратко рассмотрим три основных закона наследования признаков, которые Грегор Мендель выявил на основании своих опытов.
Закон единообразия первого поколения гибридов, или первый закон Менделя.
Для иллюстрации первого закона Менделя вос-
произведем его опыты по моногибридному скрещиванию растений гороха.
122
Скрещивание двух организмов называется гибридизацией, потомство от скрещивания двух особей с разной наследственностью называют гибридным, а отдельную особь из этого потомства - гибридом. Моногибридным называется скрещивание двух организмов, отличающихся друг от друга по одной паре альтернативных, взаимоисключающих, признаков. Следовательно, при таком скрещивании прослеживаются закономерности наследования только двух признаков, развитие которых обусловлено парой аллелей в одном локусе. Все остальные признаки, свойственные данным организмам, во внимание не принимаются.
Если скрестить растения гороха с пазушными и верхушечными цветками, то у всех полученных в результате этого скрещивания гибридов цветки будут пазушными (рис. 49).
Рис. 49. Генетическое объяснение одного из моногибридных скрещиваний, поводившихся Менделем. А - пазушные цветки (доминантный признак); а - верхушечные цветки (рецессивный признак); АА, Аа, аа -диплоидное состояние (2п), А, а - гаплоидное состояние (и)
Верхний ряд буквенных обозначений - "родительские" растения: АА - доминантная гомозигота (пазушные цветки), аа - рецессивная гомозигота (верхушечные цветки). Средний ряд: Л, а - гаметы (половые =
репродуктивные клетки). Нижний ряд: Аа - гетерозиготы, "дочерние" растения (Fу); все с пазушными цветками - внешне (фенотипически) проявляют признак, определяемый до-
минантным аллелем, однако рецессивный аллель в генотипе этих растений сохраняется.
Такая же картина будет наблюдаться при скрещивании растений, обладающих гладкой и морщинистой формой семян, все потомство первого поколения будет иметь гладкую форму семян. Следовательно, у гибрида первого поколения из каждой пары альтернативных признаков развивается только один. Второй признак как бы исчезает, не проявляется. Явление преобладания у гибрида признака одного из родителей Г. Мендель назвал доминированием. Признак, проявляющийся у гибрида первого поколения и подавляющий развитие другого признака, был назван доминантным, а противоположный, то есть подавляемый, признак - рецессивным. Если в генотипе организма два одинаковых аллеля в одном локусе, оба доминантные или оба рецессивные (АА или аа), такой организм называется гомозиготным. Если же из пары аллелей один доминантный, а другой рецессивный (Аа) то такой организм носит название гетерозиготного.
Закон доминирования - первый закон Менделя называют также законом единообразия гибридов первого поколения, так как у всех особей первого поколения проявляется один признак.
Неполное доминирование. В ряде случаев гибрид не воспроизводит полностью ни одного из родительских признаков. Все особи первого гибрид
123
ного поколения единообразны по данному признаку. Признак имеет промежуточный характер с большим или меньшим уклонением к условно доминантному или условно рецессивному состоянию. Так, в опытах Менделя при скрещивании растения с красной окраской цветков (АА) с растением, имеющим белые цветки (аа), в первом гибридном поколении (Е7) проявлялась промежуточная розовая окраска цветка (Аа).
Закон расщепления, или второй закон Менделя
Если гетерозиготных потомков первого гибридного поколения, одинаковых по изучаемому признаку, скрестить между собой, то во втором поколении признаки обоих родителей проявятся в определенном числовом соотношении: 3/4 особей будут иметь доминантный признак и 1/4 - рецессивный (рис. 50).
Рис. 50. Свободное опыление между "дочерними" растениями (Fi) и генотипы F2 (нижний ряд буквенных обозначений). Гомозиготы АА и гетерозиготы Аа фенотипически неразличимы, у них пазушные цветки. Только у рецессивных гомозигот цветки верхушечные. Численное отношение растений с пазушными и верхушечными цветками во втором гибридном поколении 3:1. Следует иметь в виду, что это отношение построено на статистически средних данных.
Явление, при котором скрещивание гетерозиготных особей приводит к образованию потомства, часть которого несет доминантный признак, а часть рецессивный, называется расщеплением. Следовательно, рецессивный признак у гибридов первого поколения не исчез, а был только подавлен и проявился во втором гибридном поколении.
Закон независимого комбинирования, или третий закон Менделя был сформулирован им после проведения следующего опыта. В ранее проведенных экспериментах было установлено, что гладкая поверхность оболочки семян (горошин) контролируется доминантным аллелем гена (А), а морщинистая - рецессивным (а). В этой же партии семян были желтые и зеленые горошины, и в опытах было установлено, что желтый цвет у тех же самых семян контролируется доминантным аллелем (Б), а зеленый - рецессивным (б).
При скрещивании растений с гладкими и морщинистыми семенами в первом гибридном поколении (Е7) были получены гибриды с гладкими семенами - доминантный аллель (А) подавил фенотипическое проявление признака морщинистости (а). Одновременно с использованием семян из той же партии скрестили растения с желтыми (доминантный аллель Б) и зелеными (рецессивный аллель б) семенами. В Е7 получили все растения с желтыми семенами. Таким образом были получены две партии семян, все горошины в которых были гладкими и желтыми. Растения, выращенные из этих двух партий семян между собой свободно скрещивались (такое скрещивание называ
124
ется дигибридным). Морфологическая характеристика семян, полученных в F2 приведена в таблице 3.
При скрещивании особей, отличающихся друг от друга по двум и более парам альтернативных признаков, гены и соответствующие им признаки наследуются независимо друг от друга и комбинируются во всевозможных сочетаниях. Например, если у одного из скрещиваемых родительских экземпляров гороха красные цветки и гладкая поверхность семян (горошин), а у второго - белые цветки и морщинистые горошины, то в гибридном потомстве будут появляться растения и повторяющие по контролируемым признакам родителей, и имеющие смешанные признаки, скажем, красные цветки и морщинистые горошины. Но при этом для каждой пары альтернативных признаков распределение особей подчиняется второму закону Менделя.
Таблица 3
Формирование фенотипов F2 при дигибридном скрещивании двух гетерозиготных особей, различающихся по двум парам контрастирующих признаков
?\d АБ Аб аБ аб
АБ АБАБ АбАБ аБАБ абАБ
Гладк. желт. Гладк. желт. Гладк. желт. Гладк. желт.
Аб АБАб АбАб аБАб абАб
Гладк. желт. Гладк. зел. Гладк. желт. Гладк. зел.
аБ АБаБ АбаБ аБаБ абаБ
Гладк. желт. Гладк. желт. Морщ. желт. Морщ. желт.
аб АБаб Абаб аБаб абаб
Гладк. желт. Гладк. зел. Морщ. желт. Морщ. зел.
Пояснение к таблице. А - гладкие семена (доминантный признак), а - морщинистые семена (рецессивный признак), Б - желтые семена (доминантный признак), б - зеленые семена (рецессивный признак). Общее отношение - 9 гладких желтых : 3 гладких зеленых : 3 морщинистых желтых : 1 морщинистый зеленый
Наследственность
Законы Г. Менделя носят статистический характер, они применимы лишь к совокупностям организмов и не раскрывают сущности и причин наследственности, что могло быть достигнуто только с помощью экспериментального изучения наследственности разными методами и прежде всего гибридологическим анализом, основы которого и были заложены Менделем. При анализе наследования количественных признаков, таких как высота деревьев, их диаметр, длина однолетних побегов, площадь листовой пластинки, отсутствовала четкая картина расщепления, что ставило под сомнение правильность выводов Менделя. (Мендель изучал качественные признаки: цвет, форма, расположение цветков). Однако в дальнейшем гибридологический и биометрический анализы наследования количественных признаков показали,
125
что и здесь наследственность, предположительно, сводится к дискретной, но наследование при этом полигенное. В этом случае расщепление трудно обнаружить, так как оно происходит по многим генам, действие которых на признак осложняется сильным влиянием условий внешней среды. Таким образом, хотя признаки и можно разделять на качественные и количественные, сами термины “качественная” и “количественная” наследственность не оправданы, так как обе категории наследственности принципиально одинаковы.
Развитие цитологии (науки о клетке) привело в конце XIX - начале XX веков к постановке вопроса о материальных основах наследственности. Важным шагом стало понимание функции ядра клетки как носителя наследственности. При изучении процесса оплодотворения была установлена индивидуальность хромосом и их качественные различия. Законы Г. Менделя получили цитологическое объяснение. Однако наиболее обстоятельное обоснование хромосомной теории наследственности было дано в работах Т. Моргана и его школы (начиная с 1911 г.), в которых было показано соответствие между генетическими и цитологическими данными. В опытах на дрозофиле (плодовой мушке - Drosophila sp.) были зафиксированы случаи нарушения закона независимого распределения признаков - сцепленное наследование. Это явление было объяснено близким расположением генов, определяющих сцепленные признаки, в одной хромосоме (и, соответственно, в ее парном гомологе). Изучение частоты рекомбинаций между сцепленными генами (в результате кроссинговера) позволило составить карты расположения генов в хромосомах.
Генетические карты хромосом отображают реально существующий линейный порядок размещения генов в хромосомах и важны как в теоретических исследованиях, так и при проведении селекционной работы, т. к. позволяют сознательно подбирать пары признаков при скрещиваниях, а также предсказывать особенности наследования и проявления различных признаков у изучаемых организмов. Последнее очень важно и в медицине.
Обмен участками хромосом с находящимися в них генами при крос-синговере происходит с различной вероятностью, зависящей от расстояния между ними (чем дальше друг от друга гены, тем выше вероятность кроссинговера и, следовательно, рекомбинации).
Выше сказанное позволило сформулировать основные положения хромосомной теории наследственности:
1. Гены располагаются в хромосомах, различные хромосомы содержат неодинаковое число генов, набор генов каждой из негомологичных хромосом уникален.
2. Гены в хромосоме расположены линейно, каждый ген занимает в хромосоме определенный локус (место).
3. Гены, расположенные в одной хромосоме, образуют группу сцепления и вместе (сцеплено) передаются потомкам, число групп сцепления равно гаплоидному набору хромосом.
4. Сцепление не абсолютно, так как в профазе мейоза может происходить кроссинговер и гены, находящиеся в одной хромосоме, разобщаются.
126
Сила сцепления зависит от расстояния между генами в хромосоме: чем больше расстояние, тем меньше сила сцепления, и наоборот.
4. Генетические карты хромосом составляют для каждой пары гомологичных хромосом. Группы сцепления нумеруют последовательно, по мере их обнаружения. Кроме номера группы сцепления, указывают полные или сокращённые названия мутантных генов, их расстояния от одного из концов хромосомы, принятого за нулевую точку, а также место центромеры.
Составить генетические карты хромосом можно только для объектов, у которых изучено большое число мутантных генов. Например, у дрозофилы идентифицировано свыше 500 генов, локализованных в её 4 группах сцепления, у кукурузы - около 400 генов, распределённых в 10 группах сцепления. В последние годы в научной литературе появились сведения о завершении работ по составлению полных генетических карт всех хромосом человека.
Генетика пола. Количество групп сцепленных генов оказалось равным количеству пар хромосом, присущих данному виду. Важнейшие доказательства хромосомной теории наследственности были получены при изучении наследования, сцепленного с полом. Ранее цитологи открыли в хромосомных наборах ряда видов животных особые, так называемые половые хромосомы, которыми самки отличаются от самцов. В одних случаях самки имеют 2 одинаковые половые хромосомы (XX), а самцы - разные (XY), в других -самцы-2 одинаковые (XX, или ZZ), а самки - разные (XY, или ZW). Пол с одинаковыми половыми хромосомами называется гомогаметным, с разными - гетерогаметным. Женский пол гомогаметен, а мужской гетерогаметен у некоторых насекомых ( в том числе у дрозофилы) и у всех млекопитающих. Обратное соотношение - у птиц и бабочек. Ряд признаков у дрозофилы наследуется в строгом соответствии с передачей потомству Х-хромосом. Самка дрозофилы, проявляющая рецессивный признак, например белую окраску глаз, в силу гомозиготности по этому гену, находящемуся в Х-хромосоме, передает белую окраску глаз всем сыновьям, так как они получают свою Х-хромосому только от матери. В случае гетерозиготности по рецессивному сцепленному с полом признаку самка передает его половине сыновей. При противоположном определении пола (самцы XX, или ZZ, сам-ки-ХУ, или ZW) особи мужского пола передают сцепленные с полом признаки дочерям, получающим свою X (=Z ) хромосому от отца. Иногда в результате нерасхождения половых хромосом при мейозе возникают самки строения ХХГ и самцы ХУУ (такие хромосомные нарушения называются трисомией). Возможны также случаи соединения Х-хромосом концами; тогда самки передают сцепленные Х-хромосомы своим дочерям, у которых и проявляются сцепленные с полом признаки. Сыновья же похожи на отцов (такое наследование называется гологеническим ). Если наследуемые гены находятся в У-хромосоме, то определяемые ими признаки (в случае гетерогаметности мужского пола) передаются только по мужской линии - от отца к сыну (такое наследование называется голандриче-ским). Хромосомная теория наследственности вскрыла внутриклеточные механизмы наследственности, дала точное и единое объяснение всех явлений наследования при половом размножении, объяснила сущность изменений наследственности, то есть изменчивости.
Теория нехромосомной наследственности. Первенствующая роль ядра и хромосом в наследственности не исключает передачи некоторых признаков и через цитоплазму, в которой обнаружены структуры, способные к самовоспроизведению. Различают ядер-ную наследственность, связанную с передачей наследственных признаков, находящихся в хромосомах ядра (иногда ее называют хромосомной наследственностью ), и внеядерную наследственность, зависящую от передачи самовоспроизводящихся структур цитоплазмы (митохондрии, пластиды). Единицы цитоплазматической (нехромосомной) наследственности при мейозе не распределяются по дочерним гаметам поровну. Поэтому при половом
127
размножении потомство при нехромосомной наследственности воспроизводит признаки только одного из родителей (чаще матери). Внеядерная наследственность реализуется и при вегетативном размножении, не сопровождаясь перераспределением генов, что обеспечивает константную передачу признаков из поколения в поколение, нарушаемую только соматическими мутациями.
Молекулярная генетика. Применение новых физических и химических методов, а также использование в качестве объектов исследования бактерий и вирусов резко повысили разрешающую способность генетических экспериментов, привели к изучению наследственности на молекулярном уровне и бурному развитию молекулярной генетики. Впервые Н. К. Кольцов (1927 г) выдвинул и обосновал представления о молекулярной основе наследственности и о матричном способе размножения “наследственных моле-кул”.В 40-х гг. XX в. была экспериментально доказана генетическая роль дезоксирибонуклеиновой кислоты ( ДНК ), а в 50-60-х гг. установлена ее молекулярная структура и выяснены принципы кодирования генетической информации.
Информация о составе, строении и характере обмена составляющих организм веществ (прежде всего белков и нуклеиновых кислот) и связанных с ними функциях, получается от предков в виде совокупности генов. У многоклеточных форм при половом размножении генетическая информация передаётся из поколения в поколение через посредство половых клеток - гамет, единственная функция которых - передача и хранение генетической информации. У микроорганизмов и вирусов имеются особые типы ее передачи. Генетическая информация заключена преимущественно в хромосомах, где она зашифрована в определённой линейной последовательности нуклеотидов в молекулах дезоксирибонуклеиновой кислоты - ДНК (генетический код).
Генетический код - это система зашифровки наследственной информации в молекулах нуклеиновых кислот, реализующаяся у животных, растений, бактерий и вирусов в виде последовательности нуклеотидов. В природных нуклеиновых кислотах - дезоксирибонуклеиновой (ДНК) и рибонуклеиновой (РНК) - встречаются 5 распространённых типов нуклеотидов (по 4 в каждой нуклеиновой кислоте), различающихся по входящему в их состав азотистому основанию. В ДНК, как нам уже известно, встречаются основания: аденин (А), гуанин (Г), цитозин (Ц), тимин (Т); в РНК вместо тимина присутствует урацил (У). Американский учёный Г. Гамов предложил (1954) модель триплетного генетического кода, т. е. такого, в котором 1 аминокислоту в составе белка кодирует группа из трёх последовательных нуклеотидов, называемых кодоном, в составе нуклеиновой кислоты.
По мере изучения наследственности на субклеточном и молекулярном уровне углублялось и уточнялось представление о гене. Если в опытах по наследованию различных признаков ген постулировался как элементарная неделимая единица наследственности, а в свете данных цитологии его рассматривали как изолированный участок хромосомы, то на молекулярном уровне ген представляет собой входящий в состав хромосомы участок молекулы ДНК, способный к воспроизведению и имеющий специфическую структуру,
128
в которой закодирована программа развития одного или нескольких признаков организма. В 50-х гг. на микроорганизмах было показано, что каждый ген состоит из ряда различных участков, которые могут мутировать и между которыми может происходить кроссинговер. Так подтвердилось представление о сложной структуре гена, развивавшееся еще в 30-х гг. А.С. Серебровским и Н.П. Дубининым на основе данных генетического анализа.
В настоящее время известно, что ген представлен длинным фрагментом ДНК (дезоксирибонуклеиновой кислоты), состоящим в среднем из 1500 кодирующих (несущих конкретную генетическую информацию) нуклеотидных пар (АТ и ГЦ) и еще большего числа нуклеотидных пар (ни) как бы балластных, не несущих никакой информации. Самое малое число ни, необходимое живой (бактериальной) клетке - 0,76-106нп, среднее число ни в эвкариотических клетках обычно на несколько порядков больше.
В 1967-1969 гг. был осуществлен синтез вирусной ДНК вне организма, а также химический синтез гена дрожжевой аланиновой транспортной РНК. Новой областью исследования стала наследственность соматических клеток в организме и в культурах тканей. Открыта возможность экспериментальной гибридизации соматических клеток разных видов. В связи с достижениями молекулярной биологии явления наследственности приобрели ключевое значение для понимания ряда биологических процессов, а также для множества вопросов практики.
Наследственность и эволюция. Эта проблема, несмотря на ее достаточно долгую историю, остается далеко не решенной. Во всем мире почти безоговорочно принята теория эволюции, базирующаяся на работах Чарльза Дарвина, со времени публикации которых прошло уже 150 лет. В свое время работы Дарвина сыграли непереоценимую положительную роль - человечество, как бы очнувшись, освободилось от примитивных теологических взглядов на окружающий мир. Правда, большинство научных положений Дарвина были неоднократно высказаны и до него, но эти, часто разрозненные, и мало доступные широкой общественности публикации долго оставались незамеченными - развитие общества к тому времени еще не достигло необходимого уровня. Однако книга Дарвина "Происхождение видов ..." (1859) и его последующие работы вышли уже своевременно - теперь общественность была готова к их восприятию. Теория эволюции по Дарвину овладела умами, это относится и к нашему времени.
Последующие разработки в области теории эволюции, а затем и открытия генетиков с большим или меньшим успехом подгонялись и часто продолжают подгоняться под дарвиновскую теорию. В результате возник неодарвинизм, который в последнее время переименовался в синтетическую теорию эволюции (СТЭ). Слабые стороны собственно дарвиновской концепции (естественный отбор случайных положительных уклонений от нормы) и СТЭ были замечены давно и неоднократно. Однако теперь общественность в силу вполне естественного консерватизма не уступает "ни пяди земли", закрепленной за дарвинизмом, точно так же, как она до Дарвина не уступала своих теологических воззрений.
129
Подробнее на этом вопросе мы остановимся в следующей главе. Здесь же следует иметь в виду, что закономерности наследственности имеют огромное значение для практики сельского хозяйства, пищевой промышленности и медицины. На них основываются выведение новых и совершенствование существующих сортов растений и пород животных. Изучение закономерностей наследственности привело к научному обоснованию применявшихся ранее эмпирических методов селекции и к разработке новых научных приёмов в практике (экспериментальный мутагенез, гетерозис, полиплоидия и др.).
Генетика человека - это отрасль генетики, тесно связанная с антропологией и медициной. Генетику человека условно подразделяют на антропогенетику, изучающую наследственность и изменчивость нормальных признаков человеческого организма, и генетику медицинскую, которая изучает его наследственную патологию (болезни, дефекты, уродства). Генетика человека связана также с эволюционной теорией, так как исследует конкретные механизмы эволюции человека и его место в природе, с психологией, философией, социологией. Из общей генетики человека вытекают и интенсивно развиваются цитогенетика, биохимическая генетика, иммуногенетика, генетика высшей нервной деятельности, физиологическая генетика.
В генетике человека вместо классического гибридологического анализа применяют генеалогический метод, который состоит в анализе распределения в семьях (точнее, в родословных) лиц, обладающих данным признаком (или аномалией) и не обладающих им, что раскрывает тип наследования, частоту и интенсивность проявления признака и т. д. При анализе семейных данных получают также цифры эмпирического риска, т. е. вероятность обладания признаком в зависимости от степени родства с его носителем. Генеалогическим методом уже показано, что более 1800 морфологических, биохимических и других признаков человека наследуется по законам Менделя. Например, тёмная окраска кожи и волос доминирует над светлой; пониженная активность или отсутствие некоторых ферментов определяется рецессивными генами, а рост, вес, уровень интеллекта и ряд др. признаков - «полимерными» генами, т. е. системами из множества генов. Многие признаки и болезни человека, наследующиеся сцеплено с полом, обусловлены генами, локализованными в X- или У-хромосоме. Таких генов известно около 120. К ним относятся гены гемофилии А и В, недостаточности фермента глюкозо-6-фосфат-дегидрогеназы, цветовой слепоты и др. Другой метод генетики человека - близнецовый метод. Однояйцовые близнецы (ОБ) развиваются из одной яйцеклетки, оплодотворённой одним спермием; поэтому набор генов (генотип) у ОБ идентичен. Разнояйцевые близнецы (РБ) развиваются из двух и более яйцеклеток, оплодотворённых разными спермиями; поэтому их генотипы различаются так же, как у братьев и сестёр.
Приведенные примеры это отдельные частности. В целом же генетика человека представляет собой самостоятельную развитую науку.
Наследственность и среда. Гены проявляют свои функции не в пустоте, а в такой высокоорганизованной системе, как клетка, которая сама находится в определенном окружении - среди других клеток или во внешней среде. Каков бы ни был генотип, его свойства проявляются лишь в той степени, в какой это позволяют окружающие условия.
Растение, выращиваемое в темноте, остается белым (бесхлорофилль-ным) и хилым ; оно неспособно извлекать световую энергию, необходимую для обмена веществ, даже в том случае, когда все его клетки содержат генетическую информацию, необходимую для развития хлоропластов, а также синтеза и деятельности хлорофилла. Генетически однородные растения (кло
130
ны), выращенные на фоне разных почвенных условий, или выращенные с применением удобрений и без них, будут существенно различаться фенотипически.
Наконец, фенотип организма представляет собой результат взаимодействий между генотипом и средой в каждый данный момент его жизни и на каждом этапе его индивидуального развития.
Действия среды могут быть отнесены к двум типам, хотя в реальной обстановке они часто налагаются друг на друга. С одной стороны, это сильные воздействия, приводящие к полному или частичному подавлению выражения генетических потенций, с другой - слабые влияния, выражающиеся лишь в небольших изменениях степени их выражения. Первый тип воздействий зависит от случайных, часто критических обстоятельств, второй обычен и неразрывно связан с функционированием живой материи.
Индивидуальное развитие высшего организма начинается со стадии зиготы (оплодотворенной яйцеклетки). Наследственные потенции, получаемые им от родителей, проявляются лишь постепенно, в ходе длительного и сложного процесса развития, и начиная с первых делений зиготы, в их реализации принимает участие среда.
Для генов будущего организма исходной средой служит цитоплазма яйца (яйцеклетки), происходящего от материнского организма и воплощающего в себе клеточную непрерывность. Этого может оказаться достаточно, чтобы ориентировать развитие эмбриона в направлении, не совпадающем с его собственным генотипом.
Сравнение внутрипарных различий между однояйцевыми и разнояйцевыми близнецами позволяет судить об относительном значении наследственности и среды в определении свойств человеческого организма.
По большому счету вопросы, связанные со взаимодействием наследственности и среды обитания организмов, подробно рассматриваются в ставшей самостоятельной науке - экологии.
Болезни человека, связанные с мутациями. Геном человека включает несколько миллионов генов, способных к тому же по-разному влиять на развитие признаков. В результате мутаций и перекомбинации генов возникает присущее человеку разнообразие по самым разным признакам. Гены человека мутируют каждый с частотой от 1 на 100 000 до 1 на 10 000 000 гамет на поколение. Распространение мутаций среди больших групп населения изучает популяционная генетика человека, позволяющая составить карты распространения генов, определяющих развитие нормальных признаков и наследственных болезней. Особый интерес для популяционной генетики человека представляют изоляты -группы населения, в которых по каким-либо причинам (географическим, экономическим, социальным, религиозным и др.) браки заключаются чаще между членами какой-либо группы. Это приводит к повышению частоты кровного родства вступающих в брак, а значит, и вероятности того, что рецессивные гены перейдут в гомозиготное состояние и проявятся фенотипически, что особенно заметно при малочисленности изолята.
Исследования в области Генетики человека продемонстрировали наличие естественного отбора в человеческих популяциях. Однако отбор у человека приобретает специфические черты: он интенсивно действует только на эмбриональной стадии (например, самопроизвольные аборты - отражение такого отбора). Отбор в человеческом обществе осуществляется посредством дифференциальной брачности и плодовитости, то есть в ре
131
зультате взаимодействия социальных и биологических факторов. Мутационный процесс и отбор обусловливают огромное разнообразие (полиморфизм) по ряду признаков, присущее человеку, что делает его с биологической точки зрения необычайно пластичным и приспособленным видом.
Широкое использование в генетике человека цитологических методов способствовало развитию цитогенетики, где основной объект исследования - хромосомы, т. е. структуры клеточного ядра, в которых локализованы гены. Установлено, что хромосомный набор в клетках тела человека (соматических) состоит из 46 хромосом, причём женский пол определяется наличием двух Х-хромосом, а мужской — Х-хромосомы и У-хромосомы. В зрелых половых клетках находится половинное (гаплоидное) число хромосом. Митоз, мейоз и оплодотворение поддерживают преемственность и постоянство хромосомного набора как в ряду клеточных поколений, так и в поколениях организмов. В результате нарушений указанных процессов могут возникать аномалии хромосомного набора с изменением числа и структуры хромосом, что приводит к возникновении хромосомных болезней, которые нередко выражаются в слабоумии, развитии тяжёлых врождённых уродств, аномалий половой дифференцировки или обусловливают самопроизвольные аборты.
Ранняя диагностика наследственных заболеваний позволяет применить необходимые методы лечения. Существенно важен учёт наследственности в реакции разных людей на лекарства и другие химические вещества, а также в иммунологических реакциях организма. Бесспорна роль молекулярно-генетических механизмов в этиологии злокачественных опухолей. Явления наследственности предстают в разной форме в зависимости от уровня жизни, на котором они изучаются (молекула, клетка, организм, популяция). Но в конечном счёте наследственность обеспечивается самовоспроизведением материальных единиц наследственности (генов и цитоплазматических элементов), молекулярная структура которых известна. Закономерный матричный характер их ауторепродукции нарушается и мутациями отдельных генов или перестройками генетических систем в целом. Всякое изменение в ауторепродуцирующемся элементе наследуется константно.
Генетическая инженерия. Что такое генетическая инженерия? Генетическая инженерия - это раздел молекулярной генетики, связанный с целенаправленным созданием новых комбинаций генетического материала. Основа прикладной генетической инженерии - теория гена. Созданный генетический материал способен размножаться в клетке-хозяине и синтезировать заданные человеком продукты обмена.
Генетическая инженерия возникла в 1972 году, в США. Тогда лаборатория П. Берга получила первую гибридную ДНК или (рек-ДНК). Она соединяла в себе фрагменты ДНК фага, кишечной палочки и обезьяньего вируса. При получении рек-ДНК образуется чаще всего несколько структур, из которых только одна является нужной. Поэтому обязательный этап составляет селекция и молекулярное клонирование рек-ДНК, введенной путем трансформации в клетку-хозяина. Существует 3 пути селекции рек-ДНК: генетический, иммунохимический и гибридизационный с мечеными ДНК и РНК.
В результате интенсивного развития методов генетической инженерии получены клоны множества генов рибосомальной и транспортной РНК, гистонов, глобина мыши, кролика, человека, коллагена, овальбумина, инсулина человека и др. пептидных гормонов, интерферона человека и прочее. Это позволило создавать штаммы бактерий, производящих многие биологически активные вещества, используемые в медицине, сельском хозяйстве и микробиологической промышленности. На основе генетической инженерии возникла отрасль фармацевтической промышленности, названная «индустрией ДНК» Это одна из современных ветвей биотехнологии.
Для лечебного применения допущен инсулин человека (хумулин), полученный посредством рек-ДНК. Кроме того, на основе многочисленных мутантов по отдельным генам созданы высокоэффективные тест-системы для выявления генетической активности факторов среды, в том числе для выявления канцерогенных соединений.
132
За короткий срок генная инженерия оказала огромное влияние на развитие молекулярно-генетических методов и позволила существенно продвинуться по пути познания строения и функционирования генетического аппарата. Генная инженерия имеет большие перспективы в лечении наследственных болезней, которых на сегодняшний день зарегистрировано около 2000. Эта наука призвана помогать в исправлении ошибок природы. Достигнуты большие успехи в клонировании. Клон, или группа клеток, образуется делением одной клетки. Каждая соматическая клетка человека несет всю необходимую для формирования организма наследственную информацию. Если она начнет делиться, то вырастет новый организм, то есть организм с таким же генотипом, что и у клетки-родоначальницы клона. В 1997 г. доктор Ян Вилмут в Шотландии в г. Эдинбурге получил с группой ученых ягненка Долли ( искусственным путем ). Этот ягненок не имеет отца, так как клетка была взята у матери. Возникло опасение, что эксперименты по генной инженерии могут быть опасны для человечества. В 1974 г. специальная Комиссия американских биологов опубликовала сообщение генетикам мира, в котором рекомендовала воздержаться от экспериментов с некоторыми видами ДНК, пока не будут разработаны меры безопасности.
Необходимо было разработать ограничительные меры. 30 июля 1997 г. комитет по науке в Конгрессе США проголосовал за полный запрет экспериментов, связанных с клонированием людей. Президент еще раньше запретил выделение денег на эти эксперименты. В России в 1996 г. Государственная Дума приняла закон о государственном регулировании в области генной инженерии.
Гены индивидуальности. Одно из "чудес", которое мы наблюдаем ежедневно и ежечасно, - неповторимая индивидуальность каждого человека, живущего на Земле. Ученым долгое время не удавалось найти ключ к этой загадке. Известно, что вся информация о строении и развитии живого организма “записана” в его геноме - совокупности генов. Считается, что внутри одного вида геномные различия очень незначительны. Например, ген окраски глаз у человека отличается от гена окраски глаз у кролика, однако у разных людей этот ген устроен одинаково и состоит из одинаковых последовательностей ДНК Существует огромное разнообразие белков, из которых построены живые организмы, и удивительное многообразие генов, кодирующих эти белки. В геноме каждого человека есть какие-то области, определяющие его индивидуальность. Некоторые гены человека отличаются от генов крысы всего несколько нуклеотидами - знаками генетического кода. Другие гены у них разные, но одинаковые у двух людей. Изменчивость, связанная с существованием генов, подобных генам группы крови у человека, также не объясняет огромного разнообразия природных белков. В 1985 г. были обнаружены в геноме человека особые сверхизменчивые участки- мини сателлиты. Эти участки ДНК оказались индивидуальными у каждого человека и с их помощью удалось получить “портрет“ его ДНК, то есть определенных генов. Этот “портрет” - сложное сочетание темных и светлых полос, похожее на слегка размытый спектр, или на клавиатуру из темных и светлых клавиш разной толщины. Это сочетание называют ДНК-отпечатками (по аналогии с отпечатками пальцев) или “ДНК-профиль”. На основе сверхизменчивых последовательностей ДНК были сконструированы специальные маркеры, или зонды ДНК Маркеры, помеченные радиоактивным изотопом, добавляют к обработанным специальным образом ДНК, с которыми первые находят сходные сверхизменчивые участки на ДНК и присоединяются к ним. Эти участки становятся радиоактивными, так что их можно выявить с помощью радиоавтографии. У каждого человека распределение таких мест индивидуально. Там, где маркеры присоединились к большому числу сверхизменчивых участков на ДНК (много радиоавтограф ических сигналов) - это широкая темная полоса. Где мало мест присоединения, - узкая темная полоса. Г де их совсем нет, - светлая полоса. Итак, ученые обнаружили, что геном человека буквально “насыщен” сверхизменчивыми последовательностями ДНК Стали обнаруживаться неуловимые прежде индивидуальные последовательности ДНК
133
После разгадки индивидуальности человека, встал вопрос: обладают ли такой же индивидуальностью другие организмы? Существуют ли у них сверхизменчивые последовательности ДНК? Ученые должны были найти универсальный маркер, одинаково пригодный как для бактерий, так и для человека. Им оказался бактериофаг (вирус бактерий). Это открытие было чрезвычайно важно для работы генетиков и селекционеров. Выяснилось, что с помощью отпечатков ДНК можно провести идентификацию личности гораздо более успешную, чем это позволяли сделать традиционные методы отпечатков пальцев и анализ крови. Вероятность ошибки - одна на несколько миллионов. Новым открытием сразу же воспользовались криминалисты, которые быстро и эффективно применили его на практике. С помощью ДНК-отпечатков можно расследовать преступления не только настоящего времени, но и глубокого прошлого.
Генетические экспертизы по установлению отцовства - наиболее частый повод обращения судебных органов к генетической дактилоскопии. В судебные учреждения обращаются мужчины, сомневающиеся в своем отцовстве, и женщины, желающие получить развод на основании того, что их муж не отец ребенка. Идентификацию материнства можно проводить по отпечаткам ДНК матери и ребенка в отсутствии отца, и наоборот, для установления отцовства достаточно ДНК-отпечатков отца и ребенка. При наличии же материала матери, отца и ребенка ДНК-отпечатки выглядят не сложнее, чем картинка из школьного учебника: каждая полоса на ДНК-отпечатке ребенка может быть “адресована” либо отцу, либо матери.
Наиболее интересны прикладные аспекты генетической дактилоскопии. Встает вопрос паспортизации по отпечаткам ДНК преступников-рецидивистов, введения в картотеки следственных органов данных об отпечатках ДНК наряду с описанием внешности, особых примет, отпечатков пальцев.
ПУТИ эволюции
Происхождение жизни на Земле, разнообразие ее проявления и объединяющая живые существа структурная и функциональная близость занимают одно из центральных мест в биологии.
В этой главе мы рассмотрим основные, наиболее распространенные теории возникновения организмов и пути их развития. Следует отметить, что данные, которыми мы будем оперировать, и выведенные из них заключения соответствуют сегодняшним взглядам; эти взгляды постоянно пересматриваются, и их достоверность ограничена уровнем наших знаний в каждый текущий момент.
Современное представление о происхождении Земли
Теории, касающиеся возникновения Земли, да и всей Вселенной, разнообразны и далеко не окончательны. Согласно одной из них - теории стационарного состояния - Вселенная существовала всегда. Согласно другим -она могла возникнуть в результате «большого взрыва», или родилась в одной из черных дыр, или же была создана Творцом. Вопреки бытующим представлениям, наука не может полностью опровергнуть идею о божественном сотворении Вселенной, так же как и некоторые теологические взгляды не обязательно отвергают возможности того, что в возникновении Земли имели место процессы, объяснимые на основе законов природы.
134
Теорию создания Вселенной и Земли волей Творца мы не рассматриваем, это компетенция религиозных учений. Впрочем, иногда нам придется вспоминать и об этой теории.
Согласно наиболее распространенным современным представлениям наша планета образовалась около 4,5 млрд, лет назад. В процессе формирования Земли из частиц протопланетного36 облака постепенно увеличивалась ее масса. Росли силы тяготения, а следовательно, и скорости частиц, падавших на планету. Кинетическая энергия частиц превращалась в тепло, и Земля все сильнее разогревалась. При ударах на ней возникали кратеры, причем выбрасываемое из них вещество уже не могло преодолеть земного тяготения и падало обратно.
Чем крупнее были падавшие объекты, тем сильнее они нагревали Землю. Энергия удара освобождалась не на поверхности, а в глубине, равной примерно двум поперечникам внедрившегося тела. А так как основная масса на этом этапе поставлялась планете телами размером в несколько сотен километров, то энергия выделялась в слое толщиной порядка 1000 км. Она не успевала излучиться в пространство, оставаясь в недрах Земли. В результате температура на глубинах 100-1000 км могла приблизится к точке плавления. Дополнительное повышение температуры, вероятно, вызвал распад короткоживущих радиоактивных изотопов.
По-видимому, первые возникшие расплавы представляли собой смесь жидких железа, никеля и серы. Расплав накапливался, а затем вследствие более высокой плотности просачивался вниз, постепенно формируя земное ядро. Таким образом, дифференциация (расслоение) вещества Земли могла начаться еще на стадии ее формирования. Ударная переработка поверхности и начавшаяся конвекция, несомненно, препятствовали этому процессу. Но определенная часть более тяжелого вещества все же успевала опустится под перемешиваемый слой. В свою очередь дифференциация по плотности приостанавливала конвекцию и сопровождалась дополнительным выделением тепла, ускоряя процесс формирования различных зон в Земле.
Предположительно ядро образовалось за несколько сотен миллионов лет. При постепенном остывании планеты богатый никелем железоникелевый сплав, имеющий высокую температуру плавления, начал кристализовы-ваться - так (возможно) зародилось твердое внутреннее ядро. К настоящему времени оно составляет 1,7 % массы Земли. В расплавленном внешнем ядре сосредоточено около 30 % земной массы.
Развитие других оболочек продолжалось гораздо дольше и в некотором отношении не закончилось до сих пор. Литосфера сразу после своего образования имела небольшую толщину и была очень неустойчивой. Она снова поглощалась мантией, разрушалась в эпоху так называемой великой бомбардировки (от 4,2 до 3,9 млрд, лет назад), когда Земля, как и Луна, подвергалась ударам очень крупных и довольно многочисленных метеоритов. На Луне и сегодня можно увидеть свидетельства метеоритной бомбардировки - много
36
Прото... - первая часть сложных слов, означающая первенство, первичность, первобытность чего-либо.
135
численные кратеры и моря (области, заполненные излившейся магмой). На нашей планете активные тектонические процессы, а также воздействие атмосферы и гидросферы практически стерли следы этого периода.
Около 3,8 млрд, лет назад сложилась первая легкая и, следовательно, "непотопляемая" в магме, гранитная кора. В то время планета уже имела воздушную оболочку и океаны; необходимые для их образования газы усиленно поставлялись из недр Земли в предшествующий период. Атмосфера тогда состояла в основном из углекислого газа, азота и водяных паров. Кислорода в ней было мало, но он вырабатывался в результате, во-первых, фотохимической диссоциации воды и, во-вторых, фотосинтезирующей деятельности простых организмов, таких как сине-зеленые водоросли, которые появились примерно 3 миллиарда лет назад (см. приложение 1).
600 млн. лет назад на Земле было несколько подвижных континентальных плит, весьма похожих на современные. Новый сверхматерик Пангея появился значительно позже. Он существовал 300-200 млн. лет назад, а затем распался на части, которые и сформировали нынешние материки.
Что ждет Землю в будущем? На этот вопрос можно ответить лишь с большой степенью неопределенности, абстрагируясь как от возможного внешнего, космического воздействия, так и от деятельности человечества, преобразующего окружающую среду, причем не всегда в лучшую сторону.
Теории возникновения жизни на Земле
Среди главных, в том числе и древнейших, теорий возникновения жизни на Земле следует назвать следующие:
1 - жизнь не возникала и не исчезала (стационарное состояние);
2 - жизнь была создана сверхъестественным Началом в определенное время (креационизм);
3 - жизнь возникала неоднократно из неживого вещества (самопроизвольное зарождение);
4 - жизнь занесена на нашу планету извне (панспермия);
5 - жизнь возникла в определенный исторический промежуток времени в результате процессов, подчиняющихся объективным химическим и физическим закономерностям (биохимическая эволюция).
Последнее положение можно рассматривать и как обращение к креационизму (кто или что определило сами объективные закономерности?).
Кратко остановимся на перечисленных теориях.
Теория стационарного состояния
Согласно этой теории, Земля никогда не возникала, а существовала вечно; она всегда поддерживала жизнь, а если и изменялась, то очень мало. Виды живых существ также существовали всегда. Животные и растения могли или менять свою численность, или вымирать. Большая часть доводов в пользу этой теории была связана с такими неясными аспектами эволюции, как наличие разрывов в палеонтологической летописи.
136
Несостоятельность этой теории доказана современными достижениями в области астрономии, геологии, палеонтологии - установлен примерный возраст Земли и примерное (± 0,5 млрд, лет) время возникновения жизни.
Креационизм
Теорию сотворения Богом Земли и жизни на ней мы просто не рассматриваем, так как она относится к области веры и не имеет прямого отношения к науке. Однако, в данном случае уместна оговорка - считается, что Святые писания носят аллегорический характер, что допускает определенную широту толкования отдельных положений Библии, Корана и других религиозных канонических документов.
Самопроизвольное зарождение
Эта теория была распространена в древнем Китае, Вавилоне и Египте, ее придерживался Аристотель (384-322 гг. до и. э.). Аристотель считал, что природа совершает плавный переход от безжизненных объектов к животным, что отдельные "частицы" вещества содержат некое "активное начало", которое при определенных условиях может создать живой организм. Надо сказать, что в такой трактовке возникновения жизни есть элементы современных воззрений, которые сформировалось после Ламарка, Дарвина и последних открытий в биологии.
В прошлом были попытки экспериментально подтвердить правильность теории самозарождения, которые в наше время выглядят комичными. Так, в начале XVII века знаменитый в то время голландский естествоиспытатель Ван Гельмонт описал эксперимент, в ходе которого он за три недели якобы создал мышей. Ему для этого потребовались лишь грязная рубашка, горсть пшеницы и темный шкаф. Активным веществом по мнению самого ученого был человеческий пот.
Панспермия
Эта теория не предлагает никакого механизма первичного возникновения жизни, считается, что жизнь на нашу планету была занесена из космоса, то есть переносит эту проблему в какое-то другое, неизвестное, место Вселенной. Поэтому ее просто нельзя считать теорией возникновения жизни. Что же касается самой возможности переноса каких-то организмов на нашу планету извне, то в последние время в научной, но чаще в научно-популярной литературе, в TV-передачах и интернете появляются в разной степени убедительные сведения об обнаружении форм живой материи в упавших на землю метеоритах. Правда, доводы в пользу биологической природы последних ученым пока не кажутся достаточно убедительными. Вообще же, по данным современных космических исследований, вероятность обнаружить жизнь в пределах Солнечной системы вне Земли ничтожна.
Биохимическая эволюция
Легкие газы в атмосфере молодой Земли из-за меньшей, чем сейчас, гравитации не удерживались и терялись в космическом пространстве. Однако простые соединения - вода, диоксид углерода, метан, аммиак могли удерживаться. Исходя из этого, академик А.И. Опарин (1923) высказал предположение, что первые органические вещества, вероятнее всего углеводороды, мог
137
ли создаваться в океане из более простых, в том числе перечисленных выше, соединений. По мнению Опарина, разнообразие находившихся в океанах простых соединений, большая площадь поверхности Земли, доступность энергии (солнечная радиация, поток космических лучей, молнии) и масштабы времени позволяют предположить, что в океанах постепенно накопились органические вещества и образовался тот «первичный бульон», в котором могла зародиться жизнь (самопроизвольное зарождение).
Теория Опарина нашла поддержку. В 1953 году Стэнли Миллер моделировал в лаборатории условия земли до возникновения жизни и в подобных модельных условиях смог синтезировать органические соединения такие, как некоторые аминокислоты, аденин, простые сахара (рис. 51). В дальнейшем, путем подобных опытов, удалось получить даже нуклеотидные цепи длиной в шесть мономерных единиц, т.е. простые нуклеиновые кислоты.
К вакуумному насосу и газовому баллону (для удаления атмосферного воздуха и введения газов)
Направление потока
Кипящая вода (поставляет водяные пары -и обеспечивает циркуляцию)
Подогрев
Газовая камера
Метан
—Д—Аммиак
Водяные пары Водород
Электрический разряд высокого разрешения (источник энергии)
Предполагаемый состав первичной атмосферы
Водяная рубашка (охлаждение)
Отделитель жидкости (накапливает продукты химических реакций)
Рис. 51. Установка Стэнли Миллера, в которой он синтезировал аминокислоты из газов, создавая условия, предположительно существовавшие в атмосфере первобытной Земли. Газы и водяные пары, циркулировавшие в установке под высоким давлением, подвергали в течение недели воздействию электрических разрядов. Из по-
лучившегося раствора было выделено 15 аминокислот, в их числе глицин, аланин, аспарагиновая кислота (из Грин, Стаут, Тейлор, 1990)
Далее, в соответствии с положениями Опарина, возникшие таким образом органические соединения могли объединяться в более или менее крупные сгустки, которые, имея коллоидную форму и будучи гидрофильными, могли обосабливаться, образуя вокруг себя водяную пленку; поглощая металлы, образовывать ферменты и сформировать при участии липидов первичную мембрану. Таким образом, могли возникнуть первые примитивные клетки.
Такое логическое построение, частично подтвержденное опытами, и являет собой принимаемое в наше время многими учеными представление о возникновении жизни на земле. Правда, следует оговориться, что принимается оно далеко не всеми. Так, американский астроном Фред Хойл сравнительно недавно высказался примерно в том духе, что вероятность возникновения живой клетки из-за случайного взаимодействия молекул соизмерима с веро
138
ятностью того, что ураган, проносящийся над мусорной кучей, может случайно собрать Боинг-747.
Современный дарвинизм или синтетическая теория эволюции
В общем виде синтетическая теория эволюции (СТЭ) представляет процесс эволюции как постепенное развитие более сложных организмов из существовавших ранее более простых. Здесь сразу же следует обратить внимание на то, что она базируется на предпосылке, будто изначально простое существо уже существовало или простые существа уже существовали.
Эволюционные гипотезы высказывались задолго до выхода в свет всемирно известных работ Чарльза Дарвина. Среди дошедших до нас первых достоверных источников можно назвать труды Аристотеля из Стагиры (384-322), например "О возникновении животных". Правда, большинство из такого рода работ, в том числе и более поздние, носили характер более или менее удачных догадок. Первой, весьма убедительной для своего времени и вполне эволюционной в современном понимании была теория французского ученого Жана Батиста Ламарка (он, кстати, впервые ввел термин "биология"), опубликованная им в 1809 году. Согласно представлениям Ламарка изменения среды могут привести к изменению формы живых организмов. Такую же творческую роль, по его мнению, играет тренировка. В случае интенсивного использования органа его эффективность и величина будут возрастать, а при неиспользовании может наступить дегенерация и атрофия. Эти изменения признаков или органов, приобретенные организмами в течении жизни, согласно Ламарку, наследуются, т.е. передаются потомкам. Например, шея у жирафа вытянулась вследствие того, что эти животные из поколения в поколение стремились дотянуться до листьев в кронах высоких деревьев.
Ламарк, вне сомнения, был прав в том, что условия среды влияют на формообразовательные процессы в мире живых существ, однако, как было установлено в дальнейшем, тренировка вызывает изменения лишь у тренирующейся особи, и совсем не влияет на потомство. Для доказательства последнего положения немецкий генетик Август Вейсман (1834-1914) на протяжении нескольких "мышиных" поколений отрезал новорожденным мышам хвосты. Рассуждая по Ламарку, бесхвостые мыши, не употребляя в жизни и не тренируя хвосты, должны были со временем совсем их лишиться, однако, мыши продолжали рождаться, как и прежде, такими же хвостатыми. Однако последние исследования в области онтогенеза животных и особенно в области иммунологии показали, что Ламарк был неправ лишь частично - наследование признаков, приобретенных в эмбриональной фазе развития, на самом деле имеет место и, вероятно, играет важную роль в процессе эволюции.
В июле 1858 года, ставший к тому времени уже известным, естествоиспытатель Чарльз Дарвин и признанный в науке зоогеограф Рассел Уоллес выступили одновременно на заседании Королевского Линнеевского общества с изложением практически одинаковых взглядов на процесс изменения живых существ во времени. А в ноябре 1859 года Чарльз Дарвин опублико
139
вал книгу «Происхождение видов путем естественного отбора». Все 1250 экземпляров этой книги были проданы в первый же день, и вполне справедливо считается, что по своему воздействию на человеческое мышление она уступала только Библии.
Естественный отбор
Согласно Дарвину и Уоллесу, механизмом, с помощью которого из ранее существовавших видов возникают новые, служит естественный отбор. Эта гипотеза (или теория) по большому счету основана на трех наблюдениях и двух выводах.
Наблюдение первое: особи, входящие в состав популяции37, обычно обладают большим репродуктивным потенциалом (число потомков часто бывает избыточным, например, число плодов у видов березы, семян у сосны и ели).
Наблюдение второе: число особей в каждой данной популяции примерно постоянно, несмотря на избыточность размножения.
Наблюдение третье: во всех популяциях существует изменчивость.
Вывод первый: многим особям не удается выжить и оставить потомство. В популяции происходит «борьба за существование».
Вывод второй: в борьбе за существование те особи, признаки которых наилучшим образом приспособлены к условиям жизни, обладают преимуществом и оставляют после себя больше потомков, чем менее приспособленные особи.
Второй вывод и содержит гипотезу о естественном отборе, который может служить механизмом эволюции.
Доказательства существования естественного отбора согласно основным положениям СТЭ
Установлено что если бы каждая женская гамета на земле была оплодотворена и смогла бы развиться в половозрелую особь, то теоретически Земля через несколько дней оказалась бы перенаселенной. Однако природа выдвигает тысячи лимитирующих факторов, которые не дают реализоваться такой теоретической возможности. К числу этих факторов относятся в первую очередь ограниченность пищи, света и пространства. Следовательно, непрерывная конкуренция между индивидуумами за необходимые организмам компоненты среды приводит к «борьбе за существование», контролирующей оптимальную численность популяций.
Наблюдения показали, что изменчивостью обладают почти все, по крайней мере очень многие, признаки живых организмов. Изменчивы они и по степени приспособленности к условиям окружающей среды. Следовательно действие естественного отбора проявляется в преимущественном выживании, а следовательно - в преимущественной возможности успешно размножаться особей, более приспособленных к условиям существования.
140
Следует отметить, что приведенные доказательства основаны преимущественно на рассуждениях, экспериментальных подтверждений выше высказанным положениям до настоящего времени не получено.
Эволюционное учение Дарвина, интерпретированное с позиций современных научных достижений в биологии и является неодарвинизмом или синтетической теорией эволюции. Неодарвинизм можно определить как теорию органической эволюции путем естественного отбора признаков, детерминированных генетически.
Для того, чтобы признать СТЭ необходимо:
1. Установить факт изменения форм жизни во времени (эволюция в прошлом). Этот факт можно считать установленным - данные палеонтологии убедительно показывают, что формы существования жизни в геологическом прошлом неоднократно менялись.
2. Выявить механизм, осуществляющий эволюционные изменения. Считалось и в основном продолжает считаться, что этим механизмом является естественный отбор. Однако это основное положение СТЭ оказалось наиболее спорным.
3. Продемонстрировать эволюцию, происходящую в настоящее время (эволюция в действии). До настоящего времени хороших, убедительных, примеров эволюции в действии не найдено.
Критика синтетической теории эволюции и другие эволюционные учения
Остановимся несколько подробнее на изложенном выше втором условии признания СТЭ.
Синтетическая теория эволюции, в основе которой лежит учение Дарвина, начала формироваться после выхода в свет работ Гуго де Фриза (начало XX века), открывшего существование наследуемых мутаций, и считавшего, что мутации лежат в основе видообразования. Далее многочисленными исследованиями было показано, что внутривидовая изменчивость включает и мутации, и изменения, возникающие под влиянием факторов внешней среды. Последние считались ненаследственными.
В дальнейшем становление СТЭ происходило в основном за счет развития генетики и, главным образом, генетики популяций. Эта наука на подопытных организмах в лаборатории, реже в полевых опытах, которые чаще всего проводили лесные генетики, а также часто и чисто математически, моделировала так называемые микроэволюционные процессы, проходящие в популяциях - мутации, миграцию и дивергенцию, изоляцию, отбор38. В ста
38 Мутации - возникающие естественно или вызываемые искусственно изменения наследственных свойств организма в результате перестроек и нарушений в генетическом материале организма - хромосомах и генах. Миграция - включение в данную популяцию генотипов из другой популяции или пространственное перемещение небольшой части исходной популяции.
Дивергенция - расхождение признаков и свойств у первоначально близких групп организмов в процессе
эволюции или в опыте.
141
новлении этого направления генетики участвовало множество всемирно известных ученых, в числе которых С.С. Четвериков, Ф.Г. Добжанский, С. Райт, Р.А. Фишер, Н.В. Тимофеев-Рессовский.
Казалось бы факторы эволюции установлены, найдены многочисленные и весьма убедительные подтверждения основных дарвиновских положений. Однако по прошествии сравнительно небольшого времени оптимизм сменился сомнениями. Стало очевидным, что так называемые микроэволю-ционные изменения вовсе не эволюционные, ни к каким эволюционным сдвигам они не приводят, а лишь меняют частоту аллелей в модельных популяциях. И интерес к генетике популяций стал заметно снижаться - кроме того, выяснилось, что лабораторным опытам нет подтверждения в природе.
Разумеется, завоевания популяционной генетики вовсе не были бесполезными, они весьма важны, например, для селекции, но в понимание процессов эволюции ничего не внесли.
Надежды объяснить процессы эволюции с позиций СТЭ были перенесены на бурно развивающуюся биохимическую или молекулярную генетику, затем на иммунологию, последняя совсем не оправдала такие надежды и, можно сказать, окончательно раскачала основы СТЭ.
Здесь следует остановиться на презумпции39 универсальности и непогрешимости СТЭ. Учение Дарвина в свое время столь сильно повлияло на образ мышления и мировосприятие цивилизованного человечества, что в последствии любое возражение против основ этого учения воспринималось как посягательство на святыню. Все открытия в биологии, особенно в генетике, имеющие отношение к эволюционной теории, так или иначе приспосабливали к концепциям СТЭ, а если приспособить было трудно или невозможно, то их просто не принимали во внимание. Надо сказать, что подобные явления наблюдались во все времена. Не случайно возник афоризм, приведенный в одной из публикаций Ю.В. Чайковского: "Наука пятится вперед", и здесь больше истины, чем шутки. Научные идеи, обгоняющие современное и общепринятое представление о каком то предмете или процессе, просто не замечают (затылком не увидишь). Именно презумпция непоколебимости дарвинизма заставляет и в наши дни, не взирая ни на какие, даже самые убедительные, прошлые и современные возражения, переносить из публикации в публикацию, из учебника в учебник взгляды 150-летней давности.
Обратимся к самому давнему научно обоснованному возражению против эволюционной роли естественного отбора случайных мелких уклонений от нормы (в современном понимании - мутаций), дающих некоторые преимущества особи. Это возражение известно как "кошмар Дженкина". (Флеминг Дженкин - был инженером, человеком разносторонним и эрудированным, он соотечественник и современник Чарльза Дарвина.)
Изоляция - возникновение барьеров (территориальных, экологических, физиологических, поведенческих, генетических), препятствующих свободному скрещиванию организмов.
Отбор - дифференциальная выживаемость и размножаемость потомства.
39 Презумпция - обычно презумпция невиновности - обвиняемый считается невиновным пока его вина не доказана. В данном случае - СТЭ верна, пока не доказана ее несостоятельность.
142
Дженкин первым высказал мысль о том, что направленный искусственный отбор, проводимый человеком для выведения новых пород скота и сортов растений, не может быть моделью естественного отбора. В нем нет места случайностям, а незначительные уклонения от нормы не принимаются во внимание. (Дарвин в результатах искусственной селекции видел пример эволюционных изменений.)
Но главное возражение - собственно "кошмар" для Дарвина и его теории - состояло в том, что любое полезное уклонение от нормы, если оно все-таки возникнет, очень скоро исчезнет, растворившись среди "нормальных", не несущих этого уклонения особей, при свободном скрещивании с ними. Дарвин не нашел достойного возражения (потому и "кошмар"). В наше же время возражение примерно таково - полезное для вида уклонение может и не исчезнуть, если оно произошло в малой изолированной популяции. Однако для реализации такого гипотетического случая (в природе его пока никто не наблюдал) необходимо совпадение множества маловероятных случайностей. Во-первых, для внешнего (фенотипического) проявления полезной для организма мутации (а вероятность такого события ничтожно мала) нужен еще хотя бы один точно такой же мутант (большинство мутаций рецессивны); во-вторых, он должен находиться, на таком расстоянии от первого, чтобы обеспечить их скрещивание; в-третьих, и в этом случае, если даже получилось потомство, несущее такое же полезное уклонение, одной пары таких организмов мало - природа не поощряет инбридинга (близкородственных скрещиваний), нужны еще несколько таких же маловероятных событий. Иными словами, мы должны многократно перемножать малые вероятности, что, естественно, приведет к практически полной невероятности. А ведь рассматриваемый нами "кошмар" относится к основному положению учения Дарвина и СТЭ!
К настоящему времени накопилось такое множество возражений против дарвинистского понимания процесса эволюции, что их трудно перечислить. Имеются в виду не только взгляды самого Дарвина, но и концепции СТЭ, основанные как на взглядах Дарвина, не всегда правильно трактуемых, так и на данных современной науки.
Наибольшей критике подверглось положение об эволюционной роли отбора случайных ненаправленных уклонений. По поводу нелепости такого допущения (именно допущения, поскольку доказательств пока не найдено) в литературе накопилось множество остроумных высказываний. Так, один из известных офтальмологов после того, как он окончательно постиг строение и функции человеческого глаза, высказался в том смысле, что только сумасшедшему могла прийти в голову мысль о возникновении такого прекрасного и совершенного творения в результате стечения случайных обстоятельств.
Совершенно очевидно и легко доказуемо, что изменчивость организмов вовсе не случайна. Существуют определенные спектры допустимых уклонений от нормы, равно как объективно существует запрет на изменчивость, не входящую в эти спектры. Например, могут быть рогатые и безрогие олени, но не может быть рогатых представителей семейства кошачьих; у зла
143
ков не бывает сетчатого жилкования листьев и т.п. Подобных примеров можно привести бесчисленное множество.
Весьма проблематично и положение СТЭ об отборе приспособительных признаков. Не ясно само понятие "приспособительный признак". С.В. Мейен по этому поводу задает вопрос: почему обитающим в одних и тех же условиях насекомым обязательно нужны три пары ног, паукообразным - четыре, многоножкам - много пар, а млекопитающим - две? Какой из этих признаков приспособительный? Почему воробью нужна защитная окраска, а снегирю и синице нет. Почему у растущих в одних и тех же условиях древесных растений разные листья, разный тип ветвления, разные репродуктивные органы? Причем здесь приспособленность? Несоответствие очевидно, однако в СТЭ все это просто замалчивается. Ортодоксальные сторонники этого научного направления ищут только подтверждения своей теории, а все ей не соответствующее стараются просто не замечать.
Действия естественного отбора никто еще не видел, а вопросов по этой проблеме великое множество. Например, если прокариоты столь стары эволюционно и предельно примитивны, а эвкариоты относительно молоды и совершенны, то почему вторые не вытеснили первых? Почему самцы оленей не потеряли в процессе эволюции свои совершенно ненужные им рога? Каким образом возникали признаки или свойства, потребности в которых раньше не было, то есть до своего появления они небыли адаптивны? Живут же все, кроме человека, животные, не умея критически мыслить, и потребности в таком мышлении у них не возникает. Как возникла система нейронных контактов в коре головного мозга человека, определяющая образ мышления - может быть тоже вследствие стечения случайных обстоятельств?
Палеонтология до настоящего времени не нашла ни единого достоверного случая превращения одного вида в другой путем отбора случайных приспособительных уклонений. Во всех учебниках биологии до последнего времени приводился и приводится один и тот же, казалось бы безукоризненный, пример превращения четырехпалой лапы гипотетического предка лошади в копыто (один палец) современной лошади. (По шуточному высказыванию Д.Н. Соболева - "Парадная лошадь эволюции".) Но та же палеонтология недавно уничтожила и этот единственный, аргумент. Выяснилось, что вымершее четырехпалое животное, считавшееся предком лошади, жило одновременно с лошадью и никак не могло быть предком последней.
На самом деле аргументов против возможности эволюции путем отбора случайных мелких уклонений очень много. Но, отрицая концепцию, следует предлагать что-то взамен. И такие предложения были представлены. На некоторых из них мы остановимся.
В 1920 году Н.П. Вавилов на 3-м Всероссийском съезде селекционеров в Саратове выступил с докладом "Закон гомологических рядов в наследственной изменчивости". Сам закон он сформулировал так:
1. Виды и роды, генетически близкие, характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных
144
форм у других видов и родов. Чем ближе генетически расположены в общей системе роды и линнеоны (виды (B.J6.)), тем полнее сходство в рядах их изменчивости.
2. Целые семейства растений в общем характеризуются определенным циклом изменчивости, проходящей через все роды и виды, составляющие семейство.
Иллюстрацией этого закона может быть, например, изменчивость злаков. Если у пшеницы встречаются растения с плотным колосом и рыхлым, остистым и безостым, если цвет колоса может быть серым, красноватым или желтым, то такая же изменчивость должна наблюдаться у представителей близкородственных родов, и она действительно наблюдается, например, у ржи и ячменя.
Н.И. Вавилов не противопоставлял этот закон учению Дарвина, однако на самом деле он убедительно показал, что изменчивость не случайна, а укладывается в строго определенные рамки, что никак не соответствует положению дарвинизма об отборе случайных уклонений.
Двумя годами позже Л.С. Берг в двух публикациях изложил свою тео-рию эволюции на основе закономерностей - теорию номогенеза . Он полагал, что изменчивость не безгранична и далеко не случайна, а пути исторического развития жизни, пути эволюции, канализованы (предопределены объективной закономерностью). Вот его слова: "Никаких случайных вариаций нет - все вариации предопределены и могут быть уложены в гомологические ряды Вавилова ...".
Заслуживает внимания также в основном номогенетический взгляд на процесс эволюции Д.Н. Соболева. Его работа "Начала исторической биогенетики" небольшим тиражом вышла в Симферополе в 1924 году и осталась мало известной. Согласно Соболеву биогенез - историческое развитие жизни на Земле - зиждется на четырех объективных законах: 1 - закон наследственности или сохранения вида; 2 - закон эволюции или органического роста (эволюция как прогрессивное развитие, параллельное развитие, онтогенез и филогенез, филогенетические гетерохронии40 41, переживание и предварение стадий развития, специализация); 3 -закон обратимости эволюции или закон биогенетических циклов (прямое и обратное развитие, обратимые процессы онтогенеза, филогенез и обратное развитие, интеративная42 изменчивость); 4 - закон прерывистости (ряды форм и коллективные типы, отсутствие переходных форм, прерывистая смена фаун (и флор - В.К.), сальтационная43 изменчивость, филогенетические превращения).
В понимании Д.Н. Соболева биологический прогресс, становление новых форм, происходят в результате двух дополняющих друг друга процессов: 1 - длительное неизменное или очень мало меняющееся состояние вида (мед-
40 Номогенез - развитие по закономерности.
41 Гетерохрония - эволюционная разновозрастность признаков одного организма или группы организмов одного вида (в ботанике синоним - гетеробатмия, т. е. разноступенчатость).
42 Интерация - цикличность, повторяемость.
43 Сальтации - внезапные, скачкообразные превращения организмов.
145
(медленное постепенное совершенствование - эволюция) и 2 - критический момент в истории группы организмов (вида или более высокой таксономической категории), при котором наблюдается быстрая деградация или обратное развитие; этот быстрый, революционный процесс сопровождается вымиранием, но он же может приводить к возникновению новых форм путем быстрого (сальтационного) превращения, с которого начинается возрождение в новой форме. Далее - вновь стационарное состояние или медленное совершенствование и новое сальтационное превращение. Циклы повторяются. По словам Соболева: "В результате исследования основ биогенеза мы приходим к воззрению, которое представляет собою синтез линнеевского принципа сохранения вида, ламаркова принципа эволюции, дополненного принципом ее обратимости, ... и принципа прерывистости, установленного Кювье и отчасти Жоффруа (см. раздел Краткая история биологии)".
"Что же касается вопроса: что лежит в основе биогенеза - случай или закон, то на этот вопрос мы вправе с полной определенностью ответить: не случай, но закон. Это значит, что законности, которые мы подметили в ходе биогенеза, не могут быть объяснены простым накоплением или подбором неупорядоченных и лишенных какой-либо тенденции случайно-полезных для вида уклонений, отбором, производимым исключительно лишь борьбою за существование и ведущим к чисто пассивному приспособлению к условиям существования. Процессы органического морфогенеза упорядочены, строго закономерны и при том тенденциозны. ...мы вынуждены признать существование присущего организмам внутреннего закона развития, по которому происходят изменения во времени. Этот закон эволюции не зависит ни от отбора, ни от прямого воздействия среды."
"Отбор не образует новых жизненных форм, он не создает, а уничтожает: отбор не есть фактор биогенеза, но он может служить одним из факторов вымирания".
В дальнейшем номогенетические воззрения в эволюционном учении развивались и другими исследователями. Однако номогенез, даже располагая вескими доказательствами (в их числе и закон гомологических рядов), не может быть безоговорочно принят в качестве все объясняющей теории эволюции - возникает ряд трудно разрешимых вопросов. Например, в чем существо закономерностей, и кто (или что) эти закономерности установил? Кстати, последний вопрос поневоле возвращает нас к креационизму.
Одна из последних, заслуживающих пристального внимания, попытка создать новую теорию эволюции принадлежит нашему современнику и соотечественнику Ю.В. Чайковскому (см. список литературы), ученику и в какой-то мере последователю известного советского палеоботаника С.В. Мейе-на (1935-1987). Ю.В. Чайковский сформулировал триединый принцип эволюции: активность, диатропическая сеть и смена норм. Все три компонента этого принципа довольно сложны и требуют разъяснения. Ниже приводим значительно упрощенное толкование этих факторов эволюции.
Активность, согласно представлениям Чайковского, является базовым понятием (философской категорией), подобной таким понятиям как про
146
странство, время, материя. Базовые понятия не имеют определения и должны восприниматься как нечто объективно существующее, безусловное и неоспоримое. Однако смысл их поддается объяснению. Согласно мнению автора новой теории активностью обладают организмы, части организмов вплоть до микроструктур и группы организмов - стая, фитоценоз44, улей (рой) и тому подобные объединения. Идея собственной активности материи и всего живого в истории развития эволюционной мысли высказывалась давно и неоднократно, например, в трудах Ламарка. Активность выступает в эволюции живого в виде нарастающей самопроизвольности - как поведения (в широком смысле этого слова), так и переработки информации. Иными словами, все живое не зависит от слепой случайности, не пассивно в своем существовании и историческом развитии, а несет в себе собственные внутренние, определенно направленные, тенденции собственной организации (самоорганизации) и прогрессивного развития.
Диатропическая сеть лучше всего иллюстрируется Периодической системой Д.И. Менделеева. В этой системе номер элемента, а также его нахождение в определенной группе и ряду, определяют свойства этого элемента. Химические элементы существуют объективно, а связь между ними отражает периодическая система. Связь тоже объективна, реально существует, просто Менделееву удалось ее обнаружить и представить в виде таблицы. Некоторые клетки этой таблицы при ее создании были пустыми, в дальнейшем они заполнялись. Мы и сейчас продолжаем заполнять таблицу Менделеева новыми трансурановыми элементами.
Примерно то же можно сказать и о представленных в виде таблицы гомологических рядах Вавилова, пустые клетки в них, как известно, тоже заполнялись. Подобных рядов к настоящему времени разными исследователями установлено много. Очевидно, все свойства, признаки и виды организмов укладываются в такие ряды или таблицы, которые и названы диатропической сетью.
Диатропические ряды или сети могут содержать признаки организмов, их свойства, сами организмы, относящиеся к разным таксономическим категориям45, биоценозы46. Ряды в диатропических таблицах указывают основные тенденции - направление эволюционных изменений. Вымирающие виды или утраченные признаки и свойства оставляют в исторически раннем участке ряда диатропической сети пустые клетки, но в ней же, в противоположных наиболее современных, частях рядов появляются новые клетки, заполняемые вновь возникающими или существенно изменившимися видами, свойствами, признаками. Таким образом задается направленность эволюционных изменений.
44 Фитоценоз - сообщество растений, так или иначе связанных между собой.
43 Таксономические категории - понятия, принимаемые в систематике для обозначения соподчиненных групп растений, грибов и животных, отличающихся различной степенью родства. Таксономические категории иерархичны - вид, род, семейство, порядок и т.д.
46 Биоценоз - совокупность животных, растений, грибов и микроорганизмов, совместно населяющих определенный участок суши или водоема.
147
Смена норм происходит путем так называемого генетического поиска. Суть его по Ю.В. Чайковскому в следующем. При утрате номы, в дискомфортных (стрессовых) условиях, большинство особей вида или гибнут, или претерпевают модификационные изменения47. Некоторые изменения могут оказаться в какой то степени адаптивными. Носители таких изменений и ведут генетический поиск. Его механизм состоит в том, что все формы активности организма (поведенческая, физиологическая и генетическая) имеют единую направленность - тенденцию к заполнению следующей клетки в ряду диатропической сети. Один из способов генетического поиска проявляется в увеличении частоты направленных мутаций, при этом направленность мутаций определяется самой диатропической сетью. В случае удачного течения такого процесса возникает новая норма, при которой дискомфортная ситуация перестает быть таковой. Наряду с созданием перспективных направлений эволюции генетический поиск может приводить и к тупиковым вариантам. В палеонтологии достаточно тому подтверждений.
В новой теории эволюции, которую автор вскользь, но весьма удачно называет "новым номогенезом”, придается большое значение еще одному фактору - преадаптации.
Преадаптация это свойство организма (или органа), имеющее приспособительную ценность для ещё не осуществленных форм взаимодействия организма со средой (или для ещё не выполняемой органом функции). Исходя из основных положений СТЭ преадаптация необъяснима и маловероятна (хотя в последнее время ее и в СТЭ перестают отрицать). На самом деле, как объяснить возникновение у организма такого свойства, потребности в котором до сих пор не было? Однако без признания возможности преадаптаций никак не объяснить возникновение в процессе эволюции организмов, обладающих принципиально новыми признаками и свойствами. Например, как возникло половое размножение, как предки теплокровных животных вышли из моря на сушу, как из пресмыкающихся получились птицы?
Для примера остановимся на последнем вопросе. В зоологии принято считать, что предками птиц в геологическом прошлом были пресмыкающиеся (рептилии). Вероятнее всего, у птиц и современных рептилий был общий предок, одни потомки которого в ходе эволюции приобрели крылья и превратились в птиц, другие стали современными рептилиями. Возникает вопрос: каким образом у рептилий появились крылья?
Если следовать положениям СТЭ, то сначала у какой-то группы древних рептилий вследствие случайных ненаправленных изменений (мутаций) постепенно, в течение исторически длительного промежутка времени, вместо нормальных передних конечностей должны были появляться некие промежуточные между передними ногами и крыльями образования, и уже из них впоследствии - крылья. Разумеется, перспективы выживания у подобных гипотетических животных были бы минимальными - они не могли бы летать
47 Модификационные изменения - морфологические или функциональные изменения организмов, вызванные факторами среды и не относящиеся к генотипическим изменениям.
148
(крылья еще не развиты) и уже не могли бы бегать (передние конечности теперь - не совсем ноги). Такой путь превращения бескрылого животного в крылатое просто невозможен. Невозможен он если стоять на позициях СТЭ -полукрылья-полуноги неадаптивны, а подобные уклонения от нормы обречены на элиминацию естественным отбором.
Новый номогенез по Чайковскому не только допускает возможность преадаптации, но и считает это явление реальным и обязательным. Появление нового свойства задается диатропической сетью, в очередной новой клетке которой появляется резко изменившийся или вновь возникший признак (свойство организма). Поскольку такое изменение не случайно, а задано тенденцией, заложенной в диатропической сети, то резкое изменение, о котором идет речь, касается не одного организма, а какого-то множества. В гено-48 ме группы изменившихся организмов резко возрастает частота мутации, направленность которых определяется опять-таки диатропической сетью. В ходе смены сравнительно небольшого числа поколений происходит "при-шлифовка" резко изменившегося или вновь возникшего свойства к организму, при которой меняется и сам организм - иными словами, происходит смена нормы. Эти изменения в масштабе времени течения эволюционных процессов происходят очень быстро. Таким способом, путем преадаптации, превращение рептилии в птицу представляется достаточно вероятным.
В целом можно сказать, что новый номогенез, предложенный Чайковским, лучше, чем синтетическая теория эволюции, объясняет многие проблемы, связанные с пониманием процессов возникновения видов живых существ и их исторического изменения. Однако ставить точку в разрешении всех вопросов теории эволюции еще рано. Новый номогенез, как и ранние теории, не лишен недостатков. К примеру, как объяснить заданную диатропической сетью направленность эволюционных изменений? По-существу, вопросы к номогенезу Берга применимы и к новому номогенезу.
Еще одна сравнительно новая теория эволюции, полностью отрицающая СТЭ, базируется на том, что мутации в историческом изменении видов не играют никакой роли. Движущим фактором эволюции, по мнению В.В. Петрашова - создателя этой теории, является редактирование (направленное изменение) ДНК под влиянием меняющейся среды существования организмов. Во взаимосвязанных понятиях "структура" и "функция" автор признает ведущую роль за функцией, адекватно изменению которой меняется и структура. Структурой в данном случае является ДНК со всеми ее свойствами. Меняющиеся факторы внешней среды неизбежно меняют функции организмов, тканей, клеток. Вследствие неизбежной же обратной связи функции изменяют (редактируют) ДНК, контролирующую эти функции. При таком течении процессов мутациям, случайным уклонениям от нормы, не остается места. Само функционирование организмов (по Чайковскому - "активность") обеспечивает их приспособительное направленное изменение. Можно ска- *
48 Геном - совокупность генов, характерных для гаплоидного набора хромосом данного вида организмов, основной гаплоидный набор хромосом. В отличие от генотипа геном представляет собой характеристику вида, а не отдельной особи.
149
зать, что это и есть эволюция по Ламарку, разумеется, с поправками на современное понимание биологических процессов.
Вероятнее всего, окончательное решение вопросов, связанных с эволюцией организмов на Земле, откладывается на неопределенно долгое время. Наше понимание этого процесса будет углубляться и уточняться по мере развития всех направлений биологической науки.
ПОНЯТИЕ ВИДА В БИОЛОГИИ
Самым убедительным доказательством реальности процесса эволюции является появление новых биогеоценозов и новых видов взамен исчезнувших в геологическом прошлом. Формы проявления жизни на нашей планете меняются, и виды организмов являются ключевыми элементами в меняющихся биогеоценозах.
Что же такое вид? Наука до сих пор не пришла к единому мнению и споры по поводу того, что же такое вид, до сих пор весьма горячи. Рассмотрим, следуя в основном за изложением С.Б. Гончаровой, наиболее общепринятые представления.
Во-первых, вид - это фундаментальная категория в таксономической49 иерархии, т.к. каждый биолог, независимо от его специализации, имеет дело с видами, любая другая таксономическая категория определена и описана на основе вида. Так род - группа видов, подвид - часть вида.
Во-вторых, вид - основная структурная единица в системе живых организмов, качественно обособленная форма живого вещества, основная единица эволюционного процесса. В зависимости от принимаемой концепции, вид определяется как: 1) наименьшая природная популяция, постоянно отделенная от любой другой популяции посредством определенной прерывности в серии {типологический виду, или 2) совокупность популяций особей, способных к скрещиванию с образованием плодовитого потомства, населяющих определенный ареал, обладающих рядом общих морфологических и физиологических признаков и типов взаимоотношений с абиотической и биотической средой, отделенных от других таких же групп особей практически полным отсутствием гибридных форм {биологический вид).
Наиболее известным в российской ботанике является ставшее уже классическим определение вида, данное В.Л. Комаровым, «вид - это морфологическая система, помноженная на географическую определенность».
Одним из удачных, является также определение академика А.Л. Тах-таджяна: вид - "система ..., объединенная общими признаками..., общим происхождением и общим географическим ареалом и достаточно четко отделенная от близких видов, как совокупностью признаков, так и изолирующими барьерами".
Основные концепции вида - номиналистическая, таксономическая (типологическая), биологическая и филогенетическая.
49
Таксономия - раздел систематики, теория и практика классификации организмов.
150
Номиналистическая концепция вида
Один из аспектов проблемы вида - объективная реальность вида как такового. Многие таксономисты полагают, что виды реальны, другие сомневаются в этом. Для того, что бы разобраться в этом вопросе, необходимо разграничить вид-категорию, и вид-таксон. Как категория, вид - созданная человеком абстракция (как любые другие философские категории: материя и т.д.), не относящаяся к конкретному организму. Виды как таксоны - некоторая группа конкретных организмов. Реальны они, или существуют только в человеческом сознании?
Номиналистическая (пдтеп - имя) концепция полагает, что природа производит только индивидуальные организмы и ничего более. Виды по своей сути - производное человеческого мозга, а не объективная реальность. Понятие вида введено как эмблема для обращения к большому количеству организмов одновременно. Виды суть классы, но не организмы. Номиналисты полагают, что индивидуумы относятся к определенным видам на основании сходства.
Таксономическая или типологическая концепция вида
Этимологически термин вид {species) происходит от латинского specere означающий «выглядеть» и относящийся к облику.
Типологическая концепция таксона состоит в том, что каждый класс организмов обладает едиными для всех составляющих его организмов характерными признаками, а члены класса отделены друг от друга индивидуальными отличиями. Концепция "типа", используемая в таксономии, жестко закрепляет имена таксонов за отдельными образцами, признанными типом. Казалось бы, тип должен отражать наиболее типичные черты, обсуждаемого таксона, однако это далеко не всегда так. По правилам Кодекса ботанической номенклатуры - это не обязательно наиболее типичный или характерный элемент таксона. В российской ботанике типологическая концепция вида широко принята. Наиболее известными представителями этой концепции были В.Л. Комаров и Н.И. Вавилов, между последователями которых до сих пор ведутся «бои». Предполагается, что В.Л. Комаров был сторонником таксономической неделимости вида (монотипической концепции), а Н.И. Вавилов считал вид сложной системой внутривидовых форм (политипическая концепция). В монотипической концепции наименьшей неделимой единицей является собственно вид, тогда как в политипической - подвид - экологогеографическая раса. Вид с позиции В.Л. Комарова приравнивается к эколого-географической расе политипического вида.
Биологическая концепция вида
Биологический вид - по наиболее общему из определений - совокупность особей (индивидов), обладающих общими биологическими признаками, способностью к скрещиванию и производству плодовитого потомства, формирующих популяционную систему, которая имеет общий ареал и общую эволюционную судьбу.
Основа биологической концепции вида - критерий нескрещиваемости, в соответствии с которым, вид - это группа особей, репродуктивно изолиро
151
ванная от других подобных групп. Биологическая концепция вида представлена, в свою очередь, двумя концепциями: концепцией одномерного и многомерного видов.
Концепция одномерного вида рассматривает взаимоотношение двух популяций, сосуществующих друг с другом, не смешивающихся в одном и том же месте и в одно и то же время, то есть симпатрические и синхронные популяции. Симпатрические популяции разных видов имеют резкие разрывы, отражающие обособленность видов. То есть, в отличие от типологической концепции, в концепции одномерного вида, виды разделяются на основе морфологических отличий, отражающих репродуктивную изоляцию данного вида от других.
В концепции многомерного вида (или концепции скрещивающихся популяций) вид рассматривается как группа популяций, фактически или потенциально скрещивающихся друг с другом. Данная концепция рассматривает пространственно разобщенные, или аллопатрические, популяции. Разобщенные в пространстве и времени популяции могут значительно отличаться друг от друга, хотя они и не изолированы репродуктивно. В данном случае мы имеем дело с видом, который представлен многими различными аллопатрическими популяциями.
Попытки представить биологическую концепцию как более естественную по сравнению с типологической так же потерпели неудачу. Причиной этому послужили:
1) . Недостаток данных. Очень часто систематик затрудняется дать ответ, является ли его выборка отдельным видом или только пространственно-генетической группировкой внутри изменчивой популяции.
2) . Бесполое размножение. Для видов, размножающихся бесполым путем или парте ногенетически, неприменим критерий скрещиваемости. В данной группе организмов предлагают вернуться к морфологическому виду. Поскольку степень генетического различия ответственна как за репродуктивную изоляцию, так и за морфологические различия, такому подходу дается и некоторое биологическое оправдание.
3) Незавершенность видообразования. Существуют популяции, которые находятся на различных этапах формирования нового вида. Возникают ситуации, которые Э. Майр делит на шесть групп.
1. Эволюционная непрерывность в пространстве и времени. В данном случае существует репродуктивная изоляция географически удаленных популяций, хотя они и связаны непрерывным рядом скрещивающихся популяций. 2. Возникновение репродуктивной изоляции без эквивалентных морфологических изменений. Такие случаи, связанные с образованием так называемых видов-двойников, создают большие трудности практического характера. 3. Возникновение морфологических различий без репродуктивной изоляции. 4. Репродуктивная изоляция, основанная на биотопической разобщенности. 5. Неполнота изолирующих механизмов. Изолирующие механизмы - это биологические свойства индивидуумов, которые препятствуют скрещиванию между действительно или потенциально симпатрическими популяциями. Существует большое число разнообразных изолирующих механизмов, в основе которых лежат особенности строения, поведения и т.д. Изолирующие механизмы создаются постепенно и до определенного уровня обратимы. 6. Достижение разными локальными популяциями различных уровней видообразования.
Эти шесть типов явлений, описанных выше, представляют собой следствие представления о постепенности процесса видообразования. Во многих из описанных случаев установление видового статуса для данной популяции - задача трудная или вовсе невыполнимая.
Филогенетическая концепция вида
Филогенетическая концепция вида основывается на положениях кладистики или филогенетической систематики (учение о происхождении одних видов организма от других). Исторической основой филогенетической сис
152
тематики является нумерическая систематика, классифицирующая организмы на основе матрицы максимально возможного количества признаков. В отличие от таксономической систематики, использующей информативные или "хорошие" признаки, филогенетическая систематика использует все доступные для наблюдения и измерения признаки.
Кладизм - теория, согласно которой организмы получают ранг и классифицируются в соответствии с родственными (сестринскими) отношениями. Современная кладистика пытается определить такие таксоны, которые являлись бы монофилетичными, то есть имеющими общее происхождение, с прослеживающейся привязкой к общему предку и включающими всех потомков этого общего предка.
Категории иные, чем вид
Как уже мы уже отмечали, при принятии политипического вида, вид подразделяется на целый ряд более мелких категорий подвидов - экологогеографических рас. В этом случае наименьшей таксономической единицей будет считаться не вид, а подвид. Вариация - совокупность особей одного вида, характеризующаяся определенными наследуемыми признаками, но не показывающая ясной географической или экологической приуроченности. Формами называют, как правило, группы, имеющие частные морфологические различия (как правило, один признак).
Все категории ранга выше вида являются собирательными таксономическими категориями, предположительно связанными родственными связями (предположительно естественные, монофилитические группы). Так, род состоит из видов, объединенных родственными связями, семейство - из родов ит.д.
Особи, принадлежащие к одному виду, редко существуют в природе в форме одной популяции. Обычно вид представлен несколькими, в той или иной мере разобщенными, небольшими или достаточно крупными популяциями, каждая из которых имеет свой генофонд. Обмен генами между отдельными популяциями внутри вида обычно возможен, хотя бы иногда. Однако, вследствие расширения ареала - района обитания вида - из-за возможных геоморфологических различий в разных участках увеличившегося ареала, отдельные популяции одного вида могут оказаться пространственно, а следовательно и репродуктивно, изолированными.
Пространственная разобщенность популяций одного вида означает что данный вид может в дальнейшем своем эволюционном развитии сталкиваться с разными условиями среды и соответственно претерпевать разные изменения. СТЭ предполагает в этом случае разные направления естественного отбора. Иными словами, две отдельные популяции одного и того же вида могут эволюционировать в различных направлениях, пока не превратятся в два самостоятельных вида, уже не способных скрещиваться между собой. Значит, для того, чтобы произошло образование двух новых видов из одного необходимо возникновение репродуктивной изоляции.
Возможны и другие пути видообразования. Так, периодически возникающие мутации вызывают репродуктивную изоляцию мутировавших осо
153
бей как бы одним большим скачком. Такие явления иногда наблюдаются у растений при образовании полиплоидов. Последние уже не могут скрещиваться с исходными диплоидными растениями. Но если они имеют и мужские, и женские репродуктивные органы, то они сами способны размножаться и дать начало новому виду. Не исключено, что таким путем получились два хороших (биологических) вида березы - Betula pendula (28 хромосом) и В. pubescens (56 хромосом).
Изложенные выше представления о виде и о путях образования видов базируются в основном на положениях синтетической теории эволюции (СТЭ). С позиций нового номогенеза большие популяции не могут быть источниками новых видов, и процесс видообразования в масштабах геологического времени не бывает длительным и постепенным. Новые виды возникают сравнительно быстро, скачкообразно, в условиях стресса в небольших ин-бредных популяциях, в семьях или даже в клонах. При этом изменяются не только отдельные признаки и свойства организмов, но и организмы в целом, и биогеоценозы. В предыдущей главе мы говорили о преадаптациях, о заполнении новых клеток в диатропической сети, сопровождающимся направленным мутагенезом и сравнительно быстрым приспособлением новых свойств к организмам и самих изменившихся организмов к новым условиям существования.
В общем и целом можно сказать, что пути видообразования сложны, разнообразны и изучены далеко не полностью.
СИСТЕМАТИКА РАСТЕНИЙ
Понятия систематика, таксономия, и классификация довольно близки. Термин "систематика" (по отношению к растениям) был впервые введен в употребление Карлом Линнеем (1751). В предисловии к 5 изданию Genera Plantarum (1787) он писал: "Использование некоторой ботанической системы я рекомендую даже начинающим, так как без системы не может быть четкости в ботанике".
Систематика - это наука, которая описывает группы организмов, дает им названия и классифицирует их; иначе говоря, систематика изучает многообразие организмов.
Таксономия - это раздел систематики, посвященный принципам, методам и правилам классификации.
Для изучения систематики очень важны понятия "таксон" и "таксономическая категория". Таксономическая категория обозначает ранг группы (например, вид, род, семейство, порядок и т.д.). Таксон - это конкретная, реально существующая группа определенного ранга, например, вид - лютик ползучий {Ranunculus repens L.), род - лютик {Ranunculus L.), семейство лютиковые {Ranunculaecae Juss) - все это таксоны.
154
Таксон - слово латинское (taxon), во множественном числе - taxa. Каждое растение принадлежит к серии таксонов последовательно соподчиненных рангов (табл. 4).
Таблица 4
Таксономические категории и таксоны с примерами из царства растений.
Уровни иерархии (таксономические категории)
Regnum Subregnum
Divisio (Phyllum) Subdivisio
Classis
Subclassis
Ordo
Subordo
Familia (Fam.) Subfamilia (Subfam.)
Tribus (Tdb.) Subtribus (Subtrib.) Genus (Gen.)
Subgenus (subgen.) Sectio (sect.) Subsectio (subsect.) Series (ser.) Subseries (subser.)
Species (sp.)
Subspecies (subsp.) Vari etas (var.) Forma (f)
Царство
Подцарство
Отдел или Филум
Подотдел
Класс
Подкласс
Порядок Подпорядок Семейство
Подсемейство
Триба или колено
Подтриба
Род
Подрод
Секция
Подсекция Серия ил и ряд
Подсерия
Вид
Подвид
Разновидност ъ
Форма
Окон мания Таксоны (на примере сосны обыкновенной) Растения - Plantae
phyta phytina Г олосеменные - Pinophyta
op si da Хвойные - Pinopsida
idae Хвойные - Pinidae
ales ineae Сосновые - Pinales
aceae Сосновые - Pinaceae
Сосна - Pinus
Сосна - Pinus
Сосна обыкновенная - Pinus sylvestris L.
В приведенной выше таблице жирным шрифтом показаны основные (обязательные) таксономические категории; последние два столбца не везде заполнены - общепринятые окончания в названиях таксономических категорий рекомендованы не для всех уровней иерархии, а необязательные таксономические категории выделяются не всегда.
Различают совершенные и несовершенные таксоны. Совершенные таксоны - это генетически закрытые системы, между которыми в норме невозможен обмен генетическим материалом (роды, семейства, порядки, классы, отделы). В составе совершенного таксона все особи обладают признаками этого таксона. Несовершенные таксоны - это генетически открытые системы, которые в норме могут обмениваться генами, например, близкие
155
подвиды, разновидности и формы. В составе несовершенного таксона не все особи, а лишь большая их часть обладает признаками данного таксона.
Принадлежность особей к тому или иному виду определяется на основании ряда критериев. Критерии вида - это разнообразные таксономические признаки, которые характерны для одного вида, но отсутствуют у других видов. При выделении видов с позиций типологической концепции в первую очередь учитываются морфологические особенности, и выделенные на основании морфологического критерия виды часто называются морфологическими видами, или морфовидами. Подразумевается, что особи, принадлежащие к одному виду, относительно однородны по анатомическому строению, физиологическим особенностям и четко отличаются от особей других видов.
Заметим, что при выделении видов многое зависит от интуиции того или иного ученого. По словам известного американского ботаника А.Д. Кронквиста: «Вид - это то, что систематик считает видом». Объем видов (набор критериев вида) неодинаков в систематике прокариот, низших эвкариот, высших и низших растений, грибов, позвоночных и беспозвоночных животных.
В начале 19 в. ботаника была главным образом описательной наукой. Ученые занимались описанием признаков растений и группированием организмов в соответствии с этими признаками. Это упорядочивание организмов в группы на основе их сходства или различия называется классификация. Продукт классификации - система. Выделяют три основных типа классификаций. Искусственные - в большинстве своем ранние классификационные системы, основанные на нескольких удобных признаках, и удобстве для использования (например, классификация Линнея). Естественные - основанные на комплексах признаков, стремящиеся отразить естественные взаимоотношения между организмами или таксонами. Филогенетические - использующие все известные критерии для интерпретации филогении (эволюции) таксона.
Термин "таксономия" был введен швейцарским ботаником Декандолем (1813) и обозначал теорию классификации растений. Позже термин стал более широко использоваться для методов и принципов классификации любых групп организмов. Например, Ч. Дарвином (1859) термины таксономия и систематика рассматривались как синонимы. И в настоящее время некоторыми ботаниками таксономия и систематика рассматриваются как синонимы, некоторыми различаются. Однако считается, что термин систематика имеет более широкое значение, чем таксономия.
После того как группы организмов были классифицированы, этим группам должны быть даны названия. Выбор названия групп организмов и правила выбора этих имен называется номенклатурой. Номенклатура регламентируется Кодексом ботанической номенклатуры.
Номенклатура - это особенно важный и очень сложный раздел систематики. Международный кодекс ботанической номенклатуры основан на следующих принципах:
156
1. Ботаническая номенклатура независима от зоологической (зоологи имеют свой Международный кодекс зоологической номенклатуры, а микробиологи - Международный кодекс номенклатуры бактерий). Кодекс применяется к названиям таксонов, определяемых как растения (включая грибы), независимо от того, рассматривались ли эти таксоны первоначально как растения или нет.
2. Ботаническая номенклатура предусматривает принцип типификации: применение названий таксонов определяется при помощи номенклатурных типов. Номенклатурный тип вида - это конкретный гербарный экземпляр (или в некоторых случаях изображение), который хранится в определенном гербарии (нахождение гербария в описании таксона указывается) и с которым связывается название данного таксона. Если вид разделяется на два или больше видов, то старое название сохраняется за той его частью, к которой принадлежит этот типовой образец, а другие виды должны получить новые названия (эпитеты). Номенклатурный тип рода - определенный вид: например, для рода дудник (Angelica L.) - вид Angelica sylvestris L., а для рода со-лонечник (Galatella Cass,) - вид Galatellapunctata (Waldst. et Kit.) Nees.
Номенклатурным типом таксонов более высокого ранга до семейства включительно является определенный род, от которого производится название семейства: семейство маковые (Papaveraceae), тип мак (Papaver L.); семейство кладофоровые (Clalophoraceae Wille), тип кладофора (Cladophora Kutzing). По отношению к семействам, правда, сделано два исключения: признаются правильными и законными старые и прочно укоренившиеся для них названия, но разрешается использовать альтернативные названия, произведенные от названия типового рода:
Пальмы - Palmae = (Агесасеае, тип Areca L.);
Злаки - Gramineae (= Роасеае, тип Pda L.);
Крестоцветные - Cruciferae (= Brassicaceae, тип Brassica L.);
Бобовые - Legumindsae (= Fabaceae, тип Faba Miller);
Зонтичные - Umbelliferae (= Apiaceae, тип Apium L );
Губоцветные - Labiatae (= Lamiaceae, тип Lamium L );
Сложноцветные - Compositae (= Asteraceae, тип Aster L.).
К названиям таксонов рангом выше семейства принцип типификации как правило не применяется.
3. Принцип приоритета: номенклатура таксонов основывается на приоритете в обнародовании, т.е. правильным считается название таксона, данное автором первой публикации. При этом время действия принципа приоритета ограничено, и для большинства групп точкой отсчета выбрано 1 мая 1753 г., когда был опубликован важнейший труд К. Линнея «Species plantarum» («Виды растений»). Имя автора, впервые опубликовавшего описание определенного таксона, приводится в конце названия этого таксона, часто в сокращенном виде, например, Plnus sylvestris L. - прописная буква "L" говорит о том, что первое описание сосны обыкновенной было опубликовано Линнеем.
157
CAROLI LINN7EI си M ns Йг.ктд Д1СИ1А1Х1, Medic & Лота я UrSALi ErjrtTLS ле> CT STZiiA Poiui, № non Аса Lt Велсь, Т&ьЗД.
UrsAj. Sr«CLH. Soc 4. Paus. Copijj.
SPECIES PLANTARUM, EX HJBENTES
PLAN TAS KITE COGNITAS,
GESERA RELATAS, CUM
Dm'ERENTIlS SpECiFicrSj TjliVIALTliVij Syxokvmjs Szlfctj^ Locis Natali aib, S L L1) i l! U Ц
SYSTEM.-! S EXHALE, Lit! ESTAS.
Tomus 1 <
<Jf4 £ Я J£tn iror C £ JL V*ii
HOL MI£t LAUKKNTIl Элиты.
Рис. 52. Титульный лист работы Карла Линнея «Виды растений» (Species Plantarum). Опубликована 1 мая 1753 г. Эта книга принята за исходный пункт ботанической номенклатуры.
Вся долиннеевская номенклатура и даже послелин-неевская, но в которой не применяются строго биноминальные названия, не считаются научной и не рассматриваются Международным кодексом ботанической номенклатуры.
4. Принцип уникальности: каждый таксон с определенными морфологическими признаками, положением и рангом может иметь только одно название - наиболее раннее и соответствующее правилам кодекса.
5. Принцип универсальности: научные названия таксонов рассматриваются как латинские (на латинском языке говорили жители Древнего Рима, в наше время этот
язык "мертвый") независимо от их происхождения и подчиняются правилам латинской грамматики. Названия растений на "живых" языках - русском, английском, китайском и др. - не считаются научными, и никаких правил,
регламентирующих их создание и применение, не существует.
(См. Приложение 3)
РЕЛЬЕФ И ПОЧВЫ
Традиционно вопросы геоморфологии и почвоведения не входят в число научных направлений, относящихся к биологии. Почва является компонентом биосферы, относящимся в основном к неживой (косной) материи. Однако лесные биоценозы теснейшим образом связаны с почвой и изучение общих положений почвоведения необходимо для понимания вопросов, рассматриваемых в дальнейшем изложении.
158
Рельеф
Слово рельеф происходит от латинского relevo - поднимаю. Рельеф -это совокупность неровностей суши, дна океанов и морей, разнообразных по очертаниям, размерам, происхождению, возрасту и истории развития. Изучением рельефа занимается наука - геоморфология. Свойства почв в значительной степени определяются рельефом.
Слагается рельеф из положительных, выпуклых, и отрицательных, вогнутых, форм. Рельеф образуется главным образом в результате длительного одновременного воздействия на земную поверхность внутренних и внешних процессов. Рельеф определяет течение почвообразовательных процессов и в конечном счете - формирование тех или иных типов растительности.
Различают 3 группы форм рельефа: макрорельеф, мезорельеф и микрорельеф.
Под макрорельефом понимают самые крупные формы рельефа, определяющие общий облик большой территории: равнины, плато, горные системы. Возникновение макрорельефа связано главным образом с тектоническими явлениями в земной коре.
Мезорельеф - формы рельефа средних размеров: увалы, холмы, лощины, долины, террасы и их элементы, плоские участки, склоны разной крутизны. Возникновение мезорельефа связано в основном с экзогенными геологическими процессами - смыв с возвышенностей и перемещение в низины продуктов выветривания земной коры (денудация), образование континентальных отложений - на которые оказывают большое влияние медленные поднятия и опускания отдельных участков суши.
Под микрорельефом понимают мелкие формы рельефа, занимающие незначительные площади - от нескольких квадратных дециметров до нескольких сотен квадратных метров, - с колебаниями относительных высот в пределах одного метра. Сюда относятся бугорки, понижения, западины, возникающие на ровных поверхностях рельефа из-за просадочных явлений, мерзлотных деформаций или по другим причинам. На склонах микрорельеф иногда определяется сползанием почвенно-грунтовых масс или почвенноэрозионными процессами.
Широко развиты склоновые формы рельефа, которые принято характеризовать по крутизне, формам и экспозиции.
Значение рельефа в формировании почв и развитии почвенного покрова велико и разнообразно. Рельеф выступает как главный фактор перераспределения солнечной радиации и осадков в зависимости от экспозиции и крутизны склонов и оказывает влияние на водный, тепловой, питательный, окислительно-восстановительный и солевой режимы почв.
Так, в горах возникает вертикальная зональность климата, растительности и почв вследствие понижения температуры воздуха с увеличением высоты и изменения в увлажнении. Воздушные массы, приближаясь к горам, медленно поднимаются и постепенно охлаждаются, что способствует выпадению осадков. Перевалив через горы, те же воздушные массы, опускаясь, нагреваются и становятся сухими.
159
Элементы мезо- и микрорельефа, и особенно склоны разной крутизны, прежде всего перераспределяют влагу осадков на земной поверхности и регулируют соотношение вод, стекающих по поверхности, просачивающихся в почву, накапливающихся в понижениях. Поверхности разного наклона и экспозиции получают неодинаковое количество солнечной радиации, что отражается на условиях температурного и водного режимов. Различия в увлажнении вызывают изменения питательного, окислительно-восстановительного и солевого режимов почв.
Все это приводит к поселению и развитию различной растительности, к существенным отличиям в синтезе и разложении органического вещества, превращении почвенных минералов и, в конечном счете, к образованию разных почв в различных условиях рельефа.
В настоящее время выделяют по положению в рельефе и по определяемому им перераспределению осадков следующие группы почв, которые называются рядами увлажнения.
Автоморфные почвы - формируются на ровных поверхностях и склонах в условиях свободного стока поверхностных вод, при глубоком залегании грунтовых вод (глубже 6 м).
Полугидроморфные почвы - формируются при кратковременном застое поверхностных вод или при залегании грунтовых вод на глубине 3-6 м; капиллярная кайма может достигать корней растений.
Гидроморфные почвы формируются в условиях длительного поверхностного застоя вод или при залегании грунтовых вод на глубине менее 3 м; капиллярная кайма может достигать поверхности почвы.
Рельеф оказывает большое влияние на развитие эрозионных процессов. В условиях склоновых форм рельефа возможно проявление водной эрозии, то есть смыва и размыва почвы. Равнинные формы в районах с засушливым и континентальным климатом благоприятствуют возникновению ветровой эрозии.
Рельеф выступает и как фактор эволюции растительности и почв при его изменении. Например, при постепенном врезе русла реки пойменная терраса превращается в надпойменную. Это приводит к изменению режима увлажнения - прекращаются пойменные и аллювиальные процессы, понижаются грунтовые воды и, как следствие, к развитию почв не в гидроморфных или полугидроморфных условиях, а в автоморфных.
На территории нашей страны, в Восточной и Северо-Восточной Сибири равнинная тайга сменяется в горах сначала горными северо-таежными редколесьями, а затем горной тундрой. Иначе выражена высотная зональность растительности в горах южной части СНГ. При достаточной высоте гор здесь развиты горные степи, горные широколиственные и хвойные леса, субальпийские и альпийские луга. Зона горных лесов полнее всего выражена в горах юго-запада (Карпаты, Кавказ) и юго-востока (Приморье), т. е, в районах повышенного увлажнения. Зона горных степей наиболее характерна для гор Средней Азии, отличающихся сухим континентальным климатом.
160
Отмеченные особенности влияния рельефа на почвообразование имеют большое значение и в лесоводстве, так как геоморфологическое разнообразие определяет и характер лесной растительности, и характер всех хозяйственных мероприятий, которые не должны нарушать целостности почвенного горизонта и не должны снижать его плодородия.
Почвы
Наука о почвах называется почвоведением. Уже в ранних, натурфилософских, научных работах подчеркивалось значение почвы как источника питательных веществ для растений. Почву изучали также с геологической точки зрения, и при этом внимание было сосредоточено на физических и химических свойствах почвы; ее описывали как инертный неорганический материал. Русский ученый В.В. Докучаев впервые стал рассматривать почву как динамическую, а не инертную среду. Он сделал это в своей классической работе о почвах России, опубликованной в 1870 году. В.В. Докучаев доказал, что почва постоянно изменяется и развивается, а в ее активной зоне идут физические, химические и биологические процессы. Он выявил пять главных почвообразующих факторов, к которым относятся климат, геологическая основа (материнская порода), топография (рельеф), живые организмы и время.
Почву часто относят к абиотическим составляющим биосферы, хотя правильнее считать ее важнейшим связующим звеном между биотическими и абиотическими компонентами наземных экосистем.
В состав почвы входят четыре важных структурных компонента: минеральная основа (обычно 50-60 % общего состава почвы), органическое вещество (до 10 %), воздух (15-25 %) и вода (25-35 %)
Минеральный скелет почвы - это неорганический компонент, который образуется из материнской породы в результате ее выветривания.
Структура почвы
Минеральные фрагменты, образующие вещество почвенного скелета, различны - от валунов и камней до песчаных крупинок и мельчайших частиц глины. Скелетный материал обычно произвольно разделяют на мелкий грунт (частицы меньше 2 мм) и более крупные фрагменты. Частицы меньше 1 мкм в диаметре называют коллоидными. Механические и химические свойства почвы в основном определяются теми веществами, которые относятся к мелкому грунту. Соотношение частиц разных размеров в мелком грунте исследуют путем механического анализа в лаборатории или при наличии опыта на ощупь в поле. Глиной считают материал с частицами меньше 0,002 мм (2 мкм) в диаметре, более крупные - песок.
Структура почвы определяется относительным содержанием в ней песка, ила и глины. Значение почвенной структуры можно отчасти уяснить, сравнивая свойства чистого песка и глины. Идеальная почва должна содержать приблизительно равные количества глины и песка в сочетании с части
161
цами промежуточных размеров. В этом случае образуется пористая, крупитчатая структура, и почвы называются суглинками. Они обычно обладают достоинствами двух крайних типов почв и не имеют их недостатков. Таким образом, легко выполнимый механический анализ почвы дает полезное представление об ее вероятных свойствах.
С сельскохозяйственной точки зрения почвы иногда относят к тяжелым - глины или легким - пески.
Химизм почв
Химизм почв определяется минеральным скелетом и органическим веществом. Большая часть минеральных компонентов представлена в почве кристаллическими структурами. Песок и алеврит состоят в основном из очень инертного минерала кварца SiO2, именуемого также кремнеземом. Кремнезем, соединяясь ионами алюминия и железа, образует электронейтраль ные кристаллы силикаты. Силикаты - преобладающие почвенные минералы.
Большую роль в удержании почвой воды и питательных веществ играет особенно многочисленная и важная группа - глинистые минералы. Большинство их встречается в виде мельчайших, плоских кристаллов, которые образуют в воде коллоидную суспензию. Важная особенность этих минералов - наличие у них постоянного отрицательного заряда, который нейтрализуется катионами, адсорбированными из почвенного раствора. Благодаря этому катионы не выщелачиваются из почвы и могут обмениваться на другие катионы из почвенного раствора и растительных тканей. Эта катионообменная способность служит одним из важных индикаторов плодородия почвы Свободные пространства между глинистыми частицами втягивают также воду, что ведет к гидратации и набуханию глины
Органическое вещество
Органическое вещество почвы образуется при разложении мертвых организмов, их частей (например, опавших листьев), экскретов и фекалий. Мертвый органический материал используется в пищу совместно детритофа-гами, которые его поедают и таким образом способствуют его разрушению, и редуцентами - грибами и бактериями, завершающими процесс разложения. Не полностью разложившиеся органические остатки называются подстилкой, а конечный продукт разложения - аморфное вещество, в котором уже невозможно распознать первоначальный материал, получило название гумуса. Цвет гумуса варьирует от темно-бурого до черного. В химическом плане это очень сложная смесь изменчивого состава, образованная органическими молекулами различных типов. В основном гумус состоит из фенольных соединений, карбоновых кислот и сложных эфиров жирных кислот. Гумус, подобно глине, находится в коллоидном состоянии; отдельные частицы его прочно прилипают к глине и образуют глино-гумусовый комплекс. Так же как и гли
162
на, гумус обладает большой поверхностью частиц и высокой катионообменной способностью. Эта способность особенно важна для почв с низким содержанием глины. Анионы в гумусе - несут электрический заряд в карбоксильных и фенольных группах. Благодаря своим химическим и физическим свойствам гумус улучшает структуру почвы и ее аэрацию, а также повышает способность удерживать воду и питательные вещества.
Одновременно с процессом гумификации (образования гумуса) жизненно важные элементы переходят из органических соединений в неорганические и становятся доступными для растений. Этот процесс называется минерализацией.
Для того чтобы формировался гумус того или иного типа, необходим достаточный дренаж почвы: в условиях переувлажнения разложение идет очень медленно, так как нехватка кислорода ограничивает рост аэробных редуцентов. В таких условиях растительные и животные остатки сохраняют свою структуру и, постепенно спрессовываясь, образуют торф, который может накапливаться вплоть до больших глубин.
Содержание воздуха и воды
Почвенный воздух, так же как и почвенная вода, находится в порах между частицами почвы. Порозность - объем пор возрастает в ряду от глин к суглинкам и пескам. Между почвой и атмосферой происходит свободный газообмен.
Общее количество воды, которое может быть удержано почвой (его определяют, добавляя избыток воды и затем ожидая, пока она не перестанет выходить каплями), называется полевой влагоемкостью.
Вода необходима всем почвенным организмам. Она поступает в живые клетки путем осмоса. Вода также важна как растворитель для питательных веществ и дыхательных газов, поглощаемых из водного раствора корнями растений. Она принимает участие в процессах разрушения материнской породы, подстилающей почву.
Образование почвы
Почва образуется в результате взаимодействия многих факторов, из которых пять наиболее важные это климат, материнская порода (геологические условия), топография (рельеф, или морфология поверхности), живые организмы и время. Рассмотрим каждый из этих факторов в отдельности.
Климат и выветривание
Физическое и химическое разрушение материнской породы под влиянием климата и в меньшей степени живых организмов называется выветриванием. Наиболее важные факторы, вызывающие выветривание, - это вода и
163
температурные колебания, поэтому роль климата удобнее рассматривать на примере именно этих факторов.
Физическое выветривание может происходить в результате изменений температуры. Горные породы попеременно расширяются и сжимаются при ее суточных колебаниях. Этот процесс ведет к постепенному разрушению горных пород. Если в трещинах имеется вода и если она периодически замерзает и оттаивает, то при образовании льда может создаваться огромное давление, также разрушающее породу. Химическое выветривание также происходит при участии биологических и климатических факторов Оно обычно ускоряется с повышением температуры. Вода действует как растворитель и как реагент в процессе гидролиза некоторых минералов. Породы, в некоторой степени, растворимые в воде, - мел и известняк, состоящие в основном из карбоната кальция, особенно подвержены эрозии. Лишайники относятся к тем немногим организмам, которые могут расти на обнаженной скальной поверхности, они тоже способствуют химической эрозии, извлекая из выветриваемых скал определенные питательные вещества.
Температурный режим и осадки - дожди и снег являются двумя главными климатическими факторами, вызывающими выветривание.
Материнская порода
Первичное происхождение всех горных пород связано с застыванием расплавленной магмы, поднимающейся в верхние зоны земной коры из глубоких недр. Породы, образовавшиеся непосредственно из застывшей магмы, называются вулканическими. Они могут быть кислыми - как гранит или основными - как базальт в зависимости от их минерального состава. При выветривании кислых пород большая часть кремния остается в виде кварца SiO2, представленного зернами песка. При выветривании основных пород значительная часть кремния связывается в различные силикаты, которые в дальнейшем формируют алевриты и глины.
Осадочные породы образуются в результате отложения продуктов выветривания других горных пород или накопления остатков живых организмов. Этот материал накапливается и уплотняется иногда в течение миллионов лет. К типичным осадочным породам относятся песчаники, мел и известняк.
Топография
Рельеф местности, влияя на такие климатические факторы, как выпадение осадков и температура, тем самым косвенно сказывается и на процессе почвообразования. Экспозиция и высота склонов в значительной мере определяют климат. На солнечных склонах температура выше, поэтому почвы развиваются быстрее; здесь они, как правило, более сухие, что нередко влияет на растительность. Наветренные склоны обычно более влажные, чем подветренные.
164
Почвообитающие организмы
При непосредственном участии живых организмов образуются органические компоненты почвы - подстилка и гумус. Растительность, так же как климат и материнская порода, влияет на развитие почв. Можно полагать, например, что и вероятность образования подзолов под степью и сообществами хвойных лесов выше, чем под сообществами лиственных лесов. Это, по-видимому, связано со свойствами веществ, вымываемых из растительной подстилки, и их ингибирующим влиянием на процессы разложения органического материала почвенными микроорганизмами. Кислая подстилка степей и сосновых лесов в отличие от подстилки лиственных лесов неблагоприятна для дождевых червей.
Деятельность почвенных детритофагов, особенно дождевых червей в зонах умеренного климата, важна во многих отношениях и в свою очередь зависит от растительности. Детритофаги ускоряют процесс разложения, размельчая частицы подстилки и таким образом увеличивая их поверхность. За счет фекалий и отходов метаболизма детритофагов увеличивается масса перерабатываемого органического и минерального вещества. Почва, проходя через пищеварительный тракт дождевого червя, измельчается, ее минеральные и органические компоненты перемешиваются, и это улучшает структуру почвы. Ходы дождевых червей улучшают аэрацию и облегчают рост корней. Точно так же способствуют перемешиванию и аэрации почвы ходы, прорытые позвоночными животными - кротами, кроликами и т. и.
Влияние человека
Хрупкое динамическое равновесие зрелой почвы может быть нарушено при использовании ее человеком для сельскохозяйственных целей, а также в процессе рубки и трелевки леса. При этом часто происходит разрушение почвенной структуры, истощение запасов питательных веществ и эрозия почв.
Структура почвы особенно легко разрушается, когда ее обрабатывают тяжелыми сельскохозяйственными или лесохозяйственными машинами. Уплотнение почвы приводит к ряду последствий; в частности, уменьшаются размеры почвенных пор, и в результате ухудшаются аэрация и дренаж. Почва может уплотняться также и при неумеренном выпасе домашнего скота. Повторное использование почвы, особенно под одни и те же культуры, может понизить содержание в ней важных питательных веществ. Распашка почвы нередко усиливает ветровую и водную эрозию, особенно на склонах. Точно так же уничтожение защитного слоя растительности при чрезмерном выпасе скота или при вырубке леса способствует эрозии. Таким образом, рациональное использование почвы является одной из ключевых проблем для почвоведов, земледельцев, лесоводов, лесозаготовителей.
165
Время
Время имеет важное, хотя и косвенное, значение для образования почвы. Скорость этого процесса в умеренном климате очень различна - может потребоваться от нескольких десятилетий (при формировании почвы на вулканическом пепле) до нескольких тысяч лет (на обнаженной поверхности скальных пород). В масштабах геологического времени происходят климатические и топографические изменения, которые, естественно, влияют на развитие почвы.
Таким образом, необходимо вновь подчеркнуть, что рассмотренные почвообразующие факторы действуют неизолированно. Каждый из них, исключая время, непосредственно влияет на образование почвы, но может оказывать также косвенное воздействие, влияя на другие факторы.
Типы почв
Как мы видели, почвы представляют собой продукт взаимодействия между биотическими и абиотическими компонентами экосистемы. Это необычайно сложные и разнообразные системы, поэтому их очень трудно классифицировать.
Почвы изучаются путем рытья шурфов или с использованием почвенных буров. При описании вертикального разреза для характеристики почвенных слоев или горизонтов применяются стандартные обозначения.
Горизонт А - элювиальный горизонт, то есть горизонт, из которого вымываются минеральные вещества.
Ао - находится над горизонтом А и называется подстилкой.
Горизонт В - иллювиальный горизонт, то есть горизонт, в котором накапливаются питательные вещества.
Горизонт С - материнская порода, подвергающаяся выветриванию.
Горизонт D - материнская порода, или коренная подстилающая порода.
Различия в строении почвенного профиля - основа классификации почв. Каждый из почвенных типов, подтипов, родов и видов характеризуется определенным строением почвенного профиля.
Лесные почвы
Лесные почвы представляют собой полнопрофильные естественные образования, формирование которых тесно связано с лесной растительностью, в первую очередь с лесными древесными растениями. Специфичность лесных почв проявляется прежде всего в значительной пятнистости почвенного покрова, обусловленной воздействием древесной растительности - видовой принадлежностью деревьев, характером их расположения в лесном пологе, неравномерной сомкнутостью.
Лиственные породы обогащают почву мягким гумусом, азотом, зольными элементами, способствуют нейтрализации почвенной кислотности, что создает благоприятные условия для развития почвенных микроорганизмов,
166
кишечнополостных, членистоногих и других представителей почвенной биоты.
Длительное произрастание хвойных лесных формаций неблагоприятно влияет на свойства почв, способствует накоплению плохо разлагающейся подстилки. Хвойные породы являются подзолообразователями. Под влиянием лиственных пород сформировались серые и бурые лесные почвы.
Подзолы. Это название происходит от слова «зола». Подзолы обычно образуются на песчаниках, отложениях гравия ледникового происхождения и аллювиальных отложениях и характерны в основном для пустошей и хвойных лесов в областях с прохладным влажным климатом. Подстилка относительно мощная, так как ее материал - хвоя и т. и. - отличается высоким содержанием фенолов, устойчивых к микробному разложению. Кальция в таких почвах мало, поэтому они слишком кислые для того, чтобы в них могло обитать много дождевых червей и других животных, заглатывающих почву. В связи с этим главную роль в разложении подстилки играют грибы, и разложение идет относительно медленно, в результате образуется толстый слой кислого гумуса, который резко отделен от нижележащих слоев.
Гумусовый горизонт подзолов по цвету напоминает серую золу и ясно выделяется среди соседних слоев. Он состоит в основном из крупинок песка Это сильно выщелоченный горизонт, содержание питательных веществ в нем невелико из-за очень большого количества осадков, вымывающих питательные вещества, или высокой водопроницаемости подстилающей материнской породы (или по обеим этим причинам).
Выщелоченный горизонт, в котором расположена большая часть корней, - кислый, содержит мало нитратов и других питательных веществ, необходимых растениям. Подзолы, таким образом, относительно неплодородны. Большая часть питательных веществ в хвойных лесах сохраняется в биомассе деревьев и подстилки, а не в почве.
Серые лесные почвы. Этот тип почв сформировался в условиях континентального умеренно влажного климата под лиственными и, реже, смешанными лесами. Эти почвы обладают относительно хорошими физическими свойствами, биологически активны, плодородны. Процесс подзолообразования в них протекает менее интенсивно, чем в подзолистых почвах. Серые лесные почвы распространены в лесостепной зоне, встречаются также в лесах Кавказа, Алтая, Саян.
Бурые лесные почвы (буроземы) - типы почв, образовавшиеся в условиях умеренно теплого и влажного климата под широколиственными, реже хвойными, лесами на различных материнских породах. Характеризуются бурой окраской, комковатой или ореховатой структурой, быстрым разложением опада, значительным запасом гумуса, высокой емкостью поглощения, слабокислой реакцией, плодородием. Они распространены на Дальнем Востоке, Северном Кавказе, встречаются на Урале, в Саянах. Подразделяются на теплые северокавказские и холодные дальневосточные.
167
ПРИРОДНЫЕ ЗОНЫ РОССИИ
Почвы и растительность природных зон
Разнообразие биосферы в целом обусловлено преимущественно широтной зональностью, она выражается в закономерной смене типов растительности от арктических пустынь на севере до влажных субтропических лесов на юге. Еще большее разнообразие дает высотная поясность. В горных районах широтная зональность растительного покрова усилена высотной зональностью, которая, как и высотная зональность почв, зависит не только от высоты гор, но и от целого ряда других факторов.
Каждой географической зоне присущ свой особый тип высотной зональности - закономерное сочетание высотных поясов, последовательно сменяющихся от подножий гор к вершинам. Границы природных зон в европейской России располагаются с запада на северо-восток. Такое расположение природных зон, не строго параллельно экватору, а под некоторым углом вызвано теплым течением Гольфстрим. В случае изменения направленности этого течения расположение природных зон может измениться.
С севера на юг на равнинах нашей страны сменяются следующие типы почв:
- арктические почвы;
- тундровые арктические почвы: тундровые глеевые почвы; типичные тундровые глеевые почвы; оподзоленные тундровые глеевые почвы;
- подзолистые почвы;
- серые лесные почвы;
- бурые лесные почвы;
- черноземные почвы: оподзоленные выщелоченные черноземы; типичные черноземы; луговые черноземы;
- каштановые почвы: темно-каштановые почвы; светло-каштановые почвы;
- серые почвы.
В высоких широтах Арктики, развиты арктические почвы. Почвы слабо выщелочены, имеют высокое содержание железа, иногда засолены. Южнее арктические почвы сменяются тундровыми', сначала тундровыми арктическими, затем тундровыми глеевыми (типичными и оподзоленными). Тундровые глеевые почвы отличаются небольшой мощностью, малым содержанием гумуса, повышенной кислотностью и оглеенностью50. Сельскохозяйственному освоению этих почв препятствуют их низкая температура, бедность питательными веществами, слабая микробиологическая активность. Внесение органических и минеральных удобрений, осушение повышают плодородие тундровых почв. Осушенные почвы лучше прогреваются, вечная мерзлота под ними летом залегает глубже, чем под заболоченными почвами. Подзоли
30 Оглеение -процесс восстановления окисных почвенных соединений, главным образом железа, в закисные, придающие почвам особые свойства, снижающие плодородие.
168
стые почвы представляют самый распространенный почвенный тип. Вместе с горно-подзолистыми и болотными почвами они занимают свыше половины территории России. На крайнем западе (Калининградская область) и юго-востоке (Среднее Приамурье и Южное Приморье) в зоне подзолистых почв появляются буроземы {бурые лесные) почвы, формирующиеся под влажными широколиственными и отчасти смешанными лесами. Серые лесные почвы распространены на стыке подзолистых почв с черноземами. Они образуются под лиственными лесами лесостепи на лёссовидных грунтах.
К югу от серых лесных почв широкой мозаичной полосой залегают черноземы (местами они перемежаются с другими типами почв). Восточнее Алтая черноземы встречаются отдельными островами до Восточного Забайкалья включительно. Черноземы богаты гумусом и зольными веществами, имеют значительную мощность, обладают прочной зернистой структурой и поэтому отличаются высоким плодородием. Черноземы - почвы открытых травянистых степей, где растительного материала для образования гумуса вполне достаточно, а процессы выноса ослаблены. Свойства черноземов существенно меняются при движении с севера на юг. Северную окраину черноземной зоны образуют оподзоленные выщелоченные черноземы. В центре зоны распространены типичные черноземы - самый плодородный подтип черноземных почв. При неглубоком залегании грунтовых вод (до 3-5 м) в степях формируются лугово-черноземные почвы. В сухих степях и полупустынях зональный почвенный покров образован каштановыми почвами. Они формируются в условиях отрицательного баланса влаги и разреженного злакового и полынно-злакового травостоя. Различают темно-каштановые, собственно каштановые и светло-каштановые почвы. Наиболее плодородны темнокаштановые почвы, граничащие на севере с черноземами. Южнее зоны каштановых почв, располагаются бурые почвы. Особым объектом природопользования являются так называемые азональные почвы, т.е. почвы не характерные для данной зоны. Они образуются в местах, где условия локально отличаются от зональных. Это болота, поймы, территории с близким залеганием засоленного горизонта и др.
Растительность различных природных зон
Большие размеры территории России определяют сравнительно большое богатство ее флоры - около 90-100 тыс. видов растений. Это основа, «фундамент», для разнообразных биологических ресурсов. Флористическая насыщенность территории возрастает при движении с севера на юг (в пустынях несколько снижается из-за сухости климата). Разнообразие флоры снижается при движении с равнинных территорий к горным.
По различным зонам растительность распределяется следующим обра
зом.
169
Растительность Арктических пустынь
В Арктических пустынях, распространенных на крайнем севере Арктики, наблюдается разорванный растительный покров - растения покрывают менее 50 % поверхности почвы. Растительный покров состоит из мхов, лишайников, водорослей и немногих видов цветковых растений.
Растительность Тундры
К югу от арктических пустынь, преимущественно на материковом побережье Северного Ледовитого океана, распространен тундровый тип растительности. В его покрове ведущая роль принадлежит мхам, лишайникам, кустарничкам и отчасти кустарникам. Древесная растительность часто представлена карликовыми и стелющимися жизненными формами.
Растительность Лесотундры
В пограничной полосе между тундровым и лесным типами растительности размещается лесотундра. В лесотундре встречаются одновременно лесные и тундровые растительные группировки. Леса здесь своеобразные. Это светлые редколесья и редины, в которых низкие деревья с разреженными кронами отстоят одно от другого на большом расстоянии.
Растительность Тайги
Зона тайги расположена к югу от лесотундры. В растительном покрове преобладают хвойные леса, произрастающие на подзолистых и мерзлотнотаежных почвах. В этой зоне большие площади заняты сфагновыми болотами. Это самая крупная по площади ландшафтная зона России.
Растительность Зоны смешанных лесов
В этой зоне распределение хвойных и широколиственных пород подчинено определенной закономерности. Леса с заметным участием широколиственных пород встречаются на суглинистых, хорошо дренированных почвах, чаще всего по южным склонам и вершинам невысоких возвышенностей. В менее благоприятных условиях (избыточная влажность почв, северные экспозиции) доля широколиственных снижается или они отсутствуют.
Растительность Зоны широколиственных лесов
Зону широколиственных лесов условно можно выделить только в пределах европейской части России. Это прерывистая узкая полоса, вытянутая в широтном направлении от западной границы страны до южного Урала, на границе между зоной смешанных лесов и лесостепью. Эдификатором лесов этой зоны является дуб. Сопутствующие породы - липа, клен, вяз, ясень, береза. Разнообразные по видовому составу подчиненные яруса лесной растительности. Почвы в основном с мощным гумусовым горизонтом, глинистые -выщелоченные черноземы, серые лесные почвы. В период вегетации наблюдается некоторый дефицит влаги.
170
За Уралом на юге Западной Сибири широколиственные леса представлены небольшими разрозненными участками, эдификатором в которых является липа. Дальше на восток широколиственные леса встречаются в южной части Дальнего Востока.
Растительность Зоны лесостепи
В лесостепи особый тип растительности, точно определяемый самим названием зоны. Здесь широколиственные леса соседствуют с разнотравными степями. Доля лесопокрытой площади небольшая. Леса представлены изолированными степью островными массивами, часто занимающими сравнительно небольшие площади. Это самая освоенная в земледельческом отношении зона.
Растительность Степной зоны
Степная зона в европейской части России простирается от лесостепи до Черного и Азовского морей, в Западной и Восточной Сибири - степи в основном расположены вдоль южных границ государства. Зональные различия выражаются в виде двух подзон: северных степей на черноземах и южных степей на темно-каштановых почвах.
В степи преобладает травянистый тип растительности ксерофильного характера, занимающий значительные пространства в умеренном поясе северного полушарии (аналог - прерии, пампа). Эта зона отличается континентальным климатом, незначительным количеством атмосферных осадков (250-450 мм) и неустойчивым их режимом, наличием засух и др. Почвы в основном представлены черноземами и каштановыми почвами. Это безлесные сообщества многолетних ксерофитных трав, представленных главным образом злаковыми ассоциациями. Лесные группировки в степях встречаются лишь по долинам крупных рек, балок, на песках надпойменных террас (сосновые боры). Доминирующей жизненной формой растений являются многолетние травы (63% от всей флоры). Широко распространены узколистные дерновинные злаки (ковыль, типчак, мятлик, овсяница и др.). На территории России по видовому составу наиболее богаты северные степи (до 80 видов на 1 м2), со значительным участием различных видов разнотравья. Для южных степей характерно господство злаков, меньшая видовая насыщенность и разреженный травяной покров.
Влияние на растительность провинциальности, или долготной зональности
Наряду с широтной зональностью растительности, обусловленной сменой климата при движении с севера на юг, существует и долготная зональность или провинциальность. Она вызвана различиями в условиях увлажнения на окраинах и внутри материка. Провинциальность внутри растительных (и ландшафтных) зон раскрывается через систему провинций, выделяемых в рамках пояса или всего материка через сектора. На территории России пред
171
ставляется целесообразным различать три основных сектора: субатлантический, внутриматериковый, тихоокеанский.
Растительность Субатлантического сектора
В субатлантическом секторе от Баренцева моря и на восток до Урала, хорошее увлажнение под влиянием Атлантического океана. Здесь широко распространены смешанные (хвойно-мелколиственные и хвойношироколиственные) и широколиственные леса. Тайга здесь темнохвойная с господством ели обыкновенной. Еловой (по составу редколесий) является и восточноевропейская лесотундра. Значительные площади заняты дубовой лесостепью.
Растительность Внутриматерикового сектора
Во внутриматериковом секторе с его континентальным климатом все меньше смешанных (хвойно-широколиственных) и широколиственных лесов. В лесотундре и тайге господствующая роль принадлежит лиственнице сибирской, а в Восточной Сибири (к востоку от Енисея) лиственнице даурской.
Дубовая лесостепь, свойственная атлантическому сектору, сменяется в Западной Сибири на березовую и березово-сосново-лиственничную лесостепь в Средней Сибири. Вместо перистых ковылей и типчака, распространенных в степях субатлантического сектора, в степных островах Средней и Южной Сибири преобладающее значение получают тырсовидные51 ковыли. Характерными типами растительности внутриматерикового сектора становятся полупустыни и пустыни, занимающие юг Казахстана и равнины Средней Азии.
Растительность Тихоокеанского сектора
Относительно неширокая полоса тихоокеанского сектора - аналог субатлантического. В этом секторе, как и на западе страны, появляются смешанные (хвойно-широколиственные) и широколиственные леса. Тайга вновь становится темнохвойной, с участием аянской ели и белокорой пихты. Очень характерны для этого сектора заросли кедрового стланика, горные березняки из каменной березы, а также поражающие пышным ростом высокотравья Камчатки и Курильских островов.
Описание природных зон России
В описании зон (рис. 53) некоторые сведения будут повторяться, это будет сделано для удобства чтения и дополнительного закрепления материала. Подробнее изложены сведения, относящиеся к Европейской части России.
31 Тырса - то же, что ковыль волосатик (Stipa capillata).
172
Зона тундры
К зоне тундры относятся арктические острова: Южный остров Новой Земли, Вайгач, Колгуев и побережье Баренцева моря. На материке южная граница тундры проходит около 67° с. ш., а северная - на Новой Земле, севернее 73° с. ш. Такое географическое положение определяет небольшую величину суммарной солнечной радиации. Летом во время непрерывного полярного дня солнечная радиация достигает более или менее значимых размеров, но подавляющая часть ее идет не на обогрев почвы и воздуха, а на испарение влаги, поэтому температура воздуха в тундре низкая даже летом. Суровость лета в тундре усиливается влиянием Баренцева и Белого морей, которые поглощают много тепла, расходуемого на таяние льдов и нагрев воды. Зато зимой моря подогревают тундру, препятствуя резкому охлаждению воздуха. Тундра принадлежит к числу молодых ландшафтов. Здесь долго держались материковые льды, и следы их пребывания повсюду хорошо выражены в рельефе. В ландшафтном отношении зона тундры распадается на три подзоны.
Арктическая тундра. Она занимает Южный остров Новой Земли, остров Вайгач и побережье Байдарацкой губы. Средняя температура июля 5-6 °C. Растительность очень бедная, над разорванным мохово-лишайниковым покровом лишь кое-где приподнимаются низкорослые травы и кустарнички. Арктические тундры сильно заболочены.
Типичная (мохово-лишайниковая) тундра. Она характерна для восточной части материкового побережья Баренцева моря. Средняя температура июля 6-9 °C
Южная (кустарниковая) тундра. Она примыкает к лесотундре. Средняя температура воздуха июля 9-11 °C. Широко распространены в южной тундре заросли из полярной березки, кустарники.
Итак, тундра - ландшафтная зона субарктического пояса, характеризующаяся избыточным увлажнением при недостатке тепла, безлесьем, широким развитием мохово-лишайникового и кустарничкового покрова. По сравнению с ледяной зоной в тундре теплее, однако средняя температура июля на юге зоны не поднимается выше 11 °C, а по-настоящему теплых дней со средней суточной температурой выше 15 °C почти не бывает. Вследствие низкого испарения тундра даже при небольшом количестве осадков избыточно увлажнена, речной сток большой и реки в летнее время многоводны. Почвы тундр маломощные, кислые, бедные гумусом.
173
ние.)
Рис. 53. Природные зоны России (Рисунок на всю страницу, альбомное расположе-
174
Зона тайги
Тайга расположена к югу от лесотундры. Европейская тайга на юго-западе она граничит с зоной смешанных лесов, на юго-востоке - с зоной лесостепи. Южная граница ее проходит по Чудскому озеру, через Псков, Новгород, Иваново, Нижний Новгород, Казань, по правобережью Камы и Белой к Уральским горам.
Зона тайги - ландшафтная зона умеренного пояса с прохладным, сравнительно влажным климатом. В растительном покрове преобладают хвойные леса на подзолистых и мерзлотно-таежных почвах и сфагновые болота. Это самая крупная по площади ландшафтная зона России. Лето в тайге теплее, а зима холоднее, чем в лесотундре и тундре. Средняя температура июля поднимается от 13° на севере зоны до 19 °C на юге. Еловые темнохвойные леса расположены в западной и восточной окраинах зоны, которые обладают умеренно континентальным климатом. Внутриматериковые азиатские части зоны с их резко континентальным климатом покрыты лиственничными лесами. В тайге много мелколиственных лесов из березы и осины. Подобно лесотундре тайга сильно заболочена.
Главнейшее богатство тайги - лес. Тайга Русской равнины имеет три особенности, которые отличают ее от других таежных районов России. Первая особенность - умеренно континентальный влажный климат с активной циклонической деятельностью. Осадков выпадает 500-600 мм в год, примерно в 2 раза больше, чем в тайге Средней Сибири. Вторая особенность тайги Русской равнины - преобладание темных еловых лесов. И в этом отношении она представляет собой полную противоположность светлой лиственничной тайге Восточной Сибири. Третья особенность - умеренная заболоченность. Это отличает ее как от бедной болотами восточносибирской тайги, так и от сильно заболоченной, лишенной естественного дренирования западносибирской тайги. В тайге сравнительно богатый животный мир.
Типичные представители животного мира этой зоны - бурый медведь, лось, рысь, росомаха, белка, бурундук, белка-летяга, заяц-беляк, куница, горностай, красная и красносерая полевки. Есть пресмыкающиеся - гадюка, уж, живородящая ящерица. Из птиц - глухарь, рябчик, клесты, различные виды куликов, различные виды дятлов. В тайгу по полям и вырубкам проникли южные виды животных: еж, заяц-русак, черный хорь, полевка обыкновенная, мыши лесная и полевая, тетерев.
В тайге Русской равнины, при движении с севера на юг, наблюдаются зональные изменения ландшафта, позволяющие различать в ней три ландшафтные подзоны: северную, типичную и южную тайгу.
Подзона северной тайги
Она лежит между зоной лесотундры и 64-й параллелью. Лето короткое и холодное (средняя температура июля от 13 до 16 °C). Характерны высокий процент заболоченности территории, низкорослость и разреженность лесов. В ельниках обычна примесь березы, в травяном и кустарниковом покрове встречаются болотно-тундровые виды. Зональные почвы - глеево
175
подзолистые и подзолистые иллювиально-гумусные. Распространены не менее широко и болотно-подзолистые почвы.
Подзона типичной (средней) тайги
Средняя температура июля достигает 18°С.. Тайга этой подзоны характеризуется сомкнутым древостоем, отсутствием березы в качестве постоянной примеси к ели, господством ельников-черничников в группе зеленомошных ельников.
На юге типичной тайги появляются первые представители широколиственных пород. В подзоне оптимальные условия для развития подзолообразовательного процесса. Ведущее значение приобретают подзолистые почвы.
Подзона южной тайги
Средняя температура июля в этой подзоне 18-19 °C. Хвойные леса находят в ней наиболее благоприятные условия для произрастания. Вместо ельников-черничников распространенной ассоциацией становятся ельники-кисличники. Появляется новый тип хвойного леса - сложные (кустарниковые) ельники и сосняки, содержащие в кустарниковом ярусе липу, вяз, лещину, и другие виды свойственные широколиственному лесу.
Деятельность человека привела к сокращению площади лесов в зоне тайги, однако и до сих пор она выделяется среди других ландшафтных зон высоким процентом лесистости от 50 до 80%. Остальная ее часть занята болотами и пашней. Кислые подзолистые почвы тайги в своем естественном состоянии мало плодородны.
Зона смешанных лесов
На северо-востоке смешанные леса граничат с тайгой по лини Чудское озеро - Псков - Новгород - Иваново - Нижний Новгород. На юго-востоке они сменяются лесостепью по линии Луцк - Житомир - Киев - Калуга - Рязань - Нижний Новгород. Распределение лесов с преобладанием широколиственных или хвойных пород подчинено определенной закономерности. Широколиственные породы предпочитают суглинистые, хорошо дренированные почвы, чаще всего по южным склонам и вершинам невысоких возвышенностей. Другие земли занимают леса с преобладанием хвойных пород. Большое разнообразие в ландшафт смешанных лесов вносят неоднородные геологогеоморфологические условия.
Зона смешанных лесов Русской равнины
Эта ландшафтная зона умеренного пояса отличается сравнительно мягким, влажным климатом, присутствием на водоразделах смешанных (темно-хвойно-широколиственных) лесов с дерново-подзолистыми почвами. На ландшафтах зоны сказывается ее западное, приатлантическое положение. Она больше, чем тайга, получает тепла и влаги. Суммы температур воздуха выше 10° равны 1800-2400°С.. Зимой здесь обычно не бывает ни сильных морозов, ни глубокого снежного покрова. Средняя температура января на западе зоны выше -5°С., а на востоке около - 12°С. Частые оттепели зимой мешают накоплению снега. Поэтому юго-запад зоны по длительности залегания снежного покрова (менее 100 дней) и его высоте (ниже 30 см) напоми
176
нает степи и полупустыни Заволжья. Западные черты климата выражаются и в обилии атмосферных осадков. В большей части зоны годовое количество осадков превышает 600 мм. Дерново-подзолистые почвы на западе зоны обладают уже некоторыми признаками, сближающими их с бурыми лесными почвами Западной Европы. В зоне смешанных лесов можно встретить следы древнего ледника. Вертикальная дифференциация ландшафтов в зоне смешанных лесов выражена намного отчетливее, чем в зоне тайги. Это обусловлено большими колебаниями относительных высот. В зоне смешанных лесов выделить несколько ландшафтных провинций.
1. Приморская провинция смешанных лесов. К этой провинции относятся побережья Балтийского моря и прилегающая к нему низменная равнина. В западных районах растут граб, тис, плющ, на крайнем западе Калининградской области - бук.
2. Провинция смешанных лесов Белорусско-Валдайского поозерья. Климат провинции влажный, продолжительность залегания снежного покрова около 150 дней. Распространены еловые, широколиственно-еловые леса, сосновые боры.
3. Среднерусская провинция смешанных лесов. В эту провинцию включаются северная лесная часть Среднерусской возвышенности. Климат Среднерусской провинции смешанных лесов более континентальный. Естественный растительный покров представлен елово-широколиственными лесами и дубравам.
4. Провинция смешанных лесов Мещеры и другие.
Зона широколиственных лесов
В зону широколиственных лесов Европейской части России, примыкающую с юга к зоне смешанных лесов, более или менее значительными частями входят следующие административные подразделения: Брянская, Курская, Орловская, Тульская, Московская (южная часть), Рязанская, Нижегородская области, республики Мордовия, Чувашия, Татарстан, Башкортостан и южная часть Челябинской области. Вся эта значительная по площади территория лишь по принятому лесорастительному районированию относится а зоне широколиственных лесов. Фактически же крупных лесных массивов на этой территории мало. Большая ее часть уже давно занята сельскохозяйственными угодьями и прочими нелесопокрытыми площадями. Из значительных, но многократно пройденных рубками, широколиственных лесных массивов можно выделить лишь Тульские засеки.
Согласно М.Г. Романовскому (см. список литературы) северн7ая граница зоны определяется исчезновением естественных ельников. Господствуют серые лесные почвы. Запасы гумуса в однометровой толще лесных почв составляют (80) 120-200 т на га. Преобладают лессовидные моренные суглинки. Песчаные и легкие супесчаные почвы связаны в основном со склоновыми и пойменными лесами и занимают 14 % общей площади зоны.
Зональной растительной формацией становятся теневые широколиственные леса: липняки с участием дуба черешчатого, клена остролистного, вя
177
за гладкого, а в среднерусской провинции - ясеня обыкновенного. Эдифика-тор лесных сообществ липа мелколистная. Во влажных и сырых экотопах единично присутствует ель. На юго-западе Брянской области встречается граб. По мере продвижения к юго-восточной границе зоны нарастает участие дуба, которому липа в сухих экотопах уступает господствующее положение. Сосняки занимают 16 % лесных площадей, концентрируясь почти исключительно на легких почвах. К востоку зоны усиливается континентальность климата и засушливость летнего периода. Из экологических рядов с атмосферным водным питанием (дожди, туманы) выпадают сырые и мокрые типы леса (почвенное водное питание), в том числе верховые болота, составляющие одну из экологических ниш сосняков.
Южная граница зоны определяется падением лесистости ландшафта ниже 20 % и господством на нелесных территориях степных черноземов.
Широколиственные леса с доминированием липы мелколистной встречаются и за Уралом, на юге Западной Сибири. В основном это также сравнительно небольшие лесные массивы.
В Восточной Сибири широколиственные леса отсутствуют. Появляются они вновь лишь на юге Дальнего Востока, частично занимая низкогорья Сихотэ-Алиня, Хора, Хехцира, и поймы горных рек.
Степная зона России
Степная зона России простирается от лесостепи до Черного и Азовского морей. На юго-востоке она граничит с зоной полупустынь. Эта граница проходит по северо-восточному побережью Цимлянского водохранилища, выходит севернее Волгограда, отсюда направляется по долине Волги к Саратову и, не дойдя до него, резко поворачивает на восток, к Уральску. Восточнее реки Урал она несколько отклоняется на юго-восток, проходит через верховья реки Илек (южнее Актюбинска). Во флоре и фауне степей Русской равнины много западных видов, родина которых находится западнее этого места. Например, среди растений - украинский ковыль и костер береговой, среди животных, обыкновенный слепыш и крапчатый суслик. В пойменных лесах зоны до долины Урала встречается дуб совместно со своими широколиственными спутниками. Здесь следует отметить такие известные дубравы как Шипов лес и Теллермановский лес с насаждениями Хоперского заповедника.
Другая ландшафтная особенность южнорусских степей - наличие зарослей степных кустарников, так называемых дерезняков. Степная вишня, терн, бобовник, дереза, этим южнорусские степи отличаются от западносибирских, которые почти лишены дерезняков. Как и в лесостепи, распределение почв, растительности и животного мира степей находится в тесной взаимосвязи с рельефом местности. В ландшафте степной зоны хорошо прослеживаются также зональные и провинциальные различия. Выражением зональных различий служит существование двух подзон: северных степей на черноземах и южных степей на темно-каштановых почвах.
178
Степная зона умеренного пояса отличается сухим континентальным климатом и безлесьем водоразделов. Здесь преобладают травянистая, преимущественно злаковая растительность, растущая на южных и отчасти обыкновенных черноземах и темно каштановых почвах. Ландшафт степной зоны это единый географический комплекс. Важную роль в формировании степного географического комплекса играет сухой континентальный климат. Лето более солнечное и жаркое, чем в лесостепи (средняя температура июля от 22 до 23,5 °C, суммы средних суточных температур выше 10° составляют 2800-3400 °C на западе зоны, 2000-2400 °C на востоке). Атмосферных осадков выпадает меньше, поэтому баланс влаги резко отрицательный.
Степная зона Русской равнины обладает огромными земельными; богатствами, используемыми преимущественно под пашни. Как и в лесостепи, на западе зоны пашня занимает 70-80 % территории. В степях возделываются лучшие сорта пшеницы, кукурузы, проса, подсолнечника, бахчевых культур; западнее Волги много садов и виноградников. Земледелие в степной зоне сочетается с высокоразвитым животноводством.
В связи с высокой распаханностью территории, легкоразмываемых почвообразующих пород (лёссовидные грунты), ливневого характера летних осадков и бурного снеготаяния весной велика опасность развития эрозии и смыва почв.
Антропогенные воздействия (распашка земель, чрезмерный выпас скота и пр.) привели к значительной деградации степных ландшафтов. Поэтому в последние годы проводятся широкие исследования по экологической оптимизации степных ландшафтов. Оптимизация ландшафтов - комплекс (система) мероприятий, направленных на достижение максимально возможной продуктивности ландшафта при сохранении экологического равновесия и его красоты для человека.
Итак, природные зоны России разнообразны и обширны, однако антропогенная деятельность в каждой из них требует большой осторожности. Методы рационального природопользования в каждой зоне должна быть сугубо индивидуальны. Нельзя механически переносить приемы использования природных ресурсов из одной зоны в другую это обычно приводит к негативным последствиям.
Некоторые особенности северной лесной зоны
Независимо от рассмотренного выше районирования в лесоведении и геоботанике часто выделяется северная лесная зона, включающая бореальные52 и суббореальные леса европейской и азиатской частей России. Эта зона объединяется многими весьма существенными общими чертами и включает большую часть лесного фонда нашей страны. Такой подход к районированию также заслуживает внимания и должен быть рассмотренным отдельно.
Северная лесная зона характеризуется в общем суровыми зимами и относительно теплым летом. В различных частях зоны все же наблюдаются большие климатические различия, причем по мере движения к востоку зима делается холоднее, а лето - теплее и амплитуда колебаний летней и зимней температур становится все значительней.
Количество осадков, в среднем, от 300 до 600 мм в год, причем оно ниже на севере и востоке и выше к югу и на западе. На северо-востоке Сибири общее количество осадков опускается до 150 мм и менее, то есть столько же, сколько и в пустыне. Максимум осадков всюду приходится на лето.
52
Бореальная лесная зона = северная лесная сонна.
179
Ветры в западной части лесной зоны более влажные и летом преобладают западные, зимой - юго-западные. В восточной же части ветры более сухие, зимою на востоке Якутии преобладают северные и северо-западные ветры.
В общем, можно сказать, что по мере продвижения с запада на восток климат становится все более континентальным, достигая крайнего своего выражения в Якутии, причем Енисей является границей, где климат значительно меняется.
Указанные большие климатические различия восточной и западной тайги сильно отражаются на составе растительности. Восточная тайга с ее суровыми, длинными, сухими зимами характеризуется сбрасывающей на зиму хвою лиственницей, тогда как для западной тайги характерны ель и другие вечнозеленые хвойные.
В отношении рельефа лесная зона также резко делится на две части: восточную - за Енисей и западную - к западу от Енисея. Последняя представляет собой в общем равнинную страну, лишь Уралом прорезываемую с севера на юг, тогда как большая часть восточной части характеризуется сильно выраженным рельефом, где простираются местами мощные хребты, которые, подобно Уралу, своими вершинами поднимаются выше предела лесной растительности.
Характерною для почв значительной части лесной зоны Сибири является так называемая вечная мерзлота, то есть наличие не успевающих за лето оттаять слоев почвы или грунта. Широко распространенное мнение, что вечная мерзлота свойственна лишь тундре, неверно. Леса всей северо-восточной части Сибири произрастают на вечной мерзлоте, тогда как в тундре европейской части России не везде имеется постоянная мерзлота. Южная граница вечной мерзлоты приблизительно проходит от горла Белого моря к городу Березову на р. Оби, отсюда - к устью Подкаменной (Средней) Тунгуски на Енисее; далее она направляется на юго-восток, пересекает оз. Байкал и через южное Забайкалье уходит в Монгольские степи.
Географически по составу древесных пород наши леса могут быть расчленены на три весьма неравномерные части:
леса таежной или лесной зоны, простирающиеся почти непрерывной полосой от западной нашей границы до Охотского моря;
леса Северного Кавказа;
леса Дальнего Востока (главным образом к югу от Амура).
Хотя на долю таежной зоны России приходится более 90 % всей покрытой лесом площади нашей страны, леса Северного Кавказа и Дальнего Востока по составу древесных пород неизмеримо богаче таежной зоны. В то время как на Кавказе и Дальнем Востоке насчитывается несколько десятков ценных хвойных и лиственных пород, во всей таежной полосе, если выбросить из нее юго-западную часть и сравнительно узкую южную полосу в европейской части России, на протяжении почти 9000 км простираются леса, в главной своей массе состоящие лишь из 7 пород - ель, сосна, лиственница, пихта, кедр, береза и осина, с явным преобладанием первых трех.
180
Леса европейской и азиатской таежных зон не разграничены резко между собой. Граница между Азией и Европой не является границей, делящей леса на естественные области. В европейской части более резко выделяется полоса дубово-широколиственных лесов, которая занимает самую южную часть таежной зоны. В состав лесов этой полосы (округа) входят: дуб, липа, клен, ясень, вяз, груша и яблоня, реже сосна. Господствует в большинстве случаев дуб. Затем несколько обособляется западная часть европейской части России, где отсутствуют сибирские хвойные породы. Этот округ отделяется линией, идущей, примерно, от Онеги к нижнему течению реки Вятки.
Остальная территория европейской части таежной зоны, будучи, в сущности, хвойной, по составу пород уже менее отличается в отдельных своих частях. По мере движения с запада на восток состав пород вообще несколько разнообразится, и если в Карелии и Мурманской области из хвойных встречается только сосна и ель, то далее на восток последовательно присоединяются лиственница, пихта и кедр. Однако лишь в Предуралье их участие в составе лесов становится более или менее значительным, а в остальной части они не играют сколько-нибудь заметной роли.
Большое значение имеет изменение качества лесов с юга на север, так как в этом направлении их бонитет постепенно падает.
Хотя Уральский хребет и не представляет собой территории, отделяющейся по составу пород от равнинных пространств, простирающихся от него к западу и востоку, однако, отличаясь разнообразием геологического строения и определяя наличие вертикальной зональности, он вызывает в лесном покрове значительное разнообразие типов леса и классов бонитета.
Все пространство к востоку от Уральского хребта и до Охотского моря характеризуется участием в лесах все тех же пород, что и на Урале, но только чем далее к востоку, тем заметнее увеличивается участие лиственницы, сначала сибирской, а за Енисеем и Байкалом - даурской, которая в Якутии становится господствующей породой.
Леса севера Сибири до сих пор недостаточно изучены, но все же можно сказать, что в них, наряду с указанным изменением с запада на восток, выражено изменение с юга на север, также связанное с падением класса бонитета. Несколько обособляются алтайские и саяно-прибайкальские леса, в состав которых хотя и входят те же породы, но резко выраженный здесь, как и на Урале, рельеф приводит к большому разнообразию типов леса и к падению бонитетов по вертикали.
Для правильной оценки огромных лесных ресурсов северной лесной зоны и планомерной организации их освоения необходимо принять во внимание следующее.
1. На значительной территории ныне еще не освоенных лесных пространств таежной зоны леса часто занимают лишь узкие полосы вдоль рек и речек, а все междуречные пространства покрыты или чистыми болотами, или заболоченными лесами весьма низких производительности и качества. Это особенно относится к Западносибирской низменности.
181
Узкие лесные полосы вдоль рек или острова леса среди болот осложняют эксплуатацию таких лесных массивов.
2. Чрезвычайно важным обстоятельством для правильной оценки лесных запасов и их качества является колоссальное развитие гарей на северо-востоке европейской части страны и в Сибири. Под гарями в Сибири находится, вероятно, около 30-40 % покрытой лесом площади, если не больше.
3. Наконец, надо учесть, что даже покрытые лесом площади, во-первых, часто имеют невысокие бонитеты вследствие заболоченности леса, а во-вторых, на этих площадях сильно выражена фаутность53 леса, одной из причин которой являются опять таки непомерно распространенные в Сибири лесные пожары.
Учитывая неоднородность наших лесов, и не касаясь полосы дубовых лесов, на южной окраине лесной зоны европейской части России, всю собственно лесную зону можно разделить на следующие округа:
1) округ хвойно-широколиственных смешанных лесов европейского типа, который, начинаясь от западных границ бывшего СССР южнее Санкт-Петербурга, постепенно выклинивается к востоку и Южному Уралу. Здесь характерна значительная примесь широколиственных пород (дуб, клен, липа и др.) к хвойным (ель и сосна);
2) округ западно-европейских хвойных лесов. Это леса расположенные к северу от предыдущего округа, на западе - до границы с тундрой, на востоке - до бассейна р. Сев. Двины. Он охватывает весь северо-запад европейской части страны;
3) округ хвойных лесов с примесью сибирских пород - лиственницы, пихты, а ближе к Уралу и сосны кедровой. Он простирается до Урала включительно;
4) округ западно-сибирских хвойных лесов, охватывающий всю Западно-сибирскую низменность до р. Енисея;
5) округ восточно-сибирских хвойных лесов, простирающийся от Енисея почти до Охотского моря. Характерными чертами его можно назвать значительное уменьшение участия в составе лесов ели, пихты, кедра и, господство лиственницы, сначала сибирской, далее даурской и сосны;
6) округ хвойных лесов охотского типа, где значительное участие принимает аянская ель;
7) округ лесов алтайско-саянского типа, где большое участие в лесах горного характера имеют пихта, ель, сосна кедровая, сибирская лиственница и сосна;
8) округ дальневосточных смешанных лесов уссурийского типа, занимающий нижнюю часть Приамурья и Приморский край, где к разнообразным хвойным - аянская ель, корейский кедр, несколько пихт - примешивается очень богатая флора широколиственных пород. Леса этого округа будут рассмотрены особо.
53
Фаутность - зараженность патогенными организмами.
182
Рассмотрим биологические особенности основных для этих округов лесообразущих пород и характерные черты образуемых ими лесов.
При описании наших древесных и кустарниковых пород остановимся лишь на тех, которые имеют большое лесохозяйственное и лесопромышленное значение. Начнем с группы так называемых темнохвойных лесов, куда входят еловые, пихтовые и кедровые леса.
ТЕМНОХВОЙНЫЕ ЛЕСА
Основные лесообразующие породы
Основными лесообразующими породами северной лесной зоны, исключая центральную и северную части Восточной Сибири, где господствует лиственница, вне сомнения, являются хвойные древесные растения с преобладанием темнохвойных. Темнохвойными называют виды ели, пихты и пятихвойных (кедровых) сосен в отличие от светлохвойных - сосны обыкновенной и видов лиственницы. Выражение "темнохвойные", вероятно, вошло в лесоводственную терминологию в связи с двумя обстоятельствами. Первое -хвоя темнохвойных действительно несколько темнее, чем у лиственницы и сосны обыкновенной, и второе - темнохвойные породы образуют густые тенистые насаждения, под пологом которых сохраняется их собственный подрост, а светлохвойные породы возобновляться не могут.
Понятие "темнохвойные леса" не исключает частичного присутствия светлохвойных и лиственных пород в виде примеси и даже чистых насаждений. Однако последние в зоне темнохвойных лесов обычно представлены временными формациями и составляют сравнительно небольшую долю от общей лесопокрытой площади.
Светлохвойные породы и образуемые ими леса будут рассмотрены отдельно.
Род ель Ptcea Dietr.
Род ель объединяет виды, к которым относятся крупные деревья с удлиненными побегами и мутовчатым ветвлением. Имеются также межмутовочные побеги, они покрыты многолетней, спирально расположенной игловидной хвоей, которая крепится на "листовых подушечках"
Ели однодомны. Опыляются ветром. Шишки созревают в тот же год, свисают с ветвей и после вылета семян остаются некоторое время на деревьях. Кроющие чешуи незаметны, Семена крылаты; при отделении от крылышка оставляют в нем ложкообразное углубление. Всходы имеют 6-9 семядолей с зубчиками по краям. Зубчики имеются и по краям первичной хвои.
Смоляные ходы встречаются в коре и древесине. Ядровая часть древесины не окрашена.
183
Представители рода ель произрастали в Европе в меловом периоде. Более ясные остатки ели обыкновенной обнаружены в плиоцене, ели сибирской - в межледниковых слоях. Во время ледникового периода ель встречалась по всей Северной Европе. Изменившиеся условия привели к обособлению европейского вида ели, ареал которой оказался разорванным по линии течения рр. Вислы, Буга и Дуная. В горах Южной и Средней Европы обособилась ель европейская. В Сибири и на северо-востоке Европы осталась ель сибирская, которая в ледниковый период заходила значительно далее на Запад, вплоть до Франции. По горным цепям Кавказа и Азии обособился ряд елей с ограниченными ареалами - ель восточная, ель тяньшанская, на Дальнем Востоке - ель аянская.
Род ель объединяет до 50 горных видов. Из них лишь немногие переходят на равнины. В странах СНГ естественно произрастают 10 видов. В северных условиях ель встречается вместе с пихтой.
Ель европейская Ptcea abies (L.) Karst.
В природных условиях ель европейская может достигать размеров крупного дерева - до 30 (50) м высоты. Ствол ели в поперечном сечении округлый, прямой, полнодревесный, боковые сучья всегда тонкие, но очень крепкие. Ствол, выросший на свободе, усажен ветвями почти до основания, а в насаждении он очищается редко более чем на одну треть своей высоты.
Крона густая, широкопирамидальная. Ветвление мутовчатое, но имеются и межмутовочные побеги и сучья. На опушках и у одиночно стоящих деревьев кроны спускаются до земли. Рост в высоту у ели европейской продолжается до глубокой старости, поэтому вершины ее всегда заострены.
Кора у ели европейской в раннем возрасте гладкая, красновато-коричневая, трещиноватость слабо выражена. Позднее начинают появляться дугообразные вертикальные трещинки. К 40 годам у основания стволов обособляются чешуйки, сначала мелкие, затем все грубее и толще, с блюдцеобразными впадинами, под которыми виднеются буровато-коричневые слои. К 100 годам толстые чешуйки выделяются уже по всей нижней части стволов, и кора становится ясно шероховатой. Для деревьев старого возраста, замедливших рост, характерно окрашивание чешуй в мутнозеленоватый или красноватый тон, что объясняется развитием водорослей, грибов и бактерий. В особенно неблагоприятных условиях ветви старых елей покрываются листоватыми или свисающими «бородатыми» лишайниками, придающими им своеобразный вид. Заостренные буроватые почки ели европейской не содержат смолы и у сильных побегов прикрыты снизу скрученными хвоинками. Деревья этого вида имеют только удлиненные побеги, вначале зеленые, с нежной хвоей. Хвоя сидит на выступах - подушечках, остающихся и после ее опадения.
Различают световую и теневую хвою. Световая хвоя встречается в верхней части кроны и торчит во все стороны. Она жесткая, колючая, че
184
тырехгранная, в разрезе ромбическая, с белыми полосками устьиц на всех сторонах, длиной 1,3-2,5 см, заостренный конец ее прозрачный. Теневая хвоя сидит неясно двухрядно в затененных местах. Она относительно плоская, менее жесткая и более зеленая.
В самых затененных местах хвоя сидит в два ряда. На таких побегах разрез ее плоскоромбический и хвоинки ориентированы горизонтально. Хвоя у ели держится до 6-7 лет, а на вершине - до 10 лет, но в городских условиях и при задымлении опадает раньше.
В центре хвои проходит сосудисто-волокнистый пучок, у краев -два смоляных хода.
Опыление у ели обыкновенной происходит в мае. Пыльцы высыпается много. Разлетается она хорошо, так как имеет воздушные мешки. Ко времени пыления на концах основных побегов появляются пурпурно-красные или изумрудно-зеленые женские стробилы; свисающие нижние побеги покрываются множеством красноватых собраний мужских стробилов. Такое расположение способствует перекрестному опылению.
Женские стробилы состоят из кроющих и семенных чешуй. Семена созревают примерно через 4-6 месяцев после опыления - к осени. Шишки с созревшими семенами остаются на деревьях зимой. Спелые шишки имеют удлиненно-цилиндрическую форму. Длина их достигает 10-15 см, ширина 3-4 см. Вначале они зеленые или фиолетовые, затем бурые, лоснящиеся. Чешуи деревянисто-кожистые, выпуклые, широкие, обратно-яйцевидной формы, по краю выемчатые или выгрызенно-зубчатые, иногда усеченные. Кроющих чешуй не видно, после опыления они не увеличиваются, (рис. 54).
Давать семена ель начинает с 25-30 лет, а на свободе - и с 15. «Пылит» на 1-2 недели ранее сосны. Ель одно домна и ветроопыляема. Семена из шишек высыпаются в конце января и до марта - апреля. Семена яйцевидной формы кофейно-бурого цвета, с легким красноватым оттенком, матовые. Кончик семени вытянут в длинное и отогнутое острие. Крыловидный придаток (крылышко) семени языковидной формы, с закругленным концом. Семя легко отделяется, оставшееся после него место - углубление на придатке -напоминает ложку. Семенные годы ели обычно наступают через 4-5 лет в более мягком климате, на севере же и в горах семенные годы наблюдаются реже. В промежуточные годы у ели часто совершенно не бывает семян, в годы же семенные плодоношение бывает очень обильным. Всхожесть семян 70-75 %, сохраняется до 9-10 лет, но удовлетворительной считается в течение 3-4 лет. Весной после посева всходы появляются через 3-5 недель.
В год посева всходы поднимаются до 4-5 см. В первые 10-15 лет, ель растет медленно, достигая к 10 годам 1-2 м. Далее дает годичные побеги до 70 и более сантиметров длины. Если в первые годы ель растет медленнее, чем сосна, то к 30-60 годам на лучших (среднесуглинистых, достаточно гумусированных) почвах она обгоняет сосну. Кульминационный рост на лучших почвах у нее около 35 лет, на худших - около 65.
Корневая система у сеянцев слабо развита, значительно слабее, чем у сосны. Первоначально главный корень ясно заметен. Обычно на 3-м году
185
жизни стержневой корень перестает развиваться, и в 10-15 лет совершенно не заметен. Сильно же развивающиеся боковые корни далеко распространяются в почве и обуславливают поверхностную корневую систему даже у старых елей. В силу этого она легко выворачивается ветром с корнями, нередко сильно страдая от ветровала.
Рис. 54. 1 - ель европейская: 1а - мужские стробилы на побеге; 16, 1в - микроспорофилл; 1г - женский стробил; 1д, 1е - мегастробил с кроющей чешуей; 1ж - раскрывшаяся и нераскрытая зрелые шишки; 1з, 1и - кроющая и семенные чешуи зрелой шишки; 1к - семена, крыловидный семенной придаток после отделения семени напоминает "ложечку"; 1л - верхняя часть побега с чешуйчатыми почками, листовыми подушечками и хвоинкой; 1м - поперечный разрез хвои. 2 - ель сибирская: 2а - зрелая шишка; 26 - семенная и кроющая чешуи зрелой шишки; с.ч - семенная чешуя; к.ч - кроющая чешуя; сз -семязачатки; л.и - листовые подушечки; лж - "ложечка" (из Зуихина, Коровин, 2006)
186
Ель не отличается особенно большой долговечностью; обычно считается, что 250-300 лет составляют ее предельный возраст. Для Западной Европы указывались случаи достижения елью 50 м высоты при 2 м толщины. У нас она достигает таких размеров (до 47 м) в самых лучших условиях произрастания. По мере продвижения на северо-восток, высота роста дерева уменьшается и деревья достигают все менее значительных высот. В средней части европейской части страны ель имеет высоту в пределах 30-35 м.
Анатомические особенности древесины. Поперечный срез. Ядро не выражено. Граница между годичными слоями (кольцами) отчетливая; переход от ранней древесины к поздней постепенный. Очертания ранних трахеид имеют почти правильную прямоугольную форму со значительно большей стороной в радиальном направлении. Поздние трахеиды квадратные или прямоугольные. Вертикальные смоляные ходы (одиночные или парные) выстланы толстостенными эпителиальными клетками. Смоляных ходов меньше, чем в древесине сосны. Осевая древесинная паренхима отсутствует. Тангенциальный срез - лучи в основном 1-рядные; средняя высота (слойность) лучей 10-15 клеток; радиальные смоляные ходы в двурядных лучах выстланы толстостенными эпителиальными клетками. Радиальный срез - поры на стенках трахеид расположены преимущественно в 1, редко 2 ряда. Краевые клетки лучей представляют собой лучевые трахеиды с гладкими стенками. Паренхимные клетки лучей с толстыми и тоже гладкими стенками. Поры на полях перекреста в ранней древесине пицеоидные (см. приложение 2).
Древесина ели желтовато-розовато-белая. Ядра нет. Древесина мягкая, легкая, блестящая, легко колется. Она легче сосновой, но уступает последней в прочности. Содержит много мелких вросших сучков. Находит широкое применение во всех тех случаях, где не требуется особенная ее прочность. В настоящее время идет в массе на целлюлозно-бумажное производство. Так называемая резонансная древесина ели используется для изготовления музыкальных инструментов.
Кора ели представляет собой хороший дубитель, так как содержит много танинов (7-12 %), из коры можно получать смолу.
Ель европейская имеет обширную область распространения, простирающуюся от Пиренеев до северо-востока европейской части России (рис. 55). В пределах нашей страны северная граница ели проходит через южную часть Кольского полуострова, не доходя, до самого северного предела древесной растительности. Южная граница ели в европейской части России довольно близко совпадает с северной границей чернозема. Распространение ели от юга Европы почти до Ледовитого океана показывает, что ее требования к климату имеют широкую амплитуду. Более чувствительна ель к высокой температуре и особенно к сухости воздуха. В сухие годы отмирают даже взрослые деревья, особенно на легко просыхающей почве. Ель принадлежит к породам, нуждающимся в значительной влажности воздуха.
Поверхностная, в общем слабая, корневая система ели обуславливает ее значительную требовательность к почвенной влаге. Она нуждается в достаточной и притом равномерной влаге в почве. Но в то же время она не выносит чрезмерного избытка влаги, особенно застаивающейся воды. В отношении минерального богатства почвы ель хотя и не отличается высокой требовательностью, но превосходит сосну, уступая, однако, пихте и лиственни
187
це. В европейской части нашей страны она образует хорошие древостои на свежей суглинистой почве, произрастая, однако, как на тяжелых глинах, так и на супесях и даже песках, если последние достаточно влажны.
Рис. 55. Распространение ели европейской, ели сибирской и ели восточной в России: 1 - ель сибирская; 2 - ель европейская; 3 - ель восточная (из Бородина, Некрасов и др.,1966)
Ель является одной из наименее требовательных к свету пород; по теневыносливости она уступает только тису и пихте, но значительно превосходит сосну, вследствие большей пластичности хвои, которая резко меняет свое положение и строение, в зависимости от того, развивается ли она на свету или в тени. С этим же надо поставить в связь и густую крону ели, медленное очищение от сучьев ствола, медленное изреживание древостоя с возрастом, долгое сохранение хвои на ветвях, относительно тонкую кору. Ель не только прекрасно развивается под пологом сосны, но она заметно не страдает, произрастая под пологом дуба. Находясь под материнским пологом, она может выносить сильное затенение. В этих случаях ель растет медленно и образует широкие зонтикообразные кроны с короткой хвоей, часто достигая в 30-40 лет, а иногда и больше, лишь 1-1,5 м высоты; но, будучи выставлена на простор, она не погибает и вырастает в нормальное для данных условий дерево.
Ель сибирская - Р. obovata Ledeb.
Ель сибирская близка к ели обыкновенной (рис. 54), образует с ней гибриды. Ареалы этих видов перекрываются, и на большей территории Ев
188
ропейской части России, в зоне хвойных лесов, ель представлена гибридными формами (рис. 55). Интрогрессивные54 гибриды елей европейской и сибирской в литературе, преимущественно иностранной, принято называть финской елью (Р. fennica) По мере приближения к Уралу сильнее проявляется преобладание признаков ели сибирской.
Морфологические отличия этих близких видов состоят в следующем: у ели сибирской шишки мельче, чем у европейской (4-8 см длины, чаще 6-7 см), овально-цилиндрической, реже яйцевидной формы; семенные чешуйки широкие, по краю закругленные, цельнокрайние; хвоя более короткая, молодые побеги опушены. Это и есть единственные достоверные морфологические отличия ели сибирской от ели европейской.
Древесина ели сибирской равно, как и других видов ели, произрастающих на территории нашей страны, по анатомическому строению не отличается от ели европейской.
Ареал ели сибирской обширен. В европейской части страны, как мы уже говорили, ель сибирская растет вместе с елью европейской и занимает северные и северо-восточные районы, заходя на юг до нижнего течения реки Камы. Растет она на Урале, в Западной и Восточной Сибири (кроме Крайнего Севера и северо-востока Азиатского континента), на Дальнем Востоке - юг Охотского побережья, бассейн верхнего и нижнего течения р. Амура.
Таким образом, ель в Сибири имеет огромную область распространения, но нигде на севере не доходит до границы произрастания древесных растений. Однако на этом протяжении ель распределена весьма неравномерно. В то время как в Западной Сибири, на Алтае и в Саянах ель - обыкновенное дерево и принимает значительное участие в сложении лесов, за Енисеем и Байкалом она встречается редко, причем ее произрастание все уменьшается по мере удаления к востоку. В пределах Якутии, по побережью Охотского моря, в Приморье, по Амуру и в Забайкалье она встречается редко. В Восточной Сибири ель чаще всего приурочена к узким ущельям гор, к долинам рек и часто сопровождает поймы ручьев и речек.
Для примера можно указать, что в низовьях р. Витима и его притоков, то есть почти в центре восточной половины ареала, ели принадлежит только 3 % общей лесной площади, тогда как лиственнице - 55 % и сосне - 13 %.
В отношении требований к условиям внешней среды ель сибирская близка к европейской. Рассмотрение ее области распространения и распределения ее внутри ареала дает возможность заключить, что она, так же, как и европейская ель, достаточно чувствительна к суровости и, по-видимому, к континентальности климата, а именно, плохо выносит сухость воздуха. Этим, по-видимому, можно объяснить убывание ели к востоку и ее преимущественную приуроченность там к более влажным экологическим разностям.
34 Интрогрессивная гибридизация - естественная гибридизация на границе ареалов двух близких видов, при которой гибридные формы периодически скрещиваются между собой и с исходными видами.
189
P. obovata не дает таких высоких стволов, как Р. abies, достигая лишь 30 м высоты, имеет более короткую хвою, обычно до 20 мм длиною, и более плотную мелкослойную древесину.
По своей роли в лесном хозяйстве ель сибирская значительно уступает европейской. Если еще в Западной Сибири она представляет значительную ценность, то в Восточной - как по малой распространенности, так и по качеству древесины - она мало интересна.
На всем обширном своем ареале ель сибирская не остается совершенно одинаковой. Если форма края чешуи у ели сибирской, в отличие от европейской характеризуется большим постоянством, то в отношении размеров и окраски шишек, консистенции и ориентировки их семенных чешуй, наблюдается большое разнообразие и намечается некоторая географическая дифференциация. Например, прибайкальская популяция ели отличается очень мелкими шишками с более тонкими семенными чешуями.
В заключение, следуя изложению данной темы академиком В.Н. Сукачевым, коснемся вопроса, чем объясняется предел распространения представителей этого рода на юге и на севере.
Есть основания считать, что современная южная граница ели находится в состоянии движения на юг. Продвижение происходит не прямым заселением степи, а постепенным внедрением ели в идущие впереди дубовые леса. В настоящее время частичная смена дубовых лесов еловыми на самом деле имеет место. И так как ель, поселившись на черноземе или близкой к нему почве, превращает ее в подзолистую, то понятно, что южная граница области ели будет совпадать с северной границей чернозема. В прошлом, когда человек начал свою энергичную деятельность по уничтожению лесов, когда дубовые леса на северной части лесостепной полосы были распространены значительнее, чем теперь, ель не успела занять всей территории, на которой она могла бы расти.
Таким образом, не отрицая того, что по своим экологическим свойствам ель не приспособлена к условиям черноземно-степной области, особенно к ее сухости, все же надо признать, что она еще не успела достигнуть своего южного предела, определяемого этими условиями. Если бы не современное влияние человека, она продолжала бы свое поступательное движение на юг, занимая ту площадь, которую она некогда, по-видимому, даже до ксеротермического периода, уже занимала.
Не менее сложен вопрос о причинах, определяющих северный предел распространения ели, имея в виду и Р. abies, и Р. obovata.
Как мы видели, в пределах европейской части России ель доходит на севере почти всюду до границы леса с тундрой, да и в Западной Сибири лишь лиственница заходит дальше ее (и то сравнительно немного), к северу. Однако, чем дальше к востоку, тем больше к югу отодвигается северный предел ели по сравнению с лиственницей, березой, осиной и тополем.
Одна из причин угнетения деревьев у границы с тундрой - в высыхании молодых побегов во время зимних месяцев, когда испаряющаяся вода не может возмещаться корнями. Сильные сухие ветры зимою, дующие в тундре,
190
вызывают особенно сильное высыхание, что и является причиной полного безлесья тундры.
Низкая температура почвы также вызывает нарушение водного баланса деревьев, поэтому не только зимою, но и в остальное время года, когда создаются, хотя бы временами, условия, сильно повышающие транспирацию древесных пород, при наличии холодной почвы они оказываются в экстремальных условиях. Поэтому и ель в пределах Европы и Западной Сибири, всюду доходя до северной границы леса, не идет далее к северу.
В Восточной Сибири лиственница, береза, осина, тополь заходят к северу значительно дальше ели, сосны, пихты, т, е. мы видим, что здесь все породы, не сбрасывающие своей листвы, в том числе и хвойные, как ель и сосна, заметно отступают от северной границы леса
Хвойные в общем на единицу испаряющей поверхности расходуют влаги значительно меньше, чем большинство лиственных. Однако для оценки рассматриваемого явления нужно принять во внимание, что у хвойных площадь поверхности побега с остающейся на зиму хвоей увеличивается, сравнительно с голым побегом лиственных пород и лиственницы, в 150-300 раз. Такая громадная поверхность дает, несмотря на малую интенсивность испарения, очень большую величину потери воды, превышающую потерю воды голых побегов в 50-100 раз. Этот расчет имеет значение при оценке того, каков должен быть ток воды к побегу, чтобы перекрыть его траты на испарение.
Большое значение имеет продолжительность вегетационного и зимнего периодов. Длительность вегетационного периода имеет значение в том смысле, что содействует вызреванию побегов и понижает их зимнее испарение, а продолжительность зимнего периода определяет промежуток времени, когда не подается вода в ткани растения и происходит высыхание побегов. В этом отношении северо-восток Азии находится в исключительно неблагоприятных условиях. Весна на северо-востоке Азиатского континента значительно запаздывает. Зимний период очень продолжителен. Если поставить все это вместе, то будет понятно, почему северная граница ели, сосны, и пихты, так значительно опускается к югу в Восточной Сибири.
Типы ельников
Ельники по своему составу, строю, генезису и производительности могут быть объединены в определенные группы. Известно, что типы леса находятся в определенной связи с условиями среды. Чтобы установить эту связь, прежде всего примем во внимание, что в пределах северной половины европейской части России можно наметить следующие пять основных типов условий произрастания еловых лесов:
1) почвы более или менее богатые, глинистые, суглинистые или супесчаные, хорошо дренированные при достаточно выраженном рельефе; на севере большей частью приуроченные к рекам;
191
2) такие же почвы, но заболачивающиеся, менее дренированные, с менее развитым рельефом, на севере большей частью расположенные дальше от реки;
3) почвы еще более заболоченные, с нерасчлененным рельефом, равнинные, расположенные еще дальше от рек; эти три типа условий произрастания составляют как бы один ряд, связанный с последовательным усилением степени заболачивания;
4) дно узких долин рек и ручьев, где появляется избыточное увлажнение, но воды в большей или меньшей степени проточные;
5) места, имеющие для севера исключительно богатые почвы, чаще всего связанные с близким залеганием известковых пород.
Этим пяти типам условий произрастания соответствуют следующие группы типов леса.
1. Рельеф, большей частью, достаточно развит, площади хорошо дренированы, почвы - то более, то менее богатые суглинки, глины или супеси, не заболочены. - Ельники-зеленомошники.
2. Рельеф менее развит, места слабо дренированы, почвы те же, но несколько заболочены. - Ельники-долгомошники.
3. Рельеф равнинный или представляет собою дно котловин, места не дренированы, почвы заболочены. - Сфагновые ельники.
4. Дно логов с заболоченными почвами, но с проточной водой. - Болотно-травяные ельники.
5. Места с богатыми, хорошо дренированными почвами, большею частью с близким залеганием известняков. - Сложные ельники.
Приведем главнейшие, более изученные типы ельников средней полосы европейской части России, не давая, однако, здесь подробной их характеристики, а указывая лишь некоторые, более типичные, отличительные черты.
Ельники-зеленомошники. Эти ельники характеризуются первым ярусом из одной ели, лишь иногда с небольшой примесью лиственных пород (осины и березы), отсутствием особого второго яруса, отсутствием сколько-нибудь значительно развитого подлеска, негустым и небогатым травяным покровом и мощным сплошным покровом из так называемых блестящих мхов. Они занимают в общем не очень богатые, но свежие или влажные почвы.
Если травяной покров ельников-зеленомошников, как упоминалось выше, не отличается значительной густотой, то, напротив, моховой покров обычно бывает совершенно сплошной, состоя, главным образом, из следующих видов: Helocomium prolifemm, Pleurdzium schreberi, Rhytidiadelphus triquetrus, Dicranum undulatum и др. Ельники этой группы по общему своему строю весьма выдержаны - организация и требования к условиям среды подчиненных ярусов строго соответствуют биологическим особенностям ели.
Главнейшие типы леса: ельник-кисличник, ельник-черничник, ельник-брусничник.
192
Ельники-долгомошники. Долгомошники характеризуются первым ярусом из ели, к которой примешана, в большей или меньшей степени, береза. Второй ярус совершенно отсутствует, подлесок еще менее развит, чем в предыдущей группе ельников. Рост ели также гораздо хуже: деревья больших размеров не достигают, сбежисты, часто увешаны бородатыми лишайниками. Травяной покров значительно беднее как в отношении видовом, так и по числу особей. Зато чрезвычайно характерен мощный, сплошной ковер из мха - кукушкина льна (Polytrichum commune). Эти ельники занимают, как правило, ровные, пониженные местоположения с избыточно увлажненными суглинистыми и супесчаными, сильно оподзоленными почвами.
Ельники этой группы довольно однообразны и не дают такого разнообразия типов, как первая группа; они очень широко распространены по всему северу. Типичным представителем этой группы будет елъник-долгомоишик, к характеристике которого относится все сказанное для всей этой группы.
Сфагновые ельники. Общими чертами типов леса этой группы являются весьма угнетенный рост ели, которая почти не дает строевого материала, малые полноты первого яруса, примесь, подчас значительная, березы, а иногда и сосны, значительное участие в моховом ковре сфагнума (Sphagnum magellanicum, S. fuscum), иногда до почти полного вытеснения всех остальных мхов, и произрастание на очень сырых почвах, часто со значительным торфянистым горизонтом.
Если в ельниках-долгомошниках роль созидателя сообщества оспаривается у ели кукушкиным льном, то здесь в этом отношении с елью конкурирует, и притом успешно, сфагнум. Ель уже не оказывает на среду значительного влияния: она больше сама во власти среды; последняя же в значительной части есть создание сфагнового ковра. Поэтому в травяном покрове участие обычных представителей елового леса сведено до минимума, напротив, преобладают болотные растения.
Болотно-травяные ельники. Это насаждения с густым, высоким травяным покровом. Характерной чертой этих ельников является приуроченность их ко дну долин небольших речек и ручьев в виде узких полос с богатой, сырой, даже мокрой почвой, весною затопляемой полыми водами, но обязательно с проточной водой. Древостой негустой, из ели с примесью березы. Подлесок развит довольно хорошо. Травяной покров густой, высокий и пышный, а моховой покров развит слабо.
Представители этой группы типов: ельник-лог, травяно-сфагновый ельник.
Сложные ельники. Эта группа типов характеризуется хорошим ростом ели, участием в составе фитоценозов широколиственных пород; они то входят в состав первого яруса, то остаются лишь во втором ярусе или в подлеске. Затем весьма характерен густой и разнообразный подлесок. Травяной покров умеренно развит, часто с представителями, свойственными широколиственным породам. Моховой ковер, большей частью, нетолстый и несплошной. Почвы в типах этой группы свежие или даже суховатые, всегда достаточно богатые.
193
Сложные ельники характерны для более южных областей, в которых они довольно распространены.
К этой группе ельников в числе прочих относятся: липовый ельник и дубовый ельник, образующие древостои высокой производительности и встречающиеся преимущественно в зоне перехода между хвойными и широколиственными лесами.
Род пихта - Abies Mill.
Род пихта объединяет виды, представленные крупными деревьями, преимущественно образователями горных лесов. Кора стволов тонкая, гладкая или слаботрещиноватая, со смоляными ходами. Крона густая, коническая, обычно низкоопущенная. Ветвление мутовчатое, с межмутовчатыми побегами. Почки округлые, реже тупоконические, засмолены или не покрыты смолой. Хвоя расположена на побегах одиночно, спирально, настильно или строго двухрядно. К стеблю хвоя прикрепляется без листовых подушечек на коре, в поперечном сечении плоская или узкоэллиптическая, мягкая, сверху большей частью желобчатая, темно-зеленая, снизу килеватая, с двумя беловатыми полосками рядов устьиц (редко устьица могут быть и на верхней поверхности); края хвои загнуты на низ; хвоя побегов, несущих шишки, четырехгранная, с белыми полосками и устьицами на всех гранях. При основании хвоя обычно сужена, а затем расширена в округлую пятку, оставляющую после опадения на побеге плоский округлый след; на конце хвоя притуплённая или раздвоенная, реже (у пихт цельнолистной, аризонской) - жесткая, заостренная. Держится на побегах 7-10 лет или более.
Микро- и макростробилы закладываются в генеративных почках (в год, предшествующий пылению), более крупных и овальных, чем почки ростовые. Опыление происходит перед распусканием молодой хвои. Женские шишечки пихт стоят вертикально, сохраняя это положение и в период созревания семян. Зрелые шишки цилиндрические, 5-12 (20) см длины, до созревания засмолены. При созревании семян шишка начинает интенсивно распадаться на чешуйки, которые вместе с семенами разносятся ветром. Обычно это происходит уже в начале-середине осени, спустя 4-4.5 месяца после пыления. После разрушения шишек на побегах остаются вертикально стоящие деревянистые стерженьки, которые довольно долго сохраняются в кроне.
Семена обратнояйцевидно-клиновидные, тупоугловатые, -плотно соединены с треугольным крылом, основание которого покрывает верхнюю сторону семени и большую часть его нижней стороны. Всходы имеют 4-5(7) плоских семядолей. Помимо семян пихта способна размножаться отводками, укореняясь нижними ветвями.
Пихта в молодости растет медленно, но затем ее рост ускоряется. Долговечность разных видов пихты составляет от 150-250 (пихты белокорая и сибирская) до 450-500 (пихта цельнолистная) и даже 800 лет (пихта кавказская).
194
Все пихты очень теневыносливы, довольно требовательны к плодородию почвы, по отношению к влажности почвы и атмосферы - мезофиты, весьма чувствительны к содержанию промышленных выбросов в атмосфере (негазостойки), часто страдают от поздних весенних заморозков (особенно в молодости), а по своей зимостойкости - различны в зависимости от географического происхождения видов и их экотипов.
Пихты имеют большое народнохозяйственное значение. Древесина их легкая, белая или желтоватая, без смоляных ходов. Используется как сырье для целлюлозно-бумажной промышленности, строительства и тарного производства. В коре пихты в особых желваках (смоловместилищах) содержится ароматная живица, богатая скипидаром. Из живицы пихты сибирской и бальзамической добывают пихтовый бальзам, применяемый в оптической промышленности, медицине и для других целей. Из хвои пихты получают масло, используемое как сырье для производства камфары. В семенах пихты содержится до 30 % растительного масла, употребляемого при изготовлении лаков.
Пихта сибирская - Abies sibirica Ledeb.
Дерево до 30 м высоты и до 0,5 м в диаметре с узко-конической заостренной кроной. Ветви расположены более или менее мутовчато, имеются и межмутовочные побеги. Верхние и средние сучья и ветви растут горизонтально, нижние обычно свисают к земле и даже укореняются, как отводки. В сомкнутых насаждениях кроны подняты более высоко, и в этом случае резко выделяются полно древесные стволы с гладкой тонкой темно серой корой. На коре образуются желваки, из которых при надрезании в жаркие летние дни вытекает душистая прозрачная смола. Часто внизу стволов проходят продольные впадины, отчего стволы, особенно на поперечном разрезе, имеют неправильную форму.
Побеги у пихты сибирской гладкие, удлиненные, с редкими толстыми волосками и спирально сидящей мягкой хвоей. Заканчиваются зеленоватобурыми шаровидными почками, залитыми светлой смолой. На большей части кроны на боковых побегах скрученная в основаниях хвоя расположена на-стильно. Хвоя узкая, тупая, длиной 1,5-3,5 см, с верхней стороны темнозеленая, блестящая, с нижней с двумя беловатыми узкими полосками, образованными каждая тремя-четырьмя (шестью) рядами устьиц. При опадании хвоя оставляет на побегах белые рубцы в виде маленьких овальных пятен. В наиболее затененной части хвоя располагается в два ряда, гребенчато. В остроконечной части кроны жесткая световая хвоя отходит от побегов в сторону почти под прямым углом и саблевидно изогнута. Смоляные ходы залегают в ее паренхиме. Хвоя держится на дереве до 10 лет; при растирании издает приятный запах (рис. 56).
Опыление у пихты сибирской происходит в мае. В это время в кроне появляются кое-где овальные желтоватые мужские стробилы, а на самой вершине - зеленоватые или буровато-красные продолговатые шишечки (женские стробилы). В период опыления микроспорофиллы раскрываются в
195
виде поперечной щели, из которой вылетает пыльца, снабженная воздушны-
ми мешками.
Рис. 56. Пихта сибирская: 1 -общий вид; 2 - побег с хвоей; 3 - хвоинка; 4 - засмоленные почки на верхушке побега; 5 - созревающая шишка; 6 - семенная и кроющая (меньше, с неровным краем) чешуи, вид с морфологически нижней стороны; 7 - семя с "крылышком"; 8 - участок побега с осью рассыпавшейся шишки (из Коровин, Зуихина, Потапова, 2003)
Шишки при созревании буреют, иногда покрываясь частично смолой. Форма их овально-цилиндрическая, с тупым концом; длина 5-9 см, ширина 2-
4 см. Семенные чешуи широкие, бархатистые, с закругленным и мелко зазубренным верхним краем, при основании с короткой ножкой. Кроющие чешуи с округлой и зазубренной пластинкой, переходящей в острие, едва достигают половины, трети длины семенных и снаружи не видны.
Семена созревают к концу августа. В сентябре - октябре шишки рассыпаются, и на ветвях остаются лишь бурые стерженьки. С чешуями опадают и семена. Семена светло-буро-желтоватого цвета неправильно обратнояйцевидной, в основании клиновидной формы с тупыми гранями. Длина 6-7 мм. Семя снабжено крыловидным придатком (крылаткой) клиновидной формы с косым верхом, длиной около 1 см. Крылатка от семени не отделяется, при обламывании часть крылатки остается при семени, окружая его на три четверти. Семена смолистые и горькие на вкус. Семена при хранении довольно быстро теряют всхожесть. 1000 семян весят 6-8 (11) г.
Пихта сибирская в пределах России занимает обширную территорию. Граница ареала на западе начинается от верховьев р. Ваги (около Вологды), идет по течению этой реки до Северной Двины, переходит ее и через среднее течение р. Пинеги и верхнее течение р. Мезени направляется на Урал. В Сибири граница ареала пересекает р. Обь около Березова и р. Енисей под 670 40' с. ш., далее на восток снижается, переходит р. Лену под 60° с ш. и охватывает р. Алдан и западную часть Забайкалья. Южная граница от верховьев р. Ваги идет восточнее г. Костромы, направляясь далее вдоль рек Волги и Камы на восток; здесь, делая поворот на Урале (до 55° 35' с. ш. ), уходит в Западную Сибирь, проходит недалеко от г. Тюмени, затем идет под 57° с. ш. и поворачивает на Алтай (рис. 57).
Отдельными участками пихта сибирская произрастает в горах Средней Азии (в Джунгарском Ала-Тау и Западном Тянь-Шане), встречается на Саянах и в Северной Монголии. В горы поднимается до 2000 м. На равнинных местах обычно растет в долинах рек, часто - на возвышенных берегах север
196
ных рек. В культуре известна с 1820 года и заходит далеко на запад, вплоть до Балтийского моря.
Рис. 57. Ареалы пихты сибирской, пихты цельнолистной, сосны обыкновенной, дуба монгольского, липы мелколистной и граба обыкновенного в пределах СНГ ( из Бородина, Некрасов и др., 1966)
Пихта сибирская весьма зимостойка и свободно переносит очень низкие температуры. Этому способствуют слабое испарение влаги зимой и залитые смолой почки. В западных областях России нередко страдает от поздних весенних заморозков при более ранней весне и более раннем начале роста, чем в условиях Сибири.
Пихта сибирская очень теневынослива, что подтверждается ее густым охвоением, теневой хвоей, наличием самосева в затененных местах, удовлетворительным ростом под пологом других пород, слабым самоизреживанием. Только остроконечные вершины пихты покрыты световой хвоей.
Затененный зонтикообразный подрост пихты высотой до 0,5 м сохраняется до 50 лет и может выжить после выставления на свет оправиться. Остроконечные вершины пихты покрыты световой хвоей.
К почве пихта сибирская довольно требовательна. Лучше всего растет на дерново-подзолистых, суглинистых почвах. Избирает достаточно дренированные местоположения или горные склоны. Хорошо растет там, где в подпочве залегают известняки. Застойно-заболоченных и сильно подзолистых почв избегает, но переносит проточное увлажнение. На бедных песчаных почвах не растет.
197
Пихта сибирская обладает глубокой корневой системой, состоящей из стержневого корня длиной до 1,5 ми нескольких боковых, косо и глубоко уходящих в почву. Корневые волоски не развиты. Имеется корневая микориза. Иногда в связи с развитием корневых грибных болезней и гнилей деревья вываливаются.
Растет пихта сибирская в первые годы довольно медленно, едва достигая в четырехлетием возрасте 10-15 см, затем ее рост усиливается и не прекращается до глубокой старости.
Удовлетворительно растет по всей лесной зоне. Доживает до 250 лет, но обычно из-за болезней отмирает раньше. Плодоносит и дает самосев. На черноземах в полосе дубрав растет удовлетворительно и к 85 годам достигает 25 м высоты. Пихта сибирская начинает семеносить на свободе с 30 лет, а в насаждении - с 50-70 лет. Часто неблагоприятные условия погоды приводят к задержке семеношения. Поэтому урожайные годы наступают через 2-3 года, на севере - через 4-6 лет. Всхожесть семян колеблется в пределах 50-60 % (до 85), а через год падает до 30 %. Всхожесть 45 % соответствует I классу качества.
Размножается пихта сибирская в основном семенами. Всходы несут обычно 4 (3-5) семядолей длиной 1, 2 см. Семядоли темно-зеленые, с неясными полосками вдоль жилки с верхней стороны и ярко-зелеными с нижней. Первичная хвоя похожа на семядоли, но имеет выемку на вершине и ясные полоски из рядов устьиц снизу. Под пологом пихты сибирской обычно присутствует густой самосев.
Пихта сибирская растет как в чистых, так и в смешанных насаждениях, с елью. Произрастая вместе с елью, вытесняет сосну. Высоко в горах растет в виде приземистого кустарника, достигая верхней границы леса.
Пихта сибирская по сравнению с другими отечественными хвойными -наименее ценная порода. Древесина ее белая, с желтоватым оттенком, без смоляных ходов. Заболонь зрительно не отличается от центральной, более спелой, древесины. Годичные кольца заметны хорошо, так как поздняя древесина выделяется более темным, цветом. Древесина малопрочная, неустойчивая к гнили, применяется для тех же целей, что и еловая, но последней всегда отдают предпочтение.
Анатомические особенности древесины. Поперечный срез. Ядро отсутствует. Граница между годичными слоями отчетливая. Более или менее постепенный переход от ранней древесины к поздней. Осевая паренхима скудная, часто отсутствует. По расположению диффузная, редко встречаются клетки терминальной паренхимы. Нормальные смоляные ходы отсутствуют; встречаются тангенциальные цепочки травматических смоляных ходов. Тангет^иалъный срез. Лучи однорядные без смоляных ходов. Высота лучей обычно 1-25 клеток. Радиальный срез. Поры на радиальных стенках ранних трахеид расположены в 1 ряд, реже - в 2 ряда супротивно. Лучевые трахеиды отсутствуют. На полях перекреста в ранней древесине таксодиоидные поры (см. приложение 2). Стенки внутренних клеток у лучей толстые; тангенциальные стенки узелковые и зубчатые, горизонтальные - гладкие и выемчато-пористые. Пограничные клетки лучей тонкостенные.
Кора молодых стволиков пихты сибирской шелушится, с возрастом становится гладкой. Из коры добывается так называемый пихтовый сибир
198
ский бальзам. Смолистость коры достигает 15 %. Живица содержит свыше 25 % скипидара. Сибирский бальзам используется главным образом в оптической промышленности для склеивания стекол, а также при изготовлении микроскопических препаратов. Большое применение имеет пихтовая лапка -мелкие ветки с хвоей. С 1 га хорошего пихтарника получается 10-12 тонн лапки. Из лапки добывается пихтовое масло, богатое борнеолом и борнилацетатом, являющееся сырьем для получения камфары, а также использующееся в парфюмерии и как антисептик.
Особая чувствительность пихты сибирской к дымовым газам и копоти делают ее непригодной для разведения вблизи фабрик и заводов, в городах с сильным задымлением.
Типология пихтарников изучена значительно меньше типологии ельников. Пихтовые леса по своей приуроченности к элементам рельефа и многим другим требованиям к условиям среды в основном повторяют еловые. Отличие определяется главным образом тем, что пихта более чем ель требовательна к плодородию почвы и еще меньше ели способна переносить заболачивание. В целом же пихтарники с некоторыми допущениями можно подразделять на те же типы леса, что и ельники.
Сосна кедровая сибирская - Pinus sibirica (Rupr) Mayr.
Характеристика рода приведена ниже в разделе "Сосновые леса".
Важнейшая лесная порода Сибири: в Восточной и Западной Сибири занимает миллионы гектаров площади. Растет в виде мощных деревьев высотой до 30-37 м и толщиной до 60-120 см. Отличается толстыми сучьями, особенно в редколесье, и густой кроной. Кора темная, с коркой в виде пластинок. Толстые удлиненные побеги, покрытые чешуевидными листочками и рыжими волосками, несут многочисленные укороченные побеги с пучками хвоинок по 5 штук. Пленчатые желто-бурые влагалища у пучков опадают в первый год. Плотная торчащая темно-зеленая хвоя достигает 13 см длины и держится 3-6 лет. Хвоя в разрезе трехгранная, по краям зубчатая. Смоляные ходы в хвое периферические или лежат в паренхиме. В центральном цилиндре помещается один сосудистый пучок.
Пылит сосна сибирская в июне - июле. Мужские стробилы оранжевокрасного цвета, собраны в крупные соцветия у основания побега. Женские стробилы прячутся в хвое у конечной почки, сначала имеют красноватую, после - фиолетовую окраску. После опыления мужские собрания стробилов опадают. Процесс оплодотворения заканчивается лишь во второй половине июня следующего года В это время шишка достигает величины небольшого лесного ореха (озимь), а затем начинает усиленно расти. Созревание семян (орешков) заканчивается в августе (рис. 58).
В сентябре начинается сбор шишек со зрелыми семенами. С середины октября по середину ноября все шишки с семенами опадают. Шишки имеют
199
в длину 6-13 см. Их плотные, прижатые чешуйки покрыты жесткими волос-
ками.
Рис. 58. Сосна сибирская кедровая: 1 - побег со зрелой шишкой; 2 - верхушка побега с мужскими стробилами; 3 - микроспорофилл с двумя микроспорангиями (пыльниками), из которых высыпается пыльца; 4 - семенная чешуя с двумя созревшими семенами ("кедровыми орешками"); 5 - вычлененный из семени зародыш (вверху -семядоли) (из Буш, 1915)
Утолщенные широкие щитки семенных чешуй заканчиваются светлым "пупком". Шишки содержат по 40-150 бескрылых семян. Длина семян 7-9 мм. Вес 1000 шт. 230-250 г. Семена состоят из твердой оболочки, пленки и эндосперма, внутри которого лежит зародыш. При благоприятной погоде в урожайные годы в хороших насаждениях добыва
ется от 100 до 400 кг семян с 1 га. Доброкачественность, или жизнеспособность, семян кедра, равная 90 %, соответствует I классу качества по ГОСТу.
Современный ареал сосны сибирской на западе доходит до верховьев р. Вычегды. На севере граница пересекает Урал под 66° с. ш. И на юге - около 57° с. ш., в Сибири идет через нижнее течение рек Оби и Енисея (68° с. ш.), направляется к г. Олекминску на р. Лене и охватывает верховья р. Алдана. Затем она круто повертывает на юго-запад, проходит через Забайкалье и уходит на территорию Монголии, захватывая Саяны и Алтай (рис. 59). В культуре встречается вплоть до Балтийского моря. Сосна сибирская очень устойчива к холодам. Растет в условиях сурового континентального климата, в горах, на равнинах, преимущественно в районах, примыкающих к рекам. Образует темнохвойный лес и хорошо переносит в благоприятной обстановке затене
ние.
Особенно хорошо растет сосна сибирская на суглинистых почвах, даже с избытком увлажнения, по берегам рек, то есть в местах с влажным воздухом и влажной почвой, достигая здесь больших размеров. Хуже растет на сухих песках и каменистых почвах, а также на заболоченных местах, где высота деревьев обычно не превышает 18 м, а чахлые кроны почти не плодоносят. Высоко в горах развивается также плохо, образуя флагообразную крону, слабо плодоносит и сильно заражена гнилями. За Байкалом уступает место кедровому стланику. Особенно сильно страдает от пожаров.
200
Корневая система у сосны сибирской развита хорошо. Кроме стержневого, не особенно длинного корня, образует большие крепкие боковые. На болотах развивает поверхностную корневую систему.
Рис. 59. Ареалы пятихвойных сосен в пределах России: 1 - сосна кедровая сибирская; 2 - кедровый стланик; 3 - сосна кедровая корейская
При смене лиственными породами поселяется под их пологом. Благодаря теневыносливости и долговечности со временем образует кедровые насаждения. Расселению сосны сибирской способствуют белка, бурундук, кедровка и другие обитатели тайги.
Всходы в первый год достигают не больше 4 см в высоту и несут 8-10 семядолей. На второй год сеянец вырастает до 7-8 см и так же медленно растет в последующие годы. После 15-20 лет рост улучшается. Несмотря на медленный рост, сосна сибирская достигает более 32 м в высоту и более 1,5 м в диаметре. Живет 400 лет и больше. На севере томской области встречались экземпляры по 475 лет с нормальной кроной и без признаков гнили в древесине ствола.
При достаточном освещении сосна сибирская начинает плодоносить с 20-25 лет, а в густом насаждении иногда только с 50-60 лет. Период усиленного плодоношения начинается с 80 лет, к 100 годам достигает максимума и к 240 годам существенно снижается, а иногда и сходит на нет. Урожайные годы чередуются с менее урожайными, в зависимости от условий погоды. Отмечена зависимость выхода семян от диаметра ствола. С увеличением
201
диаметра пропорционально увеличивается урожай семян. Урожай семян с одного дерева в урожайные годы достигает иногда 80-100 кг.
При разумном ведении лесного хозяйства прижизненное использование сосны сибирской кедровой выгоднее, нежели заготовка "кедровой" древесины. Назначать кедровники в рубку следует лишь после снижения урожайности. Местное население и существовавшие ранее госпромхозы заготавливали большое количество семян ("кедровых орехов"), которые используются в сыром виде, а также для приготовления сливок, отличающихся высокой калорийностью и содержащих большой набор витаминов. В среднем семена содержат 57 % скорлупы, 42 % ядер и около 1 % пленки. Маслянистость кедровых орешков исключительно высока: свыше 60 % веса ядра и 25 % всего веса семян.
Древесина сосны сибирской сравнительно мягкая, в отличие от древесины сосны обыкновенной хорошо полируется. Граница между поздней и весенней ранней древесиной неясная. Используется древесина в строительстве, для производства пиломатериалов, карандашей. Живица, получаемая при подсочке кедра, применяется для лечебных целей. Из кедровой хвои извлекают эфирное масло и витамин С.
Анатомические особенности. Поперечный срез. Ядро имеется. Граница между годичными слоями отчетливая. Переход от ранней древесины к поздней постепенный. Смоляные ходы с тонкими эпителиальными клетками, крупные, расположены по одиночке. Тангенциальный срез. Лучи в основном однорядные; встречаются 2 (4) -рядные лучи со смоляными ходами. Высота лучей 2-15 (20) клеток. Радиальный срез. Окаймленные поры на стенках трахеид расположены преимущественно в 1 ряд. На полях перекреста в ранней древесине в основном оконцевые поры, иногда встречаются 2-3 крупных пиноидных поры на одном поле перекреста. Имеются лучевые трахеиды. Горизонтальные клетки лучевых трахеид гладкие, волнистые. Имеются крассулы.
Леса с участием кедровой сосны - наше национальное богатство. Они удовлетворяют многообразным нуждам народного хозяйства, а также имеют большое почвозащитное и водоохранное значение, дают приют и питание ценному пушному зверю - белке, соболю и др., являются местом сбора полезных растений и т. д. Кедровая сосна весьма декоративна. Самые старые посадки ее известны под Ярославлем, где они были произведены в 70-х годах XVI века.
Типы кедровников
Многообразие географических и почвенных условий произрастания, исторических особенностей происхождения, характер взаимоотношения с другими древесными породами - все это вместе обусловило большое типологическое разнообразие кедровых лесов.
Типы кедровников объединяются в группы типов леса. Рассмотрим некоторые из них, в соответствии с классификацией известного ученого лесовода Г.В. Крылова.
Кедровники широкотравные имеют распространение в наиболее обеспеченных теплом и влагой районах Алтая и Западного Саяна. Развиваются на
202
маломощных бурых, хорошо дренированных почвах. Древостои двухъярусные, П-Ш классов бонитета с преобладанием кедра в первом ярусе и пихты во втором. Подрост с преобладанием пихты, реже кедра от 500 до 2000 экземпляров на 1 га. Подлесок сомкнут слабо и состоит из спиреи и смородины, волчьего лыка. Эта группа получила название по наличию в составе травяного покрова ряда спутников широколиственных лесов, в частности, овсяницы лесной, осморизы амурской, подлесника европейского, ясменника душистого, копытеня европейского.
Кедровники папоротниковые распространены в южной подзоне тайги, предгорьях и низкогорьях Урала, Алтая и Саян. Они достаточно разнообразны, но наиболее характерны в черневых лесах, где приурочены к склонам теневых экспозиций с заметно оподзоленными бурыми лесными почвами. Древостой двухъярусные, как правило, высокополнотные I—II бонитетов. Подрост редкий, с преобладанием пихты. Подлесок - из спиреи дубравколист-ной, рябины, жимолости алтайской, черемухи и бузины. Травяной покров хорошо развит и в основном представлен различными видами папоротников, в частности страусника обыкновенного, кочедыжника женского, щитовника игольчатого, а также борца высокого, вейника тупоколоскового, крапивы двудомной и др. Под покровом крупных трав много низкотравья из кислицы, майника, звездчатки, копытня европейского. Моховой покров развит очень слабо.
Кедровники крупнотравные (высокотравные) представлены главным образом в горах, а также в подзоне южной тайги. В горах они имеют два оптимума развития: в черневых, или низкогорных, лесах и субальпийских - высокогорных. Произрастают эти кедровники в горах на бурых, зернистых, влажных тяжелосуглинистых оподзоленных и неоподзоленных почвах, а на равнине на дерново-подзолистых и влажных серых лесных почвах. Древостой в черневом и таежном подпоясах двухъярусный с производительностью I—II классов бонитета с кедром в первом ярусе и пихтой и березой во втором. По мере поднятия в горы производительность их падает до IV класса бонитета, а в субальпийском подпоясе они становятся редкостойными и чистыми, без примеси других пород. Травяной покров этих кедровников имеет очень высокую сомкнутость и состоит из борца высокого, вейника тупоколоскового, соссюреи широколистной, чины Гмелина, скерды сибирской и других видов, свойственных лесным и субальпийским лугам.
Кедровники разнотравные (низкоразнотравные, осочковые) развиваются в южной подзоне тайги, а также в низкогорных и горных лесах на дренированных склонах световых экспозиций достаточно увлажненных супесчаных суглинистых дерново-подзолистых, скрытоподзолистых и темно-серых лесных почвах. Это очень распространенная группа типов леса. Бонитет древостоев колеблется в пределах от II до V классов в зависимости от местонахождения и типа леса. В травяном покрове здесь господствуют лесные злаки и осоки, представленные мятликом однолетним, мятликом сибирским, коротконожкой перистой, костером безостым, осокой большехвостой, осокой стоповидной, осокой сероватой, а также костяникой, подмаренником север
203
ным, геранью белоцветковой, ирисом русским, розгой золотой, кровохлебкой лекарственной.
Кедровники вейниковые встречаются небольшими участками в низкогорных и горных лесах и южной подзоне тайги на влажных и сырых супесях, суглинках, и глинистых почвах дерново-подзолистого типа. Древостой производительностью III и II классов бонитета. По плодородию почв они уступают крупнотравной группе. Характерной особенностью этих кедровников является сильное доминирование в травяном покрове вейника тупоколоскового и тростникововидного.
Кедровники травяно-болотные очень широко распространены по всей южной тайге, в низкогорьях и на средних по высоте экспозициях, а также в переувлажненных долинах рек, на иловато-перегнойных, дерново-глеевых и прочих аллювиальных илистых почвах. Древостои, как правило, редкостойные и разновозрастные с участием ели и березы. Пихта заметно примешивается только в условиях меньшей переувлажненности. Бонитет чаще всего IV, а на севере опускается до V. В среднетаежных и северных районах травяной покров содержит много представителей болотных растений, а на юге и в горах - лабазник вязолистый, вейник Лангсдорфа, живокость высокую и др. Подлесок довольно густой и состоит из черной смородины, спиреи иволи-стой, свидины, рябинника, курильского чая, березы низкой.
Кедровники зеленомошные, или мшистые, имеют наибольшее распространение в таежных лесах равнин и гор, отличаются значительным разнообразием. По экологическим условиям и преобладанию отдельных видов мхов эта группа подразделяется на три подгруппы, из которых наиболее четко выделяются подгруппы на свежих супесях с господством .в напочвенном покрове мха Шребера и брусники, на средневлажных легких суглинках - перистого и этажного мхов с участием гребенчатого мха, а также княженики, грушанки и на влажных суглинках и сырых глинах - древовидного и трехгранного мхов. Древостои этой группы имеют производительность от II до V классов бонитета в зависимости от географического и высотного положений.
Кедровники долгомошные произрастают на сырых подзолисто-глеевых или торфяно-подзолистых почвах по всей зоне тайги. В отличие от других эта группа является промежуточной от мшистых к сфагновым кедровникам. В равнинной части Западной Сибири в моховом покрове, как правило, преобладают мхи с участием кукушкина льна, а в горах к этому покрову прибавляются вейник Лангсдорфа, черника, брусника, линнея северная и др. Производительность древостоев невысокая. Чаще всего она характеризуется IV классом бонитета, иногда Va на севере и в субальпийском подпоясе. По сравнению с другими типами кедровников древостой здесь с большим преобладанием кедра, с небольшой примесью ели и березы. В подросте также преобладает кедр, меньше встречаются ель, береза и еще реже пихта.
Кедровники сфагновые распространены повсеместно в равнинных лесах на заболоченных местоположениях. Древостои этой группы разновозрастные, сильно разреженные с большой примесью сосны и березы, производительность V-Va классов бонитета. В подросте преобладает кедр, реже ветре-
204
чаются ель и сосна. Количество подроста невелико, но оно вполне обеспечивает замену отмирающего перестойного древостоя. Нередко в таких кедровниках после пожара, когда торфяной слой прогорает на значительную глубину, можно наблюдать смену этих кедровников на зеленомошные. Чаще всего это происходит через смену пород, когда на месте хвойных поселяются береза и осина, а под их пологом затем селится кедр.
Деревья второй величины, кустарники, травяной напочвенный покров
Характерными для темнохвойных лесов северной лесной зоны деревьями второй величины являются рябина и ива козья.
Рябина обыкновенная (Sorbus aucuparia L.) достигает иногда 15 м высоты и диаметра 15-18 см (в редких случаях до 40 см), довольно часто растет в виде кустарника. Широко распространена во всей лесной зоне России, доходя почти до тундры. К почве не требовательна, но предпочитает более рыхлую и влажную. Достаточно теневынослива. Дает твердую крепкую древесину, которая может использоваться на мелкие поделки и топливо. Плоды съедобны, содержат сахара и витамины.
Ива козья (Salix caprea L.) встречается в виде дерева второй величины или высокого кустарника. Растет под пологом леса или по опушкам на почвах разной степени увлажнения. Входит во второй ярус как хвойных, так и лиственных насаждений. Распространена по всей стране, доходя до крайнего севера. Растет очень быстро, особенно порослью от пня. Древесина красноватая. Довольно плотная. Кора содержит много танинов, всегда была объектом заготовки сырья для получения дубителей. Цветет рано, является хорошим медоносом.
Кустарники, свойственные этой группе лесов, входя в них как подлесок, в разных частях лесной зоны несколько различны. В европейской части России и Западной Сибири принимают участие в сложении лесных фитоценозов жимолость обыкновенная, крушина ломкая, волчье лыко. В Сибири и на Северном Урале довольно обычны кустарниковая ольха, бузина красная.
Травяной покров темнохвойного леса не очень разнообразен. Из характерных травянистых растений необходимо отметить папоротники из рода Dryopteris, плауны; в числе злаковых мятлик тенистый, луговик извилистый, среди двудольных - вороний глаз, ландыш, майник, копытень, звездчатку лесную, кислицу, грушанку, чернику, бруснику, веронику лечебную.
Экологические особенности этих растений во многом определены спецификой режима сильной затененности плотно сомкнутым пологом темнохвойного леса. Листовые пластинки большинства растений сильно развиты, темно-зеленого цвета, не имеют опушения, ориентированы горизонтально. Недостаток света привел к развитию сапрофитности или полусапрофитности, приспособленности к использованию перегнойных веществ подстилки или верхнего слоя почвы путем образования микоризы, что связано с недостаточ
205
ным обеспечением продуктами фотосинтеза. Некоторые растения целиком перешли на сапрофитный образ жизни и лишились хлорофилла, их листья превратились в желтоватые чешуйки, например, у кораллового корня, подъельника, веретляницы, гнездовки.
Испаряя мало влаги, представители травяного покрова ельников не имеют сильно развитых корневых систем. Их корни, слабо ветвясь, распределяются в самом верхнем горизонте почвы и подстилке.
Размножаются такие растения преимущественно вегетативно, поэтому в распределении напочвенного покрова хорошо выражена куртинность (копытень, кислица). Сильное затенение препятствует цветению, цветущие растения можно встретить только в «окнах» или участках разреженного древесного полога. К тому же под пологом темнохвойного леса очень мало насекомых-опылителей, особенно пчел, бабочек.
Если травяной покров этих лесов не отличается значительной густотой, то мхи часто образуют сплошные плотные «ковры». Под слоем живого мха нередко образуется довольно мощный (до 5-8 см) мертвый покров из остатков высших растений и самих мхов. Это создает условия дефицита кислорода в почве, благоприятствует анаэробным процессам с образованием подзолов. Такая ситуация особенно характерна для ельников.
СОСНОВЫЕ ЛЕСА
Сосновые леса это - светлохвойные насаждения с преобладанием сосны в древесном ярусе. Распространены в лесной и лесостепной зонах умеренного и субтропических поясов, а также в горно-лесном поясе. Растут на супесях, песках, известняках, доломитах, торфяных болотах.
Доледниковый возраст имеют сосняки на выходах мела, степные и лесостепные леса умеренного пояса. Субтропические сосновые леса растут в горах и на морских побережьях. В Евразии наибольшие площади заняты лесами из сосны обыкновенной. Среди таёжных сосняков выделяют сухие лишайниковые боры (беломошники), сосняки толокнянковые, зеленомошники (черничники, брусничники), сфагновые сосняки; для лесостепных и степных зон характерны сосняки разнотравные, остепнённые и степные боры.
По мнению В.Н. Сукачева сосновые леса сопутствуют человеку, то есть в большинстве случаев имеют антропогенное происхождение. Сосна обыкновенная как порода-пионер первой заселяет вырубки и гари, возникновение которых в большинстве случаев связано с деятельностью человека.
При естественном старении и распаде сосняков их место чаще всего занимают другие древесные растения. Под своим, хотя и довольно прозрачным, пологом сосна обыкновенная, будучи очень светолюбивой, не возобновляется. Однако после смены другими породами сосна полностью не исчезает. Она сохраняется в рефугиумах55, каковыми являются сфагновые болота и бедные сильно дренированные песчаные почвы, то есть те условия, в кото
Рефугиум - убежище.
206
рых другие древесные растения с сосной обыкновенной конкурировать не могут. Будучи крайне нетребовательной к влажности и богатству почв, сосна в подобных экстремальных условиях, превращаясь маленькие (до 1 м высоты) растения, способна давать семена и тем самым сохранять свой генофонд, "дожидаясь" очередной рубки или лесного пожара.
Вместе с тем, известны и настоящие коренные насаждения сосны обыкновенной. Примером могут служить Приобские ленточные боры, поселившиеся на песчаных дюнах среди Кулундийской степи в Алтайском крае и Казахстане.
Род сосна - Pinus L.
Род сосна включает около 100 видов, это самый большой род в семействе. Для многих народов северного полушария сосны представляют привычный компонент природы - растения, с которыми человечество знакомо с самых первых шагов своего развития. Всюду, где бы ни росли сосны, своеобразие их внешнего облика, удивительная способность при поранении ствола выделять ароматную смолу, выступающую крупными каплями, подобно слезам, запах просмоленной древесины издавна делали их, почти во всех областях естественного произрастания, объектом мифов, религиозных церемоний и поклонения.
Так, греческая мифология сохранила нам изящную легенду о нимфе Питие, которую бог ветра Борей, приревновав к лесному Пану, превратил в сосну. В Китае и Индокитае эти растения считались волшебными деревьями, приносящими счастье и отводящими беды. В Древнем Вьетнаме вера в магическую силу сосен была настолько сильна, что вошло в традицию сажать их у дворца императоров как символ долголетия и величия царствующей династии. Жители Центральной и Северной Европы тоже создали свои легенды и сказания о дереве добром, могучем и благостном, образующем светлые сухие леса - боры.
Сосны, за исключением немногочисленных карликовых или стелющихся жизненных форм, - стройные вечнозеленые деревья, достигающие в высоту 50 или даже 75 м и в диаметре до 2-4 м (сосны желтая и Ламберта).
Род Pinus, в отличие от прочих хвойных, характеризуется тем, что хвоя у них сидит только пучками по 2-5 и окружена влагалищем из видоизмененной хвои, что вместе и составляет укороченный побег, который помещается в пазухе маленького бесхлорофилльного чешуевидного листочка, часто отпадающего или малозаметного. Как эти чешуйчатые листочки, так, следовательно, и пучки хвои сидят спирально на ветвях. Мужские стробилы скучены вокруг развивающегося нового побега.
Пыльцевые зерна снабжены двумя воздухоносными мешками. Женские стробилы, в виде шишечек, в момент цветения кажутся сидящими на концах молодых побегов и состоят из кроющих чешуй, в пазухах которых сидят семенные чешуи, несущие по две семяпочки. Опыление происходит при помощи ветра. После опыления семенные чешуи разрастаются и делаются более или менее деревянистыми, кроющие же чешуи остаются маленькими и во
207
взрослой шишке малозаметными или почти незаметными. Шишки созревают на второй или даже на третий год. Чешуи шишки на конце расширены и часто утолщены, образуя так называемый щиток или апофиз, по середине которого или на конце его сидит „пупок", имеющий вид бугорка или впадины. Семена крылатые или бескрылые. Всходы несут всегда несколько (4-18) семядолей. Этими морфологическими признаками характеризуется и рассмотренная нами ранее сосна сибирская кедровая.
Сосна обыкновенная - Р. sylvestris L.
Сосна обыкновенная представляет собою мощное дерево первой величины с характерной кроной. Сосна растет вначале очень быстро, уступая среди прочих наших хвойных лишь лиственнице. Максимальный прирост у нее на лучших почвах наблюдается в 15-20 лет, а на худших - в 25 лет. Лет за 40-50 прирост значительно падает, и ель, всегда отстающая от сосны в первые десятилетия своей жизни, на лучших почвах догоняет сосну к 60-70 годам. Обычно высота на хороших почвах порядка 30-35 м. Доживает до 300-350 лет, хотя известны сосны, прожившие и дольше. Ствол отличается правильным мутовчатым ветвлением, так что в молодом возрасте (примерно до 40-50 лет), пока нижние ветви еще не окончательно сброшены, по числу мутовок можно определить возраст дерева. Нарастание побега моноподиаль-ное. Хвоя сидит по две в пучках, которые располагаются по поверхности побега спирально, и направлена радиально во все стороны, длиной 20-60 мм, плоско выпуклая, жесткая, узко-линейная, остроконечная по краю мелкопильчатая. Устьица располагаются рядами на обеих сторонах - плоской и выпуклой. На взрослых деревьях хвоя обыкновенно держится 2-3 года. Почки остро-яйцевидные, большей частью засмоленные (рис.60).
Рис. 60. Сосна обыкновенная: 1 - собрание мужских стробилов; 2 - микроспорофилл сбоку; 3 - микроспорофилл со стороны микроспорангиев (пыльников); 4 - женские стробилы; 5 - семенная чешуя с двумя семязачатками; 6 - семенная чешуя с морфологически нижней стороны; видна пленчатая кроющая чешуя, по размеру она меньше семенной; 7 - однолетняя незрелая шишка (озимь); 8 -семенная чешуя с двумя семенами из зрелой шишки; 9 - крыловидный придаток семени, видна охватывающая семя "вилочка"; 10 - семя; 11 - семя в разрезе с зародышем внутри (из Вольф, 1925)
Кора у сосны различного вида на разных частях дерева. Молодые ветви покрыты сероватой или серовато-бурой корой. Более старые ветви и верхняя
208
часть ствола несет желто-красную кору, отслаивающуюся тонкими пленками. Нижние же, более толстые части ствола покрыты толстой корою, сильно бороздчатой и отделяющей красновато-темно-бурую корку.
Сосна начинает плодоносить на свободе уже в 15 лет, но в насаждении обильное плодоношение наблюдается лишь в возрасте за 40 лет. Плодоносит обильно. Семенные годы обычно наступают через 3-5 лет.
В центре европейской России сосна начинает опыление в мае, в первой половине июня, одновременно с появлением новых побегов. После этого края семенных чешуй срастаются, и оплодотворение происходит лишь летом следующего года. Опыление осуществляется ветром. Так как мужских стробилов рассеяно по всей кроне очень много, то пыльца образуется в обилии. Там, где много растет сосны, в воздухе носится масса пыльцы, которая, падая, часто покрывает землю или поверхность водоемов тонким желтым слоем. В течение второго лета своего развития шишка значительно увеличивается, и к ноябрю в шишке созревают семена. Таким образом, от момента опыления и до созревания семян сосне нужно около 18 месяцев. Однако выпадение семян из шишки начинается лишь в конце зимы - начале весны (февраль - апрель), когда шишки раскрываются.
Чешуи зрелой шишки очень твердые, несут сильно утолщенный апофиз (щиток). Семя 3-4 мм длины, яйцевидное или продолговато-яйцевидное, вытянутое в притупленный кончик, весьма различной окраски: то почти белое, то черное, то пестрое, с одной стороны слегка блестящее. Эта изменчивость касается разных деревьев, у одного дерева все семена по цвету одинаковые. Семя несет крылышко, которое охватывает его с двух сторон своим основанием, как щипчиками. Крылышко длиннее семени в 3-4 раза, пленчатое, буроватое, со скошенной верхушкой. Прорастают семена обыкновенно через 15-20 дней. Всхожесть свежих семян обычно велика, до 90 °/о, причем она сохраняется до 3 и более лет.
Всходы несут 5-7 трехгранных, дугообразно согнутых семядолей. Уже в течение первого года развивается побег, достигающий длины 4-10 см. На нем, в противоположность тому, что наблюдается на взрослых деревьях, хвоя сидит поодиночке, она значительно тоньше и короче, чем нормальная хвоя. На второй год развивающийся побег сначала также несет одиночные хвоинки, в пазухах которых несколько позже вырастают укороченные побеги, несущие уже нормальную пару хвои. В следующие годы хвоя развивается только на укороченных побегах по две хвоинки. Ветвиться сеянец начинает обычно на третий год, но иногда бывает, что в первый год закладываются боковые почки.
Сосна обыкновенная заходит далеко на север и на восток, занимая огромные пространства в Европе и Азии, от Ирландии и почти до Тихого океана (см. рис. 57). Она растет в равнинных и горных условиях, но выше 800 м в горы обычно не поднимается.
В указанных на рисунке границах, особенно на юге, сосна обыкновенная встречается лишь в виде островных массивов. Поэтому южной границей сплошного ареала сосны следует считать линию Киев - Чернигов - Карачев
209
(Брянская область), затем Тула - Серпухов - Рязань, нижнее побережье рек Камы и Белой, Челябинск. В Сибири сплошной ареал ее ограничивается северной границей степей. Кроме того, сосна обыкновенная растет в горах Крыма, на Кавказе от Кубани до Дагестана и в северной части Грузии.
Сосна обыкновенная может переносить как суровый климат севера, так и жаркий климат степей; она не боится заморозков и первой появляется на вырубках, если этому не препятствует травяной покров. Сосна обыкновенная светолюбива, о чем говорят ее ажурная крона, наличие световой хвои, плохой рост затененного подроста, значительное самоизреживание насаждений, хорошая очищаемость от сучьев и другие особенности. На более плодородных почвах и в более южных условиях она становится менее требовательной к свету.
К почве и к почвенной влаге сосна обыкновенная не требовательна, является ксерофитом. Об этом свидетельствуют малая испаряющая поверхность и толстая кутикула у листьев, а также погруженные в ткань листа устьица. Сосна переносит физическую сухость и произрастает на очень сухих песчаных дюнах. В то же время она растет также на верховых, сфагновых болотах.
Корневая система сосны обыкновенной отличается большой пластичностью. На глубоких свежих песчаных почвах она развивает мощный стержневой и много вертикальных (до 1,5 м длиной) корней (см. рис. 42), обычно с глубины 60 см начинающих сильно ветвиться и идущих до грунтовых вод. В верхних горизонтах почвы на глубине до 25 см залегает несколько горизонтальных корней, уходящих далеко за пределы проекции кроны. Общая их длина в 10-15 раз превышает радиус кроны. Они улавливают атмосферную влагу. На очень бедных песчаных почвах при коротком стержневом корне более характерны длинные шнуровидные корни, идущие вблизи поверхности почвы. Но обычно у сосны преобладает скелетная система корней. Относительно малое количество корневых мочек и волосков восполняется наличием микоризы. Чехлы микоризы в виде разветвленных грибных нитей, оплетающих всасывающие окончания корней сосны, являются замечательным приспособлением, обеспечивающим дополнительное питание.
На суглинистых почвах корни сосны обычно недалеко выходят за пределы проекции кроны. На болотах корневая система ее поверхностная. Растет сосна в этих условиях плохо, не достигает больших размеров. При наличии нескольких водоносных горизонтов корневая система становится многоярусной.
У однолетних сосенок обычно развивается стержневой корень, в 3-4 раза превышающий по длине стебель. При нормальных условиях он растет вглубь до 40 лет, затем в основном развивает только боковую корневую систему.
Сосна обыкновенная способна формировать производительные древостои. В высокополнотных насаждениях запас древесины на гектаре иногда превышает 600 м3. Однако такие насаждения относительно редки. Чаще сосняки занимают свежие песчаные почвы. В более сухих местах здесь в по
210
крове преобладает брусника, а на несколько пониженных плато, на влажных почвах со следами оглеения, преобладание переходит к чернике. Сосна достигает в этих условиях в среднем 23-25 м высоты (II бонитет).
Хуже растет сосна на сухих дюнах и на застойно-сырых почвах. На верховых сфагновых болотах с мощностью торфяного слоя около 1 м она едва достигает высоты 10 м (V бонитет), накопляя запас около 60-100 м3.
В пределах обширного ареала сосна обыкновенная встречается с другими древесными породами, в том числе с елью, березой и широколиственными породами. На более или менее плодородных и не сильно заболоченных почвах в лесной зоне сосна вытесняется елью. Поэтому насаждения сосны обычно тяготеют к песчаным почвам. Действие пожаров сказывается на этих породах по-разному. Спелая сосна относительно легко переносит низовой пожар и после него обсеменяет гари, ель страдает значительно сильнее и уступает на пожарищах место сосне. Более сильно страдает ель также и от засух.
Сосна обыкновенная имеет большое хозяйственное значение в нашей стране. Это одна из основных лесообразующих пород в лесной зоне. Она имеет также большое водоохранное и защитное значение.
Большую ценность представляет древесина сосны, отличающаяся высокими техническими качествами и прочностью. Она имеет красноватое ядро и желтовато белую заболонь. Годичные кольца в древесине видны хорошо, так как граница между весенней (ранней) частью слоя и поздней ярко очерчена. Древесина богата смоляными ходами.
Анатомические особенности древесины. Поперечный срез. Ядро хорошо выражено. Граница между годичными кольцами отчетливая. Переход от ранней древесины к поздней иногда плавный. Толщина оболочек поздних трахеид почти вдвое больше, чем ранних. Смоляные ходы выстланы крупными тонкостенными эпителиальными клетками. Тангенциальный срез. Лучи в основном однорядные, встречаются 2 (4)-рядные лучи со смоляными ходами. Высота лучей 2-15 (20) клеток. Радиальный срез. Окаймленные поры на стенках трахеид расположены преимущественно в один, иногда в два, ряда. Имеются лучевые трахеиды. Горизонтальные стенки лучевых трахеид зубчатые. На полях перекреста в ранней древесине в основном оконцевые поры, иногда встречаются 2-3 крупных пи-ноидных поры в одном поле перекреста.
Древесина сосны широко используется в виде круглого леса - столбы, бревна, рудничная стойка; пиломатериалов - балки, брусья, доски, тес; колотого леса и щепных товаров. Она применяется в судостроении, вагоностроении, мебельном производстве, для заготовки шпал и пр.
Широко была развита у нас подсочка сосны. Из получающейся при этом живицы добывают канифоль и скипидар. Смолье и пни используют для получения скипидара и дегтя. В коре сосны содержится небольшое количество дубильных веществ и гумми. Из хвои добывают русское сосновое масло и витамин С, используемые в медицине. Сосна выделяет фитонциды, очищающие воздух от вредных микробов.
В сосновых борах находят пищу различные представители лесной фауны - белки, глухари и др., производится сбор грибов и лекарственных растений.
211
Большое значение имеет сосна также в зеленом строительстве. Зеленое кольцо вокруг Москвы с юга и востока в основном образовано сосновыми массивами. Известны пригородные сосновые рощи у многих других крупных городов. Сосняки окружают санатории и курорты. Сосна входит как одна из основных пород в большинство ландшафтных парков.
Имея обширный ареал с различными условиями среды, сосна обыкновенная в разных регионах представлена несколькими, хорошо обособляющимися морфологически, формами. Некоторые формы рассматриваются как географические расы, подвиды, трактующиеся отдельными авторами как самостоятельные виды. Из таких, ярко выраженных, географических форм следует выделить сосну лапландскую, сосну кавказскую (крючковатую), сосну кулундийскую. Последняя образует верхнеобьские ленточные боры преимущественно на территории Алтайского края.
Типы сосновых насаждений
Сосновые леса играют существенную роль в сложении растительного покрова на всем протяжении лесной зоны. Благодаря способности расти на различных почвах сосна формирует большое число типов леса. В отличие от еловых типов, сосновые в основном приурочены к песчаным и супесчаным почвам, встречаясь, кроме того, на торфянистых и редко на суглинистых и даже глинистых. Типы сосновых лесов делятся на несколько основных групп.
Боры лишайниковые. Рельеф холмистый, чаще всего в виде высоких или пологих дюнных всхолмлений. Все они характеризуются сухими и бедными почвами, сплошным лишайниковым покровом преимущественно из видов Claddnia (олений мох). Травяной напочвенный покров редкий и невысокий. Подлесок отсутствует или слабо развит. Верхний полог из одной сосны низких бонитетов (III—IV). Стволы часто корявые, плохо очищающиеся от сучьев. Эта группа представлена несколькими типами, меняющимися в зависимости от степени увлажнения почвы.
Боры-зеленомошники. Типы леса этой группы в дюнных районах занимают чаще всего склоны или неглубокие котловины между дюнами. В других рельефных разностях - более или менее равнинные места. Почвы по сравнению с предыдущей группой влажнее и несколько богаче, но все-таки весьма бедные. Для всей группы типов леса характерны сплошной моховой покров из так называемых зеленых мхов (роды Pleurozium, Hylocomium, Rhytidiadelphus, Ptilium и др.) и негустой травяной покров - грушанки, плауны. Подлесок практически отсутствует. Древостой чисто сосновый, иногда с единичной примесью березы. В зависимости от степени увлажнения почвы здесь также выделяется несколько типов леса. Наиболее важные из них бор-брусничник, бор-кисличник и бор-черничник. Первый занимает довольно сухие и бедные почвы, для второго характерны почвы более богатые, для черничника - почвы влажные, мало проточные. Наиболее высокий бонитет в этом ряду у бора-кисличника.
212
Рис. 61. Сосняк брусничник, II бонитета. Макарьевский лесхоз Костромской области (фото В. Коровина)
Сосняки-долгомошники. Эта группа типов сосняков характеризуется заметным заболачиванием, сплошным покровом из мха - кукушкина льна (Politrichum sp.) и низкой производительностью древостоев.
Сосняки сфагновые. Для них характерна далеко зашедшая стадия заболачивания. Сплошной сфагновый покров. Рост сосны плохой. Низкий бонитет.
Травяно-болотные боры. Сосняки, характеризующиеся значительной влажностью почвы и повышенным по сравнению с ранее описанными группами типов леса плодородием почвы. Это довольно широко распространенные типы леса. Данная группа типов весьма неоднородна и по существу представляет собой значительное разнообразие типов леса. Бонитет Ш-П, густой травяной напочвенный покров, слабое развитие мохового покрова, редкий или отсутствующий подлесок.
Сложные боры. Характеризуются богатством почвы и растительности. Древостой состоит из сосны, под которой развивается подлесок и часто второй ярус из дуба, иногда встречается примесь и других широколиственных пород. Подлесок хорошо развит. Травяной покров обильный и разнообразный, мхи довольно редки. Сложные боры представлены множеством типов леса. Рассмотрим наиболее распространенные.
Сосняк липовый. Этот тип характеризуется липой в подлеске, но не образующей второго яруса. Из рассматриваемых типов в этой группе, данный тип приурочен к наиболее бедным почвам, но все же они богаче, чем в предыдущих группах. Сосна растет хорошо, давая высокий бонитет.
213
Сосняк лещинный. Этот тип приурочен к лучшим почвам, чем предыдущий, отличается обычно густым подлеском из лещины, и рост сосны еще лучше, чем в сосняке липовом.
Сосняк дубовый. Этот тип, обычно приуроченный к наилучшим почвам, возможным в сосновых лесах, характеризуется тем, что под сосной развивается второй ярус из дуба, который, однако, обычно плохого вида и дает неровные низкие стволы. Подлесок из лещины, бересклета и других кустарников, обыкновенно хорошо развит. Рост сосны, можно сказать, самый лучший.
Значительный интерес представляет взаимоотношения ели и сосны, проследим за ними, следуя тексту В.Н. Сукачева (см. список литературы). По многочисленным наблюдениям, существует явно выраженная тенденция смены сосны елью в условиях произрастания благоприятных для того и другого видов. В виду важности данного вопроса не только с точки зрения общего понимания взаимоотношений между этими главнейшими породами, но и с точки зрения лесохозяйственного интереса, остановимся на этом вопросе несколько подробнее.
Наблюдая в природе сосновый лес, особенно сосновый лес с моховым покровом, можно часто видеть появление елового молодняка под пологом сосны. То мы видим его в виде единичных деревьев, то он образует под сосной самостоятельный полог разной высоты. В тех случаях, когда ель образует более или менее сомкнутый ярус, как общее правило, наблюдается отсутствие соснового подроста. Самые тщательные поиски не обнаруживают сосновых всходов или, если они изредка и встречаются, то вскоре гибнут.
Подобное явление станет для нас понятным, если мы вспомним отношение сосны и ели к свету. Сосна - порода более светолюбивая, чем ель. Поэтому еловый подрост под сосновым пологом, при наличности налета еловых семян, может появиться и успешно развиваться. Напротив, как только этот еловый подрост сомкнется и начнет затенять почву, сразу прекращается возможность возобновления сосны, даже при прочих благоприятных для этого условиях.
Следствием этого будет то, что в то время как сосновый ярус, достигнув предельного возраста, начинает умирать и вываливаться ветром, ему на смену придет не новое поколение сосен, а сплошной еловый ярус. Таким образом произойдет смена ассоциаций соснового леса на еловый. Эта смена произойдет как без изменения общих климатических условий, так и без видимого изменения почвы. По существу, конечно, изменение почвы произойдет, но уже не как причина, а как следствие смены сосняков ельниками. Все дело в том, что здесь сталкиваются две древесные породы с разными экологическими особенностями. Сосна не имеет возможности помешать вторжению ели под свой полог; скорее, можно думать, что ель под нею находит более хорошие условия. Но, появившись, ель так меняет климат ассоциации, главным образом условия освещения, что они делаются неблагоприятными для сосны, тогда как ель с ними вполне может мириться. Этим и определяется в дальнейшем господство ели за счет сосны.
214
Однако тот факт, что мы в природе наблюдаем не только разной высоты еловые пологи в сосновом лесу, но и в одном верхнем ярусе сосну и ель вместе в смеси, как бы говорит в пользу того, что сосна и ель могут устойчиво сочетаться и в одном ярусе, и в виде соснового леса с еловым подлеском, то есть что разные комбинации сосны и ели в ценозах представляют устойчивые явления. Как бы в подтверждение этого при выраженном всхолмленном ландшафте, часто на более возвышенных участках, наблюдаются одни чисто сосновые ценозы, на склонах, - сосновые фитоценозы с еловым низким или высоким пологом и, наконец, по еще более низким местам, - сосна и ель растут совместно в одном верхнем пологе.
Но если бы эта точка зрения была верна, то мы в природе часто встречали бы случаи, когда в сосновых лесах со сплошными еловыми ярусами, последние, достигнув известной высоты, прекращают рост и начинают отмирать и засыхать. Этого, однако, не наблюдается. Раз почвенные условия таковы, что позволили ели образовать сомкнутый полог, то дальнейшее развитие ели обеспечено и, напротив, прекращено возобновление сосны.
Пусть даже почвенные условия более благоприятны для сосны, нежели для ели, пусть в этих условиях сосна дает стволы особенно хорошего роста и полнодревесные, а ель растет посредственно или даже ниже среднего, тем не менее, раз ель могла образовать сомкнутый полог, вытеснение елью сосны в дальнейшем обеспечено, так как прекращается возобновление последней. А в тех сосновых лесах, где почвы сухие, и ель не образует сплошного полога, а растет единичными ослабленными растениями или вовсе отсутствует, там смена сосны елью совершенно невозможна.
На более влажных и вообще более благоприятных почвах процесс вытеснения сосны елью идет, разумеется, быстрее, нежели на почвах сравнительно сухих. Сосновая ассоциация с низкой елью, с елью средней высоты и с елью в смеси в верхнем пологе - это все разные стадии одного процесса, именно - смена сосновых ассоциаций еловыми. Это сочетание показывает также, что ель не занимает всех тех мест внутри своей области распространения, на которых по условиям почвы она может произрастать.
Здесь, однако, нельзя обойти молчанием одного обстоятельства, которое как бы противоречит вышесказанному положению. Из только что описанной картины смены сосновых ассоциаций еловыми следует, что этот процесс, в большинстве случаев, идет столь быстро, что в каких-нибудь два, максимум - три столетия он может закончиться, то есть сосновые ассоциации окончательно могут смениться еловыми. Поэтому возникает естественно вопрос: как могли дожить до нас эти различные комбинации сосны и ели в одном ценозе, почему эта смена не закончилась уже давно, до нашего времени, что помешало ели заселить все возможные для ее существования места? Ведь со времени появления ели в северной половине европейской части России прошел не малый промежуток времени, измеряемый тысячами лет, ведь остатки ели известны, например, в Ленинградской области на дне самых глубоких торфяников.
215
Чтобы это стало понятным, необходимо вспомнить, что уже прошло не одно столетие, как в жизнь природы вмешался человек. Если мы примем во внимание этот фактор, то будем в состоянии объяснить и кажущееся здесь противоречие. Скандинавские, финские и русские ученые, выяснили, что причиной, которая удержала сосну на многих таких местах, где она могла бы быть вытеснена елью, являются пожары. Дело в том, что сосна и ель относятся совершенно различно к пожарам. Низовой пожар, который не охватил самих деревьев, а прошел бегло и уничтожил сухую лесную подстилку, сухие лишайники, мхи и валежник, пройдет почти бесследно для сосны, не только старой, но и для более или менее взрослого соснового подроста, благодаря, по-видимому, более глубоко идущей корневой системе сосны. Ель, особенно же еловый подрост с поверхностной корневой системой, убивается часто даже легким низовым пожаром. Таким образом, в борьбе ели с сосной последняя находит верного союзника в лице пожаров. Сосново-еловый фитоценоз или сосновый с еловым вторым ярусом или подлеском из ели после пожара может утратить ель и остаться только сосновым. Все данные говорят, что лесные пожары, будучи характернейшим явлением там, где человеческая культура стоит низко, имели место и в докультурное время. Сейчас в Сибири можно проехать сотни километров и не найти гектара, где бы на коре деревьев не было заметно следов пожара.
Однако причиной пожаров может быть не только человек, вызывающий его сознательно или бессознательно, но и молния, поэтому во все времена деятельность огня могла сильно задерживать или полностью останавливать процесс расселения ели, выражающийся на нашей территории главным образом в вытеснении сосны.
ЛИСТВЕННИЧНЫЕ ЛЕСА
Третьим крупным объединением наших хвойных лесов являются лиственничные леса, лиственничники, называемые также „листвягами". По территории, которую они занимают в Сибири, они должны быть поставлены на первое место среди лесов всех других пород в России. Однако, простираясь, главным образом, в областях, пока еще очень мало освоенных, они остаются недостаточно изученными. В общем, можно сказать, что роль лиственницы, как лесообразователя, по мере перехода за Урал и движения далее к востоку, увеличивается, достигая наибольшего значения в этом отношении в Якутии. В нашей стране растет дико несколько видов лиственницы, из которых два имеют огромные ареалы и большое лесохозяйственное значение, а именно -сибирская и даурская лиственницы. Остальные представлены в очень небольших областях распространения и особого хозяйственного значения пока не имеют. Кроме того, часто разводится в европейской части России европейская лиственница, которая по морфологическим и по лесоводственным признакам более близка к сибирской, чем даурской.
216
Род лиственница - Larix Mill.
Род лиственница объединяет виды (около 20), представленные крупными деревьями с не ясно мутовчатым ветвлением и мягкой однолетней хвоей, сидящей пучками на укороченных побегах и поодиночке на удлиненных. Лиственницы однодомны. Опыляются ветром. Шишки небольшие, созревают в тот же год осенью. Они состоят из широких семенных и едва заметных кроющих чешуй. Семена имеют плотно приросшие крылышки. Всходы с 5-6 (10) гладкими семядолями. Древесина с ядром; как и кора, имеет смоляные ходы.
Лиственница относится к очень древнему роду хвойных: она появилась примерно в конце мелового периода. Есть предположение, что не позже середины третичного периода лиственница из Центрального Китая проникла через Алтай в Западную Сибирь и Европу. В связи с различными условиями произрастания она распалась на три отдельных вида. Ледники оттеснили лиственницу на восток, юг и запад. На юге она была вытеснена другими древесными породами, на западе обособился вид лиственницы европейской, в Сибири - сибирской. По мнению некоторых, преимущественно отечественных, ученых лиственница сибирская, заняв европейский северо-восток, образовала самостоятельный вид, выделенный и описанный известным ученым-лесоводом Н. В. Дылисом, - лиственницу Сукачева (рис. 62).
Рис. 62. Ареалы видов лиственницы
Согласно другой, принятой систематиками, точке зрения лиственница Сукачева не возводится в ранг вида, а рассматривается как географическая раса лиственницы сибирской. Морфологически лиственница Сукачева практически не отличается от лиственницы сибирской. Некоторые различия носят
217
в основном статистический характер и заключаются в большем количестве семенных чешуй у шишки, более длинной хвое, более толстой коре в комлевой части ствола. У лиственницы Сукачева шишки, семенные чешуи и семена крупнее, раскрывшиеся шишки принимают почти шаровидную форму. Кроме того, по мнению Н.В. Дылиса, лиственница Сукачева характеризуется меньшей морфологической изменчивостью, нежели лиственница сибирская.
В нашем изложении лиственница сибирская, в соответствии с позицией систематиков, также не будет дифференцироваться на два вида.
Из особой тихоокеанской ветви, идущей из Китая, за Байкалом обособился вид лиственницы даурской, и возникли японо-американские виды.
Лиственница сибирская - L. Sibirica Ledeb.
Семена лиственницы сибирской косо-обратнояйцевидные, длиной 4-6 мм, твердые, серовато- или желтовато-светлобурые, в верхней половине с буроватыми пятнышками; крылышко 4-5 мм длиной, ланцетовидное или по-луяйцевидное, с одной стороны - прямое, с другой - закругленное; вместе с семенем длина 12-13 мм, плотно прикреплено к верхней части семени и к самому краю его нижней стороны, от него не отделяется (рис. 63). Всхожесть семян сибирской лиственницы обычно не очень велика и быстро падает. Всходы имеют 5-10, чаще 5-6 семядолей. В первый год вырастает побег с одиночной спирально расположенной хвоей, в пазухах которой местами закладываются почки. К осени семядоли буреют и отмирают, точно так же, как и часть первичной хвои. Однако значительная часть хвои перезимовывает до следующего года. В этом нельзя не видеть следствия того, что лиственница произошла из вечнозеленых хвойных и на первых стадиях своего развития сохраняет на зиму свою хвою, а в более взрослом состоянии эту способность уже теряет. Первичная хвоя гладкая синевато-зеленая. На следующий год из почек развиваются укороченные побеги, несущие хвою пучками. Дальнейшее развитие идет быстро, и лиственница в первые годы растет быстрее прочих наших хвойных пород. Из части боковых почек развиваются удлиненные побеги, а остальная часть дает укороченные побеги. Такой укороченный побег живет до 10-12 лет, а потом отмирает. Нередко укороченный побег, после нескольких лет роста, дает удлиненный побег. Так как особенной приуроченности почек к концу одногодичного побега не наблюдается, то ветвление не носит мутовчатого характера. Границы годичных побегов обычно обозначаются резко, благодаря темным кольцам рубцов от отвалившихся почечных чешуй.
Растет лиственница первые годы быстро, максимум прироста между 30-40 годами, когда ее прирост в отдельные годы достигает 1 м в год. На лучших почвах она в 100-летнем возрасте достигает 30-33 м высоты, а в 200 лет - до 41 м. Крайним пределом продолжительности жизни сибирской лиственницы считается 350 лет; к этому возрасту она достигает 45 м высоты и 1,75 мв диаметре на высоте груди, утолщаясь значительно к комлю.
218
Рис. 63: Морфологические признаки видов лиственницы: 1 - лиственница сибирская: 1а - удлиненный побег текущего года, видны "ребра" и "листовые подушечки; 16 -фрагмент двулетней ветви с укороченными побегами; 1 в - ветвь состоящая из однолетнего и двулетнего удлиненных побегов (хвоя на однолетнем побеге одиночная, на приросте второго года расположена пучками на укороченных побегах); 1г -хвоя; 1д - ветвь с мужскими и женскими стробилами на укороченных побегах; 1е - мегастробил в пазухе кроющей чешуи; 1ж - микроспорофилл; 1з - поперечный разрез хвои; 1и - зрелая шишка; 1к - семенная и кроющая чешуи зрелой шишки; 1л - семенная чешуя с семенами. 1м - семя. Зрелые шишки лиственниц: 2 - лиственница европейская; 3 - л. Сукачева; 4 - л. даурская; 5 - л. японская.
Обозначения: вп - верхушечная почка; би - боковая почка; ли - листовая "подушечка"; р - "ребра"; уп - укороченный побег; кч - кроющая чешуя; сч - семенная чешуя; сз семязачатки
219
В течение всей жизни лиственницы общая форма ее кроны яйцевидноконическая, причем в молодости - более узкая, к старости - широкая, с более тупой вершиной. Однако, такою притупленной, как у сосны, она никогда не бывает. Ветви отходят под углом, более или менее приближающимся к прямому; при этом более старые характерно изгибаются: будучи сначала немного направлены вверх, они затем отгибаются книзу, а потом снова несколько приподнимаются кверху. На худшей почве крона формируется более узкой.
Кора на молодых побегах светло-серо-буроватая, потом темнеет и сереет, к старости делается очень толстой, глубоко-бороздчатой, отделяющейся значительными кусками. На старых деревьях в нижней части ствола кора достигает иногда свыше 10 см толщины. Ствол обыкновенно прямой. Корневая система сильно развита, имеет стержневой корень и значительно развитые боковые. Укрепляется прочно и неветровальна, по крайней мере, если растет на достаточно сухих почвах; на сильно влажных корневая система лиственницы делается более поверхностной, и последняя более страдает от ветровала. Поверхностной корневая система у лиственницы бывает также на местах, где близко к поверхности залегает вечная мерзлота. Побеги первого года светло-желтовато-зеленые, блестящие, голые, лишь кое-где, по преимуществу близ листовых рубцов, с редкими темноватыми, длинными волосками, бороздчатые от продолжающихся по стеблю вниз оснований листьев. Почки желтовато- или красновато-бурые, овально-полушаровидной формы, без смолы или с очень небольшим ее количеством, с чешуями по краю, более или менее реснитчатыми.
Хвоя мягкая, узко-линейная, к вершине немного расширенная, на вершине несколько туповатая, ярко зеленая с сизоватым налетом, на удлиненных побегах одиночная, прямая или слегка согнутая, на укороченных побегах сидит пучками, чаще по 30-50 штук неравной длины. В разрезе овальная, с более выпуклой верхней стороной и более плоской, но с несколько выдающимся килем снизу. Хвоя в длине колеблется очень значительно (от 12 до 45 мм), а на быстрорастущих длинных верхушечных побегах достигает даже свыше 10 см. Наиболее обычной, типичной длиной хвои надо считать 30-35 мм. Ширина хвои тоже сильно колеблется, начиная от 0,6 мм и до 1,0 мм, но на очень сильных верхушечных побегах она достигает ширины до 2,5 мм.
Устьица расположены рядами на обеих сторонах, причем на верхней стороне их обычно 3-4, а на нижней 4-6 с каждой стороны жилки; часто они прерываются.
Древесина лиственницы с ясно выраженным красноватым ядром и узкой светлой заболонью. В отличие от древесины сосны, ели и пихты у лиственницы древесина твердая, высокопрочная, смолистая - устойчивая к гнили. (Дворцы и дома Венеции стоят на лиственничных сваях). Служит прекрасным материалом для сооружений во влажных местах и под водою (шпалы, при кораблестроении и т. п.). Используется для срубов, изготовления пиломатериалов, несущих деревянных конструкций, для получения целлюлозы, спирта.
220
Анатомические особенности древесины лиственницы. Поперечный срез .Ядро хорошо выражено. Граница между годичными слоями отчетливая. В большинстве случаев резкий переход от ранней древесины к поздней. Осевая паренхима отсутствует. Смоляные ходы выстланы эпителиальными клетками со стенками средней толщины числом 8-14. Тангенциальный срез. Высота лучей 2-25 (30) клеток. Радиальные смоляные ходы выстланы эпителиальными клетками числом 7-15. Расположение смоляных ходов в лучах эксцентрическое. Радиальный срез. Лучи сложные. Стенки лучевых трахеид гладкие и волнистые. Стенки клеток лучей толстые, тангенциальные стенки узелковые. На полях перекреста в ранней древесине поры пицеоидные. Окаймленные поры на стенках ранних трахеид расположены часто в два ряда. Имеются крассулы.
Описанные особенности анатомического строения свойственны и лиственнице даурской.
Лиственница хорошо выдерживает загрязненный воздух городов, вероятно, из-за листопадности, поэтому успешно используется в озелени населенных пунктов.
Лиственница сибирская вступает в репродуктивную фазу при свободном стоянии в возрасте около 15 лет, иногда и ранее, в сомкнутом насаждении - около 20-30 лет. Стробилы появляются одновременно с началом распускания листьев. Мужские стробилы имеют вид небольших желтых полу-шарообразных, длиной 5-6 мм, шишечек, сидящих на укороченных побегах. Женские стробилы в виде шишек расположены по всей периферии кроны. Кроющие чешуи отогнуты наружу и вниз, оканчиваются длинным остроко-нечием. Семенные чешуи округло-подкововидной формы, значительно короче кроющих.
Мужские и женские репродуктивные органы размещаются по всей кроне и сидят обычно в соседстве на ветвях; первых всегда значительно больше, чем вторых. Пыльца переносится ветром, но воздушных мешков, как у ели и сосны, нет. Оплодотворение происходит в то же лето, когда лиственница цветет. Обычно уже в сентябре шишки настолько зрелы, что семена, извлеченные из них, всхожи. Если в момент «цветения» кроющие чешуи значительно превышают семенные, то к моменту созревания шишек соотношение меняется. По мере развития шишки кроющие чешуи почти не увеличиваются в размере, тогда как семенные разрастаются, и в зрелой шишке превышают кроющие. В это время семенные чешуи деревянистые, серо-коричневые, округлые или округло-яйцевидные, цельнокрайние, на верхушке обычно закругленные. Высота шишек колеблется в пределах 2,5-4,0 см.
Шишка раскрывается обычно в марте, причем чешуи раздвигаются довольно широко и семена из них легко высыпаются. Так как крылышко не отделяется легко от семени, то обыкновенно они и сеются с крылышками или их обломками. После выпадения семян шишка продолжает висеть на дереве 1-3 года.
Вегетативно не размножается. Сибирской лиственнице свойственно давать придаточные корни, которые появляются на нижней части стволиков при покрытии их землею или при обрастании мхом, если она растет на болоте.
221
Сибирская лиственница занимает обширный ареал, охватывающий северо-восток европейской части России и западную половину Сибири. В пределах европейской части России область ее распространения имеет прерывистый характер. В Сибири восточная граница ареала проходит по междуречью Енисея и Лены и через северную часть Байкала уходит в Монголию.
Южная граница от нижнего течения реки Онеги направляется севернее гор. Вологды к верхнему течению р. Унжи, затем по р. Унже, не доходя до ее устья, через нижние части р. Ветлуги и Вятки, пересекает р. Каму севернее устья р. Белой, далее направляется к верховьям р. Белой на Урале, по которой спускается до верховьев р. Урала, до 51° с. ш.. Отсюда поднимается на север, проходя далее несколько севернее г. Тюмени, идет к западным предгорьям Алтая, далее по хребтам Сауру и Тарбагатаю, где под 47° с. ш. имеет самые южные местонахождения (рис. 62).
Такой большой ареал сибирской лиственницы, простирающийся с севера на юг почти на 23°, свидетельствует об ее экологической пластичности. Перенося на севере, на границе с тундрой, весьма низкие температуры, она на юге растет при исключительно высоких температурах, спускаясь с гор почти до сухой и жаркой полупустыни. Ни одна из наших древесных пород не имеет столь разнообразных климатических условий на протяжении своего ареала, как сибирская и даурская лиственницы. Способность переносить крайности зимнего периода стоит в связи со сбрасыванием хвои на зиму. От заморозков мало страдает, если не считать нередко бывающего повреждения весною ее вегетативных органов. Если принять во внимание, что количество осадков, относительная влажность воздуха, сила ветра и другие климатические факторы в пределах ареала лиственницы сильно колеблются, то надо признать, что она является одной из самых непритязательных к климату наших древесных пород. Успех ее разведения к западу, например, в южной Финляндии, и к югу, например, в защитных насаждениях Саратовской области, от ее естественного ареала показывает, что она находит для себя подходящими климатические условия и за пределами последнего.
Если сибирскую лиственницу надо считать малотребовательной к влажности воздуха, то точно так же относится она и к влажности почвы. Уступая в этом отношении сосне, она, во всяком случае, в южной части своего ареала растет на значительно сухих почвах. Что касается других свойств почв, то в общем надо отметить известную требовательность лиственницы к богатству почвы, в частности к извести.
К избыточному увлажнению с застойной водой в пределах европейской части страны она весьма чувствительна, не произрастает на болотах, но в Западной Сибири становится менее чувствительной, заходя на моховые болота, где, однако, дает сильно угнетенные экземпляры.
Сибирская лиственница - порода светолюбивая, превосходящая в отношении потребности в свете все наши хвойные. Однако в молодости она может довольно хорошо переносить легкое отенение. В связи со светолюби-ем, ее крона редкая, мало затеняющая почву.
222
Как в пределах европейской части своего ареала, так и сибирской, лиственница часто образует северный предел древесной растительности, гранича с тундрой. Точно так же она на Алтае и в Саянах нередко поднимается до верхнего предела древесной растительности, образуя верхнюю границу леса.
Лиственница даурская - L. gmelinii (Rupr.) Rupr.
Она отличается от сибирской лиственницы следующими главнейшими признаками. Шишки длиною 1,6-2,5 см, округлые или овальные. Чешуй свыше 25, расположены в 3-4 ряда, совершенно голые, прямые, на конце обрубленные или с выемкой, большей частью отстоящие и косо вверх направленные. Семена мельче. Хвоя светло-зеленая или сизоватая. Кроющие чешуи при нижних рядах семенных чешуй всегда видны. Женские шишки в момент «цветения» бывают то красные, то зеленые, иногда розовые.
Дерево первой величины, с прямым стволом, достигающим 30 м высоты. Доживает до 400 лет. Кора такая же толстая, как и у сибирской лиственницы. Корневая система лиственницы даурской в обычных условиях ее произрастания, при близком уровне вечной мерзлоты поверхностная. Стержневой корень заметен только в раннем возрасте, а к 5-10 годам, независимо от условий произрастания, начинают развиваться боковые корни, и главный корень прекращает свой рост. Во взрослом состоянии корневая система представляет собою мощную густую сеть поверхностно идущих боковых корней. Однако в Забайкалье, в тех случаях, когда даурская лиственница растет на черноземовидных почвах с глубоким уровнем мерзлоты, у нее корневая система сохраняет до старости стержневой корень и вообще развивается значительно глубже.
Способность давать придаточные корни у даурской лиственницы еще сильнее выражена, чем у сибирской. Особенно это имеет значение для лиственницы на моховом болоте, где при обрастании сфагнумом на нижней части стволов они дают обильные придаточные корни, которые целиком заменяют первичные, с возрастом лиственницы оказывающиеся расположенными все глубже и глубже. Этим объясняется то, что на моховых болотах даурская лиственница достигает тех же возрастов - 300-400 лет, что и на сухих местах (сравнить с сосной, которая, не обладая способностью давать придаточные корни на болоте, доживает лишь в исключительных случаях до 80-90 лет, а обычно погибает ранее 50 лет).
Область ее распространения, охватывая всю Восточную Сибирь и Дальний Восток, очень велика, превосходя даже ареал лиственницы сибирской. Ее западная граница довольно близка к восточной границе сибирской лиственницы она направляется от низовья р. Енисея на Байкал и далее на юг в Монголию. Имеется полоса, меняющаяся по своей широте, где растут оба вида - и сибирская, и даурская и где они, переопыляясь, дают самые различные формы, как комбинации признаков той и другой. Эти гибридные формы обозначаются общим именем лиственницы Чекановского. На севере лист-
223
венница даурская всюду идет до границы леса, охватывая всю Якутию, Камчатку, Хабаровский и Приморский края.
По своим экологическим свойствам лиственница даурская близка к сибирской, отличаясь от нее несколько большим светолюбием, еще меньшей требовательностью к почвенным условиям, произрастанием на значительно более бедных песчаных, но не слишком сухих почвах и на сфагновых болотах. Но на юге Забайкалья она растет даже в сухих степях.
Хотя лиственница даурская вообще может расти на всевозможных почвах, исключая лишь крайне бедные и сухие, то все же она к плодородию очень чувствительна и хорошо развивается лишь на более плодородных почвах, в присутствии извести рост ее существенно улучшается.
Большая часть ареала лиственницы даурской характеризуется суровыми климатическими условиями, а, кроме того, часто избыточной увлажненностью и даже заболоченностью, поэтому в типичных экотопах она плохо растет и дает крайне низкие классы бонитета.
Древесина ее подобна древесине лиственницы сибирской и отличается теми же высокими качествами, будучи крепкой, смолистой и устойчивой к сырости, но и обладает теми же недостатками; поэтому находит такое же применение, как и лиственница сибирская.
Так как по экологическим свойствам лиственницы сибирская и даурская не настолько резко отличаются между собою, чтобы давать сильно различающиеся типы леса, то можно охарактеризовать вообще лиственничные типы леса, не подразделяя их специально на типы даурской и сибирской лиственниц, а лишь отметив некоторые их отличия, где это можно сделать. В общем, не рассматривая лесов Дальнего Востока, можно видеть, что в основном типы лиственничников могут быть разделены на те же группы, что и сосняки, и ельники, а именно: 1) листвяги-зеленомошники, 2) листвяги лишайниковые, 3) листвяги-травяноболотные, 4) листвяги сложные и 5) листвяги сфагновые.
Типы лиственничных насаждений
Группа листвягов-долгомошников приурочена к почвам среднего и ниже среднего плодородия, среднего увлажнения, с незастойной влагой и удовлетворительной аэрацией. Основным типом здесь является брусничный листвяг, который встречается в зоне распространения лиственницы сибирской и даурской. Он приурочен к сухим местам, с легкими почвами, супесями, легкими суглинками или мелкощебенчатой почвой. Характерно для него отсутствие подлеска или наличие единичных кустов. В травяном покрове господствует брусника при значительном, иногда почти сплошном, ковре из обычных зеленых мхов. Рост лиственницы относительно хороший. Эти листвяги являются одним из наиболее ценных типов лиственничного леса.
Другим значительно распространенным типом является багульниковый листвяг. Он приурочен к песчаным или щебенчато-суглинистым почвам, менее плодородным, чем в предыдущем типе, а главное - более увлажненным и
224
с малопроточной, отчасти застаивающейся влагой, то есть к почвам, уже несущим, хотя и в начальной форме, следы заболачивания. Подлесок отсутствует или очень редкий. В травяном покрове значительное участие принимает багульник, развит моховой покров.
Примером багульниковых листвягов может быть листвяг рододендровый, который широко распространен в Прибайкалье, Забайкалье и в южной части Якутии. Этот тип встречается, главным образом, на склонах, иногда даже очень крутых, с достаточно плодородными, хорошо дренированными почвами, чаще грубыми, щебенчатыми. Характерной его чертой является обычно сильно развитый подлесок из рододендрона даурского, к которому иногда примешиваются другие кустарники. Травяной покров не густой - из брусники и багульника. Моховой покров очень редкий, часто вовсе отсутствует.
Этот тип леса является очень ценным, так как, с одной стороны, в нем лиственницы обладают хорошим ростом, с другой - они обычно дают в нем здоровые стволы с хорошей древесиной.
К группе травяно-болотных листвягов относятся листвяги на проточно увлажненных плодородных почвах. Такие листвяги растут, например, в горном Забайкалье и южной части Якутии.
Лиственница здесь отличается особенно быстрым ростом, достигая наиболее значительных размеров. Однако качество ее древесины вследствие очень мощного роста хуже, чем в рододендровом листвяге. Подлесок довольно развит; в его составе ива, смородина, жимолость и некоторые другие виды кустарниковых растений. Травяной покров развит хорошо, высокий и густой; в нем наряду с брусникой, встречающейся, в сущности, во всех типах лиственничников, много крупных растений, особенно злаков. Моховой ковер развит очень слабо.
Группа сфагновых листвягов распространена особенно там, где влажность воздуха обуславливает массовое развитие сфагновых болот. Она обычно представлена сфагновым листвягом. Этот тип леса по своему облику сходен со сфагновыми сосняками, лиственница здесь дает самые низкие бонитеты.
Наконец, последняя группа листвягов, о которой необходимо сказать, -это листвяги лишайниковые. Эта группа мало распространена, в силу того, что на сухих песчаных почвах она чаще замещена сосною. Пример - толок-нянковый листвяг. Он приурочен к сухим, песчаным почвам, а следовательно, хорошо дренированным. Это обычно чистые лиственничные насаждения или с примесью сосны. Подлесок отсутствует или крайне редок. Травяной покров небогат по числу экземпляров, но довольно разнообразен по числу видов. Характерно сильное распространение толокнянки и овсяницы. Лишайниковый покров не особенно густой. Рост лиственницы здесь хуже, чем в брусничном листвяге, но лучше, чем в мшистом.
Под влиянием пожаров, из названных выше типов, которые могут относиться к категории коренных, развиваются производные, характеризующиеся чаще всего значительным участием в них березы.
225
МЕЛКОЛИСТВЕННЫЕ ЛЕСА
Мелколиственные леса (в отличие от широколиственных) образованы деревьями со сравнительно мелкими листьями - березой повислой и пушистой, осиной или тополем дрожащим, черным и белым тополями, клейкой и серой ольхой, ивой белой. Они широко распространены в северной лесной зоне Восточно-Европейской и Западно-Сибирской равнин, в горах и на равнинах Дальнего Востока, входят в состав западносибирской и среднесибирской лесостепи, образуя там полосу лиственных лесов от Урала до Енисея. Берёзовые леса распространены гораздо шире, чем осинники. Мелколиственные леса светлые, отличаются разнообразием и богатством травяного покрова. Суммарная площадь березняков составляет примерно 1,35 млн. га, осинников - 0,8. Ольховые и ивовые леса встречаются преимущественно в поймах многочисленных рек Российской Федерации и играют важную роль в регулировании их режима.
В основном, мелколиственные леса относятся к так называемым временным или производным лесным формациям, на какое-то время сменяющим коренные темнохвойные леса на севере лесной зоны и широколиственные - в южной ее части. Связано это с тем, что основные виды древесных растений, образующих мелколиственные насаждения (береза, осина), весьма светолюбивы, семена их легко разносятся ветром на большие расстояния, а семенная продуктивность взрослых деревьев очень высока. Это породы-пионеры, быстро заселяющие вырубки, гари, заброшенные сельскохозяйственные земли. Вместе с тем они не долговечны, а вследствие светолюбия практически не возобновляются под своим собственным пологом. При естественном течении лесообразовательных процессов мелколиственные леса вновь сменяются теневыносливыми темнохвойными.
Однако мелколиственные леса, особенно березняки, могут быть представлены и постоянными формациями. К таковым относятся, например, степные, обычно небольшие по площади, березовые насаждения, так называемые колки. К постоянным можно отнести и притундровые редкостойные насаждения с преобладанием березы. Весьма постоянны по составу и пойменные черноольховые леса.
Род береза - Betula L.
В нашей стране встречается порядка сорока видов этого рода, из которых одни представляют собою довольно крупные деревья, другие - карликовые формы или кустарники. Последние лесохозяйственного и, тем более, лесопромышленного значения не имеют, поэтому на них мы не будем останавливаться, а рассмотрим два вида, наиболее характерные для северной лесной зоны. Прежде чем давать подробную характеристику, укажем на их главнейшие морфологические отличия (рис. 64).
226
Рис. 64. На рисунке: А - листья, плод и чешуйка из соплодия березы повислой; Б -то же самое березы пушистой. На фотографии - слева опушение на стебле березы пушистой, справа бородавки на стебле березы повислой (фото В.В. Коровина)
1. Листья треугольно-ромбические или ромбические, с сильно оттянутой верхушкой, остродвояко-зубчатые. Ветви повислые, крона плакучая, однолетние побеги тонкие, бородавчатые или голые. Береста абсолютно белая, иногда розовая. Корка в нижней части стволов с глубокими черными трещинами, иногда из-за многочисленных трещин совсем черная. Плодовые чешуйки женских соплодий ("самолетики") с боковыми лопастями отогнутыми вниз. Крылышки плодов в 2-3 раза шире самих плодов - береза повислая.
2. Листья овальные или яйцевидные, большей частью, без оттянутой вершинки, просто зубчатые или неясно двояко-зубчатые. Ветви не повисают, крона яйцевидная, однолетние побеги более толстые, опушенные, особенно, у порослевых и молодых экземпляров. Береста у взрослых деревьев Сероватая. Кора даже у основания ствола до перестойного возраста остается без черных трещин. Плодовые чешуи женских соплодий с боковыми лопастями, направленными косо вверх, Крылышки плодов по ширине равны самим плодам или шире их в 1,5 раза - береза пушистая.
227
Береза повислая - В. pendula Roth.
Дерево до 30м высоты, до 80см в диаметре. Ствол стройный, с белой отслаивающейся тонкими пластинками корой, которая называется берестой. С возрастом кора у основания ствола утолщается и покрывается глубокими черными трещинами. Крона плакучая с тонкими свисающими ветвями. Побеги с восковыми бородавками. Листья треугольные или ромбические с оттянуто-заостренной вершинкой и дважды- или трижды-зубчатым краем. Мужские двусложные соцветия - сережки - располагаются на концах побегов и зимуют открыто. Соцветие состоит из стержня, на котором по спирали располагаются многочисленные трехцветковые дихазии. Дихазий состоит: из оси, трех прицветных листьев неравной величины (два малых и один большой, накрывающий их сверху) и трех цветков. Каждый цветок располагается под отдельным прицветным листом и состоит из двух тычинок (каждая тычинка с двумя пыльниками).
Женские двусложные соцветия - сережки, расположены в пазухах листьев укороченных побегов и зимуют в почках. На оси сережки расположены соцветия второго порядка - трехцветковые дихазии, которые состоят из короткой оси, трех прицветных листьев (самый крупный средний) и трех пестиков с двураздельными рыльцами пурпурного цвета. Из каждого пестика развивается орех с крылышками. Плодовые сережки созревают во второй половине лета (середина июля - август) и, после созревания, рассыпаются. Плод -плоский орех, 1,5-2,0 мм длиной, с остатками двух рылец на вершине и с двумя перепончатыми крылышками, которые в 2-3 раза больше семени. Из трех прицветных листьев дихазия образуется плодовая чешуйка, которая облетает вместе с плодами. Боковые лопасти плодовых чешуек отогнуты вниз, средняя лопасть длиннее и направлена вверх. В момент созревания плодов на концах побегов закладываются новые тычиночные соцветия для цветения в следующем году (рис 65).
Рис. 65. Береза повислая: 1 - побег во время цветения с мужскими и женскими сережками; 2 - мужские сережки; 3 - мужской трехцветковый дихазий; 4 - тычинка с двумя пыльниками; 5 - женские сережки; 6 - женский трехцветковый дихазий; 7 - укороченный побег с листьями и соплодием; 8 - плод (крылатый орех) ((из Strasburger, Noll, Schenck, Schimper, 1967)
Односемянные плоды, в практике часто называют семенами. Всхожесть плодов невысокая и со временем быстро теряется. Уже на следующую весну она падает всего до нескольких процентов. Всходы мелкие, нежные, с двумя овальными мелкими семядолями, голые, сверху
228
зеленого, снизу красноватого цвета. Первые 1-2 года растет не очень быстро, но потом рост значительно ускоряется, особенно к 10-20 годам. К 50-60 годам он обычно уже прекращается. Береза редко доживает до 120 и более лет.
Ареал: вся лесная зона европейской части России, Западная и Восточная Сибирь, заходит в лесостепь и в степь (рис. 66). Береза повислая морозостойка, очень светолюбива, засухоустойчива, малотребовательна к плодородию почвы, является почвоулучшающей породой, деревом - пионером, заселяющим вырубки и гари. После рубки дает обильную пневую поросль из спящих почек, которые обыкновенно в большом количестве сохраняются у корневой шейки. Такая побегопроизводительная способность сохраняется у деревьев семенного происхождения очень долго, и хотя обычно к 40 годам она ослабляется, но все же сохраняется до 60-80 лет.
Рис. 66. Ареалы двух видов березы: 1 - береза пушистая; 2 - береза повислая
Несмотря на значительное участие березы в наших лесах, подавляющее большинство березняков являются не естественными, не коренными, а производными, связанными с вмешательством человека в жизнь природы. Большинство березняков развилось на месте вырубленных или выгоревших сосновых, еловых и лиственничных или даже дубовых лесов. Указанные выше свойства березы позволяют ей быстро заселять оголяемые пожаром или рубками площади, а также и заброшенные пашни. Однако она на таких местах не способна долго удерживать за собой территорию. Если имеется налет или занос семян других пород, то они, развиваясь под пологом березняков, вскоре
229
вытесняют березу вовсе, и таким образом восстанавливается прежний характер леса на месте временного березняка.
О достоверно естественных коренных березняках можно говорить лишь в области степи и лесостепи, где береза является пионером в заселении лесом степи. По-видимому, коренными являются и некоторые березняки на заболоченных местах в лесной зоне. В остальных случаях в естественных условиях береза встречается лишь как примесь к другим породам, а самостоятельно чистых насаждений не образует.
Древесина светлая, легко колется, хорошо полируется при обработке в сухом виде, находит применение во всевозможных токарных изделиях, для изготовления ружейных лож, топорищ и пр. Березовую древесину широко используют для производства бумаги, в фанерном и мебельном производстве. Березовые дрова считаются лучшими. В результате ее сухой перегонки получают древесный спирт, уксус и уголь. Почки и молодые листья используются в медицине. Сок березы содержит до 2% сахара. Кора березы (береста) до сих пор используется для изготовления специальных емкостей для сбора грибов и ягод, а также для производства разнообразных сувениров. Продуктом сухой перегонки бересты является деготь. Вообще использование березовой древесины весьма разнообразно. В степи и лесостепи березу повислую применяют для создания полезащитных лесных полос.
Анатомические особенности древесины. Поперечный срез. Древесина белая, без выраженного ядра. Границы годичных колец слегка волнистые, часто слабо различимы. Древесина рассеяннососудистая. Просветы члеников сосудов редкие, округлые, одиночные или в радиально ориентированных группах и цепочках. Паренхима диффузная, инициальная и метатрахеальная. Тангенциальный срез. Лучи преимущественно 1-3 рядные, с небольшой долей 4-5 рядных, гомогенные. Однорядные - сложены из 1-10 клеток в высоту, многорядные - из 10-20 клеток. Могут встречаться агрегатные лучи. Радиальный срез. Перфорационные пластинки лестничные с тонкими перекладинами. Межсосудистая поро-вость очередная; поры очень мелкие, частые. Поры между сосудами и лучевой паренхимой сходны по форме и размеру с порами между сосудами. Волокнистые элементы представлены волокнистыми трахеидами. Лучи состоят из лежачих и иногда с одним краевым рядом квадратных клеток.
Особую ценность представляет собою древесина, так называемой карельской березы, стволы которой, часто образующие особые утолщения, благодаря извилистому расположению осевых элементов древесины и аномальному изменению лучевой паренхимы, на разрезах дают очень красивый рисунок, из-за которого она и ценится. Древесина карельской березы используется для изготовления токарных изделий, очень дорогой мебели, декоративной посуды, шкатулок и т. п.
Карельская береза, в пределах нашего государства, встречается: в Карелии, в также в Вологодской, Костромской, Владимирской, Калужской, Смоленской областях, и возможно в других местах. Она не образует сплошных насаждений, а встречается единично или куртинами среди березы повислой, обычно, в низкополнотных древостоях.
На кафедре селекции, генетики и дендрологии МГУЛа под руководством профессора А.Я. Любавской проведена большая и успешная работа
230
по селекции и искусственному разведению этого ценного лесного растения. Это направление исследований на кафедре продолжается.
Береза пушистая - В. pubescens Ehrh.
Береза пушистая во многом сходна с повислой, однако, имеет ряд постоянных отличительных признаков, по которым хорошо обособляется от повислой. Главнейшие отличия березы пушистой следующие.
Дерево, до 20-25 м высоты, со стройным стволом, покрытым сероватобелой нерастескивающейся коркой. Крона яйцевидная. Ветви более короткие и плотные, чем у березы повислой, и направлены вверх. Побеги, листья (особенно молодые) и черешки листьев густо опушены. У поросли опушение сохраняется в течение всего лета. Листья яйцевидной и широкояйцевидной формы с короткой, заостренной вершинкой, край листа зубчатый. Строение соцветий и плодущих сережек такое же, как и у березы повислой, но плодовые чешуйки и размеры крылышек плода отличаются. Боковые лопасти плодовой чешуйки направлены вверх. Крылышко равно семени или в 1,5 раза больше его.
После созревания соплодия держатся на дереве дольше, чем у березы повислой. Часто до поздней, или даже ранней зимы (рассыпаются значительно позже созревания семян) сережки продолжают висеть на дереве.
В природе часто бывает трудно установить принадлежность той или иной березы к определенному виду (березе повислой или березе пушистой). В ранней литературе бытовало мнение, что береза повислая легко скрещивается с березой пушистой, давая в потомстве промежуточные по морфологическим признакам экземпляры. На самом деле, возникновение таких гибридов - явление очень редкое. Эти виды имеют разное число хромосом и поэтому не могут легко скрещиваться. Кроме того, у березы пушистой и повислой на очень больших площадях ареалы перекрываются и в случае свободной гибридизации видовые признаки этих растений так или иначе должны бы были нивелироваться.
Сравнивая рассматриваемые два вида березы, следует также иметь в виду, что они обычно занимают разные экологические ниши. В пониженных местах с избыточным увлажнением явное преимущество получает береза пушистая, на более высоких местоположениях, в условиях хорошо дренированных песчаных и супесчаных почв, а так же при дефиците почвенной влаги повислая береза оказывается более конкурентоспособной.
Ареал: растет в лесной зоне европейской части России, Западной и Средней Сибири, горах Кавказа, Алтая и Средней Азии, заходит далеко на север в лесотундру и тундру (рис. 66). Подобно повислой березе, береза пушистая, на протяжении своего большого ареала имеет ряд географических форм, которые некоторыми авторами рассматриваются, как особые виды Она теневынослива, зимостойка, требовательна к плодородию и влажности почвы, может расти в условиях переувлажнения по заболоченным долинам рек и
231
на сфагновых болотах. Повышенное по сравнению с березой повислой влаголюбив делает ее непригодной для полезащитного разведения.
Древесина березы пушистой по физико-механическим свойствам практически не отличается от древесины березы повислой. Нет достоверных отличий и в анатомическом строении древесины.
Отдельные популяции березы пушистой в условиях, затрудняющих семенное размножение, в частности, в поймах рек с кочковатым микрорельефом и плотным напочвенным покровом, обладают повышенной способностью образовывать капы, особенно прикорневые (см. рис. 35). Такие насаждения известны в горной части Республики Башкортостан, на юге Кировской области, в некоторых районах Новосибирской области. В перечисленных районах капы встречаются очень часто, а величина этих образований (вес до 0,5 тонны) позволяет использовать их неповторимую по красоте древесину для изготовления подарочных изделий. Такие изделия удостаивались высоких наград на международных выставках.
Род тополь - Populus L.
Род Тополь включает в себя быстрорастущие листопадные двудомные деревья, нередко достигающие огромных размеров. Листья цельные и только у белых тополей могут быть пальчато-лопастными. Цветки закладываются в год, предшествующий цветению, в боковых цветковых почках, которые всегда крупнее ростовых и весной трогаются в рост раньше их. Цветки без околоцветника, расположены в пазухах лопастных прицветковых чешуй поникающих сережек. В мужских цветках тычинки прикреплены к расширенному дисковидному цветоложу, в женских цветоложе бокаловидное. Тополь ветроопыляемое растение, цветет одновременно с распусканием листьев или до облиствения. Плод - коробочка, семена созревают с конца весны до середины лета и сразу же разносятся ветром, когда происходит лёт «тополевого пуха». В возраст половой зрелости тополь вступает с 7- 15 лет. Размножается семенами, корневыми отпрысками, порослью от пня. В культуре тополь размножают также стеблевыми (не все) или корневыми черенками. Тополя сравнительно недолговечны; часто из-за поражения стволовой гнилью живут менее 100 лет, хотя известны отдельные долгожители в возрасте 400 лет.
Осина, тополь дрожащий - Р. tremula L.
Дерево до 35 (42) м высоты и до 1 м в диаметре. Крона округлая, высоко поднятая над землей состоит из толстых скелетных ветвей, несущих многочисленные удлиненные и укороченные побеги. Ствол цилиндрический, вверху кора гладкая, светлая, серо-зеленая, внизу темная, почти черная, про-дольно-трещиноватая. Побеги голые, блестящие буровато-коричневого цвета. Листовые почки конически-заостренные, цветочные шаровидные.
Листовые почки распускаются в конце апреля - начале мая, цветочные за две недели до распускания листьев, в апреле. Листья двух типов. На уко
232
роченных побегах они округлые, жесткие, голые или с малозаметным прижатым пушком, коротко заостренные, с городчато-зубчатым краем, до 7 см длиной, на длинных сплюснутых черешках, более тонких по середине. Такая форма листьев придает им неустойчивость, и они дрожат при малейшем ветре, отчего и произошло видовое название этого растения. Листья удлиненных побегов и поросли треугольно-эллиптические или яйцевидные, иногда очень крупные, слегка сердцевидные или плоские при основании. Жилкование листьев характерное, дланевидное (рис. 67).
Рис. 67. Осина: 1 - побег во время цветения с мужской сережкой; 2 - мужской цветок с лопастным прицветным листом; 3 - побег во время цветения с женской сережкой; 4 - женский цветок с лопастным прицветным листом; 5 - лист с длинным черешком; 6 - нераскрывшийся плод - коробочка; 7 - раскрывшийся плод; 8 - семя с пучком волосков (из Strasburger, Noll, Schenck, Schimper, 1967)
Обычно рост листьев прекращается в конце мая, в октябре опадают последние листья. Таким образом, в средних широтах вегетационный период у осины продолжается 150 дней.
Осина, как и все тополи, двудомна (часть экземпляров несет только мужские цветки, другая часть - только женские).
Мужские особи встречаются чаще женских. Процесс созревания семян про
текает быстро; сережки удлиняются иногда до 20-28 см, коробочки раскры
ваются двумя створками и к началу лета выпускают огромное количество мелких семян с волосками, обеспечивающими семенам летучесть.
Осина встречается практически во всей лесной зоне России. На севере доходит до границы леса с тундрой. В степях растет в колках; заходит даже на солончаки, принимая здесь вид кустарника. На Кавказе поднимается до альпийских лугов. В пределах обширного ареала образует значительное число форм.
К климатическим условиям осина не требовательна. Зимостойка. Не страдает от заморозков. Лишь иногда у поросли обмерзают невызревшие побеги. Первой заселяет заброшенные пашни, гари. Весьма светолюбива, в связи с чем обычно ствол ее быстро и высоко очищается от сучьев. Крона у старых осин изреживается.
К почве осина довольно требовательна, лучшие для себя условия находит на свежих, богатых перегноем, суглинках. Хорошо растет и на легких супесях при наличии богатых подстилающих пород. Способна накапливать запас древесины более 400 м3 на 1 га.
Лучшие деревья осины отличаются следующими показателями: в возрасте 10 лет имеют высоту 5 м (максимум 7 м), в 30 лет - 20 (25) м, в 50 лет -
233
21 (31) м, в 70 лет - 29 (36) м. Отдельные деревья в возрасте 75 лет иногда достигают 75 см толщины и имеют здоровую древесину.
Растет осина и на серых лесных суглинках и черноземах, в лесостепи и степи, перенося некоторую засоленность почв. Здесь она селится на месте вырубленного дуба. Однако некоторые местоположения, занятые осинниками, можно считать и их коренными местообитаниями, например, колковые леса.'
Осина развивает довольно мощную корневую систему, состоящую из корней, уходящих в глубь почвы (до 1 м). Помимо корней у осины образуются подземные стебли (корневища) с придаточными почками. Корневища расходятся далеко в стороны (до 35 м) за периферию кроны дерева, то приближаясь к поверхности почвы, то углубляясь в нее. После рубки из придаточных почек подземных стеблей развиваются побеги, так называемые «корневые отпрыски», иногда весьма многочисленные. В большинстве случаев осина возобновляется корневыми отпрысками. Однако с увеличением возраста деревьев их способность возобновляться вегетативно резко снижается.
Обычно осинники порослевого происхождения сильно заражены гнилью, вызываемой ложным трутовиком. Это заболевание резко снижает товарность древесины, делая ее не пригодной даже на дрова.
Осина вступает в пору плодоношения в возрасте около 20 лет и плодоносит почти ежегодно. Она выпускает колоссальное количество семян, но прорастают лишь единичные; остальные гибнут, попав в засушливый период лета, от затенения травами и других причин. Свежие семена имеют высокую всхожесть, но уже через 1-3 месяца полностью ее теряют. 1000 семян весят 0,13 г. На поверхности влажного субстрата проростки появляются через полтора дня после посева. Они прикрепляются волосками к почве и выпускают две мелкие семядоли. В дальнейшем развивается в первый же год корешок до 30 см длиной. В хороших условиях сеянцы могут достигнуть более 20-30 см высоты уже в первый год.
Осина долгое время считалась сорной породой. Мнение это изменилось после того, как была доказана пригодность ее древесины в качестве сырья для целлюлозно-бумажного и спичечного производства. С тех пор находят все новые области использования древесины осины.
Древесина осины белого цвета, легкая, мягкая, легко колется и режется, в сухом состоянии достаточно крепкая, твердая и прочная. Относится к типу рассеяннососудистых. Ядро не выражено. Граница годичных слоев неясная. Равномерная пористость древесины и легкость обработки делают ее незаменимой для спичечной "соломки". Из древесины осины получают целлюлозу. Она используется также для изготовления срубов в колодцах, лодок, ящичной фанеры, клепки для бочек, внутренней отделки бань.
Анатомические особенности древесины. Поперечный срез. Ядро не выражено. Годичные кольца средне отчетливые. Древесина рассеяннососудистая. Просветы сосудов крупные, редко одиночные, в основном сгруппированные в радиальные цепочки; иногда просветы сосудов составляют около 50 % площади среза. Одиночные просветы угловатые и овальные. Волокна либриформа тонкостенные. Осевая паренхима скудная, апотрахеаль-ная (диффузная и терминальная в коротких цепочках) и паратрахеальная (скудновазицен-
234
трическая). Тангенциальный срез Лучи исключительно однорядные, сложенные из 4 - 20 овальных клеток одинакового размера. У Populus alba, Р. nigra и Р. tremula встречаются перегородчатые волокна либриформа. Радиальный срез. Перфорации исключительно простые. Межсосудистая поровость очередная; поры крупные, частые, шестиугольные и округлые.. Поры между сосудами и клетками лучей слегка угловатые, простые, гораздо крупнее, чем поры между сосудами. Лучи гомогенные, редко встречаются слабо гетерогенные. Лучи состоят из лежачих клеток и иногда с одним рядом краевых квадратных. Волокнистые элементы представлены исключительно волокнами либриформа.
Типы леса с участием березы и осины
Что касается типов леса, образуемых осиной и березой, то они, будучи генетически связаны с еловыми, сосновыми или дубовыми, на месте которых возникают после рубки или пожара, естественно, являются аналогичными последним.
Так как осина относительно требовательна к плодородию почвы, то она сменяет те типы хвойных, которые приурочены к лучшим почвам. Поэтому же она, значительно чаще, чем береза, заселяется после дуба. На месте сухих с бедными почвами типов леса, таких как сосняк лишайниковый или сосняк-брусничник, осина не образует производных типов. Точно так же на сфагновых болотах, она если и встречается, то в виде небольших медленно растущих деревьев.
Хотя береза на местах лишайникового бора не дает насаждений, на сфагновых болотах ее насаждения имеются. В остальном же типы лесов этих пород будут соответствовать тем типам хвойных или широколиственных лесов, на месте которых они возникли. При этом можно различать осинники и березняки кисличные, черничники, травяно-болотные, долгомошники, лип-няковые и т. п. По своему строю, составу и растительности они сходны с соответствующими ельниками и сосняками. То же самое наблюдается и в типах березняков и осинников, производных от дубрав. Так как береза и осина более светолюбивы, чем ель, дуб и сосна, то под пологом березняков и осинников больше развит травяной покров. При этом нередко часть травяного покрова коренных типов, состоящих из тенелюбивых растений, как например, кислица, грушанка, вороний глаз, из состава травостоя выпадает.
После осины и березы, среди наших лиственных пород зоны хвойных лесов видное место занимают виды ольхи, дающие своеобразные типы леса, к рассмотрению которых и перейдем. К лесообразователям на территории северной лесной зоны относятся два вида: ольха черная и ольха серая.
Род ольха - Alnus L.
Род ольха объединяет до 20 видов, произрастающих в Европе, в горах Алжира, в Азии, Северной и Южной Америке. Это деревья, реже кустарники с очередными зубчатыми листьями с перистым жилкованием и рано опадающими прилистниками. Все они однодомны. Сережки появляются осенью. Тычиночные сережки длинные, цилиндрические, во время цветения стержни
235
сережек вытягиваются и они повисают. Пестичные сережки короткие. Цветет до распускания листьев. В наших широтах - одно из наиболее раннецветущих древесных растений. Соплодие деревянистое, шишковидное. Плод - односемянной орех со слабо развитым крылышком.
Ольха черная (клейкая) - A. glutinosa Gaertn.
Крупное дерево с небольшой кроной и полнодревесным стволом, покрытым темно-бурой трещиноватой корой. Годичные побеги трехгранной формы с треугольной сердцевиной. Почки тупые, клейкие, на ножках, молодые побеги также клейкие, с беловатыми чечевичками (рис. 68).
Рис. 68. Ольха черная: 1 - побег во время цветения с мужскими и женскими сережками; 2 - мужские сережки; 3 - мужской трехцветковый дихазий; 4 - женские сережки; 5 - женский двухцветковый дихазий; 6 -лист с выемчатой вершинкой; 7 - недозрелое шишковидное соплодие; 8 - раскрывшееся зрелое соплодие; 9 - плод - орех с узкими крылышками (из Strasburger, Noll, Schenck, Schimper, 1967)
В момент цветения тычиночные сережки, собранные по 3-5 штук, удлиняются, свисают и выделяют большое количество желтой пыльцы. Пестичные сережки собраны на концах веточек тоже по 3-5 штук, сидят на
заметных ножках После цветения и оплодотворения чешуи смыкаются и образуются зеленоватые шишки. При созревании шишки буреют и становятся деревянистыми, достигая 1,5 см длины. В них заключены плоды - сплюсну
тые орехи.
Листья ольхи черной обратноовальные или округлые, часто с выемчатой верхушкой и широко клиновидным основанием. Края листьев у основания цельнокрайние, выше - неправильно городчато-пильчатые. Размер листовой пластинки 4-9 см. Старые листья с верхней стороны темно-зеленые, гладкие, блестящие, с нижней - светлее, с бородками в углах жилок.
Ольха черная распространена на значительной территории европейской части России. Встречается в Крыму и на Северном Кавказе, в степной зоне -по речкам, на севере поднимается до Белого моря и верхнего течения Камы, в Западной Сибири местами растет около Тюмени. Одиночные острова ольхи черной известны по р. Уралу и в низовьях р. Волги.
Ольха черная достаточно зимостойка, но сильной континентальности климата не переносит. Жаркий климат хорошо переносит только при условии достаточной влажности почвы.
236
Хорошая очищаемость от сучьев свидетельствует о том, что ольха черная относится к светолюбивым породам. Однако в раннем возрасте она может переносить заметное отенение. К почве требовательна. Лучше всего растет на богатых перегноем, избыточно увлажненных почвах с проточной грунтовой водой, на так называемых черноольховых трясинах, низинных болотах, при наличии аэрации. Здесь деревья часто расположены на крупных кочках, окруженных топями. К 50 годам высота ольхи составляет в среднем 28 м, а запас на гектар превышает 300 м3. Неплохо растет и на темноцветных почвах. На торфянистых почвах рост ольхи черной заметно снижается, и она уступает место березе пушистой.
Корневая система ольхи черной поверхностная. Деревья нередко вываливаются с корнями.
Ольха черная - быстрорастущая порода. Наиболее крупные деревья к 120 годам имеют в среднем 40 см в диаметре и до 35 (40) м в высоту. Особенно быстро растет ольха черная в возрасте 5-20 лет. Первое падение прироста совпадает с началом плодоношения. Обычно ольха черная редко доживает до 150-300 лет, так как разрушается грибами, в частности, страдает от красной гнили.
Цветет ольха черная до появления листьев. Опыляется при помощи ветра, чему способствует расположение цветков в сережках. Плоды созревают к осени. Плоды переносятся водой по течению. Опадают обычно весной, при вешней воде. При спаде воды плоды прилипают к илу и прорастают, если им не мешают травы и солнцепек. Всходы ольхи имеют две мелкие гладкие темно-зеленые семядоли. Первые листочки заострены и по краям зубчатые, с редкими волосками. Иногда плоды частично опадают в сухую осень. Пустые шишечки отваливаются позднее.
Обильные урожаи зависят от погоды и повторяются через 2-3 года. Семян созревает много, всхожесть их едва превышает 40% и лишь изредка доходит до 70 %. 1000 шт. плодов весят 1,2-2 г. Ольха черная хорошо возобновляется порослью от пня и сохраняет эту способность до 80 лет. Способность к порослевому возобновлению улучшается при образовании довольно крупных прикорневых капов. Ольха семенного происхождения более долговечна.
Ольха черная является основной лесообразующей породой в условиях низинных болот и по поймам рек, способствуя мелиорации этих почв. Медоносна. Древесина ее рассеяннососудистая, заболонная (ядро слабо различимо), с широкими лучами, хорошо заметными на торцевом срезе. При срубке краснеет, в сухом виде отличается приятной розовой окраской. Древесина ольхи высоко ценится: она мягкая, легкая, хорошо колется, обрабатывается, воспринимает протравливание и окраску, стойка в воде. Используется для изготовления фанеры, в столярном, токарном, целлюлозном производствах, в строительстве гидротехнических сооружений, а также для копчения рыбы. При сухой перегонке из древесины ольхи получают уголь, древесный уксус и др. Ольховые дрова отличаются высокой теплотворной способностью. В коре содержится до 6-9 % дубильных веществ.
237
Анатомические особенности древесины. Поперечный срез. Ядро имеется. Граница годичных колец отчетливая, в месте пересечения агрегатного луча извилистая. Древесина рассеяннососудистая. Просветы довольно частые, сгруппированные в радиально ориентированные цепочки. Волокнистые элементы тонкостенные. Паренхима апотрахе-альная, диффузная, диффузно-агрегатная и терминальная в коротких тангенциальных цепочках. Клетки паренхимы трудно различимы среди трахеид. Агрегатные лучи. Часто встречаются сердцевинные повторения56. Тангенциальный срез. Лучи 1-рядные, 10-25 клеток в высоту; иногда встречаются низкие двухрядные. Агрегатные лучи состоят из 1-3-рядных. Радиальный срез. Перфорации конечных стенок сосудов лестничные с 10-20 узкими перекладинами. Межсосудистая поровость супротивная и очередная. Поры некрупные и очень частые. Поры между сосудами и члениками лучей заметно мельче, частые, округлые с зауженным окаймлением. Основная масса древесины состоит из волокнистых трахеид. Лучи гомогенные, состоящие из лежачих клеток; иногда встречаются гетерогенные, состоящие из внутренних лежачих клеток с одним рядом краевых квадратных.
Ольха серая - A. incana Moench.
Дерево средней величины или крупный кустарник с искривленным сбежистым стволом, покрытым серой гладкой корой. В отличие от ольхи черной имеет в раннем возрасте опушенные побеги, почки, цветоножки и листья. Листья овальной или яйцевидной формы по краям двоякопильчатые, к основанию цельные, на верхушке заострены, с нижней стороны с сизоватым пушком, с верхней тускло-зеленые. Тычиночные сережки светлее, чем у ольхи черной; пестичные сидят по 3-8 шт. на цветоножках. Ольха серая цветет в первой декаде апреля, до появления листьев, на неделю раньше черной. Шишечки около 1 см длиной. Раскрываются осенью. Буроватые плоды (орехи) опадают и относятся ветром. Орехи с несколько более развитым, чем у черной ольхи перепончатым краем крылышка (рис. 69).
Ольха серая очень зимостойка. На севере заходит на 4° дальше ольхи черной, достигая тундры. Южная граница ольхи серой лежит севернее лесостепи.
Ольха серая теневыносливее березы и осины. К почве менее требовательна, чем ольха черная. Хорошо растет на дерново-подзолистых суглинистых и торфянисто-подзолисто-глееватых суглинистых почвах. Ольха улучшает почву, обогащая ее азотом. Быстрое разложение подстилки из листьев увеличивает плодородие почвы, поэтому в ольшаниках всегда много трав: число видов травянистых растений доходит до сотни.
На хороших почвах ольха серая растет очень быстро. Хорошо растет также по берегам ручьев. Кислых, сильно заболоченных или очень сухих почв избегает. Застойную влагу переносит лучше, чем ольха черная.
36 Сердцевинные повторения - не очень удачное название небольшого аномального скопления раневой па-ренхимы, возникающего у некоторых древесных растений в древесине в результате повреждения камбиальной зоны насекомыми.
238
Рис. 69. Ольха серая во время цветения: 1 - мужские сережки; 2 - женские соцветия; 3 - прошлогодние соплодия (фото В. Коровина)
Корневая система ольхи серой относительно мелко сидящая, но мощная. Легко заселяет вырубки, пожарища, заброшенные пашни. Иногда достигает 15 м высоты и 30 см толщины. Доживает до 50-60 лет, единично - более 160 лет.
Древесина похожа на древесину ольхи черной, по анатомическим признакам древесины эти виды не различаются. Используется для мелких поделок, изготовления рисовального и порохового угля. Топливо посредственное. Древесина содержит камедь. Кора содержит до 10% танинов и около 3 % сахаров. Используется для дубления и крашения кож. Серьезного лесопромышленного значения не имеет.
Типы ольшаников
Черная ольха - дерево так называемых черноольховых трясин, где на богатой почве, представляющей, большею частью, хорошо разложившийся торф, с проточной водой она дает чаще всего чистые насаждения - ольшаники. Густой, сомкнутый полог, образованный ольхой, позволяет развиваться под ним теневыносливым растениям. Второй ярус обычно отсутствует; в подлеске, который редко бывает густым, встречается крушина ломкая, черная смородина, калина, реже - ива козья. Микрорельеф в таких насаждениях обычно кочковатый. Деревья растут на повышениях, между которыми часто стоит вода. Травяной покров состоит из влаголюбивых и теневыносливых растений или таких, которые цветут рано весною, пока еще ольха не затенила сильно почву. К их числу относятся калужница и селезеночник. На более сухих буграх обитают типичные лесные растения - кислица, вороний глаз, седмичник; по понижениям - сныть, таволга вязолистная.
239
В ольшаниках, подверженных сильному воздействию человека, особенно несколько осушенных, сильно разрастаются сорняки, среди которых обширные заросли образует крапива. Примерно такие ольшаники широко распространены по всему ареалу черной ольхи.
Реже встречаются несколько иные растительные ассоциации с черной ольхой в древесном ярусе. Например, на бедных почвах, на песках, покрытых слоем торфа, и развитым сфагновым ковром (ольшаник сфагновый) могут быть и некоторые другие типы ольшаников.
Ольха серая образует коренные ольховые ассоциации в виде зарослей по окраинам болот, иногда по опушкам и прогалинам. Чаще же она заселяет заброшенные пашни и суходольные луга, иногда - вырубки, пожарища, образуя кратковременные производные типы леса.
ЕВРОПЕЙСКИЕ ШИРОКОЛИСТВЕННЫЕ ЛЕСА
Современные широколиственные и хвойно-широколиственные леса сформировались пять - семь тысячелетий назад, когда на планете наступило послеледниковое потепление и широколиственные виды древесных растений смогли далеко продвинуться на север. В последующие тысячелетия климат становился более холодным и зона широколиственных лесов постепенно уменьшалась. Поскольку под этими лесами сформировались наиболее плодородные из всей лесной зоны почвы, леса интенсивно вырубались, а их место занимала пашня. Кроме того, дуб, обладающий очень прочной древесиной, широко использовался в строительстве.
Правление Петра I стало для России временем создания парусного флота. Кораблестроение требовало большого количества высококачественной древесины, поэтому так называемые корабельные рощи строго охранялись. Леса, не входившие в охраняемые территории, жители лесной и лесостепной зон активно вырубали под пашни, сенокосы и на строительство. В середине XIX века закончилась эпоха парусного флота, корабельные рощи перестали охранять, и дубовые леса стали сводить еще интенсивнее.
В настоящее время широколиственные леса Европейской части России можно считать исчезающими лесными экосистемами. Всего несколько веков назад они занимали большую часть Европы и были одними из самых богатых и разнообразных по видовому составу, а сейчас, судя по данным учета лесного фонда, на территории нашей страны их осталось не более 100 тысяч га. За несколько столетий площадь этих лесов сократилась в десятки раз.
Широколиственные леса на территории равнины европейской части России состоят в основном из дуба черешчатого. Другие породы, как липа, клен, ясень, ильмовые образуют примесь; лишь изредка в некоторых местах какая-либо из этих древесных пород начинает доминировать над другими, а дуб иногда и вовсе отсутствует. Реже встречаются и чистые насаждения, состоящие из одной, обычно сопутствующей дубу, породы. Однако в типичном виде широколиственные леса состоят именно из дуба, как господствующей породы, а другие перечисленные породы растут лишь в виде большей или
240
меньшей примеси. Поэтому область распространения широколиственных лесов на территории равнины европейской части России в сущности нигде не выходит за пределы ареала дуба.
Широколиственные леса распространены преимущественно в южной части лесной зоны и в северной части лесостепной подзоны, т. е. они тянутся в виде широкой полосы, начиная от Волыни через Киев и Чернигов на Брянск, Орел, Калугу, Тулу, Рязань, к Нижнему Новгороду и Казани. К северу от этой полосы, далее за Волгу значение широколиственных пород в лесном покрове очень сильно падает. Встречаются лишь небольшие массивы и отдельные рощицы их, или эти породы входят как единичная примесь в хвойные леса. К востоку от Урала дуб практически не встречается вплоть до широколиственных лесов южной части Дальнего Востока.
Несмотря на то, что широколиственные леса играют большую экономическую роль в хозяйстве и немало работ было посвящено их изучению, дать сколько-нибудь полную характеристику и классификацию их типов трудно, так как эти леса довольно разнообразны в типологическом отношении.
Прежде чем переходить к рассмотрению типов широколиственных лесов, остановимся на характеристике тех древесных пород, которые им свойственны. По сравнению с хвойными лесами, число древесных и кустарных пород, входящих в состав этих лесов, довольно велико. Мы начнем их рассмотрение с дуба.
Род- Quercus L.
Род Quercus (дуб) относится к семейству Fagaceae (Буковые), в которое входят еще 6 родов, в их числе Fagus (бук), и Castanea (каштан). Большинство видов семейства - вечнозеленые или листопадные деревья, часто значительной (до 60 м) высоты, и лишь немногие представители родов Castanea и Quercus - кустарники.
Палеонтологические находки свидетельствуют о том, что род Quercus довольно древний. Он появился в Европе и в Америке с нижнего мела. Однако дуб, произрастающий в европейской части России, с позиций палеоботаники не относится к древним растениям.
Многие специалисты считают дуб основным эдификатором доагри-культурных (неизмененных сельскохозяйственной деятельностью) широколиственных лесов Восточной Европы.
Современный ареал дуба существенно отличается от позднеголоценового. Примерно за последние 500 лет северная граница существенно сместилась к югу: дуб отсутствует на территории Карелии, Архангельской области и Коми. Южная граница сдвинулась к северу: дуб исчез из южных районов Украины, а восточная граница немного сместилась на запад (рис. 70).
241
Рис. 70. Ареал европейских видов дуба
Северный предел распространения дуба определяется как режимом основных ограничивающих климатических и почвенногрунтовых факторов, так и конкурентными отношениями его с другими растениями, особенно с елью. На обширных пространствах Русской равнины
самые северные местонахождения дуба - в долинах рек; по ним он нередко проникает на 200-300 км севернее, чем по междуречьям. Г.Ф. Морозов объяснял это лучшим прогреванием пойменных почв и их меньшей оподзолен-ностью. Немногим более двух веков назад дуб произрастал гораздо севернее, восточнее и южнее современной границы.
Система рода Quercus сложна, многократно подвергалась пересмотру и вряд ли может считаться окончательно устоявшейся. Представители рода Quercus (400-500 видов) - важные лесообразующие породы умеренных широт северного полушария и обычно лишь компоненты полидоминантных тропических и субтропических лесов. В Евразии богатством видов и разнообразием типов отличается Восточная и Юго-Восточная Азия. В Сибири, Центральной и Средней Азии дубов нет. Европа и Средиземноморье сравнительно бедны в видовом отношении. Значительное число видов дуба произрастает в Северной и Центральной Америке.
Большинство дубов - высокие деревья (25-30 м), на равнинах обычно выше, чем в горах. Отдельные экземпляры достигают громадной высоты (до 55 м) и имеют в возрасте 700-900 лет толстые (диаметром до нескольких метров) стволы. Немногие виды являются низкими кустарниками и даже кустарничками высотой около 0,5 м, например, дуб кустарниковый (Q. fruticosa) в Испании, дуб пушистый (Q. pubescens) на Кавказе.. Многие виды, будучи в
нормальных условиях роста высокими деревьями, в высокогорьях, в засушливых местах или при частых рубках и потраве скотом не вырастают выше
кустарника.
Корневая система дубов мощная с хорошо развитым стержневым корнем, достигающим в засушливых условиях глубины 10 (15) м. При наличии подстилающего горизонта, образованного плотными породами, а также на подзолистых или переувлажненных почвах корневая система становится поверхностной.
242
Особенности наружного слоя коры (перидермы) часто являются характерными видовыми признаками. У дуба черешчатого (Q. robur) кора с хорошо выраженными продольными глубокими извилистыми трещинами и короткими поперечными щелями, углубляющимися с возрастом; у дуба скального (Q. petraea) она разделена на отдельные пластинки.
Ветвление у дубов, особенно хорошо видное в зимний период, кажется очень резким и угловатым. Это происходит от частой смены направления роста побегов, обусловленной большим светолюбием дубов и стремлением дерева развивать листья только на освещенной части кроны. Характерный для дубов моноподиальный рост реализуется очень редко в связи с частым повреждением верхушечных почек и "перевершиниванием" побега за счет развития боковых почек. Такая смена может происходить несколько раз в течение одного вегетационного периода. У большинства видов почки пятигранные с коричневыми чешуйками, расположенными в 5 рядов. У основания почек часто развиты 2 узких прилистника.
Листья обычно скучены в верхней части побега, иногда очень густо, что приводит к так называемому ложномутовчатому листорасположению. Всегда простые, цельнокрайние, зубчатые, лопастные или перисто-рассеченные с перистонервным жилкованием, в среднем 6-15 см длины.
Листья летних ("Ивановых") побегов, появляющиеся из спящих почек у основания или верхушки прошлогоднего побега в середине вегетационного периода, часто резко отличаются от листьев весенних побегов; обычно только последние имеют характерные для вида систематические признаки.
Листья большинства видов имеют ярко-зеленую окраску сверху и более бледную снизу. Бледный цвет нижней поверхности листьев обычно обусловлен ее опушением или реже развитием воскового налета.
Молодые листья всех дубов в той или иной степени опушены, хотя бы кратковременно. Густота опушения листьев в значительной степени изменяется от вида к виду, но иногда ее степень варьирует у одного и того же вида в течение вегетационного периода, в частности, у дуба пушистого.
Мужские соцветия дубов - цилиндрические рыхлые висячие сережки. Они развиваются в пазухах чешуевидных листьев вегетативной почки или в особых цветковых почках, находящихся в верхней части прошлогоднего побега, или в пазухах чешуйчатых листьев нижней части молодого побега. Сережки имеют тонкую гибкую, как правило, опушенную ось 3-15 см длины, на которой почти сидячие цветки, имеющие при основании мелкие пленчатые прицветники, располагаются в пучках по 3-7 или по одиночке.
Околоцветники мужских цветков мелкие (1,5-4 мм длины), зеленоватые или желтоватые симметричные или чаще асимметричные опушенные изнутри или снаружи. Тычинки (6-15) обычно с длинными свободными нитями; пыльники от яйцевидно-продолговатых до почти шаровидных, 0.5-1 мм длины.
Женские соцветия дубов морфологически представляют собой простой колос. Обычно цветки развиваются на неразветвленной жесткой укороченной оси, появляющейся в верхней части весенних побегов всегда выше мужских
243
соцветий. Женские цветки сидячие, с 1-2 прицветниками при основании, располагаются на оси поодиночке, околоцветники мелкие, малозаметные, обычно 6-, реже 3-9-лопастные, сидят непосредственно на верхушке завязи.
После раскрытия мужских цветков пыльца остается жизнеспособной около 5 суток. Пыление у крупных деревьев обычно продолжается несколько дней. Пыльца дуба в массовых количествах не переносится на большие расстояния, значительная часть ее оседает в том же лесу под кронами деревьев на расстоянии 200-400 м от пылящего дерева.
Семязачатки у большинства видов с одногодичным созреванием желудей заканчивают свое развитие через 1-2 месяца после опыления, когда желудь станет величиной с горошину. Созревание и опадение желудей в зоне умеренного климата происходит осенью и обычно длится 1,5-2 месяца. Их фактический урожай обычно составляет не белее 2-4 % потенциального из-за погодных условий и повреждения на ранних стадиях развития плодожоркой, дубовым долгоносиком. Принято считать, что плодоношение дубов имеет определенную периодичность: хорошие урожаи в Средиземноморье повторяются через 2-3 года, в Центральной Европе - через 4-7 лет, а на северо-востоке ареала дуба черешчатого через 10 и более лет. Однако, специальные наблюдения показали, что правильнее говорить об эпизодичности плодоношения дуба, зависящей в основном от погодных условий в период заложения соцветий и во время цветения, а также от степени развития популяций вредителей.
Плюска дубов развивается в конце цветения. Форма плюсок очень разнообразна: блюдцевидная, чашевидная, полушаровидная, цилиндрическая, иногда с широким или даже завернутым вниз отгибом; иногда она охватывает до 2/3 зрелого желудя, но чаще только его основание. Она никогда не покрывает плод полностью и никогда целиком не срастается с ним. Ее размеры варьируют от 5-6 мм до 5-6 см в диаметре. На своей наружной поверхности плюска покрыта короткими прижатыми или длинными оттопыренными, часто отогнутыми назад (дуб каштанолистный) чешуйками.
Плод дуба - односемянный орех (желудь). Формы желудей очень разнообразны: от веретеновидных до шаровидных, полушаровидных и уплощенно-дисковидных; преобладающими, однако, являются эллипсоидальная, элипсоидально-цилиндрическая и яйцевидно-цилиндрическая формы. Снизу желудь имеет рубцовую площадку, по которой он отделяется от оси. Оболочка желудя (перикарп) древеснеющая, голая и блестящая, бывает и шелковисто опушенная, обычно светло-коричневая.
Распространение желудей производится ветром, животными, водными потоками; по крутым горным склонам они могут скатываться вниз на значительные расстояния. Птицы (сойки, голуби, фазаны) и грызуны (белки, мыши, полевки), делая запасы, довольно далеко разносят желуди; часть запасов остается неиспользованной и может прорастать. Разумеется, большая часть урожая желудей остается под кронами родительских деревьев.
Прорастание у большинства видов дуба в основном "подземное", то есть семядоли не выносятся на поверхность лесной подстилки. В первый год
244
жизни всходы дуба достигают значительной высоты 10-30 см; но после этого, примерно до двадцатилетнего возраста дуб растет медленно. Максимальный прирост в высоту у дуба черешчатого в возрасте от 20 до 80 лет, монгольского - от 65 до 75 лет, затем величина вертикального роста существенно снижается и прирост древесной массы осуществляется главным образом за счет увеличения диаметра ствола и ветвей. Плодоношение начинается в зависимости от положения в лесном пологе с 40- 60 лет и может продолжаться в течение всей жизни дерева.
Древесина дубов тяжелая, прочная средней и высокой твердости, обычно в значительной степени усыхающая, имеет хорошо выраженное бурое ядро и более светлую узкую заболонь. Сосуды ядра у большинства видов заполнены тилами.
Дуб черешчатый - Q. robur L.
Мощное красивое дерево с широкопирамидальной или шатровой кроной, крепкими сучьями и могучим стволом. В насаждениях стволы полно-древесны и высоко очищаются от сучьев. В редком стоянии, например, в поймах рек, крона широкая, раскидистая, с далеко (на 15-20 м) уходящими в сторону толстыми искривленными сучьями.
Кора толстая, в коротких продольных трещинах, темно-серая. До 20-летнего возраста кора обычно остается гладкой, называется зеркальной. У старых деревьев корка достигает толщины 10 см. Молодые побеги дуба покрыты зеленовато-красноватой гладкой корой со светлыми чечевичками.
Почки яйцевидной формы, черепитчато покрыты множеством чешуек, скученны у концов побегов (рис. 71). Рост моноподиальный. Листья дуба весьма разнообразны по форме. Более характерна перисто-лопастная форма, число лопастей обычно не превышает 5-7 пар, не считая конечной, более крупной. У основания листья сужены, часто с двумя «ушками». Длина листьев 7-15 (30) см. В середине лета листья дуба становятся кожистыми, блестящими, с верхней стороны темно-зелеными. С нижней стороны они остаются более светлыми, голыми. В сентябре начинают желтеть, в первой половине октября становятся совершенно бурыми, закручиваются и опадают, образуя рыхлый слой подстилки.
Цветет дуб черешчатый одновременно с распусканием листьев. В период цветения заметны свисающие длинные сережки с десятью и более желто-зелеными тычиночными цветками. Сережки выходят по две-три или по одной из почек на концах годичных побегов или из основания молодых растущих побегов текущего года. Пестичные цветки мелкие, с булавочную головку, сидят у верхушек молодых побегов обычно по 2-3 шт. на заметных цветоножках.
Плод - желудь. На плодоножках сидят обычно 2 (3-5) желудей. Из прицветников образуется чашевидная плюска. Обычно до июля плюска покрывает желудь целиком, затем он показывается из плюски, а ко времени созревания она охватывает его лишь на треть - половину. Длина желудей 1,5-
245
3,5 см, вес 1000 шт. 2-3 (5) кг. Созревают желуди в центральных областях в конце августа - начале сентября. В сентябре начинают опадать.
Рис. 71. Ветвь дуба черешчатого во время цветения: 1 - ветвь с мужскими и женскими соцветиями; 2 - женское соцветие на длинной цветоножке (черешке); 3 -увеличенный женский цветок; 4 - мужские соцветия (сережки); 5 - увеличенная часть сережки (из Кернер фон Марилаун, 1902)
Область распространения дуба летнего обширна, он занимает большую часть западной половины Европы. В европейской части России ареал дуба имеет форму клинообразного языка, идущего от С.-Петербурга и Одессы и направленного острием к Южному Уралу.. Северная граница дуба черешчатого начинается выше С.-Петербурга, от Ладожского озера проходит несколько южнее Вологды, через Вят
ку на Урал, но не переходит его. Дальше всего на север заходит по берегам
рек. От южных отрогов Урала и от Оренбурга граница направляется к Саратову, вдоль Волги спускается несколько южнее Волгограда, отчасти заходя в южное Заволжье. Далее по Дону доходит до Ростова, затем через степи переходит к Запорожью.
Дуб черешчатый в горы поднимается лишь до высоты 900 м по сухим освещенным склонам. Такое распространение дуба показывает, что он тяготеет к условиям умеренного климата. В суровые зимы страдает от морозов.
Светолюбив, не выносит затенения сверху. Обычно с 5 лет становится особенно требовательным к свету, однако нуждается в боковом отенении. Затененный снизу и с боков, он растет быстрее и образует более прямые ство
лы.
На обширной территории своего ареала дуб растет в разнообразных климатических и почвенно-грунтовых условиях. Лучший рост его отмечен на темно-серых и серых лесостепных суглинках, оподзоленных и выщелоченных черноземах. Неплохо растет также на увлажненных гумусированных супесях, подстилаемых суглинками.
Удовлетворительно растет на южных черноземах и темно-каштановых почвах, предпочитая супесчаные разности. На бедных песчаных почвах растет плохо. Болотных кислых почв избегает. Заходит иногда на солонцы и удерживается здесь лучше других пород. Чем богаче почвы азотом, фосфором, калием, тем лучше растет дуб при наличии достаточного количества пресных грунтовых вод. На указанных почвах дуб развивает мощную корне
246
вую систему, уходящую до 8-10 м в глубь материнской породы, поэтому может черпать питательные вещества и влагу из глубоких горизонтов и переносить засушливый климат степи. В степях дуб произрастает преимущественно по балкам и склонам оврагов.
Размножается главным образом желудями. Желуди сохраняются обычно один зимний сезон. До того, как стебелек дуба покажется над поверхностью, корень довольно глубоко уходит в землю, поэтому пересадка дуба не рекомендуется.
Первые 5-6 лет дуб растет медленно, прирастая не более чем на 15 см в год. Затем прирост становится более энергичным и достигает 30-50 см в год. В это время дуб обгоняет по высоте кустарники и становится более чувствительным к недостатку света. Рост дуба в высоту продолжается довольно интенсивно до 60 70 лет, затем ослабевает. При густом стоянии дуб в раннем возрасте растет быстрее.
Дуб начинает плодоносить при свободном стоянии с 20 лет, в насаждении с 50 60 лет. Ежегодно можно найти плодоносящие деревья, особенно среди опушечных дубов, но обильные урожаи желудей повторяются не ежегодно и тем чаще, чем благоприятнее жизненные условия для дуба. Урожаи желудей в спелых дубовых насаждениях колеблются от 300 до 800 (2000) кг на 1 га. Часть желудей далеко разносится сойками, растаскивается мелкими зверьками или прорастает под подстилкой.
К 140 годам рост дуба в высоту почти прекращается, но прирост в толщину продолжается. Дуб живет иногда более 1000 лет, образуя стволы диаметром 4 м. В мачтовых дубовых рощах высота достигает 40 м.
Дуб черешчатый возобновляется и вегетативным путем. Поросль появляется преимущественно у шейки корня. Порослевые насаждения менее долговечны и менее производительны, чем семенные.
Дуб черешчатый образует множество форм. Особое внимание привлекают к себе поздние и ранние формы дуба. Дуб поздний распускает листья на две недели позднее раннего, поэтому не подвергается воздействию ранних заморозков, объеданию гусеницами непарного шелкопряда и др., что обуславливает лучшее его плодоношение и рост. У дуба раннего верхушечный побег повреждается заморозками. Обе разновидности дуба заканчивают вегетацию почти одновременно. Более короткий период вегетации дуба позднего компенсируется более высокой энергией роста и лучшей ассимиляцией.
Дуб поздний отличается более стройным и полнодревесным стволом, сжатой, высоко поднятой кроной. Древесина его обладает более высокими техническими качествами и большим объемным весом.
Дуб - ценнейшая порода. Особую ценность представляет его кольцесосудистая древесина со светло- или темно-бурым ядром и узкой светлой заболонью. Годичные кольца резко выделяются, так как ранняя, весенняя, древесина имеет крупные, видимые невооруженным глазом, сосуды. Наличие широких и узких сердцевинных лучей в древесине дуба способствует лучшей ее раскалываемое™.
247
Тяжелая и прочная древесина дуба применяется в судостроении, вагоностроении, строительстве гидротехнических сооружении, в столярном и мебельном производствах, для изготовления сельскохозяйственных машин, повозок, паркета, лыж, шпал, телеграфных столбов, рудничной стойки, фанеры и др. Высоко ценится в мебельной промышленности древесина дуба, пролежавшего долгое время на дне лесных рек в мокром песке, известная под названием черного мореного дуба.
Анатомические особенности древесины. Поперечный срез. Ядро имеется. Древесина кольцесосудистая. Граница годичных колец слегка волнистая, четкая. Просветы крупных сосудов ранней древесины чаще одиночные, округлые или эллиптические, в поздней древесине узкопросветные сосуды сгруппированы в виде "рисунка пламени" или дендритов. Волокна либриформа распределены по годичному кольцу неравномерно, но в поздней древесине их больше. Многорядные лучи расширяются при переходе из одного годичного кольца в другое. Паренхима обильная, апотрахеальная (диффузная и метатра-хеальная в виде тангенциальных полос в 1-3 клетки шириной). Менее выражена паренхима паратрахеальная (вазицентрическая). В сосудах ранней древесины многочисленные сомкнутые тилы. В сосудах поздней древесины тилы встречаются редко. Тангенциальный срез. Лучи двух размеров: многочисленные низкие однорядные и очень широкие многорядные (по 20-30, иногда до 40 клеток). Высота широких лучей до 5 мм. Встречаются агрегатные или расчленяемые трахеальными элементами широкие лучи. В молодых побегах многорядные лучи могут отсутствовать. Радиальный срез. Перфорации члеников сосудов простые. Межсосудистая поровость очередная. Поры между сосудами и клетками лучей крупные, с узким окаймлением, округлые или слабо угловатые. Лучи гомогенные, иногда слабо гетерогенные, состоят из лежачих клеток или с одним рядом квадратных клеток. Волокнистые элементы состоят из волокон либриформа, волокнистых и сосудистых трахеид. Встречаются призматические кристаллы в клетках широких лучей и осевой паренхимы.
В коре дуба и в других частях содержатся дубильные вещества. Особенную ценность представляет зеркальная кора, содержащая более 10 % дубильных веществ.
Второй по хозяйственной значимости вид дуба в европейской части России - дуб скальный (Q. Petraea Liebl.), называемый также зимним или сидячецветным. Это более требовательное к теплу растение, растет у нас вместе с дубом черешчатым на Северном Кавказе.
Спутники дуба
Род клен - Acer L.
Деревья или кустарники с супротивным листорасположением и ветвлением. Листья простые, большей частью пальчатолопастные или сложные из 3-5 (9) листочков. Соцветие - метелка, кисть или щиток; цветки обоеполые или тычиночные с рудиментарной завязью или без нее, обычно с кольцеобразным диском, и пестичные с недоразвитыми тычинками или совсем без них; растения однодомные, двудомные или полигамные; завязь 2-гнсздная. Плод - двукрылатка, при созревании распадающаяся на 2 крылатки, каждая с продолговатым односторонним крылом с наружной стороны гнезда завязи.
248
В роде до 150 видов, распространенных в Европе, в Передней и Средней Азии, в Гималаях, в восточной Азии, на островах Зондского архипелага, в Северной и Центральной Америке.
Клены - хорошие парковые декоративные деревья и кустарники с красивой листвой, разнообразной по форме и окраске, особенно в осеннее время (ярко-желтой, оранжевой или красной самых разнообразных оттенков). У некоторых видов декоративны и цветки, распускающиеся обычно одновременно или несколько позже листьев (у немногих видов раньше). Все виды - медоносы.
Многие виды растут быстро и являются ценными лесными породами; они в большинстве случаев не образуют сплошных насаждений, а встречаются как примесь к другим широколиственным древесным породам и реже - к хвойным. Клены требовательны к плодородию почвы. Большинство видов относительно теневыносливы, особенно в молодом возрасте.
Виды клена различаются по механическим свойствам древесины. Древесина в группе рассматриваемых нами видов умеренно тяжелая, крепкая, твердая, хорошо полируется, имеет желтоватую или розоватую окраску с шелковистым блеском, довольно декоративна, но без отчетливой текстуры. При сушке мало деформируется. Используется в качестве столярного материала для изготовления мебели, деталей машин, спортивного инвентаря. Особенно важна для производства музыкальных инструментов.
Некоторые виды клена, в числе которых клены остролистный, Траут-веттера, североамериканский клен сахаристый, могут формировать аномальную древесину с очень красивой текстурой. Хорошо известна текстура "птичий глаз", "волнистость", "струйчатость", "стеганная текстура". Такие аномальные древесины представляют собой неповторимый по красоте материал для дорогой мебели, уникальных подарочных изделий и отделки помещений.
Клен остролистный - Acerplatanoides L.
Дерево до 30 м высотой и до 1м в диаметре, с густой, широкояйцевидной кроной. Кора черно-серая, трещиноватая, не отслаивающаяся. Корневая система клена состоит из неглубоко идущего стержневого корня и обильных, далеко в стороны расходящихся боковых корней. Дает обильную поросль от пня.
Побеги голые, буровато-оливковые с рассеянными чечевичками. Почки яйцевидные, 7-10 см длиной. Листья простые, пальчато-лопастные. Число лопастей варьирует от 3 до 7, на лопастях имеются отдельные редкие зубцы. Вершинки лопастей и зубцов вытянуты в тонкие остроконечия; углы между лопастями довольно широкие. Листовая пластинка вписывается в окружность от 5 до 20 см в диаметре. Сверху листья голые, блестящие, темнозеленые, снизу более светлые. Осенью листья принимают очень красивую золотисто-желтую или оранжево-красную окраску. Черешок листа длинный, от 4 до 18 см. Соцветия - конечные, прямостоячие, щитковидные метелки (рис. 72).
249
cct jdft
250
Рис. 72. 1 - клен остролистный: 1а - побег с почкой и листовыми рубцами; 16 - соцветие - щитковидная метелка; 1в - побег с листом и дробными плодами (двукрылатками). 2 - клен ложноплатановый (явор): 2а - побег с почками и листовыми рубцами; 26 -соцветие кистевидная метелка; 2в - мужской цветок; 2г -женский цветок; 2д - побег с листьями и плодами (дробными крылатками). 3 - клен полевой: За - побег с почками; 36 -побег с продольными пробковыми наростами; Зв - женский цветок; Зг - мужской цветок; Зд - побег с листьями и дробными крылатками (угол расхождения двух крылаток - 180 ) (из Зуихина, Коровин, Тимофеенко, 2009)
Цветки зеленовато-желтые, 8-12 мм в диаметре, 5-членные. На одном растении обычно бывают тычиночные, ложно обоеполые и пестичные цветки в различных сочетаниях. Тычинок 8 (5-10), пестик с плоской голой завязью и двумя отогнутыми рыльцами. Клен остролистный цветет обычно в конце апреля - начале мая, одновременно с распусканием листьев. Плод - дробная крылатка, с расходящимися под тупым углом крыльями (около 120°), общая длина 8-10 см, ширина 1-1,5 см. Семена плоские, созревают в конце августа - начале сентября. Для получения хорошей всхожести посев нужно производить осенью свежесобранными семенами.
Ареал: Европейская часть России (на юг от линии Петербург - Вологда - Пермь - Екатеринбург), Средний и Южный Урал, Кавказ, Карпаты (в Крыму отсутствует), средняя и северная часть Западной Европы до южной части Скандинавского полуострова. Клен остролистный умеренно теневынослив, особенно в молодом возрасте, требователен к плодородию почвы и умеренному увлажнению, растет быстро, умеренно морозостоек, доживает до 200-250 лет. Чистых насаждений не образует, встречается в составе смешанных и хвойно-широколиственных Европейских лесов. Его участие, обычно, не превышает 2-3 единиц от общего состава насаждения, растет он вместе с дубом, липой, елью, осиной, сосной. В северной части ареала служит показателем плодородных почв.
Хозяйственное значение клена остролистного разнообразно. Древесина светло-желтоватая или слегка красноватая, тонкого и равномерного сложения, довольно тяжелая, твердая, мало коробится и растрескивается и легко обрабатывается инструментами, используется при изготовлении струнных инструментов и мебели.
Анатомические особенности древесины. Поперечный срез. Ядро отсутствует. Годичные кольца отчетливые. Древесина рассеяннососудистая. Просветы сосудов округлые, немногочисленные, одиночные, в радиально ориентированных цепочках и мелких группах. Волокнистые элементы представлены либриформом и реже встречающимися волокнистыми трахеидами. Группы волокон с толстыми и тонкими стенками чередуются и часто образуют характерный для кленов рисунок. Паренхима скудная апотрахеальная: в основном терминальная, а также диффузная и метатрахеальная в коротких цепочках. Тангенциальный срез. Лучи в основном 1-3 (4)- рядные. Гомогенные. Клетки лучевой паренхимы округлые. Радиальный срез. Перфорации простые. Межсосудистая поровость очередная. Поры крупные, округлые и полигональные. Поры между сосудами и клетками лучей многочисленные, округлые, простые. Лучи преимущественно гомогенные, состоят из лежащих клеток или иногда с одним рядом краевых квадратных клеток. Стенки сосудов имеют спиральные утолщения.
251
Клен остролистный является хорошим ранним медоносом (с одного дерева пчелы собирают около 10 кг меда). Этот вид включает много декоративных форм и широко используется в озеленении.
Род ясень - Fraxinus L.
Крупные листопадные деревья с тонкой светлой или бурой корой. Почки конические и шаровидные, черные, коричневые или ржаво-бурые. Листья супротивные, сложные, непарноперистые. Цветки в многоцветковых метельчатых соцветиях. Цветут до распускания листьев или одновременно с ним. Опыляются ветром. Цветки обычно без околоцветника (кроме ясеня белого -F. ornus L., который отличается от других видов белыми душистыми цветками), состоят из 2 тычинок и 1 пестика, часто происходит редукция тычинок или пестика и цветки становятся однополыми, а деревья - двудомными. Плод - одногнездная крылатая семянка - созревает осенью, распространяется ветром. Все виды ясеня светолюбивы, требовательны к почве, входят в состав широколиственных лесов. В роде насчитывается более 60 видов, которые произрастают в Европе, Средней Азии, Китае и в Северной Америке, в пределах бывшего СССР естественно растут 8 видов ясеня.
Ясень обыкновенный - Fraxinus excelsior L.
Ясень обыкновенный - дерево первой величины с прямым полнодревесным стволом, высотой до 40 м и до 1 м диаметром. Крона ажурная, округло-яйцевидная, высоко поднята. Кора светло-серая, у молодых деревьев гладкая, у взрослых - с красивым рисунком неглубоких продольных и поперечных трещин. Почки черные, матовые. Побеги толстые, в верхней части слегка сплюснутые, голые. Листья крупные, неопушенные, сверху темнозеленые, состоят из 9-15 яйцевидно-ланцетных, заостренных и пильчатых по краю листочков (рис. 73). Цветки в метельчатых пазушных соцветиях, чаще ложнообоеполые, но могут быть и двуполыми. Цветет одновременно с обли-ствением в середине мая. Плод - продолговатая крылатая семянка с винтообразно изогнутым крылом. Плодоносить в насаждениях начинают около 35-40 лет, на свободе - около 20-25 лет. Семенные годы бывают почти ежегодно. Созревают плоды к концу осени, но продолжают висеть на дереве всю зиму и лишь к весне опадают, служат кормом многим кочующим птицам.
Ареал ясеня обыкновенного совпадает с зоной широколиственных европейских лесов, за Волгу не переходит, встречается в лесах Кавказа, Крыма, Малой Азии. Ясень растет быстро, светолюбив, не газоустойчив, живет до 250 лет и более. Чистые насаждения образует не часто, обычно растет в смеси с другими широколиственными породами.
252
Рис. 73. Ясень обыкновенный: 1 - побег во время цветения с соцветиями из обоеполых, мужских и женских цветков; 2 - мужской цветок; 3 - обоеполый цветок; 4 - побег с листьями и плодами; 5 - плоды - крылатки
К климату ясень требователен и особенно боится заморозков. Хорошо развивается лишь на плодородных почвах; по требовательности к богатству почвы занимает одно из первых мест среди других древесных пород; обычно относится также к породам, особенно нуждающимся в извести; относительно лучше других пород он выносит и засоленность почвы. К влаге ясень не требователен, и в области степных дубрав он, вместе с дубом, растет на довольно сухих почвах.
Корневая система ясеня сильно
развита, однако стержневой корень слабо развивается, а боковые далеко расходятся в стороны, не углубляясь значительно в почву. Растет быстро.
Древесина ядровая, кольцесосудистая, тяжелая, плотная. Ценится в кораблестроении, мебельном производстве, при производстве особенно красивого фанерного шпона с волнистой текстурой. Кора идет на дубление кож и
используется в медицине.
Анатомические особенности древесины. Поперечный срез. Древесина ядровая с широкой заболонью, кольцесосудистая. Граница годичных колец гладкая, четкая. Просветы сосудов округлые, редкие, одиночные и парные. В поздней древесине просветы окружены клетками паратрахеальной паренхимы. Стенки волокон либриформа относительно тонкие. Стенки сосудов в поздней древесине - толстые. Осевая паренхима обильная, в основном паратрахеальная (вазицентрическая, крыловидная и иногда сливающаяся). Имеется также терминальная паренхима. Тангенциальный срез. Большинство лучей 2-3-рядные., присутствуют и 4-рядные. Клетки лучей округлые, одинакового размера. 3-4-рядные лучи веретеновидные. Высота лучей 2-30 клеток. Радиальный срез. Перфорации простые. Межсосудистая поровость очередная, поры мелкие многочисленные, овальные. Поры между сосудами и клетками лучей мельче, овальные, простые. Лучи в основном гомогенные, иногда слабогетерогенные, состоят из лежачих клеток или с одним рядом краевых квадратных клеток. Основная масса древесины состоит из волокон либриформа. Клетки вази-центрической паренхимы, контактирующие с сосудами, квадратные до слабо стоячих; не контактирующие - вытянутые вдоль оси стебля.
Род липа - Tffia Juss.
Род липа (сем. Tiliaceae) представлен деревьями с опадающими листьями. Листья сердцевидные или округло-яйцевидные. Жилкование соединяет пальчатое с перистым, совершенное с несовершенным. Цветки пятерного типа, обоеполые, собраны в соцветие - плейохазий. Соцветие снабжено длин
253
ным языковидным прицветным листом бледно-зеленого цвета. Кора с сильно развитым лубом и большим количеством лубяных волокон. Древесина без ядра, белая, легкая, мягкая.
Род насчитывает около 50 видов, в нашей стране естественно растут 16 и разводится в культуре около 11 видов. Наиболее распространенным видом в европейской части России является липа мелколистная.
Липа мелколистная - Tilia cordata Mill.
Дерево первой величины, до 40м высоты, до 1,5-2 м в диаметре. При свободном стоянии крона раскидистая, шатровидная, нижние ветви свисают, в насаждении крона высоко поднята, и нижние 2/3 ствола обычно очищены от сучьев. Кора на молодых стволах гладкая, у старых деревьев - темносерая, продольно-бороздчатая. Листья округлой формы, с сердцевидным основанием, заостренной вершинкой, зубчатые по краю; базальных жилок 5-6. Сверху листья голые, зеленые, снизу - сизоватые, с бородками рыжих волосков в углах жилок. Цветки около 1 см в диаметре, беловато-кремовые, имеют 5 чашелистиков и 5 лепестков, 20-30 тычинок, собранных в 5 пучков, и один пестик; завязь густо опушена. Цветки ароматные, содержат много нектара, собраны в соцветия, состоящие из 5-11 цветков. Прицветный лист продолговатый, 6-8 см длины, 1-2 см ширины. Цветонос прикреплен к нему на 1\3 длины. Липа цветет в середине лета, в Москве в первой декаде июля. Плод -округлый или грушевидный орех, при созревании опушенный, затем почти голый, около 7 мм в диаметре (рис. 74).
ша, встречается в Крыму и на
Рис. 74. Липа мелколистная: 1 - побег с листом и пазушным соцветием; 2 - соцветие; 3 - прицветный лист соцветия; 4 - обоеполый цветок в разрезе; 5 - тычинки; 6 - плод - орех; 7 - поперечный разрез плода с семенем
Корневая система сильно развита, имеет как стержневой корень, так и сильные боковые корни; поэтому липа не ветровальна.
Ареал: растет по всей Европейской части нашей страны, северная граница проходит по линии - Петербург, Вологда, Пермь, Екатеринбург, заходит в Западную Сибирь до Ирты-е. Растет в широколиственных и сме
шанных лесах вместе с дубом, кленом, елью, сосной, является почвоулучшающей породой, морозостойка, в молодом возрасте теневынослива, требовательна к плодородию почвы, умеренно влаголюбива, долговечна (доживает до 500-600 лет). Липа лучше мирится с подзолистыми почвами, чем дуб. Может переносить временный избыток влаги, но на заболоченных почвах не
254
растет. В естественных условиях размножается семенами и порослью от пня. Часто встречается в культуре, имеет много садовых форм.
Область распространения липы показывает, что ее надо отнести к породам холодостойким. Заморозков она не боится, распускается довольно поздно. По теневыносливости она занимает одно из первых мест. Ее густое облиствение и расположение листьев содействует тому, что липа сильно затеняет почву. На свободе ствол липы очищается от сучьев медленно. Благодаря своей теневыносливости она может расти в виде второго яруса и подлеска даже под тенью ели.
Роль липы мелколистной в лесах двоякая: или она входит в состав широколиственного леса, или в виде подлеска растет в дубовых и еловых лесах. В широколиственных лесах она присутствует обычно как примесь к дубу, причем на более оподзоленных почвах ее примесь делается значительнее, а местами она дает почти чистые липовые рощицы. Липа имеет огромное значение в пчеловодстве, с одного гектара липняков пчелы собирают около 800кг меда.
Древесина у липы очень рыхлая, легкая, мягкая, мало прочная. Поэтому на постройки она идет очень мало, но в последнее время часто используется для изготовления вагонки. Применяется для разного рода мелких поделок. Из липы делают специальную тару для масла, меда и т. п. Благодаря мягкости древесины из нее вырезают сувенирные ложки, игрушки, посуду. Раньше кора липы традиционно использовалась в крестьянском быту (лапти, мочало, рогожа).
Анатомические особенности древесины. Поперечный срез. Ядро отсутствует. Граница годичных колец отчетливая, слегка волнистая. Многорядные лучи при переходе из одного годичного слоя в другой расширяются, древесина рассеяннососудистая. Просветы члеников сосудов угловатые, многочисленные, сгруппированы в радиально ориентированные группы и цепочки. Волокнистые элементы тонкостенные, осевая паренхима апот-рахеальная: диффузная, терминальная, диффузно-агрегатная в виде тангенциальных цепочек и иногда сетчатая. В сердцевине имеются масляные каналы. Тангенциальный срез. Лучи низкие однорядные и довольно высокие 2-6-рядные, высота многорядных лучей варьирует от 30-50 клеток до очень высоких (до 1-2 мм). Встречается веретеновидная паренхима. Радиальный срез. Перфорации простые. Межсосудистая поровость очередная; поры очень мелкие и частые. Поры между сосудами и клетками лучей заметно мельче, многочисленные полуокаймленные. Основная масса древесины состоит из волокон либриформа и волокнистых трахеид. Выраженные спиральные утолщения сосудов и сосудистых трахеид. Лучи гомогенные, изредка гетерогенные, состоят из лежачих клеток или с одним рядом краевых квадратных клеток.
Род граб - Carpinus L.
Относится к семейству Betulaceae. Включает деревья и крупные кустарники. Кора гладкая, серая, с возрастом слегка растрескивается. Листья эллиптические или яйцевидные, с заостренной вершинкой и дважды-зубчатым краем, жилкование совершенноперистое. Цветет одновременно с распусканием листьев. Женские соцветия располагаются на концах побегов, мужские соцветия боковые. Мужские сережки состоят из одноцветковых дихазиев, жен-
255
ские - из двухцветковых дихазиев. Плод - ребристый орех в плюске или листовой обертке.
Род граб насчитывает 30-40 видов, из которых у нас естественно растут 6 видов, из них в европейской части России наиболее распространены граб обыкновенный и граб восточный (грабинник).
Граб обыкновенный - Carpinus betulis L.
Дерево второй величины, до 25м высоты до 70см в диаметре. Стволы ребристые, покрыты гладкой серой корой. Листья эллиптические, с заостренной вершинкой и округлым основанием, жилкование совершеннонервное. Край листа дважды остро-зубчатый (рис. 75).
Рис. 75. Граб обыкновенный: 1 - побег во время цветения с мужскими и женскими сережками; 2 - мужские сережки; 3 - мужской цветок; 4 - женская сережка; 5 - женский двухцветковый дихазий; 6 - побег с листом и соплодием; 7 - плод - орех в трехлопастной листовой обертке (из Strasburger, Noll, Schenck, Schimper, 1967)
О цветении граба смотри в характеристике рода. Плод - сплюснутый ребристый орех до 9 мм длины, сидящий у основания трехлопастной листовой обертки с крупной центральной лопастью.
Ареал: растет в широколиствен
ных лесах Карпат, Крыма и Кавказа, в западной части Белоруссии в Западной Европе.
Рост граба медленный, первые три года прирастает мало, но с 4-5 лет рост становится быстрее. С 80-90 лет рост граба уже прекращается, и естественная спелость наступает в 100-120 лет. Граб теневынослив, теплолюбив, мало требователен к плодородию и влажности почвы.
Корневая система довольно развита, имеет хорошо выраженный
стержневой корень. В горах Кавказа и Крыма поднимается до верхней грани
цы леса.
Граб чаще всего растет в смеси с дубом, буком и другими породами, но нередко образует и совершенно чистые насаждения; с буком и дубом растет в одном ярусе или образует второй ярус под ними. Применяется в озеленении (в живых изгородях) и лесной мелиорации.
Порода заболонная. Древесина серовато-белого матового цвета, центральная часть ствола по цвету не отличается от периферической. Границы годичных колец хорошо заметны по тонким беловатым извилистым линиям. Невооруженным глазом сосуды неразличимы. Древесина обладает большой
256
усушкой и разбуханием, подвержена сильному короблению и растрескиванию при сушке. Вместе с тем обладает большой твердостью и прочностью. В неблагоприятных условиях легко загнивает. Строительные детали и изделия даже в сухом виде сильно повреждаются ксилофильными насекомыми.
Древесина граба очень ценится для разного рода столярных изделий и представляет собой хорошие дрова. На строевой лес, по неправильности и кривизне стволов, мало пригодна.
Анатомические особенности древесины. Поперечный срез. Ядро неразличимо. Граница годичных колец волнистая, четкая. Древесина рассеяннососудистая. Просветы сосудов немногочисленные, округлые, в радиальных цепочках, собранных в широкие радиальные ленты, отделенные друг от друга полосами из волокнистых элементов и агрегатных лучей, ели они имеются. Осевая паренхима обильная, апотрахеальная: терминальная, диффузная и диффузно-агрегатная в виде 1-2-рядных тангенциальных и косотангенциальных цепочек. Агрегатные лучи располагаются неравномерно, встречаются часто, но не всегда. Тангенциальный срез. Лучи в основном 1-2-рядные, встречаются 3-4-рядные. Однорядные лучи составлены из 10-20 клеток в высоту, многорядные лучи высотой до 40 клеток. Клетки лучевой паренхимы овальные. Радиальный срез. Перфорации простые. Межсосудистая поровость очередная, поры частые, округлые, овальные и полигональные. Поры между члениками сосудов и клетками лучей крупнее, с зауженным окаймлением. Волокнистые элементы представлены волокнистыми и сосудистыми трахеидами и волокнами либриформа. Имеются спиральные утолщения на стенках сосудов и сосудистых трахеид. Лучи в основном слабо гетерогенные, встречаются гомогенные. Состоят из лежачих клеток с 1 рядом краевых квадратных клеток или только из лежачих.
Род вяз (ильм) - Ulmus L.
Представлен листопадными деревьями различной величины, иногда до 40 м высотой и до 2 м в диаметре. Кора на стволах в молодости гладкая, с возрастом становится трещиноватой. Побеги коленчатые. Листорасположение очередное. Листья эллиптические, у основания часто неравнобокие, расположены двухрядно. Край листа зубчатый или двояко-зубчатый, жилкование совершенно-перистое.
Цветки обоеполые с колокольчатым околоцветником коричневофиолетового цвета, рассеченным на 4-9 лопастей. Пестик с двухгнездной верхней завязью и двураздельным рыльцем; количество тычинок соответствует лопастям околоцветника, тычинки выступают над краями околоцветника. Длина цветоноса и количество тычинок у разных видов различно. Плод -чечевицеобразная семянка (орех), окруженная перепончатой крылаткой с выемкой на вершине (рис. 76). Вязы цветут до распускания листьев (в средней полосе европейской части России это обычно конец апреля), плоды созревают через месяц после цветения и сразу же опадают, требуют немедленного
посева.
257
Рис. 76. Морфологические признаки 4 видов вяза: 1 - вяз гладкий: 1а - побег с цветочными и листовыми почками; 16 - побег во время цветения; 1в - цветок (ув.); 1г - побег с листьями; 1д - побег с плодами; 1е - плод - крылатая семянка (крылатый орех) с реснитчатым краем. 2 - вяз голый: 2а - побег во время цветения; 26 - цветок; 2г - плоды; 2д -крылатая семянка. 3 - вяз граболистный: За - побег с листьями; 36 - ветвь с пробковыми выростами коры; Зв - цветок; Зг - плод - крылатая семянка. 4 - вяз приземистый: 4а - побег во время цветения; 46 - отдельный цветок; 4в - побег с листьями; 4г - плод - крылатая семянка (крылатый орех) (из Зуихина, Коровин, Тимофеенко, 2009)
Кроме стержневого корня, который уходит далеко вглубь, имеются мощные боковые корни, которые уходят далеко в стороны и вниз; поэтому ильмовые не ветровальны. После срубки дают обильную поросль от пня.
258
Макроскопические признаки древесины вязов. Заболонь желтоватобелая иногда до коричневатого или красновато-коричневого цвета, включает 10-20 годичных колец. Ядро от светло-коричневого до шоколаднокоричневого оттенка. Годичные кольца разделены зонами крупных, видимых невооруженным глазом, сосудов ранней древесины. В поздней древесине сосуды мелкие.
Древесина крепкая, упругая, хорошо гнется, используется в мебельном производстве и для различных мелких поделок.
Анатомические признаки древесины. Поперечный срез. Ядро хорошо выражено. Граница годичных колец гладкая, четкая, древесина кольцесосудистая. В ранней древесине просветы сосудов редкие, крупные, одиночные или парные, реже в коротких тангенциальных цепочках. В поздней древесине просветы мелкие, всегда в группах, окруженные сосудистыми трахеидами и паратрахеальной паренхимой, образуют короткие тангенциальные и диагональные полосы ("ульмоидный" рисунок просветов). Одиночные просветы крупные. Осевая паренхима обильная, в основном паратрахеальная. В ранней древесине паренхима вазицентрическая, в поздней - крыловидная, иногда сливающаяся. Имеется и апотрахеальная (терминальная) паренхима. В сосудах ранней паренхимы встречаются ти-лы. Тангенциальный срез. Лучи довольно низкие, обычно 3-6-рядные, 1-2-рядные и 7-рядные встречаются редко. Высота многорядных лучей 30-69 клеток. В клетках лучей встречаются крупные кристаллы оксалата кальция. Радиальный срез. Перфорации конечных стенок у члеников сосудов простые. Межсосудистая поровость очередная; поры крупные, округлые. Поры между сосудами и клетками лучей мельче, овальные и округлые, простые и с зауженным окаймлением. Лучи в основном гомогенные, встречаются и слабо гетерогенные; состоят из лежачих клеток или с одним рядом краевых квадратных. Волокнистые элементы состоят из либриформа и сосудистых трахеид. Видна спиральная штриховатость сосудов и сосудистых трахеид.
Род объединяет около 30 видов. Далее рассмотрим 4 вида, из них первые два имеют наибольшее распространение на территории нашей страны.
Вяз гладкий -Ulmus laevis Pall.
Дерево высотой до 35м, до 1м в диаметре, с широкоцилиндрической кроной, тонкими ветвями и буро-коричневой, чешуйчатой коркой. Листья эллиптические или обратнояйцевидные, резко асимметричные (с сильно неравнобоким основанием), сверху гладкие, снизу опушенные. Край листа двоякопильчатый. Жилки второго порядка заканчиваются в зубцах, обычно не раздваиваясь. Цветки, мелкие, 4-5 мм в диаметре, располагаются пучками на длинных цветоносах, до 2см длины (самые длинные из всех рассматриваемых видов). Околоцветник коричневый, 8-лопастной, 8 тычинок с фиолетовыми пыльниками, беловатые концы раздвоенного рыльца, выступают из околоцветника. Крылатка плода эллиптической формы, с реснитчатым краем, располагается на длинной плодоножке, семянка (орех) размещена в центре крылатки.
Ареал: естественно растет в европейских смешанных и широколиственных лесах, а также в Крыму и на Кавказе, особенно часто встречается в пойменных лесах. Чистых насаждений не образует. Обычно спутник дуба, но встречается как редкая примесь и в хвойных насаждениях. Северная граница
259
распространения вяза гладкого в нашей стране проходит по линии - Петрозаводск, Вологда, Киров, Екатеринбург, на восток за Урал не переходит. Вяз теневынослив, требователен к влажности почвы и мало требователен к влажности воздуха, любит плодородные почвы, умеренно теплолюбив. В культуре рекомендован как сопутствующая порода для лесных полос Нижнего и Среднего Поволжья, Северного Кавказа, Приазовья и юга Украины, а также используется в озеленении, хотя довольно часто повреждается грибным заболеванием именуемым «голландская болезнь».
Вяз голый (вяз шершавый, ильм горный) - U. glabra Huds. (U. scabra Mill.; U. montana With.).
Дерево, высотой до 30м и до 2м в диаметре. Кора с неглубокими трещинами, темно-коричневая. Ветви толще, чем у вяза гладкого, побеги щетинисто-опушенные. Листья эллиптические или вытянутые обратнояйцевидные, с коротко заостренной вершинкой; часть листьев имеет на вершинке три крупных зубца. Край листа двоякопильчатый, с острыми оттянутыми к вершине зубцами. Листья сверху и снизу грубо-шероховатые, асимметричные, боковые жилки на концах обычно раздваиваются. Черешок листа очень короткий, от 1 до Змм. Короткостебельчатые цветки собраны в большие, плотные пучки. Околоцветник удлиненный, бокаловидный, буроватокоричневый, реснитчатый. Тычинок 5-6; пыльники фиолетовые. Крылатка крупная, до Зсм длиной, обратнояйцевидной или эллиптической формы, голая (без опушения). Семянка (орех) расположена в центре крылатки.
Ареал: такой же, как и у вяза гладкого, растет по поймам рек в равнинной Европе, но встречается гораздо реже. А в горах Кавказа и Крыма встречается чаще, чем вяз гладкий.. В горах он растет по ущельям и поднимается на высоту до 1700м над уровнем моря. Он более требователен к плодородию и влажности почвы и воздуха. Древесина и ее хозяйственное использование такие же, как и у вяза гладкого.
Морфологически близок к вязу шершавому вяз лопастной (U. laciniatd), который растет в лесах Приморья.
Вяз граболистный (вяз листоватый, берест, карагач) - U. carpinifolia Rupr. ex Suchow. (U. foliaceae Gilib.).
Дерево, высотой до 30м, до 70 см в диаметре, с шарообразной кроной, образуемой тонкими поникающими ветвями. Кора бурая, продольнотрещиноватая. Побеги тонкие, гладкие, окрашены в красноватый или желтоватый цвет, иногда с хорошо выраженными пробковыми наростами. Почки небольшие, тупые, черно-бурые, иногда опушенные. Листья плотные, кожистые, обратнояйцевидные, неравнобокие у основания, с заостренной вершинкой, сверху голые, снизу по жилкам жестковолосистые. По жилкам имеются красные точечные железки. Некоторые жилки второго порядка раздваиваются у края листа. Листья сидят на длинных, до 2см, голых, а иногда слабо опушенных черешках. Цветки мелкие, на очень коротких цветоносах (короче,
260
чем у ильма горного), почти сидячие. Околоцветник 4-, 5-надрезный, ржавокрасный, покрыт белыми ресничками. Тычинок 4-5, пыльники рыжеватокрасные. Крылатка обратнояйцевидной формы, до 2см длиной (иногда меньше), семянка расположена ближе к вершине крылатки, часто покрыта красноватыми железками.
Ареал: северная граница ареала вяза граболистного проходит по линии - Киев, Чернигов, Курск, Воронеж, Тамбов, Самара; растет в Крыму и на Кавказе; в степной зоне всегда растет вместе с дубом. Теплолюбив, требователен к плодородию почвы, засухоустойчив, что объясняется его мощной корневой системой, идущей вглубь до 8м. Легко образует корневые отпрыски, вследствие чего пригоден для облесения оврагов и балок и создания защитных полос. Плотная древесина береста с широким красно-бурым ядром и узкой желтоватой заболонью очень красива, используется для мелких поделок.
Вяз граболистный имеет много форм строению кроны и окраске листьев.
Вяз приземистый - U. pumila L.
Дерево, до 16м высоты. Крона шатровидная. Кора серая трещиноватая, побеги очень тонкие, голые с мелкими почками до 2мм длины. Листья мелкие, 1-5 см длины, симметричные, кожистые, по краю зубчатые, с острой вершинкой. Цветки очень мелкие, сидячие, с 3-5 тычинками, пыльники фиолетовые. Крылатка округлой формы, слегка неравнобокая, часто в ширину больше, чем в длину, около 1,2см в диаметре. Орешек расположен в центре крылатки.
Ареал: юг Восточной Сибири, Дальний Восток, Средняя Азия и Монголия. Растет по сухим каменистым почвам и галечникам в долинах рек, иногда в горах. Вяз приземистый хорошо возобновляется порослью, имеет глубокую корневую систему, выносит засоление почвы, рекомендован для создания защитных полос южного Поволжья и Заволжья.
Типы широколиственных лесов
Рассмотрев отдельные виды древесных растений лесостепной и юга лесной зон России, остановимся на типах широколиственных лесов, в состав которых входят эти породы.
В случае наибольшей сложности широколиственного леса, которая обыкновенно соответствует плодородным почвам и выражена особенно хорошо в подзоне лесостепи или на темно-серых лесных суглинках, обобщенная картина лесных растительных ассоциаций будет следующей.
Первый ярус обычно состоит из дуба, с большей или меньшей примесью липы, клена остролистного, ясеня и ильмовых. Иногда имеется примесь осины и реже - березы. Второй ярус состоит из дикой яблони, дикой груши, полевого клена, изредка черемухи и даже рябины. Второй ярус значительно
261
реже первого. Третий, подлесочный, ярус слагается из лещины, бересклета бородавчатого, бересклета европейского, крушины слабительной, клена татарского и других кустарников. Четвертый ярус - травяной покров, состоит из пролески, хохлатки, медуницы, ветреницы лютиковой, сочевичника весеннего, золотистого лютика, фиалок, сныти, папоротника мужского, колокольчика тенистого, осоки волосистой, звездчатки, копытеня, ландыша, душицы, костяники и др. Пятый ярус, моховой, довольно редкий, представлен небольшим числом видов.
Наши дубравы можно разделить на две категории, на так называемые высокоствольники - семенного происхождения, и низкоствольники, порослевые, возникшие из поросли от пней после рубки. Обычно побегов отходит от пня несколько, поэтому естественно, что порослевой низкоствольник будет иметь более густой и к тому же одновозрастный относительно быстрорастущий, но недолговечный древостой.
Строение высокоствольной дубравы отличается от низкоствольной следующими важными чертами. Высокоствольная, семенная, дубрава обычно разновозрастна. Поэтому полог старого семенного широколиственного леса очень неравномерен, что еще обуславливается тем, что и дубы, и спутники дуба неодинаково затеняют почву. Наконец, деревья второго яруса, которые распределены нередко очень неравномерно под пологом, создают еще большую неравномерность затенения. В силу этого, в таком широколиственном лесу нет такого равномерного полога и такого выдержанного затенения, какой наблюдается в чистых хвойных лесах. Это же обстоятельство приводит к большой неравномерности в сложении травяного напочвенного покрова.
При продвижении с севера на юг из лесной зоны в лесостепь, начиная от северных пределов своего распространения, дуб сначала встречается лишь как небольшая примесь к хвойным лесам, придерживаясь по преимуществу долин рек. Несколько южнее он начинает чаще встречаться и вне пойм, образуя местами целые рощи.
Чем далее к югу, тем большая доля широколиственных лесов в общей лесной площади, однако значительное участие в лесах они начинают принимать лишь начиная с условной границы, идущей от Белоруссии к Москве, Нижнему Новгороду, Казани, к югу от р. Камы и Белой.
ЛЕСА СЕВЕРНОГО КАВКАЗА
Географическое положение Кавказа, соседство двух морских бассейнов, степных и пустынных пространств, своеобразие геологического прошлого и строения, громадное разнообразие геоморфологических форм и климатических особенностей создали крайне пестрые растительные и почвенные покровы.
Главный Кавказский хребет простирается с СЗ на ЮВ и имеет в длину 1490 км; начинаясь от Анапы, он, идя сильно извилистой водораздельной линией, кончается на Апшеронском полуострове. Часть Кавказа, лежащая к се
262
веру от Главного хребта, называется Северным Кавказом; часть Кавказа, находящаяся к югу от Главного хребта, носит название Закавказья.
Для Кавказа очень характерна вертикальная зональность растительного покрова. Последняя является следствием того, что по мере поднятия местности над уровнем моря изменяется климат; он все больше и больше уклоняется в сторону холода. На Кавказе, в нескольких десятках километров от берега Черного моря, высятся горы, покрытые снегом или льдом. На этих ничтожных расстояниях климат быстро сменяется от жаркого, субтропического до холодного, арктического. Параллельно этому меняется почвенный и растительный покровы.
В более сухих и жарких местах восточного и южного Закавказья нижний пояс занят полупустынной растительностью. Лишь вдоль рек, на сравнительно влажных почвах располагаются полосы так называемых тугайных лесов.
Следующий по высоте пояс, это - пояс степей. Степи покрывают равнинные и всхолмленные части Северного Кавказа и Восточного Закавказья. Здесь, среди травянистой ксерофитной растительности, леса расположены лишь вдоль рек; иногда в оврагах или сменяются зарослями кустарников.
Выше в горы, до 300-700 м, идет пояс лесостепи. Примерно от 700 м идет лесной пояс. Он поднимается до 2000-2300 м. Однако на всем своем протяжении лес не остается однородным. В нижней полосе лесного пояса преобладают дубовые леса. Эта нижняя полоса поднимается до 600-700 м. С 600-700 м и до 1200-1400 м простирается полоса, где господствуют буковые леса. В Западном Закавказье и в западной части Северного Кавказа с 1000— 1200 м они сменяются полосой елово-пихтовых лесов, которые в свою очередь на высоте 1900-2300 м уступают место корявым лесам из бука и березы, где древесные породы часто растут как кустарники.
В этой полосе, или поясе, участки согнутого снегом леса чередуются с зарослями кустарников и луговыми пространствами. Субальпийские луга в большей части Кавказа очень высокотравны и пестры по своему сложению. С высоты 3000 м они сменяются низкорослыми травянистыми растениями, дающими плотные дернины: начинаются альпийские луга. Постепенно теряя связность дерна, они поднимаются до границы вечных снегов и льдов.
Такова общая схема зонального распределения растительности на Кавказе. Однако не следует представлять себе, что пояса растительности сменяют друг друга на вполне определенных высотах: они всегда заходят один в другой большими или меньшими языками. Это зависит в основном от рельефа, от орографии (строения горного рельефа) и создаваемых им микроклиматов; здесь играют большую роль направление отдельных хребтов, экспозиция и крутизна их склонов; значительную роль приобретают горные породы и их свойства. Эти осложняющие влияния вызывают захождение одного пояса в другой, а внутри поясов пестрое, но закономерное распределение растений, образующих во взаимоотношении с внешней средой и друг с другом определенные фитоценозы, слагающиеся в экологические ряды.
263
Лесная древесная растительность, как и экотопы Кавказа, отличается необыкновенным разнообразием. Только деревьев первой величины, образующих верхние яруса древостоев, здесь больше двадцати, еще многочисленнее виды древесных, образующие подчиненные яруса. Большинство древесных растений этого региона имеет то или иное хозяйственное значение, многие являются важными объектами лесной промышленности. К сожалению, в одной лекции, посвященной растительности Кавказа, невозможно сколько-нибудь подробно описать это разнообразие и мы вынужденно остановимся лишь на нескольких наиболее хозяйственно важных видах-лесообразователях.
Важнейшие лесообразующие породы
Пихта кавказская - Abies nordmanniana (Stev.) Spach.
Естественно распространена только на Кавказе. Пихта кавказская - дерево первой величины; достигает высоты 60-70 м и диаметра более 2 м. В нашей стране это самое крупное дерево. Формирует прекрасные колонновидные стволы со сравнительно узкой пирамидальной кроной. Кора стволов пихты кавказской уже с 80 лет начинает растрескиваться, на ветвях и в верхней части ствола кора серая гладкая, а на молодых побегах - буро-красная, с желтоватым пушком. Почки у пихты кавказской яйцевидно-шаровидные с буро-красными, несколько пушистыми чешуями, незасмоленные.
Хвоя расположена двухрядно, на побегах, растущих вверх, - сидит спирально. Хвоя широкая (2 мм); достигает длины от 20 до 40 мм, плоская, темно-зеленая сверху, с 2 белыми полосками на нижней стороне; на боковых побегах хвоя притупленная или выемчатая; прикрепляясь к побегам, не имеет свойственных ели "подушечек" (рис. 77).
Опыление происходит в начале мая. Мужские стробилы узкоцилиндрические, до 20 мм длины, образуются на побегах прошлого года. Женские -сидят по одиночке на вершине прошлогодних побегов; кроющие чешуйки длинней семенных, с шиловидным отростком, выглядывают наружу. В пазухе последних образуется по две семяпочки. После оплодотворения женские стробилы разрастаются в деревянистые шишки, прямо стоящие на ветвях. Шишки достигают величины 12-20 см, при диаметре 4-5 см; кроющие чешуйки удлиненные, с округлой зубчатой верхушкой, имеющей длинное острие, выдающееся наружу из-под семенных чешуй и загнутое вниз.
Семенные чешуи клиновидные, сильно расширенные кверху, с боковыми выступами и небольшим килем, опушены снаружи. При основании чешуй образуется по 2 сильно смолистых продолговато-клиновидных угловатых семени, почти кругом объятых основанием крылышка. Семена созревают в год «цветения»; шишка при созревании рассыпается, а на ветвях еще долго остаются торчать стержни шишек. Стробилы и шишки сосредотачиваются исключительно в верхней части кроны. Всходы имеют 4-7 линейных семядо
264
лей, несколько выемчатых на верхушке и с белыми полосками на верхней стороне.
Рис. 77. Пихта кавказская, ветвь со зрелыми шишками, хорошо видны кроющие чешуйки, выступающие из-за семенных. Справа - рассыпающаяся шишка (из Strasburger, Noll, Schenck, Schimper, 1967)
Распространена пихта в западной части Кавказа. На северо-западе она начинает встречаться единичными деревьями. На северо-восток не переходит за водораздел Кубано-Терского поднятия.
Пихта - дерево исключительно высокогорное; отдельными экземплярами плохого роста в лесах Чер
номорского округа начинает встречаться под буком и каштаном с высоты 300 м, но основные массивы ее, как и в прочих частях Кавказа, находятся на высоте 1200-1900 м, где она граничит с высокотравной альпийской областью или с субальпийской опушкой леса из бука и березы. В нижней полосе своего
распространения, придерживаясь преимущественно затененных склонов, пихта растет в смеси с буком, грабом, кленами, ильмом и реже с каштаном. Выше она господствует над буком. С высоты 1700 м к пихте начинает примешиваться клен высокогорный (Траутфеттера) рябина; увеличивается количество бука. Деревья сильно уменьшаются в росте.
Пихтовые леса на высоте около 1200 м чрезвычайно величественны и мощны, высокие, колоннообразные, серые стволы пихт поднимаются до средней высоты 40 м, имея средний диаметр 60-80 см, с высокой сомкнутостью крон. Встречаются участки с запасом до 2000 м3 на 1 га. Кроны пихты обильно одеты бородатым лишаем, свешивающимся длинными космами с ветвей. В лучших условиях произрастания на высоте 1000-1500 м на бурых лесных подзолах пихта имеет большую примесь бука, находящегося почти во втором ярусе (высота яруса бука 30-35 м), здесь же в составе есть граб и небольшая примесь явора (клена белого), клена остролистного, черешни и ильма. Благодаря наличию толстого слоя опада и сильному затенению почвы травяной и моховой покровы мало развиты. Эти мертвопокровные пихтарники наиболее производительны.
В более сухих условиях пихта образует типы леса с господством злаков в напочвенном покрове. Начиная с высоты 2000 м пихта дает очень низкий
265
бонитет. Древостой заметно обогащается кустистым буком, высокогорным кленом, рябиной, березой.
Таким образом, типы пихтовых лесов сменяют друг друга по мере возрастания высоты над уровнем моря, а также вследствие большей сухости и каменистости почв. Если от мертвопокровного пихтарника проследить изменение типов леса в соответствии с постепенным возрастанием влажности почв, их глубины и рыхлости, то увидим также ряд типов леса, имеющих свои высотнозамещающие разности.
В настоящем изложении мы остановились лишь на нескольких, часто встречающихся, типах кавказских пихтачей. На самом деле в разнообразных и часто меняющихся высотных и почвенных условиях на фоне разной атмосферной и почвенной влажности насаждения с участием пихты кавказской представлены большим множеством типов леса. Разнообразию типов леса способствует богатство видового состава древесной растительности, формирующей леса Кавказа.
Ель восточная - Picea orientalis (L.) Link.
Единственная ель, дико растущая на Кавказе. Восточная ель - дерево первой величины, с узкой пирамидальной кроной. Она достигает 2 м в диаметре, 60 м высоты и доживает до 400 лет. Ветви расположены кольцеобразно, на одном годичном побеге могут возникать дополнительные боковые межмутовочные побеги, которые маскируют мутовчатость ветвления. Молодые побеги красноватые или желтовато-серые, с бурыми волосками; со временем кора становится пепельно-серой, а на старых стволах образуется трещиноватая темно-серая корка. Почки с красноватыми несколько волосистыми чешуйками.
Ель имеет короткую (не длиннее 10 мм) тупо-четырехгранную, темнозеленую, глянцевитую хвою, сидящую спирально на невысоких четырехгранных подушечках, остающихся на побегах после опадания хвои (рис. 78). Устьица расположены на всех гранях хвои и заметных полос не дают. Мужские конические стробилы образуются на прошлогодних побегах. Женские -в виде шишечек на вершине побегов по одиночке. В пазухе каждой семенной чешуи находятся по две семяпочки. После оплодотворения шишка увеличивается в размере, достигает длины 5-15 см и повисает. Цвет ее светло-бурый. Сильно разросшиеся выпуклые семенные чешуи к этому времени одревесневают; они имеют почти округлую цельнокрайнюю вершину; кроющие чешуи мало заметны. За каждой чешуей находится 2 обратнояйцевидных бурых семени, снабженных продолговатым крылышком, имеющим ложкообразное углубление, в котором и помешается семя.
Распространена ель восточная в западной части Кавказа. Встречается по обоим склонам Главного хребта. Леса из восточной ели находятся между 1300 и 2100 м высоты над уровнем моря. Составляет древостои вместе с буком, грабом, ильмом, кленами, пихтой, реже с сосной.
266
Рис. 78. Ель восточная: 1 - побег с короткой хвоей, в нижней части побега видны листовые "подушечки"; 2 - сигарообразная шишка с выпуклыми округлыми семенными чешуйками; 3 -поперечное сечение хвои (рис. Е.И. Тимофеенко)
Типы леса, образуемые елью восточной, аналогичны типам лиса, которые формируются пихтой.
Привязанность восточной ели к западной части Кавказа и высокогорной области указывает на высокие требования ее к влажности воздуха. Почва в зоне распространения ели имеет бурый цвет и подзолистый характер. К глубине и минеральному богатству почв ель не требовательна, хотя более связана с известковыми почвами.
Древесина белая легкая, мягкая, легко колется, дает строительный и пиловочный материал; имеет прекрасные резонансные качества и потому идет на музыкальные инструменты. По анатомическим признакам практически не отличается от ели европейской. Кора пригодна для получения дубящих веществ, содержит около 8 % танинов.
Дуб скальный - Quercus petraea (Mattuschka) Liebl.
Семейство Fagaceae. Русские дополнительные синонимы дуба скального - дуб зимний, дуб сидячецветный. Близок к дубу черешчатому, морфологически отличается от него длинным (до 2 см) черешком листа и короткой плодоножкой желудя; желуди почти «сидят» на побеге, кроме того, у дуба скального более жесткий кожистый лист с клиновидным основанием и на нижних лопастях нет свойственных черешчатому дубу «ушек».
Дуб скальный к возрасту спелости может достигать больших размеров, чем дуб черешчатый; стволы при совместном произрастании часто имеют более правильную форму, но менее долговечен, чем дуб черешчатый.
На Кавказе занимает Северное Предкавказье в его лесостепной части, поднимаясь в горы до высоты около 1800 м. В Западное Закавказье он выходит между Сочи и поселком Береговая, на востоке его ареал включает и горную часть Дагестана. Кроме того, растет в Крыму, юго-западной Украине и в Западной Европе. К теплу более требователен, чем черешчатый дуб.
В Крыму и на Кавказе скальный дуб повсеместно предпочитает южные освещенные склоны нижней и среднегорной полосы. Чаще всего первый ярус древостоя сложен почти одним дубом или к нему примешиваются в нижних частях склонов и на более глубоких почвах бук, граб, ильм, клен полевой, ясень и некоторые другие виды.
В лесах на известковой почве формируется своеобразный тип леса с прекрасно развитым ярусом подлеска из скумпии. На известковых же почвах, но несколько более глубоких и увлажненных, распространен дубовый лес с ярусом из кизила. Своеобразный тип леса образуют дубовые леса Кавказа на
267
южных склонах со сравнительно неглубокой бурой оподзоленной почвой, при сильно развитом подлеске из азалии. Эти леса довольно широко распространены. Здесь же, чаще по известковым почвам, встречаются дубовые леса с господством в травяном покрове злаков, образующих почти сплошной ковер. Зимний дуб с большой долей участия входит в леса с господством граба; единичной примесью входит в состав насаждений с господством каштана и бука.
Скальный дуб столь же хозяйственно-ценная древесная порода, как и черешчатый. Древесина его отличается плотностью, крепостью, прочностью и долголетием своей службы в постройках, высокой декоративностью. Ядровая часть древесины бурого цвета, заболонная часть узкая, более светлая. Древесина применяется во всех областях строительного дела, идет на мебель, на паркет, для производства строганного шпона, дает прекрасную клепку, и т. д. Дубовая древесина в большом количестве идет на дубильный экстракт, давая выход танинов от 3 до 5 %. При изготовлении винных и коньячных бочек дубу скальному отдается предпочтение перед дубом черешчатым. По анатомическому строению мало отличается от дуба черешчатого.
Дубовая кора содержит дубильные вещества, количество которых доходит до 16 % в коре молодняка (8-25 лет). Желуди дуба используют на корм скоту.
Из-за высоких физико-механических свойств, хорошей обрабатываемости режущими инструментами, красоты текстуры древесины, а вследствие этого и высокого спроса, дуб является объектом интенсивной заготовки. Запасы дуба скального, как и дуба черешчатого, быстро сокращаются. Данная древесная порода нуждается в бережной эксплуатации и искусственном восстановлении запасов.
Бук восточный - Fagus Orientalis Lipsky
Род Бук (семейство Fagaceae) объединяет хозяйственно важные древесные породы, произрастающие в умеренной зоне. В России этот род представлен преимущественно буком восточным.
Мощное дерево с полнодревесным стройным стволом, покрытым тонкой серой гладкой корой. Крона широкая, густая. При свободном стоянии деревья уже на незначительной высоте образуют разветвления. У молодых стволов кора зеленовато-серая, гладкая, блестящая, с продолговатыми узкими чечевичками. С возрастом она становится серой, едва достигая 1,5 см толщины, твердеет и делается ломкой. В лесных насаждениях из-за развития лишаев иногда приобретает характерную пятнистость.
Листья, расположены двухрядно. При основании их черешков имеются длинноланцетные, рано опадающие, прилистники ржавого цвета. Листья обратнояйцевидной формы. Иногда встречаются деревья с перисто-лопастными листьями. Края листьев в верхней части иногда волнистые или редкозубчатые. Размер листьев колеблется от 5 до 15 см, достигая особенно боль-
268
шой величины на поросли. Жилок 7-14 пар (рис. 79). У бука восточного, как и у дуба, есть рано и поздно распускающиеся формы.
Рис. 79. 1 - бук лесной: 1а - побег с веретеновидными почками; 16 - побег с распускающимися листьями, мужскими и женскими головчатыми соцветиями; 1в - мужской цветок; 1г - женский цветок; 1д - лист; 1е - раскрытая четырехраздельная плюска с орехами. 2 - бук восточный: 2а - побег с листьями и нераскрытой плюской; 26 - плод - трехгранный орех. 3. - каштан посевной: За - побег с листьями, мужскими и женскими соцве-
269
тиями; 36 - плюска с шипами; Зв - плод - орех (каштан) (из Зуихина, Коровин, Тимофе-енко, 2009)
Бук восточный цветет одновременно с распусканием листьев, в конце апреля - в мае. В октябре созревают плоды. Они коричневые, блестящие, трехгранные, длиной 1,2-2 см, заключены в деревянистую плюску. Из плюски не выдаются и заметны лишь при раскрывании ее. Опадают с октября по декабрь. 1000 плодов весят 140-220 г.
Бук восточный - горная порода и растет на больших площадях в среднегорной полосе Кавказа от 600 до 1000 м над уровнем моря. Выше входит в полосу распространения пихты и ели, нередко образуя верхнюю границу леса. На высоте около 2200 м он приобретает вид кустообразных зарослей. Крайним пунктом его распространения на север является г. Ставрополь. На Северном Кавказе особенно широкая полоса буковых лесов тянется южнее Майкопа, Нальчика и Владикавказа, вдоль хребта, вплоть до субальпийской зоны.
Бук восточный, являясь породой горного и приморского климата, довольно теплолюбив и требователен к влажности воздуха. Страдает от заморозков. Весьма теневынослив. Поэтому придерживается северных затененных склонов и глубоких долин, в густом буковом лесу всегда полумрак, подлеска нет. Благодаря теневыносливости бук хорошо возобновляется под пологом смешанного леса, образуя местами густой самосев.
К почве бук восточный требователен. Произрастает преимущественно на горно-лесных бурых свежих почвах. Совершенно не растет на избыточно увлажненных, не выносит значительной сухости почвы.
Кроме стержневого корня, бук восточный развивает много боковых, далеко уходящих в землю. Образуется сильная, но относительно поверхностная корневая система. Это придает буку в насаждениях и группах достаточную ветроустойчивость.
Бук восточный растет медленно. В первые годы он кустится, пока ветви не закроют стволик и почву от действия солнечных лучей. С 5 лет начинает прирастать более заметно, давая иногда по два прироста в год. До 30 лет его средний прирост колеблется от 15 до 30 см в зависимости от условий произрастания, от 30 до 60 лет возрастает, достигая в лучшие годы 80 см. С началом плодоношения приросты падают до 10-20 см в год. Старые деревья достигают огромных размеров - до 40 м в высоту и 2 м в толщину. Продолжительность жизни бука 300-400 лет, но обычно из-за гнилей редко доживает и до 150.
Плодоносить начинает в 40 лет, а в сомкнутых насаждениях не ранее 60-80 лет. Обильные урожаи повторяются в зависимости от погоды через 8-10 лет, а посредственные через 3-6 лет. В урожайный год можно собрать до 0,5-1 т плодов с гектара. Семена скоро теряют всхожесть.
Бук восточный размножается семенами. При появлении всходов плод растрескивается на три части и выпускает сначала корешок, уходящий в глубь почвы. Стебелек выносит на поверхность оболочку плода, позднее она
270
опадает, обнажая две (реже три) мясистые семядоли. Размножается также вегетативно, сохраняя сравнительно сильную порослевую способность до глубокой старости.
Бук - типичная лесная порода. Образует чистые или смешанные с пихтой кавказской и елью восточной леса. Имеет исключительно важное значение как лесообразующая, укрепляющая склоны и почвозащитная порода. Его листья богаты известью и при разложении препятствуют образованию грубого гумуса.
Древесина рассеяннососудистая белая, с красноватым оттенком, усиливающимся при хранении на воздухе, довольно прочная, твердая, легко колется и после распаривания хорошо гнется. Удельный вес меньше, чем у древесины дуба. Прорезана многочисленными, хорошо различимыми невооруженным глазом, широкими и узкими лучами, образующими при отделке и полировке красивый рисунок. Иногда имеет ложное ядро буровато-красного цвета. Применяется в машиностроении, при отделке жилых и служебных помещений, в мебельной промышленности, пригоден для изготовления гнутой мебели, для изготовления шпал, ружейных лож и мелких поделок. Путем сухой перегонки получают уксус и деготь, который перерабатывается в креозот. Орешки содержат 20-30 % масла, по качеству не уступающего оливковому.
Анатомические особенности древесины. Поперечный срез. Ядро неразличимо. Часто образуется ложное ядро. Древесина рассеяннососудистая. Просветы сосудов своим расположением не образуют какого-либо рисунка, в ранней и поздней древесине мало отличаются по размерам. Очертание просветов овальное или угловатое. Граница годичных слоев хорошо выражена. Основная масса древесины состоит из волокнистых трахеид и толстостенных угловатых волокон либриформа, встречаются сосудистые трахеиды. Осевая паренхима апотрахеальная: диффузная и метатрахеальная. Последняя преимущественно в поздней древесине в виде коротких (3-4 клетки) цепочек. Многорядные лучи при пересечении границы годичных приростов расширяются. Узкие лучи возле сосудов слегка изгибаются. Тангенциальный срез. Мелкие лучи однорядные, крупные - шириной до 25 клеток. Встречаются все переходы по рядности от однорядных до широких лучей. Преобладают 3-6-рядные. Лучи гомогенные, но краевые клетки более высокие, чем внутренние. Высота всех лучей весьма изменчива. Однорядные состоят из 2-3 до 8-10 клеток; широкие - из 12-15 до 40-120 клеток. Радиальный срез. Членики сосудов средней длины, часто с небольшими заостренными клювиками. Перфорации сосудов простые (преобладают) и лестничные, простые перфорации чаще в широких сосудах. Число перекладин в лестничных перфорациях не больше 6-8. Поровость сосудов зависит от контактирующих с ним элементов: межсосудистая - сближенные окаймленные поры, расположены косо или очередно; при контакте с трахеидами или волокнами либриформа - окаймленные поры редко и свободно рассеяны по стенке членика сосуда; при контакте с лучами - группы крупных пор без видимого окаймления. Волокна либриформа встречаются реже волокнистых трахеид, они значительно длиннее трахеид, их оболочки толще, поры простые щелевидные расположены редко. Тяжи осевой паренхимы состоят из 3-6 тонкостенных высоких клеток.
Буковые леса в горах и Кавказа производят сильное впечатление стройной колоссальной громадой серых стволов (до 1-2 м диаметра при высоте до 40 м), довольно редко стоящих друг от друга, но вполне сомкнутых вверху своими кронами, дающими сильное затенение. Мрачный колорит леса
271
усиливает серый цвет гладкой коры стволов и коричневый ковер мертвой листвы на почве. К буку в незначительном количестве примешиваются граб, горный ильм, клены, каштан и черешня. Подлесок почти отсутствует. Травянистый ярус чрезвычайно беден, лишь весной здесь в большом количестве появляются эфемеры. Возобновление этого типа леса под пологом бука вполне обеспечено. Почвы, занятые им, могут быть названы или бурыми лесными подзолами, или оподзоленными буроземами.
Мертвопокровные букняки следует считать стержневым типом леса: бук в них оказывается исключительно сильным эдификатором.
При уклонении условий произрастания в сторону большой сухости почв и их возрастающей щебнистости, а склонов - в сторону большого освещения строение букового леса меняется; падает производительность древостоя; в нем постепенно увеличивается доля участия дуба. Ближе к мертвопокровному типу леса стоит орляковый букняк; он занимает несколько более сухие и освещенные склоны. На высоте около 1700 м этот букняк сменяется своим высотнозамещающим букняком, субальпийско-разнотравным. Этот последний имеет производительность III—IV класса бонитета, снижаясь до V. Уклоняясь в сторону еще большей сухости произрастания, буковый лес приобретает злаковый травяной покров. Производительность древостоя падает; в нем уже значительна примесь дуба и граба, а на бескарбонатных почвах и каштана.
В субальпийской полосе он граничит с букняком рододендровым. В этом последнем, где почва крайне камениста и мелка, под кустистым буковым древостоем разрастается целое море кавказского рододендрона. Этот тип леса встречается только на высотах 2000-2300 м.
Каштан посевной - Castanea sativa Mill.
Семейство Fagaceae. Дерево первой величины, до 35м высоты, до 2м в диаметре. Крона широкая, раскидистая, кора темно-коричневая, трещиноватая. Побеги с крупными яйцевидными темно-коричневыми почками. Листья крупные продолговато-ланцетные до 25см длиной, с 15-25 парами боковых жилок, каждая из которых входит в вершину зубца, образуя загнутое кверху острие. Цветет каштан после облиствения в первой половине лета. Тычиночные цветки, желтого цвета, собраны в дихазии, которые располагаются на длинных, до 15см, прямостоячих осях. Каждый цветок имеет пяти-, шестираздельный колокольчатый околоцветник зеленого цвета и 10-12 тычинок с длинными тычиночными нитями и желтыми пыльниками, расположенными вокруг нектароносного диска. Женские головчатые соцветия располагаются у основания мужских стерильных соцветий. Таким образом, часть соцветий чисто мужские и образуют половозрелую пыльцу, а другие мужские соцветия, у основания которых расположены женские соцветия, половозрелой пыльцы не образуют. Пестичные цветки имеют зеленый околоцветник и длинные лентовидные рыльца. Каждые три цветка (реже 1-2) собраны в дихазий и окружены колючей четырехраздельной оберткой, из которой впо
272
следствии образуется плюска. Плод - орех (каштан) коричневого цвета, блестящий, съедобный. Плоды заключены в шаровидную, колючую плюску, чаще по два - три, а иногда в плюске содержится только один, очень крупный каштан (рис. 79).
Анатомические особенности древесины. Поперечный срез. Ядро хорошо выражено. На поперечном срезе напоминает древесину дуба. Древесина кольцесосудистая, ранняя и поздняя зоны годичного прироста четко различаются. В ранней древесине крупные округлые или овальные просветы сосудов располагаются одиночно или сближено (по 2-3), в 1-3 иногда в 4 ряда. Членики сосудов заполнены тилами. В поздней древесине узкопросветные сосуды образуют радиально вытянутые группы в виде дендритов и извилистых полосок. Очертание просветов узких сосудов овальное или угловатое. Годичные слои четко ограничены - вдоль внешней границы слоя проходит полоска из 4-5 рядов сплюснутых в радиальном направлении волокон. Переход от ранней древесины к поздней резкий, основная масса древесины состоит из тонкостенных трахеид и либриформа. В ранней древесине преимущественно трахеиды, в поздней - волокна либриформа. Осевая паренхима диффузная и скудно-вазицентрическая. Тангенциальный срез. Лучи однорядные, изредка встречаются частично двухрядные, низкие в 5-7 клеток в высоту и высокие высотой до 30 клеток. Большинство лучей состоит из 10-20 клеток. Радиальный срез. Членики сосудов тонкостенные с небольшими клювиками. Перфорации сосудов простые, в широких сосудах на конечных (поперечных) стенках, у узких на боковых или скошенных конечных. Межсосудистая поровость очередная, у узких сосудов может быть - супротивная. Поры немногочисленны, расположены свободно. Окаймление округлое или овальное. Трахеиды, расположенные возле сосудов тонкостенные с заостренными или тупыми зазубренными окончаниями. Окаймленные поры на стенках трахеид расположены двухрядно. Волокна либриформа длиннее трахеид, толщина их стенок в 1,5-2 раза превышает толщину стенок трахеид, поры мелкие, плохо различимы при увеличении бООх, расположены в основном в поздней древесине.
Древесина кольцесосудистая, ядровая, с красивой текстурой, очень высоко ценится в мебельном и бондарном производствах.
Ареал: юг Западной Европы, Средиземноморье, Кавказ, Малая Азия, культивируется в Крыму и Молдавии. На Кавказе растет на высоте 300 -800м над уровнем моря. Каштан теплолюбив, светолюбив, требует плодородной и достаточно увлажненной почвы, а также повышенной влажности воздуха, любит кислые и нейтральные почвы речных долин, избегает известняков. Разводится как орехоплодное растение.
Клен ложноплатановый (клен белый, явор) - Acerpseudoplatanus
Дерево до 40 м высотой и до 2 м в диаметре. Крона раскидистая, шатровидная. Кора серовато-бежевая, отслаивается крупными пластинами, обнажая пятнами более яркую, молодую кору. Побеги голые, желто-зеленые с чечевичками, почки яйцевидные, до 15 мм длиной. Листья 5-лопастные, до 17 см в окружности, сверху тускло-зеленые, снизу беловатые, иногда с пурпурным оттенком. Лопасти яйцевидные, по краю зубчатые, угол между лопастями очень острый. Черешок листа равен или больше длины листовой пластины. Соцветие - вытянутая, кистевидная метелка, до 16 см длиной. Цветки до 8 мм в диаметре, зеленоватые, с почти равными по длине чашелистиками
273
и лепестками. Цветки также, как и у клена остролистного, могут быть мужские, женские и ложно обоеполые. Цветет в мае - июне. Плоды - дробные крылатки (двукрылатки), расходящиеся под углом от 45° до 90°, длина каждого крыла до 6 см. Семена выпуклые, шаровидные (см. рис. 72). Плоды созревают в сентябре - октябре. Лучше всего всходят при осеннем посеве свежесобранными семенами.
Ареал: юго-западная часть Украины, Кавказ, средняя и юго-восточная часть Западной Европы, черноморское побережье Малой Азии. Растет в нижнем и среднем поясе гор, до 1800 м над уровнем моря, обычно на склонах северных и западных экспозиций. Чистых насаждений не образует, а растет вместе с пихтой, буком, липой, грабом на глубоких, плодородных почвах с достаточным увлажнением. Участие этой породы в составе древостоя обычно не превышает 2-3 единиц. Явор теплолюбив, влаголюбив, особенно требователен к влажности воздуха, в молодом возрасте теневынослив, но нужно помнить, что это растение первого яруса и во втором ярусе обычно не растет. При сильном затенении подрост медленно отмирает, видимо этим объясняется редкое участие явора в составе насаждений. Растет быстро, в 60 лет может достигать 30 м высоты и 60 см в диаметре, доживает до 200-250 лет. В культуре применяется с давних пор, но успешно растет только в европейской части нашей страны, до зоны широколиственных лесов включительно, севернее обычно подмерзает.
Хозяйственное значение клена ложноплатанового велико. Он дает самую ценную древесину для изготовления музыкальных инструментов (нижние деки и обичайки струнных инструментов), используется в мебельном производстве, для столярных, резных и токарных изделий. Довольно часто образует древесину с декоративной текстурой ("птичий глаз", волнистую, струйчатую), которая используется для отделки помещений и мебели. По анатомическим признакам рассматриваемые здесь виды клена практически не различаются. Клен ложноплатановый используется как прекрасное парковое дерево.
Клен полевой - 4. cam pestre L.
Дерево 15-20 м высотой до 60 см в диаметре, крона шаровидная, кора серая, с продольными трещинами. Побеги тонкие, желтовато-бурые с продольными полосами, почки яйцевидные, до 5мм длиной. Листья цельнокрай-ные, 5-лопастные, от 4 до 14 см в диаметре, вершинки лопастей притупленные. Соцветие - конечная, прямостоячая, щитковидная метелка, до 3,5см в диаметре, цветки желто-зеленые, на одном растении бывают: пестичные, тычиночные и ложно обоеполые. Цветет через декаду после распускания листьев. Плод - дробная крылатка, длиной от 2 до 3,5см, крылья расходятся под углом 180°; семена плоские (см. рис. 72).
Ареал: от Англии до Волги по зоне широколиственных европейских лесов, лесостепь, степь, включая Крым, Кавказ, север Африки и северную часть Малой Азии. Растет вместе с дубом черешчатым, дубом скальным и
274
дубом пушистым; грабом, грабинником, древовидными можжевельниками. На Кавказе поднимается в горы до 1800 м над уровнем моря. В культуре встречается в Петербурге, Вологде, Москве, в Казахстане. Умеренно теплолюбив, засухоустойчив, требователен к плодородию почвы, растет медленно, доживает до 100-150 лет, не выносит застойного увлажнения, устойчив в условиях города. Древесина используется для столярных изделий, мелких поделок, в озеленении может применяться в аллейной и групповой посадке.
Орех грецкий - Juglans regia L.
Род орех объединяет около 20 видов листопадных древесных растений, большинство из них формируют первый ярус сложных, смешанных древостоев. Представители рода орех являются ветроопыляемыми растениями. Женские цветки располагаются на вершине молодого побега одиночно или по несколько штук вместе. Цветок находится в пазухе кроющего листа, имеет один пестик с двураздельным, крупным, бахромчатым рыльцем. Завязь нижняя, одногнездная. Тычиночные цветки собраны в сережки, появляются весной на побегах прошлого года. Цветок сидит в пазухе кроющего листа, сросшегося с двумя прицветниками. Он имеет от 8 до 40 тычинок на коротких тычиночных нитях. Плод - ложная костянка (ложный орех) зеленого цвета с кожистой внешней оболочкой, крупной косточкой и мясистыми съедобными семядолями (рис. 80).
Древесина ядровая, плотная, прочная, разнообразно и красиво окрашенная. Ядро серовато-коричневое, иногда красноватое с неравномерными темными, иногда почти черными, прожилками. Заболонь широкая сероватобурая. В нашей стране одна из самых ценных древесин для столярномебельного производства, употребляется для изготовления дорогой мебели, ружейных лож, токарных изделий. Строганный шпон грецкого ореха идет на отделку стен театров, кают пароходов.
Анатомические особенности древесины. Поперечный срез. Крупные сосуды диаметром около 250 мкм равномерно распределены по всей ширине годичного слоя, некоторые затилованы. Просветы сосудов чаще одиночные, но встречаются и радиально вытянутые группы по 2-4, изредка до 6 просветов. В ранней древесине просветы несколько шире, чем в поздней. Стенки сосудов слабо утолщенные, овальные или круглые, в группах - угловатые. Граница годичных слоев слегка волнистая. Основная масса древесины представлена тонкостенными и шорокопросветными волокнистыми трахеидами и волокнами либриформа. Осевая паренхима обильная, большей частью апотрахеальная - метатрахеаль-ная, в меньшей доле - паратрахеальная. Клетки паренхимы среди трахеид и либриформа хорошо выделяются.
275
Рис. 80. 1 - орех грецкий: 1а - побег со сложным листом и цельнокрайными листочками; 1 б - побег с мужскими и женскими соцветиями; 1 в - орех без кожистой оболочки; 1г - поперечный разрез ореха. 2 - орех маньчжурский: 2а - общий вид листа (уменьшено); 26 - фрагмент сложного листа; 2в - женский цветок; 2г - плоды (ложные костянки или ложные орехи); 2д - орех без кожистой оболочки с хорошо выраженными настоящими и ложными ребрами (из Зуихина, Коровин, Тимофеенко, 2009)
276
Тангенциальный срез. Лучи веретеновидные 1-4-рядные. Некоторые многорядные лучи имеют однорядное окончание. Клетки лучей овальные и округлые, четковидные, различной высоты. Радиальный срез. Членики широких сосудов короткие бочковидные, без клювиков или с небольшими клювиками, узких - удлиненные, без клювиков. Перфорации простые на боковых или скошенных конечных стенках. Поровость сосудов очередная и супротивная, свободная иногда сомкнутая. Свободно расположенные поры крупные овальные, с вытянутыми отверстиями, не достигающими границы окаймления. Сомкнутые поры очередные, шестиугольные. Основная масса древесины представлена либри-формом и волокнистыми трахеидами. У либриформа утолщенные оболочки и редкие щелевидные поры, трахеиды широкопросветные, тонкостенные, с порами, имеющими неясное окаймление, тяжи осевой паренхимы состоят из 8- 15 узких удлиненных клеток. Лучи гетерогенные, клетки лежачие, но разной высоты.
В нашей стране естественно растут три вида: орех грецкий, орех маньчжурский и орех Зибольда, четыре вида интродуцированы.
Орех грецкий - дерево, высотой до 35 м и до 2 м в диаметре. Стволы стройные, крона в насаждениях высоко поднята. Листья непарноперистые до 40см длиной состоят из (3) 5-9 (11) листочков. Листочки эллиптические или яйцевидные, цельнокрайние, на порослевых побегах с редкими зубцами. Верхушечный листочек крупнее остальных. Описание цветения приведено в общей характеристике рода орех. Плод - ложная костянка (ложный орех), при созревании кожистая оболочка плода растрескивается на четыре части, и плод (грецкий орех в бытовом понимании) выпадает. Форма плодов разнообразна. Скорлупа светлая относительно тонкая. Семядоли крупные, морщинистые, содержат до 50 % масла.
Родина: естественно растет в горах Средней и Малой Азии, в Закавказье. На Кавказе, в Греции и Италии является древним интродуцентом. В настоящее время растет на Северном Кавказе, в Крыму на юге Украины, в Молдавии, где часто выращивается на плантациях. Орех светолюбив, предпочитает плодородные известковые умеренно увлажненные почвы.
Орех грецкий - ценное орехоплодное растение. Кроме семян используется внешняя оболочка плода, из которой получают очень стойкие черную и коричневую краски.
Тополь белый (серебристый, белолистка) - Populus alba L.
Могучие деревья до 40м высоты и до 2м в диаметре со светлой, гладкой, у старых деревьев в нижней части ствола серой или черной трещиноватой, корой. Крона широкая раскидистая. Побеги, почки и нижняя сторона листьев с густым белым, войлочным опушением. Листья на удлиненных и порослевых побегах - пальчато-лопастные, на укороченных - округлые или овальные, с выемчато-зубчатым краем (рис. 81).
277
Рис. 81.1. Тополь черный: 1а - побег во время цветения с мужскими сережками; 16 - мужской цветок; 1в - побег с женскими сережками; 1г - женский цветок; 1д - побег с листьями и соплодием; 1е - плоды - коробочки; 1ж - семя с пучком волосков. 2. Тополь белый, удлиненный побег с пальчато-лопастными листьями (нижняя сторона листьев белая из-за войлочного опушения. 3. Чозения толокнянколистная: За - побег с листьями; 36 - женские цветок; Зв - мужской цветок; Зг - соплодие; Зд - плод - коробочка
278
Ареал: тополь белый образует пойменные леса в центральных и южных районах Европейской части нашей страны, а также на юге Западной Сибири, в Крыму, на Кавказе и в Средней Азии. Доживает до 300 лет. Широко культивируется в городских посадках, приблизительно до широты Москвы. Зимостоек, светолюбив, требователен к влажности почвы, выдерживает временное затопление. Также, как и осина образует массу корневых отпрысков, которыми обычно и разводится.
Древесина с мягкая с беловатой или слегка коричневато-белой заболонью и желтым или темным (до коричневого) ядром. Годичные кольца широкие, ровные в очертании. По анатомическим признакам мало отличается от древесины осины. Используется в качестве поделочного материала, иногда в мебельной промышленности, для производства целлюлозы.
Тополь черный (осокорь) - Populus, nigra L.
Дерево до 45м высоты и до Зм в диаметре. При достаточном освещении крона низко опущенная, густая, широкая. Часто на черном тополе образуются капы, почти сплошь покрывающие нижнюю часть ствола. Кора очень толстая (до 10 см), темная с крупными продольными трещинами. Побеги голые, цилиндрические, желтовато-серые. Почки острые, клейкие, с отогнутой вершинкой. Листья треугольные с оттянутой вершинкой. Цветет до распускания листьев, мужские сережки пурпурно-красные, благодаря окраске пыльников. Семена созревают в первой половине лета.
Ареал: распространен по поймам рек по всей Средней и Южной Европе, на Северном Кавказе, в Западной Сибири, на Алтае и в Средней Азии. Растет быстро, живет до 300-400 лет, свето- и влаголюбив, выносит паводковое затопление, к почвам нетребователен. Северные популяции зимостойки, южные теплолюбивы. Естественно размножается, чаще всего, семенами, при искусственном разведении - стеблевыми черенками.
В древесине ядро не выражено. Древесина светло-серая почти белая, легкая и мягкая. Легко обрабатывается. По анатомическим признакам мало отличается от осины. Применяется для построек, для производства целлюлозы, для изготовления мебели. Из стволов с капами получают высоко декоративный лущеный шпон, который используют для внутренней отделки помещений и изготовления дорогой мебели.
ЛЕСА ДАЛЬНЕГО ВОСТОКА
Дальний Восток в России занимает огромную территорию на крайнем востоке Азии. Он имеет значительное протяжение в широтном направлении от 42 до 70° с. ш. В него входят два больших полуострова - Камчатский и Чукотский, остров Сахалин и Курильские острова. Этот регион выделяется в особую физико-географическую Дальневосточную страну.
279
Граница между Дальним Востоком и Восточной Сибирью проходит по горным хребтам, идущим от Забайкалья до Ледовитого океана (Яблоновый, Становой, Джугджур, Колымский, Анадырский). Это преимущественно гористая страна. Помимо названных гор, сюда входят: Буреинские горы, Сихотэ-Алинь, Камчатка и Сахалин (также горные страны).
По территории Дальнего Востока протекают такие крупные реки, как Амур с притоками Зея, Бурея и Уссури, на севере - Анадырь и много небольших рек.
На климат этой страны оказывают существенное влияние с востока Тихий океан, а с запада - азиатский континент. При наличии горного рельефа и значительного протяжения в широтном направлении формируется своеобразный холодный и умеренный приморский климат: зимой - сухие холодные северо-западные ветры, летом юго-восточные влажные, тоже прохладные. Зима бывает холодная, ясная, местами малоснежная; весна затяжная, сухая; лето дождливое, особенно во второй половине, в южной части оно жаркое; осень сухая, ясная. Сумма годовых осадков в южной части колеблется от 600 до 800 мм, в северной - от 200 до 300 мм. Самая южная часть Дальневосточного края находится на широте Сухуми, и летом там достаточно влаги и тепла, однако климат не субтропический, а умеренно теплый.
Почвенный покров в северной части формируется на вечной мерзлоте и состоит из маломощных тундровых почв, которые южнее сменяются торфянистыми и слабооподзоленными суглинками на каменистых породах, а на крутых склонах переходят в каменистые гольцы. В южной части почвы подзолистые и подзолисто-глеевые, дерново-подзолистые, бурые, лесные и торфяно-глеевые.
На территории Дальнего Востока выделяются только три природные зоны: тундровая, лесотундровая и лесная. В лесной зоне встречаются несколько типов растительности: лес, растительность гольцов (мхи, лишайники, одиночные кустарники, заросли стланикового кедра), кустарниковые заросли, сфагново-моховые болота с лиственницей и луга.
Лесную зону можно разделить на четыре подзоны:
1. Северная подзона - от северной оконечности Камчатки до Аяна. В образовании лесов принимают участие лиственница даурская, береза каменная, тополь темнолистный, чозения, стланиковая сосна.
2. Средняя подзона хвойных лесов Охотского типа - от Аяна до Амура. Леса состоят из лиственницы даурской, ели аянской, пихты белокорой, березы каменной.
3. Южная подзона хвойных лесов с участием лиственных пород - от Амгуни до северного Сихотэ-Алиня, нижнее течение Амура, Северный Сахалин; леса образуют те же хвойные и, кроме того, кедр корейский и сосна обыкновенная, из лиственных - дальневосточные березы, осина и некоторые широколиственные четвертой подзоны.
4. Подзона смешанных хвойно-широколиственных лесов - Средний Амур, Уссури, Сихотэ-Алинь, Южный Сахалин характеризуется большим разнообразием видов древесных растений. Эта часть Дальнего Востока не
280
была покрыта ледником и здесь сохранились виды растений третичного периода, а также виды древесных растений, замещающие виды лесов Западной Европы.
Существуют и другие подходы к районированию территории Дальнего Востока (рис. 82).
Рис. 82. Физико-географическое районирование Дальнего Востока: А - Амуро-Сахалинская страна. Физико-географические области: I - Верхнезейская, II - Зейско-Буреинская, III - Буреинская, IV - Нижнеамурская, V - Приморье, VI - Сахалин. Б - Се-веро-Притихоокеанская страна. Физико-географические области: VII - Чукотская, VIII -Анадырско-Пенжинская, IX - Корякская, X - Охотская, XI - Камчатская, XII - Курильские острова (из Сочава, 1962)
281
В целом леса Дальнего Востока занимают площадь в 273.7 миллиона гектаров и обладают, по оценке Дальневосточного лесоустроительного предприятия, запасом древесины в 20.4 миллиарда кубометров (данные 1995 года). Потребители обычно обосновывают свои запросы этими цифрами, которые действительно производят впечатление. Общая расчетная лесосека для Дальнего Востока громадна - 95 миллионов кубометров, в том числе 79 миллионов хвойных пород. И лишь 12,8 процента из них принадлежат лесам 1 группы, 87 % - это леса коммерческого значения. Однако, реально доступными для использования при существующей инфраструктуре являются только 10 % из всех лесов 3 группы (25-30 % на юге региона). Несмотря на внушительность, размер остальных неосвоенных лесов должен быть серьезно скорректирован с учетом их глобальной значимости и крайней ранимости -параметров, опущенных в действующих методиках лесоустройства. Существенным фактором при оценке реального запаса древесины здесь следует считать и ее сравнительно низкое коммерческое качество, не позволяющее поддерживать экономически устойчивые лесозаготовительные предприятия из-за низкой цены. В связи с уменьшением доли спелых и перестойных лесов в регионе за последние десятилетия с 75-90 % до 45-50 %, применяющиеся способы оценки расчетной лесосеки неизбежно ведут к истощительному лесопользованию. Ученые Дальнего Востока неоднократно предлагали изменить методику расчета главного пользования с учетом возрастной структуры лесов.
Относительно неосвоенных лесов существуют разные количественные оценки: они варьируют в довольно широком пределе от25%до50%в зависимости от выбранных критериев. Массивы неосвоенных лесов имеют исключительное значение в сравнении с другими лесами умеренного пояса планеты.
В то время как 40 % лесов мира остается недоступными из-за горного рельефа и неразвитой дорожной сети, лесозаготовительные компании наращивают масштабы и объемы рубок в доступных районах, преимущественно вблизи дорог и населенных пунктов, не очень заботясь об улучшении методов и технологий. Проблема переруба в таких районах усугубляется неравномерным распределением лесов. Хотя специалисты-лесоводы часто утверждают, что количество лесных площадей в России растет, прямо или косвенно обосновывая увеличение лимитов лесосечного фонда, это утверждение нуждается в уточнении. Рост лесопокрытых площадей далеко не всегда означает увеличение запасов древесины, и чаще всего свидетельствует о снижении производительности и экологической пластичности древостоев. Из-за масштабных рубок и пожаров вторичные лиственные леса вытесняют спелые хвойные на большинстве лесопокрытых площадей. Большая часть территорий, подвергнутых сплошным рубкам и последующему выгоранию в ряде районов Хабаровского края, в особенности в границах зоны вечной мерзлоты, остаются безлесными из-за сильных морозов и разрушенного почвенного
слоя.
282
Сохранение и рациональное использование лесов Дальнего Востока и Восточной Сибири является проблемой глобального значения по ряду причин: сохранение биоразнообразия, включая исчезающие виды, такие как амурский тигр; сохранение мест традиционного обитания и хозяйственной деятельности для десятков тысяч представителей коренных народов региона, полностью зависимых от ресурсов леса; сохранение стабильного источника древесины для соседних стран, значительно или полностью истощивших собственные лесные ресурсы; смягчение глобальных климатических изменений и сохранение углеродного баланса в атмосфере; сохранение и развитие сбалансированной местной экономики в регионе для обеспечения социальной стабильности.
Леса горной системы Сихотэ-Алиня и Приамурья признаны Международным Союзом Охраны Природы (МСОП) как один из центров растительного многообразия, содержащий более 3000 видов сосудистых растений и множество их экзотических сочетаний. Это побудило Комиссию по Всемирному Наследию ЮНЕСКО включить ряд охраняемых территорий центрального Сихотэ-Алиня в список претендентов на получение этого высокого международного статуса. Часто этот регион рассматривают как нижнюю часть Амурского бассейна, включающую остров Сахалин. В свое время территория избежала покровного оледенения и стала своеобразным климатическим резерватом для множества видов и сообществ при очень высоком уровне эндемизма растений и беспозвоночных. Аналогичные леса когда-то покрывали Китай, Корею и Японию, но сегодня они сильно деградировали из-за чрезмерного использования.
Уникальная биогеографическая история региона привела к сочетанию в единой экосистеме таких неожиданных видов как тигр, леопард, кабарга, гималайский и бурый медведь, олени и лососи. К сожалению, большинство коммерчески значимых лесов Дальнего Востока сосредоточено в этом же регионе. Хотя здесь есть и другие территории, важные с точки зрения биоразнообразия. Так называемый "Хвойный остров" в центре Камчатки хранит водотоки богатейших в мире лососевых рек, которые в свою очередь кормят десятитысячную популяцию камчатских бурых медведей.
Не менее важную роль обширные массивы сибирских и дальневосточных лесов играют как регулятор климата планеты. Тайга - огромный резервуар углекислого газа, который ученые считают главной причиной глобального потепления. Ученые многих стран оценивают суммарный запас углерода в российских лесах в размере одной седьмой всего мирового запаса, или 75 % того, что хранят бореальные леса. Таким образом, будучи сохранены, российские леса могут сыграть решающую роль на планете в качестве ее зеленых "легких".
В то же время, интенсивное сведение российских лесов неизбежно и существенно ускорит выделение большого количества углерода в атмосферу и глобальное потепление климата. Этот процесс во многом зависит от характера ведения лесозаготовок. Если применяется технология выборочных рубок с сохранением разнообразной структуры леса, выброс СО2 оказывается
283
минимальным. Если ведутся сплошные рубки на конкретном участке, обезлесение не только ведет к высвобождению больших количеств углерода, но и резко снижает способность леса к удержанию углекислоты из-за нарушения структуры леса. На Дальнем Востоке более 90 % лесов подвергается сплошным рубкам. Кроме того, вопреки распространенному мнению, лесовосстановление далеко не всегда возвращает углерод земле. Специальные исследования на лесных плантациях показали, что они способны удерживать от одной трети до половины углерода по сравнению с тем, что мог бы удержать естественный лес на той же площади. Опасность истощительных методов лесопользования, и это доказано наукой, как раз и состоит в снижении многочисленных и взаимосвязанных защитно-экологических функций лесов - поч-во- и водозащитных, климаторегулирующих, имеющих местное, региональное и глобальное значение. Однако, для оценки этих воздействий, к сожалению, пока нет строгой научной базы, и аналитикам приходится зачастую прибегать к субъективным и опосредствованным оценкам.
Примерно 75 % преимущественно горных дальневосточных лесов расположено в зоне вечной мерзлоты, которая сохраняет почвенную влагу, обеспечивающую жизнь леса. В сухие летние месяцы мерзлота подтаивает, поставляя влагу и минеральные соли поверхностным слоям почвы и покрывающей их растительности, в том числе и лесной. Зимой все вновь замерзает, влага сохраняется, обеспечивая этим круговоротом здоровье северных лесов.
При вырубке таких лесов происходит также интенсивное выделение метана, воздействие которого на процессы глобального потепления в 10-20 раз существеннее, чем воздействие углекислоты. Крупномасштабные рубки приводят к искусственному таянию мерзлоты, открывая большие ее пространства прямым солнечным лучам и превращая ранее лесные территории в болота. В естественных условиях отмирающие растения не разрушают мерзлоту, они накапливаются на почве. Но когда мерзлота превращается в болото, этот органический материал разлагается быстрее, и свободный метан выделяется из растений и из самой мерзлоты. Постепенно эти почвы высыхают и могут превратиться в пустыню. Большинство таких зон, подвергнутых сплошной рубке, никогда не восстанавливается. Ученые также предупреждают, что значительное потепление климата Земли может в свою очередь привести к дальнейшему интенсивному таянию больших массивов вечной мерзлоты. Новые порции освобожденного метана еще больше ускорят процесс потепления, и весь цикл далее пойдет с неконтролируемым нарастанием.
В целом сохранение крупных массивов Российских лесов очевидно способствует сдерживанию происходящих климатических перемен путем сохранения углекислоты в земле за счет предотвращения вырубки лесов на весьма чувствительных к таким воздействиям мерзлотных почвах. Кроме того, сохранение естественных лесных ландшафтов обеспечивает необходимые миграционные коридоры для животных, которые вынуждены менять места обитания для адаптации к происходящим антропогенным и климатическим
изменениям.
284
В России всегда существовала определенная стратегия по сохранению особо ценных лесов. Наиболее традиционной формой было и остается создание особо охраняемых природных территорий (ООПТ). Но эта деятельность всегда противостояла индустриальной идеологии и нуждам лесной промышленности. Естественно, в этом неравном противостоянии побеждает тот, кто дает больше доходов в бюджет, то есть лесная промышленность, хотя каждый разумный человек в отдельности и согласен теоретически с природоохранительными доводами. Нынешняя Российская экономика имеет мало возможностей поддерживать даже существующие ООПТ, не говоря уже о создании новых. С другой стороны, открывшиеся зарубежные рынки спроса оказывают быстро возрастающее давление на внешне бескрайние леса Сибири, теряя все больше древесины, поступавшей ранее из тропических лесов -они тоже когда-то казались бескрайними.
В нашем учебнике, не останавливаясь на подробном рассмотрении северной части этого обширного региона, мы имеем возможность остановиться лишь на общей характеристике юга Дальневосточной страны - Амуро-Уссурийского края, в пределах которого находятся Приморский край, основная часть Хабаровского края и Амурская область. Этот регион занимает юго-восточную окраину континента, где особенно ярко проявляется взаимодействие огромной суши Евразии и вод Тихого океана. Географическое положение обусловило формирование главных особенностей природы этой части страны.
В рельефе этого региона преобладают средневысотные горы, чередующиеся с межгорными равнинами. Отмечается широкое распространение мелкосопочного рельефа. Климат носит ярко выраженный муссонный57 характер. В почвенном покрове характерно почти повсеместное устойчивое оглеение и слабое проявление подзолообразовательного процесса.
В ландшафтном отношении наиболее характерны хвойношироколиственные леса, образующие здесь вторую в России после сибирского перерыва область смешанных лесов. Наиболее специфичная черта его природы - оригинальный, нигде более не встречающийся в таком виде органический мир. Выдающийся русский географ-путешественник Н.М. Пржевальский так писал об этом крае: "Как-то странно непривычному взору видеть такое смешение форм севера и юга, которые сталкиваются здесь как в растительном, так и в животном мире. В особенности поражает вид ели, обвитой виноградом, или пробковое дерево и грецкий орех, растущие рядом с кедром и пихтою. Охотничья собака отыскивает вам медведя и соболя, но тут же рядом можно встретить тигра, не уступающего по величине и силе обитателю джунглей Бенгалии".
Амуро-Уссурийский край почти полностью входит в зоны тайги и смешанных лесов. Почвенно-растительный покров и животный мир дальневосточных лесов сильно отличаются от монотонной тайги Восточной Сиби
37 Муссоны - устойчивые сезонные ветры, направление которых резко меняется на противоположное или близкое к противоположному, 2 раза в год. Основная особенность муссонного климата - обильное осадками лето и сухая зима.
285
ри. На юге Дальнего Востока после громадного сибирского перерыва появляются смешанные хвойно-широколиственные леса с бурыми лесными почвами и оригинальным животным миром. Большие пространства в отличие от Восточной Сибири покрывают темнохвойные ценозы на буротаежных оподзоленных и неоподзоленных почвах. Наряду с ними в северо-западную часть края по многолетнемерзлым грунтам распространяются из Сибири лиственничные леса. Общая лесистость территории достигает 55 %. По долинам межгорных равнин значительные площади заняты лугами. Есть острова лесостепей - прерий.
Флора и фауна58 Амуро-Уссурийского края чрезвычайно богаты. Для них характерно наличие большого числа реликтовых видов, исключительное разнообразие с весьма типичным для юга Дальнего Востока смешением представителей флористических и фаунистических комплексов различных типов, с взаимопроникновением северных и южных форм. В растительном покрове и животном мире не только очень много отдельных реликтовых видов, но и имеются целые формации и зоокомплексы, мало отличающиеся от своих исходных родоначальных сообществ конца неогена - начала четвертичного времени (см. приложение 1). Более того, здесь встречаются растения, предки которых произрастали еще в конце мезозоя: папоротник осмун-да (Osmunda cinnamomea). оноклея (Ondclea sensibilis) и др. Обилие реликтов59 придает растительности и фауне своеобразный архаический колорит. Реликты не чувствуют себя угнетенно и широко распространены, играют важную роль в образовании современных биоценозов. Крупное древесное растение - чозения, предки которой были связующим звеном между родами ив и тополей, например, образует пойменные леса от самого юга до крайнего севера Дальнего Востока.
В распределении почвенно-растительного покрова и зооценозов наблюдается сложное сочетание широтной зональности, существенных долготных различий и хорошо выраженной высотной поясности. В северной половине юга страны господствуют таежные ландшафты, в южной - смешанные леса. Вследствие чередования горных хребтов и межгорных понижений субмеридионального направления характерно взаимопроникновение северных и южных типов леса. Тайга по хребтам продвигается до самого юга, смешанные леса доходят по равнинам и обращенным к ним склонам гор примерно до 50° с. ш.
Смешанные хвойно-широколиственные леса с бурыми лесными почвами на крайнем юге занимают склоны гор до высоты 800-900 м, к северу верхний предел их распространения снижается до 200 м и под 50-51° с. ш. в континентальных районах и 46° с. ш. на прохладном побережье эти леса постепенно исчезают.
38 Флора - исторически сложившаяся совокупность видов растений, населяющих какую-либо территорию, или населявших ее в прошлом; фауна - тоже в отношении к животным.
39 Реликты - виды растений и животных, входящие в состав растительного покрова и животного мира данной страны или области как пережитки флор и фаун минувших эпох.
286
Основу фитоценозов в смешанных лесах составляет флора так называемого маньчжурского типа. Наиболее характерны широколиственные породы: дуб монгольский, несколько видов клена, липы амурская, маньчжурская и Таке, ильмы белокорый (долинный) и горный, ясень маньчжурский, березы черная (даурская), желтая (ребристая) и железная (Шмидта), орех маньчжурский, бархат амурский, диморфант и др. Из хвойных типична сосна кедровая корейская, а на юге Приморья также пихта цельнолистная.
Строение смешанных маньчжурских лесов очень сложное. Древостой многоярусный и многовидовой. Первый ярус образует сосна кедровая корейская, чаще называемая кедром корейским (высотой до 40 м и диаметром 1,5 м), а на крайнем юге - и пихта цельнолистная (высота до 50 м, диаметр более 2 м). Из лиственных в первый ярус чаще всего выходят липа, желтая береза, монгольский дуб, в долинных лесах - ильм белокорый и ясень маньчжурский. Другие ярусы составляют прочие широколиственные. Общая сомкнутость крон в малоизмененных лесах 0,8-0,9.
Подлесок высокий, также многоярусный и разнообразный. Типичны аралия маньчжурская с колючим стволом и оригинальной декоративной листвой, ее родственник из семейства аралиевых - элеутерококк колючий, различные жимолости, бересклет, дикий жасмин-чубушник, амурская сирень, маньчжурская и разнолистная лещины, смородины. На месте гарей и рубок обычно разрастается густой подлесок из колючих аралиевых, сквозь который невозможно пройти без топора.
Специфичная особенность смешанных лесов юга Дальнего Востока -обилие крупных лиан: дикий амурский виноград с черными кисловатыми плодами, актинидии коломикта, острая (аргута) и полигамная, лимонник китайский, принадлежащий к семейству магнолиевых. Дальше всех к северу продвигаются виноград и актинидия коломикта. Наиболее крупная лиана -актинидия аргута, имеющая стебель диаметром до 15 см. Очень сладкие богатые витаминами плоды актинидий употребляются в пищу. Горьковатые похожие на бруснику плоды лимонника и его кора содержат тонизирующие лекарственные вещества, используемые в медицине. На стволах деревьев много эпифитных мхов, а на юге также и эпифитных папоротников. В наземном покрове преобладают папоротники и высокое разнотравье, на юге изредка встречается внешне невзрачный знаменитый «корень жизни» -женьшень, тоже - представитель аралиевых.
В лесах хвойно-широколиственной подзоны насчитывается свыше 200 видов деревьев и кустарников. В южном Приморье нередко встречаются лесные группировки, в составе которых на площади в 1 га имеется до 25 различных видов деревьев, до 30 кустарников, 5-6 видов лиан, до 15 видов папоротников, до 100 видов травянистых цветковых растений. Облик лесов юга Приморья не свойствен умеренному поясу. Они напоминают своеобразные джунгли в значительно большей мере, нежели субтропические леса Закавказья.
Видовой состав и строение смешанных лесов изменяются в разных районах страны. Самые богатые и наиболее сложные грабовые чернопихто-
287
во-кедрово-широколиственные леса с желтоземно-бурыми почвами южного Приморья. В первом ярусе их древостоя характерна, наряду с кедром пихта цельнолистная, во втором ярусе - граб сердцелистный. В подлеске преобладают аралиевые, много лиан, встречается женьшень. Желтоземно-бурые горно-лесные почвы отличаются сравнительно высокой гумусированно-стью, проникновением гумусовых веществ на большую глубину.
На юге от 500 до 700 м в горах и севернее предыдущей полосы, по предгорьям, распространены кедрово-широколиственные леса с бурыми лесными оподзоленными почвами. Они имеют типичные для дальневосточных смешанных лесных ценозов состав и строение. Оподзоленные бурые почвы содержат много гумуса, ненасыщенность основаниями небольшая.
На высоте от 700 до 900 м в горах крайнего юга и выше предгорных кедрово-широколиственных лесов в предыдущей полосе, а также севернее ее до границ хвойно-широколиственной зоны преобладают обедненные варианты смешанных лесов со значительным участием в древостое ели аян-ской пихты белокорой - елово-кедрово-широколиственные и еловошироколиственные леса с бурыми лесными почвами. На вырубках много белоберезово-осиновых лесов, луга распаханы. На Сахалине только пихто-во-елово-широколиственные леса, встречающиеся на крайнем юге острова; кедра в них нет. В северных смешанных лесах отсутствует целый ряд теплолюбивых растений - граб, орех и т. д., менее разнообразный подлесок и папоротниково-травяной покров, мало лиан. Бурые почвы почти не оподзо-лены, их профиль слабо дифференцирован.
В малоизмененном виде смешанные хвойно-широколиственные леса сохранились лишь в заповедниках. На всей остальной территории в результате неоднократных пожаров, а также выборочных и сплошных рубок растительность лесов нередко сильно изменена. Большие площади заняли производные мелколиственные и широколиственные леса. Самые распространенные среди последних дубняки с подлеском из разнолистной лещины или кустарника леспедецы, которая цветет почти все лето, создавая фиолетоворозовый аспект кустарникового яруса. В древостое дубовых лесов из других пород чаще всего встречается примесь черной березы, липы и ясеня.
Темнохвойные таежные леса с буро-таежными иллювиальногумусовыми оподзоленными кислыми почвами занимают около трети всей лесопокрытой площади юга страны. В северной (от 50° с. ш.) таежной половине материковой части и на Сахалине эти леса зональные, хотя и располагаются они, как правило, по горным склонам вследствие температурных инверсий60 и большой заболоченности межгорных равнин. В зону смешанных лесов они вклиниваются по горным хребтам, образуя высотный пояс, границы которого поднимаются с севера на юг. В южном Сихотэ-Алине они находятся на высоте от 900 до 1400 м (верхний предел).
60 Инверсия температуры - повышение температуры воздуха с высотой в некотором слое атмосферы вместо обычного понижения.
288
Для темнохвойных лесов Амуро-Уссурийского края характерен флористический комплекс охотско-камчатского типа. Он менее разнообразен, чем маньчжурская флора, зато полнее насыщен эндемичными видами. Темнохвойные леса без перерыва развиваются здесь с неогена, являются реликтовыми, имеют много родственных видов с таежными лесами Тихоокеанского побережья Северной Америки. Типичные представители охотско-камчатской флоры - ели аянская и мелкосемянная, пихты белокорая и сахалинская, а также несколько видов березы, объединяемых часто под общим названием каменные.
Строение темнохвойных охотских лесов намного проще маньчжурских. Первый ярус обычно составляют в Приамурье аянская ель, в Сихотэ-Алине и на Сахалине - близкая ей мелкосемянная ель. Это крупные деревья до 45 м высотой и диаметром до 1,2 м, с густой остроконечной кроной. К ели часто примешиваются каменные березы. Распространена также береза плосколистная и маньчжурская. Второй ярус обычно образуют на материке белокорая (амурская) пихта.. В этих лесах произрастает оригинальный желтый клен, продвигающийся севернее и выше всех широколиственных пород. В подлеске и кустарничково-травянистом ярусе характерны эндемичные виды: из кустарников - падуб морщинистый, бересклет большекрылый, колючая заманиха. Буро-таежные почвы пихтово-еловых дальневосточных лесов содержат много гумуса (8-11 %).
Светлохвойные таежные леса широко распространены в Приамурье и северной равнинной части Сахалина. Они занимают территорию с наиболее континентальным климатом и многолетнемерзлым грунтом на северо-западе и заболоченные низкие равнины на северо-востоке юга страны. Ценозы лесов представлены флористическим комплексом восточносибирского типа. Главную роль в древостое, как и в Восточной Сибири, играет даурская лиственница, уступающая свое место за водоразделами ближайших к тихоокеанскому побережью хребтов охотской лиственнице. В северо-западной части в лиственничных лесах почвы горные мерзлотно-подзолистые, на северо-востоке и севере Сахалина - подзолистые, подзолисто-болотные и торфяно-глеевые. В отличие от лиственничной тайги Восточной Сибири почвы часто иллювиально-железисто-гумусовые.
К хорошо дренированным участкам речных долин и предгорий приурочены травяные лиственничники с хорошим подлеском и густым травяным покровом, преимущественно из злаков - вейник Лангсдорфа. В ареале хвойных лесов благодаря пожарам и порубкам на значительных участках имеются и временные производные леса с большим участием в древостое берез: елово-березовые, лиственнично-березовые, чистые березняки.
Далее остановимся на характеристике нескольких наиболее важных в хозяйственном отношении видах лесных древесных растений, характерных для этого обширного и очень своеобразного региона.
289
Сосна кедровая корейская - Pinus koraiensis Sieb. et Zucc.
Мощное дерево уссурийской тайги, достигающее более 40 м высоты. Стволы полнодревесные, покрыты толстой темной корой. Кроны ширококонусовидные, низко посаженные и часто многовершинные. Крепкие ветви далеко простираются в стороны и концами несколько приподнимаются. Сосна корейская очень похожа на сибирскую сосну. Отличается менее толстыми побегами, покрытыми густым рыжеватым пушком. Плотная сизовато-зеленая хвоя (пять хвоинок в пучке) длиной 7-15 (20) см держится 2-3 года, но сидит менее густо. Она более шероховата (из-за зубчиков), чем у сосны сибирской, и с более сизоватым оттенком. Женские шишечки при цветении красноватые, затем фиолетовые, а при созревании становятся бурыми. Сидят на ножках. Шишки цилиндрические, очень крупные (15 см и более). Семенные чешуи густо налегают друг на друга и заканчиваются почти ромбическим щитком с острым, несколько волнистым краем, на конце с вытянутой и загнутой наружу верхушкой и пупком (апофизом). По созревании осенью второго года (в октябре) сильно оттягивают своим грузом сучья и опадают вместе с семенами. Сидящие внутри шишки бескрылые семена ("орехи") 1,5-1,7 см длиной с толстой деревянистой кожурой. 1000 семян весят до 500 г (рис. 83).
Рис. 83. Сосна кедровая корейская: 1 -укороченный побег с хвоей; 2 - поперечный разрез хвои с одним проводящим пучком в центре; 3 - зрелая шишка; 4 - семя (из Коровин, Зуихина, Потапова, 2003)
Северная граница ареала сосны корейской проходит примерно около 50° с. ш., вплоть до Буреинских гор. Она встречается также в Приморском крае и в южной части Хабаровского края, в горах Северо-Восточного Китая и Северной Кореи. Относительно зимостойка, но меньше, чем другие кедровые сосны. Теневынослива. Возобновляется под пологом леса. Растет на сухих горных склонах и гривах хребтов. Встречается на более плодородных, умеренно влажных почвах. Заболоченных мест избегает, так как повреждается здесь гнилью.
Не переносит пожаров.
Корневая система часто поверхностная, поэтому деревья при изрежи-вании вываливаются ветром. Растет в первые годы очень медленно, позднее убыстряет рост. В 20 лет едва достигает 2,5-2,8 м, к 40 годам - 5 м и к 100 годам имеет высоту 15-20 м. Живет до 500 лет и дольше.
290
Древесина кедра корейского розоватого цвета с желтовато-белой заболонью и приятным смолистым запахом. Ядро более темной окраски, составляет в среднем 83 % по диаметру. Древесина легкая; содержит большое количество смоляных ходов, особенно в наружной части ствола. Высоко ценится как строевой и поделочный материал. Она не коробится, не растрескивается, легко обрабатывается и поддается окрашиванию. По анатомическим признакам мало отличается от сосны обыкновенной.
Большая часть встречающихся в лесу кедров имеет возраст 250-300 лет. Предельные объемы 15-18 м3.
Кедр теневынослив, особенно в молодом возрасте, но меньше, чем ель аянская и пихта белокорая. С возрастом требовательность его к свету повышается.
Полог сложных насаждений кедра, являясь совокупностью разъединенных разновысотных горизонтальных пологов и изменяясь от одной группы деревьев к другой, образует вертикальную сомкнутость. Характерна, а местами и обильна, внеярусная растительность из многочисленных лиан и эпифитных споровых растений, которая подчеркивает своеобразие вертикальной сомкнутости девственных кедровников и создает весьма оригинальный ландшафт, напоминающий субтропический.
Кедрово-широколиственные леса в большинстве перестойные. Одновременно эти леса разновозрастны с колебанием возрастов растений от 1 до 500 лет.
При большой продолжительности жизненного цикла кедра одни деревья его постепенно отмирают естественной смертью, превращаясь в сухостой, другие гибнут по иным, часто случайным, причинам, не достигнув предельного возраста. Кроме того, усыхают породы-компоненты с более коротким жизненным циклом. В результате в кедровниках накапливается большое количество мертвого леса как в виде стоящих на корню сухостойных, так и упавших деревьев, составляющих почти половину общего запаса. Третья часть запаса, а местами и больше, представлена деревьями, пораженными различными гнилями и вредными насекомыми. Общие запасы насаждений, не тронутых рубкой кедровников, не так велики, как они могли бы быть, если бы за лесом осуществлялся уход, и в среднем составляют 300-400 м3 на 1 га.
Ель аянская - Picea ajanensis Fisch.
Дерево до 50 м высоты с пирамидальной кроной и серо-бурой корой, отслаивающейся пластинками. Молодые ветви блестящие, желто-бурые или буро-серые, голые, реже волосистые, с короткими тупыми смолистыми буроватыми почками. Особенно характерна плоская, слегка килеватая с обеих сторон хвоя длиной 1,2-2 см. Обращенная вверх нижняя сторона хвои темнозеленая, блестящая, с обратной стороны имеются две синевато-белые полоски с шестью-семью рядами устьиц. По настильному расположению хвои напоминает ель сибирскую. На освещенных побегах хвоя сидит изогнуто, по
291
этому открывается беловатая сторона хвоинок, и крона издалека кажется сизоватой. Листовые подушечки часто отогнуты назад. Женские шишечки при цветении зеленоватые или пурпурные, зрелые - светло-бурые. В отличие от шишек ели сибирской шишки у ели аянской рыхлы, легко сжимаются рукой. Кожистые, тонкие и довольно мягкие продолговато-эллиптические семенные чешуи заканчиваются волнисто-зубчатым или выемчатым, часто как бы обрубленным верхним краем (рис. 84).
Рис. 84. Ель аянская: 1 - общий вид дерева; 2 - зрелая раскрывшаяся шишка; 3 - побег с хвоей; 4 - схематический разрез хвои; сх - смоляной ход; пи - проводящий пучок (из Коровин, Зуихина, Потапова, 2003)
Шишки созревают и раскрываются в начале сентября, чему немало способствует сухость воздуха. Длина их 3-8 см. Небольшие, длиной 2-3,5 мм, темно-коричневые семена снабжены длинным, по краю мелкозубчатым крылышком. Отличаются невысокой всхо
жестью (около 60%).
Ель аянская - типично горная порода. Растет на Дальнем Востоке. На севере достигает склонов Колымского хребта и выходит на Охотское море около г. Охотска, занимая береговую полосу и почти не заходя в Якутскую АССР. Южнее охватывает более широкую область, проникая в северо-восточные районы Китайской народной республики. В горах Сихотэ-Алинь растет не ниже 500 м над уровнем моря, в более северных условиях и на Сахалине растет также в долинах. Встречается и на Камчатке (рис. 85).
Рис. 85. Ареал ели аянской в пределах Российской Федерации (из Бенько-ва, Швейнгрубер, 2004)
Морозостойка. Теневынослива. Лучше всего растет на свежих или умеренно влажных суглинках, но образует леса и на каменистых горных склонах. Бедных песчаных
и заболоченных торфяных почв не выносит. Избегает открытых склонов, подверженных действию морских ветров. Входит в число основных пород охотской тайги наряду с пихтой белокорой, лиственницей даурской и березой каменной.
Растет ель аянская довольно медленно, достигая в горах в 20 лет высоты 2 м, в 40 лет - 4,5 м, в 80 лет-13,1 м, в 100 лет - 16,8 м, в 200 лет - 26,3 м. Живет до 300 лет. В культуре встречается редко. В некоторых городах растет
292
в виде декоративных деревьев, плодоносит, но более других елей страдает в молодом возрасте от поздних заморозков.
По анатомическим признакам древесина ели аянской существенно не отличается от других видов ели, произрастающих на территории России. Однако вследствие медленного роста она формирует узкие годичные слои радиального прироста ствола, что положительно сказывается на механических свойствах ее древесины.
Древесина используется для строительных целей, столярных работ, как сырье для производства целлюлозы, для изготовления тары, спичечной соломки, стружечных плит.
Пихта белокорая - Abies nephrolepis (Trautv.) Maxim.
Растет в горах Дальнего Востока, Северного Китая, Кореи. Является одним из образователей темнохвойной тайги Дальнего Востока.
Стройное дерево до 30 м высотой, с густой конусовидной или пирамидальной кроной. Кора ствола гладкая, светло-серая, значительно светлее, чем у других видов (за что и получила свое название), у старых деревьев - темнеет и растрескивается. Молодые побеги желтовато-бурые, слабо-опушенные. Хвоя на концах слегка раздвоенная, сверху блестящая, темнозеленая, 1-3 см, снизу с двумя беловатыми устичными полосками, расположена гребенчато, функционирует 4-5 лет. Пыление происходит перед распусканием молодой хвои. Незрелые шишки фиолетовые, зрелые - бурые, до 6 см длины (рис. 86).
Зимостойка. Растет быстро. Требовательна к влажности воздуха и почвы, теневынослива, менее долговечна, чем другие виды, в естественных условиях доживает до 150-180 лет. В возрасте 15-20 лет нередко поражается гнилью. Удовлетворительно переносит городские условия. Благодаря изящной, темной кроне, контрастирующей со светлым стволом, несколько поникающим ветвям, декоративным шишкам заслуживает широкого использования в одиночных и групповых посадках в парках, лесопарках, в сложных композициях. В культуре недавно.
Древесина безъядровая, светло-желтая. Используется в судостроении, в строительстве мостов - сваи. Пригодна для получения пиломатериалов и целлюлозы. По анатомическому строению практически не отличается от пихты сибирской.
Пихта цельнолистная - Abies holophylla Maxim.
Встречается в южной части Приморья, Северном Китае, Корее.
Стройное, мощное дерево до 45 м высотой, с густой, широкопирамидальной, рыхлой, опущенной до земли кроной, в старости - плосковершинной. По ряду морфологических признаков хорошо отличается от других видов рода. У нее необычная для пихт темно-серая, к старости почти черная кора, сначала шелушащаяся, позднее горизонтально растрескивающаяся, на однолетних побегах охристая или желтовато-серая, продольно-желобчатая.
293
Хвоя очень плотная, жесткая, цельная, заостренная, сверху блестящая, темно-зеленая, снизу светлее, расположена на побегах гребенчато; значительно длиннее, чем у других видов пихты. Продолжительность жизни хвои 8-9 лет. Шишки цилиндрические, тупые, светло-коричневые, бархатисто-опушенные, осмоленные (рис. 86).
Рис. 86. 1 - пихта цельнолистная: 1а - зрелая шишка; 16 - семенная и кроющая чешуи зрелой шишки; 1в - семя; 1г - поперечный разрез хвои; 1д - побег с хвоей, заостренной верхушкой. 2 - пихта белокорая: 2а - зрелая шишка на побеге, верхушки хвои раздвоены; 26 - семенная и кроющая чешуи; 2в - семя; 2г - округлые слабо засмоленные почки; 2д - поперечный разрез хвои (из Коровин, Зуихина, Потапова, 2003)
До 6-10 лет растет медленно, затем рост резко усиливается, по темпам роста превосходит ряд других хвойных пород. Зимостойка. Теневынослива, но может развиваться и при полном освещении. Ветроустойчива, требовательна к богатству и влажности почвы и воздуха. Долговечна, живет 300— 400 лет. Размножается семенами и прививкой на другие виды.
Древесина по анатомическим признакам подобна ранее рассмотренным видам пихты. Используется аналогично другим видам этого рода. Заслуживает самого широкого применения в садово-парковом строительстве.
294
Тис остроконечный - Taxus cuspidata Siebold et Zucc. ex Endl.
Растет в Приморском и Хабаровском краях, на Сахалине и Курильских островах. По побережью Японского моря и Татарского пролива, а также по западным склонам и отрогам Сихотэ-Алиня, на высотах до 800-900 м над уровнем моря. При таком обширном ареале тис является на Дальнем Востоке одной из редко встречающихся пород. Растет одиночно или группами разновозрастных деревьев, обычно на склонах и в ущельях среди нетронутых тенистых смешанных лесов и с преобладанием хвойных пород. На морских островах в Приморье (о. Петрова, о. Наумова и др.) образует самостоятельные тисовые насаждения. На юге Приморья изредка образует так называемые тисовые рощи, состоящие из 10-20 рядом растущих экземпляров. Столь же редок близкородственный вид - тис ягодный (Taxus baccata) - в лесах Кавказа. Последний -один из рекордсменов долгожителей: считается, что он может доживать до 3-4 тысяч лет. Эти виды разобщены географически, морфологически же они почти не различаются.
На юге ареала деревья достигают 15-18, изредка - 20 м высоты и 0,8-1 м в диаметре. В более северных районах размеры деревьев значительно меньше, а у северной границы распространения и на предельных высотах произрастания тис приобретает низкорослую или стланиковую форму. Стволы нестройные, сбежистые, в поперечном сечении неправильной формы. Кора красно-бурая, тонкая, продольно отслаивающаяся. Крона густая, обычно неправильная, с несимметрично распростертыми ветвями. Молодые побеги зеленые, гибкие. Хвоя плоская, мягкая, 1,2-2,5 см длины и 2-2,5 мм ширины, остроконечная, сверху темно-зеленая, глянцевая, снизу - светлее, сидит одиночно, расположена гребенчато, держится 4-6 лет (рис. 87). Растение двудомное, редко - однодомное. Микростробилы мелкие, шаровидные, в виде желтоватых «букетиков» из нескольких микроспорофиллов, сидят в пазухах хвои. Женские семязачатки зеленоватые, состоят из одной семяпочки, окруженной чешуйками, сидят по одиночке в пазухах хвои. «Цветет» (пылит) в мае - июне. Пыльца без летательных мешков. Опыляется ветром. Семена созревают в год цветения - к сентябрю. В процессе развития семени из чешуек, окружающих семяпочку, образуется мясистый ярко-красный присемянник, из которого видна верхняя часть семени. Семя одиночное, яйцевидное, с заострённой верхушкой, светло-коричневатое, около 6 мм длины и 4 мм в диаметре. Корневая система неглубокая, но мощная, без заметно выраженного стержневого корня.
Требователен к плодородию и влажности почвы. Лучшие почвы - рыхлые, перегнойные и хорошо дренированные, не выносит тяжелых глинистых и кислых. Требователен к влажности воздуха. Устойчив к низким температурам, на севере ареала выдерживает морозы до 40 и более градусов. Самая теневыносливая из всех дальневосточных древесных пород. Растет медленно, к 10 годам достигает высоты 25-30 см, а к 200 годам - 11-12 м высоты и 36-40 см в диаметре. Доживает до 700-800 и более лет. Точно судить о возрасте трудно: он иногда образует ложные годичные кольца, а крупные стволы часто бывают гнилы и дуплисты. Пни срубленных деревьев образуют поросль, но обычно
295
слабую, не обеспечивающую возобновления. Возобновляется семенами и частично корневой порослью. В культурах разводится семенами.
Рис. 87. Тис остроконечный: 1 - побег со зрелыми семенами в присемянниках; 2 - побег с мужскими стробилами; 3 - микростробил в пазухе хвоинки, видны микроспорофиллы; 4 - семязачаток в пазухе хвоинки; 5 - продольный разрез семязачатка; 6 -продольный разрез семени с присемянником
Древесина мелкослойная, с узкой светло-желтой заболонью с красно-бурым ядром (отсюда народное название - «красное дерево»), плотная, твердая, упругая, трудно колется, но сравнительно хрупкая, не содержит смолы (как и кора), очень стойкая против гниения («негной-дерево»), хорошо полируется На воздухе темнеет, приобретая цвет «черного дерева»; выдержанная в воде, становится фиолетово-пунцовой, а пропитанная известковой водой - фиолетово красной.
Пригодна на детали и отделку мебели, на токарные, резные, столярные, суве
нирные изделия, деревянные музыкальные инструменты, а в виде строганного шпона может использоваться на отделку мебели, радиоприемников, телевизоров и других изделий.
Все вегетативные части тиса и семена содержат токсичный для многих животных, в том числе и для человека, алкалоид - таксин (от слова "taxus"). Видимо поэтому долговечна. Только сочные красные присемянники неядовиты, сладковаты и съедобны, отравления ими не наблюдалось.
Тис интересен для зеленого строительства: легко переносит стрижку, не страдает от задымления, загазованности и запыленности воздуха. Пригоден для озеленения в районах с влажным воздухом. Культивируется на Сахалине, в Ленинграде, Москве. В искусственных посадках Западной Европы и Америки известен с середины XIX века.
Оба, произрастающие на территории нашей страны, вида внесены в "Красную книгу". Это - и реликты, известные с Юрского периода, и ценные во многих отношениях растения. О промышленной заготовке древесины представителей семейства тисовых не может быть и речи. В настоящий учебник виды из семейства тисовых включены как пример крайне редких и заслуживающих особого внимания древесных растений.
296
Береза ребристая (желтая) - Betula costata Trautv. и другие дальневосточные виды березы
Береза ребристая или желтая - дерево до 25-27 м высоты и до 80 см в диаметре. Кора светло-желтого или розовато-желтого цвета, у крупных деревьев снизу глубоко-трещиноватая.
Листья удлиненно-овальные, с вытянутой острой верхушкой, мелкопильчатые по краям, с 12-14 парами жилок Женские сережки короткие, широкие. По форме листьев напоминает граб (рис. 88). Встречается главным образом в нетронутых горных лесах Дальнего Востока. Наиболее теневыносливая из всех берез.
Рис. 88. Береза ребристая: 1 - часть побега с листьями и соплодием (сережкой); 2 - присеменная чешуйка; 3 - семя (орех) с узкими крыловидными придатками. Чешуйка и семя увеличены
Древесина желтоватая твердая. Применяется в мебельном производстве и для изготовления паркета. По анатомическим признакам все рассматриваемые нами виды березы мало различаются.
Помимо березы ребристой в лесах Дальнего Востока встречается еще более 20 видов березы, некоторые из них представляют интерес для лесозаготовителей. Древесина этих берез используется в целлюлозно-бумажной промышленности, в мебельном производстве, в качестве дров.
Береза плосколистная - В. platyfylla Su-kacz. - наиболее распространенный на Дальнем
Востоке вид березы, замещающий там березу повислую. Растет в Амурской области, Хабаровском крае, на севере Приморья. Дерево 25-27 м высоты, до 0.6 м в диаметре.
Береза даурская - В. davurica Pall. - высота стволов 20-25 м, диаметр до 0,7. Растет в составе лиственных лесов Забайкалья, Приамурья и Приморского края. Стволы прямые, полнодревесные. Кора темно-серая иногда с коричневатым оттенком. Береста отслаивается многочисленными,
мелкими, далеко не отходящими от ствола пластинками, из-за чего поверхность ствола на ощупь мягкая. Древесина по плотности уступает лишь березам железной и ребристой.
Береза каменная (шерстистая) - В. lanata (Regel) V. Vassil. - дерево до 20 м высоты 0,5 м в диаметре. Растет на материковой части Дальнего Востока, ближе к вершинам хребтов образует чистые насаждения, но здесь ее высота доходит до 45 м. Одна из долговечных берез - доживает до 250-
297
300 лет. Древесина твердая, мелкослойная, тяжелая, колется и обрабатывается с трудом, что делает ее незаменимой для всякого рода мелких поделок.
Дуб монгольский - Quercus mongolica Fish, ex Ledeb.
Крупное дерево, достигающее высоты 25-27 м и 70 см в диаметре. Корневая система глубокая. Морфологически близок к дубу скальному. Молодые побеги ребристо-бугорчатые, голые коричневато-зеленые. Почки яйцевидные, острые. Листья почти сидячие. Отличаются от листьев скального дуба следующими признаками: 1) более плотные (кожистые); 2) менее глубоко-лопастные; 3) имеют большое число боковых лопастей (рис. 89).
Рис. 89. Дуб монгольский: 1 - мужские соцветия (рыхлые сережки); 2 - побег с листьями и плюсками после выпадения желудей; 3 - желудь (из Меницкий, 1984)
Дуб монгольский распространен по всему югу Дальнего Востока.
В кедрово-широколиственных лесах дуб входит в состав насаждений одной-двумя единицами состава. В этих древостоях он формирует наиболее пол
298
но древесные, хорошо очищенные от сучьев стволы. В чистых дубняках на восточных склонах Сихоте-Алиня и низкогорьях западных склонов дуб монгольский не бывает выше III бонитета, стволы часто неправильной формы с низко опущенной кроной и толстыми сучьями.
Очень часто дубовые леса формируются в предгорьях и на склонах под влиянием пожаров, так как сам дуб довольно устойчив к этому бедствию. Занимает он и вырубки.
До последнего времени древесина дуба монгольского невысоко ценилась и использовалась лишь на шпалы и дрова. Однако в настоящее время ситуация изменилась. Заготовители и потребители древесины обратили внимание на то, что этот материал не уступает по качеству древесине европейских дубов. Что же касается запасов, то древесина дуба монгольского составляет примерно 9/10 от общих запасов дуба в нашей стране.
Орех маньчжурский - Juglans mandshurica Maxim.
Орех маньчжурский растет в Приамурье и Приморье преимущественно по долинам рек, не поднимаясь выше 500 (600) м над уровнем моря, в древостоях хвойно-широколиственных и широколиственных лесов. Занимает плодородные аллювиальные или делювиальные глубокие рыхлые почвы с близким залеганием проточных грунтовых вод. Достигает 27-29 м в высоту и 60-70 (до 100) см в диаметре. До 80-90 лет растет быстро, затем прирост сокращается. Живет 200-250 лет. Плодоносит в насаждениях с 12-15 лет, а при одиночном произрастании - с 4-8 лет (рис. 90).
Кора темно-серая, у молодых деревьев гладкая, у старых - трещиноватая. Побеги желтовато-коричневые, опушенные; почки крупные, желтоватые, мягкочешуйчатые, бархатистоопушенные. Листья до 1 м длины из 11-19 листочков; листочки продолговато-эллиптические, со слегка оттянутой верхушкой, мелкопильчатые (рис. 90), до 20 см длиной и 12 см шириной, с резко выраженными жилками, снизу железистоопушенные, при растирании издают йодистый запах.
Мужские сережки очень длинные (до 25 см), пестичные цветки с двухлопастными ярко-розовыми рыльцами, сидят по 3-10 и более на коротких опушенных цветоносах, расположенных на концах растущих побегов. Плоды в нераскрывающейся мясистой оболочке, покрытой рыжеватыми волосками с желёзками, от продолговатых до округлых, с заостренной вершиной, созревают осенью и быстро опадают.
На земле оболочка разрушается, высвобождая костянки-орехи. Они темно-бурые, 27-45 мм длиной и 22-34 мм в диаметре, сверху заостренные, с неровной поверхностью, изрытой неправильными углублениями, и с выдающимися продольно-извилистыми гладкими ребрами.
299
Рис. 90. Плоды ореха маньчжурского (фото П.Г. Мельника)
Орехи широко используют в кондитерской промышленности, ореховое масло применяют для различных технических целей, дробленые орехи идут на корм птице и домашним животным, В лесах орехи являются кормом для кабанов, медведей, белок и других зверей. Орех маньчжурский - хороший медонос, имеет ценную древесину. По анатомическим признакам древесина близка к ореху грецкому, имеет то же применение.
Ясень маньчжурский - Fraxinus mandshurica Rupr.
Дерево первой величины. Кора светло-серая. Листья непарноперистые, из 5-11 листочков. Листочки косо-овальные, продолговатые или ланцетные, с длинным тонким заостренным концом, пильчато-зубчатые, в углах жилок с рыжеватыми бородками или пушком. Главный черешок шероховатый. Цветки однополые в густых метелках, без чашечки и венчика, расположены двудомно. Тычиночные соцветия после цветения опадают. Крылатки ланцетной формы, слегка заострены или закруглены на концах. Орешек почти равен по длине крылу.
Естественно растет на Дальнем Востоке, начиная от низовий Амура и вплоть до южных границ Приморского края. Встречается также в Северо-восточном Китае, Корее и Японии. Является замещающим видом по отно
300
шению к ясеню обыкновенному на Дальнем Востоке. Тяготеет к берегам ручьев и речек.
Имеет большое лесопромышленное значение из-за ценной древесины и значительных запасов. Древесина ядровая, кольцесосудистая, твердая, серо-коричневая. Применяется преимущественно в мебельной промышленности в виде шпона или монолита. По анатомическим признакам мало отличается от дуба черешчатого.
Виды рода липа - Tilia Mill.
Липа амурская - Т. amurensis Rupr. Дерево до 25 м высоты и до 1 м в диаметре. С темно-серой бороздчатой и продольно отслаивающейся корой. Побеги вначале шелковистые, позже голые, красновато-коричневые. Листья плодоносящих побегов овальные или широкояйцевидные, 5-7 см длины и почти равной ширины, при распускании густо шелковисто опушены. Позднее голые лишь с бородками волосков в углах жилок. Основание листа сердцевидное (рис. 91). Отличается от липы мелколистной шелковистым, белым опушением молодых побегов; зубцы по краю листа постепенно оттянуты; цветки более крупные до 1,6 см в диаметре, плоды бело-опушенные.
Рис. 91. Листья и соплодия дальневосточных видов липы: 1 - липа маньчжурская; 2 - липа Таке; 3 - липа амурская (из Усенко, 1984)
Ареал: вид замещающий липу мелколистную в широколиственных лесах Дальнего Востока. Хозяйственное использование такое же, как и у липы мелколистной.
Липа маньчжурская - Т. mandshurica Rupr. Дерево до 20 м высоты, часто от основания многоствольное. Кора старых стволов серо-бурая, трещиноватая. Побеги покрыты густым опушением из звездчатых и прямых волосков. Листья крупные, округлые или широкояйцевидные до 10-20 см в диаметре, с крупными остроконечными зубцами до 8-10 мм длиной. Листья сверху голые, снизу густо опушенные звездчатыми волосками. Базальных жилок 6-10. Прицветный лист 7-12 см длиной, 1,3-2,5 см шириной, сидячий, в нижней части закругленный, густо опушенный. Соцветие из 8-12 цветков. Цветки 10-15
мм в диаметре, лепестки лимонно-желтые, чашелистики опушенные. Плоды шаровидные до 1см в диаметре, без ребер.
Ареал: растет в Приморском и на юге Хабаровского края в кедровошироколиственных и дубовых лесах; теневынослива, более теплолюбива, чем липа амурская. Ценный медонос. В культуре встречается в основном в ботанических садах и дендрариях.
301
Липа Таке - Т. taquetii Schneid. Высота до 20-25 м, диаметр до 1 м и более. Кора серая продольно отслаивающаяся. Молодые побеги густо опушены рыжеватыми звездчатыми волосками, с возрастом волоски опадают. Черешки листьев густо опушены. Листья округлояйцевидные, 3,5- 7 см длины с сердцевидным основанием; пильчато-зубчатые, плотные, сверху голые, снизу - сизоватые. Цветки палевые, почти белые, 10-12 мм в диаметре, в 3-5-цветковых соцветиях. Прицветный лист 3-5 см длины, цветонос отделяется на 1-2 см от основания. Орехи шаровидные, 5 мм в диаметре, слаборебристые, опушенные.
Все виды липы в Приморье (насчитывается еще как минимум 6 видов) и на юге Хабаровского края - ценные медоносы (во время цветения липы один улей дает до 200 кг меда), поэтому при лесозаготовках эти деревья стараются сохранить. Старые деревья формируют очень широкие кроны, на 1 га размещается не более 4-5 экземпляров. В дуплах старых лип часто зимуют белогрудые медведи.
Виды рода клен - Acer L.
На дальнем Востоке насчитывается порядка 12 видов клена. В основном это деревья второй величины и подлесочные породы. Древесина у кленов рассеяннососудистая, заболонная (ядро не выражено), по макроскопическим признакам однородная, белая или слегка желтоватая. Тяжелая, твердая (кроме клена зеленокорого), хорошо обрабатывается и полируется. Используется древесина для столярно-токарных изделий, для изготовления фанеры, паркета, рукояток столярных инструментов, ружейных лож, спортивного инвентаря, музыкальных инструментов. По анатомическим особенностям незначительно отличается от клена остролистного. Дрова по калорийности близки к дубовым и ясеневым. В соке при весеннем "плаче" содержится до 1,2- 3 % сахара. Все виды клена - ценные медоносы.
Клен мелколистный (клен моно) - Acer mono Maxim. Типичное дерево второго или третьего, реже - первого яруса. Высота в пределах 16-24 м, диаметр до 60 см. Побеги желтоватые. Кора серая с продольными неглубокими трещинами. Листья обычно пятилопастные (иногда у основания листа имеется еще одна пара небольших лопастей); 6-12 см длины, почти такой же ширины, плотные, голые, с плоским или слегка клиновидным основанием (рис. 92). Лопасти длинноостроконечные, цельнокрайние с 1-2 зубцами на крупных лопастях. Цветки светло-желтые, 6-8 мм в диаметре, в щитковидном соцветии 15-30 цветков. Крылатки 1,5-Зсм длины, расходятся под прямым или тупым углом. Цветет в мае, плоды созревают в сентябре. К почве малотребователен, теневынослив. Растет медленно, к 100 годам достигает высоты 14-16 м. Доживает до 250 и более лет.
302
Рис. 92. Листья и плоды дальневосточных кленов: 1 -клен маньчжурский; 2 - клен мелколистный (из Усенко, 1984)
Клен маньчжурский - A. mandshuricum Maxim. Стройное дерево до 18-22 м высоты и 50-60 см в диаметре с серой или буровато-серой мелкотрещиноватой корой. Побеги голые красновато-коричневые. Листья сложные, тройчатые; листочки ланцетные или продолговато-эллиптические, по размеру все равны или средний чуть крупнее, 5-8 см длины и до 2.5 см ширины. Черешки длинные, красноватые. Однодомен. Цветки зеленовато-желтые, тычиночные и ложно-обоеполые, собраны в 3-5-цветковые щитко-
вижные соцветия. Крылатки 3-3,5 см длины, расходятся под прямым или тупым углом. Встречаются трехкрылые плоды. Тенелюбив. Сначала растет медленно, затем быстро, обгоняя клен мелколистный. Стволы хорошо очищаются
от сучьев.
Виды рода ильм - Ulmus L.
На Дальнем Востоке растут 4 вида этого рода.
Ильм сродный (долинный, японский) - U. japonica (Rehd.) Starg. Распространен в Приморском и Хабаровском краях, Амурской области, на Сахалине и Курилах. В хороших условиях это стройные деревья со светло-серой корой, достигают высоты 25-30 м и 1 м в диаметре. Листья варьируют по форме, чаще - обратнояйцевидные, неравнобокие у клиновидного основания, неравнопильчатые по краю, 3-12 см длины и 1,5-5 см. ширины, гладкие или шершавые, снизу голые или опушенные только по жилкам или же густоопушенные. Крылатка плода (ореха) 1-1.6 см длины и 0,4-0,8 см ширины, ее выемка доходит до орешка (рис.93). Доживает до 300-350 лет. Древесина кольцесосудистая, твердая, крепкая, упругая, вязкая. Заболонь желтовато-белая, узкая, ядро красновато-бурое. Лучи блестящие, хорошо заметные на радиальных поверхностях. Текстура красивая на продольных разрезах. В пропаренном виде хорошо гнется. Используется в судостроении, мебельном производстве, для изготовления лыж, паркета фанеры. В воде и сырой почве стойкая к гниению, поэтому используется для нижних венцов срубов, деталей мостов, свай. По анатомическим признакам трудно отличим от ильма (вяза) гладкого.
Ильм лопастный (горный) - U. laciniata (Trautv.) Мау г. Распространение примерно то же, что и у ильма сродного. Растет в смешанных лесах предгорий и по нижним склонам гор (до 600-700 м над уровнем моря). Деревья до 20-25 м высоты и до 55 см в диаметре. Побеги текущего года желтовато-серые. Кора светло-бурая или серая, трещиноватая, отслаивающаяся тонкими продольными пластинками.
303
Рис. 93. Особенности листьев двух ильмов Приморья: 1 -ильм сродный (долинный); 2 - ильм лопастный (горный) (из Усенко, 1984)
Листья крупные, особенно на концах побегов, до 8-20 см длины и до 8 - 12 ширины, шершавые, неправильно-пильчатые, с заметно клиновидным основанием, остроконечные, а на верхних ветках - 3-7-лопастной вершиной листовой пластинки, растение теневыносливое. Доживает до 200-230 лет. Древесина по макроскопическим признакам и механическим свойствам подобна ильму сродному, но более хрупкая.
Чозения толокнянколистная - Chosenia arbutifolia (Pall.) А. Skvorts.
Род чозения включает в себя всего 1 вид - чозению толокнянколистную, распространенную по поймам рек в Восточной Сибири и на Дальнем Востоке, включая Камчатку и Сахалин. Важнейший пионер пойменной древесной растительности от зоны тундры на севере до муссонных широколиственных лесов на юге ареала. В северных районах в высоту не превышает 8-10 м, на юге достигает 35-37 м высоты и более 1 м в диаметре (рис. 94).
Ствол малосбежистый; кора буровато-серая, отслаивающаяся пластинками. Крона пирамидальная; побеги гладкие, красные, с сизым налетом и темными поперечными полосками; почки когтеобразные, сплюснутые, розовые. Листья обратноланцетные, до 7 см длины и 2 см ширины, к основанию сужены, с заостренной или тупой верхушкой, цельнокрайние или пильчатые, молодые - сизые.
Дерево двудомное, на мужских особях сережки повислые, на женских -стоячие. Околоцветник отсутствует; тычинок 5, приросших к вздутой при-цветковой чешуе; пестик из 2 плодолистиков; завязь верхняя. Цветет чозения одновременно с облиствением, опыляется ветром; плоды - коробочки, созревают в середине-второй половине лета. Размножаются семенами, которые опадают сразу по созревании и прорастают через 2-3 дня. Однолетние сеянцы уже в первый год образуют сильно развитую корневую систему и достигают 30-40 см высоты. Это позволяет растениям выдерживать мощные потоки воды во время грозовых ливней и осенних паводков. Вегетативно чозения не возобновляется и не размножается. Растет исключительно быстро, образуя мощную корневую систему. Живет до 100-130 лет. Свето- и влаголюбива, требовательна к плодородию почвы, легко выдерживает паводковое затопление. В естественном ареале чозения вполне зимостойка, но при интродукции на Северо-западе России оказалась древесной породой, сильно обмерзающей, а в молодом возрасте даже вымерзающей совсем (например, в арборетуме Санкт-Петербургской лесотехнической академии).
304
Рис. 94. Чозения арбутолистная на юге Приморья (фото П.Г. Мельника)
Как быстрорастущее и декоративное дерево чозения представляет несомненный интерес для лесного хозяйства и озеленения в Российской Федерации. Образуемые ею пойменные леса имеют большое водоохранное и водорегулирующее значение.
Порода ядровая. Древесина легкая, мягкая, однородного тонкого строения с малой плотностью. Заболонь розовато-белая, шириной 3-6 см. Ядро буроваторозовое. Основной порок древесины - стволовая гниль. Используется в производстве спичек, мебели. По анатомическому строению древесина очень похожа на древесину представителей рода Populus.
Бархат амурский - Phello-dendron amurense Rupr.
Растение из семейства рутовые, на Сахалине встречается еще один вид из этого рода. Дерево до 25 м высотой, обычно же ниже и входит единичными экземплярами в состав второго яруса хвойношироколиственных и широколиственных насаждений Приморья, южной, Приамурской части Хабаровского края. Ствол покрыт мощным слоем морщинистой эластич
ной корки толщиной до 4 см, которую можно периодически снимать и использовать как термо-, звуко- и электроизоляционный материал.
Листья непарноперистые, очередные и супротивные на одних и тех же побегах. Цветки мелкие в верхушечных метелках, чаще раздельнополые (тогда деревья двудомные), но могут быть и обоеполыми. Цветет в начале лета. Плод -черная смолистая ягода шаровидной или грушевидной формы, 7-9 мм в диа
метре.
305
Древесина прочная и твердая, обладает высокими декоративными свойствами, заболонь светлая, почти белая, узкая; ядро темное, мягкого землистокоричневого тона. Хорошо обрабатывается, используется для сувенирных и художественных изделий.
Анатомическое строение древесины. Поперечный срез. Ядро имеется, граница годичных колец четкая, гладкая. Древесина кольцесосудистая. Просветы сосудов округлые. В ранней древесине просветы крупные, редкие, в основном одиночные; в поздней - очень мелкие, одиночные и в группах. Просветы поздней древесины окружены клетками паренхимы и сосудистыми трахеидами. Стенки сосудов тонкие. Осевая паренхима скудная, апо- и паратра-хеальная. В ранней древесине - скудновазицентрическая; в поздней - метатрахеальная и па-ратрахеальная (скудновазицентрическая, вазицентрическая и сливающаяся). Некоторые сосуды заполнены тилами с окрашенным содержимым. Тангенциальный срез. Лучи 3-5-рядные, веретеновидные, высотой до 30 клеток. Клетки лучей овальные и округлые. Тяжи осевой паренхимы состоят из 4-5 клеток. Радиальный срез. Перфорации конечных стенок члеников сосудов простые. Межсосудистая поровость очередная, поры округлые и овальные. Поры между сосудами и клетками лучей значительно мельче, частые, полуокаймленные, овальные и округлые. Лучи гомогенные и слабо гетерогенные, состоят из лежачих клеток или с одним рядом краевых квадратных клеток. Волокнистые элементы состоят из волокон либриформа, в небольшом количестве имеются сосудистые трахеиды. Видны спиральные утолщения на внутренней поверхности стенок сосудов и сосудистых трахеид.
Общие запасы древесины бархата амурского небольшие. Заготовка может вестись лишь попутно на разрабатываемых лесосеках.
Специалисты лесной промышленности в своей практической деятельности должны планировать и проводить лесозаготовительные мероприятия с учетом необходимости охраны и размножения редких представителей отечественной флоры, а всем работникам леса необходимо неукоснительно придерживаться нестареющего принципа старых русских лесоводов: рубка леса - синоним
лесовосстановления.
306
ПРИЛОЖЕНИЯ
307
Приложение 1
Геохронологическая шкала и последовательность возникновения организмов (годы х 106) (из Грин, Стаут, Тейлор, 1990)
Эра Пе- Эпоха Го- Животные Растения
риод Совре- ды
Четве менная (голо- 0,01 Господство человека
ртич-ный цен) Ледни- Современная флора
ковая 2 Появление человека
(плей-
Кайно- стоцен)
зойская Плио- 7 Распространение мле-
цен копитающих
Миоцен 26 Появление собак и „
Трети Олиго- Распространение цвет-медведеи
чный цен 38 _ ковых растении, осо- Появление человеке- г
Эоцен 54 - - бенно травянистых образных обезьян
Палео- 65 Лошади, парнокопыт-
цен ные, слоны
Исчезновение дино-
Мел 135 завров, появление со- _ ~ Доминирование цвет- временных рыб и пла- „ ковых растении
центарных млекопитающих
Мезо- Доминирование дино-
зойская Юра 195 завров, появление птиц Появление цветковых и млекопитающих, растений обилие насекомых
225 Появление динозавров, _ _ Обилие саговниковых
Триас распространение пре- и хвойных смыкающихся Распространение пре- Появление хвойных
Палео- Пермь 280 смыкающихся, появ-
зойская Кар-бон 350 ление жуков, исчезновение трилобитов Появление пресмы- Обилие древовидных кающихся и насеко- папоротников, напри-мых, распространение мер Lepidodendron, земноводных образующих "камен-
ноугольные леса"
Появление земновод- ных, аммонитов и пау- Первые мохообразные
Девон 400 кообразных, распро- и папоротникообраз-странение хрящевых и ные костных рыб Появление челюстно- „
Силур 440 Первые споровые со-ротых, первые корал- . судистые растения ловые рифы
308
Ордовик
Кембрий
Появление позвоночных бесчелюстных, 500 обилие трилобитов, моллюсков и ракообразных Появление всех типов
Археозойская
Докем брий
570 беспозвоночных и иг-
локожих
1 000 Примитивные много-
клеточные
2000 Примитивные эвка-
риоты
Сине-зеленые водо-
3000 росли, бактерии (про-
кариоты)
3500 _ ? Возникновение жизни
5000 Образование Земли
309
Приложение 2
310
Типы поровости продольных трахеид хвойных и их полей перекреста: А — агатис; Б — таксодиум (болотный кипарис); В — кипарис; Г — сосна корейская, Д — сосна обыкновенная; Е — ель обыкновенная;
а — апертура поры, а.п — араукариоидная (очередная) поровость, в.О — вырост оболочки, Д.С.п — двурядная супротивная поровость, к.п — купрессоидная поровость, л.тр — лучевая трахеида, О — окаймление, О. пр — окаймленная пора, О.С.п — однорядная супротивная поровость, ок.п — оконцевая поровость, п.к.л — паренхимная клетка луча, п.п — пиноидная поровость, п.п.п — пицеоидная поровость, п.пер — поля перекреста, пр.п — простая пора, т.п — таксодиоидная поровость, тр — трахеида (Тимонин, из Лотовой и Тимонина, 1990) (из Лотова, Нилова, Рудько, 2007)
311
Приложение 3
Дополнительные сведения по систематике
ЦАРСТВО ГРИБЫ
Грибы долгое время считались бесхлорофилльными растениями и рассматривались в курсе ботаники. В настоящее же время признано, что эта обширная группа организмов представляет собой отдельное царство, изучением представителей которого занимается самостоятельная наука - микология.
Грибы - это одна из самых больших и процветающих групп организмов; к ней относится около 80 000 описанных видов. Размеры грибов колеблются от одноклеточных дрожжей до "больших" белых, подосиновиков, и прочих всем известных съедобных грибов и так называемых "поганок" (в последних примерах речь идет не о целых грибах, толь об их плодовых телах). Грибы занимают самые разные местообитания как в воде, так и на суше. Кроме того, грибы имеют важное значение и в связи с той ролью, которую они играют в биосфере, и в связи с тем, что они используются людьми для медицинских и хозяйственных целей.
К грибам относятся бесчисленные плесени, растущие на сыром органическом материале (хлебе, коже, разлагающейся растительности), одноклеточные дрожжи, которые в изобилии появляются на сахаристой поверхности спелых фруктов, и многие паразиты растений. Последние вызывают такие опасные заболевания посевов, как мучнистая роса, головня и ржавчина. Многие грибы являются опасными, в том числе и карантинными, вредителями растущего леса и древесины.
Основные свойства и систематика грибов
Грибы - это эвкариоты, утратившие хлорофилл или никогда не имевшие его, и, следовательно, они являются такими же гетеротрофами, как животные. Вместе с тем у них имеется жесткая клеточная стенка, и они не способны передвигаться, как и растения.
Строение тела грибов уникально. Оно состоит из массы тонких ветвящихся трубчатых нитей, которые называются гифами (в единственном числе - гифа), а вся эта масса гиф называется мицелием. Каждая гифа окружена тонкой жесткой стенкой, основным компонентом которой является хитин - азотсодержащий полисахарид. Хитин является также структурным компонентом наружного скелета членистоногих. В некоторых случаях клеточная стенка содержит целлюлозу. Гифы не имеют клеточного строения. Протоплазма гиф либо совсем не разделена, либо разделяется поперечными перегородками, которые называют септами. Такие септы делят содержимое гиф на отдельные отсеки (компартмен-ты), внешне похожие на клетки. В отличие от нормальных клеточных стенок образование септ не связано с делениями ядер. В цитоплазме гиф располагаются митохондрии, аппарат Гольджи, эндоплазматический ретикулум, рибосомы, вакуоли и другие органеллы, обычные для эвкариот. В старых участках мицелия вакуоли крупнее, а цитоплазма занимает лишь небольшое место на периферии. Время от времени гифы агрегируют с образованием более плотных структур, как, например, плодовые тела, которые и собирают любители съедобных грибов.
В настоящее время царство грибы (Fungi) принято делить на 6 отделов: оомикота (Oomycota), зигомикота (Zygomycota), аскомикота (Ascomycota), базиоиомикота (Basi-diomycota), хитридио.микота (Chytridiomycota) и дебтеро.микота (Deuteromycota). Предпоследний из перечисленных отделов представлен небольшой группой микроскопических (преимущественно одноклеточных) грибов; последний - дейтеромикота - относится к так называемым несовершенным грибам, которые размножаются только бесполым путем. Это один из крупнейших отделов, в котором около 30 % видов из данного царства,
312
но их система до сих пор слабо разработана. Краткая характеристика первых четырех отделов приведена ниже в виде таблицы.
Таблица 1 Систематика и основные признаки грибов
(По Н. Грин, У. Стаут, Д. Тейлор. Биология. Том 1, с изменениями)
Oomycota
Половое размножение путем оогамии, когда оосфера (женская гамета) сливается с мужской гаметой, и образуется ооспора
Бесполое размножение посредством зооспор, образующихся в спорангиях
Гифы нечленистые
Примеры:
Phytophthora infestans -факультативный паразит, возбудитель картофельной гнили (рис. 1); Pithium - многие опасные факультативные паразиты; многие вызывают выпревание (черную ножку) проростков; Peronospora - облигатные паразиты, вызывающие ложную мучнистую росу у крестоцветных, например у капусты
Zygoniycota
Половое размножение путем конъюгации, когда два гаметангия сливаются с образованием зигоспоры
Бесполое размножение при помощи конидий или спорангиев, содержащих споры. Зооспор нет (рис.4.4)
Гифы нечленистые; мицелий обширный, ветвящийся
Примеры:
Rhizopus stolonifer -обыкновенная хлебная плесень, сапрофит;
Мисог - обычные плесени, сапрофиты
Ascomycota
При половом размножении внутри специальной структуры, которая называется сумкой или аском, образуются аскоспоры
Бесполое размножение конидиями. Спорангиев не образует
Г ифы членистые
Примеры:
Penicillium и Aspergillus - сапрофитные плесени; Saccharomyces (дрожжи) - одноклеточные сапрофиты;
Erysiphe - облигатные паразиты, вызывающие настоящую мучнистую росу, например у ячменя; Microsphaera alphitoides - паразит на листьях и побегах дуба;
Ceratocystis ulmi -паразит, вызывающий голландскую болезнь вяза
Basidiomycota
При половом размножении образуются базидии, к которым снаружи прикреплены споры (базидиоспоры)
Бесполое размножение спорами. Встречается нечасто
Г ифы членистые
Примеры:
Agaricus campestris -шампиньон обыкновенный, сапрофит (рис. 2); Boletus edulis - белый гриб, сапрофит
Пояснения терминов к таблице 4.2.
Оогамия - тип полового процесса, при котором в ходе оплодотворения половые клетки (гаметы), резко различные по размерам, форме и поведению, сливаются, образуя зиготу. Ооспоры - покоящиеся споры оомикот. Имеют значительный запас питательных веществ и толстую оболочку. После периода покоя прорастают зародышевым спорангием или вегетативно - мицелием. Зооспоры - подвижные споры, служащие для бесполого размножения и расселения. Образуются в зооспорангии. Лишены плотной оболочки. Гаметангий - вместилище гамет. Зигоспоры - образуются в месте касания копулятивных отростков мицелия в результате слияния не дифференцированных по признаку пола гаметангиев. Базидии - органы полового спороношения базидиомикот, образуются на концах гиф, имеют булавовидную форму. Базидиоспоры - в отличие от аскоспор не развиваются внутри сумки (аски), отшнуровываются от нее снаружи. Конидии - споры бесполого размножения. Образуются внутри мицелия на его выростах.
313
Ветвящийся гаусторий внутрири мезофилла клетки-хозяина
Межклеточная гифа
Спорангиеносец
Клетки мезофилла листа
Рис. 1. Фитофтора (Phytophtora in-festans\ растущая в листе, зараженного этим грибом картофеля; на нижней поверхности листа видны свешивающиеся спорангиеносцы (из Грин, Стаут, Тейлор, 1990)
Питание. Грибы - гетеротрофы, т.е. им нужны органические источники углерода. Помимо этого им необходимы источник азота (обычно органический, такой, как аминокислоты), неорганические ионы (например, К+ и Mg2+), микроэлементы (например, Fe, Zn и Си) и органические факторы роста (такие, как витамины). Грибы поглощают питательные вещества, всасывая их всей поверхностью путем диффузии. Это отличает их от животных, которые, как правило, сначала проглатывают пищу, а затем уже переваривают ее внутри своего тела, и лишь потом начинается всасывание питательных веществ. Пищеварение у грибов внешнее, осуществляемое внеклеточными ферментами.
По типу питания грибы бывают сапрофитами, паразитами и симбионтами. В этом отношении они очень похожи на бактерий.
Сапрофитные грибы вырабатывают самые разнообразные ферменты. Некоторые из них могут использовать самые разные субстраты, и их можно назвать поистине вездесущими, например какой-либо из видов PenicilUum. который образует зеленую или голубую плесень на таких субстратах, как почва, сырая кожа, хлеб или гниющие фрукты.
Грибы-сапрофиты обычно образуют большое количество легких устойчивых спор. Это позволяет им легко распространяться на другие продукты. Примерами таких грибов могут служить Miicor. PenicilUum или Agaricus.
Сапрофитные грибы и бактерии образуют вместе группу так называемых редуцентов, без которых немыслимы круговороты элементов в природе. Особенно важны те немногие грибы, которые вырабатывают целлюлазу - фермент, расщепляющий целлюлозу. Целлюлоза представляет собой важнейший структурный компонент клеточных стенок растений. Гниение древесины и других растительных остатков отчасти достигается и за счет деятельности редуцентов, секретирующих целлюлазу.
Некоторые грибы-сапрофиты имеют важное хозяйственное значение; к числу таких грибов относятся, например, дрожжи Saccharomyces или PenicilUum.
В лесу сапрофиты-редуценты способствуют быстрому разложению лесной подстилки, опада, упавших деревьев, они разрушают и минерализуют отмершую растительность, превращая ее в минеральные вещества, доступные для корней живых растений.
Спорангий
Рис. 2. Строение шампиньона обыкновенного (Agaricus campestris). Слева плодовое тело с подземным мицелием; в центре участок гименофора (три гимениальные пластин-
314
ки); справа увеличенная часть гимениальной пластинки (из Грин, Стаут, Тейлор, 1990 с изменениями)
Грибы-паразиты могут быть факультативными или облигатными; чаще они паразитируют на растениях, чем на животных. Облигатные паразиты, как правило, не вызывают гибели своих хозяев, тогда как факультативные паразиты делают это часто и потом живут сапрофитно на мертвых остатках. К облигатным паразитам относятся настоящие мучнисторосные, ложные мучнисторосные, ржавчинные и головневые грибы. Все они, как правило, ограничены узким кругом хозяев, от которых им нужен специфический набор питательных веществ. Факультативные паразиты обычно менее специализированы. Они растут и развиваются на самых разных субстратах и разных хозяевах.
Если хозяином служит растение, то гифы гриба проникают через устьица, или прямо сквозь кутикулу и эпидермис, или же через раны. Попав внутрь растения, гифы обычно ветвятся, распространяясь между клетками; иногда они выделяют пектиназы, которые переваривают ткань растения, и таким образом прокладывают себе дорогу через срединную пластинку (см. рис.1). Заболевание может быть системным, т. е. захватывать все ткани хозяина, или же может ограничиваться небольшой частью растения.
Жизненный цикл у паразитических грибов иногда бывает очень сложным. Это особенно относится к таким облигатным паразитам, как ржавчинные грибы, жизненный цикл которых состоит из нескольких стадий и при этом еще включает более одного хозяина. У облигатных паразитов в результате полового размножения образуются устойчивые споры, что обычно совпадает со смертью хозяина.
Симбиоз. Грибы участвуют в создании двух очень важных типов симбиотического союза, а именно лишайников и микоризы. Лишайник - это симбиотическая ассоциация гриба и водоросли (далее о лишайниках будет сказано подробнее).
Микориза - это симбиотическая ассоциация гриба с корнями растений. Вероятно, большинство наземных растений способно вступать в такого рода связь с почвенными грибами. Гриб образует чехол вокруг центральной части корня (эктотрофная микориза) или же проникает в ткани растения-хозяина (эндотрофная микориза). Микориза первого типа встречается главным образом у таких лесных деревьев, как хвойные, бук и дуб, и образуется при участии грибов, относящихся к отделу Basidiomycota. Их «плодовые тела» (то, что мы зовем грибами) обычно можно видеть вблизи деревьев. Гриб получает от дерева углеводы и витамины и в свою очередь расщепляет до аминокислот белки почвенного гумуса; часть аминокислот при этом поглощается и используется деревом. Кроме того, гриб обеспечивает дерево большей поверхностью всасывания, что особенно важно, когда дерево растет на бедной почве с недостатком азота.
Эндотрофная микориза встречается у самых разнообразных растений, но о ее роли в симбиозе известно очень мало.
Хозяйственное значение грибов.
Сапрофитные грибы играют важную роль в круговоротах биогенных элементов. Вместе с сапрофитными бактериями они образуют группу редуцентов, разлагающих органический материал в почве, повышая ее плодородие.
Сапрофитные грибы вместе с представителями прокариотов и сапрофитными бактериями являются составной частью той желеобразной пленки из живых существ, которая искусственно создается на поверхности фильтров для очистки питьевой воды и воды, возвращаемой производством в естественные водоемы.
Бродильное производство также основано на использовании грибов и бактерий. Старейшее бродильное производство - пивоварение. Пиво получают из ячменя, который предварительно немного проращивают, чтобы превратить крахмал, запасенный в семенах, в сахар мальтозу. Дальнейшее сбраживание проводят в больших чанах, где «работают» одноклеточные грибы-дрожжи из рода Saccharomyces. На этом этапе сахар превращается
315
в двуокись углерода и спирт, конечная концентрация которого достигает 4-8 %. На ранней стадии брожения вносят хмель, который придает пиву аромат и подавляет развитие других микроорганизмов.
Виноделие основано на сбраживании виноградного сока дикими дрожжами, находящимися на кожуре ягод. Конечная концентрация спирта достигает 8-11,5 %. После этого вино выдерживают (правда, не всегда) несколько лет, чтобы оно созрело.
Из других обычных напитков, получаемых в результате брожения, можно упомянуть национальный русский напиток - квас; сидр, приготовляемый из яблочного сока, и японское сакэ, которое делают из риса.
Другая важная отрасль бродильного производства, где тоже применяют дрожжи, хлебопекарная. В пекарнях используются специальные штаммы дрожжей, которые выделяют много двуокиси углерода, помогающей тесту подняться. Еще один продукт, который до сих пор получают из грибов, это лимонная кислота, широко применяемая в пищевой и фармацевтической промышленности. Ее образует гриб Aspergillus niger.
В сыроварении одновременно используют и бактерии, и грибы. Некоторые знаменитые сорта сыра вызревают благодаря «работе» различных видов Penicillium. это рокфор, камамбер, голубой датский сыр и итальянская горгонцола.
Из грибов получают важнейшую группу лекарств - антибиотики. Первым антибиотиком, который стали применять в клинической практике, был пенициллин. Его образуют некоторые виды Penicillium, в частности Р. notatum и Р. chrysogenum. При этом последний вид до сих пор служит источником промышленного производства этого антибиотика.
Грибы, вредные для человека
Сапрофитные грибы играют весьма важную роль в биосфере, однако они причиняют достаточно хлопот человеку, разрушая многие органические материалы. Поэтому при хранении зерна, фруктов и других продуктов необходимо применять самые различные защитные меры. Порча продуктов - это постоянная проблема, которая стоит перед человечеством. Натуральные ткани, кожа и другие товары потребления, изготовляемые из природного сырья, тоже разрушаются грибами. Так, например, грибы, живущие на целлюлозе, вызывают гниль различных лесоматериалов и тканей. На сохранение всех этих материалов расходуются большие средства.
Грибы чаще поражают растения, среди них и самые известные облигатные паразиты, а именно мучнистая роса, ржавчина и головня. Облигатные паразиты не вызывают гибели своих хозяев, но они снижают урожай, а пораженные растения становятся более уязвимыми для других болезней и более чувствительными к неблагоприятным условиям.
Патогенные и дереворазрушающие грибы существенно влияют на лесные древесные растения, заготовленную древесину и пиломатериалы. Список опасных для леса грибов велик, а изучением этих грибов и борьбой с ними занимается специальная наука -лесная фитопатология.
Дальнейшее, основное, содержание настоящего учебника относится к собственно растениям, то есть к царству Regnum vegetabile.
ЦАРСТВО РАСТЕНИЯ
Растения - живые автотрофные организмы, для которых характерны способность к фотосинтезу и наличие плотных клеточных оболочек, состоящих, как правило, из целлюлозы; запасным веществом служит обычно крахмал.
Свойственное некоторым растениям (сапрофитам, паразитам) гетеротрофное питание всегда вторичного происхождения. Другие характерные черты растений (своеобразные циклы развития, способы закладки органов, прикрепленный образ жизни и т. и.) не являются общими для всех групп растений, однако весь комплекс признаков позволяет
316
легко отличать растения (особенно высоко организованные) от представителей других царств.
Основное отличие одноклеточных растений от одноклеточных представителей других царств - наличие хлоропластов в сочетании с особенностями ультраструктуры клеток (строение оболочек, развитие вакуолей и др.). Большинство растений характеризуются сильным расчленением тела, приводящим к увеличению его поверхности, что обусловлено способом питания. Большая поверхность тела - в данном случае увеличение фотосинтезирующей поверхности в надземной части растений, и поглощающей воду с минеральными веществами в подземной.
В современном понимании царство растений включает 3 подцарства: багрянки или красные водоросли, настоящие водоросли и зародышевые или высшие растения. Эти подцарства охватывают все разнообразие мира растений с общим числом видов около 350 тысяч.
Происхождение растений связано с началом развития жизни на Земле. Еще в архее (около 3 млрд, лет назад) появились организмы, похожие на сине-зеленые водоросли или на их предшественников. Настоящие водоросли появились в протерозое. В раннем палеозое известны зеленые и красные водоросли, возможно тогда же (или в конце протерозоя) появились и другие группы настоящих водорослей. Когда растения стали завоевывать сушу - неизвестно. Первые наземные растения появились, вероятно, на границе протерозоя и палеозоя (см. приложение 1).
Особая роль растений в жизни нашей планеты состоит в том, что без них было бы невозможно существование животных и человека. Только зеленые растения, содержащие хлорофилл, способны аккумулировать энергию солнца, создавая органические вещества из неорганических. Как первичные продуценты органических веществ растения представляют собой определяющее звено в сложных цепях питания всех гетеротрофных организмов, населяющих Землю.
Наземные растения представлены самыми разнообразными жизненными формами - травы, кустарники, деревья, лианы, подушковидные растения, эпифиты, стланики и так далее. В разных условиях произрастания они образуют различные растительные сообщества (фитоценозы), обуславливая ландшафтное разнообразие Земли и бесконечное разнообразие экологических условий для других организмов.
ВОДОРОСЛИ ALGAE
Водоросли - это очень большая по численности и разнообразию группа низших растений. Представителей этой группы в быту обычно путают с высшими водными растениями такими, как кувшинка, кубышка, стрелолист и многими другими частично или полностью погруженными в воду. Водоросли не столь заметны, некоторые вообще не видны невооруженным глазом, и естественно, на них меньше обращают внимание, хотя встретить их можно практически в любом, даже очень маленьком водоеме. К ним относятся, например, крупные зеленые скопления так называемой тины, плавающей в летнее время вблизи от поверхности воды в прудах и застойных местах рек и озер. Разнообразные зеленые и сине-зеленые пленки и войлочные или ватообразные наросты на камнях, бревнах и сваях также образованы водорослями. В летнее время вода в прудах часто бывает окрашена в зеленоватый цвет, на просвет можно заметить в ней мельчайшие водоросли в форме плавающих точек, хлопьев или шариков. Здесь же нередко встречаются и более крупные водоросли, состоящие из хорошо заметных на глаз простых или ветвящихся нитей, или совсем крупные харовые водоросли, внешне похожие на хвощ, с характерными мутовками боковых побегов.
Первичную продукцию соленых вод образуют водоросли - единственные растения, которые населяют океан. Учитывая ту огромную площадь, которую занимает океан, следует ожидать, что его продуктивность должна быть еще больше, но нельзя забывать, что
317
фотосинтез возможен только в поверхностных слоях, куда проникает свет и где лимитирующим фактором является доступность биогенных элементов, особенно азота и фосфора.
Водоросли - очень важные первичные продуценты, с которых начинается большинство пищевых цепей, в том числе практически все морские и многие пресноводные цепи.
Некоторые водоросли входят в состав живого напочвенного покрова лесных фитоценозов, обитают в верхних горизонтах лесных почв, однако их роль в жизни леса, в сравнении с ролью высших растений, незначительна.
ЛИШАЙНИКИ UCHENES
Лишайники представляют собой своеобразную группу комплексных организмов, тело которых всегда состоит из двух компонентов - гриба (микобионт) и водоросли (фи-кобионт) (рис.З), традиционно их относят к низшим растениям. Ранние ископаемые лишайники предположительно относятся к верхнему мелу. Предполагается, что они произошли в результате перехода некоторых групп грибов от гетеротрофного способа питания к использованию продуктов фотосинтеза автотрофной водоросли. Долгое время взаимоотношение гриба и водоросли рассматривали как одну из форм симбиоза, в результате которого гриб "снабжает" водоросль водой и неорганическими солями, а водоросль "обеспечивает" гриб органическими веществами. В действительности же отношения между грибом и водорослью больше основаны на паразитизме, особенно со стороны гриба, который, используя продукты жизнедеятельности водоросли еще и питается ее отмершими клетками, будучи иногда сам причиной их отмирания.
Рис 3. Строение талломов некоторых лишайников: 1 - водоросль; 2 - гифы гриба; 3 -ризоиды
Гриб, образующий лишайник, как правило не проявляет избирательности по отношению к водоросли, то есть он пытается приспособить для совместного существования любую доступную ему водоросль, он обладает способностью захватывать водоросли из внешней среды, но далеко не каждая водоросль может существовать в тесном контакте с грибом. Фикобионтами некоторых лишайников являются строго опре-
деленные водоросли, но иногда у одного и того же вида лишайников обнаруживают разные виды водорослей. Некоторые виды водорослей могут быть компонентом тела лишайника и существовать самостоятельно, но другие, например требуксия, входящая в состав 7-12 тысяч видов лишайников, является специализированным компонентом и в свободножи-
вущем состоянии не встречается.
Лишайники, как правило, поселяются на обнаженных скалах или на стволах деревьев; в сырых лесах они еще и свешиваются с деревьев (бородатые лишайники - Usnea longissima. U. barbata) В сосняках лишайниковых ягель или олений мох (на самом деле это "ветвистые" лишайники - кладония оленья (Claddnia rangiferina), кладония лесная (С. sylvatica), кладония приалъпийская (С alpestris) образует плотный, почти белый с оттенками желтого, розоватого, серого, зеленого напочвенный покров (рис. 4).
318
Рис. 4. Напочвенный покров из лишайников рода Claddnia (фото. Коровина)
Форма тела лишайников весьма своеобразная, отличающаяся от свободноживущих грибов и водорослей. Специфичны и физиолого-биохимические процессы лишайников (например, водоросли в слоевище лишайников способны переносить длительное высушивание). Вегетативное тело лишайников - таллом - образовано переплетением грибных гиф и клетками водоросли, либо разбросанными среди гиф по всей толще таллома, либо образующими дифференцированный слой (см рис. 3).
Разнообразна окраска (белый, серый, серо- или желто-зеленый, оранжевый, коричневый, черный и др.) и форма лишайников в виде тонкой или толстой корочки (накипные, или корковые, - леканора, аспицилия), листовидной пластинки (листоватые - коллема, ксантория, пельтигера), прямостоячего кустика или повисающей бороды (кустистые -сферофорус, уснея) (рис. 5). Размножаются лишайники, подобно грибам, спорами, которые образуются на верхушках базидий или в асках, расположенных в плодовых телах -перитециях и апотециях. Апотеции диско- или бокаловидной формы, либо целиком образованы грибными гифами, либо в их края заходят водоросли. Перитеции кувшинообразной формы. Имеет место вегетативное размножение.
319
Рис. 5. Внешний вид некоторых лишайников: 1 - Cora pavoniw, 2 - Claddnia rangiferina (олений мох); 3 - Cladonia pyxidata, - 4 - Thamnolia vermicularis, 5 -Dermatocarpon miniatum, 6 - Grdphis script a: 7 - Parmelia acetabulum, 8 - Rhisocarpon geographicum, 9 - Roccella spec.; 10 - Usnea florida (один из бородатых лишайников)
Относительно систематического положения лишайников нет единого мнения. Одни исследователи классифицируют их в единой системе с грибами, другие - рассматривают их как самостоятельную систематическую группу - отдел в царстве растений. При этом классификации лишайников строятся на основе признаков микобионта, особенностях строения и развития их плодовых тел. Обычно выделяют два класса - сумчатые (Ascolich-enes), к ним относится подавляющее большинство видов и немногочисленные (около 20 видов) базидиальные (Basidiolichenes)
В настоящее время насчитывают свыше. 400 родов и около 26 тыс. видов этих организмов. Распространены лишайники широко, особенно велика их роль в растительном покрове тундровых, лесотундровых и лесных экосистем. Лишайники не являются паразитами деревьев, но могут причинять им косвенный вред, так как в их талломе часто поселяются насекомые-вредители. Лишайники богаты химическими веществами, из которых около 300 специфичны только для них (так называемые лишайниковые кислоты, некоторые обладают антимикробными свойствами). Лишайники участвуют в химическом выветривании пород; используются для получения антибиотиков, ароматических веществ и фиксаторов запахов, изредка как красители шерсти.
Лишайники чувствительны к загрязнению атмосферы, погибают при высоком содержании в воздухе двуокиси серы и других загрязнителей; при этом степень чувствительности варьирует у разных видов, поэтому их используют (многие виды рода уснея) в качестве биоиндикаторов степени загрязнённости окружающей среды (лихеноиндикация). Лишайники используют в геологии для определения возраста ледниковых морен, горных обвалов, так как возраст их талломов нередко насчитывает несколько сотен и тысяч лет. Лишайники - основной корм северных оленей (олений мох, исландский мох, нефрома). 29 видов лишайников занесены в Красную книгу СССР. Наука о лишайниках называется лихенологией.
ВЫСШИЕ РАСТЕНИЯ
Общая характеристика
Появление высших растений ознаменовало начало новой эры в жизни нашей планеты. Освоение растениями суши сопровождалось появлением новых, наземных, форм животных; сопряженная эволюция растений и животных привела к колоссальному разнообразию жизни на Земле, изменила ее облик. Первые наземные растения, известные толь
320
ко по спорам, и время их появления датируется началом силурийского периода. Из верхнесилурийских и нижнедевонских отложений по сохранившимся остаткам или по отпечаткам органов на камнях описаны наземные растения. Эти первые известные нам высшие растения объединены в группу риниофитов. Несмотря на анатомическую и морфологическую простоту строения, это были уже типичные наземные растения. Об этом свидетельствует наличие кутинизированной эпидермы с устьицами, развитой водопроводящей системы, состоящей из трахеид, и наличие многоклеточных спорангиев. Следовательно, можно предположить, что процесс освоения суши растениями начался значительно раньше - в кембрии или в ордовике. Однако возможно, что возникновение растений произошло в геологическом масштабе времени очень быстро.
У самых первых наземных растений, имевших еще небольшие размеры, поглощение воды осуществлялось с помощью ризоидов (аналогу корней) - одноклеточных или многоклеточных однорядных нитей. Однако по мере увеличения размеров тела происходил процесс формирования сложных специализированных органов - корней с корневыми волосками.
Выход растений на сушу сопровождался улучшением их освещения, что активизировало процесс фотосинтеза, это приводило к увеличению объема растений, что вызвало необходимость их морфологического расчленения.
Независимо от формы тела во всех группах наземных растений очень рано (с середины девона) начала проявляться тенденция к образованию плоских боковых фотосинтезирующих органов - листьев. Подобно корням листья возникали разными путями, т.е. в разных систематических группах они имеют разное происхождение. (Своеобразие происхождения листьев нашло отражение в терминологии; так, все листья мохообразных иногда называют филлидами, листья плауновидных - микрофиллами или филлоидами, папоротникообразных - макрофиллами или вайями.) Большая поверхность листьев в сочетании с наиболее совершенной, зернистой формой хлоропластов способствовала увеличению эффективности процессов фотосинтеза, т. е. образованию органического вещества. Быстрое и равномерное распределение пластических веществ по всему телу растения оказалось возможным только при наличии совершенной проводящей ткани - флоэмы, которая обнаруживается уже у нижнедевонских риниофитов.
Таким образом, у высших растений произошло формирование важнейших тканей -эпидермы, ксилемы, флоэмы, механической ткани и важнейших органов - листостебельного побега и корня.
Появившись в нижнем палеозое, высшие споровые растения стали быстро распространяться по поверхности земли, осваивая новые территории.
Многие систематики все ныне живущие высшие растения подразделяют на семь отделов: 1) мохообразные, 2) плауновидные, 3) псилотовидные, 4) хвощевидные или членистые, 5) папоротниковидные, или папоротники, 6) голосеменные, 7) цветковые, или покрытосеменные, растения. Из перечисленных 7 отделов мы кратко рассмотрим 6. Исключаемые нами из рассмотрения псилотовидные - весьма немногочисленная группа растений, произрастающих в тропиках и частично в субтропическом поясе. Это очень древние растения и в настоящее время на Земле сохранились лишь остатки этого отдела.
ОТДЕЛ МОХОВИДНЫЕ (BRIOPHYTA)
Общая характеристика
Мохообразные, представляют собой довольно крупный, насчитывающий около 20 тысяч видов, отдел царства растений. Мхи - наиболее примитивные в категории высших растений, известны с карбона. Они объединяют 2 основных класса: печеночники (Hepati-сае) и листостебельные мхи (Musci) Отдел мохообразные представлен 20-25 тысячами видов, распространенных повсеместно. Поселяются повсюду, кроме морей, засоленных почв и местообитаний , подверженных сильной эрозии.
321
Мохообразные имеют различные приспособления к наземному образу жизни, и в то же время у них сохранились черты водных растений. В большинстве случаев мохообразные слабо приспособлены к обитанию на сухих местах, они растут в среде с повышенной влажностью - болота, леса, сырые луга, где нередко образуют сплошной напочвенный покров, например, сфагновые болота или зеленомошные типы леса. Существуют виды, которые растут только в воде. Отдел моховидные подразделяется на печеночники и собственно мхи.
Мхи - автотрофные растения. В отличие от низших растений тело большинства мохообразных представлено побегом, состоящим из стебля и листьев; только у части мохообразных тело является слоевищем (талломом). Величина мхов от 1 мм до нескольких сантиметров, редко до 60 см и более.
Половое (гаметофит) и бесполое (спорофит) поколения мхов существуют совместно на одном растении. Антеридии имеют вид многоклеточного мешочка, внутри которого образуются мужские гаметы Образующиеся в антеридиях двухжгутиковые сперматозоиды передвигаются к яйцеклетке в архегонии только в капельно-жидкой среде. Архегоний имеет вид многоклеточной колбочки, в расширенной части которой - брюшке архегония -образуется женская гамета, или яйцеклетка.
После оплодотворения из зиготы формируется многоклеточный диплоидный спорофит в виде специализированного органа спороношения - спорогона, состоящего из верхней спороносящей части - коробочки и нижней - ножки со стопой, врастающей в ткань гаметофита. В коробочке спорогония формируется споровый мешок, или спорангий. Споры образуются в результате редукционного (мейотического) деления Созревшая коробочка вскрывается, и споры попадают во внешнюю среду. При наличии благоприятных условий споры прорастают и дают начало новому гаметофиту.
Выросший из спор гаметофит представляет собой многоклеточную протонему (предросток) с многочисленными почками (этим объясняется характерный для мхов рост куртинками, дерновинками, подушками), дающими начало пластинчатым талломам или облиственным побегам. В основном они выполняют функции автотрофного питания. Благодаря этому и способности гаметофита к вегетативному размножению цикл развития мхов длительное время может существовать без образования спорофита, то есть в виде гаплоидного тела.
Печеночники
По своему строению печеночники намного проще, чем мхи, и в целом они более привязаны к сырым и затененным местам. Их можно найти на берегу рек и ручьев, на влажных камнях и среди болотной растительности. У большинства печеночников видны правильные доли или вполне выраженные «стебли» с небольшими простыми «листочками». Проще всего устроены талломные печеночники, тело которых представлено плоским слоевищем и не имеет деления на стебли и листья.
Мхи (Musci или Bryopsidd)
Представители класса мхи характеризуются наличием листостебельных побегов и, как правило, радиальной симметрии. У зеленых мхов листья многослойные; у сфагнума они хотя и однослойные, но при этом дифференцированы на два типа высокоспециализированных клеток. Развивающиеся на стеблях или листьях ризоиды всегда многоклеточные, часто обильно ветвящиеся. Они образуют войлок на поверхности стеблей либо, переплетаясь, формируют шнуровидные тяжи. Коробочка спорогония в отличие от таковой у печеночников имеет сложное строение; она состоит из урночки и крышечки. Внутри ур-ночки формируется спорангий со спорами. При созревании крышечка отскакивает, способствуя активному выбрасыванию спор. Развивающаяся из споры протонема резко отличается от взрослого гаметофита; она имеет форму ветвящейся нити либо пластинки.
Подавляющее число видов мхов в отличие от печеночников приурочено к сырым местообитаниям умеренных и холодных областей, где они играют большую роль в фор
322
мировании ландшафтов северной тайги и тундры. Незначительное число видов перешло ко вторично водному образу жизни. Несмотря на широкое распространение мхов по всему земному шару, каждый род, а иногда вид, проявляет избирательность к механическому и химическому составу почв: одни мхи, предпочитающие богатые почвы, селятся на гниющих стволах, на навозе, на трупах животных; другие, а их большинство, являются олиго-трофами - они осваивают бедные песчаные или каменистые почвы. Класс мхи включает более 14,5 тыс. видов и объединяет три подкласса - сфагновые, андреевы и зеленые мхи. Мы рассмотрим некоторые особенности первого и третьего подклассов.
Подкласс сфагновые, белые, или торфяные мхи (Sphagnidae)
Подкласс сфагновые мхи представлен одним родом сфагнум (Sphagnum), объединяющим свыше 300 видов, которые морфологически довольно трудно различаются. В России произрастает 42 вида. Сфагновые мхи наиболее широко распространены в умеренной зоне Северного полушария, где на обширных пространствах они формируют верховые сфагновые болота. Поселяясь на влажных местах, многие виды сфагнума способствуют быстрому заболачиванию территорий благодаря способности активно и быстро поглощать влагу и прочно ее удерживать.
Сфагновые мхи произрастают густыми куртинками, образуя на поверхности почвы сплошной ковер. Он подобно губке способен активно поглощать влагу. Молодые побеги, густо покрыты мелкими листьями, а в основании развиваются многоклеточные ризоиды. С началом ветвления побегов образование ризоидов прекращается, и взрослые растения сфагнума полностью лишены их. На уровне каждого 4-5-го листа на побеге закладывается почка, которая, начав развиваться, сразу неоднократно ветвится, в результате чего формируется пучок из 2-7 боковых веточек. На вершине главного побега, где узлы еще сближены, пучки этих молодых веточек образуют плотную головку.
Сфагнум, будучи многолетним растением, никогда не достигает больших размеров - у разных видов они колеблются от 5 до 20 см. Это объясняется тем, что по мере нарастания молодых частей побега его прошлогодние участки отмирают. Однако разложение отмерших частей растения в пересыщенной влагой закисленной среде затруднено. В анаэробных условиях происходит процесс накопления торфа (отмерших, но не сгнивших остатков сфагнума), поэтому сфагновые мхи часто называют торфяными мхами. Разрастание сфагнового напочвенного покрова обычно приводит к образованию верхового болота.
Подкласс зеленые мхи (Bryide)
Подкласс брииды, или зеленые мхи (около 14 тыс, видов, 700 родов и 85 семейств) широко распространены по всему земному шару, однако наибольшее участие в сложении растительного покрова они принимают в умеренных и холодных областях, формируя на обширных пространствах тундровые болота, заболоченные и влажные таежные леса. Экологическая амплитуда их довольно широка, так как они могут жить в воде, на обнаженных скалах, на деревьях и даже в пустынях и степях. Однако большинство видов приурочено к тенистым, влажным местообитаниям.
Виды политрихум (Polytrichum), дикранум (Dicranum) и гипнум (Нурпит) принимают большое участие в сложении напочвенного покрова северных хвойных лесов и болот. Обширный род бриум (Вгуит) распространен в лесах и болотах всех континентов. Род тортула (Tortula) приурочен к засушливым областям; одни виды являются степными, а другие пустынными. Некоторые виды рода гриммия (Grimmia) часто выступают в качестве пионера растительности на камнях и скалах, а виды ракомитриума широко представлены в каменистых тундрах. Основное число видов зеленых мхов приурочено к лесам Северного, полушария (рис. 6).
323
Рис. 6. Морфология (типы ветвления) зеленых мхов: 1 - Diceanum’, 2 - Tortilla, 3 -Funaria, 4 - Rhodobrium, 5 - Climdcium, 6 - Hylocomium, 7 -Cratoneuron, 8- Mnium (из Еленевский, Соловьева, Тихомиров, 2006)
Зеленые мхи характеризуются различными жизненными формами, размерами, продолжительностью жизни. Немногочисленны, но весьма своеобразны виды однолетних мхов, имеющих ничтожно малые размеры до 1 мм. Наиболее крупные мхи достигают длины 20-30 см, к ним относятся кукушкин лен (Polytrichum commune), фонтиналис (Fontinalis). Некоторые австралийские виды достигают в длину 50 см. Мхи обладают разнообразным типом ветвления, причем молодые побеги возникают не в пазухе листа, а под листом, отчленяясь от его основания. Боковые ветви у одних видов нарастают неопределенно долго, у других обладают ограниченным ростом. Расположение боковых ветвей может быть очередное, мутовчатое, многоярусное. Редкие мхи не ветвятся, например кукушкин лен (рис. 7). На побегах в большом количестве возникают обильно ветвящиеся ризоиды. У некоторых видов ризоиды густо покрывают стебли в виде войлока; иногда, переплетаясь между собой, они образуют толстые прочные шнуры. Листья возникают из сегмента, который отчленяется от верхушечной клетки. Листья простые, состоят из широкого основания и цельной листовой пластинкой самой разнообразной формы - овальной, яйцевидной, ланцетовидной.
Большое биологическое значение имеет конфигурация листовой пластинки - в сечении она чаще всего бывает желобчатая, волнистая, складчатая, килеватая, очень редко плоская. Скудные лучи света попадают на неровную поверхность листа, частично отражаются и падают на другой лист, от него - на третий. В результате один солнечный луч может достичь поверхности нескольких листьев. Подобный отражательный эффект создают краевые бесцветные клетки, которые у многих видов образуют беловатый ободок.
324
Рис. 7. Кукушкин лен (Polytrichum commune}. 1 - женский гаметофит с архегония-ми; 2 - коробочка с колпачком; 3 - внешний вид коробочки; 4 - продольный разрез коробочки; 5 - поперечный разрез коробочки; 6 - вскрывшаяся коробочка, высыпаются споры; 7 - мужской гаметофит; 8 - вершина мужского гаметофита с антеридиями; Ур - урночка; кол - колонка; сиг - спорангий; к - крышечка; к - колпачек; ан - антеридий (из Еленев-ский, Соловьева, Тихомиров, 2006, с небольшими изменениями)
Развитие гаметофита начинается с прорастания спор. Споры всех зеленых мхов микроскопически мелкие, содержат хлоропласты и капли масла. Протонема подразделяется на зеленую хлоронему и подземную бесцветную ризонему, выполняющую функции корней.
Вегетативное размножение у зеленых мхов распространено широко и проявляется весьма разнообразно. Почти все части гаметофита листостебельных мхов способны производить вторичную протонему, на которой образуются молодые растения. Наиболее простой и широко распространенный способ вегетативного размножения, который наблюдается у мхов постоянно и повсеместно, связан с разрастанием моховой дернины и обособлением при этом молодых побегов, когда нижняя часть разветвленного материнского побега отмирает. Такое разрастание, ведущее к образованию целых моховых ковров, особенно типично для сфагновых, но часто наблюдается и в других группах зеленых мхов.
На развитом гаметофите формируются половые органы: женские - архегонии и мужские - антеридии Как уже отмечалось, одни виды мхов однодомные, другие - двудомные. Однако двудомность мхов носит, относительный характер, так как женские и мужские экземпляры могут развиваться на одной протонеме. Мужские растения у дву
325
домных видов часто сильно редуцированы до так называемых карликовых мужских растений, которые после образования антеридиев отмирают.
ОТДЕЛ ПЛУНОВИДНЫЕ (LYCOPHYTA)
Плауновидные представляют собой самую древнюю группу из числа ныне живущих высших растений; род плаун {Lycopodium), известный еще с верхнего девона, является поистине живым ископаемым. Все плауновидные имеют облиственные побеги и весьма своеобразные корни. Наиболее характерной особенностью плауновидных является микрофилия, под которой понимаются относительно мелкие размеры листьев. Чешуевидные листья плауновидных имеют цельную листовую пластинку и несколько расширенное основание, иногда в виде подушечки.
Спорангии плауновидных формируются на верхней стороне спорофиллов, которые у подавляющего числа видов образуют специализированные спороносные побеги - стробилы в виде колосков.
Рис. 8. Плаун булавовидный {Lycopodium clavdtus). 1 - взрослый про-таллиум; 2 - проталлиум с молодым растением; 3 - продольный разрез еще закрытого антеридия; 4 - сперматозоиды; 5 - разрез молодого, еще закрытого, архегония; 6 -зрелый оплодотворенный архегоний, высыпаются споры; 7 - общий вид растения с вертикально стоящими спорангиями ("булавами" или "колосками"); 8 - отдельный спорофилл, отделенный от спорангия; 9 -вид споры с двух сторон (из Strasburger, Noll, Schenck, Schimper, 1967)
Из представителей рода Lycopodium в наших лесах чаще всего можно встретить плаун булавовидный (рис. 8), предпочи
тающий легкие почвы сосновых насаждений.
ОТДЕЛ ХВОЩЕВИДНЫЕ {EQUISETOPHYTA)
Хвощевидные или членистые берут свое начало с верхнедевонского периода, однако расцвета они достигли в каменноугольном периоде, когда были широко представлены разнообразными древесными и травянистыми формами. Членистые принимали большое участие в сложении знаменитых каменноугольных лесов. Однако в перми начинается их угасание, и прежде всего вымирают древесные формы, так что из мезозоя известны только травянистые членистые. До настоящего времени от всей этой когда-то многочисленной группы дошел только один род хвощ {Equsetum), насчитывающий 25 видов. Поэтому хвощи, подобно плаунам, в такой же степени являются живыми ископаемыми.
Название членистых отражает специфику строения их побегов, которые расчленены на четко выраженные узлы и междоузлия, легко распадающиеся на членики. Члени-стость обусловлена мутовчатым листорасположением листьев и наличием в нижних частях междоузлий интеркалярной (вставочной) меристемы, по которой и происходит разламывание на членики.
Для современных хвощей характерны своеобразные листья - их листовые пластинки сильно редуцированы до небольших темных, иногда зеленых или бесцветных зубчи
326
ков, а хорошо выраженные влагалища срослись в общее пленчатое влагалище, Оно плотно (охватывает стебель и осуществляет защиту нижних меристематических тканей.
Для репродуктивных органов членистых характерно наличие стробилов в виде колосков. У современных хвощей спорангиофоры (т.е. органы, несущие спорангии) имеют щитковую форму.
К порядку, хвощевые относятся исключительно травянистые растения, у которых верхушечные стробилы или спороносные зоны состоят только из мутовок щитковидных спррангиофоров без чередующихся вегетативных листьев. До настоящего времени дожил только один род хвощ (Equisetum), объединяющий 25 видов. Хвощи - травянистые многолетние корневищные растения с однолетними, реже многолетними надземными побегами (рис. 9). Они широко распространены в Евразии, Северной и Южной Америке, Северной Африке. Некоторые из них, преимущественно североамериканские виды, достигают в высоту нескольких метров.
Большинство евразийских видов образуют невысокие надземные однолетние побеги, которые ветвятся с образованием боковых ветвей I, реже II порядка. Надземные побеги у ряда видов могут совмещать две функции - спороносную и вегетативную. Так, например, у хвоща болотного (Е. palustre) и приречного, или топяного (Е. fluviatile), вегетативные и спороносные побеги, возникающие одновременно, вначале не отличаются морфологически друг от друга. Только в середине лета на некоторых зеленых побегах обнаруживаются стробилы.
327
Рис. 9. Хвощи. А - хвощ полевой (Equisetum arvense). 1 -спороносный побег; 2 -вегетативный побег; 3, 4 - спорофиллы (спорангиефоры) со спорангиями; 5, 6 - споры со скрученными и развернутыми гаптерами (нитевидными приспособлениями для разбрасывания спор). Б: 1 - проталлиум с развивающимся на нем архегонием (хвощ луговой - Е. pratense); 2 - сперматозоид хвоща полевого; 3 - развивающийся из споры зародыш, на схеме он разделен на 4 зоны, в каждой из которых закладываются специализированные части будущего хвоща; 4 - проталлиум с зародышем растения (Е maximum); а - архего-ний; г - гаусторий; к - корень; л - листовая мутовка; ст - стебель (из Strasburger, Noll, Schenck, Schimper, 1967 с небольшими изменениями)
У других видов наблюдается раннее разделение функций побегов. Так, у хвоща лесного (Е. silvaticum) и хвоща лугового (Е. pratense) весной одновременно с вегетативными побегами развиваются неветвящиеся бесцветные или розоватые спороносные побеги. Однако после спороношения они зеленеют, ветвятся и не отличаются от вегетативных побегов.
У большинства видов, произрастающих в России, колосовидные стробилы развиваются только на главном побеге, реже (у хвоща болотного) они могут формироваться и на боковых побегах. Видовые различия стробилов проявляются в размерах (от 2-5 мм до 5-8 см), окраске, форме колосков.
Как лесные растения могут доминировать в напочвенном покрове, известны хво-щевые ельники, лиственничники, в Приморье - хвощевые дубняки.
ОТДЕЛ ПАПОРОТНИКООБРАЗНЫЕ (PTERIDOPHYTA)
Подобно плауновидным и хвощам папоротникообразные являются древнейшей группой, известной с девонского периода. Расцвета они достигли в конце палеозойской -начале мезозойской эры, когда были представлены большим разнообразием жизненных форм и распространены на всех континентах земного шара. Наибольшую роль в сложении растительного покрова Земли играли крупные древовидные папоротники, входившие в состав каменноугольных лесов. В настоящее время папоротники насчитывают более 10 тыс. видов и 300 родов.
Для папоротникообразных характерно сочетание ряда признаков, из которых главнейшими являются макрофилия, отсутствие камбия и отсутствие, стробилов. Под макро-филлией понимаются прежде всего относительно крупные размеры листьев, которые часто называются вайями. В отличие от плауновых и членистых листья папоротников имеют более сложное морфологическое и анатомическое строение; они состоят из основания (филлоподия), черешка и листовой пластинки, часто многократно рассеченной, с густой сетью жилок. Для листовой пластинки наиболее характерен длительный рост ее вершины.
Ныне живущие папоротники распространены на всех континентах, встречаясь в самых разнообразных экологических условиях. Однако ведущая роль и по количеству видов, и по разнообразию жизненных форм принадлежит папоротникам влажных тропических и субтропических лесов, где многие семейства находятся в расцвете формообразования. Условия обитания наложили существенный отпечаток на их анатомоморфологические и биологические особенности. Современные папоротники представлены в основном травянистыми растениями, а древовидные составляют незначительное количество. Папоротники умеренных областей, за исключением салъвитш плавающей (Salvinia nutans). - многолетние травянистые растения с подземными длинными или короткими корневищами. У длиннокорневищных папоротников - у орляка (Pteridium aquilinum), го-локучника Линнея (Gymnocarpium dryopteris) и др. - длина междоузлий измеряется сантиметрами, поэтому над землей листья располагаются на большом расстоянии друг от друга. У большинства же папоротников на коротких корневищах формируется розетка листьев, как правило, отмирающих осенью, при этом их расширенные основания длительное
328
время остаются на корневище, образуя плотный толстый чехол вокруг довольно тонкого, диаметром до 1 см, стебля.
У большинства видов папоротников листья совмещают две функции - фотосинтеза и спороношения, однако у многих видов наблюдается диморфизм листьев - одни выполняют функцию фотосинтеза, другие и функцию размножения.
Размножение папоротников осуществляется в основном за счет спор (рис. 10). Большинство видов папоротников - равноспоровые растения; количество разноспоровых
видов невелико. Для всех папоротников характерно отсутствие специализированных спороносных побегов - стробилов. У большинства папоротников спорангии сгруппированы в
сорусы, спорангии или сорусы располагаются на нижней поверхности листьев. Чаще всего они приурочены к главной и боковым жилкам, реже (например, у орляка) - близ края листа. Расположение спорангиев на нижней стороне листа оказывается биологически выгодным: во-первых, обеспечивается надежная защита спорангиев во время их созревания и одновременно не снижается интенсивность фотосинтеза. Во-вторых, обеспечивается более равномерное рассеивание спор, в-третьих, создается возможность большего продуци
рования спорангиев на всей поверхности листа, чем только по краю.
Рис. 10. Папоротник (щитовник) мужской (Dryopteris filix-masy. 1 - общий вид; 2 - сорус в разрезе (спорангии прикреплены к плаценте, некоторые спорангии раскрыты); 3 - еще не раскрывшиеся сорусы на нижней стороне листа; 4 - зрелые раскрывшиеся сорусы (из Strasburger, Noll, Schenck, Schimper, 1967)
Вегетативное размножение широко распространено среди папоротников. Часто оно осуществляется с помощью выводковых почек, возникающих на листьях, стеблях и корнях. В последнем случае корни, загибаясь, выходят на поверхность земли и на вершине формируют почку. Нередко на листьях и стеблях развиваются длинные плети, на которых возникают почки.
ОТДЕЛ ГОЛОСЕМЕННЫЕ (PINOPHYTA)
Голосеменные - древняя группа, берущая свое
начало с верхнего девона. Расцвет голосеменных приходится на мезозойскую эру, когда господствовали теплолюбивые виды. К концу мезозоя происходит сокращение таких теплолюбивых форм. Второе рождение голосеменные обрели благодаря появлению холодостойких видов в недрах мезозойской флоры. Возможно, что их возникновение было связано с местными внутриматериковыми оледенениями. Эти виды оказались способными в кайнозойскую эру осваивать обширные пространства в умеренных и умеренно-холодных областях Северного полушария, где они и поныне на больших территориях занимают гос
подствующее положение в сложении растительного покрова. На территории России голосеменные (класс хвойные) доминируют на севере европейской части страны, в Западной и Восточной Сибири и на Дальнем Востоке. Широкое распространение голосеменных в значительной степени объясняется тем, что размножение у них осуществляется семенами.
Семя как зачаток нового организма представляет собой комплексную многоклеточную форму существования растения, состоящую из зародыша, запасающих тканей эндосперма и многослойной семенной кожуры, надежно обеспечивающей сохранность зародыша. У большинства голосеменных семена обладают периодом покоя. Таким образом, молодые растения вступают в самостоятельную жизнь более жизнеспособными, чем спо
ровые растения.
329
При прорастании семени первым появляется зародышевый корешок, который формирует стержневую корневую систему; лишь у немногих видов главный корень заменяется системой боковых или придаточных корней. Наличие главного корня явилось базой для возможного формирования крупных древесных форм. Этому способствовало также наличие длительно функционирующего камбия, который формирует древесину - водопроводящую ткань, обладающую большой механической прочностью.
Среди современных голосеменных преобладают древесные формы, однако важно подчеркнуть, что среди древних палеозойских видов в разных систематических группах голосеменных встречались формы травянистые, кустарниковые, древесные со слабо развитой древесиной.
У подавляющего большинства голосеменных побеги нарастают моноподиально. Примитивные голосеменные характеризуются отсутствием ветвления, большинство же ныне живущих видов обильно ветвится. Из числа современных древние виды имеют крупные, сильно рассеченные листья, у большинства же видов листья мелкие, игловидные или чешуевидные. Древесина голосеменных состоит в основном из трахеид, паренхима развита довольно слабо, либриформ отсутствует. Характерным признаком трахеид голосеменных является наличие окаймленных пор, ровными рядами расположенных преимущественно на радиальных стенках. Для флоэмы характерно отсутствие клеток-спутниц.
Все голосеменные - разноспоровые растения; в подавляющем большинстве случаев имеются микро- и мегастробилы различной формы, которые, в свою очередь, могут объединяться в констробилы.
Помимо хвойных к голосеменным относятся малочисленные по видовому составу и практически не встречающиеся на территории нашей страны ныне живущие саговниковые, гинкговые и оболочкосеменные.
Подкласс хвойные (Pinidae)
Наряду с цветковыми хвойные принадлежат к числу наиболее нам знакомых и наиболее важных растений. В хвойных лесах накоплена огромная масса органического вещества, представляющая собой ценнейший источник древесины и других важных растительных ресурсов. Хвойные существенно влияют на экологическую ситуацию планеты. Русский быт и русская культура исторически неразрывно связаны с лесом и преимущественно с его хвойными растениями. Достаточно того, что основная масса бытовых, казенных и культовых построек были из дерева, главным образом, из сосны и лиственницы. Из них же изготовлялась мебель и многие другие предметы обихода.
Русское название "хвойные" от слова "хвоя" - игловидные листья - не совсем точное, так как далеко не у всех хвойных листья имеют игольчатую форму, встречаются хвойные с яйцевидной и ланцетной формой листа.
Хвойные - самая многочисленная и наиболее распространенная группа среди современных голосеменных. В настоящее время насчитывают 7 семейств, около 55 родов и не менее 560 видов хвойных. На обширных пространствах Северной Евразии и Северной Америки они образуют леса, часто представленные чистыми (состоящими из одного вида древесных растений) насаждениями.
По своей древности хвойные превосходят все ныне живущие группы семенных растений, в том числе даже гинкговые. В то время как от класса гинкговых сохранился лишь один вид ("живое ископаемое" - гинкго двулопастной), хвойные представляют собой процветающую группу. Среди современных хвойных самыми древними семействами являются араукариевые. подокарповые и особенно сосновые.
Многие хвойные представляют собой высокие и стройные деревья, иногда достигающие гигантских размеров (травянистых растений среди современных хвойных нет). Знаменитая калифорнийская секвойя вечнозеленая (Sequoia sempervirens) из семейства таксодиевых достигает более 100 м высоты (самый высокий из измеренных экземпляров 112 м), а диаметр стволов может достигать Им. Максимальный диаметр Мамонтова де
330
рева (Sequoiadrndron giganteum) - 12 м, но рекордсмен - тпаксодиум мексиканский (Taxodium mucronatum) - известен экземпляр с диаметром ствола 16 м.
Среди хвойных зарегистрированы и самые знаменитые долгожители - представители родов Taxus (тисс) и Sequoiadendron по некоторым сведениям могут жить до 4000 лет, но максимальный возраст из известных - 4900 и достигла его сосиа долговечная (Pinus longaeva). Эта сосна более, чем на 200 лет старше пирамиды Хеопса, "на ее глазах" прошла практически вся история человеческой цивилизации!
Ветвление у хвойных моноподиальное, молодые деревья многих хвойных имеют правильную симметричную крону. У таких растений как ель или араукария ветви располагаются сжатыми спиралями, приближающимися к мутовкам, причем ежегодно образуется лишь одно кольцо ветвей. Это дает возможность легко определить возраст дерева. Старые деревья, у которых резко замедляется прирост в высоту, имеют широкую раскидистую крону с несколькими главными ветвями. Это хорошо заметно, например, у старых сосен.
Корневая система как правило стержневая, но у некоторых видов, например у ели, главный корень довольно быстро замирает, из боковых корней формируется поверхностная корневая система. Для большинства хвойных характерны различные типы микоризы.
Анатомическое строение стеблей хвойных характеризуется слабым развитием сердцевинной паренхимы и мощным развитием вторичной ксилемы, состоящей из длинных трахеид и паренхимы, представленной преимущественно однорядными лучами. Окаймленные поры трахеид с торусами у разных видов могут располагаться как на радиальных, так и на тангентальных стенках. В древесине и коре многих хвойных имеются смоляные каналы или смоляные клетки. Листья покрыты толстой эпидермой с мощной кутикулой. По многим признакам у всех вечнозеленых хвойных листья имеют ксеро-морфную* структуру.
Стробилы у хвойных всегда однополые, они образуют собрание стробилов - конст-робилы. Мужские - у большинства хвойных мелкие, занимают пазушное положение и образуют рыхлые или плотные скопления.
На оси женских констробилов располагаются кроющие чешуи, в пазухах которых формируются семенные чешуи, на их верхней (внутренней) стороне располагаются семязачатки. Семенная чешуя это сильно видоизмененный спороносный побег - мегастробил, так как семенная чешуя располагается в пазухе кроющей, а в пазухе листа может возникнуть только побег.
Порядок хвойные (Pinales или Coniferales)
В подклассе хвойные один порядок, включающий семь семейств, из которых представители трех естественно на территории России. Представители семейства тиссовые (Тахасеае) приурочены в основном к Северному полушарию. В нашей стране естественно произрастают два вида: тисс ягодный и тисс остроконечный. Семейство кипарисовые (Cupressaceae) насчитывает 19 родов и свыше 130 видов. На территории Российской Федерации и стран СНГ естественно произрастает 21 вид из рода можжевельник (Juniperus) и 1 вид из рода микробиота (Microbiota) - дальневосточное эндемичное растение.
Семейство сосновые (Pinaceae) - самое обширное семейство среди всех голосеменных и самое важное в хозяйственном отношении. Оно насчитывает 10 родов с 250 видами, приуроченными в основном к Северному полушарию, и включают главные лесообразующие породы в умеренной и умеренно холодных областях Европы, Азии и Америки. По сравнению с другими семействами хвойных сосновые появились позже всех - род сосна (Pinus) известен из юры, ель и кедр - из нижнего мела, пихта, лиственница и другие роды - из третичных отложений кайнозойской эры. Этим, по-видимому, объясняется большой объем видов в пределах некоторых родов - сосна насчитывает 100 видов, пихта и
Ксероморфность - особенности строения, обеспечивающие существование в засушливых условиях.
331
ель - по 40-50 видов каждая. В основном это вечнозеленые, реже листопадные растения. Чаще всего это деревья высотой 20-30 м. Некоторые виды имеют форму стланикового кустарника. Сосновые, как правило, долгожители; в благоприятных условиях некоторые виды сосен доживают до 3-4 тыс. лет, хотя средний возраст - 200-400 лет. Листья у большинства видов имеют форму хвои, реже они чешуевидные или линейные. Все сосновые имеют зимующие почки, часто надежно защищенные смолистыми веществами. Корневая система характеризуется обильным ветвлением боковых всасывающих корней; почти все виды - микоризные. Древесина имеет хорошо выраженные годичные кольца и систему горизонтальных и вертикальных смоляных ходов.
Все сосновые - однодомные растения. Мужские шишки - микростробилы - одиночные или собраны в рыхлые констробилы. Они состоят из оси, на которой плотной спиралью располагаются плоские микроспорофиллы. На их нижней стороне формируется по два погруженных микроспорангия. Пылинки у большинства снабжены двумя воздушными мешками, способствующими их распространению на огромные расстояния (исключение составляют лиственница, тсуга и псевдотсуга). Женские шишки - констробилы, как правило, одиночные; на оси спирально располагаются маленькие кроющие чешуи, в пазухах которых свободно, не срастаясь с ними, сидят крупные семенные чешуи - мегастробилы. На верхней стороне семенной чешуи находятся два семязачатка. Перед опылением ось шишки немного удлиняется и семенные чешуи раздвигаются, что способствует опылению. Семена созревают либо в год образования шишки (у ели, пихты, лиственницы), либо на следующий год (у сосны). Зародыш у разных видов имеет от четырех до 18 семядолей. У сосновых наблюдается огромное разнообразие форм шишек, но при этом у всех видов семенные чешуи имеют кожистую или деревянистую консистенцию. У пихты и лжелиственницы шишки при созревании рассыпаются, у остальных родов они опадают целыми. Размеры зрелых шишек колеблются от 2-3 см (у некоторых видов лиственницы) до 30-50 см (у некоторых видов сосны).
Хозяйственное значение сосновых чрезвычайно велико. Древесина многих видов высоко ценится в качестве строительного материала, в столярном деле. Ровная древесина ели и пихты, обладающая ценнейшими акустическими свойствами, используется для изготовления лучших музыкальных инструментов. Огромное значение имеет древесина сосновых в целлюлозно-бумажной промышленности. Однако не меньшее значение сосновых заключается в оздоровлении экологической среды за счет выделения озона и бактерицидных летучих веществ. Поэтому кроме естественных лесов как в Северном, так и в Южном полушарии широко культивируются искусственные посадки сосны и ели.
ОТДЕЛ ПОКРЫТОСЕМЕННЫЕ ИЛИ ЦВЕТКОВЫЕ РАСТЕНИЯ (ANGIOSPERMAE ИЛИ MAGNOLIOPHYTA)
Общая характеристика отдела
Отдел покрытосеменные - наиболее крупный отдел растительного царства, насчитывающий более 350 семейств, 13 тыс. родов и до 240 тыс. видов. В настоящее время они распространены по всему земному шару - от тропиков до арктических и антарктических пустынь.
Автотрофные цветковые растения - важнейший активный компонент биосферы. От их жизнедеятельности зависит течение общебиосферных процессов обмена веществ и трансформации энергии, газовый состав атмосферы, климат, водный режим суши, характер процессов почвообразования.
В наземных условиях цветковые обычно доминируют в растительном покрове, «каркас» большинства экосистем образуют именно они. Только в немногих сообществах и типах растительности они уступают господство по общей массе другим группам растений - в хвойных лесах голосеменным, в некоторых типах тундр и на сфагновых болотах - мо
332
хообразным, но и в таких случаях число видов покрытосеменных обычно превышает видовое разнообразие других отделов и подотделов растительного царства.
Понятно, что покрытосеменные определяют возможность существования большинства наземных животных. Они формируют среду обитания животного населения. Животные связаны с ними самыми разнообразными трофическими и иными связями. Многие группы животных вообще могли возникнуть только тогда, когда Землю завоевали покрытосеменные и образуемые ими сообщества.
Покрытосеменные растения отличаются необычайным полиморфизмом, удивительной эволюционной пластичностью и могут существовать в самых разнообразных условиях обитания, вплоть до безводных пустынь, скал, солончаков, разбитых песков, шахтных терриконов, трещин в асфальте или бетоне. Это единственная группа среди высших растений, представители которой сумели вторично освоить морскую среду: в соленых водах морских мелководий вместе с водорослями растут десятки видов покрытосеменных.
Разнообразие размеров и жизненных форм цветковых растений, их приспособлений к конкретным условиям обитания поражает воображение. Самый крохотный их представитель, ряска вольфия (Wolffia arrhiza), едва достигает 1-1,5 мм в диаметре и похожа на свободно плавающую зеленую водоросль, а попросту - на булавочную головку. С другой стороны, гигантские эвкалипты (виды рода Eucalyptus из семейства Myrtdceae) вырастают высотой до 100 м, а длина побегов тропических лиан - ротанговых пальм измеряется сотнями метров. Преобладают среди покрытосеменных автотрофные растения, но немало среди них полупаразитов, паразитов и сапротрофов, полностью утративших хлорофилл и способность к фотосинтезу. Эпифиты (живущие на листьях других растений), деревья, кустарники, кустарнички, полукустарники и травы (многолетники и однолетники), плотоядные растения с разнообразными приспособлениями для улавливания и переваривания мелких животных - это общеизвестные примеры. А тончайшие приспособления к опылению строго определенными агентами, к распространению плодов и семян, к перенесению неблагоприятных климатических периодов. Все это дает цветковым растениям возможность в полной мере реализовать свои способности к приспособлению. Ни одна группа растений не могла выработать такого разнообразия приспособлений к определенным факторам среды.
Характернейшим признаком покрытосеменных считают цветок. Цветки покрытосеменных необычайно разнообразны, и многие считают, что нельзя допустить, будто в прошлом у них был единый прототип. Истина, возможно, заключается в том, что нужно отказаться от поисков некоторого общего для всех покрытосеменных архетипа цветка и признать, что все современное разнообразие цветков сформировалось уже в пределах этой группы в ходе ее эволюции. Покрытосеменные сначала приобрели совокупность действительно важнейших для них общих признаков - в первую очередь уникальные особенности жизненного цикла, а уже потом выработали разные цветки. При этом какого-то одного генерального направления эволюции не существовало, и в каждой эволюционной ветви покрытосеменных были осуществлены различные морфогенетические решения.
Но при любых взглядах на цветок существенные его части - это андроцей, состоящий из одной, нескольких или многих тычинок (микроспорофиллов), и гинецей - из одного, нескольких или многих плодолистиков (мегаспорофиллов).
Важнейшая часть пестика - завязь. Именно наличие завязи как замкнутой полости, в которой развиваются семязачатки (в отличие от голосеменных, у которых семязачатки расположены на поверхности открытых мегаспорофиллов), обычно считают важнейшим признаком покрытосеменных, откуда и название - Angiospermae (от греч. aggeion - сосуд, вместилище, sperma - семя). Благодаря образованию завязи расширяются и обогащаются возможности распространения зачатков: из цветка формируется плод, и теперь разлетаться и расселяться могут уже не только отдельные семена, но и целые плоды, у которых
333
возникают разнообразные приспособления к распространению семян самым широким кругом агентов.
Как и у некоторых голосеменных, у цветковых растений спермин доставляются к яйцеклетке с помощью пыльцевой трубки Пыльцевая трубка обеспечивает высокую точность попадания спермиев в зародышевый мешок. Однако коль скоро семязачатки скрыты в завязи и пыльца попасть непосредственно на микропиле не может, необходим какой-то специальный механизм, обеспечивающий улавливание пыльцы. Таким улавливающим аппаратом у покрытосеменных служит рыльце на верхушке столбика. Наличие рыльца -очень важная особенность покрытосеменных.
Одна из наиболее характерных особенностей покрытосеменных - двойное оплодотворение, при котором один из спермиев (п) сливается с яйцеклеткой (п), образуя зиготу (2п), а второй - с ядром центральной клетки зародышевого мешка (2п), давая тем самым начало триплоидному эндосперму (Зп).
Одно из следствий двойного оплодотворения - формирование своеобразной питательной ткани, триплоидного эндосперма (в отличие от гаплоидного эндосперма голосеменных, представляющего собой женский гаметофит). Он обеспечивает более благоприятные условия для развития зародыша и поэтому мог сыграть важную эволюционную роль.
В вегетативной сфере подавляющее большинство покрытосеменных характеризуется наличием в ксилеме сосудов, тогда как почти у всех голосеменных древесина состоит только из трахеид. Лишь у немногих цветковых древесина бессосудистая. Вообще специализация тканей сыграла важную эволюционную роль. Паренхиматизация позволила покрытосеменным выработать специальные запасающие ткани и разнообразные травянистые жизненные формы. В отличие от деревянистых растений у трав возможно сокращение онтогенеза и ускорение темпов эволюции; травянистые растения способны существовать в таких крайних условиях, в которых деревья и кустарники жить не могут.
Систематика покрытосеменных сложна и до настоящего времени далека от законченности. Все многочисленные, предлагавшиеся в разное время, иерархические системы предполагают происхождение одной современной группы растений из другой, тоже ныне существующей, что методически неправильно. Правда, в таких случаях уточняют, говоря о предполагаемых общих предках, но достоверных общих предков в палеоботанических материалах обычно не обнаруживается.
Следует иметь в виду, что представители разных порядков во многих случаях обнаруживают ту или иную степень внешнего сходства (например наличие колючек у кактусов и у некоторых розоцветных, скажем, у шиповника и боярышника), но такое сходство не доказывает родства. Оно может говорить только о параллелизме в историческом развитии и вовсе не предполагать существование общего предка.
Вместе с тем, отдел покрытосеменные достаточно естественно делится на два класса: двудольные и однодольные. Они обособились на заре эволюции покрытосеменных. Установление в пределах этих классов естественных, единых по происхождению, таксонов рангом выше порядка вряд ли правомерно. Можно допустить, что двудольные и однодольные образуют как бы две большие семьи, каждая из которых объединяет родных братьев и сестер и попытки их группировки по принципу "кто на кого больше похож" будут искусственными. Одновременно существуют взгляды, согласно которым некоторые порядки внутри двудольных, как и внутри однодольных, не обязательно связаны общностью происхождения. Иными словами, порядки, входящие в состав двудольных, хоть и отличаются достоверно от порядков внутри однодольных, но вовсе не обязательно являются "членами одной семьи". То же самое относится и к классу однодольные.
Что касается таксонов, относящихся к таксономическим категориям ниже порядка, то здесь просматривается закономерность, согласно которой, чем выше ранг таксономической категории, тем меньше сходства, а вместе с тем и филогенетического родства, между подчиненными таксонами. Например, род береза (Betula) включает виды, внешнее (мор
334
фологическое) сходство между которыми вызывает мало сомнений, - береза повислая (В. pendula) отличается от березы ребристой (В. costata), но и в той, и в другой все равно легко узнать березу. Вместе с тем, в семействе березовые (Betulaceae) представители родов береза и ольха (Alnus) по большинству морфологических признаков очень не похожи друг на друга, и их принадлежность к одному семейству основывается только на некотором сходстве в строении цветков и соцветий.
335
СПИСОК ИСПОЛЬЗОВАННОЙ И РЕКОМЕНДУЕМОЙ ЛИТЕРАТУРЫ
Аксенов Е.С., Аксенова Н.А. Декоративные растения, Т.1 (Деревья и кустарники). Энциклопедия природы России. М.: ABF, 1997. 560 с.
Алексеев Ю.Е., Жмылев П.Ю., Карпухина Е.А. Деревья и кустарники. Энциклопедия природы России. М.: Издат. ABF, 1997. 589 с.
АрронетН.И. Растительная клетка. - Жизнь растений. Т. 1. Гл. ред. Ал.А. Федоров. М.: Просвещение, 1974. 488 с.
Атрохин В.Г., Калуцкий К.К., Тюриков Ф.Т. Древесные породы мира. Т. 3. Древесные породы СССР. М.: Лесная промышленность, 1982. 264 с.
Бенькова В.Е., Швейнгрубер Ф.Х. Анатомия древесины растений России. Атлас для идентификации древесины деревьев, кустарников, полукустарников и деревянистых лиан России. Штудгарт, Вена.: Издат. Хаупт Берн, 2004. 456 с.
Берг Л.С. Труды по теории эволюции. Л.: Наука, 1977. 387 с.
Биологический энциклопедический словарь (БЭС). М.: Советская энциклопедия, 1989. 864 с.
Большая советская энциклопедия (БСЭ). Электронная версия. М: Советская энциклопедия, 1970-1977.
Бородина Н.А., Некрасов В.И., Некрасова Н.С., Петрова И.П., Плотникова Л.С., СмирноваН.Г. Деревья и кустарники СССР. М.: Мысль, 1966. 637 с.
Брынцев В. А., Коровин В.В. Ботаника. Учебник для средних специальных учебных заведений. М.: ООО "ЭкоСервис", 2007. 320 с.
Булыгин Н.Е. Дендрология. 2- изд. М.: МГУЛ, 2003. 528 с.
Буш Н.А. Общий курс ботаники. Систематика растений. Петроград: Издание А.Ф. Девриена, 1915. 527 с.
Вавилов Н. И. Закон гомологических рядов в наследственной изменчивости. Л.: Наука, 1987. 256 с.
Васильев А.В., Воронин Н.С., Еленевский А.Г. Серебрякова Т.Н. Ботаника. Анатомия и морфология растений. М.: Просвещение, 1978. 480 с.
Вертьянов С.Ю. Общая биология. Учебник для 10-11 классов общеобразовательных учреждений. Ред. акад. РАН Ю.П. Алтухов. М.: Свято-Троицкая Сергиева Лавра, 2006. 352 с.
Вихров В.Е. Диагностические признаки древесины главнейших лесохозяйственных и лесопромышленных пород СССР. М.: Изд. АН СССР, 1959. 132 с.
Вольф Э.Л. Хвойные деревья и кустарники Европейской и Азиатской частей СССР. Л.: Изд. Ленинградского лесного ин-та, 1925. 175 с.
Гончарова С.Б. Систематика растений: основные термины и понятия. http://botsad.ru/p papers37.htm
Гроздов Б.В. Дендрология. М.,Л.: Гослесбумиздат, 1952. 436 с.
Горшков В.Г., Макарьева А.М., 31 июля 2005 г. О видообразовании http://www.biotic-regulation.pl.ru/Russian/speciat.htm
Грин Н., Стаут У., Тейлор Д. Биология: в 3-х т. Т. 1. М.: Мир, 1990. 368 с.
Грин Н., Стаут У., Тейлор Д. Биология: в 3-х т. Т. 2. М.: Мир, 1990. 325 с.
Грин Н., Стаут У., Тейлор Д. Биология: в 3-х т. Т. 3. М.: Мир, 1990. 374 с.
Деревья и кустарники СССР. 1-6 тт. Ред. С.Я. Соколов. М.-Л.: АН СССР, 1949-1962 гг.
Джеффри Ч. Биологическая номенклатура. Перевод с англ. Е.Б. Алексеева. М.: Мир, 1980. 119 с.
336
Еленевский А.Г., М.П. Соловьева, В.Н. Тихомиров. Ботаника. Систематика высших, или наземных, растений. Учебник для студентов высших педагогических учебных заведений. - 4-е изд., испр. - М.: Издательский центр "Академия", 2006. - 464 с.
Законы наследования признаков Менделя
schools.keldysh.ru/schl952/Pages/Timokhina204/mendel.html
Зуихина С.П., Коровин В.В. Практикум по дендрологии. Учебное пособие для средних учебных заведений по специальности "Лесное и лесопарковое хозяйство". М.: ООО "Издательский дом "Лесная промышленность", 2006. 80 с.
Зуихина С.П., Коровин В.В., Тимофеенко Е.И. Покрытосеменные древесные растения. Учебное пособие. М.: Изд. МГУЛ, 2009. 176 с.
Изменчивость, http://zooznaika.ru/1399, shtml
Кернер фон-Марилаун А. Жизнь растений. Т. 2. История растений. С. Петербург, 1902. 842 с.
Козубов Г.М., Данилова М.Ф. (Ред.) Атлас ультраструктуры растительных клеток. Петрозаводск, 1972. 2006 с.
Коровин В.В. Структурные особенности коры березы пушистой (Betula pubescens Ehrh.) - Биологические науки, - № 11, 1970. - С. 55-60.
Коровин В.В. Биология. Учеб. пос. для студентов спец. 2601.00. М.: Изд. МГУЛ, 2001. 199 с.
Коровин В.В., Курносов Г.А. Строение стебля древесных растений при аномальном росте. -М.; Издат. МГУЛ, 2004. - 193 с.
Коровин В.В., Оганесянц Л.А. Дуб в лесоводстве и виноделии. М.: ДеЛи принт, 2007. 480 с.
Крамер П.Д., Козловский Т.Т. Физиология древесных растений. М.: Леси, пром-сть, 1983.464 с.
Лотова Л.И., Тимонин А.К. Определение древесных пород по анатомическим признакам древесины и коры. М.: КМК. Scientific Press, 1997. 39 с.
Лотова Л.И. Ботаника. Морфология и анатомия высших растений. 3 Изд. 3-е, испр. - М: КомКнига, 2007. - 512 с.
Лотова Л.И., Нилова М.В., Рудько А.И. Словарь фитоанатомических терминов: Учебное пособие. М.: издат. ЛКИ, 2007. 112 с.
Лесорастительное районирование.
http: //www. m suee, ru/htm!2/b ooks/wedeni e/ strani су/8, htm
Меницкий Ю.Л. Дубы Азии. Л.: Наука, 1984. 316 с.
Наследственность, http://www.ronl.ru/zabolevaniya/21341 ,htm
Новицкая Л.Л. Карельская береза: механизмы роста и развития структурных аномалий. Петрозаводск: изд. "Verso", 2008. 143 с.
Овсянников В.Ф. Лиственные породы. Пособие для учащихся и лесных специалистов. 2-изд. Владивосток.: ОГИЗ-Далькрай, 1931. 373 с.
Петрашов В.В. Глаза и мозг эволюции. Новая теория эволюции организмов. 3-е изд. М.: Прометей, 2006. 458 с.
Природные зоны России http://www. msuee. ru/htm!2/books/wedenie/straniсу/8,htm
ПчелинВ.И. Дендрология: учебник. Йошкар-Ола.: Изд. Марийского государственного технического университета, 2007. 520 с.
Рейвн П., Эверт Р., Айкхорн С. Современная ботаника. Т 1, 2. М.: Мир, 1990.
Соболев Д.Н. Начала исторической биогенетики. Симферополь: Государственное издательство Украины, 1924. 204 с.
Сочава В.Б. Опыт деления Дальнего Востока на физико-географические области и провинции. Доклады Ин-та географии Сибири и Дальнего Востока, 1962, № 1.
Сукачев В.Н. Дендрология с основами лесной геоботаники. Л.: Гослестехиздат, 1938. 576 с.
337
Усенко Н.В. Деревья, кустарники и лианы Дальнего Востока. Хабаровск: Хабаровское книжное изд., 1984. 290 с.
Фотосинтез, http://referat-5.msk.ru/fotosintez.php, http://www.krugosvet.ru/articles/l 16/1011698/1011698al.htm, http://www.ecosystema.ru/07referats/01/photosintes.htm
Царев А.П., Погиба С.П., Тренин В.В. Генетика лесных древесных пород. Петрозаводск: Издат. ПТУ, 2000. 338 с.
Чайковский Ю.В. Наука о развитии жизни. Опыт теории эволюции. М.: Товарищество научных изданий КМК, 2006. 712 с.
Чайковский Ю.В. Активный связный мир. Опыт теории эволюции жизни. М.: Товарищество научных изданий КМК, 2008. 726 с.
Чистякова О. Н., Барыкина Р. П., Вахмистров Д. Б. Корень. Большая советская энциклопедия (компьютерная версия).
Эзау К. Анатомия семенных растений. Книга 1, 2. - М.: Мир, 1980. - 558 с.
Юсуфов А.Г., Магомедова М.А. История и методология биологии. М.: Высшая школа, 2003. 238 с.
Яценко-Хмелевский А. А. Основы и методы анатомического исследования древесины.-М-Л.: Издат. АН СССР, 1954. - 337 с.
Яценко-Хмелевский А.А. Краткий курс анатомии растений. - М.:Высшая школа, 1961.-282 с.
Braune W., Leman A., Taubert Н. Pflanzenanatomisches Prakticum. - Jena. Gustav Fisher verlag, 1967. -331 S.
Molisch H, K. Hofler. Anatomie der Pflanze. - Jena, Gustav Fischer, 1965. - 202 S.
Strasburger E., F. Noll, H. Schenck, A.F.W. Schimper. - Lehrbuch der Botanik. - Jena, Gustav Fischer, 1967. - 762 S.
Wagenfuhr R. Anatomie des Holzes. - Leipzig: Fachbuchverlag, 1966. - 377.
338
ТЕРМИНОЛОГИЧЕСКИЙ СЛОВАРЬ
о Автогенез - учение, объясняющее эволюцию организмов действием только внутренних нематериальных, или пока непонятных, факторов.
о Автотрофные организмы - организмы, синтезирующие из неорганических веществ (главным образом воды, СОг, неорганических соединений азота) все необходимые для жизни органические вещества, используя энергию фотосинтеза (все зеленые растения - автотрофы) или хемосинтеза (большая часть бактерий - хемотрофы).
о Акропетальный - направленный к верхушке. Противопоставляется базипетальному -направленному к основанию.
о Алеврит - рыхлая осадочная горная порода, по составу промежуточная между песками и глинами с размерами основной массы зерен 0,01-0,1 мм.
о Аллель - одно из возможных состояний гена в данном локусе. Например: доминантный аллель или рецессивный аллель; дикий (неизмененный) аллель или мутантный аллель.
о Ангстрем (А) - внесистемная единица длины, равная 0,1 нм (нанометра).
о Антиклинальные деления - деления в плоскости перпендикулярной поверхности органа или ткани.
о Ароморфоз - усложнение организации и функций организмов в процессе их эволюции.
о Базипетальный - направленный к основанию.
о Биологические мембраны — белково-липидные структуры молекулярных размеров, до 100 А толщиной, образующие плазматическую мембрану (плазмалемму) и ограничивающие внутриклеточные образования (ядра, митохондрии, пластиды и пр.).
о Биоценоз - совокупность экологически связанных между собой животных, растений, грибов и микроорганизмов, совместно населяющих определенный участок суши или водоема.
о Выветривание - процесс разрушения и химического изменения горных пород в условиях земной поверхности или вблизи нее под влиянием колебаний температуры, химического и механического воздействия атмосферы, воды и организмов. Различают физическое выветривание, химическое и биологическое.
о Ген - элементарная единица наследственности, представляющая отрезок молекулы дезоксирибонуклеиновой кислоты - ДНК (у некоторых вирусов - рибонуклеиновой кислоты - РНК). Каждый ген определяет строение одного из белков живой клетки и тем самым участвует в формировании признака или свойства организма. Совокупность генов - генотип - несёт генетическую информацию о всех видовых и индивидуальных особенностях организма. Доказано, что наследственность у всех организмов на Земле (включая бактерии и вирусы) закодирована в последовательностях нуклеотидов гена. У высших (эукариотических) организмов ген входит в состав особых нуклеопротеидных образований - хромосом. Главная функция генов - программирование синтеза ферментных и др. белков, осуществляющегося при участии клеточных РНК (информационных - и-РНК, рибосомных - р-РНК и транспортных - т-РНК), — определяется химическим строением генов (последовательностью в них дезоксирибонуклеотидов -элементарных звеньев ДНК) При изменении структуры генов (мутации) нарушаются определённые биохимические процессы в клетках, что ведёт к усилению, ослаблению или выпадению ранее существовавших реакций или признаков.
о Геном - совокупность генов, характерных для гаплоидного набора хромосом данного вида организмов, основной гаплоидный набор хромосом. В отличие от генотипа геном представляет собой характеристику вида, а не отдельной особи.
о Генотип - генетическая (наследственная) конституция организма, совокупность всех его генов.
339
о Генофонд - сумма генотипов особей, составляющих популяцию, а генотип - генетическая (наследственная) конституция организма, совокупность всех его генов.
о Гетеротрофные организмы — получающие углерод и прочие вещества за счет других организмов, противопоставляются автотрофным зеленым (фотосинтезирующим) растениям.
о Гетерохрония - эволюционная разновозрастность признаков у одного организма или у группы организмов одного вида. В ботанике синонимом этого понятия является гете-робатмия (разноступенчатость - разные эволюционные ступени развития признака).
о Гуттация - выделение листьями растений капель воды через устьица под воздействием корневого давления.
о Дерезняки - заросли степных кустарников (дереза - Caragana frutex, степная вишня -Cerasus fruticosa, терн - Prunus spinosa, бобовник - Amigdalus папа).
о Дифференцировка (дифференциация) клеток - возникновение различий между однородными (чаще меристематическими) клетками; изменения, приводящие образованию специализированных клеток (тканей, органов). Дифференциация происходит в процессе онтогенеза: у животных большей частью на эмбриональной стадии развития, у растений - как при прорастании семени, так и при ежегодном сезонном возобновлении активности ростовых процессов (древесные и многолетние растения).
о Идиоадаптация - возникновение в процессе эволюции организмов частных изменений в строении и функциях органов при сохранении в целом уровня организации.
о Изменчивость паратипическая - изменчивость организмов, вызванная изменением факторов внешней среды.
о Изоляция - возникновение барьеров (территориальных, экологических, физиологических, поведенческих, генетических), препятствующих свободному скрещиванию организмов.
о Инверсия температуры - повышение температуры воздуха с высотой в некотором слое атмосферы вместо обычного понижения.
о Интерация - цикличность, повторяемость.
о Интрогрессивная гибридизация - естественная гибридизация на границе ареалов двух близких видов, при которой гибридные формы периодически скрещиваются между собой и с исходными видами.
о Ковалентная связь - вид химической связи. Осуществляется парой электронов, находящихся в общем владении двух атомов, образующих связь. Атомы в молекуле могут быть связаны одинарной, двойной или тройной ковалентной связью.
о Компартменты - обособленные части.
о Коферменты - органические соединения небелковой природы, входящие в состав некоторых ферментов. Вместе с ферментом образуют каталитически активные комплексы.
о Крассулы - утолщение межклеточного слоя и первичных оболочек в виде тонких полосок на радиальных стенках трахеид хвойных между парами двухрядно расположенных окаймленных пор.
о Ксерофильные растения - растения способные выдерживать недостаток влаги.
о Ксерофиты - растения, которые могут существовать при небольшом количестве влаги; также пустынные растения или растения засоленных участков, так как избыток солей затрудняет доступность для растений почвенной влаги.
о Ксилема - древесина. Этот термин чаще употребляется по отношению к первичным проводящим элементам - "первичная ксилема", но может относиться к древесине вообще.
о Кутикула - слой жирового вещества, кутина, покрывающего сплошной пленкой поверхность надземных органов многих растений (главным образом листьев, стеблей, некоторых плодов).
о Кутин - воскоподобное вещество, выделяемое клетками эпидермиса растений и от
340
кладывающееся вместе с воском в виде пленки-кутикулы на внешней поверхности клеточной оболочки.
о Ландшафт - достаточно обширный участок земной поверхности, в пределах которого различные компоненты природы (горные породы, рельеф, климат, воды, почвы, растительный и животный мир), взаимосвязаны и взаимообусловлены, составляют одно целое, образуя определенный вид местности.
о Ландшафт географический - однородная по условиям развития природная система, основная категория территориального деления географической оболочки Земли (в широком смысле синоним природного территориального комплекса любого ранга).
о Лигнин - природное полимерное вещество. Входит в состав почти всех наземных растений и по распространенности среди природных высокомолекулярных соединений уступает только полисахаридам. Содержание лигнина в древесине хвойных и лиственных пород соответственно 23-38 и 14-25 %, по массе. Расположен в клеточных стенках (инкрустирует их) и межклеточном пространстве растений и скрепляет целлюлозные волокна древесины. Вместе с гемицеллюлозами он определяет механическую прочность стволов и стеблей. Кроме того, лигнин снижает проницаемость клеточных стенок для воды и питательных веществ.
о Меристема - образовательная ткань растений, долго сохраняющая способность клеток к делению и возникновению новых клеток; отличается высокой метаболической активностью. Так называемые инициальные клетки меристемы задерживаются на эмбриональной стадии развития, и делясь, обеспечивают увеличение массы растения. Производные инициальных клеток после серии делений дифференцируются, превращаясь в клетки постоянных тканей.
о Метаболизм - весь комплекс реакций, протекающих в организме. Близкое понятие -обмен веществ.
о Миграция - включение в данную популяцию генотипов из другой популяции или пространственное перемещение небольшой части исходной популяции.
о Микориза - (грибокорень) симбиоз мицелия гриба и корня высшего растения.
о Модификационные изменения - морфологические или функциональные изменения организмов, вызванные факторами среды и не относящиеся к генотипическим изменениям.
о Муссоны - устойчивые сезонные ветры, направление которых резко меняется на противоположное или близкое к противоположному, 2 раза в год. Основная особенность муссонного климата - обильное осадками лето и сухая зима.
о Мутации — естественные или вызванные искусственно наследуемые изменения генетического материала, приводящие к изменению тех или иных признаков организма.
о Натурфилософия - философия природы, умозрительное (без эксперимента) истолкование природы в ее целостности.
о Номогенез - развитие по закономерности. Понятие относится к теории эволюции.
о Нуклеоиды - оптически неразличимые эквиваленты ядер, свойственны прокариотам.
о Нуклеотиды - нуклеозидфосфаты - фосфорные эфиры нуклеозидов. Состоят из азотистого основания (пуринового или пиримидинового), углевода (рибозы или дезоксирибозы), и остатка фосфорной кислоты. Нуклеотиды - составная часть нуклеиновых кислот, коферментов и других биологически активных соединений.
о Оглеение - процесс восстановления окисных почвенных соединений, главным образом железа, в закисные, придающие почвам особые свойства, снижающие плодородие.
о Олигосахариды - углеводы, состоящие из 2-10 моносахаридных остатков, связанных гликозидными связями. Олигосахариды с 2-5 моносахаридными остатками являются транспортными формами сахаров у растений.
о Онтогенез - индивидуальное развитие, совокупность изменений, претерпеваемых организмом от зарождения до конца жизни.
о Органеллы (органоиды) - структурно и функционально обособленные компоненты
341
клетки.
о Осмос - одностороннее проникновение (диффузия) через полупроницаемую мембрану, отделяющую раствор от чистого растворителя или раствора меньшей концентрации. Обусловлен стремлением системы к термодинамическому равновесию и выравниванию концентраций раствора по обе стороны мембраны.
о Отбор — дифференциальная выживаемость и размножаемость потомства.
о Паразитизм - форма взаимоотношения двух различных организмов, принадлежащих к разным видам и носящая антагонистический характер. Обычно один из них (паразит) использует другого (хозяина) в качестве среды обитания или источника пищи, возлагая на него регуляцию своих отношений с внешней средой.
о Периклинальные деления - деление клеток в плоскости, параллельной поверхности ткани или органа, приводящее к образованию двух смежных слоев.
о Перицикл - слой клеток первичной меристемы в корнях и иногда в стеблях, окружающий проводящий цилиндр и располагающийся под эпидермой.
о Плазмодесмы - каналы в клеточных оболочках, соединяющие протопласты соседних клеток. Состоят из плазмалеммы, выстилающей канал, слоя цитоплазмы и проходящей сквозь нее трубки эндоплазматического ретикулума. Трубка связывает эндоплазматические сети (ретикулумы) соседних клеток. Плазмодесмы объединяют протопласты растительных клеток в единую систему.
о Полигональный - многоугольный.
о Полей перекреста поровость (П.п.п.) - у хвойных растений различают несколько типов:
о П.п.п. купрессоидная - (от лат. Cupressus - кипарис) - расположение на поле перекреста нескольких окаймленных пор со щелевидными отверстиями (кипарис, можжевельник).
о П.п.п. оконцевая - наличие на поле перекреста одной крупной простой поры (сосна обыкновенная).
о П.п.п. пиноидная — (от лат. Pinus - сосна) - наличие на поле перекреста 2, реже 3 пор с едва заметными часто односторонними окаймлениями и крупными отверстиями (сосна корейская).
о П.п.п. пицеоидная - (от лат. Picea - ель) - наличие на поле перекреста мелких пор, щелевидные отверстия которых выходят за пределы окаймлений (ель, старая древесина).
о П.п.п. таксодиоидная — (от лат. Taxodium - таксодиум, болотный кипарис) - наличие пор со слабо выраженными окаймлениями и довольно крупными отверстиями (болотный кипарис)
о Поле перекреста - участок смежных стенок водопроводящего элемента и клетки древесинного луча.
о Полиморфизм - многоформенность, изменчивость морфологических признаков внутри популяции. Иногда это понятие применяют и к изменчивости функций организмов.
о Популяция - это совокупность особей одного вида, обладающая общим генофондом, занимающая определенную территорию, внутри которой осуществляется свободное скрещивание, и отделенная от соседних популяций некоторой степенью изоляции. Под генофондом понимается сумма генотипов особей, составляющих популяцию, а генотип - генетическая (наследственная) конституция организма, совокупность всех его генов.
о Презумпция - обычно презумпция невиновности - обвиняемый считается невиновным пока его вина не доказана. В данном случае - СТЭ верна, пока не доказана ее несостоятельность.
о Прото... — первая часть сложных слов, означающая первенство, первичность, первобытность чего-либо.
о Редуценты (деструкторы) - организмы питающиеся мертвым органическим вещест
342
вом и подвергающие его минерализации - разложению до простых неорганических соединений, которые затем используются растениями (продуцентами).
о Реликты - виды растений и животных, входящие в состав растительного покрова и животного мира данной страны или области как пережитки флор и фаун минувших эпох.
о Рефугиум - убежище.
о Сальтации - внезапные, скачкообразные изменения организмов. Могут касаться одного признака, но чаще - группы взаимосвязанных признаков.
о Сапротрофы (сапрофиты) - гетеротрофные организмы, использующие для питания органические соединения мертвых организмов или выделения (экскременты) животных. Участвуя в минерализации органических соединений, сапрофиты составляют важное звено в биологическом круговороте веществ и энергии.
о Сердцевидные повторения - не очень удачное название небольшого аномального скопления раневой паренхимы, возникающего у некоторых древесных растений в древесине в результате повреждения камбиальной зоны насекомыми.
о Симбиоз - различные формы совместного существования организмов разных видов, составляющих симбионтную систему. Симбиоз бывает факультативным, когда каждый из организмов при отсутствии партнера может жить самостоятельно; и облигатным, когда один из организмов (или оба) оказывается в такой зависимости от другого, что самостоятельное существование невозможно.
о Стробилы - органы размножения некоторых высших растений: плауновидных, хвощевидных, голосеменных. Это видоизмененный укороченный побег, несущий специализированные листья - спорофиллы, на которых формируются спорообразующие органы - спорангии. У разноспоровых растений стробилы дифференцированы на микростробилы (мужские) и мегастробилы (женские стробилы).
о Строма — коллоидная белковая масса, заполняющая внутренние полости пластид.
о Таксон - Группа организмов, связанных той или иной степенью родства и достаточно обособленная, чтобы ей можно было присвоить определенную таксономическую категорию того или иного ранга - вид, род, семейство и т.д. В отличие от таксономической категории таксон всегда подразумевает конкретные биологические объекты.
о Таксономические категории - понятия, принимаемые в систематике для обозначения соподчиненных групп растений, грибов и животных, отличающихся различной степенью родства. Таксономические категории иерархичны - вид, род, семейство, порядок и т.д.
о Таксономия - раздел систематики, теория и практика классификации организмов.
о Тилы (тиллы) - выросты протопластов паренхимных (обычно лучевых) клеток, проникающих через пары пор в полость смежного членика сосуда или трахеиды.
о Транспирация - испарение воды растением.
о Трофия от греч. питание. Трофический уровень - совокупность организмов, объединяемых типом питания.
о Тырса - то же, что ковыль волосатик (Stipa capillata).
о Фауна - исторически сложившаяся совокупность видов животных, населяющих какую-либо территорию, или населявшая ее раньше.
о Фаутность - зараженность патогенными организмами.
о Ферменты - (энзимы) - биологические катализаторы, присутствующие во всех живых клетках. Осуществляют превращение веществ в организме, направляя и регулируя обмен веществ. По химической природе - белки.
о Филогенез - процесс исторического развития мира организмов, их видов, родов, семейств, порядков, классов, отделов, царств.
о Фитоценоз — сообщество растений на одной территории, в той или иной степени и форме, связанных между собой.
о Флора - исторически сложившаяся совокупность видов растений, населяющих какую-
343
либо территорию, или населявшая ее раньше.
о Хемосинтез - процесс образования некоторыми (не фотосинтезирующими) бактериями органических веществ из двуокиси углерода за счет энергии, полученной при окислении неорганических соединений (аммиака, водорода, соединений серы, закисного железа и др.). Хемосинтезирующие бактерии вместе с фотосинтезирующими растениями и микробами составляют группу автотрофных организмов.
о Ценоз - то же, что биоценоз.
о Цитоплазма - внеядерная часть протоплазмы животных и растительных клеток. Состоит из гиалоплазмы, в которой содержатся органеллы (органоиды) и другие включения.
о Эдификаторы - средообразующие виды, виды, определяющие облик биогеоценозов.
о Экотон - комбинация экологических факторов и их режимов в пределах определенного однородного местоположения, обусловленная сочетанием и взаимодействием компонентов неживой природы.
о Экссудат - раствор, транспортируемый по сосудам ксилемы (ксилемный экссудат) и ситовидным трубкам (флоэмный экссудат).
о Эндемики - виды (роды, семейства и т. д.) растений и животных, свойственных относительно небольшой конкретной территории. Вне этой территории они не встречаются.
о Эпифиты - растения, поселяющиеся на других растениях, преимущественно на стволах и ветвях (в отличие от эпифиллов, живущих на листьях), и получающие питательные вещества из окружающей среды (но не из растения, на котором поселились).